
Exercise induces an increase in muscle UCP3 as a component of the increase in mitochondrial...
23
1 Exercise induces an increase in muscle UCP3 as a component of the increase in mitochondrial biogenesis Terry E. Jones, Keith Baar, Edward Ojuka, May Chen, John O. Holloszy Department of Medicine, Washington University School of Medicine, St. Louis, Missouri 63110 Running Title: Exercise, UCP3 and mitochondrial biogenesis Please send correspondence to: John O. Holloszy, MD Washington University School of Medicine Department of Medicine Campus Box 8113 4566 Scott Avenue St. Louis, MO 63110 Phone: 314-362-3506 Fax: 314-362-7657 E-Mail: [email protected] Copyright 2002 by the American Physiological Society. AJP-Endo Articles in PresS. Published on September 17, 2002 as DOI 10.1152/ajpendo.00316.2002
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Exercise induces an increase in muscle UCP3 as a component of the increase in mitochondrial...

1
Exercise induces an increase in muscle UCP3 as a component of the increase in mitochondrial
biogenesis
Terry E. Jones, Keith Baar, Edward Ojuka, May Chen, John O. Holloszy
Department of Medicine, Washington University School of Medicine, St. Louis, Missouri 63110
Running Title: Exercise, UCP3 and mitochondrial biogenesis
Please send correspondence to:
John O. Holloszy, MD Washington University School of Medicine Department of Medicine Campus Box 8113 4566 Scott Avenue St. Louis, MO 63110
Phone: 314-362-3506 Fax: 314-362-7657 E-Mail: [email protected]
Copyright 2002 by the American Physiological Society.
AJP-Endo Articles in PresS. Published on September 17, 2002 as DOI 10.1152/ajpendo.00316.2002

2
ABSTRACT
Previous studies have indicated that exercise acutely induces large increases in UCP3 in
skeletal muscle, while endurance training results in marked decreases in muscle UCP3. Because
UCP3 expression appears to be regulated by the same mechanism as other mitochondrial
constituents, it seemed unlikely that exercise would result in such large and divergent changes in
mitochondrial composition. The purpose of this study was to test the hypothesis that major
changes in UCP3 protein concentration do not occur independently of mitochondrial biogenesis
and that UCP3 increases as a component of the exercise-induced increase in mitochondria. We
found a large increase in UCP3 mRNA immediately and 3h after a bout of swimming. UCP3
protein concentration was increased ~35% 18h after a single exercise bout, ~63% after 3d and
~84% after 10d of exercise. These increases in UCP3 roughly paralleled those of other
mitochondrial marker proteins. Our results are consistent with the interpretation that endurance
exercise induces an adaptive increase in mitochondria that have a normal content of UCP3.
gene expression; cytochrome c; cytochrome oxidase; skeletal muscle

3
INTRODUCTION
The mitchondrial protein UCP3 was identified on the basis of its homology with the
uncoupling protein UCP1 present in brown adipose tissue (14). It has been proposed on the basis
of its homology with UCP1 that UCP3 also functions as an uncoupling protein (5, 14).
Furthermore, some investigators have interpreted the results of studies of the heterologous
expression of UCP3 in yeast (36) and of overexpression or knockout of UCP3 in transgenic mice
(8, 9, 34) as providing evidence for uncoupling.
UCP3 is expressed primarily in skeletal muscle (14). Because of its possible role in
energy metabolism, a number of investigators have studied the adaptive response of UCP3 in
skeletal muscle to exercise (11, 23, 29, 33, 37). Tsuboyama-Kasaoka et al. (33) reported that
UCP3 mRNA was increased 7-fold in skeletal muscle of mice three hours after a single bout of
treadmill running, and 16-fold 3 h after exercise in mice that had been trained for 3 wk by
swimming. They speculated that “upregulation of UCP3 mRNA may be a defense mechanism
against extra energy supply to consume extra energy in skeletal muscle.” Similarly, Zhou et al.
(37) reported that UCP3 mRNA was increased ~7-fold immediately after a 200 min long bout of
swimming. A similar response was seen following a bout of treadmill running. They also
reported that UCP3 protein concentration was increased 3.5-fold immediately after 100 min of
treadmill running and 5.6-fold immediately after 200 min of running (37). In studies in humans
Pilegaard et al. (23) found a ~2.5-fold increase in UCP3 mRNA 4 h after a 4 h bout of cycling,
and Schrauwen et al. (29) reported a ~2-fold increase in UCP3 mRNA 4 h after exercise in men
who performed cycle exercise for 2 h in the fasting state.
Although there were differences in the time course and magnitudes of the increases, all of
the studies on the acute effect of exercise have shown that UCP3 mRNA is increased after a bout

4
of exercise. However, Boss et al. (6) reported that in contrast to the acute effects of exercise,
endurance exercise training by means of an 8 week long program of treadmill running resulted in
76% and 59% decreases in UCP3 mRNA in tibialis anterior and soleus muscles of rats. They
speculated that “a need for high metabolic efficiency is associated with decreased mRNA
expression of UCPs in skeletal muscle which would decrease energy dissipation” in the trained
state. Similarly, Schrauwen et al. (29) reported that endurance trained men had significantly
reduced UCP3 mRNA levels in quadriceps muscles compared to untrained men, and that UCP3
mRNA was negatively correlated with maximal oxygen uptake capacity in these subjects.
Taken together the results of these studies have lead to the hypotheses that the “acute
regulation of UCP3 gene expression has immediate and functionally important consequences”
(37), while the decrease in UCP3 mRNA with long term training results in an increase in the
metabolic efficiency of muscle (6, 30). These hypotheses imply that exercise results in major
changes in mitochondrial composition with a great short-term increase in UCP3, and a large
long-term decrease in the amount of UCP3 relative to the other mitochondiral constituents.
Because UCP expression appears to be regulated by the same mechanisms as other
mitochondrial constituents (25, 35), it seemed improbable to us that major changes in UCP3
protein concentration would occur independently of mitochondrial biogenesis. Therefore, the
purpose of the present study was to test the hypothesis that UCP3 protein concentration increases
as a component of the increase in skeletal muscle mitochondria induced by exercise (4, 18).

5
MATERIALS AND METHODS
Materials. Reagents for ECL were obtained from Amersham Pharmacia Biotech.
Reagents for SDS-polyacrylamide gel electrophoresis and Zeta-Probe membranes were from
Bio-Rad. TRIzol Reagent, for isolation of RNA was purchased from Invitrogen. [ 32P]dATP
was purchased from NEN Life Science Products. ULTRAhyb and the Strip EZ DNA labeling kit
were obtained from Ambion. A rabbit polyclonal antibody directed against the 20 carboxy-
terminal amino acids of citrate synthases was generated by Alpha Diagnostic International. A
mouse anti-human cytochrome oxidase (COX) subunit I monoclonal antibody was purchased
from Molecular Probes. A mouse anti-cytochrome c monoclonal antibody was purchased from
Pharmingen International. Horseradish peroxidase-conjugated secondary antibodies were from
The Jackson Laboratory. All other reagents were purchased from Sigma.
Animals. Male wistar rats (~100 g) were purchased from Charles River. Purina chow
and water were provided ad libitum. This study was approved by the Animal Studies Committee
of Washington University School of Medicine.
Exercise. The rats were accustomed to swimming for 2 days, 10 min/day before
performing the exercise protocol of swimming for two-3 h long periods separated by a 45 min
long rest period, as described previously (24, 26). One group of rats performed the exercise
protocol for one day, one group of rats performed three daily bouts of exercise, and a third group
performed 10 daily bouts of exercise. Animals were anesthetized with pentobarbital sodium (5
mg/100 g body wt) immediately, 3h or 18h after the last bout of swimming and triceps and
epitrochlearis muscles were collected. In a separate experiment, soleus, and white and red
portions of quadriceps muscles were harvested from sedentary rats. All muscles were clamp-
frozen and stored at -80°C. The anesthetized rats were killed by exsanguination.

6
Northern blot analysis. Triceps muscle was homogenized in TRIzol. Total RNA was
precipitated using chloroform and isoproopanol. 25 µg of total RNA was size-fractionated in a
1% formaldehyde agarose gel and transferred to Zeta-Probe membrane. Hybridization was
carried out overnight in ULTRAhyb containing a cDNA probe with specific radioactivity of 106
cpm/ml. The cDNA utilized in this study was rat UCP3 (a generous gift from G. Lynis Dohm)
labeled by using a Strip-EZ DNA kit and [ 32P]dATP. Autoradiographs of the membranes were
made using Kodak Biomax MR film.
Western immunoblotting. Epitrochlearis, triceps, soleus, white quadriceps, or red
quadriceps muscle was homogenized in a buffer containing 20 mM HEPES, 1 mM EDTA, 250
mM sucrose, pH 7.4. Protein content was measured using BCA (Pierce). Aliquots of
homogenate were solubilized in Laemmli sample buffer, and subjected to SDS-polyacrylamide
gel electrophoresis. Proteins were transferred to polyvinylidene fluoride membranes.
Membranes were blocked in PBS or TBS containing 5% nonfat dry milk. Blots were probed
with antibodies directed against UCP3, cytochrome c, citrate synthase and COX subunit I. Blots
were then incubated with secondary antibody conjugated to horseradish peroxidase. Antibody
bound protein was detected by ECL. Films were scanned and analyzed using Sigma Scan
(Jandel Scientific).
Statistical analyses. Results are expressed as mean ± SE. Statistical comparisons were
made using unpaired t-tests with the level of statistical significance set at p<0.05.

7
RESULTS
Response of UCP3 to a single bout of exercise. We were unable to quantify the
magnitude of the increase of UCP3 mRNA, as a UCP3 mRNA signal was not detected in non-
exercised muscles. However, Fig. 1A clearly shows that UCP3 mRNA expression was increased
in triceps muscles of rats immediately after, and three hours after, a bout of swimming. This
finding confirms the results of Zhou et al. (37). However, in contrast to the finding of Zhou et al.
(37) that UCP3 protein was increased ~ 5 fold immediately after 200 min of exercise, there was
no increase in UCP3 protein in muscles of our rats ~10 min after 6h of exercise. (Fig. 1B)
Adaptive response of UCP3 and mitochondrial marker enzymes to a program of daily
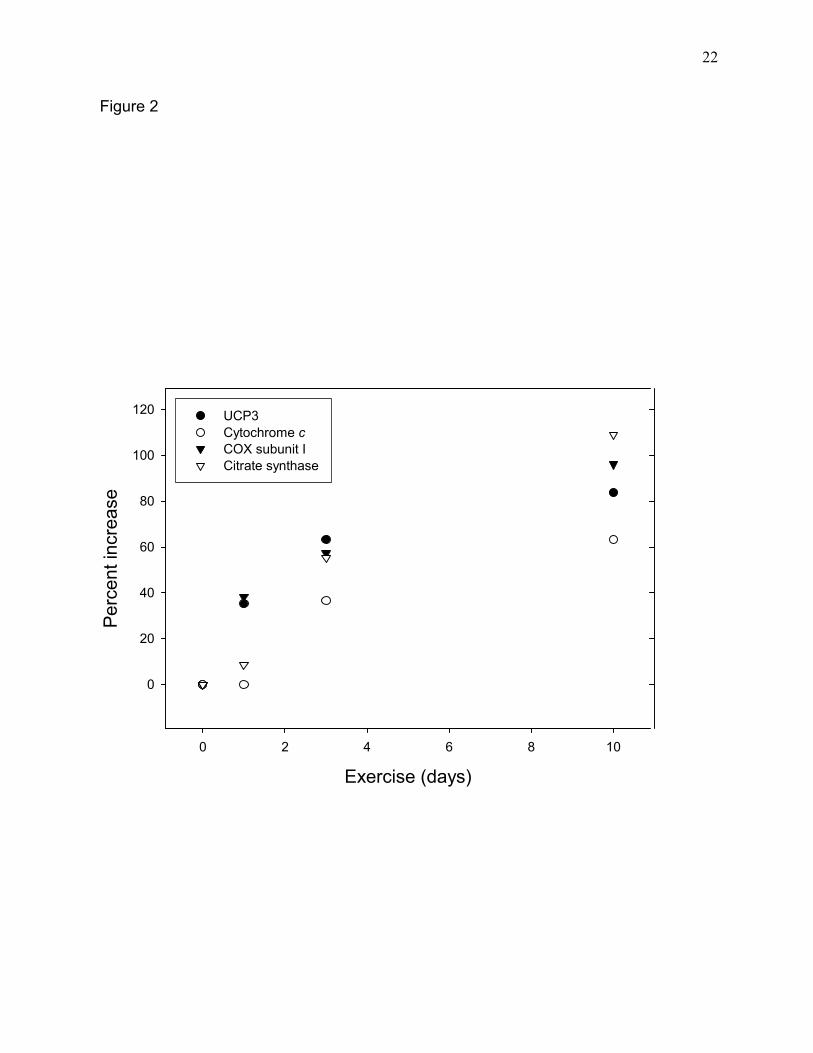
swimming. Eighteen hours after a bout of swimming UCP3 protein concentration was increased
~35% in epitrochlearis muscle (Fig. 1B, Fig. 2). There was a further progressive increase in
UCP3 protein concentration in the epitrochlearis muscle in response to 3 and 10 days of daily
swimming (Fig. 2). This pattern of increase was similar to that of cytochrome oxidase subunit I.
Except for a more rapid initial increase, the increments in UCP3 also paralleled those of two
other mitochondrial markers, cytochrome c and citrate synthase. A similar pattern of increase of
mitochondrial marker enzymes was seen in triceps muscle (data not shown). Thus, it appears
that the adaptive increase in UCP3 in response to exercise reflects the overall increase in
mitochondrial biogenesis.
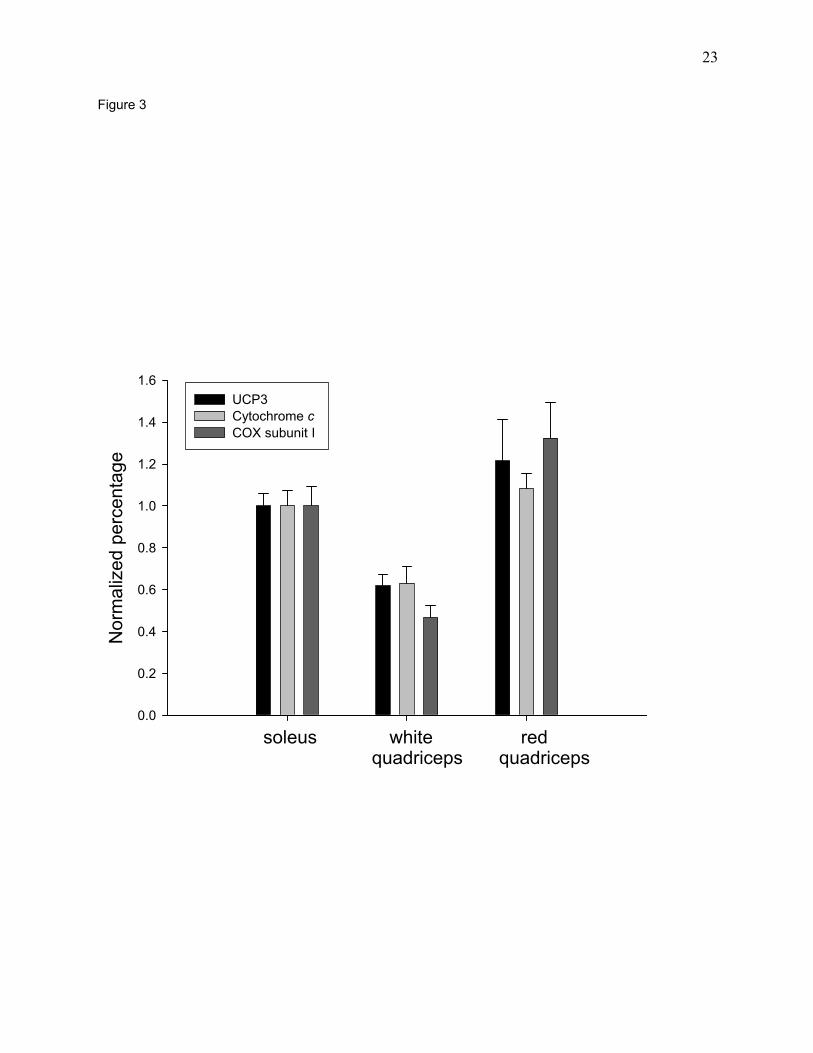
Relationship between UCP3 and other mitochondrial constituents in the three muscle
fiber types. It has been reported that expression of UCP3 protein is most abundant in type IIb
fibers, less in type IIa fibers, and lowest in type I fibers (17). This report appears to be in
conflict with our hypothesis that the UCP3 content of skeletal muscles is proportional to their
content of mitochondria, because type IIb fibers have a lower content of mitochondria than type

8
IIa or type I fibers. We therefore compared the pattern of UCP3 expression with that of two
mitochondrial marker proteins, cytochrome oxidase subunit I and cytochrome c in the
superficial, white and deep, red portions of the quadriceps muscle, and in the soleus muscle. The
superficial, white portion of the quadriceps contains a high proportion of type IIb fibers, the deep
red portion of the quadriceps is made up predominantly of type IIa fibers, while the soleus has a
high proportion of type I fibers (1). As shown in Fig. 3, the concentration of UCP3 protein
roughly parallels those of two other mitochondrial marker proteins.

9
DISCUSSION
The results of this study support our hypothesis that the increase in UCP3 protein that
occurs in response to exercise does not represent an increase in UCP3 protein per se. Instead it
appears that UCP3, a mitochondrial constituent, increases as a component of the exercise-
induced increase in muscle mitochondria. Previous studies have shown that mitochondria from
endurance-trained muscle are tightly coupled (2, 18, 22). An increase in tightly coupled muscle
mitochondria of normal composition, such as occurs with endurance training, does not result in
an increase in resting metabolic rate (31), because there is no increase in the energy requirement
of trained resting muscle, or, therefore, in the availability of ADP and inorganic phosphate (10,
12)
Previous studies of the acute effects of exercise showed that UCP3 mRNA is increased
immediately or shortly after a bout of exercise (23, 33, 37). This finding led to speculation
regarding the functional significance of increased UCP3 expression. Tsuboyama-Kasaoka, et al.
(33) hypothesized that the large up-regulation of UCP3 mRNA may play a role in “fine
adjustments in energy expenditure that may be a defense mechanism against extra supply to
consume extra energy in skeletal muscles”. They also speculated that this increase in UCP3
expression might play a role in the increase in glucose uptake seen three hours after exercise.
While increases in mRNA are frequently referred to as increases in gene expression,
mRNA only provides the information that can lead to increased gene expression. Gene
expression can be controlled at various points beyond transcription in the sequence of events
leading to an increase in the concentration of a protein and modification of the phenotype.
Increased gene expression and its physiological consequences do not occur until there is an
increase in the concentration of the protein encoded by the gene. As illustrated by the response of

10
UCP3 to exercise, the magnitude of an adaptive increase in the concentration of a protein can be
very different from that of the increase in its mRNA, i.e., 7-16 fold increases in mRNA after a
bout of exercise (33, 37) versus a ~35 % increase in UCP3 protein concentration 18 hours after
one bout and a ~ 84 % increase after 10 daily exercise bouts (Fig. 2).
Zhou et al. (37) hypothesized that the rapid increase in UPC3 expression induced by
exercise functions to decrease production of reactive oxygen species in contracting muscles. This
hypothesis seemed reasonable in the context of their finding that the amount of UCP3 protein
was increased 5.6-fold in leg muscles of rats immediately after a single, 200 min long bout of
treadmill running. This large and very rapid increase in protein concentration, which paralleled
the increase in UCP3 mRNA, is, to the best of our knowledge, unprecedented. The largest and
most rapid increases in mitochondrial proteins in skeletal muscle in response to exercise reported
previously are probably those of GLUT4 and ALA synthase both of which can increase as much
as 2-fold during an 18 hour post-exercise period (19, 26). In any case, we were unable to detect
an increase in UCP3 protein immediately post-exercise and found only a 35% increase 18 h after
one bout of exercise. We have no explanation for this discrepancy.
Boss et al. (6) reported that a strenuous 13 wk long program of treadmill exercise training
resulted in decreased expression of UCP3 (an average decrease of ~65 %) in rat skeletal muscles.
They concluded that, in keeping with their hypothesis, this “decreased expression of uncoupling
protein in skeletal muscle would allow for a higher level of metabolic efficiency . . . which
would . . . favor energy storage . . . recovery.” Similarly Schrauwen et al. (29) reported that
UCP3 expression was significantly lower in muscles of trained, as compared to untrained, men,
and that maximal oxygen uptake capacity was negatively correlated with UCP3. In these studies
UCP3 mRNA, rather than actual UCP3 expression, was measured. Nevertheless the finding of a

11
decrease in UCP3 mRNA concentration is puzzling and seems incompatible with our finding of a
progressive increase in UCP3 protein concentration during 10d of exercise-training (Fig. 2).
This finding of Boss et al. (6) is also in disagreement with the results of Tsuboyama-Kasaoka et
al. (33), who found a large increase in UCP3 mRNA after 3 wk of exercise training. Our results
provide evidence that exercise does not result in major changes in the ratio of UCP3 to other
mitochondrial constituents. This in contrast to the picture provided by previous studies of
marked alterations in mitochondrial composition, with an initial large disproportional increase in
UCP3 with one exercise bout (33, 37) and a large decrease in UCP3 with training (6, 29).
There is considerable controversy regarding the functional role of UCP3. UCP1 mediates
adaptive thermogenesis in brown adipose tissue by uncoupling mitochondrial respiration from
ATP synthesis (27). Because UCP3 is a homologue of UCP1, it has been proposed that UCP3
also functions as an uncoupling protein that plays a role in skeletal muscle thermogenesis and
energy expenditure (6, 9, 14, 20).
The results of two studies on UCP3 knockout mice were interpreted to indicate that
mitochondria were more tightly coupled, with an increased state 3/state 4 ratio in one study (34),
and, in another study (9), a 4-fold increase in the rate of ATP synthesis with no change in citrate
cycle flux, CO2 production or energy expenditure. The latter finding is surprising, since the
theoretical maximum P/O ratio (i.e. number of ATP’s generated per oxygen atom) is 3; while
more recently values of ~2.5 have been obtained (3). These findings also do not seem to fit with
the apparent absence of an altered phenotype in UCP3 knockout mice. They do not have a
decreased metabolic rate and are not obese (7, 9, 15, 34), and respond normally to thyroid
hormone (15). A subsequent study on UCP3 knockout mice by Cadenas et al. (7) failed to
confirm the earlier findings of more tightly coupled mitochondria (9, 34); instead they found that

12
respiratory control and proton conductance were unchanged in mitochondria lacking UCP3.
There is evidence that UCP3 can uncouple mitochondria when expressed in yeast (36) or
overexpressed in mouse muscle (7, 8). However, as discussed by Brand and co-workers (7, 16)
this uncoupling appears to be an expression artifact due to disruption of mitochondrial function
by the enormous, 20 to 66-fold, unphysiological increases in UCP3. Although studies in which
UCP3 expression was altered by physiological interventions also do not support an uncoupling
role for UCP3 (21, 28, 32), further research is needed to firmly establish whether or not UCP3
functions as an uncoupling protein muscle under some physiologic conditions such as, for
example, cold exposure.
Another hypothesis regarding the biological role of UCP3 in skeletal muscle is that it has
no significant uncoupling function under physiological conditions, but serves to transport
superoxide anions as a component of the defense mechanism against reactive oxygen species
(ROS) (5, 13). If this concept is correct, our results would indicate that endurance exercise-
training results in an increase in skeletal muscle mitochondria that have a normal capacity to
protect against ROS by the UCP3 mediated transport of superoxide anions.
In conclusion, our results provide evidence that endurance exercise results in an increase
in UCP3 protein in skeletal muscle as a component of the exercise-induced increase in
mitochondrial biogenesis. In contrast to the results of previous studies, they indicate that this
adaptive response results in mitochondria with a normal content of UCP3, rather than in
mitochondria with a markedly increased or decreased compliment of UCP3.

13
ACKNOWLEDGEMENTS
We are grateful to Mrs. Victoria Reckamp for expert technical assistance with
preparation of this manuscript, and to Dr. Lorraine Nolte for technical assistance. We thank Dr.
Lynis Dohm for kindly providing the UCP3 cDNA.
This research was supported by National Institutes of Health Grant AG00425. Terry E.
Jones, Keith Baar and Edward Ojuka were supported by National Institute on Aging Institutional
National Research Service Award AG00078.

14
REFERENCES
1. Armstrong, R. B. and R. O. Phelps. Muscle fiber type composition of the rat hindlimb.
Am.J.Anat. 171: 259-272, 1984.
2. Barnard, R. J., V. R. Edgerton, and J. B. Peter. Effect of exercise on skeletal muscle. I.
Biochemical and histochemical properties. J.Appl.Physiol. 28: 762-766, 1970.
3. Beattie, D. S. Bioenergetics and oxidative metabolism. In Devlin, T. M., ed. Textbook of
Biochemistry. New York, Wiley-Liss. 2002, 578.
4. Booth, F. W. and K. M. Baldwin. Muscle plasticity: energy demanding and supply
processes. In Rowell, L. B. and J. T. Shephard, eds. Handbook of Physiology, Section 12,
Exercise Regulation and Integration of Multiple Systems. New York, Oxford University
Press. 1997, 1075-1123.
5. Boss, O., T. Hagen, and B. B. Lowell. Uncoupling proteins 2 and 3. Potential regulators
of mitochondrial energy metabolism. Diabetes 49: 143-156, 2000.
6. Boss, O., S. Samec, D. Desplanches, M.-H. Mayet, J. Seydoux, P. Muzzin, and J.-P.
Giacobino. Effect of endurance training on mRNA expression of uncoupling proteins 1, 2,
and 3 in the rat. FASEB J 12: 335-339, 1998.
7. Cadenas, S., K. S. Echtay, J. A. Harper, M. B. Jekabsons, J. A. Buckingham, E. Grau,
A. Abuin, H. Chapman, J. C. Clapham, and M. D. Brand. The basal proton
conductance of skeletal muscle mitochondria from transgenic mice overexpressing or
lacking uncoupling protein-3. J.Biol.Chem. 277: 2773-2778, 2002.

15
8. Clapham, J. C., J. R. S. Arch, H. Chapman, A. Haynes, C. Lister, G. B. T. Moore, V.
Piercy, S. A. Carter, I. Lehner, S. A. Smith, L. J. Beeley, R. J. Godden, N. Herrity, M.
Skehel, K. K. Changani, P. D. Hockings, D. G. Reld, S. M. Squires, J. Hatcher, B.
Trail, J. Latcham, S. Rastan, J. A. Harper, S. Cadenas, J. A. Buckingham, M. D.
Brand, and A. Abuin. Mice overexpressing human uncoupling protein-3 in skeletal
muscle are hyperphagic and lean. Nature 406: 415-418, 2000.
9. Cline, G. W., A. J. Vidal-Puig, S. Dufour, K. S. Cadman, B. B. Lowell, and G. I.
Shulman. In vivo effects of uncoupling protein-3 gene disruption on mitochondrial energy
metabolism. J.Biol.Chem. 276: 20240-20244, 2001.
10. Constable, S. M., R. J. Favier, J. A. McLane, R. D. Fell, M. Chen, and J. O. Holloszy.
Energy metabolism in contracting rat skeletal muscle: Adaptation to exercise-training.
Am.J.Physiol. 253: C316-C322, 1987.
11. Cortright, R. N., D. Zheng, J. P. Jones, J. D. Fluckey, S. E. DiCarlo, D. Grujic, B. B.
Lowell, and G. L. Dohm . Regulation of skeletal muscle UCP-2 and UCP-3 gene
expression by exercise and denervation. Am.J.Physiol:Endocrin.Metab. 276: E217-E221,
1999.
12. Dudley, G. A., P. C. Tullson, and R. L. Terjung. Influence of mitochondrial content on
the sensitivity of respiratory control. J.Biol.Chem. 262: 9109-9114, 1987.
13. Echtay, K. S., D. Roussel, J. St-Pierre, M. B. Jekabsons, S. Cadenas, J. A. Stuart, J. A.
Harper, S. J. Roebuck, A. Morrison, S. Pickering, J. C. Clapham, and M. D. Brand.
Superoxide activates mitochondrial uncoupling proteins. Nature 415: 96-99, 2002.

16
14. Gong, D.-W., Y. He, M. Karas, and M. Reitman. Uncoupling protein-3 is a mediator of
thermogenesis regulated by thyroid hormone, 3-adrenergic agonists, and leptin.
J.Biol.Chem. 272: 24129-24132, 1997.
15. Gong, D.-W., S. Monemdjou, O. Gavrilova, L. R. Leon, B. Marcus-Samuels, C. J.
Chou, C. Everett, L. P. Kozak, C. Li, C. Deng, M.-E. Harper, and M. L. Reitman.
Lack of obesity and normal response to fasting and thyroid hormone in mice lacking
uncoupling protein-3. J.Biol.Chem. 275: 16251-16257, 2000.
16. Harper, J. A., J. A. Stuart, M. B. Jekabsons, D. Roussel, K. M. Brindle, K. Dickinson,
R. B. Jones, and M. D. Brand. Artifactual uncoupling by uncoupling protein 3 in yeast
mitochondria at the concentrations found in mouse and rat skeletal-muscle mitochondria.
Biochem.J. 361: 49-56, 2002.
17. Hesselink, M. K. C., H. A. Keizer, L. B. Borghouts, G. Schaart, C. F. P. Kornips, L. J.
Slieker, K. W. Sloop, W. H. M. Saris, and P. Schrauwen. Protein expression of UCP3
differs between human type 1, type 2a, and type 2b fibers. FASEB J 10: 1096-1099, 2001.
18. Holloszy, J. O. Biochemical adaptations in muscle. Effects of exercise on mitochondrial
O2 uptake and respiratory enzyme activity in skeletal muscle. J.Biol.Chem. 242: 2278-
2282, 1967.
19. Holloszy, J. O. and W. W. Winder. Induction of -aminolevulinic acid synthetase in
muscle by exercise or thyroxine. Am.J.Physiol:Regulatory 236: R180-R183, 1979.

17
20. Jaburek, M., M. Varecha, R. E. Gimeno, M. Dembski, P. Jezek, M. Zhang, P. Burn,
L. A. Tartaglia, and K. D. Garlid. Transport function and regulation and mitochondrial
uncoupling proteins 2 and 3. J.Biol.Chem. 274: 26003-26007, 1999.
21. Jucker, B. M., J. Ren, S. Dufour, X. Cao, S. F. Previs, K. S. Cadman, and G. I.
Shulman. 13C/31P NMR assessment of mitochondrial energy coupling in skeletal muscle of
awake fed and fasted rats. J.Biol.Chem. 275: 39279-39286, 2000.
22. Molé, P. A., L. B. Oscai, and J. O. Holloszy. Adaptation of muscle to exercise. Increase
in levels of palmityl CoA synthetase, and in the capacity to oxidize fatty acids.
J.Clin.Invest. 50: 2323-2330, 1971.
23. Pilegaard, H., G. A. Ordway, B. Saltin, and P. D. Neufer. Transcriptional regulation of
gene expression in human skeletal muscle during recovery from exercise.
Am.J.Physiol:Endocrin.Metab. 279: E806-E814, 2000.
24. Ploug, T., B. M. Stallknecht, O. Pedersen, B. B. Kahn, T. Ohkuwa, J. Vinten, and H.
Galbo. Effect of endurance-training on glucose transport capacity and glucose transporter
expression in rat skeletal muscle. Am.J.Physiol:Endocrin.Metab. 259: E778-E786, 1990.
25. Puigserver, P., Z. Wu, C. W. Park, R. Graves, M. Wright, and B. M. Spiegelman. A
cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 92:
829-839, 1998.
26. Ren, J.-M., C. F. Semenkovich, E. A. Gulve, J. Gao, and J. O. Holloszy. Exercise
induces rapid increases in GLUT4 expression, glucose transport capacity, and insulin-
stimulated glycogen storage in muscle. J.Biol.Chem. 269: 14396-14401, 1994.

18
27. Ricquier, D., L. Casteilla, and F. Bouillaud. Molecular studies of the uncoupling protein.
FASEB J 5: 2237-2242, 1991.
28. Samec, S., J. Seydoux, and A. G. Dulloo. Role of UCP homologues in skeletal muscles
and brown adipose tissue: mediators of thermogenesis or regulators of lipids as fuel
substrate? FASEB J 12: 715-724, 1998.
29. Schrauwen, P., M. K. C. Hesselink, I. Vaartjes, E. Kornips, W. H. M. Saris, J.-P.
Giacobino, and A. Russell. Effect of acute exercise on uncoupling protein 3 is a fat
metabolism-mediated effect. Am.J.Physiol:Endocrin.Metab. 282: E11-E17, 2002.
30. Schrauwen, P., F. J. Troost, E. Ravussin, and W. H. M. Saris. Skeletal muscle UCP2
and UCP3 expression in trained and untrained male subjects. Int.J.Obes. 23: 966-972,
1999.
31. Schulz, L. O., B. L. Nyomba, S. Alger, T. E. Anderson, and E. Ravussin. Effect of
endurance training on sedentary energy expenditure measured in a respiratory chamber.
Am.J.Physiol:Endocrin.Metab. 260: E257-E261, 1991.
32. Short, K. R., J. Nygren, R. Barazzoni, J. Levine, and K. S. Nair. T3 increases
mitochondrial ATP production in oxidative muscle despite increased expression of UCP2
and -3. Am.J.Physiol:Endocrin.Metab. 280: E761-E769, 2001.
33. Tsuboyama-Kasaoka, N., N. Tsunoda, K. Maruyama, M. Takahashi, H. Kim, S.
Ikemoto, and O. Ezaki. Up-regulation of uncoupling protein 3 (UCP3) mRNA by exercise
training and down-regulation of UCP3 by denervation in skeletal muscles.
Biochem.Biophys.Res.Commun. 247: 498-503, 1998.

19
34. Vidal-Puig, A. J., D. Grujic, C.-Y. Zhang, T. Hagen, O. Boss, Y. Ido, A. Szczepanik, J.
Wade, V. Mootha, R. Cortright, D. M. Muoio, and B. B. Lowell. Energy metabolism in
uncoupling protein 3 gene knockout mice. J.Biol.Chem. 275: 16258-16266, 2000.
35. Wu, Z., P. Puigserver, U. Andersson, C. Zhang, G. Adelmant, V. Mootha, A. Troy, S.
Cinti, B. Lowell, R. C. Scarpulla, and B. M. Spiegelman. Mechanisms controlling
mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell
98: 115-124, 1999.
36. Zhang, C.-Y., T. Hagen, V. K. Mootha, L. J. Slieker, and B. B. Lowell. Assessment of
uncoupling activity of uncoupling protein 3 using a yeast heterologous expression system.
FEBS Letts. 449: 129-134, 1999.
37. Zhou, M., B.-Z. Lin, S. Coughlin, G. Vallaga, and P. F. Pilch. UCP-3 expression in
skeletal muscle: effects of exercise, hypoxia, and AMP-activated protein kinase.
Am.J.Physiol:Endocrin.Metab. 279: E622-E629, 2000.

20
FIGURE LEGENDS
Fig. 1. Effects of one bout of exercise on triceps muscle UCP3 mRNA and protein content. A.
Northern blots were performed on RNA from triceps muscle of sedentary rats, and on muscles of
exercised rats obtained ~10 min, 3h or 18h after exercise, as described in Materials and Methods.
B. Western blots were performed on homogenates of epitrochlearis muscles from sedentary rats,
and on muscles of exercised rats obtained ~10 min and 18h after exercise, as described in
Materials and Methods. Each bar represents the mean ± SE for muscles from 6-11 rats.
Fig. 2. Time course of the increases in UCP3 protein and a number of other mitochondrial
proteins in response to exercise. Epitrochlearis muscles were obtained from sedentary rats and
from rats exercised by swimming either once, for 3d or for 10d. Muscles were taken ~18-20h
after the last exercise bout. Enzyme protein levels were determined by Western blot analysis as
described under Materials and Methods. Each point represents the mean obtained on muscles
from 3-12 rats.
Fig. 3. Comparison of UCP3, cytochrome c and cytochrome oxidase subunit I (COXI) levels in
the soleus and the deep, red and superficial, white portions of the quadriceps muscle. Muscle
homogenates were used to determine UCP3, cytochrome c and cytochrome oxidase subunit I
protein levels as described under Materials and Methods. Bars are means ± SE for muscles from
3-4 rats.

21
Control 10 min 18 h
UC
P3 (n
orm
aliz
ed to
tal i
nten
sitie
s)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
A
Control 10 min 3 h 18 h Post-exercise
B
Control 10 min 18 h Post-exercise
Figure 1
UCP3 mRNA
UCP3 Protein
Post-exercise
*
22
Exercise (days)0 2 4 6 8 10
Perc
ent i
ncre
ase
0
20
40
60
80
100
120 UCP3Cytochrome cCOX subunit ICitrate synthase
Figure 2
23
Nor
mal
ized
per
cent
age
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6UCP3Cytochrome cCOX subunit I
soleus white red quadriceps quadriceps
Figure 3




















