
Hepatic resection and transplantation for peripheral cholangiocarcinoma

Static lung compliance and body pressures in Tupinambismerianae with and without post-hepatic septum
Wilfried Klein a,*, Augusto S. Abe b, Steven F. Perry a
a Institut fur Zoologie, Universitat Bonn, Poppelsdorfer Schloß, 53115 Bonn, Germanyb Departamento de Zoologia, Universidade Estadual Paulista-Rio Claro, c.p. 199, 13506-900 Rio Claro, SP, Brazil
Accepted 14 February 2003
Abstract
The surgical removal of the post-hepatic septum (PHS) in the tegu lizard, Tupinambis merianae , significantly reduces
resting lung volume (VLr) and maximal lung volume (VLm) when compared with tegus with intact PHS. Standardised
for body mass (MB), static lung compliance was significantly less in tegus without PHS. Pleural and abdominal
pressures followed, like ventilation, a biphasic pattern. In general, pressures increased during expiration and decreased
during inspiration. However, during expiration pressure changes showed a marked intra- and interindividual variation.
The removal of the PHS resulted in a lower cranio-caudal intracoelomic pressure differential, but had no effect on the
general pattern of pressure changes accompanying ventilation. These results show that a perforated PHS that lacks
striated muscle has significant influence on static breathing mechanics in Tupinambis and by analogy provides valuable
insight into similar processes that led to the evolution of the mammalian diaphragm.
# 2003 Elsevier Science B.V. All rights reserved.
Keywords: Mechanics of breathing; compliance; ventilation; post-hepatic septum; Reptiles; tegu lizard (Tupinambis merianae );
Ventilation; reptile
1. Introduction
The lungs of the tegu lizard, Tupinambis mer-
ianae , are single-chambered and their respiratory
surface is made up of homogeneously distributed
faveoli (Perry, 1983). The lungs and the liver are
enclosed in a pleuro-hepatic cavity and are sepa-
rated from the remaining viscera by a post-hepatic
septum (PHS), a membranous structure containing
bundles of smooth muscle (Broman, 1904; Hoch-
stetter, 1906; Klein et al., in press). The PHS is
highly developed in the teiid genera Tupinambis
and Crocodilurus (Klein et al., 2000). It is trans-
versely oriented in the body cavity and, except for
a persistent dorso-lateral opening on the left side,
resembles the mammalian diaphragm. Recently
(Klein et al., in press), it was shown that the
removal of the PHS has profound effects on the
topology of the viscera: in tegus with a surgically
ruptured PHS, intestine and stomach are displaced
cranially, thereby reducing the space for the lungs.
* Corresponding author. Tel.: �/49-228-735489; fax: �/49-
228-735458.
E-mail address: [email protected] (W. Klein).
Respiratory Physiology & Neurobiology 135 (2003) 73�/86
www.elsevier.com/locate/resphysiol
1569-9048/03/$ - see front matter # 2003 Elsevier Science B.V. All rights reserved.
doi:10.1016/S1569-9048(03)00063-6

The effects of the PHS on the mechanics ofbreathing are unknown.
A measure of the mechanical properties of lungs
is its compliance (distensibility), which is defined
as the change in intrapulmonary volume for a
given intrapulmonary pressure. Surface tension
and elastic properties of the lung tissue are the
main determinants of lung compliance. In the tegu
lung, with its homogeneously distributed anddense parenchyma, a high work of breathing is
necessary to inflate the low-compliance lungs
(Perry and Duncker, 1980). Such a small, par-
enchyma-rich single-chambered lung is considered
an evolutionary dead end with respect to gas
exchange, as the space for elaboration of parench-
ymal gas exchange surfaces is limited to the lung
wall and because the deep faveolar parenchyma isnot accessible to convective air movement (Perry
and Duncker, 1980). Therefore, the PHS may have
evolved as an accessory structure in conjunction
with supporting inflation of the stiff lungs, thereby
partially overcoming the structural limitations set
by the single-chambered lungs.
In mammals, similar functional pressures may
have lead to the evolution of their unique lung�/
diaphragm complex (Perry and Duncker, 1978,
1980). Mammalian lungs show the lowest compli-
ance of all vertebrates and a high work of breath-
ing is necessary to ventilate them (Crosfill and
Widdicombe, 1961; Perry and Duncker, 1980). If
the pleural cavity were opened, the lungs would
collapse completely. In intact mammals an oppos-
ing negative pressure is applied in the pleural spacepreventing the lungs from collapsing (Radford,
1964) while at the same time allowing their free
movement within the closed pleural cavity. There-
fore an undivided pleuro-peritoneal body cavity,
as can be found in lizards, is inappropriate for
mammals and consequently, the evolution of the
mammalian lung seems impossible in the absence
of a complete diaphragm. A functional model forthe stepwise evolution of the diaphragm from an
incomplete septum therefore appears implausible.
The PHS of the tegu could represent such a model.
Breathing mechanics was studied in Tupinambis
as part of comparative studies (Perry and
Duncker, 1978, 1980), but these authors examined
mechanical properties only in the closed chest
preparation. These studies, however, reported thelowest values of total compliance (CT) found in a
lizard. A surgical rupture of the PHS not only may
affect lung volumes simply as a result of the
reduced pleuro-hepatic space (Klein et al., in
press), but also could alter the amount and
distribution of elastic fibres in the lung itself,
either stiffening the lungs against the pressure of
the viscera or balancing the lungs at a smallerresting volume. Such a change may be reflected in
decreased lung compliance.
During aspiration breathing, rib movement
generates positive pressure during expiration and
negative pressure during inspiration. In an undi-
vided body cavity, these pressures should apply
equally to all regions. Gans (1970) postulated that
the forces produced by respiratory muscles duringbreathing act directly on the visceral organs and
that these forces are transmitted from one com-
partment to another by the interaction of local
forces between the compartments (linkage) rather
than by hydrostatic forces alone. Therefore the
pressures in the visceral part of an undivided
pleuro-peritoneal cavity could be different from
those found in the pulmonary part (Templeton,1964).
In the present study we test the hypothesis that
the transverse division of the body cavity by the
PHS in Tupinambis is important for lung ventila-
tion and that in spite of the left-side foramen a
large transseptal pressure difference is created,
thereby facilitating breathing.
2. Materials and methods
2.1. Breathing mechanics
2.1.1. Animals and experimental design
Investigations of static breathing mechanics
were performed on neonatal tegus (N�/8; MB:
17.69/5.3 g; SVL: 87.49/7.6 mm), juvenile tegus(N�/5; MB: 120.39/27.6 g; SVL: 160.29/11.0 mm),
sham-operated (SO) adult tegus (N�/6; MB:
707.09/422.5 g; SVL: 267.59/44.3 mm) and adult
tegus maintained with removed PHS (SR-tegus)
for 11 months (N�/8; MB: 1193.89/263.0 g; SVL:
316.49/20.0 mm). The removal of the PHS has
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/8674

been described in detail elsewhere (Klein et al., inpress). All tegus were killed by intra-peritoneal
injection of 60 mg kg�1 pentobarbital sodium
(SagatalTM), which causes analgesia and cessation
of spontaneous breathing within 15�/20 min, while
the heart continues beating for hours. Experiments
were performed at room temperature (�/28 8C).
In the supine animal the trachea was intubated
to obtain volume�/pressure relationships. Thistube was connected to a three-way stopcock
from which one connection led to a plastic syringe
and the other, to a water-filled tube (PE 60)
connected to the DELTRAN pressure transducer
of a strain gauge. The dead space of all tubing
combined was less than 2 ml. The signal of the
pressure transducer was amplified (1000�/) by a
custom-made pre-amplifier and then sent to ananalogue-to-digital (A�/D) converter and recorded
through a computerised data-acquisition system
(DAC, Sable System). To calibrate the pressure
transducer, the water column was moved in a
stepwise fashion in a range between �/30 and �/30
cm and the regression line derived from this
calibration was used to obtain a regression line
for the V�/P relationships. The experimental pro-tocol followed closely that of Perry and Duncker
(1978). Briefly, resting lung volume (VLr) was
defined as the amount of air that can be removed
from the lungs in a freshly killed animal until a
negative pressure of �/10 cmH2O was reached.
Lung volumes were determined by adding air in
increments to the lungs of an animal with closed
body cavity, starting at a pressure of �/10 cmH2Oand increasing to �/20 cmH2O. The total amount
of air necessary to fill the lungs from the state at
greatest negative pressure until the V�/P diagram
passed its upper inflection point was defined as
maximal lung volume (VLm). To stabilise the V�/P
diagrams (Bernstein, 1957), the lungs were inflated
and deflated several times (usually four) over the
whole pressure range before measurements com-menced. After each determination of VLm, the
system was opened to the atmosphere and lung
volume was allowed to return spontaneously to
VLr. V�/P diagrams were obtained by (1) stepwise
removal of air until a pressure of �/10 cmH2O was
reached, (2) adding of air until pressure reached �/
20 cmH2O, and (3) returning stepwise to VLr.
Pressure was allowed to stabilise for 15�/30 secbefore taking the reading. After each V�/P dia-
gram, air was added until VLm was reached and
the system was opened to return to VLr again. V�/P
diagrams were first made for the animals with
intact body cavity, and then for the in situ lungs
after cutting the mesopneumonia, and widely
opening or resecting the body wall. The lungs
were moistened with saline (0.9% NaCl) through-out. The mean values of three V�/P diagrams were
taken for analysis.
2.1.2. Data handling and statistics
Compliance (C) was defined as the slope (b) of
the straight portion of the inflation curve and
obtained by fitting a line (y�/a�/ bx) through the
points given by the V�/P diagram. Compliance ofthe lungs (CL) and total compliance (CT) were
determined from V�/P diagrams where lung vo-
lume was expressed in ml (non-standardised), as
well as after standardisation of lung volume
against MB, VLr and VLm. Body wall compliance
was defined as 1/CB�/1/CT�/1/CL. The index of
hysteresis (H) was defined as the length of a
perpendicular line connecting the midpoints ofthe inflation curve and the deflation curve, and
was calculated by the Pythagorean method (for
details see Perry and Duncker, 1978).
ANOVA was used examine significant differ-
ences between the groups, followed by an all
pairwise multiple comparison procedure (Stu-
dent-Newman�/Keuls Method) to determine which
groups differ significantly from each other. Adifference was considered significant at P B/0.05.
2.2. Pressure measurements
2.2.1. Animals, surgery and experimental protocol
Four tegus without PHS (MB: 1015.259/193.61
g) and four tegus with intact PHS (MB: 1150.09/
298.86 g) were used in this study.
After exposing the animals to CO2 until theydisplayed no reaction to handling or pinching, the
tegus were placed in a prone position on an
operation table and a constant low CO2-flow was
provided to their nostrils to maintain anaesthesia.
The abdomen was disinfected with alcohol and
with an injection needle (¥ 2 mm) a piece of PE
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/86 75
tubing (Portex, 800/110/240/100) was inserted on
the right dorso-lateral side into the pleural cavity
at the level of the 3th�/5th rib and into the
abdominal cavity 2�/3 cm anterior to the pelvic
girdle. Each tube extended 4�/6 cm into the body
cavity. To prevent blocking of the opening by the
viscera two additional holes were made opposing
each other near the tip of the tube and the tubes
were flushed from time to time with saline (0.9%
NaCl). Pressures were measured using two DEL-
TRAN pressure transducers from the strain gauge
type, each connected to a custom-made pre-
amplifier (amplifying voltage 1000�/).After surgery, which was carried out before
09:00, a mask for ventilation measurements was
put over the tegus head and the animal was placed
into a climatic chamber at 25 or 35 8C to recover.
The mask was connected by tygon tubing to a
pressure transducer (Sable PT-100) for ventilatory
recordings. The signals from pressure transducer
and pre-amplifiers were sent to an A�/D converter
and recorded through a computerised data-acqui-
sition system (DAC, Sable System).
Pressures and spontaneous ventilation data were
measured between 17:00 and 20:00. When these
measurements were completed, the temperature
was increased or decreased and pressures together
with ventilation were measured under the new
conditions the following day between 17:00 and
20:00.
2.2.2. Data handling
To analyse the data, a time was chosen when
tegus appeared to be resting quietly, and trans-
mural pleural and abdominal pressures were
determined: the baseline before each breathing
bout was taken and the change from the baseline
to the maximal or minimal peak was taken as
pressure created during expiration and inspiration,
respectively. Transseptal pressure was obtained bysubtraction of abdominal pressure from pleural
pressure.
3. Results
3.1. Lung volumes
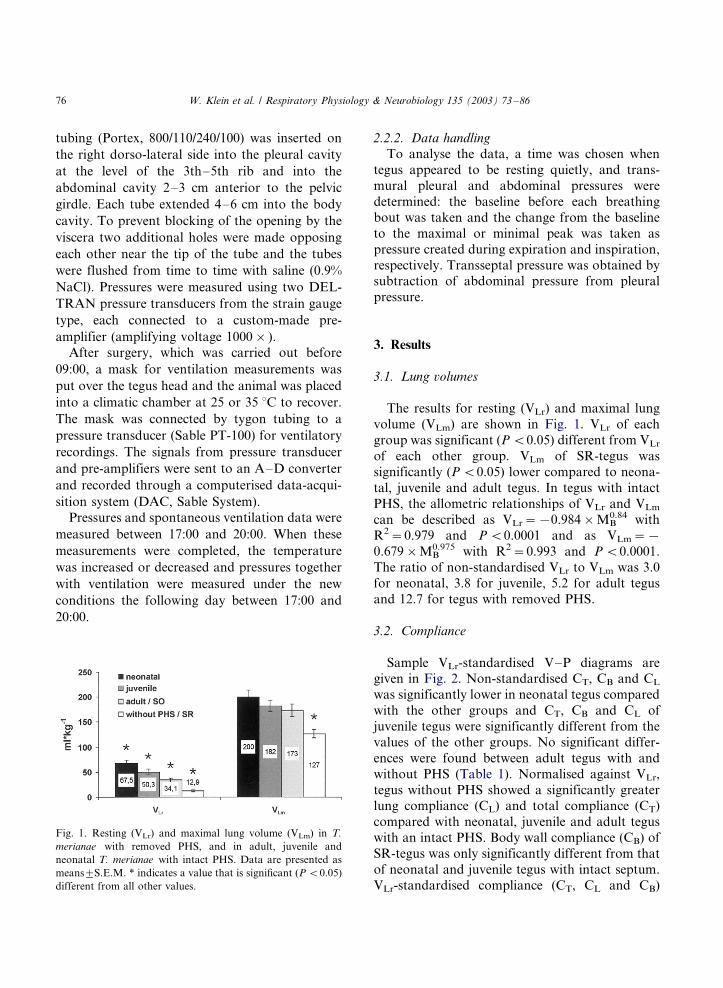
The results for resting (VLr) and maximal lung
volume (VLm) are shown in Fig. 1. VLr of each
group was significant (P B/0.05) different from VLr
of each other group. VLm of SR-tegus was
significantly (P B/0.05) lower compared to neona-
tal, juvenile and adult tegus. In tegus with intact
PHS, the allometric relationships of VLr and VLm
can be described as VLr�/�/0.984�/MB0.84 with
R2�/0.979 and P B/0.0001 and as VLm�/�/
0.679�/MB0.975 with R2�/0.993 and P B/0.0001.
The ratio of non-standardised VLr to VLm was 3.0
for neonatal, 3.8 for juvenile, 5.2 for adult tegus
and 12.7 for tegus with removed PHS.
3.2. Compliance
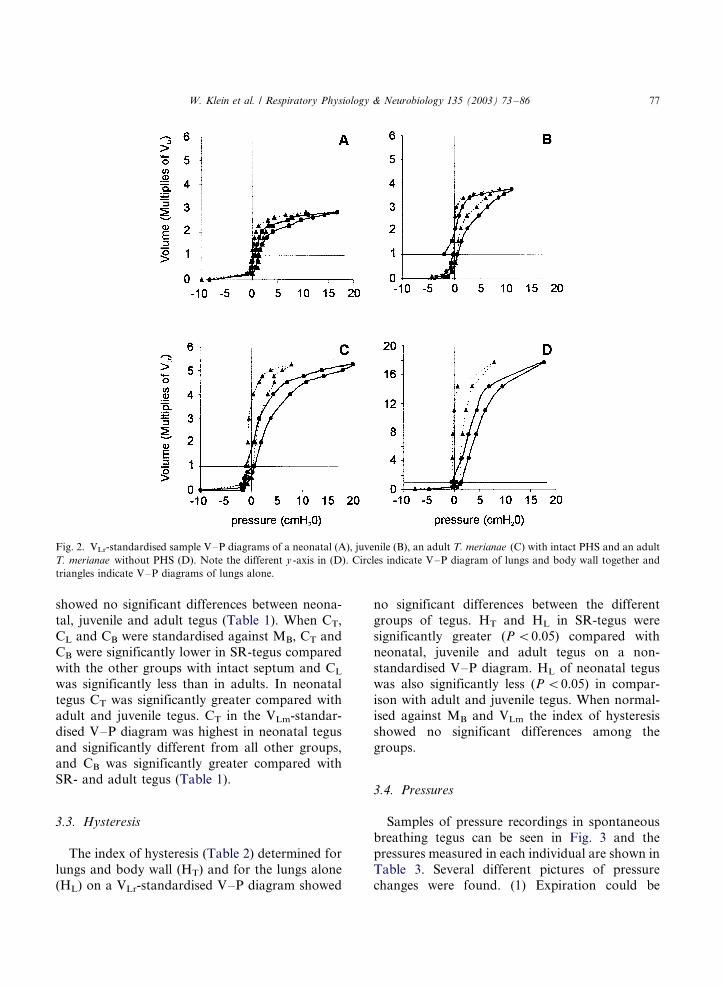
Sample VLr-standardised V�/P diagrams aregiven in Fig. 2. Non-standardised CT, CB and CL
was significantly lower in neonatal tegus compared
with the other groups and CT, CB and CL of
juvenile tegus were significantly different from the
values of the other groups. No significant differ-
ences were found between adult tegus with and
without PHS (Table 1). Normalised against VLr,
tegus without PHS showed a significantly greaterlung compliance (CL) and total compliance (CT)
compared with neonatal, juvenile and adult tegus
with an intact PHS. Body wall compliance (CB) of
SR-tegus was only significantly different from that
of neonatal and juvenile tegus with intact septum.
VLr-standardised compliance (CT, CL and CB)
Fig. 1. Resting (VLr) and maximal lung volume (VLm) in T.
merianae with removed PHS, and in adult, juvenile and
neonatal T. merianae with intact PHS. Data are presented as
means9/S.E.M. * indicates a value that is significant (P B/0.05)
different from all other values.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/8676
showed no significant differences between neona-
tal, juvenile and adult tegus (Table 1). When CT,
CL and CB were standardised against MB, CT and
CB were significantly lower in SR-tegus compared
with the other groups with intact septum and CL
was significantly less than in adults. In neonatal
tegus CT was significantly greater compared withadult and juvenile tegus. CT in the VLm-standar-
dised V�/P diagram was highest in neonatal tegus
and significantly different from all other groups,
and CB was significantly greater compared with
SR- and adult tegus (Table 1).
3.3. Hysteresis
The index of hysteresis (Table 2) determined for
lungs and body wall (HT) and for the lungs alone
(HL) on a VLr-standardised V�/P diagram showed
no significant differences between the different
groups of tegus. HT and HL in SR-tegus were
significantly greater (P B/0.05) compared with
neonatal, juvenile and adult tegus on a non-
standardised V�/P diagram. HL of neonatal tegus
was also significantly less (P B/0.05) in compar-
ison with adult and juvenile tegus. When normal-ised against MB and VLm the index of hysteresis
showed no significant differences among the
groups.
3.4. Pressures
Samples of pressure recordings in spontaneous
breathing tegus can be seen in Fig. 3 and the
pressures measured in each individual are shown in
Table 3. Several different pictures of pressure
changes were found. (1) Expiration could be
Fig. 2. VLr-standardised sample V�/P diagrams of a neonatal (A), juvenile (B), an adult T. merianae (C) with intact PHS and an adult
T. merianae without PHS (D). Note the different y -axis in (D). Circles indicate V�/P diagram of lungs and body wall together and
triangles indicate V�/P diagrams of lungs alone.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/86 77
accompanied by a large increase in pressure in the
abdominal and pleural part of the body cavity, (2)
only an increase in pressure in the pleural cavity or
(3) no marked change in pressure at all. (4) In
some cases the pleural pressure decreased at the
beginning of an expiration, followed by a slight
increase at the end of the expiration. During
inspiration, on the other hand, a large decrease
in pressures was always found in the pleural as well
as in the abdominal part of the body cavity.The recorded pressures were similar at 25 and
35 8C, and only SO-tegus showed a larger decrease
in pressure during inspiration at 35 8C compared
with 25 8C. Tegus with a surgically ruptured PHS
showed some differences compared with those
having an intact PHS. Those differences were
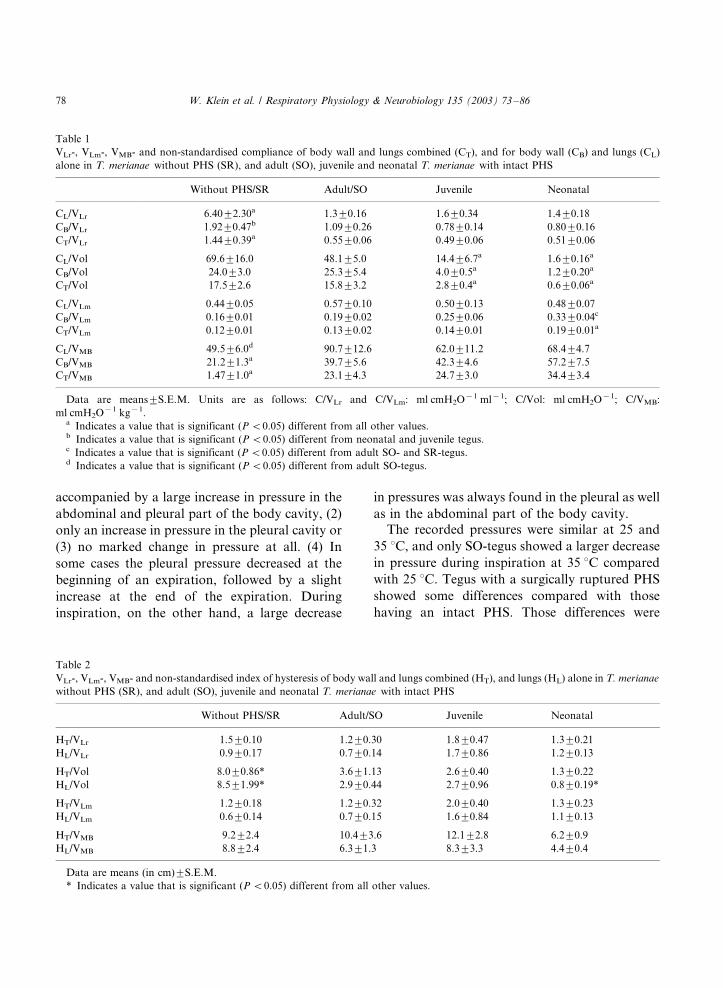
Table 1
VLr-, VLm-, VMB- and non-standardised compliance of body wall and lungs combined (CT), and for body wall (CB) and lungs (CL)
alone in T. merianae without PHS (SR), and adult (SO), juvenile and neonatal T. merianae with intact PHS
Without PHS/SR Adult/SO Juvenile Neonatal
CL/VLr 6.409/2.30a 1.39/0.16 1.69/0.34 1.49/0.18
CB/VLr 1.929/0.47b 1.099/0.26 0.789/0.14 0.809/0.16
CT/VLr 1.449/0.39a 0.559/0.06 0.499/0.06 0.519/0.06
CL/Vol 69.69/16.0 48.19/5.0 14.49/6.7a 1.69/0.16a
CB/Vol 24.09/3.0 25.39/5.4 4.09/0.5a 1.29/0.20a
CT/Vol 17.59/2.6 15.89/3.2 2.89/0.4a 0.69/0.06a
CL/VLm 0.449/0.05 0.579/0.10 0.509/0.13 0.489/0.07
CB/VLm 0.169/0.01 0.199/0.02 0.259/0.06 0.339/0.04c
CT/VLm 0.129/0.01 0.139/0.02 0.149/0.01 0.199/0.01a
CL/VMB 49.59/6.0d 90.79/12.6 62.09/11.2 68.49/4.7
CB/VMB 21.29/1.3a 39.79/5.6 42.39/4.6 57.29/7.5
CT/VMB 1.479/1.0a 23.19/4.3 24.79/3.0 34.49/3.4
Data are means9/S.E.M. Units are as follows: C/VLr and C/VLm: ml cmH2O�1 ml�1; C/Vol: ml cmH2O�1; C/VMB:
ml cmH2O�1 kg�1.a Indicates a value that is significant (P B/0.05) different from all other values.b Indicates a value that is significant (P B/0.05) different from neonatal and juvenile tegus.c Indicates a value that is significant (P B/0.05) different from adult SO- and SR-tegus.d Indicates a value that is significant (P B/0.05) different from adult SO-tegus.
Table 2
VLr-, VLm-, VMB- and non-standardised index of hysteresis of body wall and lungs combined (HT), and lungs (HL) alone in T. merianae
without PHS (SR), and adult (SO), juvenile and neonatal T. merianae with intact PHS
Without PHS/SR Adult/SO Juvenile Neonatal
HT/VLr 1.59/0.10 1.29/0.30 1.89/0.47 1.39/0.21
HL/VLr 0.99/0.17 0.79/0.14 1.79/0.86 1.29/0.13
HT/Vol 8.09/0.86* 3.69/1.13 2.69/0.40 1.39/0.22
HL/Vol 8.59/1.99* 2.99/0.44 2.79/0.96 0.89/0.19*
HT/VLm 1.29/0.18 1.29/0.32 2.09/0.40 1.39/0.23
HL/VLm 0.69/0.14 0.79/0.15 1.69/0.84 1.19/0.13
HT/VMB 9.29/2.4 10.49/3.6 12.19/2.8 6.29/0.9
HL/VMB 8.89/2.4 6.39/1.3 8.39/3.3 4.49/0.4
Data are means (in cm)9/S.E.M.
* Indicates a value that is significant (P B/0.05) different from all other values.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/8678

Fig. 3. Sample recordings of ventilation (upper trace), abdominal pressure (middle trace) and pleural pressure (lower trace) at 25 8C(A) and 35 8C (B�/C) of T. merianae with intact PHS. B and C show the variation in the pressure changes accompanying expiration.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/86 79
more pronounced at 35 than at 25 8C. During
expiration at 35 8C, for example, the increases in
pressures in the pleural and the abdominal part of
the body cavity were greater in SR- than in SO-
tegus. During inspiration at 35 8C, however,
pressure changes in the pleural cavity were smaller
and pressure changes in the abdominal cavity were
larger in SR-tegus than in SO-tegus. The resulting
intracoelomic pressure gradient was about 2
cmH2O greater in SO- than in SR-tegus (Table 3).
4. Discussion
4.1. Lung volumes
Perry and Duncker (1978) reported for similar
sized Tupinambis teguixin (earlier T . nigropuncta-
tus , Avila-Pires, 1995) a resting lung volume of 43
ml kg�1 and a maximum lung volume of 151
ml kg�1. The slightly lower values found in this
study on T . merianae may be due to differences in
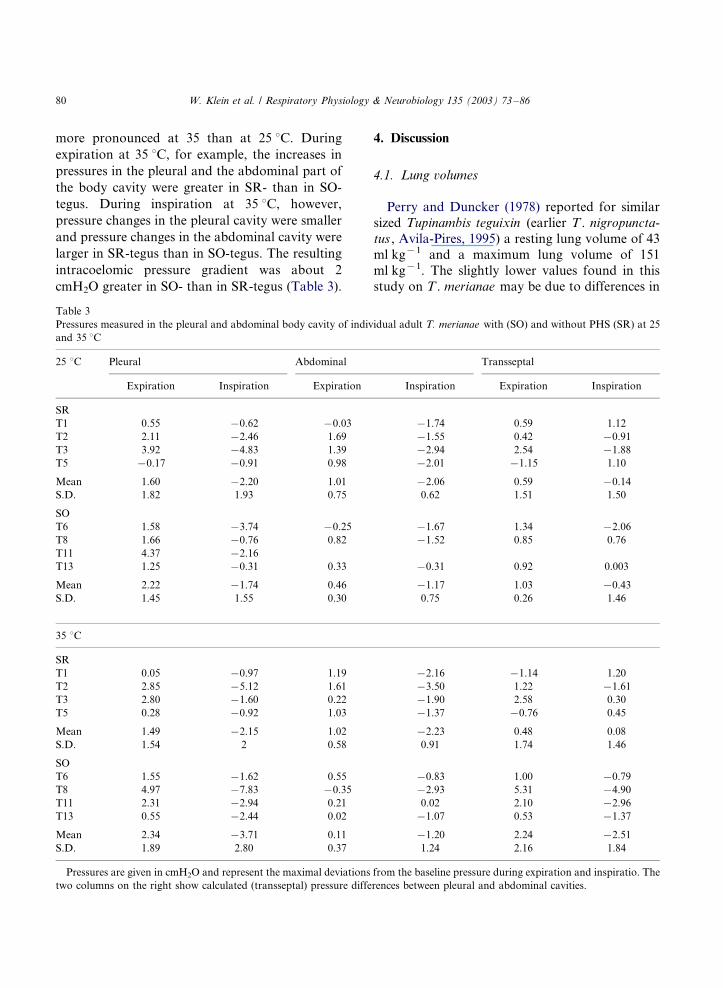
Table 3
Pressures measured in the pleural and abdominal body cavity of individual adult T. merianae with (SO) and without PHS (SR) at 25
and 35 8C
25 8C Pleural Abdominal Transseptal
Expiration Inspiration Expiration Inspiration Expiration Inspiration
SR
T1 0.55 �/0.62 �/0.03 �/1.74 0.59 1.12
T2 2.11 �/2.46 1.69 �/1.55 0.42 �/0.91
T3 3.92 �/4.83 1.39 �/2.94 2.54 �/1.88
T5 �/0.17 �/0.91 0.98 �/2.01 �/1.15 1.10
Mean 1.60 �/2.20 1.01 �/2.06 0.59 �/0.14
S.D. 1.82 1.93 0.75 0.62 1.51 1.50
SO
T6 1.58 �/3.74 �/0.25 �/1.67 1.34 �/2.06
T8 1.66 �/0.76 0.82 �/1.52 0.85 0.76
T11 4.37 �/2.16
T13 1.25 �/0.31 0.33 �/0.31 0.92 0.003
Mean 2.22 �/1.74 0.46 �/1.17 1.03 �/0.43
S.D. 1.45 1.55 0.30 0.75 0.26 1.46
35 8C
SR
T1 0.05 �/0.97 1.19 �/2.16 �/1.14 1.20
T2 2.85 �/5.12 1.61 �/3.50 1.22 �/1.61
T3 2.80 �/1.60 0.22 �/1.90 2.58 0.30
T5 0.28 �/0.92 1.03 �/1.37 �/0.76 0.45
Mean 1.49 �/2.15 1.02 �/2.23 0.48 0.08
S.D. 1.54 2 0.58 0.91 1.74 1.46
SO
T6 1.55 �/1.62 0.55 �/0.83 1.00 �/0.79
T8 4.97 �/7.83 �/0.35 �/2.93 5.31 �/4.90
T11 2.31 �/2.94 0.21 0.02 2.10 �/2.96
T13 0.55 �/2.44 0.02 �/1.07 0.53 �/1.37
Mean 2.34 �/3.71 0.11 �/1.20 2.24 �/2.51
S.D. 1.89 2.80 0.37 1.24 2.16 1.84
Pressures are given in cmH2O and represent the maximal deviations from the baseline pressure during expiration and inspiratio. The
two columns on the right show calculated (transseptal) pressure differences between pleural and abdominal cavities.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/8680

the species or to better maintenance conditionsand the resulting greater body mass in the
Brazilian animals. With increasing body mass,
VLr increased less than VLm, increasing the ratio
VLm to VLr from 3.0 in neonatals to 5.1 in adults.
Perry and Duncker (1978) found a ratio of 3.4 in
T . teguixin , similar to those of neonatals and
juveniles in the present study. The ratios reported
for man (3.0; Rahn et al., 1946), mouse (4.8;Crosfill and Widdicombe, 1961) and some lizards
(Lacerta 4.0; Gecko 4.7; Chamaeleon 5.0; Varanus
4.2; Perry and Duncker, 1978) are, despite lung
types varying from single- to multichambered
lungs, all in the same range, whereas SR-tegus
(VLm/VLr�/12.7) lie outside this range.
Maximum lung volume was found to increase as
MB0.975 in tegus, which is similar to the slopes
reported by Schmidt-Nielsen (1984) for mammals
and birds (MB0.94 and MB
1.06, respectively) and
found by Tenney and Tenney (1970) for amphi-
bians (MB1.05). Thus, maximum lung volume seems
to be proportional to body mass in tetrapods in
general.
VLr as well as VLm were significantly less in SR-
tegus than in other groups. VLr reaches 37.8% andVLm 73.4% of the respective volumes shown by
adults with intact PHS. The large difference in VLr
can be explained by the cranial movement of the
viscera in PHS-ectomotised tegus as described by
Klein et al. (in press). The smaller difference
between tegus with intact and surgically ruptured
PHS in VLm could be explained by the high
pressure (�/20 cmH2O) which was used to inflatethe lungs. The maximal change in lung volume
occurred in tegus at pressures between 0 and �/6
cmH2O. An excursion to �/20 cmH2O only reflects
the maximal possible distension of the lung tissues
and this may not change significantly in one year
without PHS. The decrease in lung volume seen in
tegus without a PHS does not necessarily lead to a
significant decrease in respiratory surface areabecause of the way in which faveolar parenchyma
responds to volume changes. When lung volume
decreases, the walls of the faveoli form accordion
pleats until parts of the epithelia come into contact
with each other (Daniels et al., 1994, 1995). If the
faveolar walls in tegus without PHS do not
collapse completely, the area for gas exchange
remains unaffected and oxygen uptake may re-main unchanged.
4.2. Compliance
The value for total compliance from the
non-standardised V�/P diagram found in this
study is similar to CT reported by Perry and
Duncker (1978) for T . teguixin (15.8 vs. 13.65
ml cmH2O�1, respectively). Perry and Duncker(1978) reported 0.47 ml cmH2O�1 for VLr-
standardised CT, which is similar to the value
found in this study (CT: 0.55 ml cmH2O�1). In
comparison with other lizards studied by Perry
and Duncker (1978) CT and CB of tegus are most
similar to those of the emerald lizard Lacerta
viridis , the tokay Gekko gecko , and the monitor
Varanus exanthematicus , but the values for CL
found in the present study are the lowest ones for
lizards reported so far. Only the emerald lizard has
a CL as low as that of the tegu. Both species belong
to the superfamily of lacertiform lizards (Estes et
al., 1988), which characteristically possess single-
chambered lungs with dense, homogeneously dis-
tributed parenchyma (Duncker, 1978). No differ-
ences were found in compliance between tegus ofdifferent sizes, which indicates that not only lung
volume but also static mechanical properties are
proportional to MB throughout development.
Further evidence for this hypothesis comes from
the values of compliance obtained from non-
standardised V�/P diagrams. Here, neonatal tegus
showed the lowest values of CT, CB and CL which
are comparable with those reported by Perry andDuncker (1978) for similar sized emerald lizards
(MB: 28 g; CT: 0.51 ml cmH2O�1; CB: 1.39
ml cmH2O�1; CL: 1.74 ml cmH2O�1). The allo-
metric relationships for CT, CL and CB of tegus
with intact PHS are shown in Table 4. With a mass
exponent close to 1.0, CL shows a similar propor-
tionality to MB as VLm, whereas CB and CT show
the same allometric relationship as VLr. VLr is aresult of forces interacting between lungs and
thorax. Due to surface tension phenomena and
elastic recoil, lungs tend to collapse, whereas the
rib cage shows a tendency to spring outwards. The
resulting balance of forces determines VLr (Agos-
tini et al., 1959). Apparently, neonatal tegus have a
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/86 81

stiffer body wall than adults, which leads to a
relatively greater VLr and thus to a smaller VLm/
VLr ratio than in adult tegus with intact PHS.
Tegus with ruptured PHS showed, on a VLr-
standardised V�/P diagram, significantly greater
CT, CB and CL than tegus with intact PHS and
especially CL is very high, exceeding CL of the
chameleon Chamaeleo chamaeleon (6.4 vs. 5.7
ml cmH2O�1, respectively; Perry and Duncker,
1978). This apparently high compliance in SR-
tegus was the result of the significantly lower VLr,
which leads to a steeper slope in the V�/P diagram
and thus to a greater VLr-standardised compliance.
Therefore, to compare the effect of a removed
PHS on static breathing mechanics, compliance
was also determined on V�/P diagrams standar-
dised with MB and VLm. On a body mass basis, CT,
CB and CL were significantly less in tegus without
PHS than in neonatal, juvenile and adult tegus
with intact PHS. In particular, CB was only half as
great as in the other groups. Not only the
abdominal wall but also the viscera belong to the
category ‘body wall’. There is no reason to believe
that in tegus without PHS the elastic properties of
the body wall changed due to the removal of the
PHS, as dissection did not reveal a marked
hypertrophy of the intercostal and abdominal
musculature. Therefore, the significantly lower
CB is solely due to the lack of a PHS. Tegus
without PHS must inflate the lungs not only
against the resistance of the abdominal wall but
also must displace the viscera which occupy a large
part of the former pleuro-hepatic cavity.
The VLm-standardised V�/P diagram gave no
significant differences in the compliance, which
may be expected from the proportionality of VLm
with MB and the small difference in VLm betweenoperated and intact tegus.
4.3. Hysteresis
The index of hysteresis (H) for the total
respiratory system of intact adult tegus found in
the present study is similar to HT reported by
Perry and Duncker (1978; 1.2 vs. 1.6 cm, respec-tively) and in the same range like HT of Lacerta .
HL in tegus, however, is less than HL of the
emerald lizard, but somewhat greater than HL of
the tokay and the chameleon (Perry and Duncker,
1978). The removal of the PHS had no significant
effect in H, based on VLr-, VLm- or MB-standar-
dised V�/P diagrams. Perry and Duncker (1978)
postulated that HL depends upon the degree ofpartitioning of the lungs, being greater in lungs
with uniformly distributed, fine parenchyma as
seen in Lacerta and less in lungs with ventral and
caudal dilatations as seen in Chamaeleo or Gecko .
The present results are therefore surprising, be-
cause the surface-to volume ratio for Lacerta and
Tupinambis are nearly identical (Perry, 1983).
Tegus in this study showed no difference inhysteresis over a body mass ranging from 17 to 700
g. The non-standardised H, however, showed a
markedly greater HT and HL in tegus without PHS
compared to the other groups.
During the determination of open-body V�/P
diagrams some lungs showed alternating contrac-
tions at mid-inflation causing a slight increase in
pressure. Such peristaltic waves are a long knowncharacteristics for turtles (Francois-Franck, 1906)
and lizards (Saalfeld, 1934) and have been attrib-
uted to spontaneous contractions of trabecular
smooth muscle (Perry and Duncker, 1978). They
may be of significant importance in the dense
parenchyma of tegus and could promote mixing of
air in the faveoli under apneic conditions and
allow more effective use oxygen stores in thecentral lumen of the lungs.
4.4. Pressures
In human or veterinary medicine, internal body
pressures are measured by inserting balloon cathe-
ters into the esophagus and stomach to measure
Table 4
Allometric relationships for non-standardised CT, CL and CB of
T. merianae with intact PHS expressed as y�/aMBb with a as
proportionally coefficient, b as body-mass exponent and y as
dependent variable
y a b P 9/S.E.M. R2
CT �/1.27 0.851 B/0.0001 0.05 0.954
CL �/1.06 0.996 B/0.0001 0.1 0.89
CB �/1.13 0.884 B/0.0001 0.08 0.916
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/8682

e.g. transdiaphragmatic pressures (e.g. Gilbert etal., 1979; Desmecht et al., 1992). Some problems
that arise with this technique are like different
pressures depending on whether or not the gastric
balloon lies in a fluid-filled part of the stomach, or
pressure decreases when the balloon is moved
caudally (Desmecht et al., 1992). Therefore, pres-
sures were measured with small, open tubes which
could be implanted quickly with minor surgery,and tegus seemed to be unaffected by this proce-
dure. Although the exact position of the tip of the
tubes could not be determined, their position in
the middle of the pleural or abdominal cavity,
respectively, was sufficient to reflect pressures in
these regions of the body cavity. It is possible, that
the lack of large pressure changes during expira-
tion may be caused by blocking of the tube by theviscera. In this case, there would be no signal at all
during expiration or inspiration. It is unlikely that
both the abdominal and pleural tubes were
blocked at the same time throughout the entire 3
h of recording and despite several flushing events.
In general, expiration in tegus was accompanied
by an increase and inspiration by a decrease in
pressure compared with the breath-hold period.The pressure changes accompanying expiration,
however, were not continuous throughout a mea-
suring period in an individual tegu, and when the
animals were moving, the signals could change
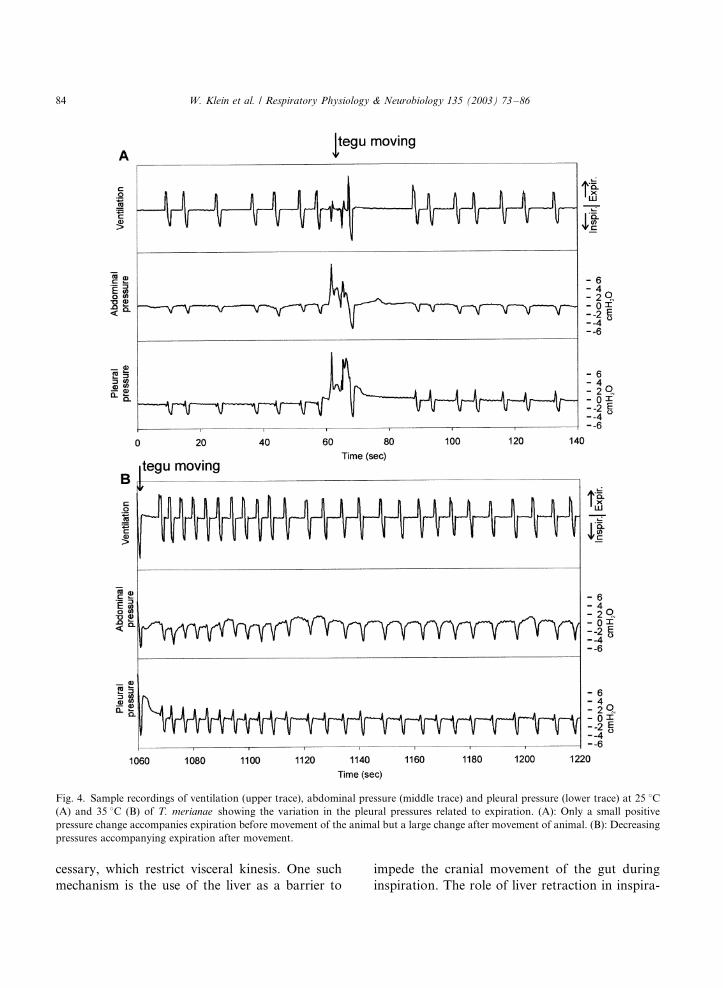
(Fig. 4A). Other tegus showed a continuous
decrease in expiratory pressures after an activity
bout (Fig. 4B). During rest, expiration was mainly
an active process and the increase in pressure wascaused by a contraction of the body wall muscles,
thus moving the ribs caudad and mediad and
reducing the volume of the body cavity. The small
pressures recorded in some animals during expira-
tion may be explained by the elastic recoil or
surface tension of the lungs. A decrease in lung
volume together with an unchanged volume in rib
cage position could cause a small negative pressurein the pleural cavity at the beginning of expiration.
Afterwards, the chest wall also moved inward,
either by its elastic recoil or by active contraction,
and caused a small increase in pressure. Whenever
the expiratory air flow stopped and inspiration
began, pressures fell dramatically, causing a nega-
tive pressure in the pleural cavity, and air flowed
into the lungs. Termination of inspiration wascoincident with abolition of the transthoracic
pressure differential.
The changes in pressures in the abdominal
cavity were, in general, smaller than those in the
pleural cavity. This held true even for tegus
without PHS, which agrees with the hypothesis
of Gans (1970), that pressures on the viscera are
transmitted by local interaction of forces betweenthe different compartments rather than by the
whole body cavity acting as a single fluid filled
space. The residual transseptal pressure difference
in SR-tegus could be induced by the liver, which
partially protects the right lung from the viscera.
At the greater temperature, the transseptal pres-
sure was much greater in SO- than in SR-tegus:
i.e., the pressures in the body cavity were moreevenly distributed in animals that lacked a PHS.
Farmer and Hicks (2000) measured pressures in
the abdominal part of the body cavity in Iguana
iguana . They recorded an increase in pressure in
the body cavity during exercise caused by the
activation of the trunk muscles during locomotion,
and further that elevated pressures provoke col-
lapsing of abdominal veins and thereby reduce thevenous return from the caudal body.
In tegus with intact PHS, the greatest part of the
pressure changes necessary for ventilation may be
restricted by the PHS to the thoracic region,
particularly under conditions of elevated meta-
bolic demand. This situation would maintain low
pressures in the abdominal part of the body cavity,
and thus would not reduce venous return evenduring light exercise. The fact that the role of the
PHS in increasing the intracoelomic pressure
gradient at rest is equivocal, however, suggests
that this is not the primary function of the septum.
Rather, its main function appears to be preventing
the craniad migration of the viscera during in-
spiration (Klein et al., in press).
The respiratory system of T. merianae givesvaluable insights in the breathing mechanics in
lizards and allows us to construct a functional
morphological scenario about the evolution of the
mammalian lung/diaphragm complex. In lizards,
costal aspiration breathing may be efficient when
the lungs are highly compliant. With reduced lung
compliance some mechanisms appears to be ne-
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/86 83
cessary, which restrict visceral kinesis. One such
mechanism is the use of the liver as a barrier to
impede the cranial movement of the gut during
inspiration. The role of liver retraction in inspira-
Fig. 4. Sample recordings of ventilation (upper trace), abdominal pressure (middle trace) and pleural pressure (lower trace) at 25 8C(A) and 35 8C (B) of T. merianae showing the variation in the pleural pressures related to expiration. (A): Only a small positive
pressure change accompanies expiration before movement of the animal but a large change after movement of animal. (B): Decreasing
pressures accompanying expiration after movement.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/8684

tion in crocodiles and testudines is well known(Gans and Clark, 1976; Gaunt and Gans, 1969),
and ‘abdominal breathing’ in mammals also in-
volves liver displacement. The present studies
suggest that dynamic fixation of the liver during
inspiration may precede hepatic kinesis as an
accessory respiratory mechanism in general.
The superficial similarity of the teiid PHS and
the embryonic mammalian diaphragm as a trans-verse perforated barrier, which fixes the liver in the
tegu and divides the body cavity between lungs
and liver in the mammal, is striking. Later in
ontogeny the mammalian diaphragm closes its
perforations whereas in the tegu an opening
persists, suggesting that the basic function of
PHS and diaphragm may have been similar during
early phylogenetic stages in mammalian ancestors.In both cases the presence of this transverse barrier
allows the efficient inflation of low-compliance
lungs. In the mammal striated muscle may have
replaced smooth muscle (Brink, 1956), or may
have been present from the beginning (Keith,
1905; Graper, 1928) allowing breath-by-breath
participation of the diaphragm, whereas the
smooth muscle of the teiid PHS is expected toallow only tonal increase of the efficiency of costal
breathing.
Acknowledgements
Financial support was provided by a PRO-
BRAL (CAPES/DAAD) grant to ASA and SFP
and a Graduiertenstipendium des Landes NRW to
WK. We also thank two anonymous reviewers for
helpful comments on the manuscript.
References
Agostini, E., Thimm, F.F., Fenn, W.O., 1959. Comparative
features of the mechanics of breathing. J. Appl. Physiol. 14,
679�/683.
Avila-Pires, T.C.S., 1995. Lizards of Brazilian Amazonia
(Reptilia; Squamata). Zool. Verh. (Leiden) 299, 1�/706.
Bernstein, L., 1957. The elastic pressure�/volume curves of the
lungs and thorax of the living rabbit. J. Physiol. (Lond.)
138, 473�/487.
Brink, A.S., 1956. Speculations on some advanced mammalian
characteristics in the higher mammalian-like reptiles. Pa-
laeontol. Afr. 4, 77�/96.
Broman, I., 1904. Die Entwicklungsgeschichte der Bursa
omentalis und ahnlicher Rezessbildungen bei den Wirbel-
tieren. Entwicklungsgeschichtliche Monographien, 1�/611.
Crosfill, M.L., Widdicombe, J.G., 1961. Physical characteristics
of the chest wall and lungs and the work of breathing in
different mammalian species. J. Physiol. Lond. 158, 1�/14.
Daniels, C.B., McGregor, L.K., Nicholas, T.E., 1994. The
dragon’s breath: a model for the dynamics of breathing and
faveolar ventilation in agamid lizards. Herpetologica 50,
251�/261.
Daniels, C.B., Orgeig, S., Smits, A.W., 1995. The composition
and function of reptilian pulmonary surfactant. Respir.
Physiol. 102, 121�/135.
Desmecht, D., Rollin, F., Lekeux, P., 1992. Transdiaphrag-
matic pressure measurement in cattle: technical data. Res.
Vet. Sci. 53, 148�/153.
Duncker, H.R., 1978. Coelom-Gliederung der Wirbeltiere�/
Funktionelle Aspekte. Verh. Anat. Ges. 72, 91�/112.
Estes, R., de Queiroz, K., Gauthier, J., 1988. Phylogenetic
relationships within Squamata. In: Estes, R., Pregill, G.
(Eds.), Phylogenetic Relationships in Lizard Families.
Stanford University Press, pp. 119�/281.
Farmer, C.G., Hicks, J.W., 2000. Circulatory impairment
induced by exercise in the lizard Iguana iguana . J. Exp.
Biol. 203, 2691�/2697.
Francois-Franck, C.E., 1906. La mecanique respiratoire des
Cheloniens. I.- Contractilite de l’appareil pulmonaire de
tortue terrestre. In: Comptes Rendus Hebdomadaires des
Seances et Memoires Libraires de la Societe de Biologie.
Libraires de L’academie de Medecine, Paris, pp. 968�/970.
Gans C., 1970. Strategy and sequence in the evolution of the
external gas exchangers of ectothermal vertebrates. forma et
functio 3, 61�/104.
Gans, C., Clark, B.D., 1976. Studies in ventilation of Caiman
crocodilus (Crocodilia: Reptilia). Respir. Physiol. 26, 285�/
301.
Gaunt, A.S., Gans, C., 1969. Mechanics of respiration in the
snapping turtle, Chelydra serpentina (Linne). J. Morphol.
128, 195�/228.
Gilbert, R., Peppi, D., Auchincloss, J.H., Jr., 1979. Measure-
ments of transdiaphragmatic pressure with a single gastric-
esophageal probe. J. Appl. Physiol. 47, 628�/630.
Graper, L., 1928. Zwerchfell, Lunge und Pleurahohlen in der
Tierreihe. Anat. Anz. Erg. Hefte 66, 71�/77.
Hochstetter F., 1906. Uber die Entwicklung der Scheidewand-
bildungen in der Leibeshohle der Krokodile. Voeltzkows
Reise in Ostafrika; Wissenschaftliche Ergebnisse 4, 141�/
205.
Keith, A., 1905. The nature of the mammalian diaphragm and
pleural cavities. J. Anat. Physiol. 39, 243�/284.
Klein, W., Bohme, W., Perry, S.F., 2000. The mesopneumonia
and the post-hepatic septum of the Teiioidea (Reptilia:
Squamata). Acta Zool. (Stockholm) 81, 109�/119.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/86 85

Klein, W., Abe, A.S., Andrade, D.V., Perry, S.F. Structure of
the post-hepatic septum and its influence on visceral
topology in the Tegu lizard, Tupinambis merianae (Teiidae:
Reptilia). J. Morphol., in press.
Perry, S.F., 1983. Reptilian lungs. Functional anatomy and
evolution. Adv. Anat. Embryol. Cell Biol. 79, 1�/81.
Perry, S.F., Duncker, H.R., 1978. Lung architecture, volume
and static mechanics in five species of lizards. Respir.
Physiol. 34, 61�/81.
Perry, S.F., Duncker, H.R., 1980. Interrelationship of static
mechanical factors and anatomical structure in lung evolu-
tion. J. Comp. Physiol. B 138, 321�/334.
Radford, E.P., Jr., 1964. Static mechanical properties of
mammalian lungs. In: Fenn, W.O. (Ed.), Handbook of
Physiology: Respiration (Chaptor 15). Am. Physiol. Soc,
Washington D.C..
Rahn, H., Otis, A.B., Chadwick, L.E., Fenn, W.O., 1946. The
pressure�/volume diagram of the thorax and lung. Am. J.
Physiol. 146, 161�/178.
Schmidt-Nielsen, K., 1984. Why is Animal Size So Important?.
Cambridge University Press, p. 241.
Saalfeld, E.V., 1934. Die Mechanik der Atmung bei Uromastix
(Lacertilia). Pflugers Archiv 223, 431�/448.
Templeton, J.R., 1964. Cardiovascular responses during buccal
and thoracic respiration in the lizard Sauromalus obesus .
Comp. Biochem. Physiol. 11, 31�/43.
Tenney, S.M., Tenney, J.B., 1970. Quantitative morphology of
cold�/blooded lungs: amphibia and reptilia. Respir. Physiol.
9, 197�/215.
W. Klein et al. / Respiratory Physiology & Neurobiology 135 (2003) 73�/8686
Copyright © 2022 FDOKUMEN




















