
Roles of DCL4 and DCL3b in rice phased small RNA biogenesis
13
Roles of DCL4 and DCL3b in rice phased small RNA biogenesis Xianwei Song 1,2,† , Pingchuan Li 1,3,4,† , Jixian Zhai 1,3,4,† , Ming Zhou 1,2,† , Lijia Ma 1 , Bin Liu 5 , Dong-Hoon Jeong 3,4 , Mayumi Nakano 3,4 , Shouyun Cao 1 , Chunyan Liu 1 , Chengcai Chu 1 , Xiu-Jie Wang 1 , Pamela J. Green 3,4 , Blake C. Meyers 3,4 and Xiaofeng Cao 1,* 1 State Key Laboratory of Plant Genomics and National Center for Plant Gene Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China, 2 Graduate University of the Chinese Academy of Sciences, Yuquan Road, Beijing 100039, China, 3 Department of Plant and Soil Sciences, University of Delaware, Newark, DE 19711, USA, 4 Delaware Biotechnology Institute, Newark, DE 19711, USA, and 5 Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China Received 17 August 2011; revised 27 September 2011; accepted 29 September 2011. * For correspondence (fax +86 10 64873428; e-mail [email protected]). † These authors contributed equally to this work. SUMMARY Higher plants have evolved multiple proteins in the RNase III family to produce and regulate different classes of small RNAs with specialized molecular functions. In rice (Oryza sativa), numerous genomic clusters are targeted by one of two microRNAs (miRNAs), miR2118 and miR2275, to produce secondary small interfering RNAs (siRNAs) of either 21 or 24 nucleotides in a phased manner. The biogenesis requirements or the functions of the phased small RNAs are completely unknown. Here we examine the rice Dicer-Like (DCL) family, including OsDCL1, -3a, -3b and -4. By deep sequencing of small RNAs from different tissues of the wild type and osdcl4-1, we revealed that the processing of 21-nucleotide siRNAs, including trans-acting siRNAs (tasiRNA) and over 1000 phased small RNA loci, was largely dependent on OsDCL4. Surprisingly, the processing of 24-nucleotide phased small RNA requires the DCL3 homolog OsDCL3b rather than OsDCL3a, suggesting functional divergence within DCL3 family. RNA ligase-mediated 5¢ rapid amplification of cDNA ends and parallel analysis of RNA ends (PARE)/degradome analysis confirmed that most of the 21- and 24-nucleotide phased small RNA clusters were initiated from the target sites of miR2118 and miR2275, respectively. Furthermore, the accumulation of the two triggering miRNAs requires OsDCL1 activity. Finally, we show that phased small RNAs are preferentially produced in the male reproductive organs and are likely to be conserved in monocots. Our results revealed significant roles of OsDCL4, OsDCL3b and OsDCL1 in the 21- and 24-nucleotide phased small RNA biogenesis pathway in rice. Keywords: phased small RNA, tasiRNA, miRNA, OsDCL4, OsDCL3b, OsDCL1. INTRODUCTION Higher plants are rich in two types of endogenous small RNAs, namely small interfering RNAs (siRNAs) and microRNAs (miRNAs), which are important in regulating genome stability, plant development and various biotic and abiotic stress responses (Baulcombe, 2004; Ramachandran and Chen, 2008). Most miRNAs are 20–22 nucleotides in length, whereas siRNAs are mostly 21 and 24 nucleotides in length. The biogenesis of small RNAs, which typically have 5¢-monophosphate and 3¢-OH groups, requires RNase III-like enzymatic activities. The small RNAs repress target genes in a sequence-specific manner at either transcriptional or post-transcriptional levels (Carthew and Sontheimer, 2009). The miRNAs and siRNAs originate from different types of precursors: miRNA precursors are endogenous single-stranded RNA molecules, which form hairpin-like secondary structures that are imperfectly mat- ched and predominately generate one species of small RNAs (Lagos-Quintana et al., 2001; Lau et al., 2001; Lee and Ambros, 2001; Reinhart et al., 2002; Meyers et al., 2008); siRNAs, on the other hand, are processed from long double-stranded RNAs (dsRNAs) with perfect or nearly perfect complementarity (Zamore et al., 2000; Elbashir et al., 2001). The Dicer and Dicer-like (DCL) proteins are core compo- nents involved in the biogenesis of small RNAs and are evolutionarily conserved in a diverse set of eukaryotes. ª 2011 The Authors 1 The Plant Journal ª 2011 Blackwell Publishing Ltd The Plant Journal (2011) doi: 10.1111/j.1365-313X.2011.04805.x
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Roles of DCL4 and DCL3b in rice phased small RNA biogenesis

Roles of DCL4 and DCL3b in rice phased small RNA biogenesis
Xianwei Song1,2,†, Pingchuan Li1,3,4,†, Jixian Zhai1,3,4,†, Ming Zhou1,2,†, Lijia Ma1, Bin Liu5, Dong-Hoon Jeong3,4,
Mayumi Nakano3,4, Shouyun Cao1, Chunyan Liu1, Chengcai Chu1, Xiu-Jie Wang1, Pamela J. Green3,4, Blake C. Meyers3,4
and Xiaofeng Cao1,*
1State Key Laboratory of Plant Genomics and National Center for Plant Gene Research, Institute of Genetics and Developmental
Biology, Chinese Academy of Sciences, Beijing 100101, China,2Graduate University of the Chinese Academy of Sciences, Yuquan Road, Beijing 100039, China,3Department of Plant and Soil Sciences, University of Delaware, Newark, DE 19711, USA,4Delaware Biotechnology Institute, Newark, DE 19711, USA, and5Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China
Received 17 August 2011; revised 27 September 2011; accepted 29 September 2011.*For correspondence (fax +86 10 64873428; e-mail [email protected]).†These authors contributed equally to this work.
SUMMARY
Higher plants have evolved multiple proteins in the RNase III family to produce and regulate different classes of
small RNAs with specialized molecular functions. In rice (Oryza sativa), numerous genomic clusters are
targeted by one of two microRNAs (miRNAs), miR2118 and miR2275, to produce secondary small interfering
RNAs (siRNAs) of either 21 or 24 nucleotides in a phased manner. The biogenesis requirements or the functions
of the phased small RNAs are completely unknown. Here we examine the rice Dicer-Like (DCL) family, including
OsDCL1, -3a, -3b and -4. By deep sequencing of small RNAs from different tissues of the wild type and osdcl4-1,
we revealed that the processing of 21-nucleotide siRNAs, including trans-acting siRNAs (tasiRNA) and over
1000 phased small RNA loci, was largely dependent on OsDCL4. Surprisingly, the processing of 24-nucleotide
phased small RNA requires the DCL3 homolog OsDCL3b rather than OsDCL3a, suggesting functional
divergence within DCL3 family. RNA ligase-mediated 5¢ rapid amplification of cDNA ends and parallel analysis
of RNA ends (PARE)/degradome analysis confirmed that most of the 21- and 24-nucleotide phased small RNA
clusters were initiated from the target sites of miR2118 and miR2275, respectively. Furthermore, the
accumulation of the two triggering miRNAs requires OsDCL1 activity. Finally, we show that phased small
RNAs are preferentially produced in the male reproductive organs and are likely to be conserved in monocots.
Our results revealed significant roles of OsDCL4, OsDCL3b and OsDCL1 in the 21- and 24-nucleotide phased
small RNA biogenesis pathway in rice.
Keywords: phased small RNA, tasiRNA, miRNA, OsDCL4, OsDCL3b, OsDCL1.
INTRODUCTION
Higher plants are rich in two types of endogenous small
RNAs, namely small interfering RNAs (siRNAs) and
microRNAs (miRNAs), which are important in regulating
genome stability, plant development and various biotic
and abiotic stress responses (Baulcombe, 2004;
Ramachandran and Chen, 2008). Most miRNAs are 20–22
nucleotides in length, whereas siRNAs are mostly 21 and
24 nucleotides in length. The biogenesis of small RNAs,
which typically have 5¢-monophosphate and 3¢-OH groups,
requires RNase III-like enzymatic activities. The small RNAs
repress target genes in a sequence-specific manner at
either transcriptional or post-transcriptional levels (Carthew
and Sontheimer, 2009). The miRNAs and siRNAs originate
from different types of precursors: miRNA precursors are
endogenous single-stranded RNA molecules, which form
hairpin-like secondary structures that are imperfectly mat-
ched and predominately generate one species of small
RNAs (Lagos-Quintana et al., 2001; Lau et al., 2001; Lee
and Ambros, 2001; Reinhart et al., 2002; Meyers et al.,
2008); siRNAs, on the other hand, are processed from long
double-stranded RNAs (dsRNAs) with perfect or nearly
perfect complementarity (Zamore et al., 2000; Elbashir
et al., 2001).
The Dicer and Dicer-like (DCL) proteins are core compo-
nents involved in the biogenesis of small RNAs and are
evolutionarily conserved in a diverse set of eukaryotes.
ª 2011 The Authors 1The Plant Journal ª 2011 Blackwell Publishing Ltd
The Plant Journal (2011) doi: 10.1111/j.1365-313X.2011.04805.x

Genetic and biochemical analyses have shown both speci-
ficity and redundancy of Dicer proteins in small RNA
metabolism (Henderson et al., 2006). There are four DCLs
encoded in the Arabidopsis thaliana genome (DCL1–4) and
six DCLs in rice (Oryza sativa), with rice containing one extra
copy of both DCL2 and DCL3 (Liu et al., 2005; Margis et al.,
2006).
OsDCL1 and DCL1 are major enzymes accounting for
miRNA maturation in rice and Arabidopsis, respectively
(Park et al., 2002; Liu et al., 2005). In Arabidopsis, DCL1
processes primary miRNAs (pri-miRNAs) into pre-miRNAs
and subsequently to a miRNA/miRNA* duplex (Park et al.,
2002; Kurihara et al., 2006). In rice, strong loss-of-function
OsDCL1 transgenic lines lead to lethality and weak lines
show pleiotropic phenotypes, similar to the phenotypes of
strong and weak DCL1 loss-of-function alleles in Arabid-
opsis (Schauer et al., 2002; Liu et al., 2005). DCL2 is
mainly responsible for 22-nucleotide siRNA related viral
defenses in Arabidopsis (Xie et al., 2004; Garcia-Ruiz et al.,
2010). DCL3 produces 24-nucleotide siRNAs as triggers for
transcriptional silencing of repeat sequences, mobile
elements and transgenes (Xie et al., 2004). However, dcl2
and dcl3 mutants in Arabidopsis have no obvious devel-
opmental defects under normal growth conditions. In rice,
knock-down of OsDCL2 produced a dwarf plant with
significantly lower spikelet fertility and negatively affected
maintenance of an endogenous dsRNA virus by increas-
ing the accumulation of virus siRNAs (Urayama et al.,
2010). Rice contains two genes that are homologous to
DCL3 of Arabidopsis, OsDCL3a and OsDCL3b. OsDCL3a is
responsible for the biogenesis of 24-nucleotide long
miRNAs (lmiRNA), which can direct cytosine DNA meth-
ylation both in cis and in trans (Wu et al., 2010). However,
the function of OsDCL3b, which exhibited panicle- and
early seed-specific expression (Kapoor et al., 2008),
remains unknown.
Besides processing a few evolutionarily ‘young’ miRNAs
(Rajagopalan et al., 2006), DCL4 is mainly responsible for
processing endogenous 21-nucleotide siRNAs including
trans-acting siRNAs (tasiRNAs) (Dunoyer et al., 2005; Gas-
ciolli et al., 2005; Xie et al., 2005; Yoshikawa et al., 2005) as
well as virus-derived siRNAs (Garcia-Ruiz et al., 2010).
Trans-acting siRNAs are a special type of siRNAs whose
biogenesis is initiated by miRNAs that target the non-coding
TAS transcripts for cleavage. After cleavage, RNA DEPEN-
DENT RNA POLYMERASE 6 (RDR6) converts the sliced
single-stranded RNA into double-stranded with the assis-
tance of SUPPRESSOR OF GENE SILENCING 3 (SGS3).
Subsequently, DCL4 performs a ‘phased’ cleavage at
21-nucleotide intervals starting from the miRNA-directed
cleavage site using the dsRNA as substrate (Vazquez et al.,
2004; Allen et al., 2005; Gasciolli et al., 2005; Xie et al., 2005).
In this context, ‘phased’ indicates that the small RNAs are
generated in a fixed pattern, separated by 21-nucleotides on
both strands, with each duplex of 21-mers having a two-
nucleotide 3¢ overhang.
In Arabidopsis, three DCL1-dependent miRNAs trigger the
initial cleavage event in four TAS families (TAS1–TAS4), with
miR173 acting on TAS1 and TAS2, miR390 on TAS3 and
miR828 on TAS4, respectively (Allen et al., 2005; Rajagopa-
lan et al., 2006). The dual miR390-binding sites present on
TAS3 (only the 3¢ site is cleavable) are conserved in higher
plants and were proposed to explain TAS3 tasiRNA forma-
tion (Axtell et al., 2006). Phylogenetic analysis revealed that
the TAS3 loci are conserved in higher plants (Axtell et al.,
2006) and both OsDCL1 and OsDCL4 are required for
OsTAS3 tasiRNA biogenesis, indicating that the tasiRNA
biogenesis pathway is conserved between rice and Arabi-
dopsis (Liu et al., 2005, 2007). Recently, two independent
lines of evidence showed that both 21- and 22-nucleotide
forms of the same miRNAs can guide cleavage; however,
only the 22-nucleotide miRNAs were capable of triggering
secondary siRNA production, reflecting the unique role of
22-nucleotide RNA in small RNA metabolism (Chen et al.,
2010; Cuperus et al., 2010). We and Nagasaki et al. (2007)
showed that OsDCL4 mutations cause the loss of tasiRNA-
AUXIN RESPONSE FACTORs (tasiR-ARF) and result in
up-regulation of ARFs and miR165/166, which are thought
to establish the adaxial–abaxial axis in lateral organs (Liu
et al., 2007; Nagasaki et al., 2007). Moreover, we also showed
that endogenous 21-nucleotide siRNAs derived from long
hairpin structures are OsDCL4-dependent (Liu et al., 2007).
In rice, many 21- and 24-nucleotide phased small RNA
clusters and superclusters were identified from inflores-
cence small RNA libraries, and miR2118 and miR2275/
miR2775 (known as miR2275 in http://www.mirbase.org/,
and we refer to it as miR2275 hereafter) were predicted to
target the 21- and 24-nucleotide phased small RNA loci,
respectively (Johnson et al., 2009). The biogenesis of
phased small RNAs, especially the 24-nucleotide ones that
were not found in Arabidopsis, is still unknown.
Here, six small RNA libraries were constructed from
seedlings and panicles of 93-11 (a wild-type indica rice
variety) and osdcl4-1, as well as from Nipponbare (a wild-
type japonica rice variety). Sequencing data derived from
93-11 and osdcl4-1 indicated that OsDCL4 has a stronger
impact on 21-nucleotide siRNA accumulation in panicles
than in seedlings. Consistent with previous results (Liu
et al., 2007), tasiRNAs were OsDCL4-dependent in both
tissues tested, and osdcl4 loss-of-function mutants
showed great and moderate reduction of 21- and
24-nucleotide phased small RNAs, respectively. In addi-
tion, we found that the processing of 24-nucleotide
phased small RNAs requires OsDCL3b whose function
was previously unknown, rather than OsDCL3a, which
processes long miRNAs (Wu et al., 2010). The target sites
for cleavage of 21- and 24-nucleotide phased small RNA
loci, which match two families of RNase III-dependent
2 Xianwei Song et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

22-nucleotide small RNAs (miR2118 and miR2275), respec-
tively, were confirmed by the RNA ligase mediated-5¢rapid amplification of cDNA ends (RLM 5¢-RACE) and a
parallel analysis of RNA ends (PARE)/degradome library.
Both miR2118 and miR2275 were dependent on OsDCL1
and preferentially expressed in stamen and they are likely
to be conserved in monocots. Our results showed
genome-wide effects of osdcl4 on small RNA biogenesis
in rice and revealed the roles of OsDCL4, OsDCL3b and
OsDCL1 in the phased small RNA pathway.
RESULTS
Deep sequencing of small RNA populations
Our previous work showed that OsDCL4 is the major Dicer-
like protein generating 21-nucleotide siRNAs from exoge-
nous transgenic inverted repeats, endogenous inverted
repeats and TAS3 (Liu et al., 2007), indicating that OsDCL4
processes multiple substrates in rice. To further determine
the extent of OsDCL4 action on a genome-wide scale, six
small RNA libraries were constructed from panicles and
seedlings of the wild-type 93-11, the osdcl4-1 null allele in
the 93-11 background (Liu et al., 2007) and another wild-
type, Nipponbare. These libraries were used for deep
sequencing on the Illumina 1G system.
In total, over 20 million raw reads (18–26 nucleotides)
were generated from these six libraries, with each library
represented by 2.3–4.5 million reads. Approximately 73–82%
of the reads, varying by library, could be perfectly mapped to
the reference genomes (93-11 or Nipponbare), and the
numbers of genome-matched non-redundant small RNA
sequences ranged from 692 568 to 2 135 639 (Table S1 in
Supporting Information).
OsDCL4 preferentially affects 21-nucleotide siRNA
biogenesis in panicles
In 93-11 and Nipponbare, 21- and 24-nucleotide small RNAs
were the most predominant sizes (Figure 1a,b), consistent
with the small RNA size distribution typically observed in
Arabidopsis and rice (Henderson et al., 2006; He et al., 2010).
To determine the effect of OsDCL4 loss-of-function on small
RNA metabolism, we compared the small RNA populations
in panicles and seedlings of the osdcl4-1 null mutant with
those of the wild-type 93-11 plants. A marked reduction the
proportion of 21-nucleotide small RNAs was observed in
osdcl4-1 panicles (Figure 1a), but this reduction was smaller
in seedlings (Figure 1b). In the osdcl4-1 panicle sample,
21-nucleotide small RNAs accounted for 6.86% of the total
population, corresponding to an approximately two-fold
reduction compared to the 93-11 panicle library (18.9% of
total). In seedlings, however, the proportion of 21-nucleotide
small RNAs in wild-type 93-11 and osdcl4-1 was 18.6 and
14.3%, respectively, which showed much less of a change
compared with that in the panicles (Figure 1a,b). This is
consistent with two-fold higher expression level of OsDCL4
in panicles compared with seedlings (Figure 1c) and sug-
gests that more OsDCL4-dependent small RNAs exist in
panicles. Further analysis showed that these 21-nucleotide
small RNAs, which exhibited the most significant reduction
in the panicle of osdcl4-1, were preferentially derived from
unannotated regions in the rice genome (Figure 1d);
whereas the 24-nucleotide small RNAs were preferentially
from both repeat-associated and unannotated regions (Fig-
ure 1e), consistent with the previous analysis of rice small
RNA massively parallel signature sequencing (MPSS) data
(Nobuta et al., 2007).
Figure 1. Size distribution and annotation of
small RNAs from various rice genotypes.
(a, b) Small RNA size distribution of 93-11,
osdcl4-1 and Nipponbare in panicles (a) and
seedlings (b). The abundance of small RNA at
each size class is represented by their percentage
relative to the total abundance.
(c) Real-time PCR analysis of the expression level
of OsDCL4 in seedlings and panicles. OsACTIN1
was used for normalization.
(d, e) Annotation of 21- (d) and 24-nucleotide (e)
small RNAs.
Phased small RNA clusters biogenesis in rice 3
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

OsDCL4-dependent conserved and non-conserved
TAS3 loci in rice
Previous studies showed that OsDCL4 can produce
21-nucleotide tasiRNAs from TAS3 in a phased manner (Liu
et al., 2007). To further investigate whether other siRNAs
were processed in a phased pattern by OsDCL4, we per-
formed a computational analysis to identify phased siRNAs,
using an algorithm modified from a previous study (Howell
et al., 2007) to deal with the higher background caused by
deeper sequencing capacity (Figure S1). Small RNA data
from a published Arabidopsis Col-0 inflorescence study
(Lister et al., 2008) were used to test this modified algorithm.
From the published data, seven phased loci were found with
a phasing score (P-score) higher than 22.5, and all of them
had been previously reported as tasiRNA loci expressed in
flowers (Chen et al., 2007; Howell et al., 2007) (Table S2).
This result indicated that the modified algorithm efficiently
identified phased small RNA patterns within a deep
sequencing dataset. So we adopted a P-score of 25, a more
stringent cut-off, for the identification of phased small RNA.
Applying this algorithm to our six rice libraries, all of the
known rice TAS genes (TAS3a, TASb3b, TAS3c) were
identified with P-scores higher than 25 (Figure S2a) in the
wild types. Notably, TAS3b has two copies (TAS3b1 and
TAS3b2) within an 18 kb region in 93-11 (Figure S2) but one
copy in Nipponbare (data not shown). In addition, two
additional tasiRNA loci, TAS3d and TAS3e, with dual
miR390-binding sites but lower scores (�20) were identified
from both 93-11 and Nipponbare (Figure S2 and data not
shown). The abundances of most tasiRNA loci were dra-
matically reduced in osdcl4-1, and the phasing patterns of
some tasiRNA loci were also impacted (Figure S2a), con-
firming that OsDCL4 is key for tasiRNA biogenesis in rice (Liu
et al., 2007).
Roles of OsDCL4 in the biogenesis of 21- and 24-nucleotide
phased small RNAs
Consistent with a previous report (Johnson et al., 2009),
many 21- and 24-nucleotide phased small RNAs were iden-
tified in wild type panicles, whereas only a few phased small
RNA loci with a score of 25 or higher (mainly known tasiR-
NAs) were identified in seedlings and grains (Figure 2a,c,
Table S3). No other intervals were detected with a signifi-
cant phased pattern (Table S2). The 21-nucleotide phased
small RNAs, represented by 1136 clusters in 93-11 and 1540
clusters in Nipponbare with P-scores over 25 (Figure 2a),
were mainly derived from unannotated regions (Figure 2b).
In osdcl4-1, both the abundance and cluster number of
21-nucleotide phased small RNAs were significantly reduced
compared to the wild type (Table S3, Figure 2a,b). For
example, at 21-nucleotide phased locus No. 1000 in 93-11,
there is a dominant phasing pattern of each cycle to be
coincident with the cleavage site, which was identified by
5¢-RACE (Figure 3a) and further confirmed by PARE/degra-
dome library analysis (Figure 3b) of a degradome library
constructed from young panicles of 93-11 (Zhou et al., 2010).
The phased level within the cluster is reduced in osdcl4-1
due to lower small RNA abundance from each phased po-
sition (Figure 3a). These results suggested that the accu-
mulation of 21-nucleotide phased small RNAs is largely
dependent on the activity of OsDCL4.
The 24-nucleotide phased small RNAs, most of which also
originated from unannotated regions, were identified in
panicles of 93-11 (74 clusters) and Nipponbare (30 clusters)
with P-scores over 25 (Figure 2c,d). Unlike the 21-nucleotide
phased small RNAs, whose abundances and cluster num-
bers were both dramatically impacted in osdcl4-1 (Fig-
ure 2a,b), the abundances of some 24-nucleotide phased
small RNAs were moderately decreased in the osdcl4-1
Figure 2. Cluster numbers and annotation of 21-
and 24-nucleotide (nt) phased small RNAs.
(a, c) Cluster numbers of 21- (a) and 24-nucleo-
tide (c) phased small RNAs in different tissues of
93-11, osdcl4-1 and Nipponbare with a phasing
score (P-score) over 25. Grain data were derived
from (Zhu et al., 2008).
(b, d) Annotation for 21- (b) and 24-nucleotide (d)
phased small RNAs.
4 Xianwei Song et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

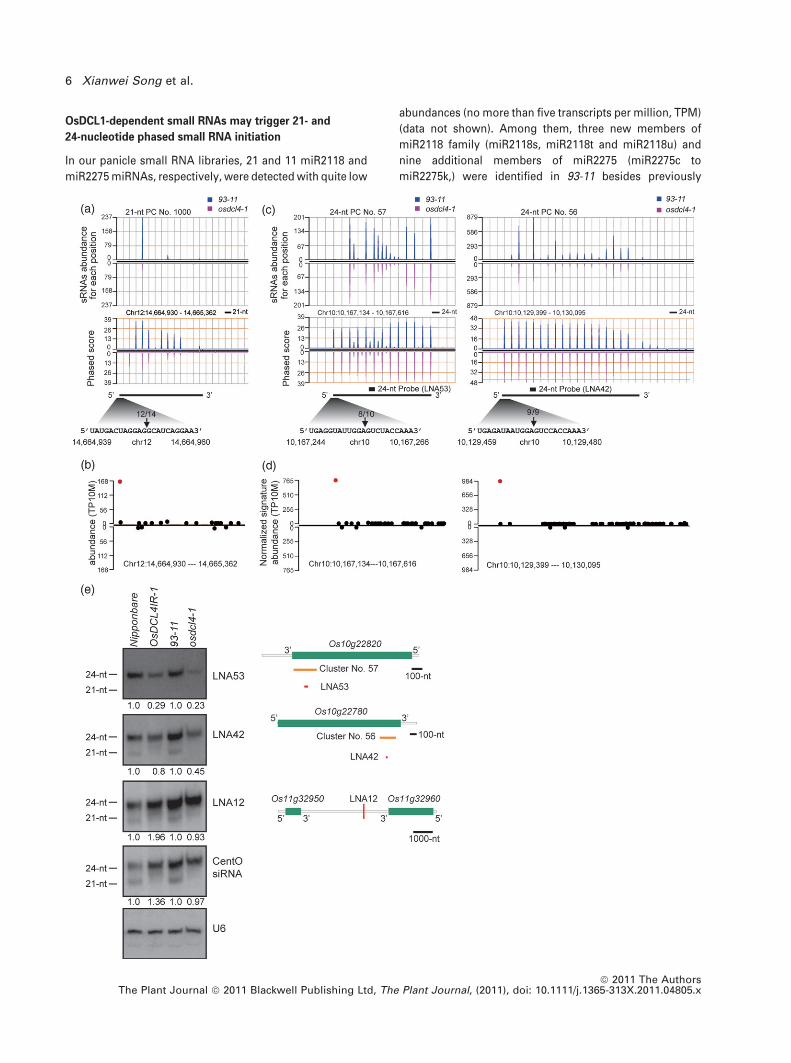
mutant and the cluster numbers were unaffected
(Figure 2c,d). To determine the accumulation pattern of
24-nucleotide phased small RNAs in osdcl4-1, two phased
clusters of No. 57 and No. 56 were selected for further
analysis. Each phased small RNA cluster was validated by
5¢-RACE (Figure 3c) and the PARE/degradome library anal-
ysis (Figure 3d), showing that the predominant cleavage
sites coordinate well with the phasing pattern (Figure 3c,d).
Consistent with the genome-wide data (Table S3, Figure 2c),
the overall phasing patterns of cluster No. 57 and No. 56
were mostly unchanged but with less accumulation of
phased small RNAs in each locus in osdcl4-1 (Figure 3c). In
addition, a moderate reduction of 24-nucleotide phased
small RNAs in osdcl4-1 was also detected by locked nucleic
acid (LNA) labeled probes, LNA53 and LNA42 derived from
cluster No. 57 and No. 56, respectively (Figure 3c,e). The
influence by osdcl4-1 on 24-nuclotide phased small RNAs
might be specific, because 24-nucleotide unphased small
RNAs including one derived from a single copy locus
(detected by LNA12) and another one derived from the
centromeric repeats (CentO siRNAs) (Zhang et al., 2005)
showed no obvious changes in osdcl4-1(Figure 3e). In
addition, the distribution of 21-nucleotide clusters showed
little overlap with that of 24-nucleotide phased small RNA
clusters in the 93-11 genome (Figure S3).
The 24-nucleotide phased small RNAs are processed
by OsDCL3b
Deep sequencing data combined with RNA gel blot results
showed the accumulation of 24-nucleotide phased small
RNAs was partially dependent on OsDCL4, suggesting other
DCL proteins might also be involved. In Arabidopsis, DCL3 is
the major Dicer that processes 24-nucleotide siRNAs (Xie
et al., 2004). Compared with Arabidopsis, two copies of
DCL3, namely OsDCL3a and OsDCL3b, are found in the rice
genome (Figure S4a) (Kapoor et al., 2008). OsDCL3a is
involved in non-canonical long miRNA biogenesis (Wu et al.,
2010). To determine whether OsDCL3a and OsDCL3b play a
role in the production of 24-nucleotide phased small RNAs,
we used RNA interference (RNAi) to knock down either
OsDCL3a or OsDCL3b (Figure S4b). Two OsDCL3a and
OsDCL3b knock-down lines, in which the transcript levels of
OsDCL3a and OsDCL3b were reduced to �20 and �50% of
wild type (WT; Nipponbare) (Figure S4c), respectively, were
used for northern blots to detect phased small RNAs. In the
OsDCL3a RNAi lines (OsDCL3aIRs), the 24-nucleotide
phased small RNAs detected by LNA53 and LNA42 showed
no significant changes (ranging from 0.72 to 1.15-fold)
compared with WT (Figure 4), whereas the signals for 24-
nucleotide unphased small RNAs including the one detected
by LNA12 and CentO siRNAs (Zhang et al., 2005), sharply
decreased (Figure 4). In contrast, the OsDCL3b RNAi lines
specifically affected 24-nucleotide phased but not unphased
small RNAs (Figure 4), suggesting that OsDCL3b is involved
in the processing of 24-nucleotide phased small RNAs and
acts distinctly from OsDCL3a. In addition, we also found that
the knock-down of OsDCL1 did not result in obvious defects
in the accumulation of both 24-nucleotide phased and
unphased small RNAs (Figure 4).
Cleavage sites of 21- and 24-nucleotide phased small RNA
loci share conserved motifs as target sites of miRNAs
Johnson et al. have predicted that miR2118 and miR2275 act
as triggers to initiate formation (Johnson et al., 2009) of 21-
and 24-nucleotide phased small RNAs, respectively. 5¢-RACE
analysis of a 21-(Figure 3a) and two 24-nucleotide phased
small RNAs (Figure 3c), suggested that these phased small
RNAs were initiated from a fixed cleavage site like tasiRNAs.
To explore whether this is common for all phased small
RNAs, we performed 5¢-RACE to locate the cleavage sites;
TAS3c, for which the transcripts are cleaved by miR390, was
used as a positive control (Table S4). Because the tran-
scribed strand of the phased small RNA loci is unknown
(RDR can produce the secondary strand and thus form a
dsRNA template), PCR primers were chosen to amplify only
one of the two strands. Among 30 21-nucleotide phased
small RNA loci chosen based on their abundances, 15 were
successfully mapped with a fixed cleavage site (Figure 5a,
top 15 lines). The cleavage sites exhibit a high degree of
sequence similarity, which defined a 22-nucleotide motif
targeted by members of miR2118 family (Figure 5b). Using
the sequence motif, 9 more 21-nucleotide phased small RNA
loci with this motif were identified and examined by RLM 5¢-RACE; the cleavage sites on these nine transcripts mapped
exactly to the motif region (Figure 5a, bottom 9 lines). MEME
was used to examine whether this motif was common for all
21-nucleotide phased small RNA originating sites (Bailey
and Elkan, 1994). Out of the 1136 clusters, 1116 (98.2%) were
found to contain this 22-nucleotide motif. From the PARE/
degradome library constructed from young panicles of the
93-11 library (Zhou et al., 2010), the cleavage sites of more
than 97% (1084 out of 1116) of the 21-nucleotide phased
small RNA clusters could be confirmed at the 22-nucleotide
motif (Figure 3b, Table S5).
The 24-nucleotide phased small RNA clusters were also
tested by RLM 5¢-RACE to identify cleavage sites (Figure 5c,
Table S6). A different 22-nucleotide motif was found to be
shared by all the tested cleavage sites and can be targeted by
members from the miR2275 family (Figure 5d). We used the
motif-identification program MEME to reveal that almost all
(73 of 74 in 93-11 and 30 of 30 in Nipponbare) 24-nucleotide
phased small RNA clusters contained such a motif
sequence. Moreover, cleavage sites in 67 of 73 (91.8%)
motif-containing clusters in 93-11 could be identified by the
PARE/degradome library (Figure 3d, Table S7). Therefore,
our results confirmed that miR2118 and miR2275 mediate
the biogenesis of 21- and 24-nucleotide phased small RNAs,
respectively, by initiating cleavage of their transcripts.
Phased small RNA clusters biogenesis in rice 5
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x
OsDCL1-dependent small RNAs may trigger 21- and
24-nucleotide phased small RNA initiation
In our panicle small RNA libraries, 21 and 11 miR2118 and
miR2275 miRNAs, respectively, were detected with quite low
abundances (no more than five transcripts per million, TPM)
(data not shown). Among them, three new members of
miR2118 family (miR2118s, miR2118t and miR2118u) and
nine additional members of miR2275 (miR2275c to
miR2275k,) were identified in 93-11 besides previously
6 Xianwei Song et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

described ones (Johnson et al., 2009) (Figure 6a,b, Tables
S8 and S9). The miR2118 members were mapped to two
small regions on chromosome 4 and 11 and found to be
tandem aligned in 93-11 and Nipponbare (Figure 6a). On
chromosome 4, 17 miR2118 members form nine clusters
within a 20 kb region. On chromosome 11, four members of
the miR2118 family were present in two clusters. In 93-11,
supported by degradome data and bioinformatics predic-
tions, each miR2118 and miR2275 family member could
potentially cleave transcripts and thus trigger the formation
of many 21- or 24-nucleotide phased small RNA clusters
(Figure 6a, b).
In addition, to exclude the possibility that low abundance
of miR2118 and miR2275 family members is a cloning
artifact in our deep sequencing, we treated small RNAs with
5¢-phosphate-dependent exonuclease (Terminator), which
specifically digests RNA containing a 5¢-monophosphate but
does not digest RNAs with a 5¢-triphosphate, 5¢-cap or
5¢-hydroxyl (see Experimental Procedures). The signals of
miR2118 and miR2275 were eliminated after exonuclease
treatment compared with untreated small RNAs from pan-
icles of three rice varieties (93-11, Nipponbare and Zhong-
hua11) (Figure 6c). As a negative control, the signal of U6,
which has a 5¢-triphosphate, was not affected (Figure 6c).
Therefore, we confirmed these motif-targeting small RNAs
were RNase III products with a 5¢-monophosphate.
To determine which DCL protein generates the triggers of
phased small RNA biogenesis, small RNA gel blot analysis
was performed using the LNA probes to detect the levels of
miR2118 (LNA58) and miR2275 (LNA64). As expected,
miR168, the well-recognized and conserved miRNA, was
greatly reduced in OsDCL1 knock-down transgenic plants
(OsDCL1IRs) (Figure 6d). Consistently, both miR2118 and
miR2275 were less accumulated in OsDCL1IRs. We also
noted that the accumulations of both miRNAs were affected
in osdcl4-1 (Figure 6d), which might account for the lower
accumulation of 24-nucleotide phased small RNAs in osdcl4-
1 (Figures 2d, 3c,e) owning to its epistatic influence on
miR2275. Therefore, it seems that the biogenesis of both
triggering miRNAs for 21- and 24-nucleotide siRNAs is
dependent on OsDCL1 and OsDCL4. However, it is worth
noting that loss-of-function osdcl4-1 shows severe spikelet
defects including thread-like lemma and male sterility. It is
also hypothesized that the loss of some organs or cell types
harboring abundant miR2118 and miR2275 in osdcl4-1
might results in losing of both miRNAs.
Figure 4. Effects of OsDCLs on the biogenesis of 24-nucletiode (nt) phased
small RNAs.
Northern blot analysis of the accumulation of the 24-nucleotide phased small
RNAs detected with probes LNA53, LNA42 in Nipponbare, OsDCL1IR1-1,
OsDCL3aIRs and OsDCL3bIRs. LNA12 labels a 24-nucleotide unphased small
RNA. CentO siRNA labels siRNAs derived from 165-bp CentO satellite. U6 was
used as the loading control. The intensity ratio of each signal relative to wild-
type Nipponbare is indicated below each image. The bottom panels represent
expression analysis of OsDCL3a and OsDCL3b in Nipponbare and RNA
interference (RNAi) lines by RT-PCR. The OsACTIN gene was used as an
internal control.
Figure 3. OsDCL4 has different effects on accumulation of 21- and 24-nucleotide (nt) phased small RNAs.
(a, c) The abundance and the phased patterns of selected 21- (a) and 24-nucleotide (c) phased small RNA clusters in the panicles of 93-11 and osdcl4-1. The top panel
represents the abundance of each position. The bottom panel represents the phasing score, plotted in a 10-cycle window. The schematic below the phase plot
represents the genomic position of the phased small RNA clusters. The black arrow indicates a site verified by RNA ligase mediated-5¢ rapid amplification of cDNA
ends (RLM 5¢-RACE) with the frequency of cloned RACE products. The black boxes in (c) indicate the locked nucleic acid (‘LNA’) probe, LNA53 and LNA42, detecting
the most abundant 24-nucleotide phased small RNA in both clusters, respectively.
(b, d) Validation of the cleavage by analysis of a parallel analysis of RNA ends (PARE)/degradome library of a 21- (b) and 24-nucleotide (d) phased small RNA clusters
as shown in (a) and (c), respectively. The red dot represents the cleavage site mapped by highly abundant degradome tag.
(e) Northern blot analysis of the accumulation of the 24-nucleotide phased small RNAs detected with probes LNA53, LNA42 in Nipponbare, OsDCL4IR-1, 93-11,
osdcl4-1. LNA12 labels a 24-nucleotide unphased small RNA. CentO siRNA labels siRNAs derived from 165-bp CentO satellite. U6 was used as the loading control.
The intensity ratio of each signal relative to wild-type Nipponbare is indicated below each image. The genomic locations of LNA53, LNA42 and LNA12 are shown on
right panel. Protein-coding genes and phased small RNA clusters are indicated by green and golden bars, respectively.
Phased small RNA clusters biogenesis in rice 7
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

The triggers and phased small RNAs are preferentially
expressed in stamen and are conserved in cereal
To determine the hypothesis, we investigated the expres-
sion pattern of both triggering miRNAs (miR2118 and
miR2275). The deep sequencing data from us and a previous
report showed that both 21- and 24-nucleotide phased small
RNAs are a class of panicle-specific siRNAs (Figure 2a,c,
Table S3) (Johnson et al., 2009). Northern blotting signals
for miR2118 and miR2275 as well as 24-nucleotide phased
small RNAs were consistently present in rice panicles but
absent in seedlings (Figure 7a). Furthermore, we found that
in contrast to tasiR-ARFs that accumulated in all detected
tissues and organs, the trigger miR2275 and 24-nucleotide
phased small RNAs were exclusively expressed in the sta-
men (Figure 7a). Compared to miR2275, miR2118 displayed
a distinct pattern and was present predominantly in stamens
but also in the palea and the pistils at low levels (Figure 7a).
Northern blot analysis in maize also revealed that both
miR2118 and miR2275 accumulated in stamens from various
developmental stages (Figure 7b), which was consistent
with the previous identification of both miR2118 and
miR2275 in deep sequencing data of small RNAs from early
developmental stages of maize anthers (Johnson et al.,
2009). Taken together, these findings indicate that 21- and
24-nucleotide phased small RNAs might be stamen-specific
small RNAs conserved in cereal species. So, the develop-
mental defects of osdcl4-1, in which the stamen identity is
disrupted, may account for the lower accumulation of
miR2118 and miR2275 in it.
DISCUSSION
Here we sequenced six small RNA libraries, including
93-11, osdcl4-1 (in 93-11 background) and Nipponbare,
from both panicles and seedling tissues, using Illumina’s
SBS technology. The deep sequencing data showed that
OsDCL4 is responsible for the biogenesis of 21-nucleotide
siRNAs including tasiRNAs, and its role is more promi-
nent in panicles than in seedlings. A genome-wide
phasing analysis revealed that 21-nucleotide phased small
RNAs are completely OsDCL4-dependent, whereas
24-nucleotide phased small RNAs only partially depend
on OsDCL4. 5¢-RACE and PARE/degradome library con-
firmed two 22-nucleotide conserved sequence motifs as
target sites of miR2118 and miR2275, which act to trigger
the biogenesis of 21- and 24-nucleotide phased small
RNAs, respectively. Further experiments demonstrated the
OsDCL1-dependent miR2118 and miR2275 are mainly or
were exclusively expressed in the stamen. In addition, we
revealed that OsDCL3b but not OsDCL3a is the Dicer that
directly processes 24-nucleotide phased small RNAs. To
sum up, our work revealed the roles of OsDCL4, OsDCL3b
and OsDCL1 in the biogenesis of 21- and 24-nucleotide
phased small RNAs.
Figure 5. Cleavage sites of 21- and 24-nucleotide phased small RNAs loci share conserved motifs as target sites of microRNAs (miRNAs).
(a, c) Alignments of sequences flanking the cleavage sites revealed by RNA ligase mediated-5¢ rapid amplification of cDNA ends (RLM 5¢-RACE) for 21- (a) and 24-
nucleotide phased small RNA clusters (c). The black arrows indicate cleavage sites verified by RLM 5¢-RACE. The frequencies of cloned RACE products are listed on
the right. The cleavage sites are marked in yellow. The top 15 sequences (above the black line) in (a) are from the randomly selected phased small RNA loci; the
bottom nine (below the black line) were selected based on the motif sequences for further validation.
(b, d) Conserved 22-nucleotide motifs of the 21- (b) and 24-nucleotide phased small RNA clusters (d) analyzed by WebLogo (Crooks et al., 2004). The diagrams for
conserved motifs were generated from 1084 21-nucleotide and 67 24-nucleotide sequences. The sequence complementarity between miR2118a and the motif of 21-
nucleotide phased small RNAs (b), miR2275a and the motif of 24-nucleotide phased small RNAs (d) are displayed at the bottom. The black arrows indicate cleavage
sites.
8 Xianwei Song et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

The roles of Dicers in the biogenesis of phased small RNAs
In rice, we and others previously showed that OsDCL1,
OsDCL3a and OsDCL4 are responsible for the production of
canonical miRNAs, non-canonical long miRNAs and
21-nucleotide siRNAs, respectively (Liu et al., 2005, 2007; Wu
et al., 2010). Moreover, both OsDCL1 and OsDCL4 are
required for tasiRNA generation (Liu et al., 2007). In rice
Figure 7. Expression patterns of trigger microRNAs
(miRNAs) and 24-nucleotide phased small RNA in
different tissues.
(a, b) Northern blot analyses of trigger miRNAs in
different tissues of rice (a) and maize (b). Stamen (S),
(M) and (L) represent the anther lengths of �3.0–
3.5 mm, �3.6–4.1 mm and �4.2–5 mm, respectively.
LNA53 detects 24-nucleotide phased small RNAs.
Figure 6. The triggers of 21- and 24-nucleotide phased small RNAs.
(a, b) Genomic locations of miR2118 (a) and miR2275 (b) in the 93-11 genome. The blue box indicates the exon of a coding gene, the light blue box indicates the
intron, the gray line shows the intergenic region and the arrow shows the transcriptional direction. Illustrations below show the hairpin structures of microRNA
(miRNA) precursors. The mature sequences of miRNA are marked in red. The number in each semicircle represents the predicted target clusters by each miRNA.
(c) A northern blot of low molecular weight RNA treated with (+) or without ()) Terminator Exonuclease (TE) was hybridized to detect miR2118 and miR2275. Trans-
acting small interfering RNA-AUXIN RESPONSE FACTORs (tasiR-ARFs) and miR390 were used as the mono-phosphate control, with this 5¢ end conferred via
cleavage by Dicer-like enzymes. U6 was used as the loading control. ZH11 and Nipp represent Zhonghua 11 and Nipponbare accessions.
(d) RNA gel blot analysis of miR2118 and miR2275 in panicles of OsDCL1IRs and osdcl4-1. U6 and 5S/tRNA were used as the loading controls; miR168 was used as
the control for OsDCL1IRs.
Phased small RNA clusters biogenesis in rice 9
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

phased small RNA biogenesis, our results demonstrated
that the OsDCL1-dependent 22-nucleotide miRNAs trigger
the cleavage for a vast number of phased small RNA pre-
cursors (Figures 5, 6 and 8). After the initial cleavage direc-
ted by the triggers, it would be reasonable to expect that
RDR might be required for second-strand RNA synthesis,
providing suitable substrates for DCLs. Comparing the
moderate and sharp decrease of triggering miR2118 and
mature 21-nucletiode phased small RNAs in osdcl4-1,
respectively (Figures 2b, 3a and 6d), we proposed that
OsDCL4 is the direct DCL to process the maturation of
21-nucleotide phased small RNAs, while the moderate
reduction of triggering miR2118 might be indirect effect
which could be due to lack of certain types of cells in osdcl4-
1 mutant (Figure 8). For 24-nucleotide phased small RNA
biogenesis, the processing was carried out by OsDCL3b but
not OsDCL3a (Figure 4, 8). This is also consistent with Os-
DCL3b being expressed specifically in panicle and early seed
development stages (Kapoor et al., 2008). In addition, that
the 24-nucleotide phased small RNAs show no obvious
change in OsDCL1IRs (Figure 4) may be attributed the weak
RNAi lines that we used, in which the triggering miR2275 is
only moderately decreased (Figure 6d).
Convergent evolution of DCL3b and 24-nucleotide phased
small RNAs in monocots
In rice, two DCL3 family members have distinct roles in the
generation of 24-nucleotide small RNAs. OsDCL3a plays
conserved roles in producing 24-nucleotide unphased
small RNAs, including long miRNAs and heterochromatin
associated siRNAs (hc-siRNAs) such as CentO siRNAs
(Figure 4) (Wu et al., 2010), whereas OsDCL3b specifically
acts in 24-nucleotide phased small RNA biogenesis (Fig-
ure 4). Arabidopsis has a single DCL3, which is more
similar to OsDCL3a (Figure S4a). In addition, both maize
and Brachypodium genomes encode two DCL3 homologs,
which are similar to OsDCL3a and OsDCL3b (Figure S4a).
Clusters of 21- and 24-nucleotide phased small RNAs have
only been reported to exist in the developing young pan-
icles of rice and Brachypodium distachyon (Figure S4a)
(Johnson et al., 2009; Vogel, 2010). Considering that maize
has miR2118 and miR2275 as well as DCL102, the homolog
of OsDCL3b, we speculate that 21- and 24-nucleotide
phased small RNAs also exist in maize (Johnson et al.,
2009) (Figure S4a). We propose that the ancestral DCL3
gene, which has the same functions as that of Arabidopsis,
was subjected to a gene duplication event in monocots
after the divergence of dicots and monocots. During
monocot genome evolution, one paralog, DCL3a, with
constitutive expression patterns may have retained the
original functions of DCL3 in the production of 24-nucleo-
tide hc-siRNAs; whereas DCL3b, which is preferentially
expressed in panicles and early seeds, has acquired the
function of producing stamen-specific 24-nucleotide
phased small RNAs. Therefore, the divergent evolution of
DCL3b might be a prerequisite for the generation of 24-
nucleotide phased small RNAs in monocots, suggesting
the coevolution of DCL3b and 24-nucleotide phased small
RNAs.
Phased small RNAs in rice reproductive development
To date, several types of small RNAs have been reported
to be expressed and function specifically in the reproduc-
tive organs of animals and plants. In animals, piwi-inter-
acting RNAs (piRNAs) and 21U small RNAs are
preferentially expressed in the germline and play impor-
tant roles in gametogenesis (Girard et al., 2006; Batista
et al., 2008). In Arabidopsis, a class of retrotranspo-
son-derived 21-nucleotide siRNAs in pollen regulates
transposon activity in gametes (Slotkin et al., 2009), AGO9-
dependent small RNAs play a crucial role in specifying cell
fates in the Arabidopsis ovule (Olmedo-Monfil et al., 2010),
and a sperm-specific natural cis-antisense siRNA (cis-nat-
siRNA) is essential for double fertilization (Ron et al.,
2010). In addition, a type of 24-nucleotide RNA polymerase
IV (Pol IV)-dependent siRNA is expressed uniparentally in
the developing endosperm, providing a link between
genomic imprinting and RNA silencing (Mosher et al.,
2009). In rice, 21- and 24-nucleotide phased small RNAs
are preferentially expressed in panicles (Figure 7a) (John-
son et al., 2009). Furthermore, we show that the trigger
miRNAs for panicle-specific 21- and 24-nucleotide phased
Figure 8. A model of 21- and 24-nucleotide phased small RNA biogenesis in
rice.
The trigger precursors are processed by OsDCL1 into 22-nucleotide mature
miR2118 and miR2275. The 22-nucleotide triggers can initiate the cleavage of
21- and 24-nucleotide phased small RNA primary transcripts generated by
RNA polymerase II (Pol II). Unknown RDR protein(s) may act on the cleavage
transcripts to synthesize the second strand to form double-stranded RNAs as
a substrates for DCLs. OsDCL4 produces 21-nucleotide phased small RNAs
and OsDCL3b is responsible for 24-nucleotide phased small maturation.
10 Xianwei Song et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

small RNAs are mainly or exclusively expressed in sta-
mens (Figure 7a), suggesting that both sizes of phased
small RNAs are preferentially present and play roles in
stamens. Here, we tested more than 50 of the potential
21-nucleotide phased small RNA targets by 5¢-RACE and
also did a genome-wide scan using PARE/degradome data;
this analysis found that none of the predicted cleavages on
mRNAs were confirmed, which indicated that 21-nucleo-
tide phased small RNAs might function in a non-cleavage-
based manner. Although the direct targets of phased small
RNAs in rice remain to be elucidated, their tissue-speci-
ficity suggests that they may act at a specific stage during
male reproductive development. What the phased small
RNAs that match to thousands of genomic loci might
contribute to plant reproduction remains a major challenge
for the future.
EXPERIMENTAL PROCEDURES
Plant materials
Rice plants including O. sativa ssp. indica cv. 93-11, osdcl4-1(backcrossed to 93-11 three times from L16S), O. sativa ssp.japonica cv. Nipponbare, OsDCL1IRs, OsDCL3aIRs, OsDCL3bIRs andOsDCL4IRs were grown in the field as previously described (Liuet al., 2005, 2007). For the small RNA libraries, seedling materialsincluded the roots and aerial parts at 6 days after germination(DAG); panicle tissue was �4 cm in length, mainly comprising thefloral organs at differentiation stage In7 (Itoh et al., 2005). All thecollected materials were instantaneously frozen in liquid nitrogenand stored at )80�C until total RNA was extracted. Floral organs ofrice were separated from Zhonghua11 (the same subspecies withNipponbare) in stage An8 (Itoh et al., 2005), and those of maize isfrom B73.
Construction of RNAi vector and plant transformation
The binary OsDCL3a and OsDCL3b RNAi vectors were constructedas previously described (Liu et al., 2005).The fragments to targeteach homologous region of rice OsDCL3a and OsDCL3b wereobtained using primers as follows. OsDCL3a, cx0022: 5¢-CGCGGATCCAGTTCCAGAAATTGTA-3¢. cx0023: 5¢-CCGCTCGAGCATTAGGTACTTTTGATCT-3¢; OsDCL3b, cx0024: 5¢-CGCGGATCCTACTACATGTGTTCTTGCTT-3¢; cx0025: 5¢-CGCGGATCCTACTACATGTGTTCTTGCTT-3¢. Rice transformation and regeneration were performed aspreviously published (Hiei et al., 1994). The generated transgenicplants were confirmed by polymerase chain reaction (PCR) as pre-viously described (Liu et al., 2005).
Small RNA library construction, sequencing and
data analysis
Total RNA was extracted from seedlings and panicles using TRIZOL
reagent (Invitrogen, http://www.invitrogen.com/). Small RNA prep-aration was as previously described (Liu et al., 2005). The purifiedproduct was submitted for sequencing on an Illumina 1G analyzerafter a quality control (QC) assay; sequencing was performed at theBeijing Genomics Institution at Shenzhen.
The data for this article have been deposited at the National Centerfor Biotechnology Information Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo/) under accession number GSE22763.The Illumina sequencing raw data were trimmed by removingadaptor sequences to generate a set of ‘clean’ reads. All the reads
were mapped to either the 93-11 (http://rice.genomics.org.cn/) orNipponbare (TIGR release version 5; http://www.tigr.org/) genomeusing SOAPaligner (http://soap.genomics.org.cn). Reads in the sizerange of 18–26 nucleotides and perfectly matched to the ricegenome, excluding those matching tRNAs, rRNAs, snRNA andsnoRNA (t/r/sn/snoRNA), were retained for further analysis. Annota-tion was done with Rfam6 (http://www.sanger.ac.uk/Software/Rfam), miRBase 13.0 (http://microrna.sanger.ac.uk/), RepeatMasker(http://www.repeatmasker.org) and the TIGR rice genome annota-tion (TIGR release version 5; http://www.tigr.org/).
After removing t/rRNA matching reads and those with 20 or morematches to the 93-11 or Nipponbare genomes, respectively, theremaining ‘clean’ small RNA reads were used for phasing analysisas described previously but with modifications (Howell et al., 2007;De Paoli et al., 2009).
Phylogenetic analysis
Full-length protein sequences were used for phylogenetic analysis.The sequences were downloaded from ChromDB (http://www.chromdb.org). Alignments of protein sequences were per-formed using ClustalX version 1.81 with default parameters (gapopening, 10.00; gap extension, 0.20; delay divergent sequences,30%; DNA transition weight, 0.50). A bootstrapped phylogenetictree was constructed by the MEGA2 program with the unweightedpair group method and the arithmetic mean method. The number ofbootstrap replicates was 1000.
RNA ligase mediated 5¢-RACE
RNA ligase-mediated 5¢-RACE was performed as previouslydescribed (Yoshikawa et al., 2005). Primers used are listed inTables S4 and S6.
RT-PCR and real-time PCR
Total RNA extraction and reverse transcription were performed aspreviously described (Liu et al., 2005). Transcripts levels weremeasured by RT-PCR and real-time PCR. Real-time PCR analysis wasperformed using the CFX96� Real-Time PCR System (Bio-Rad,http://www.bio-rad.com/) and SYBR Green I (Invitrogen, S-7567).The PCR was performed using hot-start Taq DNA polymerase(DR007B, TaKaRa, http://www.takara-bio.com/). For each sample,quantifications were made in triplicate. Melt curves were read at theend of each amplification by steps of 0.3�C from 65 to 95�C to ensurethat the quantifications were derived from real PCR products andnot primer dimmers. Primers used were as follows: for OsACTIN,cx909 (5¢-CCAATCGTGAGAAGATGACCCA-3¢) and cx910 (5¢-CCATCAGGAAGCTCGTAGCTCT-3¢), for OsDCL4, cx0758 (5¢-GGACTAGTTACACGAACGTCCTCTTCTTTTGGTAGGT-3¢) and cx0857 (5¢-CGATGAGAGAACTTCGAGAGCT-3¢), for OsDCL3a, cx0022(5¢-CGCGGATCCAGTTCCAGAAATTGTA-3¢) and cx0274(5¢-CACTCGCAGTGTTCCACACAAA-3¢), for OsDCL3b, cx0024(5¢-CGCGGATCCTACTACATGTGTTCTTGCTT-3¢) and cx0275(5¢-GCAAGTACATGCTTTCCGTGT-3¢).
Small RNA recognizing motif analysis, folding and
Weblogo generation
The most conserved nucleotides (5¢-TGGGAGGCATCAGGA-3¢, fromthe 7th to 21st position of the motif sequence) of the 21-nucleotidephased small RNA motif were used to scan against all the smallRNAs in our libraries having a size between 20 and 24 nucleotidesand cloned in the set of four 93-11 and Nipponbare panicle libraries.Patscan (Dsouza et al., 1997) was used to identify a conserved motifwith the maximum score at 4.5 (in which a G:U mismatch counts as
Phased small RNA clusters biogenesis in rice 11
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

0.5 points, and other mismatches count for 1 point). A similarstrategy was applied to search for small RNAs matching the24-nucleotide phased small RNA motif; in this case, ‘GAGAT-ATTGGAG’ was used as the conserved sequence. All the smallRNAs were mapped to both 93-11 and TIGR5 genomes, andpotential precursor sequences were derived using the small RNAsequence plus 150 nucleotides upstream and 150 nucleotidesdownstream; this potential precursor was passed to Mfold (Zuker,2003) for hairpin structure folding. Cleavage sites experimentallyvalidated or predicted by MEME were extracted with 42 nucleotidesupstream and 40 nucleotides downstream to illustrate the con-served motif using Weblogo online tool (http://weblogo.berke-ley.edu/).
Identification of signature derived from motif-containing
phased small RNA clusters in PARE/Degradome library
Small RNA and PARE/degradome signatures (Zhou et al., 2010) weremapped to the 93-11 genome and the most abundant of degradomesignatures in the phasing RNA clusters were kept. From the center ofthe degradome signatures, 82-nucleotide sequence strings weregenerated, using the 22-nucleotide signature plus 30 nucleotidesupstream and 30 nucleotides downstream. The most conserved 17-nucleotide seed sequence, derived from the Weblogo analysis, wasused to scan the tags for candidates with a Perl script performing thepattern search using a maximum penalty score of six (based on themismatches described above). Subsequently, the candidates forgood miRNA/target matches were further filtered by restricting thescore as previously described (Jones-Rhoades and Bartel, 2004).
Small RNA gel blot analysis
All of the small RNA hybridization experiments were carried out aspreviously described (Liu et al., 2005). The LNAs used in this paperwere as follows: LNA53 (gatcaggCccaaCaAggaCa), LNA42(5¢-gtgcttgtcctTgtTgggcCtGata-3¢), LNA12 (5¢-gcttctCtcCtCcccgaag-3¢), LNA58 (5¢-tagGaatggGaggcatcagGaa-3¢), LNA64 (5¢-tgaGatattGgagaaaaaCaag-3¢), LNA3 (tasiR-ARFs) (5¢-gtctcgcCgcttCtcagtCt-ggc-3¢). The uppercase letters represent the bases which weremodified LNAs. Other end-labeled oligos were used to detect miR168(cx0161 5¢-GTCCCGATCTGCACCAAGCGA-3¢) and U6 (cx32565¢-TGTATCGTTCCAATTTTATCGGATGT-3¢), respectively. The probecorresponding to CentO siRNA was prepared as previous described(Liu et al., 2005).
Quantification of the RNAs was carried by scanning the phosphorscreens with Typhoon Trio (GE Healthcare, http://www.gehealth-care.com/) and generating the density readings of RNAs bands withthe help of ImageQuant 5.2 software (GE Healthcare). The relativelevels were then determined by setting the value of Nipponbare as 1and calculate the values of other samples accordingly.
5¢-Phosphate-dependent exonuclease (Terminator) was pur-chased from Epicentre Biotechnologies (http://www.epibio.com/)and experiments were performed according to the manufacturer’sinstructions.
ACKNOWLEDGEMENTS
The authors would like to thank Shouhong Sun and Lianfeng Gufrom the Institute of Genetics and Developmental Biology for theirtechnical support. This work was supported by National BasicResearch Program of China (grant no. 2009CB941500 to XC), byNational Natural Science Foundation of China (grant nos. 30921061and 30870534 to XC), by Department of Agriculture of China (grantnos. 2008ZX08009-001 and 2011ZX08009-001 to CL), by 863 project(grant no. 2007AA10Z142 to CL) and by NSF PGRP award 0701745 toBCM and USDA grant no. 2007-01991 to PJG.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Flowchart for identification of phased small RNA clusters.Figure S2. Abundance and phasing scores for each position of TAS3in seedlings and panicles of 93-11 versus osdcl4-1.Figure S3. Distribution of 21- and 24-nucleotide phased small RNAclusters in 93-11.Figure S4. The knock down of OsDCL3a and OsDCL3b.Table S1. Summary of small RNA profiling.Table S2. Summary of phased small RNA clusters from 19- to25-nucleotide intervals in rice and Arabidopsis.Table S3. Profiles of phased small RNA clusters with intervals of 21and 24 nucleotides.Table S4. Primers of RNA ligase mediated-5¢ rapid amplification ofcDNA ends (RLM 5¢-RACE) for 21-nucleotide phased small RNAclusters.Table S5. The validation of motif-mediated cleavage for 21-nucle-otide phased small RNAs from the parallel analysis of RNA ends(PARE)/degradome data. (Please see separate Excel file for thistable).Table S6. Primers of RNA ligase mediated-5¢ rapid amplification ofcDNA ends (RLM 5¢-RACE) for 24-nucleotide phased small RNA clusters.Table S7. The validation of motif-mediated cleavage for 24-nucleo-tide phased small RNAs from the parallel analysis of RNA ends(PARE)/degradome data. (Please see separate Excel file for this table)Table S8. Location of miR2118 in riceTable S9. Location of miR2275 in ricePlease note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Allen, E., Xie, Z., Gustafson, A.M. and Carrington, J.C. (2005) microRNA-
directed phasing during trans-acting siRNA biogenesis in plants. Cell, 121,
207–221.
Axtell, M.J., Jan, C., Rajagopalan, R. and Bartel, D.P. (2006) A two-hit trigger
for siRNA biogenesis in plants. Cell, 127, 565–577.
Bailey, T.L. and Elkan, C. (1994) Fitting a mixture model by expectation
maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst.
Mol. Biol. 2, 28–36.
Batista, P.J., Ruby, J.G., Claycomb, J.M. et al. (2008) PRG-1 and 21U-RNAs
interact to form the piRNA complex required for fertility in C. elegans. Mol.
Cell, 31, 67–78.
Baulcombe, D. (2004) RNA silencing in plants. Nature, 431, 356–363.
Carthew, R.W. and Sontheimer, E.J. (2009) Origins and Mechanisms of
miRNAs and siRNAs. Cell, 136, 642–655.
Chen, H.M., Li, Y.H. and Wu, S.H. (2007) Bioinformatic prediction and exper-
imental validation of a microRNA-directed tandem trans-acting siRNA
cascade in Arabidopsis. Proc. Natl. Acad. Sci. USA, 104, 3318–3323.
Chen, H.M., Chen, L.T., Patel, K., Li, Y.H., Baulcombe, D.C. and Wu, S.H. (2010)
22-Nucleotide RNAs trigger secondary siRNA biogenesis in plants. Proc.
Natl. Acad. Sci. USA, 107, 15269–15274.
Crooks, G.E., Hon, G., Chandonia, J.M. and Brenner, S.E. (2004) WebLogo: a
sequence logo generator. Genome Res, 14, 1188–1190.
Cuperus, J.T., Carbonell, A., Fahlgren, N., Garcia-Ruiz, H., Burke, R.T., Takeda,
A., Sullivan, C.M., Gilbert, S.D., Montgomery, T.A. and Carrington, J.C.
(2010) Unique functionality of 22-nt miRNAs in triggering RDR6-dependent
siRNA biogenesis from target transcripts in Arabidopsis. Nat. Struct. Mol.
Biol. 17, 997–1003.
De Paoli, E., Dorantes-Acosta, A., Zhai, J., Accerbi, M., Jeong, D.H., Park, S.,
Meyers, B.C., Jorgensen, R.A. and Green, P.J. (2009) Distinct extremely
abundant siRNAs associated with cosuppression in petunia. RNA, 15,
1965–1970.
12 Xianwei Song et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x

Dsouza, M., Larsen, N. and Overbeek, R. (1997) Searching for patterns in
genomic data. Trends Genet. 13, 497–498.
Dunoyer, P., Himber, C. and Voinnet, O. (2005) DICER-LIKE 4 is required for RNA
interference and produces the 21-nucleotide small interfering RNA com-
ponent of the plant cell-to-cell silencing signal. Nat. Genet. 37, 1356–1360.
Elbashir, S.M., Lendeckel, W. and Tuschl, T. (2001) RNA interference is
mediated by 21- and 22-nucleotide RNAs. Genes Dev. 15, 188–200.
Garcia-Ruiz, H., Takeda, A., Chapman, E.J., Sullivan, C.M., Fahlgren, N.,
Brempelis, K.J. and Carrington, J.C. (2010) Arabidopsis RNA-dependent
RNA polymerases and dicer-like proteins in antiviral defense and small
interfering RNA biogenesis during Turnip Mosaic Virus infection. Plant
Cell, 22, 481–496.
Gasciolli, V., Mallory, A.C., Bartel, D.P. and Vaucheret, H. (2005) Partially
redundant functions of Arabidopsis DICER-like enzymes and a role
for DCL4 in producing trans-acting siRNAs. Curr. Biol. 15, 1494–1500.
Girard, A., Sachidanandam, R., Hannon, G.J. and Carmell, M.A. (2006) A
germline-specific class of small RNAs binds mammalian Piwi proteins.
Nature, 442, 199–202.
He, G., Zhu, X., Elling, A.A. et al. (2010) Global epigenetic and transcriptional
trends among two rice subspecies and their reciprocal hybrids. Plant Cell,
22, 17–33.
Henderson, I.R., Zhang, X., Lu, C., Johnson, L., Meyers, B.C., Green, P.J. and
Jacobsen, S.E. (2006) Dissecting Arabidopsis thaliana DICER function in
small RNA processing, gene silencing and DNA methylation patterning.
Nat. Genet. 38, 721–725.
Hiei, Y., Ohta, S., Komari, T. and Kumashiro, T. (1994) Efficient transformation
of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis
of the boundaries of the T-DNA. Plant J. 6, 271–282.
Howell, M.D., Fahlgren, N., Chapman, E.J., Cumbie, J.S., Sullivan, C.M.,
Givan, S.A., Kasschau, K.D. and Carrington, J.C. (2007) Genome-wide
analysis of the RNA-DEPENDENT RNA POLYMERASE6/DICER-LIKE4 path-
way in Arabidopsis reveals dependency on miRNA- and tasiRNA-directed
targeting. Plant Cell, 19, 926–942.
Itoh, J., Nonomura, K., Ikeda, K., Yamaki, S., Inukai, Y., Yamagishi, H., Kitano,
H. and Nagato, Y. (2005) Rice plant development: from zygote to spikelet.
Plant Cell Physiol. 46, 23–47.
Johnson, C., Kasprzewska, A., Tennessen, K., Fernandes, J., Nan, G.L., Wal-
bot, V., Sundaresan, V., Vance, V. and Bowman, L.H. (2009) Clusters and
superclusters of phased small RNAs in the developing inflorescence of rice.
Genome Res. 19, 1429–1440.
Jones-Rhoades, M.W. and Bartel, D.P. (2004) Computational identification of
plant microRNAs and their targets, including a stress-induced miRNA. Mol.
Cell, 14, 787–799.
Kapoor, M., Arora, R., Lama, T., Nijhawan, A., Khurana, J.P., Tyagi, A.K. and
Kapoor, S. (2008) Genome-wide identification, organization and phyloge-
netic analysis of Dicer-like, Argonaute and RNA-dependent RNA Polymer-
ase gene families and their expression analysis during reproductive
development and stress in rice. BMC Genomics, 9, 451.
Kurihara, Y., Takashi, Y. and Watanabe, Y. (2006) The interaction between
DCL1 and HYL1 is important for efficient and precise processing of pri-
miRNA in plant microRNA biogenesis. RNA, 12, 206–212.
Lagos-Quintana, M., Rauhut, R., Lendeckel, W. and Tuschl, T. (2001) Identi-
fication of novel genes coding for small expressed RNAs. Science, 294,
853–858.
Lau, N.C., Lim, L.P., Weinstein, E.G. and Bartel, D.P. (2001) An abundant class
of tiny RNAs with probable regulatory roles in Caenorhabditis elegans.
Science, 294, 858–862.
Lee, R.C. and Ambros, V. (2001) An extensive class of small RNAs in Ca-
enorhabditis elegans. Science, 294, 862–864.
Lister, R., O’Malley, R.C., Tonti-Filippini, J., Gregory, B.D., Berry, C.C., Millar,
A.H. and Ecker, J.R. (2008) Highly Integrated Single-Base Resolution Maps
of the Epigenome in Arabidopsis. Cell, 133, 523–536.
Liu, B., Li, P., Li, X., Liu, C., Cao, S., Chu, C. and Cao, X. (2005) Loss of function
of OsDCL1 affects microRNA accumulation and causes developmental
defects in rice. Plant Physiol. 139, 296–305.
Liu, B., Chen, Z., Song, X. et al. (2007) Oryza sativa dicer-like4 reveals a key
role for small interfering RNA silencing in plant development. Plant Cell, 19,
2705–2718.
Margis, R., Fusaro, A.F., Smith, N.A., Curtin, S.J., Watson, J.M., Finnegan, E.J.
and Waterhouse, P.M. (2006) The evolution and diversification of Dicers in
plants. FEBS Lett. 580, 2442–2450.
Meyers, B.C., Axtell, M.J., Bartel, B. et al. (2008) Criteria for Annotation of
Plant MicroRNAs. Plant Cell, 20, 3186–3190.
Mosher, R.A., Melnyk, C.W., Kelly, K.A., Dunn, R.M., Studholme, D.J. and
Baulcombe, D.C. (2009) Uniparental expression of PolIV-dependent siRNAs
in developing endosperm of Arabidopsis. Nature, 460, 283–286.
Nagasaki, H., Itoh, J., Hayashi, K. et al. (2007) The small interfering RNA
production pathway is required for shoot meristem initiation in rice. Proc.
Natl Acad. Sci. USA, 104, 14867–14871.
Nobuta, K., Venu, R.C., Lu, C. et al. (2007) An expression atlas of rice mRNAs
and small RNAs. Nat. Biotechnol. 25, 473–477.
Olmedo-Monfil, V., Duran-Figueroa, N., Arteaga-Vazquez, M., Demesa-Are-
valo, E., Autran, D., Grimanelli, D., Slotkin, R.K., Martienssen, R.A. and
Vielle-Calzada, J.P. (2010) Control of female gamete formation by a small
RNA pathway in Arabidopsis. Nature, 464, 628–632.
Park, W., Li, J., Song, R., Messing, J. and Chen, X. (2002) CARPEL FACTORY, a
Dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in
Arabidopsis thaliana. Curr. Biol. 12, 1484–1495.
Rajagopalan, R., Vaucheret, H., Trejo, J. and Bartel, D.P. (2006) A diverse and
evolutionarily fluid set of microRNAs in Arabidopsis thaliana. Genes Dev.
20, 3407–3425.
Ramachandran, V. and Chen, X. (2008) Small RNA metabolism in Arabidopsis.
Trends Plant Sci. 13, 368–374.
Reinhart, B.J., Weinstein, E.G., Rhoades, M.W., Bartel, B. and Bartel, D.P.
(2002) MicroRNAs in plants. Genes Dev. 16, 1616–1626.
Ron, M., Saez, M.A., Williams, L.E., Fletcher, J.C. and McCormick, S. (2010)
Proper regulation of a sperm-specific cis-nat-siRNA is essential for double
fertilization in Arabidopsis. Genes Dev. 24, 1010–1021.
Schauer, S.E., Jacobsen, S.E., Meinke, D.W. and Ray, A. (2002) DICER-LIKE1:
blind men and elephants in Arabidopsis development. Trends Plant Sci. 7,
487–491.
Slotkin, R.K., Vaughn, M., Borges, F., Tanurdzic, M., Becker, J.D., Feijo, J.A.
and Martienssen, R.A. (2009) Epigenetic reprogramming and small RNA
silencing of transposable elements in pollen. Cell, 136, 461–472.
Urayama, S., Moriyama, H., Aoki, N., Nakazawa, Y., Okada, R., Kiyota, E.,
Miki, D., Shimamoto, K. and Fukuhara, T. (2010) Knock-down of OsDCL2 in
rice negatively affects maintenance of the endogenous dsRNA virus, Oryza
sativa endornavirus. Plant Cell Physiol. 51, 58–67.
Vazquez, F., Vaucheret, H., Rajagopalan, R., Lepers, C., Gasciolli, V., Mallory,
A.C., Hilbert, J.L., Bartel, D.P. and Crete, P. (2004) Endogenous trans-acting
siRNAs regulate the accumulation of Arabidopsis mRNAs. Mol. Cell, 16, 69–
79.
Vogel, J. (2010) Genome sequencing and analysis of the model grass
Brachypodium distachyon. Nature, 463, 763–768.
Wu, L., Zhou, H., Zhang, Q., Zhang, J., Ni, F., Liu, C. and Qi, Y. (2010) DNA
methylation mediated by a microRNA pathway. Mol. Cell, 38, 465–475.
Xie, Z., Johansen, L.K., Gustafson, A.M., Kasschau, K.D., Lellis, A.D., Zilber-
man, D., Jacobsen, S.E. and Carrington, J.C. (2004) Genetic and functional
diversification of small RNA pathways in plants. PLoS Biol. 2, E104.
Xie, Z., Allen, E., Wilken, A. and Carrington, J.C. (2005) DICER-LIKE 4 functions
in trans-acting small interfering RNA biogenesis and vegetative phase
change in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA, 102, 12984–
12989.
Yoshikawa, M., Peragine, A., Park, M.Y. and Poethig, R.S. (2005) A pathway
for the biogenesis of trans-acting siRNAs in Arabidopsis. Genes Dev. 19,
2164–2175.
Zamore, P.D., Tuschl, T., Sharp, P.A. and Bartel, D.P. (2000) RNAi: double-
stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23
nucleotide intervals. Cell, 101, 25–33.
Zhang, W., Yi, C., Bao, W., Liu, B., Cui, J., Yu, H., Cao, X., Gu, M., Liu, M. and
Cheng, Z. (2005) The transcribed 165-bp CentO satellite is the major func-
tional centromeric element in the wild rice species Oryza punctata. Plant
Physiol. 139, 306–315.
Zhou, M., Gu, L., Li, P., Song, X., Wei, L., Chen, Z. and Cao, X. (2010) Degra-
dome sequencing reveals endogenous small RNA targets in rice (Oryza
sativa L. ssp. indica). Front. Biol. 5, 67–90.
Zhu, Q.H., Spriggs, A., Matthew, L., Fan, L., Kennedy, G., Gubler, F. and
Helliwell, C. (2008) A diverse set of microRNAs and microRNA-like small
RNAs in developing rice grains. Genome Res 18, 1456–1465.
Zuker, M. (2003) Mfold web server for nucleic acid folding and hybridization
prediction. Nucleic Acids Res. 31, 3406–3415.
Phased small RNA clusters biogenesis in rice 13
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04805.x




















