
Molecular basis of usher pore gating in Escherichia coli pilus biogenesis
9
Molecular basis of usher pore gating in Escherichia coli pilus biogenesis Ender Volkan a,b,c,d , Vasilios Kalas a,b , Jerome S. Pinkner a , Karen W. Dodson a , Nadine S. Henderson e , Thieng Pham f , Gabriel Waksman g , Anne H. Delcour f , David G. Thanassi e,1 , and Scott J. Hultgren a,b,1 a Department of Molecular Microbiology and b Center for Women’s Infectious Disease Research, Washington University School of Medicine, Saint Louis, MO 63110; c Biotechnology Research Centre and d Faculty of Pharmacy, Department of Pharmaceutical Microbiology, Cyprus International University, Nicosia, Northern Cyprus 98258, Mersin 10, Turkey; e Center for Infectious Diseases, Department of Molecular Genetics and Microbiology, Stony Brook University, Stony Brook, NY 11794; f Department of Biology and Biochemistry, University of Houston, Houston, TX 77204; and g Institute of Structural and Molecular Biology, Birkbeck College, University College London, London WC1E 7HX, United Kingdom Contributed by Scott J. Hultgren, November 7, 2013 (sent for review July 31, 2013) Extracellular fibers called chaperone-usher pathway pili are critical virulence factors in a wide range of Gram-negative pathogenic bac- teria that facilitate binding and invasion into host tissues and mediate biofilm formation. Chaperone-usher pathway ushers, which catalyze pilus assembly, contain five functional domains: a 24-stranded trans- membrane β-barrel translocation domain (TD), a β-sandwich plug do- main (PLUG), an N-terminal periplasmic domain, and two C- terminal periplasmic domains (CTD1 and 2). Pore gating occurs by a mechanism whereby the PLUG resides stably within the TD pore when the usher is inactive and then upon activation is translocated into the periplasmic space, where it functions in pilus assembly. Using antibiotic sensitivity and electrophysiology experiments, a single salt bridge was shown to function in maintaining the PLUG in the TD channel of the P pilus usher PapC, and a loop between the 12th and 13th beta strands of the TD (β12–13 loop) was found to facilitate pore opening. Mutation of the β12–13 loop resulted in a closed PapC pore, which was unable to efficiently mediate pilus assembly. Dele- tion of the PapH terminator/anchor resulted in increased OM perme- ability, suggesting a role for the proper anchoring of pili in retaining OM integrity. Further, we introduced cysteine residues in the PLUG and N-terminal periplasmic domains that resulted in a FimD usher with a greater propensity to exist in an open conformation, resulting in increased OM permeability but no loss in type 1 pilus assembly. These studies provide insights into the molecular basis of usher pore gating and its roles in pilus biogenesis and OM permeability. outer membrane usher | macromolecular assembly | bacterial pathogenesis T he β-barrel membrane proteins of Gram-negative bacteria participate in a variety of outer membrane (OM) functions, including physiological maintenance, protein folding, transport, and organelle assembly (1–5). Some OM pore proteins are in- volved in protein secretion and assembly of virulence-associated surface-exposed appendages required for adherence to host surfaces (3, 5, 6). One such class of proteins is the ushers of the chaperone-usher pathway (CUP) found in various phyla of bac- teria ranging from Proteobacteria to Cyanobacteria to Dein- ococcus-Thermus (7). Ushers are gated OM channels that function as molecular machines that convert subunit binding and folding energy into work to assemble highly stable macromo- lecular CUP fibers and facilitate their extrusion to the bacterial surface in Gram-negative bacteria. In uropathogenic Escherichia coli (UPEC), type 1 pili are critical in causing bladder infection (8), whereas P pili are important in pyelonephritis (9, 10). Type 1 and P pili are encoded by the fim and pap operons, respectively, and have been extensively used as model systems to elucidate the structural basis of chaperone-usher pilus assembly. We used both the type 1 and P pilus ushers in this study to elucidate funda- mental insights into the gating mechanism of OM ushers. P pilus assembly requires the function of a dedicated chaper- one (PapD) (11, 12) and the usher (PapC) (3, 13). The P pilus comprises a flexible-tip fibrillum made up of minor pilins with the two-domain adhesin PapG at the distal end where it can recognize Galα1–4Gal disaccharide-containing glycolipids found in the human kidney (14, 15). The P pilus tip is joined to a right- handed, helical pilus rod made up of PapA pilins (16, 17) and is anchored in the OM via the terminator/anchoring subunit PapH (18). Type 1 pili are composed of FimA subunits making up the pilus rod joined to the tip fibrillum, which contains the FimH tip adhesin joined to FimG and adapted to the FimA rod by FimF. FimC and FimD are the chaperone and usher, respectively. CUP pilus subunits are incomplete Ig-like folds missing their C-terminal β-strand. Through a process called donor strand complementation, the chaperone transiently completes the Ig fold of a subunit in the periplasm to form a binary complex, which facilitates the subunit’s proper folding and stability and results in a subunit primed for assembly into a pilus fiber (19– 21). Chaperone-subunit complexes are targeted to the OM usher, which catalyzes donor strand exchange (DSE), whereby the chaperone is displaced and the Ig fold of the polymerizing subunit is completed by its neighbor via an N-terminal exten- sion in an ATP-independent manner (19, 22). The usher has five domains: an N-terminal periplasmic domain (NTD), a trans- membrane β-barrel domain (TD), a β-sandwich plug domain (PLUG), and two periplasmic, β-sandwich C-terminal domains (CTD1 and 2) (3, 5). During the initiation of pilus assembly, a chaperone-adhesin is initially targeted to the usher NTD and then transferred from the NTD to the CTDs via catalytic dis- sociation by CTD2 (23). Significance Gram-negative bacteria use chaperone-usher pathway (CUP) pili to colonize host tissues and mediate biofilm formation. CUP ushers are outer membrane (OM) pore proteins that catalyze pilus assembly and mediate pilus extrusion in a gated fashion to maintain OM homeostasis. Using antibiotic sensitivity and electrophysiology experiments, specific residues of two ushers, FimD of type 1 pili and PapC of P pili, were identified as crucial for proper usher gating. Further, deletion of the P pilus anchor- ing/terminator subunit resulted in an open usher conformation, indicating this subunit may be a target for antivirulence com- pounds to potentiate antibiotic treatment. Identification of mechanisms that can be used to increase bacterial sensitivity to antibiotics is crucial for development of novel compounds to fight infections. Author contributions: E.V. and S.J.H. designed research; E.V., J.S.P., K.W.D., N.S.H., and T.P. performed research; E.V., V.K., J.S.P., K.W.D., N.S.H., T.P., G.W., A.H.D., and D.G.T. contributed new reagents/analytic tools; E.V. and K.W.D. analyzed data; and E.V., V.K., K.W.D., and S.J.H. wrote the paper. The authors declare no conflict of interest. 1 To whom correspondence may be addressed. E-mail: [email protected] or [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1320528110/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1320528110 PNAS Early Edition | 1 of 6 MICROBIOLOGY
Transcript of Molecular basis of usher pore gating in Escherichia coli pilus biogenesis

Molecular basis of usher pore gating in Escherichia colipilus biogenesisEnder Volkana,b,c,d, Vasilios Kalasa,b, Jerome S. Pinknera, Karen W. Dodsona, Nadine S. Hendersone, Thieng Phamf,Gabriel Waksmang, Anne H. Delcourf, David G. Thanassie,1, and Scott J. Hultgrena,b,1
aDepartment of Molecular Microbiology and bCenter for Women’s Infectious Disease Research, Washington University School of Medicine, Saint Louis, MO63110; cBiotechnology Research Centre and dFaculty of Pharmacy, Department of Pharmaceutical Microbiology, Cyprus International University, Nicosia,Northern Cyprus 98258, Mersin 10, Turkey; eCenter for Infectious Diseases, Department of Molecular Genetics and Microbiology, Stony Brook University,Stony Brook, NY 11794; fDepartment of Biology and Biochemistry, University of Houston, Houston, TX 77204; and gInstitute of Structural and MolecularBiology, Birkbeck College, University College London, London WC1E 7HX, United Kingdom
Contributed by Scott J. Hultgren, November 7, 2013 (sent for review July 31, 2013)
Extracellular fibers called chaperone-usher pathway pili are criticalvirulence factors in a wide range of Gram-negative pathogenic bac-teria that facilitate binding and invasion into host tissues and mediatebiofilm formation. Chaperone-usher pathway ushers, which catalyzepilus assembly, contain five functional domains: a 24-stranded trans-membrane β-barrel translocation domain (TD), a β-sandwich plug do-main (PLUG), an N-terminal periplasmic domain, and two C-terminal periplasmic domains (CTD1 and 2). Pore gating occurs bya mechanism whereby the PLUG resides stably within the TD porewhen the usher is inactive and then upon activation is translocatedinto the periplasmic space, where it functions in pilus assembly. Usingantibiotic sensitivity and electrophysiology experiments, a single saltbridge was shown to function in maintaining the PLUG in the TDchannel of the P pilus usher PapC, and a loop between the 12thand 13th beta strands of the TD (β12–13 loop) was found to facilitatepore opening. Mutation of the β12–13 loop resulted in a closed PapCpore, which was unable to efficiently mediate pilus assembly. Dele-tion of the PapH terminator/anchor resulted in increased OM perme-ability, suggesting a role for the proper anchoring of pili in retainingOM integrity. Further, we introduced cysteine residues in the PLUGand N-terminal periplasmic domains that resulted in a FimD usherwith a greater propensity to exist in an open conformation, resultingin increased OM permeability but no loss in type 1 pilus assembly.These studies provide insights into the molecular basis of usher poregating and its roles in pilus biogenesis and OM permeability.
outer membrane usher | macromolecular assembly | bacterial pathogenesis
The β-barrel membrane proteins of Gram-negative bacteriaparticipate in a variety of outer membrane (OM) functions,
including physiological maintenance, protein folding, transport,and organelle assembly (1–5). Some OM pore proteins are in-volved in protein secretion and assembly of virulence-associatedsurface-exposed appendages required for adherence to hostsurfaces (3, 5, 6). One such class of proteins is the ushers of thechaperone-usher pathway (CUP) found in various phyla of bac-teria ranging from Proteobacteria to Cyanobacteria to Dein-ococcus-Thermus (7). Ushers are gated OM channels thatfunction as molecular machines that convert subunit binding andfolding energy into work to assemble highly stable macromo-lecular CUP fibers and facilitate their extrusion to the bacterialsurface in Gram-negative bacteria. In uropathogenic Escherichiacoli (UPEC), type 1 pili are critical in causing bladder infection(8), whereas P pili are important in pyelonephritis (9, 10). Type 1and P pili are encoded by the fim and pap operons, respectively,and have been extensively used as model systems to elucidate thestructural basis of chaperone-usher pilus assembly. We used boththe type 1 and P pilus ushers in this study to elucidate funda-mental insights into the gating mechanism of OM ushers.P pilus assembly requires the function of a dedicated chaper-
one (PapD) (11, 12) and the usher (PapC) (3, 13). The P piluscomprises a flexible-tip fibrillum made up of minor pilins withthe two-domain adhesin PapG at the distal end where it can
recognize Galα1–4Gal disaccharide-containing glycolipids foundin the human kidney (14, 15). The P pilus tip is joined to a right-handed, helical pilus rod made up of PapA pilins (16, 17) and isanchored in the OM via the terminator/anchoring subunit PapH(18). Type 1 pili are composed of FimA subunits making up thepilus rod joined to the tip fibrillum, which contains the FimH tipadhesin joined to FimG and adapted to the FimA rod by FimF.FimC and FimD are the chaperone and usher, respectively.CUP pilus subunits are incomplete Ig-like folds missing their
C-terminal β-strand. Through a process called donor strandcomplementation, the chaperone transiently completes the Igfold of a subunit in the periplasm to form a binary complex,which facilitates the subunit’s proper folding and stability andresults in a subunit primed for assembly into a pilus fiber (19–21). Chaperone-subunit complexes are targeted to the OMusher, which catalyzes donor strand exchange (DSE), wherebythe chaperone is displaced and the Ig fold of the polymerizingsubunit is completed by its neighbor via an N-terminal exten-sion in anATP-independentmanner (19, 22). The usher has fivedomains: an N-terminal periplasmic domain (NTD), a trans-membrane β-barrel domain (TD), a β-sandwich plug domain(PLUG), and two periplasmic, β-sandwich C-terminal domains(CTD1 and 2) (3, 5). During the initiation of pilus assembly,a chaperone-adhesin is initially targeted to the usher NTD andthen transferred from the NTD to the CTDs via catalytic dis-sociation by CTD2 (23).
Significance
Gram-negative bacteria use chaperone-usher pathway (CUP)pili to colonize host tissues and mediate biofilm formation. CUPushers are outer membrane (OM) pore proteins that catalyzepilus assembly and mediate pilus extrusion in a gated fashionto maintain OM homeostasis. Using antibiotic sensitivity andelectrophysiology experiments, specific residues of two ushers,FimD of type 1 pili and PapC of P pili, were identified as crucialfor proper usher gating. Further, deletion of the P pilus anchor-ing/terminator subunit resulted in an open usher conformation,indicating this subunit may be a target for antivirulence com-pounds to potentiate antibiotic treatment. Identification ofmechanisms that can be used to increase bacterial sensitivity toantibiotics is crucial for development of novel compounds tofight infections.
Author contributions: E.V. and S.J.H. designed research; E.V., J.S.P., K.W.D., N.S.H., andT.P. performed research; E.V., V.K., J.S.P., K.W.D., N.S.H., T.P., G.W., A.H.D., and D.G.T.contributed new reagents/analytic tools; E.V. and K.W.D. analyzed data; and E.V., V.K.,K.W.D., and S.J.H. wrote the paper.
The authors declare no conflict of interest.1To whom correspondence may be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1320528110/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1320528110 PNAS Early Edition | 1 of 6
MICRO
BIOLO
GY
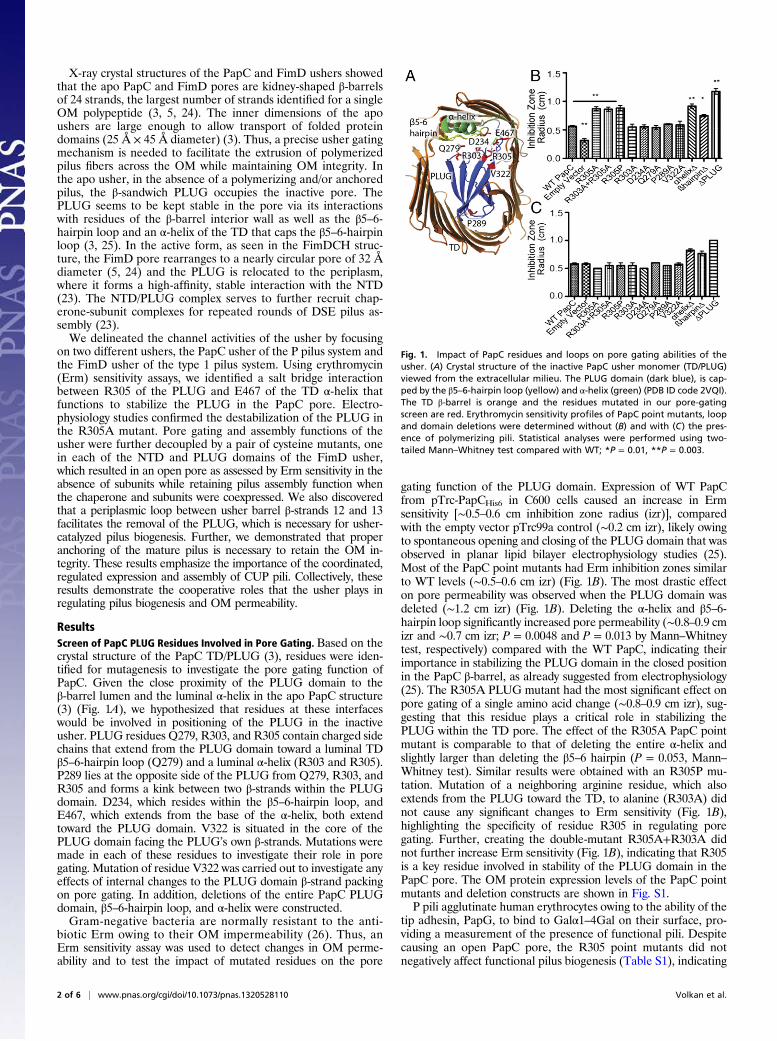
X-ray crystal structures of the PapC and FimD ushers showedthat the apo PapC and FimD pores are kidney-shaped β-barrelsof 24 strands, the largest number of strands identified for a singleOM polypeptide (3, 5, 24). The inner dimensions of the apoushers are large enough to allow transport of folded proteindomains (25 Å × 45 Å diameter) (3). Thus, a precise usher gatingmechanism is needed to facilitate the extrusion of polymerizedpilus fibers across the OM while maintaining OM integrity. Inthe apo usher, in the absence of a polymerizing and/or anchoredpilus, the β-sandwich PLUG occupies the inactive pore. ThePLUG seems to be kept stable in the pore via its interactionswith residues of the β-barrel interior wall as well as the β5–6-hairpin loop and an α-helix of the TD that caps the β5–6-hairpinloop (3, 25). In the active form, as seen in the FimDCH struc-ture, the FimD pore rearranges to a nearly circular pore of 32 Ådiameter (5, 24) and the PLUG is relocated to the periplasm,where it forms a high-affinity, stable interaction with the NTD(23). The NTD/PLUG complex serves to further recruit chap-erone-subunit complexes for repeated rounds of DSE pilus as-sembly (23).We delineated the channel activities of the usher by focusing
on two different ushers, the PapC usher of the P pilus system andthe FimD usher of the type 1 pilus system. Using erythromycin(Erm) sensitivity assays, we identified a salt bridge interactionbetween R305 of the PLUG and E467 of the TD α-helix thatfunctions to stabilize the PLUG in the PapC pore. Electro-physiology studies confirmed the destabilization of the PLUG inthe R305A mutant. Pore gating and assembly functions of theusher were further decoupled by a pair of cysteine mutants, onein each of the NTD and PLUG domains of the FimD usher,which resulted in an open pore as assessed by Erm sensitivity in theabsence of subunits while retaining pilus assembly function whenthe chaperone and subunits were coexpressed. We also discoveredthat a periplasmic loop between usher barrel β-strands 12 and 13facilitates the removal of the PLUG, which is necessary for usher-catalyzed pilus biogenesis. Further, we demonstrated that properanchoring of the mature pilus is necessary to retain the OM in-tegrity. These results emphasize the importance of the coordinated,regulated expression and assembly of CUP pili. Collectively, theseresults demonstrate the cooperative roles that the usher plays inregulating pilus biogenesis and OM permeability.
ResultsScreen of PapC PLUG Residues Involved in Pore Gating. Based on thecrystal structure of the PapC TD/PLUG (3), residues were iden-tified for mutagenesis to investigate the pore gating function ofPapC. Given the close proximity of the PLUG domain to theβ-barrel lumen and the luminal α-helix in the apo PapC structure(3) (Fig. 1A), we hypothesized that residues at these interfaceswould be involved in positioning of the PLUG in the inactiveusher. PLUG residues Q279, R303, and R305 contain charged sidechains that extend from the PLUG domain toward a luminal TDβ5–6-hairpin loop (Q279) and a luminal α-helix (R303 and R305).P289 lies at the opposite side of the PLUG from Q279, R303, andR305 and forms a kink between two β-strands within the PLUGdomain. D234, which resides within the β5–6-hairpin loop, andE467, which extends from the base of the α-helix, both extendtoward the PLUG domain. V322 is situated in the core of thePLUG domain facing the PLUG’s own β-strands. Mutations weremade in each of these residues to investigate their role in poregating. Mutation of residue V322 was carried out to investigate anyeffects of internal changes to the PLUG domain β-strand packingon pore gating. In addition, deletions of the entire PapC PLUGdomain, β5–6-hairpin loop, and α-helix were constructed.Gram-negative bacteria are normally resistant to the anti-
biotic Erm owing to their OM impermeability (26). Thus, anErm sensitivity assay was used to detect changes in OM perme-ability and to test the impact of mutated residues on the pore
gating function of the PLUG domain. Expression of WT PapCfrom pTrc-PapCHis6 in C600 cells caused an increase in Ermsensitivity [∼0.5–0.6 cm inhibition zone radius (izr)], comparedwith the empty vector pTrc99a control (∼0.2 cm izr), likely owingto spontaneous opening and closing of the PLUG domain that wasobserved in planar lipid bilayer electrophysiology studies (25).Most of the PapC point mutants had Erm inhibition zones similarto WT levels (∼0.5–0.6 cm izr) (Fig. 1B). The most drastic effecton pore permeability was observed when the PLUG domain wasdeleted (∼1.2 cm izr) (Fig. 1B). Deleting the α-helix and β5–6-hairpin loop significantly increased pore permeability (∼0.8–0.9 cmizr and ∼0.7 cm izr; P = 0.0048 and P = 0.013 by Mann–Whitneytest, respectively) compared with the WT PapC, indicating theirimportance in stabilizing the PLUG domain in the closed positionin the PapC β-barrel, as already suggested from electrophysiology(25). The R305A PLUG mutant had the most significant effect onpore gating of a single amino acid change (∼0.8–0.9 cm izr), sug-gesting that this residue plays a critical role in stabilizing thePLUG within the TD pore. The effect of the R305A PapC pointmutant is comparable to that of deleting the entire α-helix andslightly larger than deleting the β5–6 hairpin (P = 0.053, Mann–Whitney test). Similar results were obtained with an R305P mu-tation. Mutation of a neighboring arginine residue, which alsoextends from the PLUG toward the TD, to alanine (R303A) didnot cause any significant changes to Erm sensitivity (Fig. 1B),highlighting the specificity of residue R305 in regulating poregating. Further, creating the double-mutant R305A+R303A didnot further increase Erm sensitivity (Fig. 1B), indicating that R305is a key residue involved in stability of the PLUG domain in thePapC pore. The OM protein expression levels of the PapC pointmutants and deletion constructs are shown in Fig. S1.P pili agglutinate human erythrocytes owing to the ability of the
tip adhesin, PapG, to bind to Galα1–4Gal on their surface, pro-viding a measurement of the presence of functional pili. Despitecausing an open PapC pore, the R305 point mutants did notnegatively affect functional pilus biogenesis (Table S1), indicating
Fig. 1. Impact of PapC residues and loops on pore gating abilities of theusher. (A) Crystal structure of the inactive PapC usher monomer (TD/PLUG)viewed from the extracellular milieu. The PLUG domain (dark blue), is cap-ped by the β5–6-hairpin loop (yellow) and α-helix (green) (PDB ID code 2VQI).The TD β-barrel is orange and the residues mutated in our pore-gatingscreen are red. Erythromycin sensitivity profiles of PapC point mutants, loopand domain deletions were determined without (B) and with (C) the pres-ence of polymerizing pili. Statistical analyses were performed using two-tailed Mann–Whitney test compared with WT; *P = 0.01, **P = 0.003.
2 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1320528110 Volkan et al.

that the pore gating function of the PLUG domain is distinct fromits involvement in the catalysis of pilus assembly. Importantly, theErm sensitivity phenotypes of the PapC point mutants R305A,R305A+R303A, and R305P were reversed under pilus-inducingconditions, likely owing to mature/polymerizing pili blocking thePapC pore and preventing entry of the antibiotic Erm. However,the PapC PLUG deletion mutant remained Erm-sensitive, con-sistent with its inability to assemble pili. Interestingly, α-helix andβ5–6-hairpin loop deletion constructs remained Erm-sensitiveeven under pilus-inducing conditions (Fig. 1C). This may suggestthat the α-helix and β5–6-hairpin loop are involved in retaining pilion the cell surface or that their deletion may cause a change in theoccluded shape/size of the transmembrane barrel such that Ermcan still be transported even when a polymerizing pilus fiber istransiting the channel.
Electrophysiology Experiments Suggest a Possible Destabilization ofthe PLUG Domain in the R305A Mutant. As previously documented(25), the WT PapC channel is characterized by many shortopenings of various sizes, seen as downward spikes in (Fig. 2A).Few of these transitions have a conductance greater than 1 ns,and those that do are extremely transient (a few milliseconds).These small transitions have been interpreted as originating fromthe “jiggling” of the PLUG domain and/or the dynamic in-terference of the N- and C-terminal domains with the ion con-duction pathways identified at the interface of the PLUG and theTD. However, in 7 out of 10 experiments performed on R305APapC the channels displayed occasional large transitions thatlasted for several seconds (3–30 s) (Fig. 2B), in addition to theopenings of small conductance similar to those of the WTchannel. These large openings have a conductance of ∼4 ns,which is similar to the monomeric conductance of the PLUG-less PapC mutant (25), and thus could represent PLUG release.Just as was observed for the PLUG-less mutant (25), numerousflickers to lower conductance levels occur within the large openings,
probably representing a combination of the dynamic behavior ofthe released PLUG and the mobility of various other elements(loops or the β-barrel itself) within the conductance pathway.Therefore, although the behavior is not consistently observedin every sweep, the R305A channels seem to be more prone tospontaneously adopt a state in which the PLUG domain has beendisplaced and the pore is fully open for a significant amount of time.
Salt Bridge Between the Usher PLUG and α-Helix Is Important forStable Positioning of the PLUG in the PapC Pore. In the PapC crys-tal structure, the PLUG residue R305 forms a salt bridge withresidue E467, which extends from the base of the α-helix towardthe PLUG domain (Fig. 1A). An E467A mutation increased Ermsensitivity equivalent to R305A or deletion of the α-helix, sug-gesting that much of the α-helix’s contribution to PLUG’s posi-tioning in the pore is mediated through this salt bridge (Fig. 3).Consistent with this hypothesis, mutation of R305 to the acidicresidue glutamate similarly increased Erm sensitivity (∼0.8–0.9cmizr), suggesting that the observed changes in pore gating are likelydue to charge–charge interactions. We thus switched the locationsof the charged residues by creating the R305E+E467R doublemutant. This double mutant restored Erm sensitivity to WT levels(∼0.5–0.6 cm izr) (Fig. 3), revealing that reestablishment of thecharge–charge interaction between these two residues restoresstability of the PLUG in the closed usher state.
Usher Periplasmic Loop β12–13 Is Involved in Pore Gating and PilusAssembly. In the crystal structure of the FimD usher in complexwith the FimCH chaperone-adhesin complex, the usher peri-plasmic loop residues S421 and L422, found between FimD’sβ-barrel strands 12 and 13, stack against the PLUG domain,suggesting that translocation of the PLUG to the periplasm maybe stabilized by these interactions. To test this hypothesis, wereplaced the mature PapC β12–13 loop residues 430–433 witha glycine and expressed the mutant PapC from the arabinose-inducible plasmid pNH305. This resulted in a decreased Erm
Fig. 2. The R305C PapC channel displays frequent prolonged openings oflarge conductance. Electrophysiological recordings of the activity of PapCWT (A) and PapC R305A mutant (B) were obtained in 1 M KCl and 5 mMHepes, pH 7.2, at −50 mV. The current level corresponding to closed channelsis marked by C. Downward deflections represent opening events, as shownby the arrows. The segment of the R305A trace highlighted by a gray box isshown under the trace at an expanded time scale to illustrate a typicalprolonged opening of ∼4 ns.
Fig. 3. Salt bridge interaction between R305 and E467 residues is importantin stability of PLUG in PapC pore. Erm sensitivity assays of PapC salt bridgemutants reveal that disrupting the residues involved in salt bridge in-teraction gives rise to a more open pore (R305A, R305A+R303A, E467A, andR305E) compared with WT PapC. The locations of the residues forming thesalt bridge can effectively be switched to reverse the increased pore openingphenotype back to WT levels (R305E+E467R). Significance compared withWT; *P < 0.05, **P < 0.03, ***P < 0.005, Mann–Whitney test.
Volkan et al. PNAS Early Edition | 3 of 6
MICRO
BIOLO
GY

inhibition zone radius (∼0.1–0.2 cm izr) compared with thecorresponding WT PapC, which had an izr of ∼0.6, indicatinga reduction in the open conformation of the PLUG. This effectwas not due to a defect in PapC expression because immunoblotsof isolated OM carrying WT PapC or β12–13 loop mutant withanti-PapC antisera revealed similar expression levels (Fig. 4C).The mutation did not affect the ability of the usher to form astable β-barrel in the OM as determined by heat-modifiablemobility assay for β-barrel stability (27) (data not shown). Thismutation decreased the efficiency of P pilus biogenesis as de-termined by HA titers (Fig. 4D) and by analysis of pilus prepa-rations, which showed reduced levels of polymerized major pilusrod subunit PapA (Fig. S2). Our results suggest that the usherperiplasmic β12–13 loop is important to open the PLUG and/orstabilize it in an open state required for efficient pilus assembly.
Usher PLUG Domain and NTD Work in Synergy. Biophysical studiessuggested that the translocation of the PLUG to the periplasm,where it subsequently serves to recruit further chaperone-subunitcomplexes, is stabilized by an interaction with NTD (23). In theFimDCH crystal structure, NTD residue T114 and PLUG residueP281 are in close proximity (5) (Fig. 5A). Thus, we probed theNTD/PLUG interaction by mutating these residues to cysteines,hypothesizing that this would potentiate an open conformation.The point mutants did not affect FimD OM expression levels asdetermined by immunoblotting with anti-FimD antisera (Fig.5D). However, expression of the NTD and PLUG double cysteinemutant (P281C+T114C) resulted in a dramatic increase in Ermsensitivity (∼1 cm izr) (Fig. 5B) compared with theWT or a P281CFimD point mutant (∼0.5–0.6 cm izr), demonstrating that the
cysteine mutations resulted in an usher with a greater propensityto exist in an open pore conformation. Thereafter, we investigatedthe ability of P281C + T114C FimD to assemble pili. Despite theincrease in pore permeability, the P281C + T114C FimD was ableto assemble pili as determined by mannose-sensitive HA assays,similar to WT FimD (Fig. 5C). These results further suggest thatthe pore gating function of the PLUG domain can be decoupledfrom its function in catalysis.
Anchored, Mature Pili Are Necessary to Retain OM Integrity of E. coli.Previous studies revealed the importance of PapH (the terminatorsubunit of P pili) in anchoring and terminating P pili at the OM(18, 28). Biophysical studies further revealed that the PapDHcomplex was specifically recruited by the PLUG domain or theintramolecular NTD/PLUG complex but did not bind to CTD2(23). This suggested that PapH may anchor pili on the cell surfacevia interaction with the usher’s PLUG domain (23). Thus, we hy-pothesized that the PapDH–PLUG interaction may result inPLUG positioning that has ramifications on OM impermeabilityand thus deletion of PapH would affect OM permeability. Weinvestigated this hypothesis by expressing the pap operon lackingpapH on plasmid pEMM13 in MC1061 cells and providing eitherpapH or an empty vector control in trans (pEVpapH and pTrc99aplasmids, respectively) in MC1061 cells and then analyzing Ermsensitivity. EM analysis showed that the papH deletion resulted inlong pili that were shed, whereas providing papH in trans usingpEVpapH plasmid restored assembled pili to their expectedlength and abundance (Fig. 6B). Erm sensitivity assays revealedthat deletion of papH resulted in a significant increase in Ermsensitivity compared with WT (∼0.9 cm izr vs. ∼0.4 cm izr, re-spectively) (Fig. 6A). Thus, incorporation of the PapH terminatorresults in maintainence of OM impermeability, likely owing in
Fig. 4. PapC Periplasmic loop β12–13 is important for PLUG domain releaseand pilus biogenesis. (A) Crystal structure of activated FimD highlighting theβ12–13 periplasmic loop (green) stacking against the PLUG domain (darkblue). The NTD domain is removed for clarity (PDB ID code 3RFZ). (B) Ermsensitivity assays were carried out on LB agar plates (P < 0.003 for bothcomparisons). (C) Western blots carried out on normalized OM preparationsusing anti-PapC antibody reveal PapC expression in the OM (D) The β12–13periplasmic loop is required for efficient functional pilus assembly as de-termined by HA assays using human red blood cells. Statistical analyses wereperformed using two-tailed Mann–Whitney test.
Fig. 5. Pore gating and pilus assembly functions of the usher are decoupledwhile revealing in vivo NTD and PLUG cooperation. (A) FimD usher residuesT114 from NTD (cyan) and P281 from PLUG domain (dark blue) were tar-geted for cysteine mutagenesis (PDB ID code 3RFZ). Targeted residues areshown in yellow. (B) Erythromycin sensitivity assays ( Mann–Whitney test, *P <0.018). (C) The ability of FimD mutants to complement a fimD- operon(pETS6) was determined using mannose-sensitive hemagglutination assays(Mann–Whitney test, P > 0.05). (D) Immunoblot carried out using anti-FimDantibody with normalized, isolated OM fractions.
4 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1320528110 Volkan et al.

part to preventing the dissociation of pili from the cell surface andleaving behind an activated, open pore (23).
DiscussionProtein secretion across the OM of Gram-negative bacteria isa critical step in bacterial pathogenesis. We investigated PapC andFimD in P and type 1 CUP pilus biogenesis, respectively, to betterunderstand the mechanisms used by Gram-negative bacteria toassemble pili while regulating pore gating. The transmembraneβ-barrel domains of PapC and FimD are composed of 24 β-strandssurrounding a large central pore that is gated by a PLUG domain,as determined by crystal structures and planar lipid bilayer elec-trophysiology studies (3, 5, 25). The PapC central channel is 45Å ×25Å in diameter and thus would potentially allow passage ofmolecules, such as antibiotics, from the extracellular milieu acrossthe OM. Thus, strict pore gating is critical for bacterial homeo-stasis. Other β-barrel proteins that shuttle constituents across orinto the outer membrane use the energy of ATP or the membranepotential across the inner membrane to regulate a gated channel,for instance the TonB-dependent translocators (29), or as in thecase of the recently solved BamA, seem to facilitate insertion ofproteins into the outer membrane by opening/closing of the BamAβ-barrel (30). In CUP ushers, OM integrity is maintained via theexistence of a β-sandwich PLUG domain, which resides in thetransmembrane channel in the absence of pilus assembly (Fig. 7).The PLUG is also needed for catalysis of pilus assembly, workingin concert with the NTD and CTD periplasmic domains of theusher (23, 25). Polymerization of subunits from chaperone-subunitcomplexes into a final pilus structure and extrusion of the fiberacross the OM does not require either the energy of ATP or thatof the proton-motive force (22), but rather seems to be the con-sequence of concerted binding events starting with the binding ofa chaperone/adhesin complex to the usher (23, 31, 32).Our structure–function analyses revealed that the PLUG is
maintained in the pore via interactions that involve the α-helix andβ5–6-hairpin loop. Furthermore, a specific PLUG residue mutation,R305A, interferes with PLUG stabilization as determined by planarlipid bilayer electrophysiology studies, where the mutant channelis characterized by an increased tendency to produce large and
persistent openings. Residue R305 forms a salt bridge interactionwith residue E467 (Fig. 7). Disruption of this salt bridge interactionpromotes an open usher state, whereas flipping the residue iden-tities across the two positions maintains electrostatic contact andthe WT antibiotic sensitivity phenotype, confirming the criticalnature of this salt bridge as part of a mechanism of pore gating. Saltbridges were also shown to be important factors in the gating ofmuch smaller pores without a PLUG domain such as OmpC andOmpA (33, 34). Despite increased pore permeability of the R305Amutant, functional pilus biogenesis was not affected. This was alsothe result for the P281C + T114C double cysteine mutant of FimD.Although we were unable to verify that P281C formed a disulfidebond with T114C, the results imply that usher activity in pilusbiogenesis is independent of the ability of the PLUG to be stablymaintained in the usher pore, because this mutant is mostly in anopen conformation as determined by Erm sensitivity.The translocation of the PLUG domain to the periplasm seems
to be stabilized by the periplasmic loop extending from β-strands12 and 13, which stacks against the PLUG in the open confor-mation (Fig. 7) in the FimDCH structure and may help stabilizethe PLUG in an orientation that binds to the NTD in the activeusher. Mutating the β12–13 loop significantly decreased the openconformation of the pore, resulting in Erm sensitivity levels equalto vector control levels, despite mutant PapC expression levelsequal to the WT. The β12–13 loop mutation also caused a signifi-cant decrease in functional pilus assembly, presumably owing to itsinability to stably relocate the PLUG and open the pore. However,a low level of pilus assembly was still observed (Fig. 4 and Fig. S2)in the presence of chaperone-subunit complexes, indicating thatthe mutant usher is capable of carrying out the necessary confor-mational changes to open the pore and function in pilus biogenesis,albeit less efficiently.We found that deletion of the terminator subunit, PapH, caused
a significant increase in OM permeability, presumably owing tothe shedding of pili from the cell surface and the NTD/PLUGdomain interaction that leaves the usher in an open state. Inthe presence of WT pilus expression, Erm sensitivity is signifi-cantly lower. This finding suggests proper PapH-mediated pilus
Fig. 6. Importance of mature pili for OM integrity of Gram-negative bac-teria. (A) Bacteria that express P pili without PapH have significantly in-creased Erm sensitivity compared with bacteria carrying full-length pili. Nosignificant change was observed between the izr of vector controls and WTPapH (Mann–Whitney test, P > 0.5) (B) Electron micrographs revealing thedifferent pili morphologies in the presence and absence of PapH. WT pililength and abundance is observed when papH is expressed; in the absenceof PapH, scarce, longer, and shedding pili are seen. Vector control shows theabsence of P pili.
Fig. 7. Structural view of the determinants of PLUG positioning in usherstates. (A) A single salt bridge between R305 (blue surface) and E467 (orangesurface) respectively emanating from the PLUG domain (marine cartoon)and β-barrel wall (orange cartoon) extending from underneath the α-helix(green) and above the β5–6-hairpin loop (magenta) stabilizes the PLUG inthe closed, apo usher state. Results were mapped onto the structure of thePapC translocation pore (PDB ID code 2VQI). (B) The β12–13 loop (orangesurface) along with the engineered cysteine mutants at position 114 (cyansurface) within the NTD (cyan cartoon) and position 281 (blue surface) withinthe PLUG domain promotes PLUG stability in the open, active usher state.Results were mapped onto the structure of the FimD usher in complex withthe chaperone-adhesin FimCH (PDB ID code 3RFZ).
Volkan et al. PNAS Early Edition | 5 of 6
MICRO
BIOLO
GY

termination is necessary to retain OM integrity. These resultshighlight the necessity of organized CUP operons to ensurestrictly regulated expression of all subunits and suggest thatcompounds that could prevent this termination event couldincrease antibiotic efficacy and thus have therapeutic value.Overall, our studies reveal mechanisms by which OM usher
proteins regulate channel conductivity and pilus biogenesis.Results obtained in this study indicate important interactionsneeded for the usher to retain OM integrity while catalyzingefficient pilus biogenesis and extrusion to the cell surface.
Materials and MethodsBacterial Strains, Plasmids, and Culture Conditions. Bacterial strains weregrown in LB agar, Tryptic Soy Agar (TSA), or LB broth with antibiotics as ap-propriate. Detailed strain and plasmid information is provided in Table S2.
Erm Sensitivity Assays. Erm sensitivity was determined using disk diffusionassays on agar plates using discs containing Erm (15 μg; BD Biologicals). SIMaterials and Methods gives further details.
HA Titer Assays. HA titers for type 1 pili were carried out using cells after2 × 24 h of 10 mL static bacterial growth with appropriate antibiotics asdescribed previously. HA titers for P pili were performed on bacteria grownon TSA plates overnight as previously described (35).
Pili and protein expression/purification, electrophysiology, and pili assayswere done as previously described (23, 25, 36). Details can be found in SIMaterials and Methods.
ACKNOWLEDGMENTS.We gratefully acknowledge the expertise ofW. Beatty(Imaging Facility, Washington University School of Medicine). This study wasfunded by National Institutes of Health R01 Grants AI029549 and AI049450(to S.J.H.) and Grant GM062987 (to D.G.T.).
1. Ricci DP, Silhavy TJ (2012) The Bam machine: A molecular cooper. Biochim BiophysActa 1818(4):1067–1084.
2. Wimley WC (2003) The versatile beta-barrel membrane protein. Curr Opin Struct Biol13(4):404–411.
3. Remaut H, et al. (2008) Fiber formation across the bacterial outer membrane by thechaperone/usher pathway. Cell 133(4):640–652.
4. Meng G, Surana NK, St Geme JW, 3rd, Waksman G (2006) Structure of the outermembrane translocator domain of the Haemophilus influenzae Hia trimeric auto-transporter. EMBO J 25(11):2297–2304.
5. Phan G, et al. (2011) Crystal structure of the FimD usher bound to its cognate FimC-FimH substrate. Nature 474(7349):49–53.
6. Allen WJ, Phan G, Hultgren SJ, Waksman G (2013) Dissection of pilus tip assembly bythe FimD usher monomer. J Mol Biol 425(5):958–967.
7. Nuccio SP, Bäumler AJ (2007) Evolution of the chaperone/usher assembly pathway:Fimbrial classification goes Greek. Microbiol Mol Biol Rev 71(4):551–575.
8. Martinez JJ, Mulvey MA, Schilling JD, Pinkner JS, Hultgren SJ (2000) Type 1 pilus-mediated bacterial invasion of bladder epithelial cells. EMBO J 19(12):2803–2812.
9. Roberts JA, et al. (1994) The Gal(alpha 1-4)Gal-specific tip adhesin of Escherichia coliP-fimbriae is needed for pyelonephritis to occur in the normal urinary tract. Proc NatlAcad Sci USA 91(25):11889–11893.
10. Melican K, et al. (2011) Uropathogenic Escherichia coli P and Type 1 fimbriae act insynergy in a living host to facilitate renal colonization leading to nephron obstruc-tion. PLoS Pathog 7(2):e1001298.
11. Holmgren A, Bränden CI (1989) Crystal structure of chaperone protein PapD revealsan immunoglobulin fold. Nature 342(6247):248–251.
12. Lindberg F, Tennent JM, Hultgren SJ, Lund B, Normark S (1989) PapD, a periplasmictransport protein in P-pilus biogenesis. J Bacteriol 171(11):6052–6058.
13. Norgren M, Båga M, Tennent JM, Normark S (1987) Nucleotide sequence, regulationand functional analysis of the papC gene required for cell surface localization of Pappili of uropathogenic Escherichia coli. Mol Microbiol 1(2):169–178.
14. Hultgren SJ, et al. (1989) The PapG adhesin of uropathogenic Escherichia coli containsseparate regions for receptor binding and for the incorporation into the pilus. ProcNatl Acad Sci USA 86(12):4357–4361.
15. Dodson KW, et al. (2001) Structural basis of the interaction of the pyelonephriticE. coli adhesin to its human kidney receptor. Cell 105(6):733–743.
16. Båga M, et al. (1984) Nucleotide sequence of the papA gene encoding the Pap pilussubunit of human uropathogenic Escherichia coli. J Bacteriol 157(1):330–333.
17. Gong M, Makowski L (1992) Helical structure of P pili from Escherichia coli. Evidencefrom X-ray fiber diffraction and scanning transmission electron microscopy. J Mol Biol228(3):735–742.
18. Båga M, Norgren M, Normark S (1987) Biogenesis of E. coli Pap pili: papH, a minorpilin subunit involved in cell anchoring and length modulation. Cell 49(2):241–251.
19. Sauer FG, et al. (1999) Structural basis of chaperone function and pilus biogenesis.Science 285(5430):1058–1061.
20. Kuehn MJ, Normark S, Hultgren SJ (1991) Immunoglobulin-like PapD chaperone capsand uncaps interactive surfaces of nascently translocated pilus subunits. Proc NatlAcad Sci USA 88(23):10586–10590.
21. Kuehn MJ, et al. (1993) Structural basis of pilus subunit recognition by the PapDchaperone. Science 262(5137):1234–1241.
22. Jacob-Dubuisson F, Striker R, Hultgren SJ (1994) Chaperone-assisted self-assembly ofpili independent of cellular energy. J Biol Chem 269(17):12447–12455.
23. Volkan E, et al. (2012) Domain activities of PapC usher reveal the mechanism of actionof an Escherichia coli molecular machine. Proc Natl Acad Sci USA 109(24):9563–9568.
24. Geibel S, Procko E, Hultgren SJ, Baker D, Waksman G (2013) Structural and energeticbasis of folded-protein transport by the FimD usher. Nature 496(7444):243–246.
25. Mapingire OS, Henderson NS, Duret G, Thanassi DG, Delcour AH (2009) Modulatingeffects of the plug, helix, and N- and C-terminal domains on channel properties of thePapC usher. J Biol Chem 284(52):36324–36333.
26. Mao JC, Putterman M (1968) Accumulation in gram-postive and gram-negative bac-teria as a mechanism of resistance to erythromycin. J Bacteriol 95(3):1111–1117.
27. Henderson NS, Ng TW, Talukder I, Thanassi DG (2011) Function of the usher N-ter-minus in catalysing pilus assembly. Mol Microbiol 79(4):954–967.
28. Verger D, Miller E, Remaut H, Waksman G, Hultgren S (2006) Molecular mechanism ofP pilus termination in uropathogenic Escherichia coli. EMBO Rep 7(12):1228–1232.
29. Bradbeer C (1993) The proton motive force drives the outer membrane transport ofcobalamin in Escherichia coli. J Bacteriol 175(10):3146–3150.
30. Noinaj N, et al. (2013) Structural insight into the biogenesis of β-barrel membraneproteins. Nature 501(7467):385–390.
31. Saulino ET, Thanassi DG, Pinkner JS, Hultgren SJ (1998) Ramifications of kinetic par-titioning on usher-mediated pilus biogenesis. EMBO J 17(8):2177–2185.
32. Dodson KW, Jacob-Dubuisson F, Striker RT, Hultgren SJ (1993) Outer-membrane PapCmolecular usher discriminately recognizes periplasmic chaperone-pilus subunit com-plexes. Proc Natl Acad Sci USA 90(8):3670–3674.
33. Hong H, Szabo G, Tamm LK (2006) Electrostatic couplings in OmpA ion-channelgating suggest a mechanism for pore opening. Nat Chem Biol 2(11):627–635.
34. Liu N, Delcour AH (1998) The spontaneous gating activity of OmpC porin is affectedby mutations of a putative hydrogen bond network or of a salt bridge between theL3 loop and the barrel. Protein Eng 11(9):797–802.
35. Slonim LN, Pinkner JS, Brändén CI, Hultgren SJ (1992) Interactive surface in the PapDchaperone cleft is conserved in pilus chaperone superfamily and essential in subunitrecognition and assembly. EMBO J 11(13):4747–4756.
36. Henderson NS, Thanassi DG (2013) Purification of the outer membrane usher proteinand periplasmic chaperone-subunit complexes from the P and type 1 pilus systems.Methods Mol Biol 966:37–52.
6 of 6 | www.pnas.org/cgi/doi/10.1073/pnas.1320528110 Volkan et al.

Supporting InformationVolkan et al. 10.1073/pnas.1320528110SI Materials and MethodsBacterial Strains, Plasmids, and Culture Conditions. Strains used inthis study include Escherichia coli C600, E. coli MC1061, andE. coli AAEC185 (1). For storage, strains were kept in −80 °C in15% (vol/vol) glycerol. Ampicillin (100 μg/mL), kanamycin (50μg/mL), chloramphenicol (20 μg/mL), and tetracycline (10 μg/mL) were used for plasmid selection. The plasmid pTrc-Pap-CHis6, used to express and mutate C-terminally His-tagged PapC,was obtained by cloning PapC gene from pPap5 into pTrc99avector using standard PCR and recombinant techniques. Site Di-rected Mutagenesis Kit (Quikchange; Invitrogen) was used to in-troduce the point mutations in the papC gene in pTrc-PapCHis6plasmid. To express these mutants with the rest of the pap op-eron, pEV33 plasmid was used, which contains all pap operongenes from pPap33 in which the papC gene is inactivated byinsertion of an XhoI linker. This construct was cloned into thetetr, TSA inducible backbone vector from pFJ3 obtained byBglII and BamH1 digestion.A version of pSH2, termed pETS6, was used to express fim
operon lacking fimD under static conditions. FimD was com-plemented in trans using C-terminally His-tagged fimD genecloned on pTrc99a vector from pSH2 plasmid termed pEV-fimD. pEVfimD was used for all mutagenesis studies carriedout on fimD gene.pEMM13, which contains the pap operon without papH on the
pMMB91 backbone, was used to express PapH− P pili on TSA.Isopropyl β-D-1-thiogalactopyranoside (IPTG)-inducible pEV-papH plasmid containing papH gene was used to complementthe lack of PapH, whereas pTrc99a was used as a negativecontrol in MC1061 cells. pEVpapH was generated by removingthe papD gene from pTrcDH (2) by digestion with BamHI andXba enzymes. The plasmid lacking papD was treated with DNApolymerase Klenow fragment to generate blunt ends and re-ligated to create pEVpapH, which carries papH gene under theIPTG-inducible pTac promoter.
Erythromycin Sensitivity Assay. For erythromycin (Erm) sensitivityassays, 3-mL cultures of strains containing plasmids encoding theindicated PapC or FimD constructs were grown, shaking, withappropriate antibiotics for 6–8 h. Sterile cotton swabs were usedto spread bacteria on large (150- ×15-mm plates) plates (Falcon),each of which had 50 mL of LB agar along with the appropriateantibiotics for plasmid selection and 10−3 mM IPTG to inducePapC or FimD gene expression. Subsequent to spreading on theplates, tablets of the antibiotic Erm (15 μg; BD Biologicals) wereplaced on the plates and incubated overnight at 37 °C. The an-tibiotic sensitivity of each PapC mutant was obtained by mea-suring the radius of the inhibition zone, subtracting the radius ofthe tablet itself. The same procedure was used to assess eryth-romycin sensitivity under pilus-inducing conditions using pEV33and pTrc-PapCHis6 plasmids. TSA agar was used instead of LB toinduce the pap operon off of pEV33 plasmid. The same experi-mental method was used to test Erm sensitivity of C600 cellscarrying pDG2, pNH305, and pMON6235, where plates con-tained 0.1% arabinose for induction.
Membrane Extraction and Pili Preparations. C600 cells carrying WTPapC or vector control pTrc99a or PapC β12–13 loop mutant
pNH305 were transformed with pMJ2, which carries the papoperon lacking papC, and were grown overnight in LB broth. Piliwere heat-extracted as previously described (3). The pili prepswere boiled in 8 M urea SDS loading buffer and separated by gelelectrophoresis, transferred to nitrocellulose membrane. Immu-noblotting analysis was carried out using a 1:10,000 concentra-tion of WU93 antibody raised against whole P pili, whichrecognizes PapA. Outer membrane (OM) extraction for all PapCconstructs was carried out from 10−3 mM IPTG-induced 250-mLcultures, as previously described (4). Levels of PapC mutantvariants were compared with WT PapC for expression level andability to fold into a stable β-barrel in the OM. OM isolation,analysis of usher protein levels, and the heat-modifiable mobilityassay for β-barrel stability were done as previously described (5).
HA Titer Assays. HA titers for type 1 pili were carried out usingguinea pig red blood cells after 2 × 24 h, 10-mL static bacterialgrowth with appropriate antibiotics as described previously (6).HA titers for P pili were performed on bacteria grown on TSAplates overnight by serial dilution in microtiter plates as pre-viously described (7). pEVfimD plasmid encoding WT fimD orpoint mutants and empty vector control pTrc99a were trans-formed in AAEC185 E. coli cells along with the pETS6 vectorthat carries fim operon without fimD to induce pilus assembly.FimD carrying P281C + T114C mutations at N-terminal peri-plasmic domain and β-sandwich plug (PLUG) domains can as-semble pili without a significant defect as determined bymannose-sensitive HA assays. HA titers for P pili were per-formed on bacteria grown on TSA plates overnight by serialdilution in microtiter plates as previously described (7).
Electrophysiology Experiments. WT or R305A PapC dimers werepurified as described (8) and incorporated into planar lipid bilayersfor electrophysiology analysis (9). The planar lipid bilayer setupconsisted of a 0.01-mm-thick Teflon film (Goodfellows) with ∼100-μm aperture pretreated with a solution of 1% of hexadecane (TCI)in pentane (Burdick & Jackson) and sandwiched between twoTeflon chambers filled with 1.5 mL of buffer (1 M KCl and 5 mMHepes, pH 7.2). Five microliters of a 5 mg/mL asolectin (L-α-phosphatidylcholine Type II-S; Sigma) solution made in pentanewas added on each side, and the bilayer was formed over theaperture using the Montal and Mueller technique (10). Once thebilayer was formed and stabilized, 4–8 μg of pure protein dilutedin either 0.5% or 2% N-octyl-oligo-oxyethylene (octyl-POE; Ax-xora) was added to one chamber (defined as the cis side), andgentle stirring was used to promote spontaneous insertions ofproteins in the bilayer. Once channel activity was observed (<10min), stirring was stopped to limit further insertions.Electrophysiology recordings were made by measuring cur-
rent under voltage-clamp conditions (trans side grounded) withthe use of Ag/AgCl electrodes connected to an Axopatch 1D am-plifier (Axon Instruments) with a CV-4B headstage. The currentwas filtered at 500 Hz, digitized at 1.25-ms sampling intervals (ITC-18; Instrutech), and stored on a PC computer using the Acquiresoftware (Bruxton). The analysis and reading of the traces wereperformed with the Clampfit program from Axon Instruments.
Volkan et al. www.pnas.org/cgi/content/short/1320528110 1 of 3
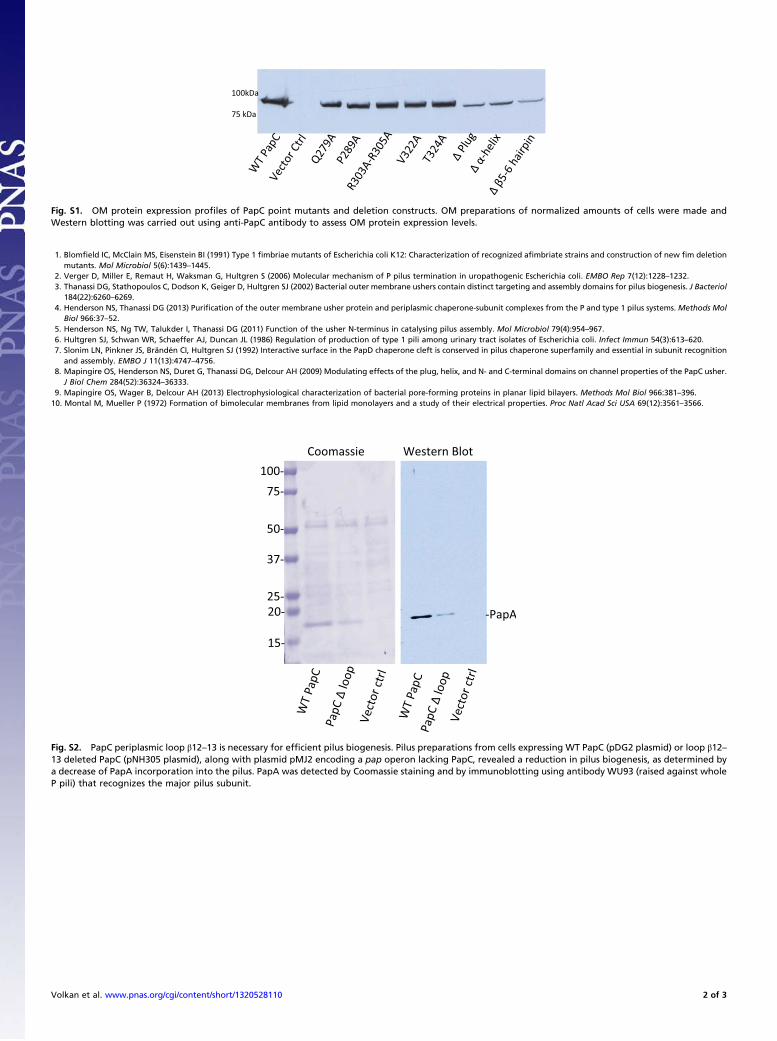
Fig. S1. OM protein expression profiles of PapC point mutants and deletion constructs. OM preparations of normalized amounts of cells were made andWestern blotting was carried out using anti-PapC antibody to assess OM protein expression levels.
1. Blomfield IC, McClain MS, Eisenstein BI (1991) Type 1 fimbriae mutants of Escherichia coli K12: Characterization of recognized afimbriate strains and construction of new fim deletionmutants. Mol Microbiol 5(6):1439–1445.
2. Verger D, Miller E, Remaut H, Waksman G, Hultgren S (2006) Molecular mechanism of P pilus termination in uropathogenic Escherichia coli. EMBO Rep 7(12):1228–1232.3. Thanassi DG, Stathopoulos C, Dodson K, Geiger D, Hultgren SJ (2002) Bacterial outer membrane ushers contain distinct targeting and assembly domains for pilus biogenesis. J Bacteriol
184(22):6260–6269.4. Henderson NS, Thanassi DG (2013) Purification of the outer membrane usher protein and periplasmic chaperone-subunit complexes from the P and type 1 pilus systems. Methods Mol
Biol 966:37–52.5. Henderson NS, Ng TW, Talukder I, Thanassi DG (2011) Function of the usher N-terminus in catalysing pilus assembly. Mol Microbiol 79(4):954–967.6. Hultgren SJ, Schwan WR, Schaeffer AJ, Duncan JL (1986) Regulation of production of type 1 pili among urinary tract isolates of Escherichia coli. Infect Immun 54(3):613–620.7. Slonim LN, Pinkner JS, Brändén CI, Hultgren SJ (1992) Interactive surface in the PapD chaperone cleft is conserved in pilus chaperone superfamily and essential in subunit recognition
and assembly. EMBO J 11(13):4747–4756.8. Mapingire OS, Henderson NS, Duret G, Thanassi DG, Delcour AH (2009) Modulating effects of the plug, helix, and N- and C-terminal domains on channel properties of the PapC usher.
J Biol Chem 284(52):36324–36333.9. Mapingire OS, Wager B, Delcour AH (2013) Electrophysiological characterization of bacterial pore-forming proteins in planar lipid bilayers. Methods Mol Biol 966:381–396.10. Montal M, Mueller P (1972) Formation of bimolecular membranes from lipid monolayers and a study of their electrical properties. Proc Natl Acad Sci USA 69(12):3561–3566.
Fig. S2. PapC periplasmic loop β12–13 is necessary for efficient pilus biogenesis. Pilus preparations from cells expressing WT PapC (pDG2 plasmid) or loop β12–13 deleted PapC (pNH305 plasmid), along with plasmid pMJ2 encoding a pap operon lacking PapC, revealed a reduction in pilus biogenesis, as determined bya decrease of PapA incorporation into the pilus. PapA was detected by Coomassie staining and by immunoblotting using antibody WU93 (raised against wholeP pili) that recognizes the major pilus subunit.
Volkan et al. www.pnas.org/cgi/content/short/1320528110 2 of 3
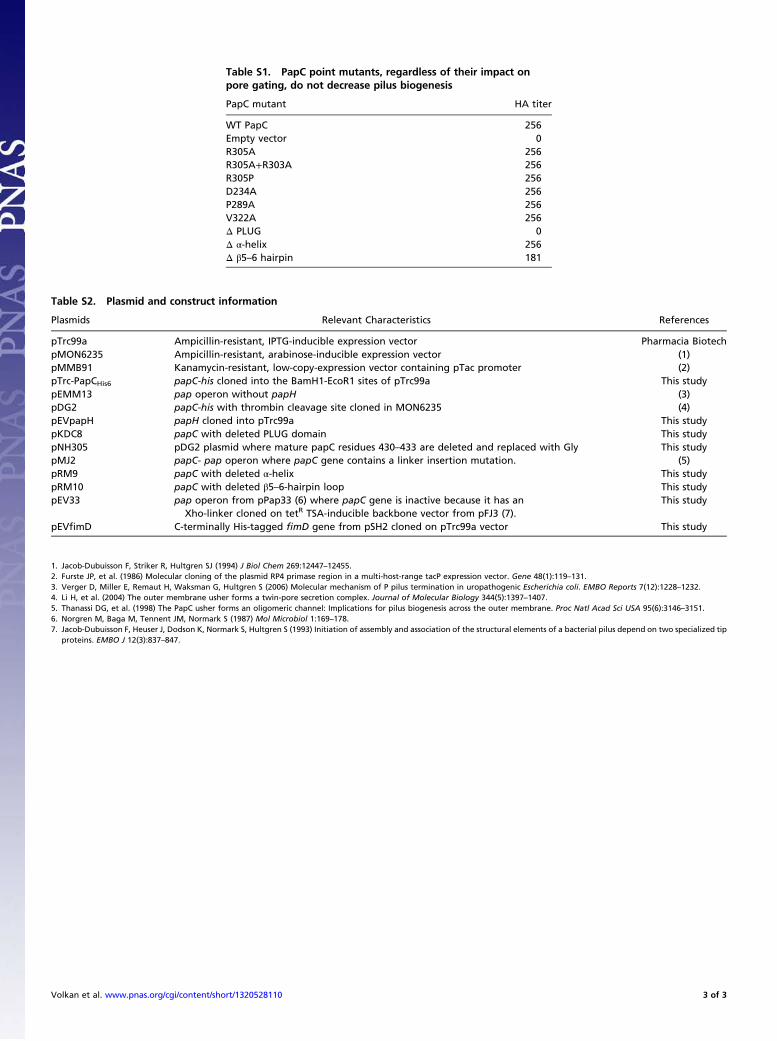
Table S1. PapC point mutants, regardless of their impact onpore gating, do not decrease pilus biogenesis
PapC mutant HA titer
WT PapC 256Empty vector 0R305A 256R305A+R303A 256R305P 256D234A 256P289A 256V322A 256Δ PLUG 0Δ α-helix 256Δ β5–6 hairpin 181
Table S2. Plasmid and construct information
Plasmids Relevant Characteristics References
pTrc99a Ampicillin-resistant, IPTG-inducible expression vector Pharmacia BiotechpMON6235 Ampicillin-resistant, arabinose-inducible expression vector (1)pMMB91 Kanamycin-resistant, low-copy-expression vector containing pTac promoter (2)pTrc-PapCHis6 papC-his cloned into the BamH1-EcoR1 sites of pTrc99a This studypEMM13 pap operon without papH (3)pDG2 papC-his with thrombin cleavage site cloned in MON6235 (4)pEVpapH papH cloned into pTrc99a This studypKDC8 papC with deleted PLUG domain This studypNH305 pDG2 plasmid where mature papC residues 430–433 are deleted and replaced with Gly This studypMJ2 papC- pap operon where papC gene contains a linker insertion mutation. (5)pRM9 papC with deleted α-helix This studypRM10 papC with deleted β5–6-hairpin loop This studypEV33 pap operon from pPap33 (6) where papC gene is inactive because it has an
Xho-linker cloned on tetR TSA-inducible backbone vector from pFJ3 (7).This study
pEVfimD C-terminally His-tagged fimD gene from pSH2 cloned on pTrc99a vector This study
1. Jacob-Dubuisson F, Striker R, Hultgren SJ (1994) J Biol Chem 269:12447–12455.2. Furste JP, et al. (1986) Molecular cloning of the plasmid RP4 primase region in a multi-host-range tacP expression vector. Gene 48(1):119–131.3. Verger D, Miller E, Remaut H, Waksman G, Hultgren S (2006) Molecular mechanism of P pilus termination in uropathogenic Escherichia coli. EMBO Reports 7(12):1228–1232.4. Li H, et al. (2004) The outer membrane usher forms a twin-pore secretion complex. Journal of Molecular Biology 344(5):1397–1407.5. Thanassi DG, et al. (1998) The PapC usher forms an oligomeric channel: Implications for pilus biogenesis across the outer membrane. Proc Natl Acad Sci USA 95(6):3146–3151.6. Norgren M, Baga M, Tennent JM, Normark S (1987) Mol Microbiol 1:169–178.7. Jacob-Dubuisson F, Heuser J, Dodson K, Normark S, Hultgren S (1993) Initiation of assembly and association of the structural elements of a bacterial pilus depend on two specialized tip
proteins. EMBO J 12(3):837–847.
Volkan et al. www.pnas.org/cgi/content/short/1320528110 3 of 3




















