
General Secretion Pathway (eps) Genes Required for Toxin Secretion and Outer Membrane Biogenesis in...
11
1997, 179(22):6994. J. Bacteriol. Bagdasarian, M Koomey, V J DiRita and M Bagdasarian M Sandkvist, L O Michel, L P Hough, V M Morales, M membrane biogenesis in Vibrio cholerae. required for toxin secretion and outer General secretion pathway (eps) genes http://jb.asm.org/content/179/22/6994 Updated information and services can be found at: These include: CONTENT ALERTS more» cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new articles http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on January 28, 2014 by guest http://jb.asm.org/ Downloaded from on January 28, 2014 by guest http://jb.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of General Secretion Pathway (eps) Genes Required for Toxin Secretion and Outer Membrane Biogenesis in...

1997, 179(22):6994. J. Bacteriol.
Bagdasarian, M Koomey, V J DiRita and M BagdasarianM Sandkvist, L O Michel, L P Hough, V M Morales, M membrane biogenesis in Vibrio cholerae.required for toxin secretion and outer General secretion pathway (eps) genes
http://jb.asm.org/content/179/22/6994Updated information and services can be found at:
These include:
CONTENT ALERTS more»cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new articles
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
on January 28, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

JOURNAL OF BACTERIOLOGY,0021-9193/97/$04.0010
Nov. 1997, p. 6994–7003 Vol. 179, No. 22
Copyright © 1997, American Society for Microbiology
General Secretion Pathway (eps) Genes Required for ToxinSecretion and Outer Membrane Biogenesis in Vibrio choleraeMARIA SANDKVIST,1† LINDA OVERBYE MICHEL,2 LLOYD P. HOUGH,2 VICTOR M. MORALES,3
MIRA BAGDASARIAN,2 MICHAEL KOOMEY,4 VICTOR J. DIRITA,4 AND MICHAEL BAGDASARIAN2*
Department of Microbiology and NSF Center for Microbial Ecology, Michigan State University, East Lansing,Michigan 488242; Oral Infection and Immunity Branch, National Institute of Dental Research, NationalInstitutes of Health, Bethesda, Maryland 20892-43501; Gastroenterology Division, Brigham and Women’sHospital, Boston, Massachusetts 021153; and Department of Microbiology and Immunology, University of
Michigan Medical School, Ann Arbor, Michigan 481094
Received 10 June 1997/Accepted 16 September 1997
The general secretion pathway (GSP) of Vibrio cholerae is required for secretion of proteins includingchitinase, enterotoxin, and protease through the outer membrane. In this study, we report the cloning andsequencing of a DNA fragment from V. cholerae, containing 12 open reading frames, epsC to -N, which aresimilar to GSP genes of Aeromonas, Erwinia, Klebsiella, Pseudomonas, and Xanthomonas spp. In addition to thetwo previously described genes, epsE and epsM (M. Sandkvist, V. Morales, and M. Bagdasarian, Gene 123:81–86, 1993; L. J. Overbye, M. Sandkvist, and M. Bagdasarian, Gene 132:101–106, 1993), it is shown here thatepsC, epsF, epsG, and epsL also encode proteins essential for GSP function. Mutations in the eps genes resultin aberrant outer membrane protein profiles, which indicates that the GSP, or at least some of its components,is required not only for secretion of soluble proteins but also for proper outer membrane assembly. Several ofthe Eps proteins have been identified by use of the T7 polymerase-promoter system in Escherichia coli. One ofthem, a pilin-like protein, EpsG, was analyzed also in V. cholerae and found to migrate as two bands onpolyacrylamide gels, suggesting that in this organism it might be processed or otherwise modified by a prepilinpeptidase. We believe that TcpJ prepilin peptidase, which processes the subunit of the toxin-coregulated pilus,TcpA, is not involved in this event. This is supported by the observations that apparent processing of EpsGoccurs in a tcpJ mutant of V. cholerae and that, when coexpressed in E. coli, TcpJ cannot process EpsG althoughthe PilD peptidase from Neisseria gonorrhoeae can.
In Vibrio cholerae, the general secretion pathway (GSP) isrequired for extracellular secretion of several proteins includ-ing chitinase, cholera toxin (CT), and protease (51, 60). Thispathway likely plays a significant role in the survival of Vibrio indifferent environments as well as in its pathogenicity. For in-stance, secreted chitinase and protease may assist in the de-tachment of V. cholerae from chitinous waterborne particlesand from host epithelial cells, respectively, thus facilitatingdissemination (14, 71). CT is the major component responsiblefor inducing the diarrheal disease cholera and is therefore alsoinvolved in the dissemination process (for a review, see refer-ence 30).
Escherichia coli heat-labile enterotoxin (LT) is very similarto CT in its structure, biochemical function, and ability totranslocate across the outer membrane of V. cholerae (66). It isa multimeric protein which consists of a single A subunit anda pentamer of identical B-subunit polypeptides (18, 64, 65). Itssecretion from V. cholerae proceeds in two steps, similar to thesecretion of CT. The first step is translocation of the individualsubunit polypeptides across the cytoplasmic membrane. This ismediated by a pathway which is presumably similar to the secsystem of E. coli (63). During this step, the signal peptides ofthe A and B precursor polypeptides are cleaved off (8, 25, 52)and the mature subunits are transported to the periplasm.
Here they undergo folding and assembly to form the holotoxin(13, 23, 61). The second step is the translocation of the holo-toxin (or the B-subunit pentamer if the A subunits are notpresent) through the outer membrane (22, 56). This requiresthe function of the GSP and, as shown in this work, occurs withthe assistance of several gene products. This process seems tobe highly conserved: GSP genes have been identified in avariety of bacteria, including Aeromonas (25), Erwinia (6, 21,38, 57), Klebsiella (9), Pseudomonas (73), and Xanthomonas(11, 27) spp.
We report here the cloning and sequencing of a cluster ofgenes which constitute part of the GSP of V. cholerae. Thesegenes appear to play a role not only in the secretion of solubleproteins through the outer membrane but also in the assemblyof the outer membrane itself.
MATERIALS AND METHODS
Bacterial strains and plasmids. Bacterial strains and plasmids used are listedin Table 1 and Fig. 1.
DNA sequence analyses. Nucleotide sequence determination was performedby the dideoxy chain termination method (62) with the Sequenase kit (U.S.Biochemical Corp.) or by the Taq polymerase cycle methods (2,7-deaza-GTP kit;U.S. Biochemical Corp.) on single- or double-stranded DNA. Products wereanalyzed either manually or with an ABI 373A automatic sequencer at theMSU-DOE-PRL Plant Biochemistry Facility. The insertion point of Tn5 in theV. cholerae TRH7000 epsC mutant was determined by sequencing the PCR-amplified fragments containing the insertion sequence (IS) and the adjacentchromosomal DNA by the method of Thien (72).
Construction of eps mutants. A StuI DNA fragment carrying the Kmr gene ofTn903 was inserted into the epsF, epsG, and epsL genes by in vitro manipulationand introduced into the chromosome of V. cholerae by homologous recombina-tion as described previously (58). An epsE-lacZ operon fusion was constructed bycloning a PCR fragment containing a promoterless lacZ gene into the epsE gene.The resulting fusion was introduced into the chromosomal epsE gene by homol-
* Corresponding author. Mailing address: Department of Microbi-ology and NSF Center for Microbial Ecology, S-110 Plant BiologyBuilding, Michigan State University, East Lansing, MI 48824. Phone:(517) 353-8619. Fax: (517) 353-1926. E-mail: [email protected].
† Present address: Jerome H. Holland Laboratories, Department ofBiochemistry, American Red Cross, Rockville, MD 20855.
6994
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

ogous recombination. To easily screen for recombination of this fusion onto thechromosome, a Kmr gene from pACYC177 was inserted downstream of the lacZgene.
PCR amplification of mutant DNA containing Tn5. In order to determine thelocation of Tn5 in the V. cholerae TRH7000 epsC mutant, PCR was performedwith converging primers. Primer MMB41 (59-CGCACGATGAAGAGCAGA-39) was located in the IS of Tn5, and primer MMB73 (59-TGCTTGGCTTCCATCTG-39) was located in the epsD gene. PCR mixtures (100-ml final volume)contained 10 mM Tris-HCl (pH 8.3) (at 25°C), 50 mM KCl, 4 mM MgCl2,0.001% (wt/vol) gelatin (Sigma), 0.5 mM each primer, 200 mM each deoxynucleo-side triphosphate, and 1 mg of genomic DNA. The reaction mixture was overlaidwith 100 ml of mineral oil (Sigma) and heated to 95°C for 5 min, at which point2.5 U of AmpliTaq DNA polymerase (Perkin-Elmer Cetus) was added. TheDNA was then amplified in a Perkin-Elmer thermal cycler for 30 cycles eachconsisting of the following steps: (i) 30 s at 94°C, (ii) 30 s at 64°C, and (iii) 30 sat 72°C, with the last cycle having the 72°C incubation extended to 7 min.
Expression of genes under bacteriophage T7 f10 promoter control. Deriva-tives of pT7-5 or pT7-6 (70) containing different fragments of the eps gene cluster(Fig. 1) were introduced into E. coli MC1061[pGP1-2]. The expression of pro-teins encoded by the recombinant plasmids was performed as described previ-ously (51).
Expression of EpsG protein in V. cholerae was determined as follows. StrainsTRH7000 and J71K-1 containing plasmid pMMB586, expressing the T7 RNApolymerase, and plasmid pMMB564, carrying the epsG gene (coordinates 5808to 6609 of the eps sequence), were used. Cells were grown at 30°C overnight inM9 medium (plus 100 mg of thymine per ml, 20 mg of thiamine per ml, and 0.01%concentrations of each of 19 amino acids [without Met]) with 50 mg of ampicillinand kanamycin per ml. Two hundred microliters of this culture was then pelleted,resuspended in 1 ml of the same medium, and incubated for 2 h at 30°C followedby 30 min of incubation at 42°C. Rifampin was added to a final concentration of300 mg/ml, and the incubation was continued for another 20 min, when the
temperature was lowered to 30°C for 10 min. The cells were labeled for 10 minwith [35S]Met (10 mCi/ml, 1,000 Ci/mmol), pelleted, resuspended in samplebuffer, and heated at 95°C for 5 min. Proteins were separated on a 0.1% sodiumdodecyl sulfate (SDS)–15% polyacrylamide gel, which was then dried and auto-radiographed. For pulse-chase labeling of V. cholerae J71K-1, containingpMMB586 and pMMB564, the following conditions were used. The culture wasgrown in the same way as described for the preceding experiment. One milliliterof the preculture was added to 11.0 ml of the same medium and grown at 30°C,induced at 42°C, and treated with rifampin as described above. It was pulsed with10 mCi of [35S]Met per ml for 30 s. Unlabeled Met was added to a finalconcentration of 0.6 mM, and 2.0-ml samples were withdrawn at 0.5, 1, 2, 5, and10 min into 2.0 ml of cold 10% trichloroacetic acid. After 15 min on ice, thesamples were centrifuged, washed with acetone, suspended in sample buffer,heated at 95°C for 10 min, and separated on a 0.1% SDS–17% polyacrylamidegel and subjected to autoradiography.
Isolation of outer membrane fractions from V. cholerae. Cells of V. choleraeTRH7000 and of eps mutant strains were grown overnight in Luria-Bertani (LB)medium or LB medium containing 0.4% maltose at 37°C (to induce ompSexpression). Cells from 1.0 ml of culture were pelleted and resuspended in 250ml of 10 mM phosphate buffer (pH 7.3)–150 mM NaCl (phosphate-bufferedsaline [PBS]) containing 4,000 U of polymyxin B sulfate (Sigma) per ml andincubated on ice for 25 min to release periplasmic proteins. The cells were thencentrifuged at 16,000 3 g in an Eppendorf microcentrifuge at 4°C for 5 min,resuspended in 250 ml of PBS, placed on ice, and broken by sonication with two15-s pulses. After a low-speed centrifugation (2,000 3 g for 2 min) to remove anyunbroken cells, the lysate was centrifuged at 16,000 3 g at 4°C for 30 min topellet the membrane fraction. This pellet was resuspended in 250 ml of PBScontaining 1.0% Triton X-100, held at room temperature for 30 min, and cen-trifuged at room temperature for 30 min at 16,000 3 g. The pellet was resus-pended in 60 ml of sample buffer, and 14 ml was loaded on a 0.1% SDS–15.0%polyacrylamide gel (35). The pH of the buffer in the upper reservoir was adjusted
TABLE 1. Strains and plasmids used in this study
Strain orplasmid Relevant characteristic(s) Source or
reference
StrainsV. cholerae
0395 wt classical strain 42JJM43 0395 DtoxR43 DctxA1 22TRH7000 ElTor thy Hgr D(ctxA-ctxB) 24J71K-1 0395 tcpJ::Tn5 31
E. coliMC1061 F2 araD139 D(ara-leu)7697 D(lac)X74 rpsL hsdR2 mcrA mcrB1 4BL21(DE3) F2 ompT T7 gene 1 under lacp control on l prophage 68
PlasmidsE26 Cosmid pLAFR5::epsC to -N This studypGP1-2 Kmr T7 gene 1 under lpL control, cI857 70pMMB67EH Apr Ptac mob1 17pMMB68 pMMB67EH etxB under Ptac control 59pMMB206 Same as pMMB67EH but Cmr 45pMMB356 pMMB67EH::SacI fragment epsF This studypMMB531 pT7-5, KpnI fragment epsH to -M This studypMMB547 pT7-5::EcoRI fragment epsE to -N This studypMMB551 pT7-5::EcoRI-BamHI fragment epsE to -J This studypMMB560 pT7-6::HindIII-EcoRI fragment epsC This studypMMB564 pT7-5::HindII-XmnI fragment coordinates 5808–6609, epsG This studypMMB574 pMMB206::HindII-XmnI fragment epsG This studypMMB586 pGP1-2 with Kmr replaced by Cmr This studypMMB608 pT7-5::HindIII fragment Bal 31 treated, epsE This studypMS10 pMMB67HE::HindIII-EcoRI fragment epsC This studypMS13 pMMB207::KpnI-EcoRI fragment region downstream of epsN This studypMS14 Same as pMS13, but fragment in reverse orientation This studypMS19 pT7-5::XbaI-BamHI, epsF to -J This studypMS21 pMMB207::tcpJ This studypMS37 pT7-5::EcoRV fragment epsH to -K 58pMS38 pT7-6::BamHI-SspI, epsL 58pMS44 pMMB67HE::BamHI-SspI, epsL This studypT7-5/6 Apr T7 f10 promoter 70pTKK4 pT7-5::HpaI-SspI epsKL This studypTKK5 pT7-6::BamHI-KpnI epsLM This studypWD615 Tcr etxB 7
VOL. 179, 1997 EXTRACELLULAR SECRETION IN V. CHOLERAE 6995
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

to 6.7 to optimize separation of OmpS and OmpT (36). The proteins wereelectroblotted onto a nitrocellulose membrane and probed with anti-OmpS (do-nated by E. Tapio Palva [36]) or with anti-OmpU antibodies.
Quantitative determination of LT B-subunit pentamer secretion. The geneetxB, encoding the B subunit of LT, was introduced into V. cholerae on plasmidpMMB68 (59) or pWD615 (7). LT B-subunit pentamers present in the growthmedium and sonicated cells were determined by GM1 enzyme-linked immu-nosorbent assay (69) as described previously (51).
Enzyme assays. b-Galactosidase and b-lactamase production was determinedas described in references 43 and 23, respectively.
For determination of extracellular DNase, the mixture contained the follow-ing: culture medium passed through a Sephadex G-25 column (3.0 to 12.0 mg ofprotein/ml), salmon sperm DNA (400 mg/ml), and buffer (25 mM HEPES, 4 mMMgCl2, 4 mM CaCl2, pH 7.4). Incubation was at 37°C for 0, 15, and 30 min. Anequal volume of 6% HClO4 was added to the samples to stop the reaction andprecipitate the high-molecular-weight DNA. After 15 min on ice, the sampleswere centrifuged at 20,000 3 g and the absorption at 260 nm was determined.One optical density unit was considered equivalent to 50 mg of DNA per ml.
Extracellular protease was determined in the same samples of culture mediumby a modification of the method of Julius et al. (29). The mixture contained, in10 ml, HEPES, Tris (100 mM; pH 7.0), and N-tert-butoxy-carbonyl-Gln-Ala-Arg-7-amido-4-methyl-coumarin (Sigma). The mixture was placed in the microcu-vette of the fluorometer (DynaQuant 2000 from Hoefer-Pharmacia), and thesignal was recorded every minute for 10 min at room temperature.
Nucleotide sequence accession number. The nucleotide sequence of the epsC-to-epsN cluster has been deposited in the GenBank database under accession no.L33796.
RESULTS
Nucleotide sequence of the epsC to -N gene cluster. Approx-imately 12 kb of the DNA fragment of the V. cholerae chro-mosome, containing two previously described genes, epsE andepsM (51, 60, 61), has been sequenced. In addition, the nucle-otide sequence of approximately 1.5 kb upstream of epsC (Fig.1), present in the original cosmid insert, has been determined.
Computer analysis (10) of the sequence obtained revealed12 open reading frames (ORFs) (Table 2). Table 3 shows theidentities of putative Eps proteins from V. cholerae (obtainedby translation of the appropriate ORFs) and GSP proteins ofother organisms. Similarities were also noticed between someof the Eps proteins and the Pil proteins of Pseudomonas aerugi-nosa and Neisseria gonorrhoeae (1, 2, 37, 39, 48) and the Tcpproteins of V. cholerae, which are required for processing andsecretion of type IV pilin (32, 50). Recently, genes homologousto some of the eps genes, required for the biogenesis of V.cholerae mannose-sensitive hemagglutinin, have also beenidentified (20, 41).
Hydrophobic regions that could represent potential mem-
brane-spanning domains were found in 11 of the predicted Epsproteins by computer analysis (34). The exception was EpsE,which is a cytoplasmic protein associated with the cytoplasmicmembrane by virtue of its interaction with the EpsL protein(58, 60). EpsC, -L, and -M each possess one putative hydro-phobic transmembrane region, whereas EpsF has three. Notypical signal sequence capable of being cleaved was detectedin any of these proteins. Fractionation experiments using V.cholerae (46) and E. coli (unpublished data) demonstrated thatEpsM is localized to the cytoplasmic membrane, showing thatthe hydrophobic region of this protein likely acts as a mem-brane localization signal.
Stretches of hydrophobic residues typical of those found insignal peptides (74) were found at the N-terminal domains ofEpsD and EpsN. It is probable, therefore, that both of theseproteins are translocated through the cytoplasmic membrane.Outer membrane location has recently been demonstrated forthe EpsD homologs PulD and pIV (19, 40). Hydrophobic se-quences similar to those recognized by the prepilin peptidaseand present in type IV pilin subunits were found at the Ntermini of EpsG, -H, -I, and -J. These signal sequences consistof a short leader sequence followed by a conserved region(G-¹F-T-L/I-Q) surrounding the prepilin peptidase cleavagesite (¹), which is followed by a region of hydrophobic aminoacids (54, 55, 67). Similar proteins are known to be part of theGSPs in other bacteria and have been shown to adopt trans-membrane conformations suitable for processing by prepilinpeptidase (49, 56). It is considered probable, therefore, thatproteins EpsG through EpsJ are also recognized and processedby a prepilin peptidase encoded by V. cholerae, although sucha peptidase has yet to be identified.
The epsC gene is preceded by the sequence GTGGAA, apossible ribosome binding site. One hundred twenty base pairsupstream of epsC, the sequence ..TTGAGACACTTCGCTCCACTATGCTTGTGTTATATT could represent a s70-specificpromoter, although the distance between the 235 and 210regions is 23 bp rather than the 14 to 19 bp usually found in thes70 promoters. Further DNA sequencing, insertional mutagen-esis, and primer extension experiments will have to be per-formed on the region upstream of epsC to map the promoterand make certain that no additional genes or long untranslatedsequences are present in this region of the eps cluster.
The epsC to -N gene cluster is followed by an inverted repeat(ATTTCACGCCCAACTGGGCGTGAAAT) which may rep-resent a transcriptional terminator. Analysis of the DNA se-
FIG. 1. Physical and genetic map of the eps gene cluster. The extent of DNAfragments inserted into pT7-5 or pT7-6 vectors and used for deletion mappingand visualization of gene products is indicated by horizontal lines. Restrictionsite abbreviations: B, BamHI; E, EcoRI; H, HindIII; K, KpnI; S, SmaI; X, XbaI.
TABLE 2. Location and characteristics of predicted Eps readingframes in the sequence of the epsC to -N gene cluster
Protein ORF No. ofresidues
Predictedmol wt
EpsC 213–1130 305 33,592EpsD 1176–3200 674 73,337EpsEa 3197–4708 503 56,358EpsFa 4708–5928 406 44,916EpsG 5969–6409 146 16,063EpsH 6443–7027 194 21,739EpsIa 7017–7370 117 13,493EpsJa 7364–8022 210 23,757EpsKa 8012–9022 336 37,599EpsLa 8991–10202 403 45,343EpsM 10209–10709 166 18,521EpsNa 10711–11466 251 27,322
a Initiation codon of ORF overlaps termination codon of preceding ORF or isseparated by 2 nucleotides or less.
6996 SANDKVIST ET AL. J. BACTERIOL.
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
quence 600 bp downstream of epsN did not reveal any ORFswith homology to known GSP genes but did identify a partialORF, in the opposite orientation from epsN, with similarity tothe cysQ gene of E. coli (46). This is in contrast to most otherGSP operons, in which an additional gene, encoding a prepilinpeptidase required for secretion, is present directly down-stream of the epsN gene (3, 12, 55).
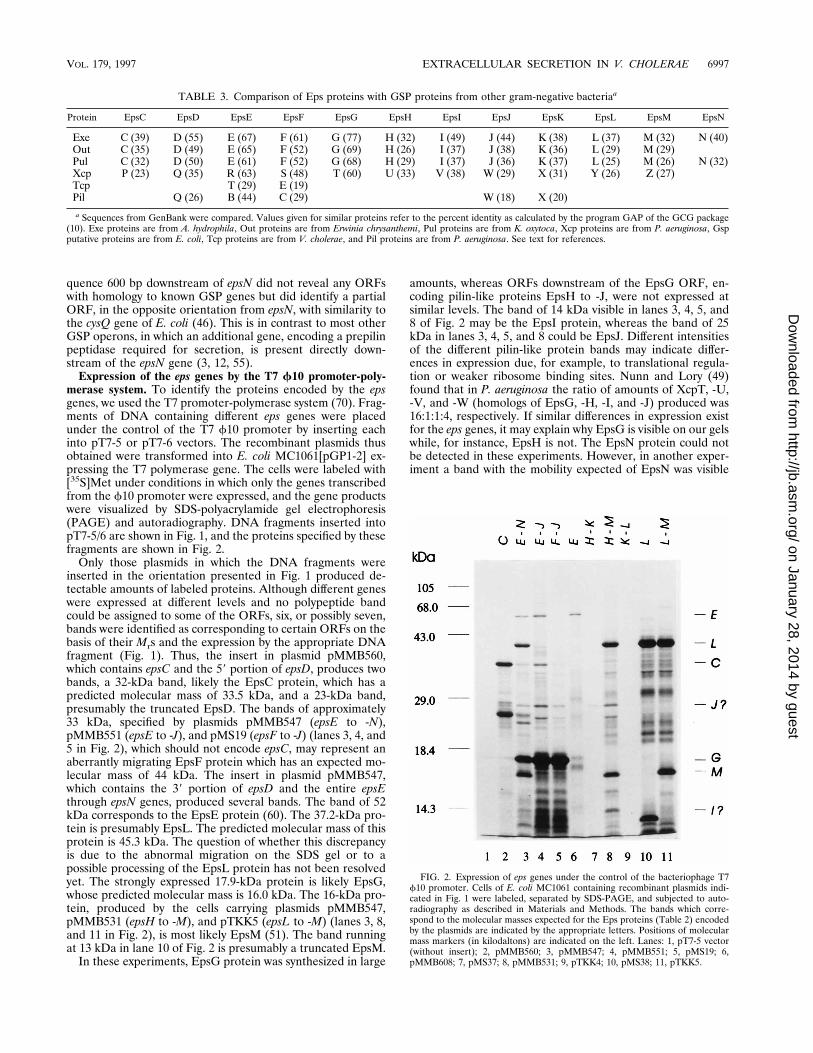
Expression of the eps genes by the T7 f10 promoter-poly-merase system. To identify the proteins encoded by the epsgenes, we used the T7 promoter-polymerase system (70). Frag-ments of DNA containing different eps genes were placedunder the control of the T7 f10 promoter by inserting eachinto pT7-5 or pT7-6 vectors. The recombinant plasmids thusobtained were transformed into E. coli MC1061[pGP1-2] ex-pressing the T7 polymerase gene. The cells were labeled with[35S]Met under conditions in which only the genes transcribedfrom the f10 promoter were expressed, and the gene productswere visualized by SDS-polyacrylamide gel electrophoresis(PAGE) and autoradiography. DNA fragments inserted intopT7-5/6 are shown in Fig. 1, and the proteins specified by thesefragments are shown in Fig. 2.
Only those plasmids in which the DNA fragments wereinserted in the orientation presented in Fig. 1 produced de-tectable amounts of labeled proteins. Although different geneswere expressed at different levels and no polypeptide bandcould be assigned to some of the ORFs, six, or possibly seven,bands were identified as corresponding to certain ORFs on thebasis of their Mrs and the expression by the appropriate DNAfragment (Fig. 1). Thus, the insert in plasmid pMMB560,which contains epsC and the 59 portion of epsD, produces twobands, a 32-kDa band, likely the EpsC protein, which has apredicted molecular mass of 33.5 kDa, and a 23-kDa band,presumably the truncated EpsD. The bands of approximately33 kDa, specified by plasmids pMMB547 (epsE to -N),pMMB551 (epsE to -J), and pMS19 (epsF to -J) (lanes 3, 4, and5 in Fig. 2), which should not encode epsC, may represent anaberrantly migrating EpsF protein which has an expected mo-lecular mass of 44 kDa. The insert in plasmid pMMB547,which contains the 39 portion of epsD and the entire epsEthrough epsN genes, produced several bands. The band of 52kDa corresponds to the EpsE protein (60). The 37.2-kDa pro-tein is presumably EpsL. The predicted molecular mass of thisprotein is 45.3 kDa. The question of whether this discrepancyis due to the abnormal migration on the SDS gel or to apossible processing of the EpsL protein has not been resolvedyet. The strongly expressed 17.9-kDa protein is likely EpsG,whose predicted molecular mass is 16.0 kDa. The 16-kDa pro-tein, produced by the cells carrying plasmids pMMB547,pMMB531 (epsH to -M), and pTKK5 (epsL to -M) (lanes 3, 8,and 11 in Fig. 2), is most likely EpsM (51). The band runningat 13 kDa in lane 10 of Fig. 2 is presumably a truncated EpsM.
In these experiments, EpsG protein was synthesized in large
amounts, whereas ORFs downstream of the EpsG ORF, en-coding pilin-like proteins EpsH to -J, were not expressed atsimilar levels. The band of 14 kDa visible in lanes 3, 4, 5, and8 of Fig. 2 may be the EpsI protein, whereas the band of 25kDa in lanes 3, 4, 5, and 8 could be EpsJ. Different intensitiesof the different pilin-like protein bands may indicate differ-ences in expression due, for example, to translational regula-tion or weaker ribosome binding sites. Nunn and Lory (49)found that in P. aeruginosa the ratio of amounts of XcpT, -U,-V, and -W (homologs of EpsG, -H, -I, and -J) produced was16:1:1:4, respectively. If similar differences in expression existfor the eps genes, it may explain why EpsG is visible on our gelswhile, for instance, EpsH is not. The EpsN protein could notbe detected in these experiments. However, in another exper-iment a band with the mobility expected of EpsN was visible
FIG. 2. Expression of eps genes under the control of the bacteriophage T7f10 promoter. Cells of E. coli MC1061 containing recombinant plasmids indi-cated in Fig. 1 were labeled, separated by SDS-PAGE, and subjected to auto-radiography as described in Materials and Methods. The bands which corre-spond to the molecular masses expected for the Eps proteins (Table 2) encodedby the plasmids are indicated by the appropriate letters. Positions of molecularmass markers (in kilodaltons) are indicated on the left. Lanes: 1, pT7-5 vector(without insert); 2, pMMB560; 3, pMMB547; 4, pMMB551; 5, pMS19; 6,pMMB608; 7, pMS37; 8, pMMB531; 9, pTKK4; 10, pMS38; 11, pTKK5.
TABLE 3. Comparison of Eps proteins with GSP proteins from other gram-negative bacteriaa
Protein EpsC EpsD EpsE EpsF EpsG EpsH EpsI EpsJ EpsK EpsL EpsM EpsN
Exe C (39) D (55) E (67) F (61) G (77) H (32) I (49) J (44) K (38) L (37) M (32) N (40)Out C (35) D (49) E (65) F (52) G (69) H (26) I (37) J (38) K (36) L (29) M (29)Pul C (32) D (50) E (61) F (52) G (68) H (29) I (37) J (36) K (37) L (25) M (26) N (32)Xcp P (23) Q (35) R (63) S (48) T (60) U (33) V (38) W (29) X (31) Y (26) Z (27)Tcp T (29) E (19)Pil Q (26) B (44) C (29) W (18) X (20)
a Sequences from GenBank were compared. Values given for similar proteins refer to the percent identity as calculated by the program GAP of the GCG package(10). Exe proteins are from A. hydrophila, Out proteins are from Erwinia chrysanthemi, Pul proteins are from K. oxytoca, Xcp proteins are from P. aeruginosa, Gspputative proteins are from E. coli, Tcp proteins are from V. cholerae, and Pil proteins are from P. aeruginosa. See text for references.
VOL. 179, 1997 EXTRACELLULAR SECRETION IN V. CHOLERAE 6997
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
after longer exposure (results not shown). The difference indetection level could also reflect different stabilities of the Epsproteins when expressed in a different host in the absence ofother Eps products. Such a situation has been observed in thecase of EpsE and PulD, when produced in the absence of EpsL(58) and PulS (18), respectively.
DNA inserts carried by the plasmids pMS37 and pTKK4,containing genes epsH to -K and epsK to -L, respectively, didnot direct the synthesis of any proteins in the T7 f10 promot-er-T7 RNA polymerase system (lanes 7 and 9 in Fig. 2). Thereason for this is not clear, particularly since other fragmentsdirected the synthesis of bands corresponding in molecularweight to EpsL or EpsI. Moreover, EpsL can be detectedimmunologically in strains containing pTKK4 (data notshown). At least in the case of EpsI, it is possible that trans-lation from the regions upstream of the epsH gene is requiredfor successful expression of the epsI gene or for the stabiliza-tion of its product.
Although the results obtained with the T7 f10 promoter arenot a conclusive proof that these Eps proteins are expressed inVibrio, rigorous confirmation has been obtained for EpsE (53)and EpsL and EpsM (data not shown) by purification andimmunodetection. Work on identification of other Eps pro-teins is in progress.
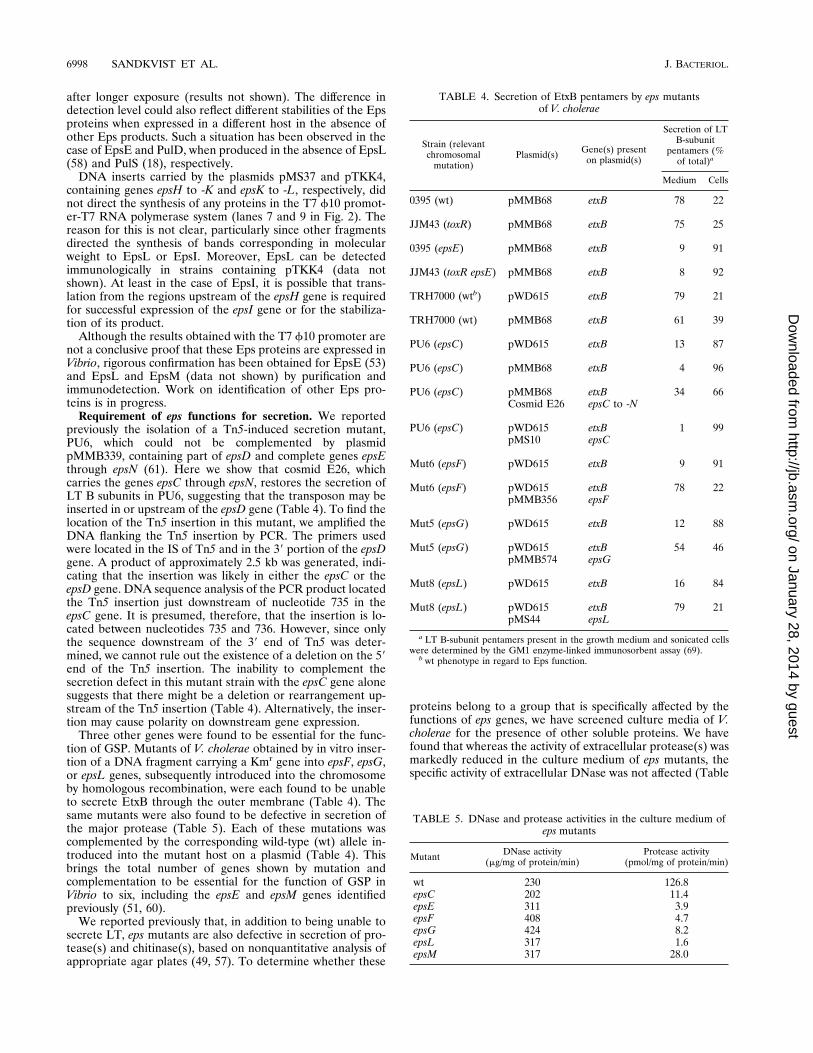
Requirement of eps functions for secretion. We reportedpreviously the isolation of a Tn5-induced secretion mutant,PU6, which could not be complemented by plasmidpMMB339, containing part of epsD and complete genes epsEthrough epsN (61). Here we show that cosmid E26, whichcarries the genes epsC through epsN, restores the secretion ofLT B subunits in PU6, suggesting that the transposon may beinserted in or upstream of the epsD gene (Table 4). To find thelocation of the Tn5 insertion in this mutant, we amplified theDNA flanking the Tn5 insertion by PCR. The primers usedwere located in the IS of Tn5 and in the 39 portion of the epsDgene. A product of approximately 2.5 kb was generated, indi-cating that the insertion was likely in either the epsC or theepsD gene. DNA sequence analysis of the PCR product locatedthe Tn5 insertion just downstream of nucleotide 735 in theepsC gene. It is presumed, therefore, that the insertion is lo-cated between nucleotides 735 and 736. However, since onlythe sequence downstream of the 39 end of Tn5 was deter-mined, we cannot rule out the existence of a deletion on the 59end of the Tn5 insertion. The inability to complement thesecretion defect in this mutant strain with the epsC gene alonesuggests that there might be a deletion or rearrangement up-stream of the Tn5 insertion (Table 4). Alternatively, the inser-tion may cause polarity on downstream gene expression.
Three other genes were found to be essential for the func-tion of GSP. Mutants of V. cholerae obtained by in vitro inser-tion of a DNA fragment carrying a Kmr gene into epsF, epsG,or epsL genes, subsequently introduced into the chromosomeby homologous recombination, were each found to be unableto secrete EtxB through the outer membrane (Table 4). Thesame mutants were also found to be defective in secretion ofthe major protease (Table 5). Each of these mutations wascomplemented by the corresponding wild-type (wt) allele in-troduced into the mutant host on a plasmid (Table 4). Thisbrings the total number of genes shown by mutation andcomplementation to be essential for the function of GSP inVibrio to six, including the epsE and epsM genes identifiedpreviously (51, 60).
We reported previously that, in addition to being unable tosecrete LT, eps mutants are also defective in secretion of pro-tease(s) and chitinase(s), based on nonquantitative analysis ofappropriate agar plates (49, 57). To determine whether these
proteins belong to a group that is specifically affected by thefunctions of eps genes, we have screened culture media of V.cholerae for the presence of other soluble proteins. We havefound that whereas the activity of extracellular protease(s) wasmarkedly reduced in the culture medium of eps mutants, thespecific activity of extracellular DNase was not affected (Table
TABLE 4. Secretion of EtxB pentamers by eps mutantsof V. cholerae
Strain (relevantchromosomal
mutation)Plasmid(s) Gene(s) present
on plasmid(s)
Secretion of LTB-subunit
pentamers (%of total)a
Medium Cells
0395 (wt) pMMB68 etxB 78 22
JJM43 (toxR) pMMB68 etxB 75 25
0395 (epsE) pMMB68 etxB 9 91
JJM43 (toxR epsE) pMMB68 etxB 8 92
TRH7000 (wtb) pWD615 etxB 79 21
TRH7000 (wt) pMMB68 etxB 61 39
PU6 (epsC) pWD615 etxB 13 87
PU6 (epsC) pMMB68 etxB 4 96
PU6 (epsC) pMMB68 etxB 34 66Cosmid E26 epsC to -N
PU6 (epsC) pWD615 etxB 1 99pMS10 epsC
Mut6 (epsF) pWD615 etxB 9 91
Mut6 (epsF) pWD615 etxB 78 22pMMB356 epsF
Mut5 (epsG) pWD615 etxB 12 88
Mut5 (epsG) pWD615 etxB 54 46pMMB574 epsG
Mut8 (epsL) pWD615 etxB 16 84
Mut8 (epsL) pWD615 etxB 79 21pMS44 epsL
a LT B-subunit pentamers present in the growth medium and sonicated cellswere determined by the GM1 enzyme-linked immunosorbent assay (69).
b wt phenotype in regard to Eps function.
TABLE 5. DNase and protease activities in the culture medium ofeps mutants
Mutant DNase activity(mg/mg of protein/min)
Protease activity(pmol/mg of protein/min)
wt 230 126.8epsC 202 11.4epsE 311 3.9epsF 408 4.7epsG 424 8.2epsL 317 1.6epsM 317 28.0
6998 SANDKVIST ET AL. J. BACTERIOL.
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

5). This indicates that extracellular DNase may be secreted bya pathway different from the GSP.
Outer membrane changes in eps mutants. During pheno-typic analysis of eps mutant V. cholerae, we observed throughmicroscopic examination that the cells form filaments thatmust have arisen as the consequence of a division defect (re-sults not shown). In addition, several of the eps mutants dem-onstrated lower growth rates. These observations are consis-tent with the possibility that eps mutants may have defectivecell envelopes. To examine whether this is so, we analyzedouter membrane proteins from wt (TRH7000) and eps mutantderivatives by SDS-PAGE. The wt TRH7000 expressed sixdetectable outer membrane protein species ranging in sizefrom 27 to 43 kDa. The largest of these proteins, approxi-mately 43 kDa, is induced by maltose and reacts with antibod-ies against OmpS, a homolog of the E. coli LamB protein (Fig.3A) (36). The other major band, which is not induced bymaltose, is likely the OmpT protein, as judged by its migrationon SDS-PAGE and its relative intensity (44). OmpT is a porinmolecule whose expression is negatively controlled by theToxR virulence regulator (5).
Membrane preparations from the eps mutants, exemplifiedby those of the epsC and epsM mutants shown in Fig. 3A, didnot differ from the wt extracts in the amounts of the 27-kDa
polypeptide, but the content of the putative OmpT protein aswell as the amounts of the minor bands was dramatically lower,and the amount of the OmpS protein was visibly diminished.When the amount of OmpT was compared among different epsmutants, we found that the epsE, epsF, and epsL mutants didnot contain any visible OmpT protein, whereas the epsC, epsG,and epsM mutants produced detectable levels of OmpT, albeitlower than those of the wt strain (Fig. 3B).
V. cholerae with a wt toxR gene typically produces the OmpUprotein, which is under positive control by ToxR, and does notproduce OmpT (10). As TRH7000 was considered to be wt fortoxR, we were therefore surprised to find that it expressedOmpT, but not OmpU (compare lanes 1, 8, and 9 in Fig. 3B).Western blotting with anti-ToxR serum demonstrated thatTRH7000 produces no detectable ToxR. The reason thatTRH7000 does not produce ToxR is not known; however, thisfact explains why it expresses OmpT and not OmpU.
In order to test whether the biogenesis of OmpU also re-quires an intact Eps apparatus, we analyzed the toxR1 strain,0395, and an epsE mutant thereof. The amount of OmpUrelative to that of other proteins detected in the outer mem-brane of the epsE mutant strain was lower than that in the wtstrain, indicating that, as with OmpT in the toxR mutant back-ground, the biogenesis of the mature OmpU is also dependenton a functioning Eps apparatus (Fig. 3B, lanes 8 and 10). Thelower level of OmpU in the outer membrane of the mutantstrain was not due to a difference in fractionation behavior dueto a more fragile membrane, since the amount of OmpU de-tected by immunoblot analysis in total cell lysate of the epsEmutant was also considerably lower than that in the wt strain(Fig. 3B, lanes 12 and 14; for comparison, a sample from thetoxR mutant strain JJM43 in lane 13 is also included). Analysisof ompU RNA by primer extension experiments demonstratedthat the mutation in epsE does not affect the level of ompUmRNA (data not shown). We conclude, therefore, that thelower amount of OmpU detected in the mutant strain is mostlikely due to degradation of OmpU due to its inability toproperly insert into the outer membrane. Alternatively, theepsE mutation may effect the translation of the OmpU protein.
Since the reason for the lack of ToxR production inTRH7000 is not known, the possibility exists that this straincarries other, unknown mutations. Therefore, we introduced amutation into the epsE gene of a toxR mutant and confirmedthat OmpT is dependent on the Eps machinery (Fig. 3B, lane11), consistent with the results obtained with TRH7000.
ToxR does not regulate the secretion of toxin. Because bio-genesis of the ToxR-regulated OmpU protein requires an in-tact Eps apparatus, we considered the possibility that ToxRmay also be involved in regulating eps gene expression. Toaddress this possibility, we assessed both the secretion of toxinsubunits and the expression of an epsE-lacZ transcriptionalfusion in wt and toxR mutant backgrounds.
For the secretion experiment, we used a plasmid (pMMB68)that expresses the B subunit of the E. coli LT, because the toxRmutant used in this experiment (JJM43) does not express CT.The amount of B subunit secreted by JJM43[pMMB68] wascompared to that of wt strain 0395 containing the same plas-mid, by using the monoclonal antibody 118-8, which is specificfor LT B subunits (therefore, CT B subunits were not detectedin this assay). The same amount of LT B subunits was pro-duced and secreted from these strains (Table 4), suggestingthat ToxR does not regulate the toxin secretion process.
To determine whether ToxR controls eps transcription, weconstructed a chromosomal epsE-lacZ operon fusion as de-scribed in Materials and Methods. This strain is deficient forEpsE production, which was confirmed by immunoblotting
FIG. 3. Outer membrane protein profiles. (A) Proteins of outer membranefractions from cells of wt V. cholerae TRH7000 and epsM and epsC mutantsgrown in LB with or without maltose were resolved by SDS-PAGE as describedin Materials and Methods. The Coomassie blue-stained gel (left panel) and theimmunoblot with anti-OmpS antibodies (right panel) are shown. (B) Outermembrane fractions from cells of wt V. cholerae TRH7000 (lane 1); mutants epsC(lane 2), epsE (lane 3), epsF (lane 4), epsG (lane 5), epsL (lane 6), and epsM (lane7); wt 0395 (lanes 8 and 12); toxR mutant JJM43 (lanes 9 and 13); epsE mutantof 0395 (lanes 10 and 14); and epsE mutant of JJM43 (lane 11). Left panel,Coomassie blue-stained gel; right panel, immunoblot with anti-OmpU antibod-ies.
VOL. 179, 1997 EXTRACELLULAR SECRETION IN V. CHOLERAE 6999
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

(not shown), due to the insertion of the fusion by homologousrecombination. The epsE-lacZ strains were no longer able tosecrete the B subunits to the extracellular environment, irre-spective of the toxR status (Table 4). When these strains wereanalyzed for b-galactosidase production, they were found toproduce similar amounts (approximately 600 and 800 Millerunits for wt and toxR mutant strains, respectively), demonstrat-ing that the expression of epsE is not regulated by ToxR.
Processing of EpsG in V. cholerae. As stated earlier, thepredicted amino acid sequences of EpsG through EpsJ pro-teins contain hydrophobic regions which resemble the signalpeptides of type IV pilin.
A gene encoding a prepilin peptidase with sequence simi-larities to genes such as pulO of Klebsiella oxytoca or pilD of P.aeruginosa has not been found in the eps gene cluster se-quenced in this study. The available sequences upstream of theepsC gene and downstream of the epsN gene do not containORFs with any homology to the sequences for known prepilinpeptidases.
In order to determine whether EpsG is processed in V.cholerae, the epsG gene was inserted downstream of the T7f10 promoter in pT7-5 to create pMMB564. This plasmid wasintroduced into E. coli MC1061 and V. cholerae TRH7000expressing the T7 polymerase gene. The EpsG protein pro-duced by the plasmid was labeled with [35S]Met and visualizedby SDS-PAGE and autoradiography (Fig. 4A). It was foundthat in E. coli, the EpsG protein was seen as a single band,whereas in V. cholerae, an additional band of lower Mr wasproduced. This suggested that the second band was a pro-cessed form of EpsG. The inefficient processing of radiola-beled EpsG in V. cholerae was most likely due to the continu-ous labeling conditions, since in experiments in which proteinswere pulse-labeled for only 30 s and then chased with unla-beled methionine most of the EpsG was present in the pro-cessed form at the end of the chase (Fig. 4B).
It could be imagined that the prepilin peptidase encoded bythe tcpJ gene of V. cholerae (31) might be involved in theprocessing of the EpsG through EpsJ proteins. This would be
analogous to the dual function of the prepilin peptidase PilD inPseudomonas, in which PilD is responsible for the processingof both the type IV prepilin subunits and the pilin-like proteinsXcpT to XcpW required for extracellular secretion (44, 62).However, the expression of epsG in a tcpJ mutant of V. choleraeresulted in a two-band pattern of EpsG similar to that in the wtV. cholerae strain (Fig. 4A). In addition, coexpression of thetcpJ and epsG genes in E. coli did not result in an alteration inthe migration of the EpsG protein (Fig. 5, lane 5), indicatingthat EpsG is not a substrate for TcpJ. On the other hand,expression of the N. gonorrhoeae gene pilD in the same E. colihost as epsG resulted in the processing of EpsG to a smallerprotein (Fig. 5, lane 6), demonstrating that EpsG can be thesubstrate for a prepilin peptidase. A slight degree of EpsGproteolytic processing detectable in E. coli in the absence ofany exogenous genes (Fig. 5, lanes 2 to 5) is probably due toendogenous prepilin peptidase, since some strains have beendemonstrated to express proteolytic activity analogous to thatof prepilin peptidases (33). The K-12 genome has been re-ported to carry at least two genes which by all criteria encodehomologs of the prepilin peptidases associated with the GSPand type IV pilus biogenesis systems (15). The failure to detectthis endogenous processing in the T7 expression studies mayreflect differences in strain backgrounds.
As noted earlier, no sequence homologous to other prepilinpeptidase genes could be observed 600 bp downstream of theepsN gene. Since DNA of approximately 3.5 kb downstream ofepsN was available, we tested this for the presence of a pepti-dase gene. The KpnI-EcoRI fragment containing DNA down-stream of epsN was cloned in both orientations (Fig. 1) and wascoexpressed with epsG in E. coli. No processing of EpsG couldbe observed in the presence of this insert, demonstrating thatno prepilin peptidase able to process EpsG is encoded by DNAsequences adjacent to the eps gene cluster (Fig. 5, lanes 3 and4).
FIG. 4. Processing of EpsG protein in V. cholerae. (A) Cells of E. coli and V.cholerae strains containing plasmid pMMB586, expressing the T7 RNA polymer-ase, and plasmid pMMB564, expressing the epsG gene from the T7 f10 pro-moter, were labeled as indicated in Materials and Methods. Total cell proteinswere subjected to SDS-PAGE and autoradiography. Lanes: 1, V. choleraeTRH7000[pMMB564; pMMB586]; 2, E. coli MC1061[pMMB564; pMMB586]; 3,V. cholerae tcpJ mutant[pMMB564; pMMB586]. (B) Cells of V. cholerae 0395tcpJ::Tn5 containing plasmids pMMB586 and pMMB564 were pulse-labeled andchased as described in Materials and Methods. Lanes: 1, 0.5 min; 2, 1 min; 3, 2min; 4, 5 min; 5, 10 min after the chase.
FIG. 5. Processing of EpsG by gonococcal prepilin peptidase PilD. Totalmembrane proteins from E. coli BL21(DE3), expressing the T7 RNA polymer-ase, grown in LB and induced with 0.1 mM isopropyl-b-D-thiogalactopyranosidefor 2 h in the absence (lane 1) or presence (lanes 2 to 6) of plasmid pMS19expressing preEpsG, were analyzed by SDS-PAGE and Coomassie blue staining.Samples in lanes 3 to 6 correspond to strains with the additional plasmids pMS13(lane 3), pMS14 (lane 4), pMS21 (lane 5), and pPilD (lane 6). Numbers at leftshow molecular mass in kilodaltons.
7000 SANDKVIST ET AL. J. BACTERIOL.
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

DISCUSSION
CT is secreted via a two-step pathway that requires a specificset of genes, eps, for the outer membrane translocation. Herewe report the cloning and sequencing of these genes. Muta-tions in the epsC, epsE, epsF, epsG, epsL, and epsM genes havebeen introduced, either through random transposon mutagen-esis or through homologous recombination, and have resultedin the accumulation of assembled toxin in the periplasm. Withthe exception of the epsC mutation, in each case, the secretiondefect could be complemented by the corresponding wt alleleon a plasmid, indicating that each gene is essential for secre-tion and that the secretion defect obtained by introducingmutations in upstream genes is not due to polarity on down-stream genes. Since several eps ORFs overlap each other,suggesting that they may be part of a single operon, it is likelythat also epsD, -H, -I, -J, -K, and -N are required for secretion.Collmer and coworkers have demonstrated the requirementfor secretion for all -C to -N genes, except for the -H homolog,in Erwinia (38, 39). Other genes, in addition to the eps genesidentified in this study, are most likely also needed for thetranslocation of proteins through the outer membrane sinceexpression of the epsC to -N genes in E. coli does not lead toextracellular secretion of LT or EtxB (results not shown). Thisis in contrast to the secretion of pullulanase, which can beobtained in E. coli in the presence of the complete pul genecluster, pulC through pulO, and pulS (19, 53). It is likely,therefore, that eps genes homologous to pulO and pulS areessential for secretion in V. cholerae and that these are absentfrom the epsC to -N cluster. It is, however, also possible thatthe eps genes are not expressed properly in E. coli. A cluster ofgenes homologous to eps, with similar organization, has beendetected in the E. coli chromosome, but at least two of thesegenes, gspG and gspO, appear not to be expressed under nor-mal laboratory growth conditions (16).
In addition to defects in extracellular secretion, eps mutantsalso demonstrate altered outer membrane profiles. In partic-ular, the biogenesis of two major outer membrane proteins,OmpU and OmpT, is severely affected. Defects in the outermembrane were previously observed in exe mutants of Aero-monas (26, 28). It seems likely, therefore, that the GSP func-tions are required not only for extracellular secretion of solu-ble proteins but also for proper assembly of the outermembrane in V. cholerae. They may be responsible for trans-port of the affected components, such as OmpU and OmpT,into the outer membrane or for the transport of other proteinsrequired for proper assembly and anchoring of the affectedproteins in the membrane. A correlation was observed be-tween the amount of OmpT or OmpU detected in the variousmutant strains and the severity of the growth defect: the lessOmpT and OmpU protein produced, the greater the growthdefect. The reason for this is not known. We speculate thatsince OmpT and OmpU are major outer membrane proteins inV. cholerae, they may have stabilizing effects on the membraneand, when they are missing, the integrity of the cell envelopemight be altered, resulting in a growth defect. Consistent withthis is that the outer membranes in some of the mutants areleaky. For instance, 25 to 30% of the periplasmic protein b-lactamase is found in the growth medium of certain mutants,whereas the level of this enzyme in medium from the wt strainis less than 5%.
Although the eps mutants appear to be defective in a num-ber of different functions, the defect in extracellular secretionof toxin and protease is not simply a secondary result from ageneral defect in the outer membrane, since the secretion ofyet another extracellular protein, DNase, is not affected. In
addition, our findings demonstrate that at least two differentpathways for extracellular secretion exist in V. cholerae. WhileCT and protease use the general secretion pathway, the secre-tion of DNase occurs by a different, as yet unidentified secre-tion mechanism. V. cholerae is known to secrete two differentDNases into the extracellular environment (15, 47). In ourassay for total DNase activity, we cannot determine whetherwe are detecting one or the other, or both DNases. However,our data suggest that the secretion of at least one of theDNases is not affected in our mutant strains.
No gene encoding a prepilin peptidase active in processingEpsG to EpsJ was identified adjacent to the epsC-to-epsN genecluster. This is different from most other GSP operons, inwhich a prepilin peptidase gene is present directly downstreamof the -N gene (3, 12, 55). However, it demonstrates the sim-ilarity of the V. cholerae GSP system to GSP systems in P.aeruginosa and Aeromonas hydrophila, in which the peptidasegene is found elsewhere on the chromosome. In contrast to P.aeruginosa, in which the prepilin peptidase PilD is required forthe processing of both the type IV prepilin subunits and theEpsG-to-EpsJ homologs, XcpT to XcpW, it was demonstratedthat in V. cholerae a prepilin peptidase different from TcpJ isable to cleave EpsG. On the other hand, prepilin peptidasePilD of N. gonorrhoeae is able to cleave EpsG of V. cholerae,demonstrating that this protein can be a substrate for a prepilinpeptidase, thus suggesting that it is likely that processing ofEpsG is required for its function. The signal sequence of EpsGmore closely resembles the signal sequences of other type IVprepilins and other pilin-like proteins than the one in the V.cholerae type IV pilin, TcpA. The reason that TcpJ cannotprocess EpsG might be a shorter signal sequence and thepresence of phenylalanine instead of methionine at position11 in the mature portion of EpsG. Since the tcpJ gene does notseem to be involved in either the secretion of CT (31) or theprocessing of the EpsG protein, it is probable that a secondgene, encoding a prepilin peptidase active in the GSP and witha specificity similar to that of the gonococcal PilD, exists in V.cholerae. In fact, prior evidence for a second prepilin peptidasein V. cholerae can be found in the observation that TcpAprocessing is not completely abolished in a tcpJ mutant (29).Moreover, the precedent for multiple prepilin peptidase geneshas already been established in E. coli (15). Although ourexperiments do not show that the observed modification of theEpsG is required for its function in the GSP, they constitute anindication, making the search for a second prepilin peptidasein V. cholerae interesting and important.
ACKNOWLEDGMENTS
We thank Molly Brennan for technical assistance. We are grateful toTapio Palva for the gift of anti-OmpS antibodies, Ron Taylor for thetcpJ mutant and clone, and Darren Higgins for a DNA fragmentencoding a promoterless lacZ gene.
This work was supported by grants from the U.S. Department ofAgriculture (92-37204-7839), the NSF Center for Microbial Ecology(BIR 9120006), and the Research Excellence Fund of the State ofMichigan to M.B. and by Public Health Service grant AI-31645 toV.J.D. M.S. was an NRSA fellow of the Public Health Service undergrant T32 AI-07360 to the University of Michigan. Sequence analysiswas supported in part by grant MO1 RR00042 to the Clinical ResearchCenter at the University of Michigan.
REFERENCES
1. Alm, R. A., J. P. Hallinan, A. A. Watson, and J. S. Mattick. 1996. Fimbrialbiogenesis genes of Pseudomonas aeruginosa: pilW and pilX increase thesimilarity of type 4 fimbriae to the GSP protein-secretion systems and pulY1encodes a gonococcal PilC homologue. Mol. Microbiol. 22:161–173.
2. Bally, M., G. Ball, A. Badere, and A. Lazdunski. 1991. Protein secretion inPseudomonas aeruginosa: the xcpA gene encodes an integral inner membrane
VOL. 179, 1997 EXTRACELLULAR SECRETION IN V. CHOLERAE 7001
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

protein homologous to Klebsiella pneumoniae secretion function proteinPulO. J. Bacteriol. 173:479–486.
3. Bally, M., A. Filloux, M. Akrim, G. Ball, A. Lazdunski, and J. Tommassen.1992. Protein secretion in Pseudomonas aeruginosa: characterization of sevenxcp genes and processing of secretory apparatus components by prepilinpeptidase. Mol. Microbiol. 6:1121–1131.
4. Casadaban, M. C., and S. N. Cohen. 1980. Analysis of gene control signalsby DNA fusion in Escherichia coli. J. Mol. Biol. 138:179–207.
5. Chakrabarti, S. R., K. Chaudhuri, K. Sen, and K. Das. 1996. Porins of Vibriocholerae: purification and characterization of OmpU. J. Bacteriol. 178:524–530.
6. Condemine, G., C. Dorel, N. Hugovieux-Cotte-Pattat, and J. Robert-Bau-douy. 1992. Some of the out genes involved in the secretion of pectate lyasesin Erwinia chrysanthemi are regulated by kdgR. Mol. Microbiol. 6:3199–3211.
7. Dallas, W. S. 1983. Conformity between heat-labile toxin genes from humanand porcine enterotoxigenic Escherichia coli. Infect. Immun. 40:647–652.
8. Dallas, W. S., D. M. Gill, and S. Falkow. 1979. Cistrons encoding Escherichiacoli heat-labile toxin. J. Bacteriol. 139:850–858.
9. d’Enfert, C., A. Ryter, and A. P. Pugsley. 1987. Cloning and expression inEscherichia coli of the Klebsiella pneumoniae genes for production, surfacelocalization and secretion of the lipoprotein pullulanase. EMBO J. 6:3531–3538.
10. Devereux, J., P. Haeberli, and O. Smithies. 1984. A comprehensive set ofsequence analysis programs for the VAX. Nucleic Acids Res. 12:387–395.
11. Dums, F., G. M. Dow, and M. J. Daniels. 1991. Structural characterization ofprotein secretion genes of the bacterial phytopathogen Xanthomonascampestris pathovar campestris: relatedness to secretion systems of othergram-negative bacteria. Mol. Gen. Genet. 229:357–364.
12. Dupuy, B., and A. P. Pugsley. 1994. Type IV prepilin peptidase gene ofNeisseria gonorrhoeae MS11: presence of a related gene in other piliated andnonpiliated Neisseria strains. J. Bacteriol. 176:1323–1331.
13. Findlay, G., J. Yu, and T. R. Hirst. 1993. Analysis of enterotoxin synthesis ina Vibrio cholerae strain lacking DsbA, a periplasmic enzyme involved indisulphide bond formation. Biochem. Soc. Trans. 21:212S.
14. Finkelstein, R. A., M. Boesman-Finkelstein, Y. Chang, and C. C. Hase. 1992.Vibrio cholerae hemagglutinin/protease, colonial variation, virulence, anddetachment. Infect. Immun. 60:472–478.
15. Focareta, T., and P. A. Manning. 1991. Distinguishing between the extracel-lular DNases of Vibrio cholerae and development of a transformation system.Mol. Microbiol. 5:2547–2555.
16. Francetic, O., and A. P. Pugsley. 1996. The cryptic general secretory pathway(gsp) operon of Escherichia coli K-12 encodes functional proteins. J. Bacte-riol. 178:3544–3549.
17. Furste, J. P., W. Pansegrau, R. Frank, H. Blocker, P. Scholz, M. Bagdasar-ian, and E. Lanka. 1986. Molecular cloning of the plasmid RP4 primaseregion in a multi-host-range tacP expression vector. Gene 48:119–131.
18. Gill, D. M., J. D. Clements, D. C. Robertson, and R. A. Finkelstein. 1981.Subunit number and arrangement in Escherichia coli heat-labile enterotoxin.Infect. Immun. 33:677–682.
19. Hardie, K. R., S. Lory, and A. P. Pugsley. 1996. Insertion of an outermembrane protein in Escherichia coli requires a chaperon-like protein.EMBO J. 15:978–988.
20. Hase, C. C., M. E. Bauer, and R. A. Finkelstein. 1994. Genetic character-ization of mannose-sensitive hemagglutinin (MSHA)-negative mutants ofVibrio cholerae derived by Tn5 mutagenesis. Gene 150:17–25.
21. He, S. Y., M. Lindeberg, A. K. Chatterjee, and A. Collmer. 1991. ClonedErwinia chrysanthemi out genes enable Escherichia coli to selectively secretea diverse family of heterologous proteins to its milieu. Proc. Natl. Acad. Sci.USA 88:1079–1083.
22. Herrington, D. A., R. H. Hall, G. Losonsky, J. J. Mekalanos, R. K. Taylor,and M. M. Levine. 1988. Toxin, toxin-coregulated pili, and the ToxR regulonare essential for Vibrio cholerae pathogenesis in humans. J. Exp. Med. 168:1487–1492.
23. Hirst, T. R., and J. Holmgren. 1987. Conformation of protein secreted acrossbacterial outer membranes: a study of enterotoxin translocation from Vibriocholerae. Proc. Natl. Acad. Sci. USA 84:7418–7422.
24. Hirst, T. R., L. L. Randall, and S. J. S. Hardy. 1984. Cellular location ofheat-labile enterotoxin in Escherichia coli. J. Bacteriol. 157:637–642.
25. Hofstra, H., and B. Witholt. 1984. Kinetics of synthesis, processing, andmembrane transport of heat-labile enterotoxin, a periplasmic protein inEscherichia coli. J. Biol. Chem. 259:15182–15187.
26. Howard, P. S., J. Critch, and A. Bedi. 1993. Isolation and analysis of eight exegenes and their involvement in extracellular protein secretion and outermembrane assembly in Aeromonas hydrophila. J. Bacteriol. 175:6695–6703.
27. Hu, N.-T., M.-N. Hung, S.-J. Chiou, F. Tang, D.-C. Chiang, H.-Y. Huang, andC.-Y. Wu. 1992. Cloning and characterization of a gene required for thesecretion of extracellular enzymes across the outer membrane by Xanthomo-nas campestris pv. campestris. J. Bacteriol. 174:2679–2687.
28. Jiang, B., and S. P. Howard. 1992. The Aeromonas hydrophila exeE gene,required both for protein secretion and normal outer membrane biogenesis,is a member of a general secretion pathway. Mol. Microbiol. 6:1351–1361.
29. Julius, D., A. Brake, L. Blair, R. Kunisawa, and J. Thorner. 1984. Isolation
of the putative structural gene for the lysine-arginine-cleaving endopeptidaserequired for processing of yeast prepro-a-factor. Cell 37:1075–1089.
30. Kaper, J. B., J. J. G. Morris, and M. M. Levine. 1995. Cholera. Clin.Microbiol. Rev. 8:48–86.
31. Kaufman, M. R., J. M. Seyer, and R. K. Taylor. 1991. Processing of TCP pilinby TcpJ typifies a common step intrinsic to a newly recognized pathway ofextracellular protein secretion by Gram-negative bacteria. Genes Dev. 5:1834–1846.
32. Kaufman, M. R., C. E. Shaw, I. D. Jones, and R. K. Taylor. 1993. Biogenesisand regulation of the Vibrio cholerae toxin-coregulated pilus: analogies toother virulence factor secretory systems. Gene 126:43–49.
33. Koomey, M., S. Bergstrom, M. Blake, and J. Swanson. 1991. Pilin expressionand processing in pilus mutants of Neisseria gonorrhoeae: critical role ofGly-1 in assembly. Mol. Microbiol. 5:279–287.
34. Kyte, J., and R. F. Doolittle. 1982. A simple method for displaying thehydropathic character of a protein. J. Mol. Biol. 157:105–132.
35. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature (London) 227:680–685.
36. Lång, H., and E. T. Palva. 1993. The ompS gene of Vibrio cholerae encodesa growth-phase dependent maltoporin. Mol. Microbiol. 10:891–901.
37. Lauer, P., N. H. Albertson, and M. Koomey. 1993. Conservation of genesencoding components of a type IV pilus assembly/two-step protein exportpathway in Neisseria gonorrhoeae. Mol. Microbiol. 8:357–368.
38. Lindeberg, M., and A. Collmer. 1992. Analysis of eight out genes in a clusterrequired for pectic enzyme secretion by Erwinia chrysanthemi: sequencecomparison with secretion genes from other gram-negative bacteria. J. Bac-teriol. 174:7385–7397.
39. Lindeberg, M., G. Salmond, and A. Collmer. 1996. Complementation ofdeletion mutations in a cloned functional cluster of Erwinia chrysanthemi outgenes with Erwinia carotovora out homologues revealed OutC and OutD ascandidate gatekeepers of species-specific secretion of proteins via type IIpathway. Mol. Microbiol. 20:175–190.
40. Linderoth, N. A., P. Model, and M. Russel. 1996. Essential role of a sodiumdodecyl sulfate-resistant protein IV multimer in assembly-export of filamen-tous phage. J. Bacteriol. 178:1962–1970.
41. Marsh, J. W., D. Sun, and R. K. Taylor. 1996. Physical linkage of the Vibriocholerae mannose-sensitive hemagglutinin secretory and structural subunitgene loci: identification of the mshG coding sequence. Infect. Immun. 64:460–465.
42. Mekalanos, J. J., D. J. Swartz, G. D. N. Pearson, N. Harford, F. Groyne, andM. de Wilde. 1983. Cholera toxin genes: nucleotide sequence, deletion anal-ysis and vaccine development. Nature (London) 306:551–557.
43. Miller, J. H. 1972. Experiments in molecular genetics. Cold Spring HarborLaboratory Press, Cold Spring Harbor, N.Y.
44. Miller, V. L., and J. J. Mekalanos. 1988. A novel suicide vector and its usein construction of insertion mutations: osmoregulation of outer membraneproteins and virulence determinants in Vibrio cholerae requires toxR. J.Bacteriol. 170:2575–2583.
45. Morales, V. M., A. Backman, and M. Bagdasarian. 1991. A series of wide-host-range low-copy-number vectors that allow direct screening for recom-binants. Gene 97:39–47.
46. Neuwald, A. F., B. R. Krishnan, I. Brikun, S. Kulakauskas, K. Suziedelis, T.Tomcsanyi, T. S. Leyh, and D. E. Berg. 1992. cysQ, a gene needed forcysteine synthesis in Escherichia coli K-12 only during aerobic growth. J.Bacteriol. 174:415–425.
47. Newland, J. W., B. A. Green, J. Foulds, and R. K. Holmes. 1985. Cloning ofextracellular DNase and construction of a DNase-negative strain of Vibriocholerae. Infect. Immun. 47:691–696.
48. Nunn, D., and S. Lory. 1991. Product of the Pseudomonas aeruginosa genepilD is a prepilin leader peptidase. Proc. Natl. Acad. Sci. USA 88:3281–3285.
49. Nunn, D., and S. Lory. 1993. Cleavage, methylation, and localization of thePseudomonas aeruginosa export proteins XcpT, -U, -V, and -W. J. Bacteriol.175:4375–4382.
50. Ogierman, M. A., S. Zabihi, L. Mourtzios, and P. A. Manning. 1993. Geneticorganization and sequence of the promoter-distal region of the tcp genecluster of Vibrio cholerae. Gene 126:51–60.
51. Overbye, L. J., M. Sandkvist, and M. Bagdasarian. 1993. Genes required forextracellular secretion of enterotoxin are clustered in Vibrio cholerae. Gene132:101–106.
52. Palva, E. T., T. R. Hirst, S. J. S. Hardy, J. Holmgren, and L. L. Randall.1981. Synthesis of a precursor to the B subunit of heat-labile enterotoxin inEscherichia coli. J. Bacteriol. 146:325–330.
53. Possot, O., C. d’Enfert, I. Reyss, and A. P. Pugsley. 1992. Pullulanase secre-tion in Escherichia coli K-12 requires a cytoplasmic protein and a putativepolytopic cytoplasmic membrane protein. Mol. Microbiol. 6:95–105.
54. Pugsley, A. P. 1993. Processing and methylation of PulG, pilin-like compo-nent of the general secretory pathway of Klebsiella oxytoca. Mol. Microbiol.9:295–308.
55. Pugsley, A. P., and B. Dupuy. 1992. An enzyme with type IV prepilin pep-tidase activity is required to process components of the general extracellularprotein secretion pathway of Klebsiella oxytoca. Mol. Microbiol. 6:751–760.
56. Reeves, P. J., P. Douglas, and G. P. C. Salmond. 1994. Beta-lactamase
7002 SANDKVIST ET AL. J. BACTERIOL.
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

topology probe analysis of the OutO NMePhe peptidase, and six other Outprotein components of the Erwinia carotovora general secretion pathwayapparatus. Mol. Microbiol. 12:445–457.
57. Reeves, P. J., D. Whitcombe, S. Wharam, M. Gibson, G. Allison, N. Bunce,R. Barallon, P. Douglas, V. Mulholland, S. Stevens, D. Walker, and G. P. C.Salmond. 1993. Molecular cloning and characterization of 13 out genes fromErwinia carotovora subspecies carotovora: genes encoding members of ageneral secretion pathway (GSP) widespread in Gram-negative bacteria.Mol. Microbiol. 8:443–456.
58. Sandkvist, M., M. Bagdasarian, S. P. Howard, and V. J. DiRita. 1995.Interactions between the autokinase EpsE and EpsL in the cytoplasmicmembrane is required for extracellular secretion in Vibrio cholerae. EMBOJ. 14:1664–1673.
59. Sandkvist, M., T. R. Hirst, and M. Bagdasarian. 1987. Alterations at thecarboxyl terminus change assembly and secretion properties of the B subunitof Escherichia coli heat-labile enterotoxin. J. Bacteriol. 169:4570–4576.
60. Sandkvist, M., V. Morales, and M. Bagdasarian. 1993. A protein requiredfor secretion of cholera toxin through the outer membrane of Vibrio cholerae.Gene 123:81–86.
61. Sandkvist, M., L. J. Overbye, T. K. Sixma, W. G. J. Hol, and M. Bagdasar-ian. 1993. Assembly of Escherichia coli heat-labile enterotoxin and its secre-tion from Vibrio cholerae, p. 293–309. In C. Kado, J. Crossa, and L. Sequeira(ed.), Molecular mechanisms of bacterial virulence. Academic Publishers,Dordrecht, The Netherlands.
62. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencing withchain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467.
63. Schatz, P. J., and J. Beckwith. 1990. Genetic analysis of protein export inEscherichia coli. Annu. Rev. Genet. 24:215–248.
64. Sixma, T. K., K. H. Kalk, B. A. M. van Zanten, Z. Dauter, J. Kingma, B.Witholt, and W. G. J. Hol. 1993. Refined structure of Escherichia coli heat-
labile enterotoxin, a close relative of cholera toxin. J. Mol. Biol. 230:890–918.65. Sixma, T. K., S. E. Pronk, K. H. Kalk, E. S. Wartna, B. A. M. Van Zanten,
B. Witholt, and W. G. J. Hol. 1991. Crystal structure of a cholera toxin-related heat-labile enterotoxin from E. coli. Nature (London) 353:371–377.
66. Spangler, B. D. 1992. Structure and function of cholera toxin and the relatedEscherichia coli heat-labile enterotoxin. Microbiol. Rev. 56:622–647.
67. Strom, M. S., D. N. Nunn, and S. Lory. 1993. A single bifunctional enzyme,PilD, catalyzes cleavage and N-methylation of proteins belonging to the typeIV pilin family. Proc. Natl. Acad. Sci. USA 90:2404–2408.
68. Studier, F. W., A. H. Rosenberg, J. J. Dunn, and J. W. Dubendorf. 1990. Useof T7 RNA polymerase to direct expression of cloned genes. Methods En-zymol. 185:60–89.
69. Svennerholm, A.-M., and J. Holmgren. 1978. Identification of Escherichiacoli heat-labile enterotoxin by means of a ganglioside immunosorbent assay(GM1-ELISA) procedure. Curr. Microbiol. 1:19–23.
70. Tabor, S., and C. C. Richardson. 1985. A bacteriophage T7 RNA polymer-ase/promoter system for controlled exclusive expression of specific genes.Proc. Natl. Acad. Sci. USA 82:1074–1078.
71. Tamplin, M. L., A. L. Gauzens, A. Huq, D. A. Sack, and R. R. Colwell. 1990.Attachment of Vibrio cholerae serogroup O1 to zooplankton and phytoplank-ton of Bangladesh waters. Appl. Environ. Microbiol. 56:1977–1980.
72. Thien, S. L. 1989. A simplified method of direct sequencing of PCR ampli-fied DNA with Sequenase T7 DNA polymerase. USB Editorial Comments16:8,18.
73. Tommassen, J., A. Filloux, M. Bally, M. Murgier, and A. Lazdunski. 1992.Protein secretion in Pseudomonas aeruginosa. FEMS Microbiol. Rev. 103:73–90.
74. von Heijne, G. 1986. A new method for predicting signal sequence cleavagesites. Nucleic Acids Res. 14:4683–4690.
VOL. 179, 1997 EXTRACELLULAR SECRETION IN V. CHOLERAE 7003
on January 28, 2014 by guesthttp://jb.asm
.org/D
ownloaded from




















