
Screening for the Optimal Specificity Profile of Protein Kinase C Using Electrospray...
10
http://jbx.sagepub.com/ Journal of Biomolecular Screening http://jbx.sagepub.com/content/10/4/320 The online version of this article can be found at: DOI: 10.1177/1087057104274353 2005 10: 320 J Biomol Screen Mart Loog, Bo Ek, Nikita Oskolkov, Ale Närvänen, Jaak Järv and Pia Ek Screening for the Optimal Specificity Profile of Protein Kinase C Using Electrospray Mass-Spectrometry Published by: http://www.sagepublications.com On behalf of: Journal of Biomolecular Screening can be found at: Journal of Biomolecular Screening Additional services and information for http://jbx.sagepub.com/cgi/alerts Email Alerts: http://jbx.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: http://jbx.sagepub.com/content/10/4/320.refs.html Citations: What is This? - Jun 10, 2005 Version of Record >> by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from by guest on October 11, 2013 jbx.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Screening for the Optimal Specificity Profile of Protein Kinase C Using Electrospray...

http://jbx.sagepub.com/Journal of Biomolecular Screening
http://jbx.sagepub.com/content/10/4/320The online version of this article can be found at:
DOI: 10.1177/1087057104274353
2005 10: 320J Biomol ScreenMart Loog, Bo Ek, Nikita Oskolkov, Ale Närvänen, Jaak Järv and Pia Ek
Screening for the Optimal Specificity Profile of Protein Kinase C Using Electrospray Mass-Spectrometry
Published by:
http://www.sagepublications.com
On behalf of:
Journal of Biomolecular Screening
can be found at:Journal of Biomolecular ScreeningAdditional services and information for
http://jbx.sagepub.com/cgi/alertsEmail Alerts:
http://jbx.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://jbx.sagepub.com/content/10/4/320.refs.htmlCitations:
What is This?
- Jun 10, 2005Version of Record >>
by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from by guest on October 11, 2013jbx.sagepub.comDownloaded from

10.1177/1087057104274353Loog et al.Optimal Specificity Profile of Protein Kinase C
Screening for the Optimal Specificity Profile ofProtein Kinase C Using Electrospray Mass-Spectrometry
MART LOOG,1 BO EK,2 NIKITA OSKOLKOV,3 ALE NÄRVÄNEN,4 JAAK JÄRV,3 and PIA EK1
A peptide library approach based on electrospray mass-spectrometric (ESI-MS) detection of phosphopeptides was designedfor rapid and quantitative characterization of protein kinase specificity. The kcat/Km values for the protein kinase Cβ (PKCβ)were determined for a systematically varied set of individual substrate peptides in library mixtures by the ESI-MS method.The analysis revealed a complex structural specificity profile in positions around the phosphorylated serine with hydrophobicand/or basic residues being mostly preferred. On the basis of the kinetic parameters, a highly efficient peptide substrate forPKCβ (Km value below 100 nM) FRRRRSFRRR and its alanine substituted pseudosubstrate-analog inhibitor (Ki value of 76nM) were designed. The quantitative specificity profiles obtained by the new approach contained more information aboutkinase specificity than the conventional substrate consensus motifs. The new method presents a promising basis for design ofsubstrate-site directed peptide or peptidomimetic inhibitors of protein kinases. Second, highly specific substrates could be de-signed for novel applications such as high-throughput protein kinase activity screens on protein kinase chips. (Journal ofBiomolecular Screening 2005:320-328)
Key words: protein kinases, protein kinase inhibitors, protein kinase C, mass-spectrometry, peptide library
PHOSPHORYLATION OF TARGET PROTEINS by protein kinases isthe most abundant type of molecular regulation, involved in
virtually every cellular process, and the fidelity of this regulationmechanism is ensured by the substrate specificity of these en-zymes. Detailed knowledge of the substrate specificity profile en-ables the design of the specific inhibitors of protein kinases that canbe used for therapeutic intervention with different diseases. Sec-ond, specific substrates of protein kinases can be designed for useas molecular probes for diagnostic purposes. The knowledge aboutthe substrate specificity profile may also enable the prediction ofthe potential phosphorylation targets in protein sequencedatabases.
In earlier studies, the minimal consensus motifs of substraterecognition sites for a number of protein kinases have been definedby using synthetic peptide substrates.1 However, the consensusmotifs contain a limited amount of information because they show
only the most favorable amino acids in a few key positions aroundthe phosphorylatable amino acid residue. Because of the high re-dundancy of such sequence motifs, they have no particular valuefor phosphorylation site predictions. It has now been largely recog-nized that the substrate site specificity of protein kinases involves ahigher degree of complexity,2 which can be fully characterizedonly by extensive and systematic structural variations in the criticalregion around the phosphorylation site. In recent years, combina-torial peptide library mixtures have been successfully used for thistask in a number of cases.3-5 The advantage of these methods is thereduction in time and cost by performing simultaneous analysis ofmultiple peptides in a mixture. However, if compared with theclassical kinetic assay using individual peptide substrates, the li-brary technologies reported so far do not provide quantitative ki-netic data for individual components of the mixture. Moreover, inmost cases, no attention has been paid to the fact that the compara-ble quantitative data of peptide phosphorylation can be obtainedonly if the substrate turnover is followed under the initial velocityconditions.
In recent years, the electrospray mass-spectrometry (ESI-MS)technology has been repeatedly shown to be among the most sensi-tive methods for quantitative detection and resolution of complexpeptide mixtures6,7 providing a linear range of detection of morethan 4 orders of magnitude and a detection limit reaching to lowfemtomolar concentrations. Here, we demonstrate that the appli-cation of ESI-MS for resolution and quantitative detection of a ra-tionally designed peptide substrate library gives new perspectives
320 www.sbsonline.org © 2005 The Society for Biomolecular Screening
1Department of Medical Biochemistry and Microbiology, Uppsala University, 75123 Uppsala, Sweden2Swedish Agricultural University, 750 07 Uppsala, Sweden3Institute of Organic and Bioorganic Chemistry, Tartu University, 51014 Tartu,Estonia4Department of Organic Chemistry, Kuopio University, 70211 Kuopio, Finland
Received Jul 30, 2004, and in revised form Oct 13, 2004. Accepted for publica-tion Oct 29, 2004.
Journal of Biomolecular Screening 10(4); 2005DOI: 10.1177/1087057104274353

for quantitative and high-throughput characterization of proteinkinase specificity.
The general aim of the current study was to develop a quantita-tive ESI-MS–based method for studying the protein kinase speci-ficity with peptide libraries. In our previous studies, we haveshown that a peptide structure LLRLHSLRER is an efficient sub-strate for at least 2 relatively distant representatives of 2 largekinase groups; the mammalian protein kinase C (PKC)β (repre-sentative of the PKC family in ACG group) and the plant calcium-dependent protein kinase (CaMK group).8,9 Such overlappingspecificity suggests that this structure may serve as a universaltemplate for kinases from these 2 groups. Using the split synthesisstrategy, a mixture of amino acids was introduced into each subse-quent position of the peptide, leaving the rest of the structure un-changed. The amino acids were chosen by taking into account thatthe mass per charge ratios in positive ion mode of the spectrawould not be overlapping.
With the help of the new method, the sensitivity of structuralvariation for substrates to PKC could be characterized with highprecision in terms of kcat/Km values for the catalyzed reaction. Byevaluating the specificity determinants at each position, we wereable to design a highly efficient peptide substrate for PKC, as wellas its alanine-substituted pseudosubstrate-analog inhibitor. Fi-nally, we show that the obtained specificity profile may provide areliable tool for predicting the presence of phosphorylation sites ofPKC from protein sequence databases.
MATERIALS AND METHODS
Synthesis of the peptide libraries
The peptide libraries were synthesized using PerSeptive 9050Plus automated peptide synthesizer with Fmoc strategy, usingTBTU/DIPEA as the coupling reagent and dimethylformamide(DMF) as solvent. For the synthesis, NovaSyn TGA with 4-hydroxymethylphenoxyacetic acid linker was used(Novabiochem, Läufelfingen, Switzerland). The side-chain pro-tecting groups used in synthesis were 2,2,4,6,7-pentamethyl-dihydrogenbenzofurane-5-sulfonyl (Pbf) for Arg, trityl (Trt) forAsn and His, O-tert-butyl (OtBu) for Glu, t-butyloxycarbonyl(t-Boc) for Lys, and tert-butyl (tBu) for Ser. The 8 different aminoacids (A, N, E, G, I, K, F, and V) were coupled to the position Xmanually. Each of the 9 libraries had the following overall struc-tures: XLRLHSLRER, LXRLHSLRER, LLXLHSLRER,LLRXHSLRER, LLRLXSLRER, LLRLHSXRER,LLRLHSLXER, LLRLHSLRXR, and LLRLHSLREX. The syn-thesizer was programmed to stop at position X, and the resin wasremoved to 8 different columns. Amino acids were activated man-ually, then added separately to the columns and stirred overnight atroom temperature. After washing 3 times with DMF, the couplingwas monitored by a ninhydrin test.10 Following the manual cou-pling, resins were pooled and returned to the synthesizer to com-
plete the synthesis. Peptides were verified using a MALDI-TOFmass spectrometer (Bruker, Germany).
PKC
PKCβ was isolated from pig spleen according to the method ofParker and Marais11 with the modifications of Ferrari andThomas.12 Human recombinant PKCβII was obtained fromCalbiochem and was used as a control in the kinetic studies withthe newly designed peptides.
Phosphorylation of the peptide libraries
Phosphorylation of the peptide libraries by PKC was performedin a reaction mixture containing 50 mM Tris-HCl (pH 7.5), 5%DMSO, 0.75 mM CaCl2, 60 µg/ml phosphatidylserine, 1 µg/mldiolein, 0.5 mg/ml bovine serum albumin, 10 mM MgCl2, 0.2 mM[γ-32P]ATP (Amersham Biosciences), and 0.5 to 10 nM PKC. Thereaction was carried out at room temperature and was initiated byaddition of [γ-32P]ATP (100-200 cpm/pmol) and MgCl2. Aliquotesof 500 µl were taken at several time points, and the reaction wasstopped with 0.3 volumes of acidic stop solution composed of 1MHCl added to a solution of 50 mM MES (pH 5.5)/1M NaCl, in avolume ratio of 1:40. The pH of the aliquots was titrated back to pH5.5 with a predetermined amount of the same MES/NaCl buffercontaining NaOH as the basic component. The phosphopeptideswere subsequently isolated using a ferric chelating column basedon Chelating-Sepharose 6B as described in Muszynkska et al.13
The 250 µl ferric column was equilibrated with MES buffer (pH5.5) containing 1M NaCl. Ten column volumes of the same bufferwere used to wash the column, and the phosphopeptides wereeluted using 1.8 ml of 20 mM sodium phosphate buffer (pH 7.7)containing 1M NaCl. A defined amount of the standard peptideLLRLHSLRE was added to the eluted phosphopeptides, and theresultant mixture was desalted using a 10-µl column of R3 beads(Poros) packed into a gel-loading pipette tip, equilibrated with0.5% formic acid. After loading the sample, the column waswashed with 200 µl of 1% formic acid and 20 µl of 0.5% formicacid. The peptides were eluted with 30 µl of 25% acetonitrile in0.5% formic acid, and 1.5 µl of the obtained eluate was transferredinto the nanospray needle for quantification of the phosphopeptidepatterns using ESI-MS. All initial velocity experiments were per-formed a minimum of 2 times. The same sample preparation pro-cedure was performed for the phosphopeptide calibration experi-ments. For these experiments, the libraries were phosphorylatedusing larger amounts of enzyme and longer time periods to achievemaximal phosphorylation of all components. Different volumes ofthe obtained solutions were then mixed with appropriate volumesof stop solution and nonphosphorylated reaction mixture contain-ing the same library. In this way, the calibration mixtures were gen-erated, containing equally phosphorylated library components inconcentrations covering the concentration range of the phospho-peptides observed in aliquots taken at initial velocity conditions. In
Optimal Specificity Profile of Protein Kinase C
Journal of Biomolecular Screening 10(4); 2005 www.sbsonline.org 321

the case of 2 libraries, LLXLHSLRER and LLRLHSLXER, someof the peptides were not 100% phos- phorylated over the time peri-ods used. In these cases, the values corresponding to 100% werecalculated by taking into account the relative amount of remainingnonphosphorylated peptides compared to their relative amounts innonphosphorylated libraries. In parallel, the initial velocity mea-surements were also followed by the phosphocellulose papermethod as in Loog et al.,9 to quantitatively estimate the extent of li-brary phosphorylation. The phosphocellulose paper method wasalso used for determination of Km and Vmax values for peptides thatwere synthesized separately. For each peptide, the reaction timeand amount of enzyme used was adjusted so that the initial rates ofthe phosphorylation reaction could be measured. The reactionswere initiated by addition of the [γ-32P]ATP (specific radioactivity100-200 cpm/pmol).
ESI-MS analysis of the libraries
The mass-spectrometric analyses were carried out on a Micro-mass Q-TOF hybrid mass spectrometer (Micromass, Manchester,UK) equipped with an orthogonal electrospray source (Z-spray),operated in the electrospray positive ion mode. The nanosprayneedle containing 1.5 µl of desalted peptide library solution wasused for injection. The optimized conditions were capillary volt-age 800 V, source temperature 80° C, and cone voltage 25 V. TheESI gas was nitrogen. Q1 was set to broad bandwith and wasscanned between 200 and 400 with a cycle time of 1 s. The TOFanalyzer collected data between m/z 200 and 800. The spectrawere analyzed using Masslynx software 3.5. The background ofthe spectra was subtracted, and centered peak heights of the 1stpeaks of isotopic clusters of both +3 and +4 (when present) chargevariants were summarized for each peptide. Because of the partialspectral overlap, the intensities for peptides containing N or E invaried positions were obtained similarily except that the predictedvalue of the 2nd isotopic peak of peptides containing I and K, re-spectively, were previously subtracted from the 1st peak values ofpeptides containing N and E in varied positions. The concentrationof the desalted library used for injection to MS was always kept solow that the total ion current of the most abundant peak in a singlespectra was below 1000, so that dead time correction was notnecessary to obtain a linear calibration curve.
Determination of kinetic constants for the peptide libraryphosphorylation reaction
At saturating MgATP concentrations, the total initial velocity ofthe library phosphorylation (vt) can be expressed as the sum of in-dividual initial velocities in the form of a modified Michaelis-Menten equation,14 in which the different library components re-late to each other as competitive inhibitors. In this situation, the ap-parent Km for an individual peptide A can be expressed as its realKm multiplied by a factor (1 + [B]/KmB + [C]/KmC . . . etc.), where B,C, and so forth denote other library components. It is easy to dem-
onstrate that the values of vo/[S] for each library component relateto each other as a profile of constants kcat/Km. Thus, dividing therelative spectral intensity values obtained for each phosphopeptideat initial velocity conditions with the corresponding values ob-tained from the calibration experiment, where the same librarymixture forms a spectral pattern characteristic for its fully phos-phorylated state, gives a set of values relating to each other as thekcat/Km values of the library components. The kcat/Km, or the “speci-ficity constant,” but not the Km values alone, are the best character-istics for the specificity of a substrate toward a particular enzyme,representing the specificity in the sense of discriminating between2 or more competing substrates in a mixture.15 This is analogous tothe situation in vivo in which several possible substrate sites arepotentially present.
Working in initial velocity conditions is important for obtainingthe correct kcat/Km values for each library component. According tothis criterion, at the point of taking an aliquot, the extent ofphosphorylation of any individual component of interest shouldnot exceed 10%. In several libraries, the initial velocity differencesfor individual peptides were large, and additional aliquots at higherphosphorylation levels were taken for accurate quantification ofthe signals of the peptides with lower activity. In these cases, atleast 1 peptide must be within the range of 10% of turnover in bothof the 2 subsequent aliquots, to serve as a reference value to obtainthe full initial velocity profile for all library components.
RESULTS
Design of the peptide library
We focused on developing the ESI-MS–coupled library ap-proach for kinases, which require 1 or multiple basic residues asthe main specificity determinants in the minimal consensus se-quence. These enzymes, also classified as the basophilic proteinkinases,16 belong mainly to the ACG and CaMK groups of the pro-tein kinase superfamily17 and have the common requirement forthe basic residue in position –3 from the phosphorylated aminoacid. Nine libraries were designed and synthesized, each contain-ing a nearly equimolar mixture of 9 amino acids in 1 of the 9 posi-tions flanking the serine in the template structure LLRLHSLRER.The varied mixture contained amino acids with basic, acidic, hy-drophilic, and hydrophobic side chains, providing minimal butsufficient structural variation for the study of structure-activity re-lationships. The other natural amino acids were excluded becauseof possible phosphoacceptor hydroxyl groups (S, T, Y), low stabil-ity (C, M, W), or overlapping of mass-spectral signals fromphosphorylated or nonphosphorylated forms from other librarycomponents (L, Q, H, D). For primary evaluation of the methodand to enable a comparison of the data in terms of kcat/Km, the tem-plate peptide and the peptides with arginine substitutions in eachposition were synthesized separately (Table 1). The Km and Vmax
values of these peptides were determined by the conventionalphosphocellulose paper method.
Loog et al.
322 www.sbsonline.org Journal of Biomolecular Screening 10(4); 2005
Optimal Specificity Profile of Protein Kinase C
Journal of Biomolecular Screening 10(4); 2005 www.sbsonline.org 323
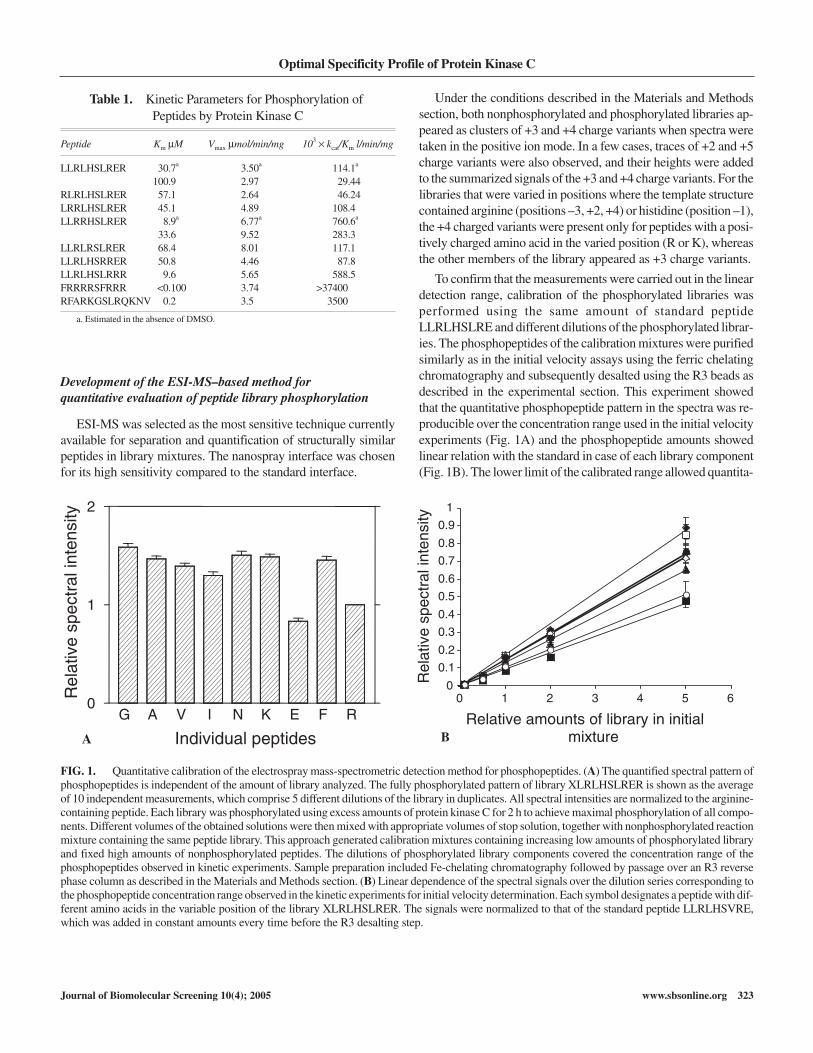
Development of the ESI-MS–based method forquantitative evaluation of peptide library phosphorylation
ESI-MS was selected as the most sensitive technique currentlyavailable for separation and quantification of structurally similarpeptides in library mixtures. The nanospray interface was chosenfor its high sensitivity compared to the standard interface.
Under the conditions described in the Materials and Methodssection, both nonphosphorylated and phosphorylated libraries ap-peared as clusters of +3 and +4 charge variants when spectra weretaken in the positive ion mode. In a few cases, traces of +2 and +5charge variants were also observed, and their heights were addedto the summarized signals of the +3 and +4 charge variants. For thelibraries that were varied in positions where the template structurecontained arginine (positions –3, +2, +4) or histidine (position –1),the +4 charged variants were present only for peptides with a posi-tively charged amino acid in the varied position (R or K), whereasthe other members of the library appeared as +3 charge variants.
To confirm that the measurements were carried out in the lineardetection range, calibration of the phosphorylated libraries wasperformed using the same amount of standard peptideLLRLHSLRE and different dilutions of the phosphorylated librar-ies. The phosphopeptides of the calibration mixtures were purifiedsimilarly as in the initial velocity assays using the ferric chelatingchromatography and subsequently desalted using the R3 beads asdescribed in the experimental section. This experiment showedthat the quantitative phosphopeptide pattern in the spectra was re-producible over the concentration range used in the initial velocityexperiments (Fig. 1A) and the phosphopeptide amounts showedlinear relation with the standard in case of each library component(Fig. 1B). The lower limit of the calibrated range allowed quantita-
Table 1. Kinetic Parameters for Phosphorylation ofPeptides by Protein Kinase C
Peptide Km µM Vmax µmol/min/mg 103 kcat/Km l/min/mg
LLRLHSLRER 30.7a 3.50a 114.1a
100.9 2.97 29.44RLRLHSLRER 57.1 2.64 46.24LRRLHSLRER 45.1 4.89 108.4LLRRHSLRER 8.9a 6.77a 760.6a
33.6 9.52 283.3LLRLRSLRER 68.4 8.01 117.1LLRLHSRRER 50.8 4.46 87.8LLRLHSLRRR 9.6 5.65 588.5FRRRRSFRRR <0.100 3.74 >37400RFARKGSLRQKNV 0.2 3.5 3500
a. Estimated in the absence of DMSO.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 1 2 3 4 5 6
Relative amounts of library in initial mixture
ytisnetni l art ceps evit al eR
G A V I N K E F R0
1
2
Individual peptides
ytisnetni l art ceps evit al eR
FIG. 1. Quantitative calibration of the electrospray mass-spectrometric detection method for phosphopeptides. (A) The quantified spectral pattern ofphosphopeptides is independent of the amount of library analyzed. The fully phosphorylated pattern of library XLRLHSLRER is shown as the averageof 10 independent measurements, which comprise 5 different dilutions of the library in duplicates. All spectral intensities are normalized to the arginine-containing peptide. Each library was phosphorylated using excess amounts of protein kinase C for 2 h to achieve maximal phosphorylation of all compo-nents. Different volumes of the obtained solutions were then mixed with appropriate volumes of stop solution, together with nonphosphorylated reactionmixture containing the same peptide library. This approach generated calibration mixtures containing increasing low amounts of phosphorylated libraryand fixed high amounts of nonphosphorylated peptides. The dilutions of phosphorylated library components covered the concentration range of thephosphopeptides observed in kinetic experiments. Sample preparation included Fe-chelating chromatography followed by passage over an R3 reversephase column as described in the Materials and Methods section. (B) Linear dependence of the spectral signals over the dilution series corresponding tothe phosphopeptide concentration range observed in the kinetic experiments for initial velocity determination. Each symbol designates a peptide with dif-ferent amino acids in the variable position of the library XLRLHSLRER. The signals were normalized to that of the standard peptide LLRLHSVRE,which was added in constant amounts every time before the R3 desalting step.
BA

tive detection of each phosphopeptide at picomolar concentration.Such results confirmed that the described simple 2-step samplepreparation could be reproducibly used for quantitative analysis ofpeptide phosphorylation in the library mixtures by ESI-MS.
The spectra taken for initial velocities and calibration mixturesfor the libraries XLRLHSLRER and LXRLHSLRER are shown asthe +4 charged components in Figures 2A and 2B. The compo-nents of these 2 libraries have the same molecular masses and showvery similar spectral patterns when fully phosphorylated. In con-
trast, profoundly different peak heights were observed for theindividual peptide components in the initial velocity spectra. Fur-thermore, large differences were observed when the initial velocityspectra of the 2 libraries were compared, demonstrating strong se-quence specificity of PKC for these 2 adjacent positions.
Specificity profile for PKC obtainedfrom the peptide library experiments
The obtained specificity profiles presenting the kcat/Km valuesfor each library component in the PKCβ-catalyzed phosphory-lation reaction are given in the form of diagrams in Figure 3. In ad-dition, the Km and Vmax values for template peptide and for the pep-tides in which amino acids were successively replaced by argininewere determined separately (Table 1). The position showingthe highest sensitivity for amino acid substitutions was position –3upstream from the serine. It was not surprising because it is knownto be a conserved substrate recognition feature in basophilic kin-ases.18 According to our data, arginine is exclusively preferred overlysine in this position. The most important specificity determinantat the C-terminal side of the peptide was found to be arginine in po-sition +2. These results are in agreement with a number of previousstudies reviewed by Kennelly and Krebs.1 In addition, arginine wasstrongly preferred over the other amino acids in positions –5, –2,+3, and +4. However, the specificity in these positions was not asstringent as that of position –3, and lysine was well favored, thoughalways less than arginine. The only position in which positiveamino acids were not preferred over the hydrophobic ones was po-sition –5, where a phenylalanine substitution showed strong effect.However, arginine was still effective, demonstrating only about a2-fold lower activity relative to phenylalanine. Position +1 wasshown to tolerate both basic and bulky hydrophobic amino acids,whereas amino acids with small side chains such as glycine andalanine produced a strong negative effect. In contrast, glycine andalanine, in position –1, contributed a positive effect, comparable tothe dominating arginine and lysine residues. In general, comparedto other varied positions, position –1 showed the lowest overallspecificity for amino acid substitutions.
In all varied positions, the positively charged arginine residuesshowed strong preference over negatively charged glutamic acidsubstitutions, pointing to a universal importance for electrostaticinteractions in the vicinity of the phosphorylation site. Aside fromthe exceptions in position –1 and +1, this result was in good agree-ment with sequence alignment-based prediction of amino acidcontacts between the peptide substrate side chains and residues inthe active site of PKCβ isoenzymes.19
Design of the optimal peptide substrate for PKC
By combining the most favorable amino acids in each positionthat flanked the serine, we designed an optimal peptide structure,FRRRRSFRRR, and tested it as a substrate for PKCβ. Thephenylalanine in position +1 was included instead of arginine to re-duce the net positive charge and because it did not appreciably
Loog et al.
324 www.sbsonline.org Journal of Biomolecular Screening 10(4); 2005
329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 m/z0
100
%
0
100
%
344.2
329.9340.4333.4 343.9
347.9
347.7
352.4354.7
352.4
343.9
340.4329.9333.4
347.7
354.7R
G A V N
F
K
G A V
I
N
IK E
FR
E
Phosphopeptide calibration XLRLHSLRER
Initial velocity conditions
347.9344.2
0
329.9344.2
343.9 352.4
100
%
0354.7
347.9
VN
E
329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358m/z
347.7
332.4 334.7340.4333.4
G A F
KI
100
%
344.2
343.9333.4329.9
332.4
340.4
334.7
347.9
347.7
354.7
352.4
R
G A VN
IK
E
F
R
LXRLHSLRERPhosphopeptide calibration
Initial velocity conditions
FIG. 2. Examples of initial velocity spectra and phosphopeptide cali-bration spectra, for +4 charged components of libraries XLRLHSLRER(A) and LXRLHSLRER (B). Each letter designates a peptide with the cor-responding amino acid in the varied position. The components of these 2libraries have the same molecular masses and show very similar spectralpatterns when fully phosphorylated (phosphopeptide calibration panel).In contrast, profoundly different peak heights are observed for the individ-ual peptide components in the initial velocity spectra, showing strong se-quence specificity of protein kinase C in these 2 adjacent positions, withthe optimal amino acids determined to be phenylalanine and arginine. The2 undesignated peaks on the left side of the spectra correspond to traceamounts of the nonphosphorylated library components that remain boundto the Fe-chelating column.
B
A

change the specificity of substrate peptide (Fig. 3). PKCβ showedvery high activity toward the new peptide with a Km value in theconcentration range lower than 100 nM (Table 1). It is not possibleto accurately measure the exact value of Km at such a low concen-tration range. The new substrate has at least 2-fold higher specific-ity for PKC than the best known peptide substrate, thepseudosubstrate-based peptide RFARKGSLRQKNV. It was alsofound that the Vmax values of the 2 peptides were in comparablerange and that the combined specificity determinants contributedmainly to the value of Km (Table 1). To test the relative specificityof these peptides, a phosphorylation assay of a mixture of 3 pep-tides, FRRRRSFRRR, RFARKGSLRQKNV, and the templatepeptide LLRLHSLRER, was performed using the ESI-MSmethod for phosphopeptide detection. As shown in Figure 4, thenewly designed peptide was phosphorylated much more effi-ciently and suppressed the phosphorylation of the other 2 specificsubstrates in the 1st phase of the reaction. Such delay in the sub-strate mixture turnover can happen when 1 of the substrates have avery high (1+ [S]/Km) factor and it acts as a competitive inhibitorrelatively to others until it has been mostly consumed. Such
behavior is an additional indicator of a very high affinity of thenewly designed peptide.
Substrate specificity-based design ofpseudosubstrate inhibitor for PKC
Next, we tested if the obtained quantitative specificity profilecan be used for the design of pseudosubstrate inhibitor structures.Pseudosubstrate inhibitors are generally the substrate analogs withthe phosphoacceptor amino acid replaced by alanine. We designeda new peptide by replacing the serine in FRRRRSFRRR withalanine to create a highly efficient pseudosubstrate inhibitor with aKi value of 76 nM. It was shown that the action mode of the newcompound was competitive in respect to both peptidesLLRLHSLRER and FRRRRSFRRR (Figs. 5A, 5B, respectively),suggesting the pseudosubstrate mechanism of inhibition. It alsoconfirms that the high efficiency of the peptide substrateFRRRRSFRRR is not due to some undefined activator effect re-sulting from binding to some other part of the enzyme moleculethan the active site substrate binding region.
Optimal Specificity Profile of Protein Kinase C
Journal of Biomolecular Screening 10(4); 2005 www.sbsonline.org 325
G A V I L N K E F R0
50
100
150
200
250
300
G A V I N K E F R0
10
20
30
G A V I N K E F R0
100
200
300
400
500
600
700
G A V I N K E F R0
10
20
30
103
k II
(l/m
in/m
g)
Position +4
103
k II
(l/m
in/m
g)
Position +3
103
k II
(l/m
in/m
g)
Position +2
G A V I L N K E F R0
25
50
75
100
103
k II
(l/m
in/m
g)
Position +1
G A V I H N K E F R0
25
50
75
100
125
150
103
k II
(l/m
in/m
g)
Position -1
103
k II
(l/m
in/m
g)
Position -2
G A V I N K E F R0
10
20
30
103
k II
(l/m
in/m
g)
Position -3
G A V I L N K E0
25
50
75
100
125
103
k II
(l/m
in/m
g)
Position -4
G A V I L N K E F R0
25
50
75
100
12510
3k
II (
l/min
/mg)
Position -5
Figure 3
FIG. 3. Positional specificity profiles for protein kinase C β obtained from the library phosphorylation experiments. The quantitative patterns ofthe 2nd-order rate constant kcat/Km (designated as kII) were determined from initial velocity measurements of the phosphorylation reaction using theelectrospray mass-spectrometric method as described in the Materials and Methods section. The values presented in the figure were calculated usingthe kinetic data of the separately measured values of kcat/Km for peptides containing arginine in position X, as shown in Table 1.

DISCUSSION
Our study shows that ESI-MS can be successfully applied forquantitative screening of peptide libraries in protein kinase studies.ESI-MS analysis has very high quantitative detection sensitivity,allowing for the detection of each single phosphopeptide in a li-brary mixture under initial velocity conditions. This can even per-mit the measurement of phosphopeptide formation in the pico-molar concentration range. The ability to study the effects ofamino acid variations in kinetic detail offers a great advantage overthe relatively unquantitative random libraries containing a largenumber of components.
The novel peptide library approach is based on the use of an ef-ficiently phosphorylated template peptide, which contains theminimal consensus motif for recognition by the kinase. This strat-egy enabled us to characterize the structural requirements foramino acid side chains in each position of the substrate under con-ditions, where the remaining unvaried portion of the peptide pro-vides a sufficient number of specificity determinants to maintainefficient phosphorylation. Such a positionally oriented approachallowed us to correctly determine the kinetic constants for individ-ual substrates, even when very detrimental substitutions wereintroduced at the varied positions.
Based on several previous studies, one can define a minimalconsensus sequence for PKC as R/KXXS/TXR/K,1,2,16,19 which
points out the most important specificity determinants as the basicresidues in positions –3 and +2. Although the experiments pre-sented here confirmed the importance of this minimal motif, it wasalso found that other positions in the vicinity of the phosphory-lation site had strikingly sensitive specificity requirements. Recentstudies have demonstrated that primary structure specificity deter-minants may provide useful information for the prediction of invivo substrates of protein kinases using protein sequence data-bases.20-22 However, it is not clear if a minimal consensus motif issufficient for such predictions or whether it is necessary to employa detailed and quantitative specificity profile of the sequence sur-rounding the phosphoacceptor amino acid. Therefore, it will be ofbenefit to determine more precisely how often the minimal con-sensus sequence elements and the additional specificity determi-nants in other positions are present around the phosphorylation
Loog et al.
326 www.sbsonline.org Journal of Biomolecular Screening 10(4); 2005
0
5
10
15
20
25
30
35
40
45
0 10 20 30 40 50
Time (min)
ytisnet ni l art ceps evit al eR
FIG. 4. Relative specificity of substrate peptides of protein kinase Cdemonstrated in a phosphorylation assay of a mixture of FRRRRSFRRR,RFARKGSLRQKNV, and LLRRHSLRER, using the electrospray mass-spectrometric method for phosphopeptide detection. The newly designedpeptide FRRRRSFRRR was phosphorylated much more efficiently andsuppressed the phosphorylation of the other 2 specific substrates in the 1stphase of the reaction. � = FRRRRSFRRR; � = RFARKGSLRQKNV;� = LLRRHSLRER.
0
0.5
1
1.5
2
-0.025 0 0.025 0.05 0.075 0.1
0
0.5
1
1.5
2
2.5
3
3.5
-0.05 0 0.05 0.1 0.15 0.2 0.25
A
B
1/[Peptide] ( µM-1)
1/v
opm
ole-
1 x m
in
1/v
opm
ole-
1 x m
in
1/[Peptide] ( µM-1)
FIG. 5. Inhibition of protein kinase C by the pseudosubstrate peptideFRRRRAFRRR. (A) Lineweaver-Burk plot for inhibition of phosphory-lation of substrate peptide LLRLHSLRER.�= no inhibitor;�= 100 nMof FRRRRAFRRR. (B) Lineweaver-Burk plot for inhibition of phos-phorylation of the newly designed substrate peptide FRRRRSFRRR bydifferent concentrations of the inhibitor peptide. � = 30 µMFRRRRAFRRR; � = 100 µM FRRRRSFRRR; � = 300 µMFRRRRSFRRR.

Optimal Specificity Profile of Protein Kinase C
Journal of Biomolecular Screening 10(4); 2005 www.sbsonline.org 327
sites in established protein substrates of PKC. We analyzed the 9surrounding positions of PKC phosphorylation sites from proteinsequences retrieved from the phosphoprotein databasePhosphoBase (http://www.cbs.dtu.dk/databases/PhosphoBase/)described in Kreegiupuu et al.23 and Blom et al.24 Every uniqueserine or threonine shown in the database to be a bona fide PKCphosphorylation site was categorized for the presence of the 2
main specificity elements of the minimal consensus sequence R/KXXS/TXR/K and also for the presence of additional specificitydeterminants shown in the full specificity profile B/R/K-R/K-R/K-R/K-X-S/T-B/R/K-R/K-R/K-R/K. Taken together, the resultsshown in Table 2 reveal the surprising result that only 20% of theknown 173 substrate sites possess the full minimal consensus se-quence R/KXXS/TXR/K. Furthermore, 26% of all the sites did notcontain this motif at all. Of these sites, the majority had 2 or moreadditional specificity determinants. Additional determinants werealso important in cases in which full or partial consensus motifswere present.
It can be concluded from our analysis that additional to the con-sensus sequence, the specificity elements determined by the ESI-MS method are also used in natural protein phosphorylation sites,and for prediction of these sites from the sequence databases, thewhole specificity matrix has to be used. Availability of such de-tailed specificity matrices will enable the identification of potentialsites for phosphorylation site-to-alanine mutagenesis, an approachwidely used for studying the functionality of protein phosphory-lation. In addition, such information can be very useful for assign-ing candidate kinases for in vivo phosphorylated sites identified inmass-spectrometric analysis of protein digests.
Generally, the requirement of the initial velocity measurementis a limitation in the methods of library mixtures because as thenumber of the components increases, the detection threshold low-ers. However, in the present study, we have successfully demon-strated, despite the few limitations in the library design, such as thepossible spectral overlap that decreases the number of availablevariations, that ESI-MS is an excellent and highly sensitive tech-nique for such analysis, because the phosphopeptide detection waslinear down until the picomolar range and theoretically allows thismethod to be applied to mixtures with much larger componentnumbers.
By combining the optimal specificity determinants for PKCaround the serine residue, a peptide structure with the highestknown specificity so far known for PKC was created; furthermore,the serine to alanine substituted analog of this peptide behaved as apseudosubstrate inhibitor with a nanomolar Ki value. Interestingly,highly basic stretches of amino acids can be found in several pro-teins from general protein databases indicating that the highly ba-sic nature of the designed peptides is not something entirelynonphysiological.
We propose that this positionally oriented ESI-MS approachimproves the potential for design of substrate site–directed peptideor peptidomimetic inhibitors for protein kinases in general. Highlyspecific and selective substrates could be designed for novelapplications such as high-throughput protein kinase activityscreens on protein kinase chips.25-27 The potent pseudosubstrateinhibitors can be linked to membrane-permeant signal peptidesfor modulation of protein kinase activities in vivo, as described inrecent studies.28,29
Table 2. Analysis of the Frequency of the ConsensusSequence and Additional Specificity Determinants in thePhosphorylation Sites of Established Protein Substrates
for Protein Kinase C
Full specificity profile –5 –4 –3 –2 –1 +1 +2 +3 +4B/R/K-R/K-R/K-R/K-X-S/T-B/R/K-R/K-R/K-R/K
Minimal consensus motif –3 –2 –1 +1 +2R/K-X-X-S/T-X-R/K
Substrate statistics
Number of AdditionalPos–3 R/K Pos+2 R/K Determinants Number of Sites
+ + 6 –+ + 5 6+ + 4 7+ + 3 6+ + 2 8+ + 1 7+ + 0 1
35 (20.2% of total)
+ – 6 –+ – 5 2+ – 4 9+ – 3 9+ – 2 14+ – 1 6+ – 0 4
44 (25.4% of total)
– + 6 –– + 5 –– + 4 3– + 3 16– + 2 12– + 1 14– + 0 4
49 (28.3% of total)
– – 6 –– – 5 2– – 4 4– – 3 11– – 2 17– – 1 9– – 0 2
45 (26.0% of total)
The 9 amino acid positions surrounding the phosphorylated residue were analyzed in pro-tein kinase C phosphorylation sites retrieved from the phosphoprotein databasePhosphoBase (http://www.cbs.dtu.dk/databases/PhosphoBase/) described by Dostmannet al.
23,24Every unique serine or threonine was categorized for the presence of the 2 main
specificity elements of the consensus sequence and also for the presence of additional spec-ificity determinants shown in the full specificity profile (B denotes a large hydrophobicamino acid).

ACKNOWLEDGMENTS
This work was supported by the EU INCO Copernicus grant IC15-CT96-0900; Swedish Medical Research Council grant K98-03X; Swedish Agricultural and Forestry Research Board grant729.1181/97; grant KAW 1998.0028; Estonian Science Founda-tion grants ETF 3916, 5214, 5479; and the Estonian Ministry ofEducation and Science Grant 2592. We thank Dr. David Randle forcritical comments on the article.
REFERENCES
1. Kennelly PJ, Krebs EG: Consensus sequences as substrate specificity deter-minants for protein kinases and protein phosphatases. J Biol Chem
1991;266:15555-15558.
2. Kreegipuu A, Blom N, Brunak S, Jarv J: Statistical analysis of protein kinasespecificity determinants. FEBS Lett 1998;430:45-50.
3. Songyang Z, Cantley LC: The use of peptide library for the determination ofkinase peptide substrates. Methods Mol Biol 1998;87:87-98.
4. Turk BE, Cantley LC: Peptide libraries: at the crossroads of proteomics andbioinformatics. Curr Opin Chem Biol 2003;7:84-90.
5. Wu JJ, Phan H, Lam KS: Comparison of the intrinsic kinase activity and sub-strate specificity of c-Abl and Bcr-Abl. Bioorg Med Chem Lett 1998;8:2279-2284.
6. Till JH, Annan RS, Carr SA, Miller WT: Use of synthetic peptide libraries andphosphopeptide-selective mass spectrometry to probe protein kinase sub-strate specificity. J Biol Chem 1994;269:7423-7428.
7. Huyer G, Kelly J, Moffat J, Zamboni R, Jia Z, Gresser MJ, et al: Affinity se-lection from peptide libraries to determine substrate specificity of protein ty-rosine phosphatases. Anal Biochem 1998;258:19-30.
8. Lindblom S, Ek P, Muszynska G, Ek B, Szczegielniak J, Engstrom L:Phosphorylation of sucrose synthase from maize seedling. Acta Biochim Pol1997;44:809-817.
9. Loog M, Toomik R, Sak K, Muszynska G, Jarv J, Ek P: Peptidephosphorylation by calcium-dependent protein kinase from maize seedlings.Eur J Biochem 2000;267:337-343.
10. Kaiser E, Colescott RL, Bossinger CD, Cook PI: Color test for detection offree terminal amino groups in the solid-phase synthesis of peptides. AnalBiochem 1970;34:595-598.
11. Parker PJ, Marais RM: Purification of protein kinase C isotypes from bovinebrain. Methods Enzymol 1991;200:234-241.
12. Ferrari S, Thomas G: Micro- and macropurification methods for protein kin-ases. Methods Enzymol 1991;200:159-169.
13. Muszynska G, Dobrowolska G, Medin A, Ekman P, Porath JO: Model studieson iron(III) ion affinity chromatography: II. Interaction of immobilizediron(III) ions with phosphorylated amino acids, peptides and proteins. JChromatogr 1992;604:19-28.
14. Segel IH: Enzyme Kinetics. New York: John Wiley & Sons.
15. Fersht A: Enzyme Structure and Mechanism. New York: W. H. Freeman.
16. Pinna LA, Ruzzene M: How do protein kinases recognize their substrates?Biochim Biophys Acta 1996;1314:191-225.
17. Hanks SK, Hunter T: The protein kinase superfamily. In Hardie DG, HanksSK (eds): The Protein Kinase Factsbook. London: Academic Press, 1995:7-47.
18. Songyang Z, Lu KP, Kwon YT, Tsai LH, Filhol O, Cochet C, et al: A struc-tural basis for substrate specificities of protein Ser/Thr kinases: primary se-quence preference of casein kinases I and II, NIMA, phosphorylase kinase,calmodulin-dependent kinase II, CDK5, and Erk1. Mol Cell Biol 1996;16:6486-6493.
19. Nishikawa K, Toker A, Johannes FJ, Songyang Z, Cantley LC: Determinationof the specific substrate sequence motifs of protein kinase C isozymes. J BiolChem 1997;272:952-960.
20. Songyang Z: Analysis of protein kinase specificity by peptide libraries andprediction of in vivo substrates. Methods Enzymol 2001;332:171-183.
21. Obenauer JC, Cantley LC, Yaffe MB: Scansite 2.0: proteome-wide predictionof cell signaling interactions using short sequence motifs. Nucleic Acids Res2003;31:3635-3641.
22. Manning BD, Cantley LC: Hitting the target: emerging technologies in thesearch for kinase substrates. Sci STKE 2002:PE49.
23. Kreegipuu A, Blom N, Brunak S: PhosphoBase, a database ofphosphorylation sites: release 2.0. Nucleic Acids Res 1999;27:237-239.
24. Blom N, Kreegipuu A, Brunak S: PhosphoBase: a database ofphosphorylation sites. Nucleic Acids Res 1998;26:382-386.
25. Williams DM, Cole PA: Kinase chips hit the proteomics era. Trends BiochemSci 2001;26:271-273.
26. Houseman BT, Mrksich M: Towards quantitative assays with peptide chips: asurface engineering approach. Trends Biotechnol 2002;20:279-281.
27. Houseman BT, Huh JH, Kron SJ, Mrksich M: Peptide chips for the quantita-tive evaluation of protein kinase activity. Nat Biotechnol 2002;20:270-274.
28. Dostmann WR, Taylor MS, Nickl CK, Brayden JE, Frank R, Tegge WJ:Highly specific, membrane-permeant peptide blockers of cGMP-dependentprotein kinase Ialpha inhibit NO-induced cerebral dilation. Proc Natl Acad
Sci U S A 2000;97:14772-14777.
29. Dostmann WR, Tegge W, Frank R, Nickl CK, Taylor MS, Brayden JE. Ex-ploring the mechanisms of vascular smooth muscle tone with highly specific,membrane-permeable inhibitors of cyclic GMP-dependent protein kinaseIalpha. Pharmacol Ther 2002;93:203-215.
Address reprint requests to:Mart Loog
Department of PhysiologyGenentech Hall, Box 2200
University of California600 16th Street
San Francisco, CA 94143-2200
E-mail: [email protected]
Loog et al.
328 www.sbsonline.org Journal of Biomolecular Screening 10(4); 2005




















