
Beta-specificity: The turnover of host species in space and another way to measure host specificity
Upload
independentCategory
view
8download
0
AllCHIVES OF BIOCHEMISTRY .\ND BIOPHYSICS 137, 483-493 (1970)
Substrate Specificity of Pullulanase’
MUKHTAR ABDULLAH2 AND DEXTER FRENCH
Department of Biochemistry and Biophysics, Iowa State University, Ames, Iowa 50010
Received February 25, 1969; accepted January 8, 1970
Action of pullulanase from Aerobacter aerogenes has been tested on a number of starch oligosaccharides. Hydrolysis is confined to cleavage of a-1 -+ 6-int,erchain links between oligosaccharide chains containing a minimum of two glucose units per chain. Starch oligosaccharides and dextrins containing two or more a-1 + 6-links are also cleaved. Glucose has not been observed as a product. A single (~-1 ---t 6-linked glucose “stub” does not interfere with pullulanase action, and oligosaccharides con- t.aining such st,ubs are found in the products of pullulanase action. Under special conditions, pullulanase preparations exhibit condensation and transferase reactions.
Pullulanase (bacterial R-enzyme, EC 3.2.1.9, from Aerobacter aerogenes) speci- fically cleaves certain a-l --f 6-links in starch, glycogen, pullulan, and oligosac- charides related to these polysaccharides (l-3). The potential value of pullulanase in complex saccharide structure analysis and enzymology depends on our knowledge of its specificity. In this research we have examined the action of pullulanase prepa- rations on various oligosaccharides and dextrins derived from starch and glycogen. Previous workers showed that pullulanase (Aerobacter debranching enzyme) action occurs at a-l +6-links between malto- oligosaccharide chains provided each chain contains a minimum of two a-1 + 4-linked glucose units. Our qualitative results cor- roborate these findings. We find also that oligosaccharides containing a reducing iso- maltose moiety are cleaved provided that both glucose units of isomaltose are sub- stituted at position 4 by one or more glu- cose units. Action of pullulanase is reversible,
1 Journal Paper No. J-5645 of the Iowa Agricul- tural and Home Economics Experiment Station, Ames, Iowa, Project No. 1116, Supported in part by grants from t,he Corn Industries Research Foundation, Agricultural Research Service 12-14. 100-7688 (71), and US Public Health Service (GM-08822).
2 Present address: Corn Products Co., Argo, Illinois.
and at high substrate concentrations con- densation reactions occur (4). Thus, G$ can be condensed through an a-1 -+6-link to Gz, G,, amylose, or Schardinger dextrins. Reduction of the glucose unit at the reduc- ing terminus of a branched oligosaccharide to a sorbitol unit may partially inhibit the characteristic hydrolytic activity of pullu- lanase, and in some cases transferase ac- tivity has been observed.
A recent report by Walker (5) describes a pullulanase from cell extracts of Strepto- coccus mitis. This enzyme is similar in many respects to the Aerobacter enzyme but differs in being much less active on small oligosaccharides and in failing to show thermal stability or reversible thermal, inactivation (6).
3 Abbreviations: Gz, Ga, etc., for maltose, maltotriose, etc.; B4, B,, etc., for “branched” oligosaccharides containing a single a-1 --) 6.link in an otherwise a- 1 + 4-linked t,et.rasaccharide, pentasaccharide, etc.; BBs, etc., for “doubly branched” oligosaccharides containing two 01- 1+ 6-linksin a hexasaccharide, etc.Useof the term “branch” connotes an a-1 + g-linked offshoot, from the main, or-1 --) 4-linked chain, rather than necessarily a bifurcation. In t,his sense one could have a branch at either end of a main chain. “Fast Bb” and “slow B5” are 63-a-glucosyl- maltotetraose and 63-a-maltosylmaltotriose, re- spectively. The designations fast and slow refer to appreciable differences in their chromato- graphic mobility (2).
483

484 ABDULLAH AND FRENCH
EXPERIMENTAL
Paper chromatography. The high-temperature, multiple ascent method (7) was used. Solvents employed were water:pyridine: butanol (3:4:6 or 4:4:6 parts by volume) or aqueous propanol (7076 by volume). The propanol solvent was useful to give rapid and clean separation of linear oligo- saccharides. However, the pyridine-containing solvents gave superior discrimination between a-l + 4- and or-1 -+ 6-linked oligosaccharides. The retardation by an or-1 -+ 6-link was about. one-half to two-thirds that of an additional a-1 --t /l-linked n-glucose unit. Most compounds were detected using the silver dip method (8). High concentra- tions of the nonreducing oligosaccharide alcohols and Schardinger dextrins gave unusual spots with a dark gray rim around the border and a light gray center. With low concentrations, only light gray spots were observed. The Schardinger dextrins could also be detected by dipping in methanolic IrKI solution (10 ml of aqueous 0.2% 12, 2% KI plus 200 ml of methanol). The 01 dextrin gave a blue or purple spot; the p and y dextrins gave yellow-brown spots.
Two-dimensional chromatography: To survey the action of pullulanase on oligosaccharide mix- tures, the two-dimensional chromatographic method (9) was employed. The oligosaccharide mixture was applied near one corner of a sheet of chromatography paper and subjected to multiple ascents (usually 24). The area containing oligo- saccharides was sprayed until damp (but not run- ning wet) with a pullulanase solution and allowed to incubate in a damp atmosphere for from 30 min to overnight. After drying and spot’ting with a reference mixture, the chromatogram was again subjected to multiple-ascent. chromatography, and the carbohydrates were revealed by the silver dip method.
Enzymes: Pullulanase was prepared by the method of Bender and Wallenfels (1) as modified by Abdullah et al. (3). We could not detect any or-amylase activity in the preparations. The ace- tone-dried powder assayed on pullulan (3) con- tained 195 U4 per g.
For reactions in solution, digests usually con- tained 5 mg of substrate and 0.25 U of pullulanase (dialyzed against pH 6.9 phosphate buffer) per milliliter. An undialyzed 1% water extract of the acetone powder was used as a spray reagent (2 U/ml). Aspergillus niger amyloglucosidase (Dex-
4 One unit (U) according to the International Union of Biochemistry is that amount of enzyme that, when acting at 30” and optimal pH and sub- strate concentration, will hydrolyze 1 pmole of glycosidic bonds per minute.
trinase 7159) was a gift from Miles Chemical Co., Elkhart,, Indiana. The enzyme was purified from small amounts of reducing sugars by passing through Sephadex G-25. The treated enzyme had excellent amyloglucosidase (a-l + 4) action but also showed low a-1 + 6-glucosidase activity, resembling the amyloglucosidase reported by Abdullah, Fleming, Taylor, and Whelan (lo), as well as transferase activity. For example, extended action on panose gave glucose, maltose, isomal- tose, isomaltotriose, 4-~-is0ma1t0tri0sy1g1uc0se, and other compounds. Action on panitol gave glucose and maltitol. Crystalline human salivary amylase was prepared as by Fischer and Stein (11). Crystalline sweet potato D-amylase prepared ac- cording to Balls, Walden, and Thompson (12) was purchased from Worthington Biochemical Corp., Freehold, New Jersey. Bacillus macerans trans- glycosylase was prepared by the method of Tilden and Hudson (13). Aspergillus oryzae enzyme mix- ture containing transferase (Clarase concentrate) was a gift of Miles Chemical Co., Elkhart, Ind. Crystalline Aspergillus oryzae amylase was pre- pared by the method of Toda and Akabori (14).
Isomaltose and panose coupling reactions: Chro- matographically pure isomaltose and panose were laboratory samples obtained by yeast fermenta- tion and charcoal chromatography of an A. oryzae enzyme digest of maltose. Coupling reaction de- tails were similar to those reported by French, Taylor, and Whelan (15), except that the reaction was conducted at 37” rather than at about 15”. Isomaltose (1 g) and cyclohexa-amylose (6 g) were treated with 84 Tilden units of B. macerans enzyme in a total volume of 20 ml for 5 days at 37”. The mixture was heated briefly to boiling to inactivate the enzyme and cooled. To remove cyclodextrin, 1 ml of 1, 1,2,2-tetrachloroethane was added. The mixture was shaken, stored at 0” for 2 days, and centrifuged. The clear supernatant was evaporated to dryness.
To the coupled reaction product (250 mg, 25 ml) was added 0.2 M pyridine acetate buffer pH 4.7 (4.9 ml) and P-amylase (0.1 ml, 828 U). After 24 hr at 37”, the digest was heated in a boiling water bath for 5 min. It was evaporated to dryness and fractionated by paper chromatography. The branched oligosaccharides were eluted from the paper, dissolved in Hz0 (2 ml), and t,reated with pyridine acetate buffer (1 ml.) and P-amylase (0.4 ml, 3640 U). After 48 hr at 37”, the digest was heated to inactivate the enzyme. Paper chroma- tography showed that the final mixture contained n-glucose, Gz, and branched oligosaccharides through Bs. A portion (0.2 ml, 1 mg) was examined by action of pullulanase (two-dimensional chro- matography; Fig. 1).

SUBSTKATE SPECIFICITY OF PULLULANASE 485
The H. maceruns coupling reaction with panose was carried out in a similar fashion except that pure crystalline panose was used in place of iso- maltose. In this case, action of amylases on the digests gave branched oligosaccharides through Bg (with salivary amylase) or B7 (with fl-amylase).
FIc+s-la., lb.
FIG. 1. Two-dimensional chromatogram show- ing action of pullulanase on P-amylase-trimmed isomaltose-coupled products. (a) Actual chromat- ogram; (b) idealized drawing showing chromato- graphic arrangement and identification of spot.s. All tInreacted components of the original mixture fall on a line, extending from the point of sample application to glucose (the line may be curved owing to difference between the first, and second sets of chromatographic irrigation). Any spots elsewhere indicate reaction with the sprayed enzyme. The B4 spot to t,he left, below the band of unreacted compomlds, is the tetrasaccharide mixture resulting from condensation of two mole- cules of maltjose (pullulanase Bo). Such a tetrasac- charide spot is Ilsually seen in chromatograms having a large amount of maltose.
Results of pullulanase action on the salivary amy- lase product are shown in Fig. 2.
Salivary amylase limit de&ins: Shellfish glyco- gen (Sigma Chemical Co., St. Louis, Missouri) was exhaustively treated with salivary amylase into t,he “second stage” region of Walker and Whelan (16). At this degree of hydrolysis, singly branched oligosaccharides are iti the range of B4 to Bi. Doubly branched oligosaccharides are mainly in the range BBS-BBI~, and there are also present resistant dextrins of high molecular weight and low chromatographic mobilit,y ( “macrodext,rins”) (17). Action of pullulanase was examined by the two-dimensional chromatographic technique using about 1 mg of the mixture (Fig. 3).
Trimmed salivary dextrins: Branched oligosac- charides obtained as just described were partially separated from o-glucose and GB by chromatog-
FIGS-2a., 2b FIG. 2. Two-dimensional chromatogram show-
ing action of pullulanase on salivary amylase- resistant panose-coupled products. (a) Act,ual chromatogram; (h) idealized drawing; t,he spot labelled BBS is probably a transfer product of B.,.

486 ABDULLAH AND FRENCH
raphy on Sephadex G-10 and then “t,rimmed” by treatment with Dextrinase. Singly branched oligo- saccharides were converted mainly into Bd, with small amounts of other oligosaccharides; doubly branched oligosaccharides were converted into a well-defined set consist,ing of BB,, BB,, and BBs. The dextrin mixture was submitted to pullulanase action on paper by the two-dimensional chromat- ographic technique (Fig. 4).
Salivary amylase macrodextrin: Macrodextrin was obtained from glycogen essentially as by Heller and Schramm (19) except that salivary amylase was used in place of pancreatic amylase. Following repeated precipitation with 66yo ethanol the macrodextrin showed no chromatographically visible oligosaccharides. The resistant macro- dextrin (5.6 mg) was treated with 0.1 ml of pullu- lanase solution for 24 hr at 37”. After deionization,
a portion of the digest was examined by one- dimensional chromatography (Fig. 5a). In a similar experiment, macrodextrin was simultane- ously treated with pullulanase plus an excess of P-amylase (Fig. 5b).
Pullulanase condensation reactions. Maltose and maltolriose: Chromatographically pure GZ in 5- 254” solution was t,reat,ed with pullulanase, and the reaction was followed by paper chromatog- raphy. With the higher substrate concentrations, a condensation product in the tetrasaccharide region (pullulanase-B& gradually increased in amount, and a very weak hexasaccharide spot appeared (BBC). When the amount of Bd appeared to be levelling off, the mixture was separated by charcoal chromatography, and t,he Ba was obtained in chromatographically pure form (yield, 3.4%). Similar reactions with G3 as substrate gave a hexasaccharide (pullulanase-Ba; yield, 2.17,).
- FIRST lRuIG*TNJN
/ @ @ O0 @ 0, ----- ________________ o- G? -
AREA SPRAYEO WITH POLLULANASE
P @> SAMPLE CONTROL
FIGS-3a., 3b.
FIG. 3. Action of pullulanase on limit dext.rins from action of salivary amylase on glycogen. (a) Actual chromatogram; (b) idealized drawing. The designations fBs and sBs refer to “fast Bs” and “slow BS” respectively.3
c FIRST IRRIGATION
I PROWCTS FROM
AREA SPRAYED WITH PUUl.MNASE
/@ @> SAMPLE CONTROL
FIGs4a., 4b.
FIG. 4. Action of pullulanase on Dextrinase- “trimmed” salivary amylase limit dextrins. (a) Actual chromatogram; (b) idealized drawing.

SUBSTRATE SPECIFICITY OF PULLULANASE 487
FIGS-~., 5b.
FIG. 5. Action of pullulanase on glycogen macro- dextrin. (a) Action of pullulanase only; (b) com- bined action of pullulanase and @-amylase. Note formation of small amounts of fl-amylase-resistant “stubbed” oligosaccharides.
Pullulanase condensation reaction of maltose plus amylose: GZ (177,) plus amylose (0.2y0) was treated with pullulanase (1.6 U/ml of digest) as in the preceding paragraph. After 72 hr at 37”, the reaction was terminated by heating to boiling, and the mixture was treated with 1, 1,2,2-tetrachlo- roethane to precipitate the least-modified amylose fraction. The precipitat,ed complex was removed by centrifugation. On incubation with @-amylase, it gave a limit of 76C0 malt.ose as opposed to the control amylose (treated with pullulanase in absence of Gz), which gave an 857, P-amylase limit. The supernatant and washings from the tetrachloroethane precipitation were treated with 2 volumes of 95yc ethanol to precipitate the poly- saccharide component. The precipitate was re- moved in the centrifuge and washed with several changes of 66% (by volume) ethanol. The resulting polysaccharide was essentially free from oligosac- charides, as judged by paper chromatography. Exhaustive reaction with Aspergillus oryzae amylase gave glucose and G2 together with small amounts of oligosaccharides identified chromato- graphically as B, and Bg with lesser amounts of Bq and B7. A control digest of untreated amylose gave no branched oligosaccharides.
Pullulanase condensation of maltose plus cyclo- hexa-amylose: Gz (177,) was condensed with cyclo- hexa-amylose (1.7yc) using pullulanase as in the
preceding paragraph. Reaction produced a new compound detected on paper chromatograms by the silver dip method (light gray spot). The enzyme was inactivated by heating to boiling, and the reaction mixture was treated with 1,1,2,2- tetrachloroethane at 3”. The precipitated complex was recovered by centrifugation, washed with tetrachloroethane-saturated water twice in the centrifuge, and redissolved by boiling in water. The precipitated and redissolved sample on paper chromatography now showed only two spots: unchanged cyclohexa-amylose and the condensa- tion product. On reacting with pullulanase the condensation product disappeared and gave strong G2 and cyclohexa-amylose spots.
Branched oligosaccharide alcohols: Oligosac- charide (10 mg) and sodium borohydride (10-20 mg) were dissolved in 1 ml of water and allowed to stand overnight. More sodium borohydride (lO- 20 mg) was added, and the mixture was allowed to react 4 hr. After neutralization with 3 N acetic acid to pH about 5-G, the solution was treated with Amberlit,e MB (carbonated), evaporated to dry- ness, and evaporated several times with methanol (to remove boric acid). Results of pullulanase action are shown in Fig. 6. Macrodextrin was reduced using sodium [3H] borohydride (New England Nuclear Co.) using essentially the same procedure on a small scale.
Pullulanase action on branched cyclic oligosac- charides: “Limit dextrins” were obtained by
FIG. 6 FIG. 6. Action of pullulanase on branched
oligosaccharide alcohols. (a) Reference compounds (glucose, Gz, Ga, etc.); (b) pullulanase Ba-itol; (c) pullulanase B&01; (d) pullulanase Be, sali- vary amylase-treated, then reduced; (e) salivary amylase Bb-itol; (f) salivary amylase Bg; (g) enzyme alone (control).

488 ABDULLAH AND FRENCH
AREA SPRAYED WITH PULLULANASE
SAMPL&&!WL
Fms--‘la., 7b.
FIG. 7. Pullulanase action on branched cyclic molecules. (a) Limit dext.rins from Schardinger dextrin preparation; (b) idealized drawing. Cycle-G6 (Schardinger or-dextrin) is indicated on the drawing in shaded circles; it is easily visible only with the iodine dip; only the products from Gz-cycle-G6, G3-cycle-G6, etc., are indicated.
action of B. maceruns transglycosylase on starch (18). After removal of Schardinger dextrins by complex-forming precipitants, the limit dextrins were precipitated by ethanol and dried. Action of pullulanase on the chromatographed material is shown in Fig. 7.
DISCUSSION OF RESULTS
Interpretation of the two-dimensional chromatograms. Recognition and applica- tion of several basic factors makes it possible to obtain a great deal of information from a
single two-dimensional chromatogram (9). All compounds that have not reacted with the sprayed enzyme during the reaction on paper fall along a single line (usually a cqrve). This line extends from the point of application of sample to whatever unre- acted compound present (usually glucose) has the highest Rf. All compounds deviating from this line are reaction products. It does not necessarily follow that all com- poundp remaining on the line are unreactive to the enzyme (for example, the enzyme to substrate ratio may be too low to give com- plete reaction), or that all reaction products have moved significantly from the line. It is possible to obtain approximate Rf values of a given reactant and its products, and by use of appropriate controls, in- ferences may be drawn regarding their molecular sizes and structures. Rf values in butanol-pyridine-water permit easy distinction between linear, singly branched, and dbubly branched oligosaccharides of given molecular size (19). Finally, the total number of glucose residues in the reactants must appear in the products. For example, if pullulanase action on a substrate in the heptasaccharide range gives Ba as one product, one should look for a trisaccharide as another product. Alternatively, if the reaction involved transferase, the other product might be a decasaccharide. Con- densation reactions lead to whole multiples of the original substrate (Gz -+ Bd, BBC, etc.).
Action of pullulanase cm isomaltose- and panose-coupled products. In the B. macerans transglycosylase coupling reactions, a glu- cose unit or chain of maltosidically linked glucose units becomes attached by an a-1 + 4-link to an appropriate acceptor (glucose, glucosides, or oligosaccharides). With iso- maltose, both glucose units can act as ac- ceptors. Similarly, with panose, both non- reducing glucose units can accept glucose or oligosaccharide chains. Thus by adding glucose units to the a-1 + 6-linked glucose units of isomaltose or panose, it is possible
to build up “branched” oligosaccharides of interest as test substrates for pullulanase. Such coupling products consist of mixtures of homologous and isomeric oligosac-

SUBSTRATE SPECIFICITY OF PULLULANASE 489
I NO ACTION
. . . . - 3 . . . . O-3 9 ---9 . . . .
73
a
. . . . 3 -9 ------o--4 SCHEME 1
charides, and except in the simplest cases, it is not feasible to isolate individual com- pounds. However, the higher homologs can be “trimmed” (for example, by amylases) to give well-defined sets of compounds. Such sets are more suitable as test sub- strates for pullulanase, since relatively few products are produced and the reactions can be readily identified. Results with beta- amylase trimmed isomaltose-coupled prod- ucts are shown in I”ig. 1 and interpreted in Scheme 1.” These results indicate that
5 Symbolic formulas used are O,n-glucose units; -, a-1 + 4-link; 1, a-1 + 6-link; $3, reducing D- glucose unit; S, sorbitol (o-glucitol) unit; hori- zontal dotted lines, points of cleavage by pullu- lanase.
pullulanase can act on oligosaccharide configurations in which the a-1 + Clink is to the reducing n-glucose unit of the chain, as suggested by Abdullah et al. (3).
Results with “trimmed” panose-coupled products are shown in Fig. 2 and Scheme 2.
The results of the isomaltose coupling with the B. macerans transferase show definitively that coupling occurs at both glucose units in isomaltose, as previously indicated (20). Failure to obtain significant coupling at the reducing glucose unit of isomaltose in the experiments of French
4 NO ACTION
74 . . . . -- & Fig+--?&
. . . . 24 -kJ+---&
z---z SCHEME 2

490 ABDULLAH AND FRENCH
A. Branching configurations resistant to B. Branching configurations resistant. to human salivary amylase and sweet- sweet-potato beta amylase but non- potato beta amylase. resistant to human salivary amylase.
SCHEME 3
L -84 . . . . b-a 6
ii-9
. . . . -
SCHEME 4
et al. (15) must be attributed to differences in reaction conditions used (specifically, the relatively low temperature of reaction) which led to the precipitation of an in- soluble polysaccharide and, possibly, differ- ences in the relative amounts of transferase and hydrolase in the B. macerans enzvme
The product distribution in the amylase- treated coupling products of isomaltose and panose confirms the limiting specificities of amylases on the distal (nonreducing) side of a-1 -+ 6-branch points (Scheme 3).
Action of pullulanase on the salivary amylase limit dextrins frona glycogen. Action of pullulanase on various resolved and partly resolved components of a salivary amylase-glycogen digest is shown in Fig. 3. With singly branched substrates, speci- ficity conforms to the general pattern es- tablished previously (2, 3). From more
complex dextrins in the range of 9-13 glucose units (BBg-BBr3)” pullulanase gave large amounts of Gz, Ga and Gh, with minor amounts of unidentified higher molecular weight products. The macrodextrins, con- taining about 30 glucose units and having an average chain length of about 4, are also essentially completely degraded to Gz, Ga, and Cd, with much smaller amounts of higher oligosaccharides. At the “second stage” of cr-amylolysis, where the macro- dextrins were isolated, the average length of A-chains is only about two glucose units and the exterior length of B and C chains is less than one glucose unit (16). The fact that pullulanase gave nearly total cleavage of such compounds indicates that compact- ness of branching per se does not block pullulanase action, provided that the ex- terior chains are not too long.
Action of Dextrinase (glucoamylase, syn. amyloglucosidase) on branched dextrins follows a specificity pattern (10) such that terminal a-l, 4-links are selectively cleaved in preference to terminal a-1, g-links. Careful regulation of the extent of reaction gives primarily terminally stubbed dextrins (21). Dextrinase treatment of the salivary amylase limit dextrins gave a mixture con- taining trimmed dextrins. Action of pullu- lanase on the chromatographed materials is illustrated in Fig. 4. The two-dimensional chromatogram is necessarily complicated, and the higher molecular weight substrates and products are poorly resolved. However, it is clear that BB,, BB,, and BB, are the principal doubly branched trimmed dextrins, and that these are converted by pullulanase
9 “\
--La (Bla; 86%) rather than that based on the
%” isomaltose structure (Bhb; 14 %) (22). Action of pullulanase on branched oliqo-
saccharide alcohols. Reduction of the free
\ 3 reducing group of an oligosaccharide pro-
64b vides a convenient method of identifying or of tritium-labelling such groups. Therefore, it is of considerable interest to examine pullulanase action and specificity on such compounds. Judging from qualitative re- sults, such as the two-dimensional chromn-
\
tographic method, reducing the terminal
\
n-glucose unit to a n-glucitol unit greatly
x 6gb
retards pullulanase action. This effect may be related to the polyol inhibition of gly- cosidases as reported by Kelemen and Whelan (23). For example, with conditions \ such that “slow B5” (Ccr-maltosylmalto-
66” triose) is completely degraded to Gz + Ga, the corresponding sugar alcohol is only slightly attacked. Moreover, as discussed below, a substantial amount of transfer activity occurs apparently according to the following reactions” :
AMYLOSE
SCHEME 5 Under similar conditions, a component of the pullulanase-coupled maltose product after reduction and pullulanase treatment
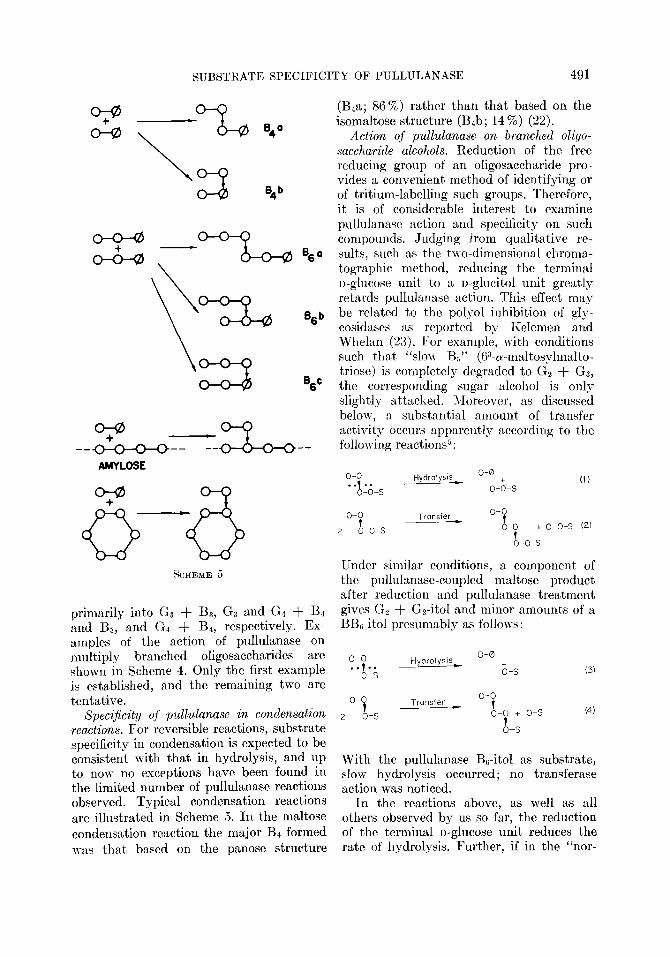
primarily into G, + B3, Gt and Gh + B, gives Gz + Gz-itol and minor amounts of a and BB, and Gq + Bq, respectively. Ex- BBC-it01 presumably as follows: amples of the action of pullulanase on multiply branched oligosaccharides are O-0 o-0
shown in Scheme 4. Only the first example . . . . is
Hydrolysis-
6-S
is established, and the remaining two are tentative.
Specificity of pullulanase in condensation 2 “-1-s
Transfer _ o-c
t
reactions. For reversible reactions, substrate o-0 + o-s (4)
b-s specificity in condensation is expected to be consistent with that in hydrolysis, and up to now no exceptions have been found in
Wibh the pullulanase B,-itol as substrate,
the limited number of pullulanase reactions slow hydrolysis occurred; no transferase action was noticed.
observed. Typical condensation reactions In the reactions above, as well as all are illustrated in Scheme 5. In the maltose others observed by us so far, the reduction condensation reaction the major Bk formed of the terminal n-glucose unit reduces the was that based on the panose structure rate of hvdrolvsis. Further. if in the “nor- _ ~~ ~ .,
SUBSTRATE SPECIFICITY OF PULLULANASE 491
0-o . . . . t
o-o-s
Hydrolysis _ o-0
0-h-s (1)
o-o TfOW+3 o-o
2 L-0-s
t 0-f + o-o-s (2)
b-o-s

492 ABDULLAH AND FRENCH
mal” hydrolysis reaction, Gz would be produced (from the distal or nonreducing side of the 1 + 6 bond), there is a tendency for 1,6 + 1,6 transfer reactions to occur, with disproportionation of the parent sub- strate. In fact it is possible to envision the ultimate hydrolysis reaction as proceeding through initial transfer reaction as follows:
O-O 1 Transfer o-
2 o-s e -7 + o-s (5)
6-S
o-o
-5 o-s
o-0 Hydrolysis
- + o-o (6)
x-s
of B. macerans transglycosylase on various branched starches or dextrins gives, in addition to variable amounts of the purely a-1 -+ 4-linked cyclic Schardinger dextrins, both “exe-branched” and “endo-branched” cyclic oligosaccharides (28) (see Scheme 6). Following removal of the cy-, p-, and y-dex- trins by use of selective complex-forming precipitants (such as 1 , 1,2,2-tetrachloro- ethane), the high molecular weight polysac- charides (limit dextrins) may be precipi- tated by ethanol, leaving branched cyclic oligosaccharides in solution. To the extent that the transglycosylase preparation con- tains hydrolytic activity, there will also be present various open-chain linear and branched oligosaccharides.
Since we have not used a crystalline pul- lulanase preparation (24, 25), it cannot be stated with certainty whether such transferase reactions are catalyzed by pullulanase itself (as opposed to a con- taminating enzyme). Actually these trans- ferase reactions are similar to those reported by Taylor and Whelan (26) using a sweet- corn enzyme preparation. On the other hand, the reactions are analogous to many other recognized and authenticated systems wherein a hydrolytic enzyme is also capable of effecting transferase reactions. A case in point is the transferase action of lysozyme acting on low molecular weight oligosac- charides too small to be effective substrates for hydrolysis (27).
Action of B. macerans transglycosylase on “stubbed glucan” (partial hydrolysate of amylopectin, containing single glucose side chains or “stubs”) has been reported by Stark (21) to give considerable amounts of “stubbed” Schardinger dextrins, totally resistant to pullulanase but easily suscep- tibIe to amylo-l-+6-glucosidase. In principle, such stubbed substrates could also yield a variety of other endo-branched and exo- branched cyclic oligosaccharides, through re- peated transfer reactions.
In connection with the structure analysis of glycogen macrodextrin, all the reducing groups were converted into n-glucitol units using sodium [3H] borohydride. The reduced macrodextrin was then hydrolyzed by pul- lulanase, and the individual reduced oligo- saccharides were separated by paper chroma- tography. A time study of the rate of release of these reduced oligosaecharides, measured by counting 3H, showed that the release of Gn-itol was very slow, G4-itol somewhat slow, and higher reduced oligosaccharides (through G8-itol) were released relatively rapidly. These results show that the re- tarding effect of a sorbitol unit is most pronounced with C-chains of only two or three units in length.
The two-dimensional chromatogram (Fig. 7) shows the action of pullulanase on B. nzacerans limit dextrins. Of particular in- terest is the series of exo-branched cyclic molecules based on cyclohexa-amylose, with side chains of 2-6 glucose units clearly resolved, and still higher unresolved con- stituents. The chromatogram suggests the presence of comparable series, based on cyclohepta-amylose, cycle-octa-amylose, and higher cyclic compounds. As is expected,
Branched cyclic oliqosaccharides. Action SCHEME 6

SUBSTRATE SPECIFICITY OF PULLULANASE 493
such chromatograms do not show if cyclic molecules with single glucose side chain stubs are present. However, it is clear that with exo-branched cyclic substrates, pul- lulanase can remove side chains (A-chains) in the range of 2-8 glucose units and prob- ably higher.
In previous work (%), it was shown that endo-branched cyclic molecules were hy- drolyzed by pullulanase to give the cor- responding open-chain linear oligosac- charides (Scheme 6). Although endo- branched cyclic molecules have never been isolated, models of them are easily con- structed with as few as four glucose units. Hydrolysis (ring opening) of such com- pounds by pullulanase would be expected to decrease the chromatographic mobility to some degree. We therefore suggest that the spots below the line of unchanged com- pounds in l:ig. 7 result from ring opening of endobranched molecules in the range of G-S glucose units per molecule. The type of reaction shown in the lower half of Scheme G would be expected to be reversible. By the reverse reaction, pullulanase would form endo-branched cyclic molecules from straight chains with the same number of glucose units. Such ring closure would be expected t’o increase the chromatographic mobility. We suggest that some of the spots in l’ig. 7 immediately above the line of unreacted compounds result from such ring closure by pullulanase action.
ACKNOWLEDGMENT
The authors are grateful to Dr. Johu F. Robyt for assistauce dttring the investigation and prep- aration of the manuscript. Some of the experi- ments were carried out in collaboration with Mr. M. Aslam Khwaja and I)r. Walter Verhue.
REFERENCES
1. BESDER, H., AND WALLENFELS, K., Biochem. is. 334, i9 (1961).
2. BROIVN, n. H., ILLING~ORTH, B., AND KORN- FELD, It., Hioche?nislry 4, 486 (1965).
3. ABDULL~~H,M., CATLEY, B. J., LEE, E. Y. C., ROBYT, J., WALLENFELS, K., AND WHEI,.~N, W. J., Cereal Chem. 43, 111 (1966).
4. ABDULLAH, M., SND FRENCH, D., Nature (London) 210, 200 (1966).
5. WALKER, G. J., Biochem. J. 108, 33 (1968). 6. ABDULLAH, M., CATLEY, B.J., CUTHBERTSON,
W. F. J., AND WHEL.\N, W. J., Biochem. J. 100, 8 P (1966).
7. FRENCH, D., MANCUSI, J. L., ABDULLAH, M., AND BRAMMER, Cr. L., J. Chromatogr. 19, 445 (1965).
8. ROBYT, J., END FRENCH, Il., Arch. Biochem. Biophys. 100, 451 (1963).
9. FRENCH, D., PULLEY, A. O., ABDULL.4H, M., END LINDEN, J. C., J. Chromatogr. 24, 271 (1966).
10. ABDULL~~H, M., FLEMING, I. l)., TAYLOR, P. M., BND WHEL.~N, W. J., Biochem. J. 89, 35 P (1963).
11. FISCHEH, E. H., AND STEIN, E. A., Biochem. Prep. 8, 27 (1961).
12. BJLLLS, A. K., WALDEN, M. K., MXD THOMPSON, R. It., J. Biol. Chem. 173,9 (1948).
13. TILDEN, E. B., .\XD HUDSON, C. S., J. Bacler- iol. 43, 627 (1942).
14. ToD:~, H., A\~~ AK.\BOKI, S., J. Biochem. (Tokyo) 63, 102 (1963).
15. FRENCH, I>., T~YL~I~, P. hf., .~ND WHELSN, W. J., Biochem. J. 90, 616 (19G4).
16. WIILKER, G. J., .\XD WHEL.\N, W. J., Biochern. J. 76, 257 (1960).
17. HELLER, J., AND SCHHXVN, M., Biochim. Bio- phys. dcta 81, 96 (1964).
18. FRENCH, D., PULLEY, A. O., .~ND WHEL.IS, W. J., Die Stiirke 16, 280 (1963).
19. FRENCH,L).,INDWILD,G.M., J.Amer.Chem. Sot. 76,2612 (1953).
20. SIJMMER, R., L4~~ FRENCH, D., J. Biol. Chem. 222, 469 (1956).
21. ST.\IX, J. R., Biochem. J. 102, 27 P (1967). 22. KHW~ZJA, A., Unpublished M.S. thesis, Iowa
Stat,e University (1967). 23. KELEMEN, M. V., .\ND WHELAN, W. J., Arch.
Biochem. Biophys. 11’7, 423 (1966). 24. WM~LENFELS, K., BENDER, H., END RACHED,
I. It., Biochem. Biophys. Res. Commun. 22, 254 (1966).
25. W-~LLENFELS, K., .~XD RXHED, I. R., Biochem. Z. 344, 524 (1966).
26. T.~YLOR, P. M., .\ND WHELBX, W. J., Biochem. J. 96, 26 P (1965).
27. ItUPLEY, J. A., .\ND (;.~TEs, V., Proc. Kat. Acad. Sci. G. S. 67,49G (1967).
28. FRENCH, I)., PGLLEY, A. O., EFFENBERGER, J. A., ROUGVIE, M. A., AND ABDULLAII, M., ilrch. Biochem. Biophys. 111, 153 (1965).
Copyright © 2022 FDOKUMEN




















