
Solubilization of Minerals by Bacteria - NCBI
9
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 1994, p. 3349-3357 Vol. 60, No. 9 0099-2240/94/$04.00+0 Solubilization of Minerals by Bacteria: Electrophoretic Mobility of Thiobacillus ferrooxidans in the Presence of Iron, Pyrite, and Sulfur ROBERT C. BLAKE II,* ELIZABETH A. SHUTE, AND GARY T. HOWARDt Department of Biochemistry, Mehany Medical College, Nashville, Tennessee 37208 Received 2 May 1994/Accepted 17 June 1994 ThiobaciUus ferrooxidans is an obligate acidophile that respires aerobically on pyrite, elemental sulfur, or soluble ferrous ions. The electrophoretic mobility of the bacterium was determined by laser Doppler velocimetry under physiological conditions. When grown on pyrite or ferrous ions, washed cells were negatively charged at pH 2.0. The density of the negative charge depended on whether the conjugate base was sulfate, perchlorate, chloride, or nitrate. The addition of ferric ions shifted the net charge on the surface asymptotically to a positive value. When grown on elemental sulfur, washed cells were close to their isoelectric point at pH 2.0. Both pyrite and colloidal sulfur were negatively charged under the same conditions. The electrical double layer around the bacterial cells under physiological conditions exerted minimal electrostatic repulsion in possible interactions between the cell and either of its charged insoluble substrates. When Thiobacilusferrooxidans was mixed with either pyrite or colloidal sulfur at pH 2.0, the mobility spectra of the free components disappeared with time to be replaced with a new colloidal particle whose electrophoretic properties were intermediate between those of the starting components. This new particle had the charge and size properties anticipated for a complex between the bacterium and its insoluble substrates. The utility of such measurements for the study of the interactions of chemolithotrophic bacteria with their insoluble substrates is discussed. Thiobacillus ferrooxidans is the most extensively character- ized member of a group of chemolithotrophic bacteria that inhabit ore-bearing geological formations exposed to the at- mosphere and obtain all of their energy for growth from the dissolution and oxidation of minerals within the ore. The possibility of exploiting this activity to extract metals for commercial gain, a process known as bacterial leaching, has gained attention in recent years. A widespread application of bioleaching is its use in the pretreatment of refractory gold- bearing ores (16, 19). It is estimated that $10 to $50 million worth of gold was recovered by biooxidation in 1988 and that this value would rise to $2 to $3 billion by 1998 (22). Further- more, the global market for the bioleaching of base metals (mainly copper) and uranium amounted to $2 billion in 1988 and should reach $8 billion by 1998. Despite the environmental and possible economic impor- tance of these bacterial activities, very little fundamental information concerning chemical and biochemical events at the mineral surface during dissolution in the presence of bacteria is available. Although there is electron microscopic (1, 2, 20, 23) and physical (7, 9, 18, 21, 33) evidence for the adsorption of T. ferrooxidans onto mineral surfaces, the relative contribution to the leaching process of sessile [by a direct contact mechanism(s)] versus planktonic (by indirect chemical reactions with soluble ferric ions) organisms remains unclear. Major obstacles include difficulties in enumerating particle- bound bacteria (autotrophic bacteria that are notoriously difficult to culture on solid media) and the microheterogeneity of natural mineral samples. The present paper describes electrophoretic mobility mea- * Corresponding author. Present address: College of Pharmacy, Xavier University, New Orleans, LA 70125. Phone: (504) 483-7489. Fax: (504) 488-3108. t Department of Biological Sciences, Southeastern Louisiana Uni- versity, Hammond, LA 70402. surements of the interaction of T. ferrooxidans with ferric ions, pyrite (FeS2), and colloidal sulfur by laser Doppler velocim- etry. Surfaces of metal sulfide minerals react with anions and cations in solution to acquire a net charge. Bacteria are also amphoteric by virtue of the ionogenic residues on their cell surfaces and the preferential adsorption of ions. Aqueous suspensions containing both types of amphoteric particles may be viewed, in principle, as a mixture of colloids. The goal of these experiments was to investigate whether techniques com- monly employed to study the interactions and stability of colloidal systems could be applied to the interactions of chemolithotrophic bacteria with their insoluble mineral sub- strates. The binding of ferric ions, pyrite, and sulfur to T. ferrooxidans was observed under physiological conditions for this acidophilic bacterium. The net surface charge on T. ferrooxidans was such as to minimize charge repulsive forces in its interactions with any mineral surface. MATERIALS AND METHODS Cell culture. T. ferrooxidans ATCC 23270 was grown au- totrophically on soluble ferrous ions in an apparatus that permitted in situ electrochemical reduction of the product ferric ions to achieve enhanced yields of the bacteria (3). The ferrous sulfate medium (28) was adjusted to pH 1.8 and amended with 1.6 mM cupric sulfate. The bacterial growth chamber was a 12-gallon (ca. 45-liter) glass carboy filled to capacity. Iron-grown cells were harvested by removal and centrifugation of small portions (1 to 2 liters) of the culture, typically containing 1 to 2 g of wet cell paste per liter. The soluble iron in the medium was electrochemically reduced and maintained in a completely reduced state 1 h prior to harvest- ing. The volume in the growth chamber was maintained at a constant level by the addition of fresh sterile medium. Cultivation of T. ferrooxidans on either pyrite (catalog no. 8455; National Institute of Standards and Technology, Gaith- 3349
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Solubilization of Minerals by Bacteria - NCBI

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 1994, p. 3349-3357 Vol. 60, No. 90099-2240/94/$04.00+0
Solubilization of Minerals by Bacteria: ElectrophoreticMobility of Thiobacillus ferrooxidans in the Presence
of Iron, Pyrite, and SulfurROBERT C. BLAKE II,* ELIZABETH A. SHUTE, AND GARY T. HOWARDtDepartment of Biochemistry, Mehany Medical College, Nashville, Tennessee 37208
Received 2 May 1994/Accepted 17 June 1994
ThiobaciUus ferrooxidans is an obligate acidophile that respires aerobically on pyrite, elemental sulfur, orsoluble ferrous ions. The electrophoretic mobility of the bacterium was determined by laser Dopplervelocimetry under physiological conditions. When grown on pyrite or ferrous ions, washed cells were negativelycharged at pH 2.0. The density of the negative charge depended on whether the conjugate base was sulfate,perchlorate, chloride, or nitrate. The addition of ferric ions shifted the net charge on the surface asymptoticallyto a positive value. When grown on elemental sulfur, washed cells were close to their isoelectric point at pH 2.0.Both pyrite and colloidal sulfur were negatively charged under the same conditions. The electrical double layeraround the bacterial cells under physiological conditions exerted minimal electrostatic repulsion in possibleinteractions between the cell and either of its charged insoluble substrates. When Thiobacilusferrooxidans wasmixed with either pyrite or colloidal sulfur at pH 2.0, the mobility spectra of the free components disappearedwith time to be replaced with a new colloidal particle whose electrophoretic properties were intermediatebetween those of the starting components. This new particle had the charge and size properties anticipated fora complex between the bacterium and its insoluble substrates. The utility of such measurements for the studyof the interactions of chemolithotrophic bacteria with their insoluble substrates is discussed.
Thiobacillus ferrooxidans is the most extensively character-ized member of a group of chemolithotrophic bacteria thatinhabit ore-bearing geological formations exposed to the at-mosphere and obtain all of their energy for growth from thedissolution and oxidation of minerals within the ore. Thepossibility of exploiting this activity to extract metals forcommercial gain, a process known as bacterial leaching, hasgained attention in recent years. A widespread application ofbioleaching is its use in the pretreatment of refractory gold-bearing ores (16, 19). It is estimated that $10 to $50 millionworth of gold was recovered by biooxidation in 1988 and thatthis value would rise to $2 to $3 billion by 1998 (22). Further-more, the global market for the bioleaching of base metals(mainly copper) and uranium amounted to $2 billion in 1988and should reach $8 billion by 1998.
Despite the environmental and possible economic impor-tance of these bacterial activities, very little fundamentalinformation concerning chemical and biochemical events atthe mineral surface during dissolution in the presence ofbacteria is available. Although there is electron microscopic (1,2, 20, 23) and physical (7, 9, 18, 21, 33) evidence for theadsorption of T. ferrooxidans onto mineral surfaces, the relativecontribution to the leaching process of sessile [by a directcontact mechanism(s)] versus planktonic (by indirect chemicalreactions with soluble ferric ions) organisms remains unclear.Major obstacles include difficulties in enumerating particle-bound bacteria (autotrophic bacteria that are notoriouslydifficult to culture on solid media) and the microheterogeneityof natural mineral samples.The present paper describes electrophoretic mobility mea-
* Corresponding author. Present address: College of Pharmacy,Xavier University, New Orleans, LA 70125. Phone: (504) 483-7489.Fax: (504) 488-3108.
t Department of Biological Sciences, Southeastern Louisiana Uni-versity, Hammond, LA 70402.
surements of the interaction of T. ferrooxidans with ferric ions,pyrite (FeS2), and colloidal sulfur by laser Doppler velocim-etry. Surfaces of metal sulfide minerals react with anions andcations in solution to acquire a net charge. Bacteria are alsoamphoteric by virtue of the ionogenic residues on their cellsurfaces and the preferential adsorption of ions. Aqueoussuspensions containing both types of amphoteric particles maybe viewed, in principle, as a mixture of colloids. The goal ofthese experiments was to investigate whether techniques com-monly employed to study the interactions and stability ofcolloidal systems could be applied to the interactions ofchemolithotrophic bacteria with their insoluble mineral sub-strates. The binding of ferric ions, pyrite, and sulfur to T.ferrooxidans was observed under physiological conditions forthis acidophilic bacterium. The net surface charge on T.ferrooxidans was such as to minimize charge repulsive forces inits interactions with any mineral surface.
MATERIALS AND METHODS
Cell culture. T. ferrooxidans ATCC 23270 was grown au-totrophically on soluble ferrous ions in an apparatus thatpermitted in situ electrochemical reduction of the productferric ions to achieve enhanced yields of the bacteria (3). Theferrous sulfate medium (28) was adjusted to pH 1.8 andamended with 1.6 mM cupric sulfate. The bacterial growthchamber was a 12-gallon (ca. 45-liter) glass carboy filled tocapacity. Iron-grown cells were harvested by removal andcentrifugation of small portions (1 to 2 liters) of the culture,typically containing 1 to 2 g of wet cell paste per liter. Thesoluble iron in the medium was electrochemically reduced andmaintained in a completely reduced state 1 h prior to harvest-ing. The volume in the growth chamber was maintained at aconstant level by the addition of fresh sterile medium.
Cultivation of T. ferrooxidans on either pyrite (catalog no.8455; National Institute of Standards and Technology, Gaith-
3349

APPL. ENVIRON. MICROBIOL.
ersburg, Md.) or elemental sublimed sulfur (Fisher Scientific,Norcross, Ga.) was accomplished in the liquid medium de-scribed previously (15) containing 0.8 g of monobasic potas-sium phosphate per liter and a 1% (wt/vol) amendment of theinsoluble inorganic substrate. The elemental sulfur was steril-ized by heating the dry powder for 1 h in a boiling-water bathfor each of 3 successive days. The pyrite was autoclaved as thedry powder. Bacterial growth on pyrite or sulfur was monitoredby the solubilization of iron or the acidification of the medium,respectively.
Pyrite- or sulfur-grown cells were harvested in an identicalmanner. The inorganic solids remaining in a 2- to 4-week-old19-liter culture were permitted to settle by gravity, and the bulkof the clarified liquid was decanted. The supernatant was thenconcentrated by tangential flow filtration on a Millipore Pelli-con (Millipore Corporation, Bedford, Mass.) equipped with a100,000-molecular-weight cutoff membrane. Centrifugation ofthe retentate in a Sorvall Refrigerated Superspeed centrifugeat 10,000 x g for 15 min yielded only traces of cell massregardless of whether the growth substrate was pyrite or sulfur,indicating that the majority of the cells remained associatedwith the insoluble substrate. Substrate-associated cells weresubsequently recovered by bringing the filtrate obtained aboveto 0.01% (wt/vol) in Triton X-100 (Sigma Chemical Co., St.Louis, Mo.), adding the amended filtrate back to the particu-late substrate, and repeating the steps described above. Typicalyields varied from 1 to 3 g of wet cell paste per 19 liters.
Harvested cells were routinely washed three times with 0.001N sulfuric acid and stored (1 g of wet cell paste per 4 ml of0.001 N sulfuric acid) at 4°C.
Electrokinetic measurements. Electrophoretic mobility wasdetermined with a DELSA 440 instrument (Coulter ScientificInstruments, Inc., Hialeah, Fla.). Electrophoretic motion wasdetected by the Doppler shift in frequency of scattered coher-ent light by a heterodyne method. Doppler frequency spectrawere obtained simultaneously at four different scattering an-gles. Values for the corresponding mobilities and zeta poten-tials were determined from the Doppler frequency spectra byusing operating and analysis software provided by Coulter.Instrument control and data analysis were accomplished with aGateway 2000 486DX-33 computer (North Sioux City, S. Dak.)interfaced to the DELSA 440.
Samples for analysis were diluted at least 100-fold into thedesired solution and placed in the observation cell so that noair bubbles were visible. The observation cell was positioned inthe instrument so that all measurements were conducted at theupper stationary layer. The frequency shift range was 250 Hz inall experiments. The instrument was operated in the constant-current mode with field strengths, indicated below, that variedwith the conductivity of the analyte. All measurements wereperformed at 25°C.
Bacterial suspensions (108 cells per ml, final concentration)were prepared in 0.01 N solutions of either sulfuric, perchloric,hydrochloric, or nitric acid immediately prior to the measure-ment. Ferric ions, when present, were added as the salt of thecorresponding conjugate base so that each solution containedonly one major species of anion. Colloidal sulfur (RugerChemical Co., Irvington, N.J.), when present, was added froma 1% (wt/vol) suspension in deionized water to a final concen-tration of 0.4 g of sulfur per ml. Ground pyrite (NIST catalogno. 8455) was further reduced by amalgamation on a Silametdental amalgamator (Vivadent, Schaan, Austria) and grindingwith a mortar and pestle. The resulting powder was suspendedin distilled water, and colloidal pyrite was taken as the fractionof suspended fine particles that were decanted after thesuspension had settled for 2 h. The suspension of colloidal
pyrite was brought to pH 2.0 with sulfuric acid for measure-ments that involved pyrite in acid.
Gel electrophoresis. The cell wall and outer membranecomponents of T. ferrooxidans grown on each of the threesubstrates were prepared as described elsewhere (4). Thecomponents of each preparation were separated by sodiumdodecyl sulfate-polyacrylamide gel electrophoresis on a Phast-System with PhastGel Gradient 8 to 25% polyacrylamide gelsand PhastGel SDS Buffer Strips (Pharmacia LKB Biotechnol-ogy, Piscataway, N.J.). The molecular weight standards werethe Dalton Mark VII-L kit from Sigma Chemical Co. The gelswere double-stained according to a silver-Coomassie brilliantblue R-250 staining strategy that permitted color-coded differ-entiation of proteinaceous and sugar-containing components(8). Silver staining was performed on the PhastSystem byadapting the manual protocol for the Bio-Rad silver stain kit(catalog no. 161-0443; Bio-Rad Laboratories, Hercules, Calif.),while Coomassie staining was conducted by the standardautomated protocol suggested by Pharmacia.
Iron determinations. The total iron associated with washedcells of T. ferrooxidans was determined on whole-cell lysates byatomic adsorption spectrometry on a Perkin Elmer 2100(Norwalk, Conn.) operated in the flame mode. Cell numberswere obtained with a Multisizer Ile (Coulter Scientific Instru-ments, Inc.) fitted with a 30-,um aperture. The instrument wasprogrammed to siphon 100 ,ul of sample volume containing0.8% (wt/vol) NaCl as the electrolyte. Cells for iron analysiswere boiled in concentrated nitric acid (10 ml of HNO3 per gof wet cell paste). The resulting solutions were diluted with 1.0N sulfuric acid for iron analyses by atomic adsorption.
RESULTS AND DISCUSSION
Zeta potential. An electrical potential due to charge sepa-ration frequently develops at the interface where two dissimilarmaterials are in contact. Particles immersed in aqueous solu-tions acquire a surface charge for a variety of reasons, the mostprominent being the ionization of surface molecular groupsand the preferential adsorption of ions. Ions of opposite chargeto the charge on the particle surface (counterions) are at-tracted to the surface, while ions of like charge to those on thesurface (co-ions) are repelled. The surface charge plus theexcess of counterions over co-ions near the surface constitutesthe electrical double layer.
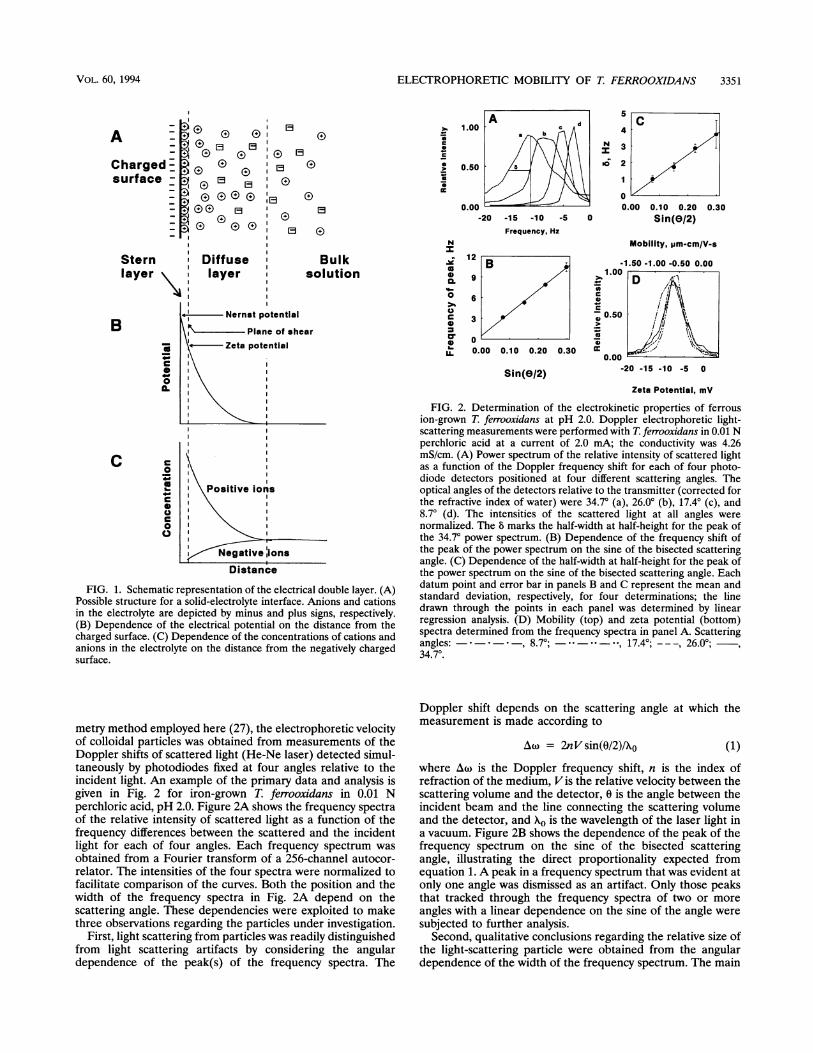
Figure 1 illustrates those features of the electrical doublelayer of immediate interest (14). Most of the surface charge isneutralized by tightly bound counterions in the Stern layer; theremaining charge is balanced by the diffuse or Gouy layer ofcounterions whose concentration decreases as a function of thedistance from the surface. When the charged particle is set intomotion under the influence of an applied electric field, ions inthe Stern layer move with the particle, while ions in the diffuselayer are constantly changing as the particle moves through thecontinuous phase. The imaginary plane inside which the tightlyassociated ions migrate with the particle surface, the shearboundary, lies just outside the Stern layer. The electricalpotential determined at the shear boundary with respect to thebulk liquid is the zeta (t) potential. A negative ; potentialindicates that the particle is negatively charged and migratestoward the anode in an electric field, while a positive ;potential indicates just the opposite. The greater the absolutevalue of the t potential, the greater is the charge density on thesurface.
Electrophoretic mobility. Zeta potential is measured byparticle electrophoresis methods. In the laser Doppler veloci-
3350 BLAKE ET AL.
ELECTROPHORETIC MOBILITY OF T. FERROOXIDANS 3351
*@ 03 03' 0(
(3E (3E 2(
(i) (3 (315:
*(3 (i
Diffuse Bilayer soli
, < Nernst potential
Plane of shear- Zeta potential
Positive ions
Negative ,ions
1.00
0.50c
.-I!
0.00-20 -15 -10 -5 0
N
010
Sin(e/2)Frequency, Hz
ulkution
Distance
FIG. 1. Schematic representation of the electrical double layer. (A)Possible structure for a solid-electrolyte interface. Anions and cationsin the electrolyte are depicted by minus and plus signs, respectively.(B) Dependence of the electrical potential on the distance from thecharged surface. (C) Dependence of the concentrations of cations andanions in the electrolyte on the distance from the negatively chargedsurface.
metry method employed here (27), the electrophoretic velocityof colloidal particles was obtained from measurements of theDoppler shifts of scattered light (He-Ne laser) detected simul-taneously by photodiodes fixed at four angles relative to theincident light. An example of the primary data and analysis isgiven in Fig. 2 for iron-grown T. ferrooxidans in 0.01 Nperchloric acid, pH 2.0. Figure 2A shows the frequency spectraof the relative intensity of scattered light as a function of thefrequency differences between the scattered and the incidentlight for each of four angles. Each frequency spectrum wasobtained from a Fourier transform of a 256-channel autocor-relator. The intensities of the four spectra were normalized tofacilitate comparison of the curves. Both the position and thewidth of the frequency spectra in Fig. 2A depend on thescattering angle. These dependencies were exploited to makethree observations regarding the particles under investigation.
First, light scattering from particles was readily distinguishedfrom light scattering artifacts by considering the angulardependence of the peak(s) of the frequency spectra. The
N
U
0.k
0
0*S
U-
Mobility, pm-cm/V-s
0.00 - 0
Sin(e/2) -20 -15 -10 -5 0
Zeta Potential, mV
FIG. 2. Determination of the electrokinetic properties of ferrousion-grown T. ferrooxidans at pH 2.0. Doppler electrophoretic light-scattering measurements were performed with T. ferrooxidans in 0.01 Nperchloric acid at a current of 2.0 mA; the conductivity was 4.26mS/cm. (A) Power spectrum of the relative intensity of scattered lightas a function of the Doppler frequency shift for each of four photo-diode detectors positioned at four different scattering angles. Theoptical angles of the detectors relative to the transmitter (corrected forthe refractive index of water) were 34.70 (a), 26.00 (b), 17.40 (c), and8.70 (d). The intensities of the scattered light at all angles werenormalized. The 8 marks the half-width at half-height for the peak ofthe 34.7° power spectrum. (B) Dependence of the frequency shift ofthe peak of the power spectrum on the sine of the bisected scatteringangle. (C) Dependence of the half-width at half-height for the peak ofthe power spectrum on the sine of the bisected scattering angle. Eachdatum point and error bar in panels B and C represent the mean andstandard deviation, respectively, for four determinations; the linedrawn through the points in each panel was determined by linearregression analysis. (D) Mobility (top) and zeta potential (bottom)spectra determined from the frequency spectra in panel A. Scatteringangles: - * * , 8.7°; - --- -- 17.4; ---, 26.00;34.70.
Doppler shift depends on the scattering angle at which themeasurement is made according to
Aw = 2nVsin(0/2)/X0 (1)where Aw is the Doppler frequency shift, n is the index ofrefraction of the medium, Vis the relative velocity between thescattering volume and the detector, 0 is the angle between theincident beam and the line connecting the scattering volumeand the detector, and Ao is the wavelength of the laser light ina vacuum. Figure 2B shows the dependence of the peak of thefrequency spectrum on the sine of the bisected scatteringangle, illustrating the direct proportionality expected fromequation 1. A peak in a frequency spectrum that was evident atonly one angle was dismissed as an artifact. Only those peaksthat tracked through the frequency spectra of two or more
angles with a linear dependence on the sine of the angle were
subjected to further analysis.Second, qualitative conclusions regarding the relative size of
the light-scattering particle were obtained from the angulardependence of the width of the frequency spectrum. The main
A -Chargedsurface-
Sternlayer
B
0
.
C c0a
c0aaC.
VOL. 60, 1994
3352 BLAKE ET AL.
Mobility, pm-cm/V-sec
-1 0 I
-10 0 10
Zeta Potential, mV
E
*0
00.co
0N
0
-2
-4
-6
-80 5 10 15 20 25 30
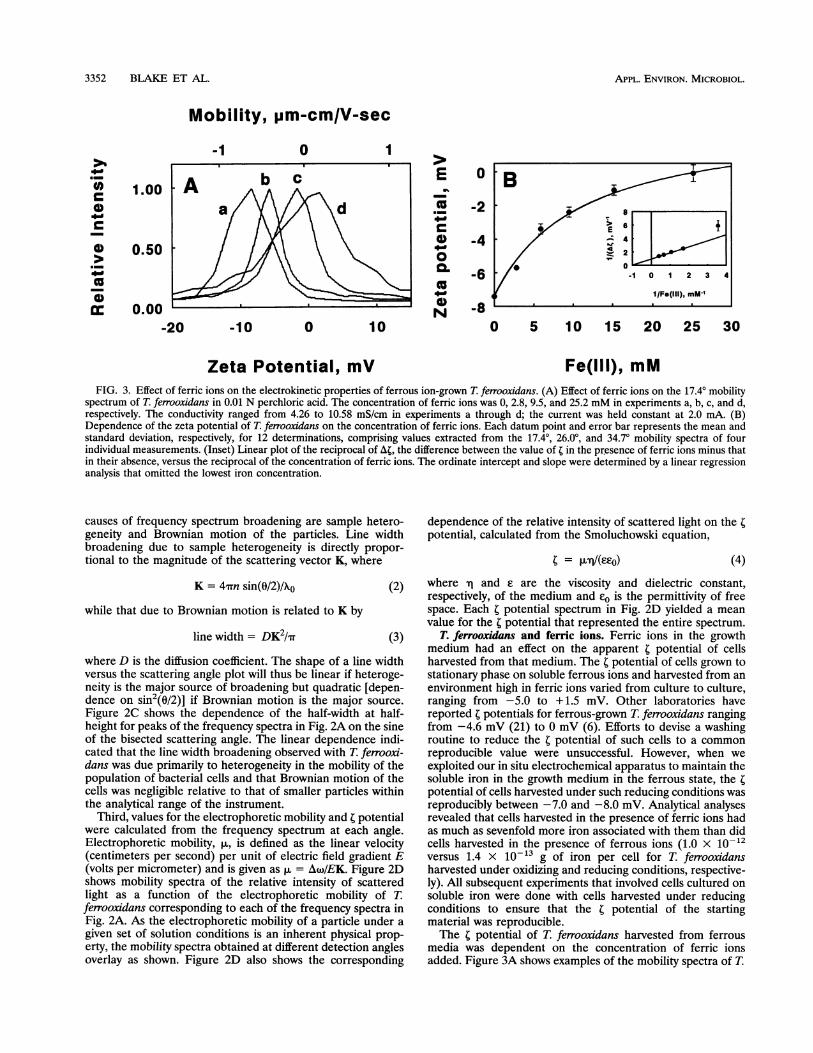
Fe(lIl), mMFIG. 3. Effect of ferric ions on the electrokinetic properties of ferrous ion-grown T. ferrooxidans. (A) Effect of ferric ions on the 17.40 mobility
spectrum of T. ferrooxidans in 0.01 N perchloric acid. The concentration of ferric ions was 0, 2.8, 9.5, and 25.2 mM in experiments a, b, c, and d,respectively. The conductivity ranged from 4.26 to 10.58 mS/cm in experiments a through d; the current was held constant at 2.0 mA. (B)Dependence of the zeta potential of T. ferrooxidans on the concentration of ferric ions. Each datum point and error bar represents the mean andstandard deviation, respectively, for 12 determinations, comprising values extracted from the 17.40, 26.0°, and 34.70 mobility spectra of fourindividual measurements. (Inset) Linear plot of the reciprocal of At, the difference between the value of t in the presence of ferric ions minus thatin their absence, versus the reciprocal of the concentration of ferric ions. The ordinate intercept and slope were determined by a linear regressionanalysis that omitted the lowest iron concentration.
causes of frequency spectrum broadening are sample hetero-geneity and Brownian motion of the particles. Line widthbroadening due to sample heterogeneity is directly propor-tional to the magnitude of the scattering vector K, where
K = 4-rrn sin(0/2)/10 (2)
while that due to Brownian motion is related to K by
line width = DK2/I7 (3)
where D is the diffusion coefficient. The shape of a line widthversus the scattering angle plot will thus be linear if heteroge-neity is the major source of broadening but quadratic [depen-dence on sin2(0/2)] if Brownian motion is the major source.Figure 2C shows the dependence of the half-width at half-height for peaks of the frequency spectra in Fig. 2A on the sineof the bisected scattering angle. The linear dependence indi-cated that the line width broadening observed with T. ferrooxi-dans was due primarily to heterogeneity in the mobility of thepopulation of bacterial cells and that Brownian motion of thecells was negligible relative to that of smaller particles withinthe analytical range of the instrument.
Third, values for the electrophoretic mobility and 4 potentialwere calculated from the frequency spectrum at each angle.Electrophoretic mobility, ,u, is defined as the linear velocity(centimeters per second) per unit of electric field gradient E(volts per micrometer) and is given as ,u = Aw/EK. Figure 2Dshows mobility spectra of the relative intensity of scatteredlight as a function of the electrophoretic mobility of T.ferrooxidans corresponding to each of the frequency spectra inFig. 2A. As the electrophoretic mobility of a particle under agiven set of solution conditions is an inherent physical prop-
erty, the mobility spectra obtained at different detection anglesoverlay as shown. Figure 2D also shows the corresponding
dependence of the relative intensity of scattered light on the 4potential, calculated from the Smoluchowski equation,
C = rl'/(EcO) (4)where -q and £ are the viscosity and dielectric constant,respectively, of the medium and e0 is the permittivity of freespace. Each 4 potential spectrum in Fig. 2D yielded a meanvalue for the 4 potential that represented the entire spectrum.
T. ferrooxidans and ferric ions. Ferric ions in the growthmedium had an effect on the apparent 4 potential of cellsharvested from that medium. The t potential of cells grown tostationary phase on soluble ferrous ions and harvested from anenvironment high in ferric ions varied from culture to culture,ranging from -5.0 to + 1.5 mV. Other laboratories havereported t potentials for ferrous-grown T. ferrooxidans rangingfrom -4.6 mV (21) to 0 mV (6). Efforts to devise a washingroutine to reduce the t potential of such cells to a commonreproducible value were unsuccessful. However, when weexploited our in situ electrochemical apparatus to maintain thesoluble iron in the growth medium in the ferrous state, the tpotential of cells harvested under such reducing conditions wasreproducibly between -7.0 and -8.0 mV. Analytical analysesrevealed that cells harvested in the presence of ferric ions hadas much as sevenfold more iron associated with them than didcells harvested in the presence of ferrous ions (1.0 x 1012versus 1.4 x 10-13 g of iron per cell for T. ferrooxidansharvested under oxidizing and reducing conditions, respective-ly). All subsequent experiments that involved cells cultured onsoluble iron were done with cells harvested under reducingconditions to ensure that the t potential of the startingmaterial was reproducible.The t potential of T. ferrooxidans harvested from ferrous
media was dependent on the concentration of ferric ionsadded. Figure 3A shows examples of the mobility spectra of T.
>,A._
0
0
(a._
m
1.00
0.50
0.00-20
APPL. ENVIRON. MICROBIOL.
ELECTROPHORETIC MOBILITY OF T. FERROOXIDANS 3353
> 2E
0
-2
0. -4 /
-6/N
-80 5 10 15 20 25 30
Fe(III), mM
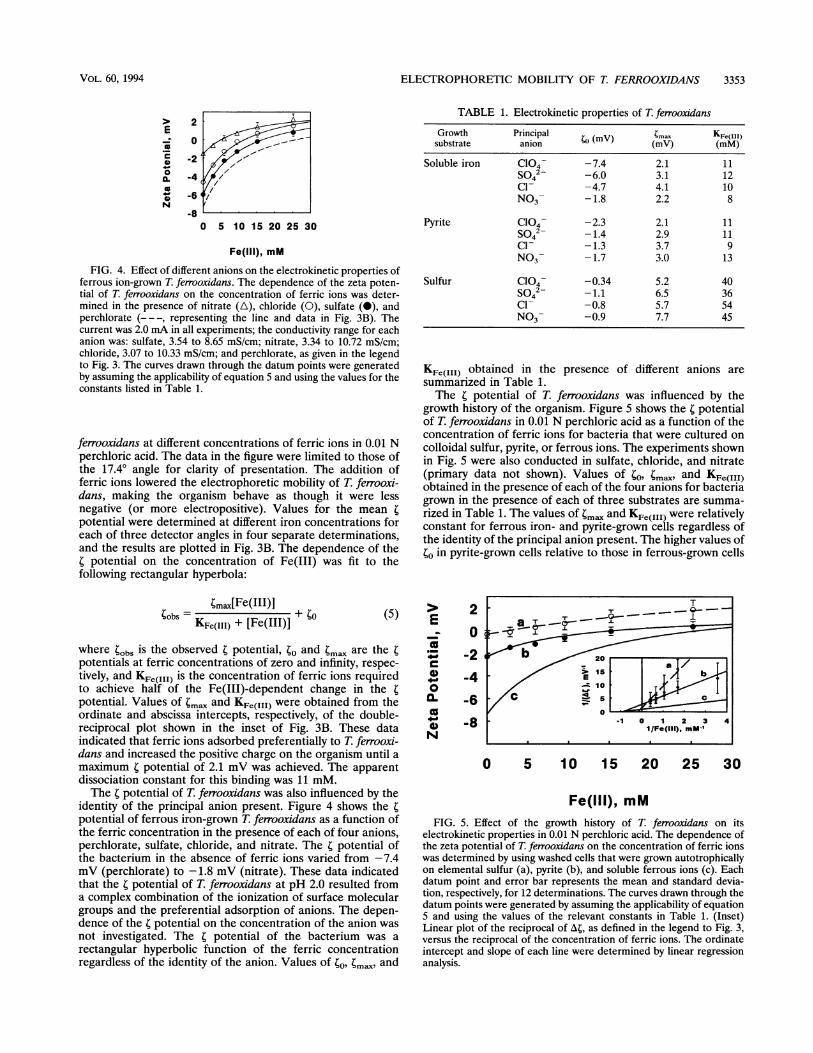
FIG. 4. Effect of different anions on the electrokinetic properties offerrous ion-grown T. ferrooxidans. The dependence of the zeta poten-tial of T. ferrooxidans on the concentration of ferric ions was deter-mined in the presence of nitrate (A), chloride (0), sulfate (0), andperchlorate (- - -, representing the line and data in Fig. 3B). Thecurrent was 2.0 mA in all experiments; the conductivity range for eachanion was: sulfate, 3.54 to 8.65 mS/cm; nitrate, 3.34 to 10.72 mS/cm;chloride, 3.07 to 10.33 mS/cm; and perchlorate, as given in the legendto Fig. 3. The curves drawn through the datum points were generatedby assuming the applicability of equation 5 and using the values for theconstants listed in Table 1.
ferrooxidans at different concentrations of ferric ions in 0.01 Nperchloric acid. The data in the figure were limited to those ofthe 17.40 angle for clarity of presentation. The addition offerric ions lowered the electrophoretic mobility of T. ferrooxi-dans, making the organism behave as though it were lessnegative (or more electropositive). Values for the mean 4potential were determined at different iron concentrations foreach of three detector angles in four separate determinations,and the results are plotted in Fig. 3B. The dependence of thet potential on the concentration of Fe(III) was fit to thefollowing rectangular hyperbola:
tmax[Fe(III) +
KFe(III) + [Fe(III)]
where tbobs iS the observed t potential, to and tmax are the tpotentials at ferric concentrations of zero and infinity, respec-tively, and KF,(tII) is the concentration of ferric ions requiredto achieve half of the Fe(III)-dependent change in the tpotential. Values of tma. and KFe(III) were obtained from theordinate and abscissa intercepts, respectively, of the double-reciprocal plot shown in the inset of Fig. 3B. These dataindicated that ferric ions adsorbed preferentially to T. ferrooxi-dans and increased the positive charge on the organism until amaximum t potential of 2.1 mV was achieved. The apparentdissociation constant for this binding was 11 mM.The t potential of T. ferrooxidans was also influenced by the
identity of the principal anion present. Figure 4 shows the tpotential of ferrous iron-grown T. ferrooxidans as a function ofthe ferric concentration in the presence of each of four anions,perchlorate, sulfate, chloride, and nitrate. The t potential ofthe bacterium in the absence of ferric ions varied from -7.4mV (perchlorate) to -1.8 mV (nitrate). These data indicatedthat the t potential of T. ferrooxidans at pH 2.0 resulted froma complex combination of the ionization of surface moleculargroups and the preferential adsorption of anions. The depen-dence of the r potential on the concentration of the anion wasnot investigated. The t potential of the bacterium was arectangular hyperbolic function of the ferric concentrationregardless of the identity of the anion. Values of to, tmax and
TABLE 1. Electrokinetic properties of T. ferrooxidans
Growth Principal (mV) )max KFe(mMl)substrate anion (mV) (mM)
Soluble iron C104- -7.4 2.1 11S042- -6.0 3.1 12Cl- -4.7 4.1 10NO3- -1.8 2.2 8
Pyrite ClO4- -2.3 2.1 11S042- -1.4 2.9 11Cl- -1.3 3.7 9NO3- -1.7 3.0 13
Sulfur Cl04- -0.34 5.2 40S042- -1.1 6.5 36Cl- -0.8 5.7 54NO3- -0.9 7.7 45
KFe(III) obtained in thesummarized in Table 1.
presence of different anions are
The t potential of T. ferrooxidans was influenced by thegrowth history of the organism. Figure 5 shows the r potentialof T. ferrooxidans in 0.01 N perchloric acid as a function of theconcentration of ferric ions for bacteria that were cultured oncolloidal sulfur, pyrite, or ferrous ions. The experiments shownin Fig. 5 were also conducted in sulfate, chloride, and nitrate(primary data not shown). Values of to, ;mav and KFe(III)obtained in the presence of each of the four anions for bacteriagrown in the presence of each of three substrates are summa-rized in Table 1. The values of tm. and KFe(III) were relativelyconstant for ferrous iron- and pyrite-grown ceils regardless ofthe identity of the principal anion present. The higher values ofto in pyrite-grown cells relative to those in ferrous-grown cells
E
4._oNa
20-2-4
-6-8
0 5 10 15 20 25 30
Fe(III), mMFIG. 5. Effect of the growth history of T. ferrooxidans on its
electrokinetic properties in 0.01 N perchloric acid. The dependence ofthe zeta potential of T ferrooxidans on the concentration of ferric ionswas determined by using washed cells that were grown autotrophicallyon elemental sulfur (a), pyrite (b), and soluble ferrous ions (c). Eachdatum point and error bar represents the mean and standard devia-tion, respectively, for 12 determinations. The curves drawn through thedatum points were generated by assuming the applicability of equation5 and using the values of the relevant constants in Table 1. (Inset)Linear plot of the reciprocal of A4, as defined in the legend to Fig. 3,versus the reciprocal of the concentration of ferric ions. The ordinateintercept and slope of each line were determined by linear regressionanalysis.
VOL. 60, 1994
(5)
APPL. ENVIRON. MICROBIOL.
Mobility, pm-cm/V-sec-1 0 1
66,000
45,000 -36,000 -29,000 -24 000 -20,10014,200
1 2 3
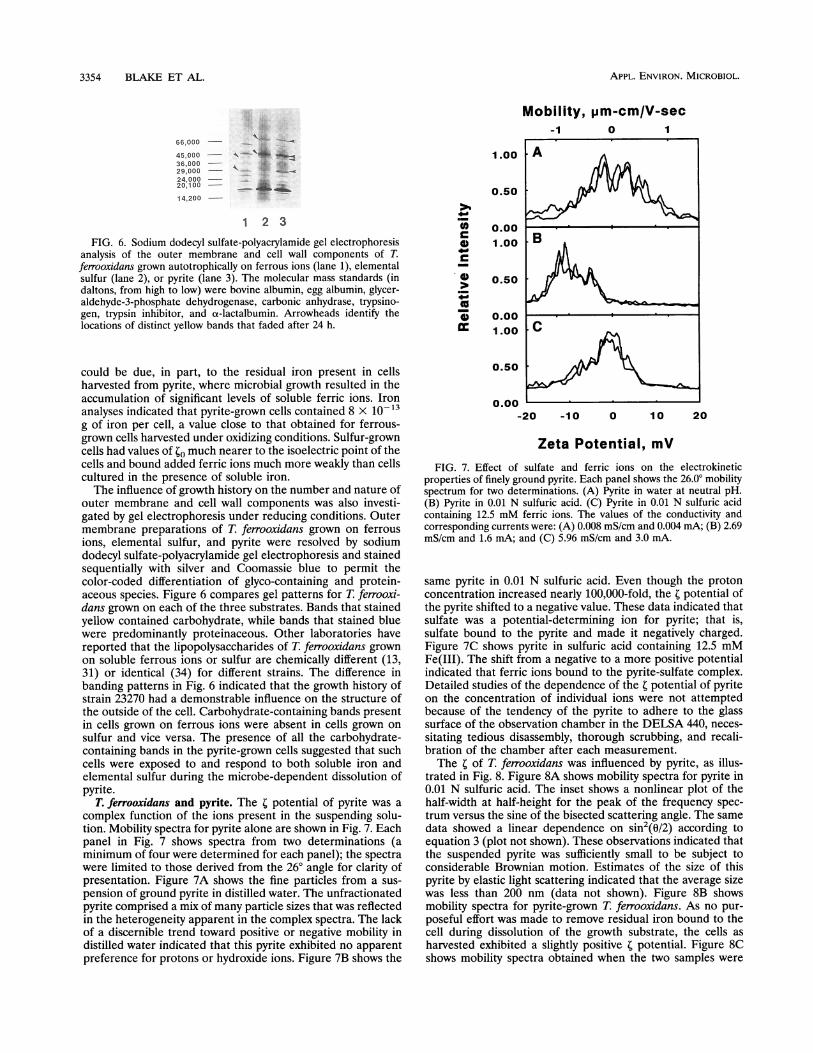
FIG. 6. Sodium dodecyl sulfate-polyacrylamide gel electrophoresisanalysis of the outer membrane and cell wall components of T.ferrooxidans grown autotrophically on ferrous ions (lane 1), elementalsulfur (lane 2), or pyrite (lane 3). The molecular mass standards (indaltons, from high to low) were bovine albumin, egg albumin, glycer-aldehyde-3-phosphate dehydrogenase, carbonic anhydrase, trypsino-gen, trypsin inhibitor, and ot-lactalbumin. Arrowheads identify thelocations of distinct yellow bands that faded after 24 h.
could be due, in part, to the residual iron present in cellsharvested from pyrite, where microbial growth resulted in theaccumulation of significant levels of soluble ferric ions. Ironanalyses indicated that pyrite-grown cells contained 8 x 10-13g of iron per cell, a value close to that obtained for ferrous-grown cells harvested under oxidizing conditions. Sulfur-growncells had values of O much nearer to the isoelectric point of thecells and bound added ferric ions much more weakly than cellscultured in the presence of soluble iron.The influence of growth history on the number and nature of
outer membrane and cell wall components was also investi-gated by gel electrophoresis under reducing conditions. Outermembrane preparations of T. ferrooxidans grown on ferrousions, elemental sulfur, and pyrite were resolved by sodiumdodecyl sulfate-polyacrylamide gel electrophoresis and stainedsequentially with silver and Coomassie blue to permit thecolor-coded differentiation of glyco-containing and protein-aceous species. Figure 6 compares gel patterns for T. ferrooxi-dans grown on each of the three substrates. Bands that stainedyellow contained carbohydrate, while bands that stained bluewere predominantly proteinaceous. Other laboratories havereported that the lipopolysaccharides of T. ferrooxidans grownon soluble ferrous ions or sulfur are chemically different (13,31) or identical (34) for different strains. The difference inbanding patterns in Fig. 6 indicated that the growth history ofstrain 23270 had a demonstrable influence on the structure ofthe outside of the cell. Carbohydrate-containing bands presentin cells grown on ferrous ions were absent in cells grown onsulfur and vice versa. The presence of all the carbohydrate-containing bands in the pyrite-grown cells suggested that suchcells were exposed to and respond to both soluble iron andelemental sulfur during the microbe-dependent dissolution ofpyrite.
T. ferrooxidans and pyrite. The C potential of pyrite was acomplex function of the ions present in the suspending solu-tion. Mobility spectra for pyrite alone are shown in Fig. 7. Eachpanel in Fig. 7 shows spectra from two determinations (aminimum of four were determined for each panel); the spectrawere limited to those derived from the 260 angle for clarity ofpresentation. Figure 7A shows the fine particles from a sus-pension of ground pyrite in distilled water. The unfractionatedpyrite comprised a mix of many particle sizes that was reflectedin the heterogeneity apparent in the complex spectra. The lackof a discernible trend toward positive or negative mobility indistilled water indicated that this pyrite exhibited no apparentpreference for protons or hydroxide ions. Figure 7B shows the
1.00
0.50
0.001.00
0.50
0.001.00
4)0
. )
co
0.50
0.00 1
-20 -10 0 10 20
Zeta Potential, mVFIG. 7. Effect of sulfate and ferric ions on the electrokinetic
properties of finely ground pyrite. Each panel shows the 26.0° mobilityspectrum for two determinations. (A) Pyrite in water at neutral pH.(B) Pyrite in 0.01 N sulfuric acid. (C) Pyrite in 0.01 N sulfuric acidcontaining 12.5 mM ferric ions. The values of the conductivity andcorresponding currents were: (A) 0.008 mS/cm and 0.004 mA; (B) 2.69mS/cm and 1.6 mA; and (C) 5.96 mS/cm and 3.0 mA.
same pyrite in 0.01 N sulfuric acid. Even though the protonconcentration increased nearly 100,000-fold, the C potential ofthe pyrite shifted to a negative value. These data indicated thatsulfate was a potential-determining ion for pyrite; that is,sulfate bound to the pyrite and made it negatively charged.Figure 7C shows pyrite in sulfuric acid containing 12.5 mMFe(III). The shift from a negative to a more positive potentialindicated that ferric ions bound to the pyrite-sulfate complex.Detailed studies of the dependence of the 4 potential of pyriteon the concentration of individual ions were not attemptedbecause of the tendency of the pyrite to adhere to the glasssurface of the observation chamber in the DELSA 440, neces-sitating tedious disassembly, thorough scrubbing, and recali-bration of the chamber after each measurement.The 4 of T. ferrooxidans was influenced by pyrite, as illus-
trated in Fig. 8. Figure 8A shows mobility spectra for pyrite in0.01 N sulfuric acid. The inset shows a nonlinear plot of thehalf-width at half-height for the peak of the frequency spec-trum versus the sine of the bisected scattering angle. The samedata showed a linear dependence on sin2(0/2) according toequation 3 (plot not shown). These observations indicated thatthe suspended pyrite was sufficiently small to be subject toconsiderable Brownian motion. Estimates of the size of thispyrite by elastic light scattering indicated that the average sizewas less than 200 nm (data not shown). Figure 8B showsmobility spectra for pyrite-grown T. ferrooxidans. As no pur-poseful effort was made to remove residual iron bound to thecell during dissolution of the growth substrate, the cells asharvested exhibited a slightly positive C potential. Figure 8Cshows mobility spectra obtained when the two samples were
3354 BLAKE ET AL.
ig,v*k4..
j

ELECTROPHORETIC MOBILITY OF T. FERROOXIDANS 3355
Mobility, pm-cm/V-sec Mobility, pm-cm/V-sec-4 -2 0
0 1
0.50
0.001 .00
>b
cSC
NWS'U
0.50
0.001 .00
0.50
0.001 .00
0.50
0.001 .00
0.50
0.00 --60 -40 -20 0 20
Zeta Potential, mVI** FIG. 9. Effect of colloidal sulfur on the electrokinetic properties of
-10 0 10 20 sulfur-grown T. ferrooxidans. Each panel shows the 26.00 mobilityspectrum for two determinations in 0.01 N sulfuric acid. The conduc-tivity in samples containing colloidal sulfur was 5.21 mS/cm; the
Zeta Potential, mV current was 2.0 mA in all experiments. (A) Colloidal sulfur alone; (B)T. ferrooxidans alone; (C through E) T ferrooxidans plus colloidalFIG. 8. Effect of pyrite on the electrokinetic properties of pyrite-
grown T. ferrooxidans. Each panel shows the 26.00 mobility spectrumfor two determinations in 0.01 N sulfuric acid. The current was 2.0 mAin all experiments. (A) Pyrite alone. Inset, nonlinear dependence ofthe half-width at half-height for the peak of the frequency spectrum onthe sine of the bisected scattering angle. (B) T. ferrooxidans alone. (C)T. ferrooxidans plus pyrite 5 min after mixing. Inset, linear dependenceof the half-width at half-height for the peak of the frequency spectrumon the sine of the bisected scattering angle. Each datum point anderror bar in the insets represent the mean and standard deviation,respectively, for 18 determinations.
mixed and incubated for 5 min. The pyrite- and cell-dependentmobility spectra disappeared and were replaced with new,broad mobility spectra that corresponded to a colloidal particlewith an electrophoretic mobility intermediate between those ofthe free components. These new spectra exhibited the heter-ogeneity of the free pyrite but the size of the free bacteria, asindicated by the angular dependence shown in the inset of Fig.8C. This intermediate-mobility particle thus exhibited theproperties anticipated for a complex between the bacterialcells and the particulate pyrite.
T. ferrooxidans and sulfur. The 4 potential of T. ferrooxidanswas influenced by colloidal sulfur. Mobility spectra for theinteraction between T. ferrooxidans and colloidal sulfur arepresented in Fig. 9. Figure 9A shows mobility spectra forcolloidal sulfur alone in 0.01 N sulfuric acid. The average 4potential of -40 mV indicated that there was significantcharge repulsion among the individual sulfur particles, contrib-uting to the stability of the colloidal suspension. Figure 9Bshows mobility spectra for sulfur-grown T. ferrooxidans. Cells
sulfur at 5 min (C), 10 min (D), and 20 min (E) after mixing.
cultured autotrophically on elemental sulfur were slightlynegative. Figures 9C through E show mobility spectra obtainedwhen the two samples were mixed and incubated for 5, 10, and20 min, respectively. The mobility spectra of colloidal sulfurgradually diminished, while those of T ferrooxidans becameprogressively more negative. After 20 min, only one spectrumremained, with an electrophoretic mobility intermediate be-tween those of the free starting components. Once again, thisintermediate-mobility particle exhibited the properties antici-pated for a complex between the bacterial cells and thecolloidal sulfur.
T. ferrooxidans double layer. The long-range forces betweenparticles in solution are an important subject in colloid andsurface chemistry. Particles in suspension experience both vander Waals' forces and electrostatic interactions. The van derWaals' interaction is attractive until the particles are suffi-ciently close for overlap of their atomic orbitals to createsubstantial repulsive forces. The electrostatic interaction isusually repulsive because both bacteria and surfaces are pre-dominantly negatively charged in nature. The balance betweenthese attractive and repulsive forces determines the ease andhence the rate with which the particles can approach suffi-ciently closely for possible short-range forces (hydrophobicand hydrogen bonds, etc.) to contribute to adherence.The adsorptive interactions between particles of the same
charge have frequently been considered in terms of the Der-jaguin-Landau-Verwey-Overbeck (DLVO) theory of colloidstability (24). Although the DLVO theory has been applied to
-1
1.00
0.50
>._
0
.0
00
0.001.00
0.50
0.001.00
0.50s
0.00 L-20
VOL. 60, 1994
0-

APPL. ENVIRON. MICROBIOL.
studies of viral, bacterial, and eukaryotic cell adhesion (5, 14,17, 29, 30), numerous complications arise because cells are notnonpermeable objects of well-defined structure and surfacetopography. One must know the value of the surface potentialto calculate the electrostatic repulsion forces. The surfacepotential is usually assumed to be equal to the t potential forcolloidal systems in which t is less than 50 mV (11), but withcells, such an assumption is doubtful. The fixed ionogenicgroups that are distributed at different levels throughout thegel-like structure of outer layers of the cell all contribute to thetotal effective surface charge. Ionic groups far from the shearboundary may not possess counterions that move with the cellas a unit. Cells may not be uniformly charged on their surfacebut may possess regions of higher and lower charge density.Also, the transformation of electrophoretic mobility to tpotential by the Smoluchowski equation is strictly applicableonly to impenetrable spherical particles. The more penetrableis a particle by counterions, the more inaccurate is the calcu-lation of t from ,u (12). These and other factors make theDLVO theory at best a qualitative guide for predicting celldeposition onto surfaces.Washed cells of T. ferrooxidans grown in the presence of
reduced iron were negatively charged. The thickness of thediffuse layer of counterions (protons in the absence of addedferric ions) around T ferrooxidans was estimated from therelationship
1/K = [(4re21/ckT) E CiZ2 I
where e is the charge on an electron, k is Boltzmann's constant,T is absolute temperature, and c and z are the concentrationand charge, respectively, of the ith ion. The value of 1I/K,technically the distance from the shear boundary where thepotential has reached 1/exp of the t potential, was about 30 A(ca. 3 nm) for iron-grown T. ferrooxidans in sulfate at pH 2.0.Calculations of charge density in the diffuse layer by integrat-ing curves such as those shown in Fig. 1C gave an excess of 1.0x 10-12 mol of protons in a column of solution of 1 cm2 crosssection and a deficiency of 0.6 x 10-12 mol of anions for a tpotential of -7 mV. There was, correspondingly, a compen-sating negative surface charge of 1.6 X 10-12 mol of electriccharge per cm2. Similar diffuse layers of even greater chargedensity existed around particulate pyrite and sulfur under thesame conditions. The initial long-range force between T.ferrooxidans and its insoluble substrates in the absence of ironwould thus be repulsive.The addition of Fe(III) to the Stern layer neutralized the net
charge on the surface of T. ferrooxidans and removed anydouble-layer repulsive barrier to the cell's interaction withnegatively (or positively) charged particles. High concentra-tions of iron (7 to 200 mM) are typical of acid mine drainageand mining environments (10, 32). The net charge on T.ferrooxidans under physiological conditions is thus neutral orslightly positive. Indeed, the t potential before washing ofbacteria harvested from pyrite or oxidized soluble iron waszero or slightly positive. Similarly, when T. ferrooxidans wasgrown on elemental sulfur, the structure of the bacterium wasaltered so as to impart a net charge of about zero on thesurface. T. ferrooxidans thus appears to regulate the net chargeon its surface to minimize double-layer repulsive forces with itsparticulate, charged substrates.The binding of T. ferrooxidans with pyrite and sulfur was
observed instrumentally. Since the immediate output of theseDoppler velocimetry experiments is relative, not absolute,light-scattering intensities, the binding results must be inter-preted qualitatively. That is, one cannot use this method to
quantitatively characterize the binding of T. ferrooxidans to itsparticulate substrates in terms of dissociation constants andbinding stoichiometries. One can only conclude that binding isevident. Other investigators have studied the sorption betweenbacterial cells and clay particles by changes in particle size anddistribution using an electrical impedance method commonlyemployed in particle characterization (25, 26). It is anticipatedthat these and other physical measurements used to character-ize colloidal systems may be exploited to quantify the physico-chemical characteristics of the binding of T. ferrooxidans andrelated organisms to minerals. Once quantitative assays for thebinding of bacteria to mineral surfaces have been developed,fundamental questions concerning the interaction can be ad-dressed.
ACKNOWLEDGMENTSWe thank Stephen McGinness for preliminary experiments on the
DELSA 440.This research was supported by grants DE-FG05-85ER13339 and
DE-FG05-92ER20087 from the United States Department of Energy.
REFERENCES1. Bennet, J. C., and H. Tributsch. 1978. Bacterial leaching patterns
on pyrite crystal surfaces. J. Bacteriol. 134:310-317.2. Berry, V. K., and L. E. Murr. 1978. Direct observations of bacteria
and quantitative studies of their catalytic role in the leaching oflow-grade, copper-bearing waste, p. 103-136. In L. E. Murr, A.Torma, and J. A. Brierley (ed.), Metallurgical applications ofbacterial leaching and related microbiological phenomena. Aca-demic Press, New York.
3. Blake, R. C., II, G. T. Howard, and S. McGinness. Enhanced yieldsof iron-oxidizing bacteria by in situ electrochemical reduction ofsoluble iron in the growth medium. AppI. Environ. Microbiol.60:2704-2710.
4. Clark, J. M., Jr., and R. L. Switzer. 1977. Transport studies withbacterial membrane vesicle preparations, p. 195-198. In Experi-mental biochemistry, 2nd ed. W. H. Freeman and Co., NewYork.
5. Collins, Y. E., and G. Stotzky. 1992. Heavy metals alter theelectrokinetic properties of bacteria, yeasts, and clay minerals.Appl. Environ. Microbiol. 58:1592-1600.
6. Devasia, P., K. A. Natarajan, D. N. Sathyanarayana, and G.Ramananda Rao. 1993. Surface chemistry of Thiobacillusferrooxi-dans relevant to adhesion on mineral surfaces. AppI. Environ.Microbiol. 59:4051-4055.
7. DiSpirito, A. A., P. R. Dugan, and 0. H. Tuovinen. 1983. Sorptionof Thiobacillus ferrooxidans to particulate material. Biotechnol.Bioeng. 25:1163-1168.
8. Dzandu, J. K., M. H. Deh, D. L. Barratt, and G. E. Wise. 1984.Detection of erythrocyte membrane proteins, sialoglycoproteins,and lipids in the same polyacrylamide gel using a double-stainingtechnique. Proc. Natl. Acad. Sci. USA 81:1733-1737.
9. Espejo, R. T., and P. Romero. 1987. Growth of Thiobacillusferrooxidans on elemental sulfur. Appl. Environ. Microbiol. 53:1907-1912.
10. Filipek, L. H., D. K. Nordstrom, and W. H. Ficklin. 1987.Interaction of acid mine drainage with waters and sediments ofWest Squaw Creek in the West Shasta mining district, California.Environ. Sci. Technol. 21:388-396.
11. Haydon, D. A. 1960. Adsorption at the oil-water interface and thecalculation of electrical potentials in the aqueous phase. I. Neutralmolecules and a simplified treatment of ions. Phil. Trans. R. Soc.London A252:225-248.
12. Haydon, D. A. 1964. The electrical double layer and electrokineticphenomena, p. 94-158. In Recent progress in surface science.Academic Press, New York.
13. Hirt, W. E., and J. R. Vestal. 1975. Physical and chemical studiesof Thiobacillus ferrooxidans lipopolysaccharides. J. Bacteriol. 123:642-650.
14. James, A. M. 1991. Charge properties of microbial cell surfaces, p.223-262. In N. Mozes, P. S. Handley, H. J. Busscher, and P. G.
3356 BLAKE ET AL.

ELECTROPHORETIC MOBILITY OF T. FERROOXIDANS 3357
Rouxhet (ed.), Microbial cell surface analysis. VCH Publishers,Inc., New York.
15. Johnson, D. B., and S. McGinness. 1991. A highly efficient anduniversal solid medium for growing mesophilic and moderatelythermophilic, iron-oxidizing, acidophilic bacteria. J. Microbiol.Methods 13:113-122.
16. Lindstrom, E. B., E. Gunneriusson, and 0. H. Tuovinen. 1992.Bacterial oxidation of refractory sulfide ores for gold recovery.Crit. Rev. Biotechnol. 12:133-155.
17. Marshall, K. C. 1991. The importance of studying microbial cellsurfaces, p. 5-19. In N. Mozes, P. S. Handley, H. J. Busscher, andP. G. Rouxhet (ed.), Microbial cell surface analysis. VCH Publish-ers, Inc., New York.
18. McCready, R. G. L., and B. P. Le Gallais. 1984. Selectiveadsorption of 32P-labelled Thiobacillus ferrooxidans in finelyground coal suspensions. Hydrometallurgy 12:281-288.
19. Miller, D. M., and G. S. Hansford. 1992. Batch bio-oxidation of agold-bearing pyrite-arsenopyrite concentrate. Min. Eng. 5:613-629.
20. Murr, L. E., and V. K. Berry. 1976. Direct observation of selectiveattachment of bacteria on low-grade sulphide ores and othermineral surfaces. Hydrometallurgy 2:11-24.
21. Ohmura, N., K. Kitamura, and H. Saiki. 1993. Selective adhesionof Thiobacillus ferrooxidans to pyrite. Appl. Environ. Microbiol.59:4044-4050.
22. Rawlings, D. E., D. R. Woods, and N. P. Mjoli. 1991. The cloningand structure of genes from the autotrophic biomining bacteriumThiobacillus ferrooxidans, p. 215-237. In P. J. Greenaway (ed.),Advances in gene technology. JAI Press, Ltd., London.
23. Rodriguez-Leiva, M., and H. Tributsch. 1988. Morphology ofbacterial leaching patterns by Thiobacillus ferrooxidans on syn-thetic pyrite. Arch. Microbiol. 149:401-405.
24. Rutter, P., and B. Vincent. 1980. The adhesion of microorganismsto surfaces: physicochemical aspects, p. 79-92. In R. C. W.Berkeley, J. M. Lynch, J. Melling, P. R. Rutter, and B. Vincent(ed.), Microbial adhesion to surfaces. Ellis Horwood, Chichester,United Kingdom.
25. Santoro, T., and G. Stotzky. 1967. Effect of electrolyte compositionand pH and particle size distribution of microorganisms and clayminerals as determined by the electrical sensing zone method.Arch. Biochem. Biophys. 122:664-671.
26. Santoro, T., and G. Stotzky. 1968. Sorption between microorgan-isms and clay minerals as determined by the electrical sensing zoneparticle analyzer. Can. J. Microbiol. 14:299-307.
27. Sugrue, S., T. Oja, and S. Bott. 1989. Doppler electrophoretic lightscattering analysis using the Coulter DELSA 440. Int. Lab.19:34-45.
28. Tuovinen, 0. H., and D. P. Kelly. 1973. Studies on the growth ofThiobacillus ferrooxidans. I. Use of membrane filters and ferrousiron agar to determine viable numbers, and comparison with14C02-fixation and iron oxidation as measures of growth. Arch.Microbiol. 88:285-298.
29. Valentine, R. C., and A. Allison. 1960. Virus particle adsorption.III. Adsorption of viruses by cell polymers and effects of somevariables on adsorption. Biochim. Biophys. Acta 40:400-410.
30. van Loosdrecht, M. C. M., J. Lyklema, W. Norde, and A. J. B.Zehnder. 1990. Influence of interfaces on microbial activity.Microbiol. Rev. 54:75-87.
31. Vestal, J. R., D. G. Lundgren, and K. C. Milner. 1973. Toxic andimmunological differences among lipopolysaccharides from Thio-bacillus ferrooxidans grown autotrophically and heterotrophically.Can. J. Microbiol. 19:1335-1339.
32. Wakao, N., H. Tashibana, Y. Tanaka, Y. Sakurai, and N. Shiota.1985. Morphological and physiological characteristics of streamersin acid mine drainage water from a pyritic mine. J. Gen. Appl.Microbiol. 31:17-28.
33. Yeh, T. Y., J. R. Godshalk, G. J. Olson, and R. M. Kelly. 1987. Useof epifluorescence microscopy for characterizing the activity ofThiobacillus ferrooxidans on iron pyrite. Biotechnol. Bioeng. 30:138-146.
34. Yokota, A., Y. Yamada, and K. Imai. 1988. Lipopolysaccharides ofiron-oxidizing Leptospirillum ferrooxidans and Thiobacillus fer-rooxidans. J. Gen. Appl. Microbiol. 34:27-37.
VOL. 60, 1994




















