
The Analogs of Furanyl Methylidene Rhodanine Exhibit Broad ...
Upload
independentCategory
view
0download
0
PESTICIDE BIOCHEMISTRY AND PHYSIOLOGY 18, 235-246 (1982)
A Carboxylesterase with Broad Substrate Specificity Causes Organophosphorus, Carbamate and Pyrethroid Resistance in
Peach-Potato Aphids (Myzus persicae)
ALAN L. DEVONSHIRE AND GRAHAM D. MOORES
Department of Insecticides and Fungicides, Rothamsted Experimental Station. Harpenden. Hertfordshire, Great Britaia
Received March 24, 1982; accepted June 15, 1982
Carboxylesterase E4. the enzyme previously shown to cause resistance to organophosphorus insecticides in peach-potato aphids, Myzus persicae, was purified and characterized by elec- trophoretic and enzyme kinetic techniques. Its insecticidal substrate specificity, determined by following the rate of recovery of esterase activity after inhibition by a range of acylating inhibitors, included a variety of carbamates and organophosphates, although the catalytic center activity for these substrates was low. Radiometric measurement of hydrolysis of the pyrethroid, permethrin, showed that E4, whether purified or in crude aphid homogenates, hydrolyzed the (1S)trans enan- tiomer rapidly but hydrolysis of the other three isomers could not be detected. Such absolute specificity for one enantiomer of a pyrethroid is rare. The rates of hydrolysis of the various insecticidal classes correlated well with the relative degrees of resistance to them, and no other resistance mechanisms have been detected. Although the enzyme is relatively inefficient in de- grading insecticidal esters, it is produced in very large quantity, accounting for approximately 3% of the total protein in very resistant aphids. Its effect is thus mediated not only by hydrolysis but also by sequestering a substantial proportion of a toxic dose of insecticide. It is effective in this respect because the molar amount present is similar to that of a lethal dose of insecticide. These results support earlier indirect evidence for ‘*overproduction” of E4, probably because of struc- tural gene duplication or amplification and have direct implications for strategies to delay the buildup of resistance or for developing synergists to overcome resistance.
INTRODUCTION
Peach-potato aphids (Myzus persicae) have developed resistance to many of the insecticides used to control them both in glasshouses and in the field. Although these aphids show cross-resistance between or- ganophosphorus, carbamate, and pyre- throid insecticides (1, 2), only one bio- chemical difference has been detected be- tween strains (3); resistant insects have a more active carboxylesterase (E4), which also hydrolyzes paraoxon (4). This in- creased activity is caused by the production of more enzyme, rather than the same amount of a mutant, more active form (4). The molar concentrations of E4 in aphids were shown to double between each of the
seven known resistant aphid variants, sug- gesting that this increased production of E4 is caused by a series of duplications of the structural gene for the enzyme, the most resistant variant having 64 times as many copies as the susceptible (5). The activity of the enzyme is readily estimated in indi- vidual aphids by a range of techniques (6), which have been used widely (7, 8), both in fundamental laboratory studies and by ag- ricultural advisers, to monitor the resis- tance of field populations.
The present work is aimed at establishing whether the variations in quantity of this one enzyme are responsible for the ob- served wide spectrum of resistance, and if so, which classes of insecticide are likely, from biochemical considerations, to be
235 0048-3575182/050235-12$02.00/O Copyright @ 1982 by Academic Press, Inc. All rights of reproduction in any form reserved.

236 DEVONSHIRE AND MOORES
most effective against resistant aphids, or which might be used as synergists because they inhibit the enzyme.
MATERIALS AND METHODS
Aphids. The origins and maintenance of the aphid clones used in this work have been described previously (9).
Chemicals. m-Cresyl n-propylcarbamate, m-cresyl iso-propylcarbamate, m-cresyl n-butylcarbamate, and O-methyl O-(2,5- dichloro-4-iodophenyl) N-iso-propylphos- phoroamidate and the insecticides O-ethyl 0-(2-chloro-4-bromophenyl) S-n-propyl phosphorothiolate (profenofos) and thio- dicarb (at that time, an experimental com- pound) were gifts from Dr. G. Voss, Ciba- Geigy, Basel, Switzerland.
O,S-Dimethyl S-(N-methylcarbamoyl- methyl) phosphorodithioate (isodimetho- ate) was synthesized from dimethoate by dealkylation with di-iso-propylphosphoro- dithioate (10) followed by alkylation of the silver salt of the monomethyl ester by methyl iodide. Methylation occurred pref- erentially, and exclusively, on the sulfur to give the isomeric form of dimethoate; it had NMR peaks (in CDCl,) at 6 2.4 (d, 3H, 16 Hz, CH,S), 2.8 (d, 3H, 5 Hz, CH,N), 3.5 (d, 2H, 20 Hz, CH,), and 3.9 (d, 3H, 14 Hz, CH,O).
The N-ethyl analog of aldicarb was syn- thesized by stirring 2-methyl-2-(methyl- thio)propionaldehyde oxime (3 mmol, from a sample kindly supplied by Dr. R. H. Bromilow), ethyl isocyanate (3.3 mmol), and triethylamine (50 ~1) in dry hexane (3.5 ml) at room temperature for 20 hr. The colorless crystalline product was re- moved and washed with dry hexane. Its NMR spectrum had peaks (in Ccl,) at 6 1.2 (t, 3H, 7 Hz, CH,CH,), 1.5 (s, 6H, C(CH,),), 2.0 (s, 3H, CH,S), 3.4 (dq, 2H, 6, 7 Hz, CHJ, 6.1 (broad s, lH, NH), and 7.7 (s, lH, CH=).
Other insecticides were analytical stan- dards supplied by manufacturers.
Radiochemicals. 1-14C-acid-labeled (50
mCi/mmol) and methylene-‘“C-alcohol- labeled (59.7 mCi/mmol) permethrin were gifts from Wellcome Research, Berkham- sted, United Kingdom. The (lRS)-cis and (lRS)-trans isomers of each were separated and purified by thin-layer chromatography (TLC) on Merck 0.25-mm Kieselgel 60 plates in benzene:carbon tetrachloride (1: 1) and, after eluting and respotting, in hex- ane:diethyl ether (9: 1).
The meth?/lene-14C-alcohol-labeled es- ters, (lR)tvans-permethrin and (lS)trans- permethrin were synthesized by the method of Elliott et al. (11) from the resolved acids (supplied by Dr. M. Elliott (12) as their salts with D(-)-1-p-nitrophenyl-2-dimethyl- amino- 1,3-propanediol) and [methylene- 14C]3-phenoxybenzyl alcohol (3-POBA, 59.7 mCi/mmol, prepared by Soderlund (13) by hydrolysis of [14C]permethrin). Free acids were prepared from their salts by acidifying and extracting into chloroform, and after drying, each (10 mg) was dissolved in dry benzene (0.5 ml) together with thionyl chloride (4 ~1) and pyridine (4 ~1). After 3 hr at room temperature, each mixture was added to dry benzene (0.5 ml) containing pyridine (4 ~1) and 3-POBA (40 PCi, equiv- alent to 0.67 pmol) and left for 16 hr at room temperature. TLC purification as described above gave yields based on radio- activity of 56% for (lR)trans-permethrin and 65% for (lS)trans-permethrin. The stereochemistry of each isomer was con- firmed by a simple discriminating dose bio- assay in which groups of 20 susceptible (strain 608) house flies were treated with 10 pg (3.4 x lo5 dpm) insecticide/fly in 1 ~1 acetone. The (IR) isomer killed all the flies, and the (IS) isomer none, as would be expected from their published toxicities ( 12).
[ 1,3-“HIDi-isopropylphosphorofluoridate (DFP), specific activity 3.0 Ci/mmol, was obtained from the Radiochemical Centre. Amersham, United Kingdom.
Enzyme preparations and assay. Crude aphid homogenates and purified E4 were prepared in 20 mM phosphate buffer, pH 7.0, as described previously (4) except that

BIOCHEMICAL CAUSE OF CROSS RESISTANCE OF Myzus prrsicoe 237
the concentrated enzyme from DEAE- cellulose was further purified by gel filtra- tion on Sephacryl S-200 Superfine in the above buffer containing 0.1 M sodium chloride to minimize adsorption effects.
Hydrolysis of naphth-l-y1 acetate by these preparations was monitored spec- trophotometrically by a method based on that of Rosenberry and Bernhard (14) which relies on the small difference between the uv spectra of naphth-l-y1 acetate and naphth-l-01. To prepare a calibration curve, stock solutions (10 n-&I) of substrate and product were prepared in acetone, mixed in various proportions, maintaining a com- bined concentration of 10 mh4, and then diluted loo-fold with buffer. The calibration curve measured at 234 nm against a similar dilution of 10 mM naphth-l-y1 acetate in buffer was linear. Subsequent enzyme as- says were restricted to a maximum absor- bance change of 0.26 eq to 10% conversion of naphth-l-y1 acetate to naphth-l-01. All assays were at 0.1 mM substrate because at significantly greater concentrations the blank absorbed too strongly. The rate at this concentration was converted to V,,, by multiplying by 2.3 (see Ref. 4). The technique is less sensitive than the diazo coupling assay used earlier (4) but has the advantage of being a continuous, rather than a destructive, point assay.
Hydrolysis of organophosphorus atzd cavbamate esters. Only radiometric meth- ods would be sensitive enough to measure the hydrolysis of organophosphorus and carbamate esters by E4 directly because of its very slow turnover of these substrates. Since a suitable range of high specific ac- tivity radiolabeled insecticides was not available, an alternative indirect approach was used (4) based on the generally ac- cepted reaction mechanism (15) between acylating inhibitors or substrates and es- terases:
k, k, EH + ABSEHAB- EA
k-,
k3 EA-EH + AOH
Hz0
where EH is active enzyme, AB is the in- hibitor or substrate, EHAB is a Michaelis complex between the two, and EA is acyl- ated enzyme. Acylation of the enzyme oc- curs rapidly and the rate-limiting step in the overall reaction sequence is the hydrolysis of the acylated enzyme to regenerate free enzyme. The first-order rate constant (kJ of this hydrolysis represents the catalytic center activity (or turnover number) of the en- zyme for a particular type of substrate ( 15).
The first-order rate constants were mea- sured as follows. E4 was incubated in buffer (1 ml) containing insecticide ( 10d3 M) and bovine serum albumin (0.25%, BSA) for 10 min to give more than 95% inhibition. The inclusion of BSA stabilized the enzyme and improved the linearity of the sub- sequent enzyme assays. The excess of in- hibitor was removed by passing the mixture through a column (2.5 x 35 cm) of Seph- adex G-25, and the acylated enzyme was collected in 25 ml of eluent; preliminary experiments with [14C]paraoxon and [3H]DFP established that this column completely separated E4 from unbound insecticide, with a volume of 30 ml between the end of the E4 peak and the start of the insec- ticide peak. As the enzyme activity recov- ered, samples (1 ml) were assayed at in- tervals with naphth-l-y1 acetate, and k, calculated by fitting (percentage remaining inhibited) to a standard exponential func- tion (4) using the maximum likelihood com- puter program (MLP) (16).
Methamidophos, a poor esterase inhib- itor in vitro, was oxidized by m-chloroper- benzoic acid (17) immediately before being incubated with E4.
Hydrolysis of I-‘“C-acid-labeled permeth- rin. [‘“Clpermethrin (cis or trans) in acetone was added to purified E4 or to aphid homogenates (5 mg/ml) in 20 mh4 phosphate buffer, pH 7.0, to a final con- centration of 1 ,uM permethrin and 1%

238 DEVONSHIRE AND MOORES
acetone and incubated at 25°C. At intervals, samples (300 ~1) were taken from the incu- bations and mixed well with 1 ml chloro- form:methanol (1:2) to give a monophasic solution and inhibit further enzyme activ- ity. The radiolabeled hydrolysis product, 3-(2,Zdichlorovinyl)-2,2-dimethylcyclopro- panecarboxylate, was estimated by a tech- nique based on that of Nicholson (18); chlor- oform (300 ~1) and buffer (150 ~1) were added to the inhibited enzyme in chloro- form:methanol and mixed well and the phases separated by centrifuging at 2000g for 2 min. After removing the organic phase, more chloroform (300 ~1) was added, mixed, and centrifuged and the organic phase again discarded. The radioactive acid remaining in the aqueous phase was deter- mined by liquid scintillation spectrometry in Rialuma (LKB).
Hydrolysis of methylene-‘“C-alcohol- labeled permethrin. Enzyme and substrate (1 PM) were incubated as described for acid-labeled permethrin, and the reaction stopped by adding samples (100 ~1) of the incubation to acetone (200 ~1) at -78°C. After warming to room temperature, a sample of the mixture (200 ~1) was spotted on the concentrating zone of silica gel 60 (Merck catalog No. 11798) TLC plates and developed in toluene (saturated with formic acid):diethyl ether (10:3). Radioactive zones were located by autoradiography and the plates then impregnated with Neatan (Merck). When dry, the radioactive areas were cut out, peeled from the glass, and counted in Rialuma containing 20% water. Under these conditions, the counting effi- ciency of 3-POBA decreased to 93% of that when counted in homogeneous solution in the cocktail whereas the counting efficiency of the esters was unaffected.
Electrophoresis. Individual aphids were homogenized in water (15 ~1) containing (by weight) sucrose (10%) and Triton X-100 (0.5%). and samples (10 ~1) were analyzed by electrophoresis on 7.5% polyacrylamide gel slabs in a buffer system based on that of Williams and Reisfeld (19). The composi-
tion of the reservoir buffer and running gel buffer were as described, but the homoge- nates were added to pockets cast in the spacer gel (2.5% acrylamide) which had the same buffer as the running gel (4). After 1.5 hr electrophoresis at 250 V, gels were stained for esterase activity as described previously (20) or fixed and stained for protein with Coomassie brilliant blue R-250.
Insect homogenates and purified E4 were also characterized by SDS electrophoresis as described by Laemmli (21), after de- naturing by heating at 60°C for 5 min in 2% SDS, 5% &mercaptoethanol, and 4.25 M urea.
Radioactivity was located by slicing gels (4), incubating each slice for 90 min at 50°C in 30 ~1 water and 270 ~1 NCS (Amer- sham/Searle) and then counting by liquid scintillation spectrometry in toluene con- taining 0.4% PPO and 0.01% POPOP, mixed 2: 1 with Triton X-100.
RESULTS AND DISCUSSION
The only biochemical change known to be associated with resistance of M. per- sicae to a wide range of insecticides is in- creased E4 activity. Other possible resis- tance mechanisms have been investigated including speed of cuticular penetration (22), detoxication by microsomal oxidation and glutathione transferases (23), and al- tered sensitivity of acetylcholinesterase (Devonshire, unpublished), but in all these respects, resistant and susceptible aphids were indistinguishable.
In the context of acetylcholinesterase, one of the naphth-l-y1 acetate-hydrolyzing esterases identified after electrophoresis has been classified as a cholinesterase be- cause it is eserine sensitive (24). However, we have shown that this enzyme does not hydrolyze acetylcholine, and it is therefore not acetylcholinesterase. The only acetyl- cholinesterase present on gels runs as a very sharp band with a mobility (R,) of 0.05, half that of El, the eserine-sensitive esterase (Devonshire, unpublished) and is

BIOCHEMICAL CAUSE OF CROSS RESISTANCE OF Myzus prrskae 239
detectable only by a very sensitive radio- metric assay (25).
Purification and Identity of E4
E4 was purified from homogenates of 5- to 10-g aphids (strain TlV, PirR or G6) by gel filtration and ion-exchange chroma- tography after which it was shown by SDS electrophoresis (Fig. 1) to behave as a single protein. Its mobility both as the native protein on Sephacryl S-200, and on SDS electrophoresis after digestion for 5 min at 60°C with SDS (2%) mercaptoetha- no1 (5%), and urea (4.25 M), indicate that the enzyme consists of a single polypeptide with a MW of 65,000.
To establish whether the SDS protein band corresponds to E4, a measured amount of the purified enzyme (35 pmol, calculated from its activity with naphth- l-y1 acetate as substrate and its established catalytic center activity (4)) was incubated with [3H]DFP (0.15 mM) to inhibit the enzyme irrevers- ibly by more than 99%. After removing excess free DFP on a Sephadex G-25 col- umn, the amount of bound “H was 96% of that expected, confirming the catalytic center activity previously determined (4) by “titration” with [l”C]paraoxon and that only the E4 in the purified preparation was bind- ing DFP. The inhibited enzyme from an identical incubation was next examined by SDS electrophoresis on gel rods. The gels were stained for protein, cut into slices (of approx 1 mm), and solubilized in NCS, and the 3H was determined. Radioactivity was located entirely in the only protein band detected confirming that this corres- ponds to E4. This preparation was used in all subsequent kinetic studies.
We concluded (4) from catalytic center activity measurements on E4 purified from different aphid clones that resistant insects have more E4 than susceptible aphids, rather than the same molar quantity of a more efficient mutant enzyme. We now have more direct evidence; Figs. 1 and 2 show gels stained for protein after electro- phoresis of aphid homogenates and purified
FIG. 1. SDS rlectrophoresis of aphid homogenates, purifed E4. and molecular bcvight standards. E.rperi- mental details are given in the text. The gel was stained for protein tvith Coomassie brilliant blue R-250. (a) The homogenate of susceptibk aphids (US/L). (b) The homogenate of resistant aphids (G6). (c) Molecular rrseight standards (Sigma, SDS-6H): I. nyosin (206.000); 2, P-gulactosidase (116.000); 3. phosphor.vlase B (97,400): 4, albumin (66.000); 5, oval- bumin (45,000); 6. carbonic anhj,drase (29,000). (d) purified E4.
E4 in the SDS buffer system and the Tris/ barbitone buffer system used previously for characterizing esterase activity. It is clear that the increased activity in the re- sistant clone is due to a larger amount of the E4 protein.
Hydrolysis of Organophosphorus and Carbamate Esters
E4 is extremely sensitive to inhibition; for example, 10 nM paraoxon inhibits it al- most completely in 10 min, even when the enzyme is “protected” by substrate (4). To
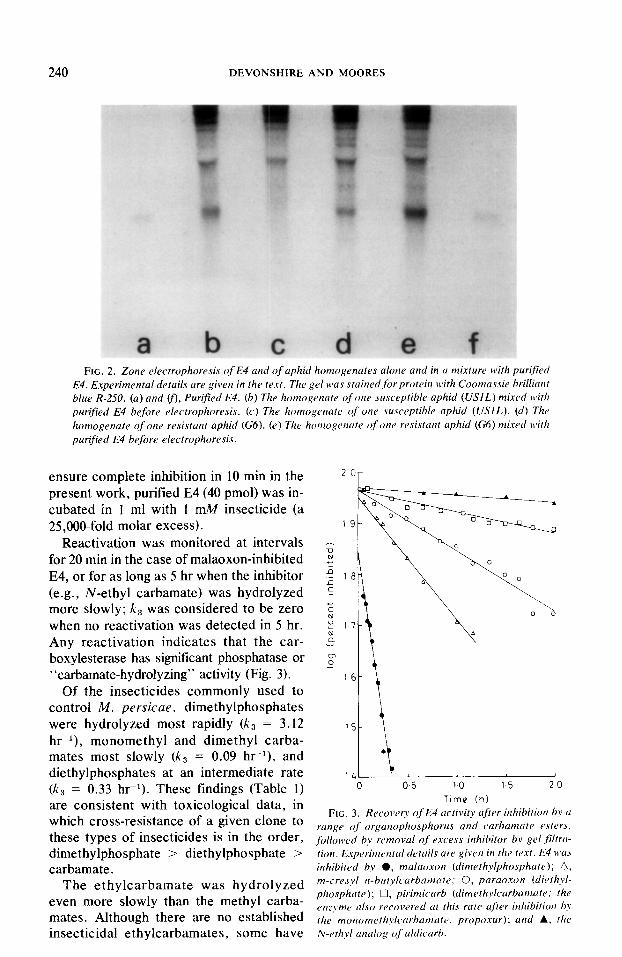
240 DEVONSHIRE AND MOORES
c FIG. 2. Zone electrophoresis of E4 and of aphid homogenates alone and in a mixture with purified
E4. E.xperimental details are given in the text. The gel was stained for protein with Coomassie brilliant blue R-250. (a) and v), Purified E4. (b) The homogenate of one susceptible aphid (US/L) mixed itith purified E4 before electrophoresis. (c) The homogenate of one susceptible aphid (USIL). (d) The homogenate of one resistunt aphid (G6). (e) The homogenate of one resistant aphid (G6) mixed bc?th purij?ed E4 before electrophoresis.
ensure complete inhibition in 10 min in the present work, purified E4 (40 pmol) was in- cubated in I ml with 1 mM insecticide (a 25 ,OOO-fold molar excess).
Reactivation was monitored at intervals for 20 min in the case of malaoxon-inhibited E4, or for as long as 5 hr when the inhibitor (e.g., N-ethyl carbamate) was hydrolyzed more slowly; k, was considered to be zero when no reactivation was detected in 5 hr. Any reactivation indicates that the car- boxylesterase has significant phosphatase or “carbamate-hydrolyzing” activity (Fig. 3).
Of the insecticides commonly used to control M. persicae, dimethylphosphates were hydrolyzed most rapidly (li3 = 3.12 hr-l), monomethyl and dimethyl carba- mates most slowly (k, = 0.09 hr-I), and diethylphosphates at an intermediate rate (k, = 0.33 hr’). These findings (Table 1) are consistent with toxicological data, in which cross-resistance of a given clone to these types of insecticides is in the order, dimethylphosphate > diethylphosphate > carbamate.
The ethylcarbamate was hydrolyzed even more slowly than the methyl carba- mates. Although there are no established insecticidal ethylcarbamates, some have
2 Or
14 b ,
0 05 10 15 20
Ttme (h)
FIG. 3. Recovery of E4 activity ufter inhibition by a range of organophosphorus and carbamate esters. followed by removal of excess inhibitor by gel filtra- tion. E.xperimental detuils are given in the text. E4 was inhibited by 0, malaoxon (dimethvlphosphate); A, m-cresvl n-butvlcarbamate; 0, paraoron (dieth.vl- phosphate); 0, pirimicarb (dimethvlcurbamate; the enzyme also recovered at this rate after inhibition b? the tnonometh~lcarbanz~~t~~, proposur); and A. the N-ethyl unalog of aldicarb.

TABL
E 1
Firs
t-Ord
er
Rate
Co
tzst
atlts
(k,
) fo
r th
e Re
activ
ation
of
E4
In
hibi
ted
by
a Ra
nge
of O
rgan
opho
spbo
rus
and
Carb
amat
e Es
ters
m
Inse
ctic
idal
cl
ass
inhi
bitin
g E4
a
k,
i%
RI
R2
Inhi
bito
rs
used
(h
r-’
2 SD
) SC
R,R
,P(O
).E4
E
CHBO
C
H,O
m
alao
xon
3.12
2
0.24
(1
3Y
F
C&O
C&
O
para
oxon
0.
33
2 0.
03
(6)
2 n-
C,H
,O
n-&H
,0
n-pr
opylp
arao
xon
0 C
iso-&
H,O
iS
o-C
3H,0
iso
-pro
pylp
arao
xon,
D
FP
0 K
CH
30
NH
* m
etha
mid
opho
s su
lfoxi
de
(17)
0
%
CH
,O
CH
,S
isodi
met
hoat
e 0
0 C&
O
n-C
3H,S
pr
ofen
ofos
0
g C
H,O
is
o-C
BH,N
H
O-m
ethy
l 0-
(2,5
-dich
loro
-4-
0 z?
io
doph
enyl)
N
-isop
ropy
lpho
s-
phor
oam
idat
e R
z
n-C
,H$
n-C4
HSS
S,S,
S-tri
buty
lpho
spho
rotri
thio
- 0
Y
late
(D
EF)
R,R
ZNC
(O).E
4 C
Hz
CH
a C
d-b
n-C
&H,
iso-C
3H7
n-C$
HS
(S-m
etho
myl
)
a Nu
mbe
r of
det
erm
inat
ions
.
CH
, pi
rimica
rb
H
prop
oxur
H
et
hyl
anal
ogue
of
ald
icarb
H
m
-cre
syl
n-pr
opylc
arba
mat
e H
m
-cre
syl
iso-p
ropy
lcarb
amat
e H
m
-cre
syl
n-bu
tylc
arba
mat
e
CH
, th
iodi
carb

242 DEVONSHIRE AND MOORES
recently been shown to have strong juvenile hormone activity, and to control stored product pests resistant to conventional in- secticides (26).
Of the other esters examined (Table 1) two are well-established insecticides al- though only methamidophos has been used widely to control M. persicae; this com- pound and profenofos, both 0-alkyl S-alkyl phosphate esters, inhibited the enzyme ir- reversibly. Methamidophos is a poor es- terase inhibitor, considered (17) to act through the formation in vivo of its sul- foxide. Consequently, oxidized methami- dophos was used in the present in vitro work. Since M. persicae is only slightly resistant to such O,S-dialkylphosphate de- rivatives they are a good choice for con- trolling resistant aphids if their field per- formance is adequate. Profenofos is known to control effectively Heliothis virescens resistant to other organophosphorous in- secticides, although the biochemical basis of this is not known (27). It also synergizes strongly the toxicity of some pyrethroids to cabbage looper larvae and housefly adults (28).
Other compounds inhibiting E4 irrevers- ibly were the well-established esterase in- hibitor, S,S,S-tributylphosphorotrithiolate (DEF), the O,O-dipropylphosphates, the O-methyl N-iso-propylphosphoroamidate, and the N-propylcarbamates. Again, resis- tance to such classes of chemical should be slight but they have not been used widely as insecticides. However, N-propylcarba- mates have given effective control of Neph- otettix cincticeps resistant to N-methyl- carbamates, because the mutant acetyl- cholinesterase responsible for resistance to the latter is more susceptible than the nor- mal enzyme to inhibition by the propyl analogs (29).
Thiodicarb, essentially two molecules of methomyl joined by a N-S-N bridge (30), inhibited E4 irreversibly. This indicates that it acts in vitro as the intact molecule, rather than through the formation of methomyl when the inhibited enzyme would have been expected to reactivate at a
rate characteristic of monomethyl carba- mates. Although the insecticide is a poor aphicide, it shows synergistic activity with cypermethrin against Aphis fabae and also with methomyl against soya fly larvae (30).
As well as predicting which classes of chemical should be as effective against re- sistant aphids, these studies should also define compounds as potential synergists when they inhibit E4 irreversibly. How- ever, since E4 has a very poor catalytic ef- ficiency and causes resistance only because it is present in such large amounts, syner- gists are unlikely to be effective in this in- stance; they have the greatest potential when they disrupt an insecticide-degrading enzyme having a high catalytic center ac- tivity. Rice and Sawicki (personal com- munication) have demonstrated that al- though the fungicide Kitazin P (O,O-di- iso-propyl S-benzylphosphorothiolate) strongly synergizes demeton-S-methyl, more so against resistant than susceptible M. persicae, its effect was apparent in leaf-dip bioassays only at a concentration of 0.05-O. l%, close to that killing aphids di- rectly and also causing phytotoxicity. Such concentrations are approximately 500 times and 5 times, respectively, the L&‘s for demeton-S-methyl alone against suscepti- ble and R, aphids and is unlikely to be of practical significance. Similarly, synergistic effects of analogous phosphates were only apparent when doses of synergists close to their LD,,‘s were used (31). Kitazin P has been observed to synergize malathion well in the green rice leafhopper (IV. cincticeps) in which resistance is caused by an esterase hydrolyzing the carboxyl ester in this in- secticide (29) as opposed to the nonspecific phosphatase activity of E4 in M. persicae.
Hydrolysis of Perrnethrin b-y E4
Preliminary experiments in which aphid homogenates (5 mg/ml buffer) were incu- bated with 14C-acid-labeled cis- or trans- permethrin (1 PM) indicated that the cis isomer was unaffected while the tram iso- mer was hydrolyzed at a rate proportional

BIOCHEMICAL CAUSE OF CROSS RESISTANCE OF h4.m~ prrsiuw 243
to the E4 activity in susceptible, R, and R2 aphids. Permethrin metabolism was studied further with the purified E4 preparation using both acid-labeled and alcohol-labeled permethrin. Because the enzyme was very active, assays of either type typically lasted 5-10 min. during which they were linear.
It was not possible to measure the K,,] of E4 for (IRS)tmns-permethrin. At concen- trations up to 1 @4, the rate of hydrolysis was directly proportional to the concentra- tion of trans-permethrin indicating no sig- nificant accumulation of Michaelis com- plex. Attempts to assay the enzyme with greater substrate concentrations (with con- stant 1% acetone in the assay) gave very erratic results, often with hydrolysis ceas- ing virtually completely, probably caused by the limited solubility of trans-permeth- rin. All subsequent in vitro assays were therefore done at 1 pM.
No matter how much E4 was used for in Ivitro incubations, no hydrolysis of (IRS) cis-permethrin was detected (detection lim- it < 1% of rate observed for rruns-per- methrin). Furthermore, with (lRS)frans- permethrin as substrate, regardless of the amount of E4 used, no more than 50% was hydrolyzed suggesting that E4 might be specific for one of the trans enantiomers.
Each was therefore synthesized (see Materials and Methods) and incubated with the purified enzyme. (lS)frans-Permethrin was readily hydrolyzed but no hydrolysis of its enantiomer could be detected even with large amounts of E4 or longer assays. While IUULS isomers are usually hydrolyzed more rapidly than cis isomers, the absolute spe- cificity of E4 for this one enantiomer is un- usual (32) and was surprising since it is the least toxic of the four isomers and its me- tabolism would be expected to have little effect on overall resistance to the mixture. The possibility of resistance to the other isomers arising by their being sequestered by the very large amount of E4 protein in resistant aphids is presently being investi- gated.
To relate the activity of E4 in hydrolyzing
(lS)truns-permethrin with its activity against organophosphates and carbamates, samples of the enzyme were assayed using naphth-l-y1 acetate and the pyrethroid as substrates. The ratio of activities (naphth- l-y1 acetate at V,,, :( lS)rruns-permethrin at 1 pM) was 1.41(?0.16) x lo3 (*SD, mean of four determinations). From the known (4) catalytic center activities (k3) of E4 for naphth-l-y1 acetate and insecticides, (lS)frans-permethrin (at 1 PM) is therefore hydrolyzed approximately 250, 2500, and 8000 times faster than dimethylphosphates, diethylphosphates, and methylcarbamates (at V,,,), respectively.
E4 is thus responsible for cross-resis- tance of M. persicue to permethrin and probably also to the other pyrethroids pre- viously examined toxicologically. In sup- port, clones with increased E4 activity con- sistently show cross-resistance to pyre- throids. There is limited evidence that resistance to the pyrethroid, deltamethrin, can occur independently of E4 and or- ganophosphorus resistance, but this was only in a clone generated in the laboratory by treating aphids with the mutagen ethyl- methane sulfonate (33). Its relevance to naturally occurring populations is uncer- tain. Furthermore, there is evidence that the insecticide used in this work, delta- methrin, affects the aphids’ behavior (34) distorting its toxological assessment in bio- assays using treated leaves.
Summury of the Role of E4 in Resistance
At first sight. it seems surprising that such an inefficient enzyme could cause re- sistance. Its catalytic center activity (k3) is extremely small, each molecule of enzyme hydrolyzing in an hour only three molecules of dimethylphosphate, 0.3 of diethylphos- phate. or 0.1 of mono- or dimethylcarba- mate. However, the very large amount of E4 produced by resistant aphids is suffi- cient to detoxify a significant proportion of a toxic dose, not only by hydrolysis, but also by sequestering the insecticide. Table 2 illustrates that E4 affects the performance

244 DEVONSHIRE AND MOORES
TABLE 2 Muximum Amount (ng) of LUI lllseclicide (oj. MW 250) Thut Could Be Detoxified by Hydrolysis urui
Sequestration by the Catalytic Center of E4 in Susceptible and Resistant Aphids
Aphid type: s R, R2 G6 pmol E4/400 yg aphid: 0.15 0.7 2.7 10
Inhibitor Seq. Hyd. Seq. Hyd. Seq. Hyd. Seq. Hyd.
Dimethylphosphate Diethylphosphate Mono- or
dimethylcarbamate
0.04 0.12 0.18 0.56 0.68 2.1 2.5 7.8 0.04 0.01 0.18 0.06 0.68 0.22 2.5 0.83
0.04 0.004 0.18 0.02 0.68 0.07 2.5 0.25
Note. The amounts of insecticide sequestered and hydrolyzed were calculated thus: Seq., ng sequestered (nmol E4 x MW of insecticide); Hyd., ng hydrolyzed/hr (ng sequestered x k, for each inhibitor class).
of carbamates only slightly, primarily by sequestering, and that a comparable amount to this would be hydrolyzed only after 10 hr. However, for dimethylphos- phates sequestering is of lesser importance since every hour three times the amount sequestered is hydrolyzed.
In the context of the amount of insec- ticides required to kill an aphid, these fig- ures are very significant. In topical applica- tion bioassays, the LD,,,‘s of insecticides are typically of the order of 1 ng, of which only lo-25% penetrates and the remainder is lost by evaporation (22). Thus, even in susceptible insects, of the 200 pg or so pen- etrated, approximately 80% of a dimethyl- phosphate can be removed by E4 within an hour. With the larger doses needed to kill very resistant aphids, an even smaller pro- portion of the applied dose actually pene- trates (39, possibly amplifying the effect of the increased amount of E4 in these aphids.
It is clear from Figs. 1 and 2 that E4 rep- resents a substantial proportion of the total protein in very resistant aphids. Gel filtra- tion and SDS electrophoresis indicate the MW of E4 to be 65,000, so that the lo-12 pmol E4 per aphid in the most resistant clone (5) represents 700-850 ng protein. Aphids each have 30-40 pg protein, of which E4 is therefore approximately 3%. Despite this large diversion of resources into producing an enzyme that is apparently redundant in the absence of insecticides,
these aphids show no “lack of fitness” under laboratory rearing conditions.
E4 is classified as a B-esterase because it hydrolyzes carboxyl esters and this activity is readily inhibited by lo-* M paraoxon but is unaffected by lo-” M eserine (36). How- ever, such rigid definitions are not appro- priate here since its ability to cause resis- tance relies on the weak phosphatase activity of E4, and even weaker hydrolytic activity with carbamate esters, coupled with its high affinity for sequestering these com- pounds.
E4 tends to be concentrated in the em- bryos (7) and it has been suggested (37) that aphids have evolved a novel way of “ex- creting” insecticide bound to the E4 in their offspring and “the adult is then free to synthesise new stocks of unbound es- terase.” This is unlikely, because the in- secticide is detoxified as soon as it binds to E4 and so the parent stands to gain nothing by eliminating it; on the contrary, di- methylphosphates remain “bound” for only 20 min, so that in very resistant aphids, as much as 10 pmol unbound E4 is regenerated every 20 min, a very definite advantage to the aphid, not relying on de llovo protein synthesis. However, the loca- tion of an increased concentration of E4 in the embryos, which is also apparent in early instar nymphs (8), might have a more al- truistic explanation; in leaf dip bioassays, the test aphids may all be killed by a partic-

BIOCHEMICAL CAUSE OF CROSS RESISTANCE OF My,-us pmiccrr 245
ular insecticide concentration while their offspring produced during the bioassay may survive and feed with no apparent ill effects (1) perhaps because of the protection af- forded by their increased E4 content.
The work described here forms part of a project aimed at formulating strategies to counter resistance. Results of such funda- mental work have important practical con- sequences; for M. persicae in which resis- tance relies on increased production of E4, there is no value in alternating the use of different classes of insecticide because any one will select the same mechanism. Fur- thermore, since this one mechanism gives very strong resistance to dimethylphos- phates, but only weak resistance to carba- mates, the latter (any of the group, not only those used here) will impose the least selection pressure on this particular resis- tance mechanism. Zf this holds under$eld conditions, then carbamates should be used preferentially, even if organophosphorus insecticides are apparently performing ad- equately. Such a strategy should help to delay the buildup of resistance in M. per- sicae populations.
ACKNOWLEDGMENTS
We thank A. D. Rice, R. M. Sawicki, and M. F. Stribley for useful discussions throughout this work, permission to refer to unpublished results, and for providing a plentiful supply of aphids.
REFERENCES
1. R. M. Sawicki and A. D. Rice, Response of sus- ceptible and resistant peach-potato aphids Mv;us persicar (Sulz.) to insecticides in leaf- dip bioassays, Prstic. Sci. 9, 513 (1978).
2. D. Otto, Nachweis von Kreuzresistenz eines Dimethoat-resistenten Mvius-persicar-Sulz.- Stammes gegenuber Pyrethroiden, Arch. Phytopmthoi. Pjkwzrnschutz. Brrlirl 16 (4). 283
(1980).
3. P. H. Needham and R. M. Sawicki, Diagnosis of resistance to organophosphorus insecticides in My:us prrkac (Sulz.), Nature (London) 230, 125 (1971).
4. A. L. Devonshire. The properties of a car- boxylesterase from the peach-potato aphid. M.v~.rr~ persicue (Sulz.), and its role in confer-
5.
6.
8.
9.
10.
11.
12.
13.
14.
1.5.
16.
ring insecticide resistance. Biochenr. J. 167.675
(1977). A. L. Devonshire and R. M. Sawicki, Insec-
ticide-resistant MyiUs persicae as an example of evolution by gene duplication, Natrtre (Len- don) 280, 140 (1979).
R. M. Sawicki, A. L. Devonshire. A. D. Rice, C. D. Moores, S. M. Petzing, and A. Cameron. The detection and distribution of organophos- phorus and carbamate insecticide-resistant Myzrrs pcrsicaa (Sulz.) in Britain in 1976. Pes- tic. Sci. 9, 189 (1978).
H. Takada, Esterase variation in Japanese popu- lations of My;us prrsicac (Sulzer) (Homoptera: Aphididae), with special reference to resistance to organophosphorus insecticides, Appl. Ettt. Zoo/. 14, 245 (1979).
U. Wachendorff and F. Klingauf, Esterasetest zur Diagnose von Insektizidresistenz bei Aphiden. 2. P~crtz;~,t~XrrrnXheit~~tl PJlun;rnschut: 85, 2 18 (1978).
R. M. Sawicki, A. L. Devonshire, R. W. Payne. and S. M. Petzing. Stability of insecticide re- sistance in the peach-potato aphid, Myzm twr- sicaa (Sulzer), Pestic. Sri. 11, 33 (1980).
S. D. Curry, R. W. Young, A. Berkelhammer. and D. E. Ailman, Organophosphorothioate com- pounds and methods of preparing same, U.S. Patent Number 3,309,371 (1967).
M. Elliott, A. W. Farnham, N. F. Janes, D. M. Johnson. and D. A. Pulman, The Pyrethrins and related compounds. XXXIV. Synthesis, ‘“C- nuclear magnetic resonance spectra and insec- ticidal activity of cycloalkyl analogues of fen- valerate, Prstk. Sci. 11, 513 (1980).
P. E. Burt. M. Elliott. A. W. Farnham, N. F. Janes. P. H. Needham, and D. A. Pulman. The Pyrethrins and related compounds. XIX. Geometrical and optical isomers of 2,2- dimethyl-3-(2,2-dichlorovinyl)-cyclopropane- carboxylic acid and insecticidal esters with 5-benzyl-3-furylmethyl and 3-phenoxybenzyl alcohols, Pestic. SC;. 5, 791 (1974).
D. M. Soderlund, Pharmacokinetic behaviour of enantiomeric pyrethroid esters in the cock- roach, Periplc~nrtu atnericatla L.. Prstic-. Biochem. Physid. 12, 38 (1979).
T. L. Rosenberry and S. A. Bernhard, Studies of catalysis by acetylcholinesterase. Synergistic effects of inhibitors during the hydrolysis of acetic acid esters, Biochemistry 11,4308 (1972).
W. N. Aldridge and E. Reiner, “Enzyme Inhib- itors as Substrates,“ North-Holland. Am- sterdam/London, 1972.
G. J. S. Ross. Simple non-linear modelling for the general user in “Proc. Sess. Int. Stat. Inst. 40th.” Vol. 2. p. 585, 1975.

246 DEVONSHIRE AND MOORES
17. M. Eto, S. Okabe, Y. Ozoe, and K. Maekawa. Oxidative activation of O,S-dimethyl phos- phoramidothiolate, Pesfic. Biochrm. Physiol. 7, 367 (1977); C. M. Thompson and T. R. Fukuto, Mechanism of cholinesterase inhibition by methamidophos, J. Agr. Food Chem. 30, 282 (1982).
18. R. A. Nicholson and R. M. Sawicki, Genetic and biochemical studies of resistance to permethrin in a pyrethroid-resistant strain of houseflies, Pesfic. Sci.. in press (1982).
19. D. E. Williams and R. A. Reisfeld, Disc electro- phoresis in polyacrylamide gels: Extension to new conditions of pH and buffer, Ann. N.Y. Acud. Sci. 121, 373 (1964).
20. A. L. Devonshire, Studies of the carboxyles- terases of Myzus persicae resistant and sus- ceptible to organophosphorus insecticides, in “Proc. 8th Br. Insectic. Fungic. Conf.,” Vol. 1, p. 67, 1975.
21. U. K. Laemmli, Cleavage of structural proteins during the assembly of the head of bac- teriophage T4, NatLdre (London) 227, 680 (1970).
22. A. L. Devonshire and P. H. Needham, The fate of some organophosphorus compounds applied topically to peach-potato aphids (M.vzus per- sitar (Sulz.)) resistant and susceptible to in- secticides, Pesric. Sci. 5, 161 (1974).
23. A. L. Devonshire, The biochemical mechanisms of resistance to insecticides with especial refer- ence to the housefly, Muscu tionwstictr and aphid, Myzus persicctr. Pestic. Sci. 4, 521 (1973).
24. A. P. Beranek, Esterase variation and organo- phosphate resistance in populations of AphiJ
fabae and M,v;us persicue. En/. Exp. Appl. 17, 129 (1974).
25. C. D. Johnson and R. L. Russell, A rapid, simple radiometric assay for cholinesterase, suitable for multiple determinations, At7ul. Biochem. 64, 229 (1975’).
26. S. Dorn, M. L. Frischknecht, V. Martinez, R. Zurfluh, and U. Fischer, A novel non- neurotoxic insecticide with a broad activity spectrum, 2. pflanzerlkrankheiter2 ef/an- xwschur; 88, 269 (1981).
27. J. B. Nosky, J. A. Harding. and D. A. Wolfen-
barger. Activity of certain O-ethyl S-propyl phosphorothioates and phosphorodithioates and oxime carbamates against organophos- phorus resistant and susceptible strains of the tobacco budworm. SW. Enromol., 5,245 (1980).
28. L. C. Gaughan, J. L. Engel, and J. E. Casida, Pesticide interactions: Effects of organophos- phorus pesticides on the metabolism, toxicity. and persistence of selected pyrethroid insec- ticides. Pesfic. Biochem. Physic/. 14,81 (1980).
29. H. Hama, Mechanism of insecticide resistance in green rice leafhopper and small brown plant- hopper. Rev. Plant. Pro&c. Res. 13, 54 (1980).
30. H. S. Yang and D. E. Thurman, Thiodicarb-A new insecticide for integrated pest manage- ment. in “Proc. Br. Crop Prot. Conf.,” Vol. 3, p. 687, 1981.
31. F. J. Oppenoorth and S. Voerman, Hydrolysis of paraoxon and malaoxon in three strains of M.vzus persicae with different degrees of para- thion resistance, Pestic. Biochem. Physiol. 5, 431 (1975).
32. D. M. Soderlund and J. E. Casida. Stereo- specificity of pyrethroid metabolism in mam- mals. Amer. Chem. Sot. Symp. 42, 173 (1977).
33. R. Bikhi, Evidence that resistance against pyre- throids in aphids Myzus persicae and Phorodon humuli is not correlated with high carboxyles- terase activity, Z. P~anlerrkrat7klleirer7 Pjlatr- zwchrcti 88, 631 (1981).
34. R. W. Gibson, A. D. Rice and R. M. Sawicki, Ef- fects of the pyrethroid deltamethrin on the ac- quisition and inoculation of viruses by Myzus persicae. AH~. Appl. Bid. 100, 49 (1981).
3.5. P. H. Needham and A. L. Devonshire, Labora- tory techniques for evaluating insecticides for aphid control and their application to studies on resistance, in “Crop Protection Agents” (N. R. McFarlane, Ed.), p. 413, Academic Press, New York/London, 1977.
36. W. N. Aldridge. Serum esterases, Bioc,hrm. J. 53, 110 (1953); R. S. Holmes and C. J. Masters. The developmental multiplicity and isoenzyme status of cavian esterases, Biochim. BiophyA. Actu 132, 379 (1967).
37. S. Bunting, Sexless aphids-The secret of their success, Nell, Sci. 91, 602 (1981).
Copyright © 2022 FDOKUMEN




















