
Radiosensitive populations and recovery in X-ray-induced apoptosis in the developing cerebellum
10
Acta Neuropathol (1993) 86:491 - 500 Acta . . Heuropatho Ka Springer-Verlag1993 Radiosensitive populations and recovery in X-ray-induced apoptosis in the developing cerebellum I. Ferrer 1, T. Serrano 1, R. Rivera 1, M. Oliv61, M. J. Zfijar 1, E Graus 2 1 Unidad de Neuropatologia, Servicio de Anatomia Patol6gica, Hospital Principes de Espafia, Universidad de Barcelona, E-08907 Hospitalet de Llohregat, Spain 2 Servicio de Neurologfa, Hospital Clfnico, Barcelona, Spain Received: 19 April 1993/Revised, accepted: 14 June 1993 Abstract. Sprague-Dawley rats received a single dose of 2 Gy X-rays at the age of 1 or 3 days and were killed at different intervals. Dying cells with the morphological characteristics of apoptosis appeared in the external and internal granular layers (EGL and IGL) and white matter (WM) of the cerebellum, mainly 3--6 h after irradiation, and decreased thereafter to reach normal values between 48 h and 5 days later. This process was curbed by the injection of cycloheximide at a dose of 1 ~g/g body weight. In addition, the number of mitoses in EGL rapidly decreased after irradiation and did not reach normal values until a few days later. Proliferating cell nuclear antigen (PCNA)-immunoreactive cells, which were chiefly found in EGL but also in IGL and WM, dramatically decreased in number from 3 to 48 h after irradiation. PCNA-immunoreactive cells reap- peared and reached age-matched values in the following days. Hu (considered as an early neuronal marker) and vimentin immunocytochemistry disclosed that Hu-non- reactive cells in the upper level of EGL, Hu-immunore- active cells in the inner level of EGL, Bergmann glia and many astrocytes in WM, as well as many non-typified cells in WM, were radiosensitive populations, whereas Purkinje cells were not. The present results indicate that irradiation at P1 or P3 blocks mitosis in EGL and kills sensitive cells mainly in the late G1 and S phases of the cell cycle, probably by apoptosis through a protein synthesis-mediated process. Radiosensitive cells are germinal cells and neuroblasts in EGL, Bergmann glia, astrocytes in WM, and non-typified cells, probably glial cell precursors, in WM. Surviving cells in EGL and PCNA-immunoreactive cells in other cortical layers and white matter reconstitute the cerebellum following a single dose of X-rays. This work was supported in part by CEC project FI3P-CqX)2-0015 and FIS grant 93-131. M. Oliv6 is the recipient of a grant from the Pi i Sunyer Foundation. R. Rivera has a grant from the CIRIT Correspondence to: I. Ferrer Key words: X-rays - Hu - Vimentin - Proliferating cell nuclear antigen - Cerebellum Cerebellar granule cells, and most particularly the cells of the external granular layer, are sensitive to irradiation [1, 2, 12, 21, 22, 24, 25]. Although repeated doses of X-rays at appropriate times of gestation and early postnatal period may produce cerebellar malformations [2, 12], external granular cells that survive the effects of a single dose of X-rays can proliferate and reconstitute the external granular layer (EGL) [1, 2]. Recent observations have suggested that X-ray- induced cell death in the developing cerebellum is reminiscent of apoptosis in other systems [20], and that the process is curbed by the injection of protein synthesis inhibitors [26]. In the present study we have examined, by means of different immunocytochemical markers, those cerebel- lar populations that are radiosensitive and those phases, of the cell cycle that are vulnerable to X-rays. It has also been our aim to investigate the distribution of cells participating in the process of cerebellar reconstitu- tion. Material and methods Sprague-Dawley rats of both sexes were irradiated with a single dose of 2 Gy X-rays at postnatal ages of I or 3 days (P1 or P3). The source of X-rays was a 300 Kvp Stabilipan (Siemens) with an HVL of 3.3 mm Cu. The animals irradiated at lal were killed at 30 and 60 min, 3, 6, 24, 48 h, 5, 15 and 25 days. The animals irradiated at P3 were killed at 30 and 60 min, 3, 6, 24, 48 h, 4 and 18 days post-irradiation (n = 3 for every interval). Another group of rats (n = 8 at P1, and n = 6 at P3) received cycloheximide i.p. (Actid- ione; ICN Biochemicals) at a dose of 1 ~g/g body wt., dissolved in saline, immediatly after irradiation, and was allowed to survive for 6 h. Age-matched rats (n = 26) were used as controls (three animals at the stages of P1--P5, P7, P15, P21 and two rats at P25). The animals were killed with an overdose of diethyl ether and the brains were fixed with Carnoy for 6 h. Sagital samples of the cerebellum
-
Upload
ciecs-conicet -
Category
Documents
-
view
0 -
download
0
Transcript of Radiosensitive populations and recovery in X-ray-induced apoptosis in the developing cerebellum

Acta Neuropathol (1993) 86:491 - 500 Acta . .
Heuropatho Ka �9 Springer-Verlag 1993
Radiosensitive populations and recovery in X-ray-induced apoptosis in the developing cerebellum
I. Ferrer 1, T. Serrano 1, R. Rivera 1, M. Oliv61, M. J. Zfijar 1, E Graus 2
1 Unidad de Neuropatologia, Servicio de Anatomia Patol6gica, Hospital Principes de Espafia, Universidad de Barcelona, E-08907 Hospitalet de Llohregat, Spain 2 Servicio de Neurologfa, Hospital Clfnico, Barcelona, Spain
Received: 19 April 1993/Revised, accepted: 14 June 1993
Abstract. Sprague-Dawley rats received a single dose of 2 Gy X-rays at the age of 1 or 3 days and were killed at different intervals. Dying cells with the morphological characteristics of apoptosis appeared in the external and internal granular layers ( E G L and IGL) and white mat te r (WM) of the cerebellum, mainly 3 - -6 h after irradiation, and decreased thereaf ter to reach normal values be tween 48 h and 5 days later. This process was curbed by the injection of cycloheximide at a dose of 1 ~g/g body weight. In addition, the number of mitoses in E G L rapidly decreased after irradiation and did not reach normal values until a few days later. Proliferating cell nuclear antigen (PCNA)- immunoreac t ive cells, which were chiefly found in E G L but also in I G L and WM, dramatically decreased in number f rom 3 to 48 h after irradiation. PCNA-immunoreac t ive cells reap- peared and reached age-matched values in the following days. H u (considered as an early neuronal marker ) and vimentin immunocytochemis t ry disclosed that Hu-non-
�9 reactive cells in the upper level of E G L , Hu- immunore- active cells in the inner level of E G L , Bergmann glia and many astrocytes in WM, as well as many non-typified cells in WM, were radiosensitive populat ions, whereas Purkinje cells were not. The present results indicate that irradiation at P1 or P3 blocks mitosis in E G L and kills sensitive cells mainly in the late G1 and S phases of the cell cycle, probably by apoptosis through a protein synthesis-mediated process. Radiosensitive cells are germinal cells and neuroblasts in E G L , Bergmann glia, astrocytes in WM, and non-typified cells, probably glial cell precursors, in WM. Surviving cells in E G L and PCNA-immunoreac t ive cells in other cortical layers and white ma t t e r reconsti tute the cerebel lum following a single dose of X-rays.
This work was supported in part by CEC project FI3P-CqX)2-0015 and FIS grant 93-131. M. Oliv6 is the recipient of a grant from the Pi i Sunyer Foundation. R. Rivera has a grant from the CIRIT
Correspondence to: I. Ferrer
Key words: X-rays - H u - Vimentin - Proliferating cell nuclear antigen - Cerebel lum
Cerebellar granule cells, and most particularly the cells of the external granular layer, are sensitive to irradiation [1, 2, 12, 21, 22, 24, 25]. Al though repeated doses of X-rays at appropr ia te times of gestat ion and early postnatal per iod may produce cerebellar malformat ions [2, 12], external granular cells that survive the effects of a single dose of X-rays can proliferate and reconsti tute the external granular layer (EGL) [1, 2].
Recent observations have suggested that X-ray- induced cell dea th in the developing cerebel lum is reminiscent of apoptosis in other systems [20], and that the process is curbed by the injection of protein synthesis inhibitors [26].
In the present study we have examined, by means of different immunocytochemical markers , those cerebel- lar populat ions that are radiosensitive and those phases, of the cell cycle that are vulnerable to X-rays. I t has also been our aim to investigate the distribution of cells participating in the process of cerebellar reconstitu- tion.
Material and methods
Sprague-Dawley rats of both sexes were irradiated with a single dose of 2 Gy X-rays at postnatal ages of I or 3 days (P1 or P3). The source of X-rays was a 300 Kvp Stabilipan (Siemens) with an HVL of 3.3 mm Cu. The animals irradiated at lal were killed at 30 and 60 min, 3, 6, 24, 48 h, 5, 15 and 25 days. The animals irradiated at P3 were killed at 30 and 60 min, 3, 6, 24, 48 h, 4 and 18 days post-irradiation (n = 3 for every interval). Another group of rats (n = 8 at P1, and n = 6 at P3) received cycloheximide i.p. (Actid- ione; ICN Biochemicals) at a dose of 1 ~g/g body wt., dissolved in saline, immediatly after irradiation, and was allowed to survive for 6 h. Age-matched rats (n = 26) were used as controls (three animals at the stages of P1--P5, P7, P15, P21 and two rats at P25). The animals were killed with an overdose of diethyl ether and the brains were fixed with Carnoy for 6 h. Sagital samples of the cerebellum
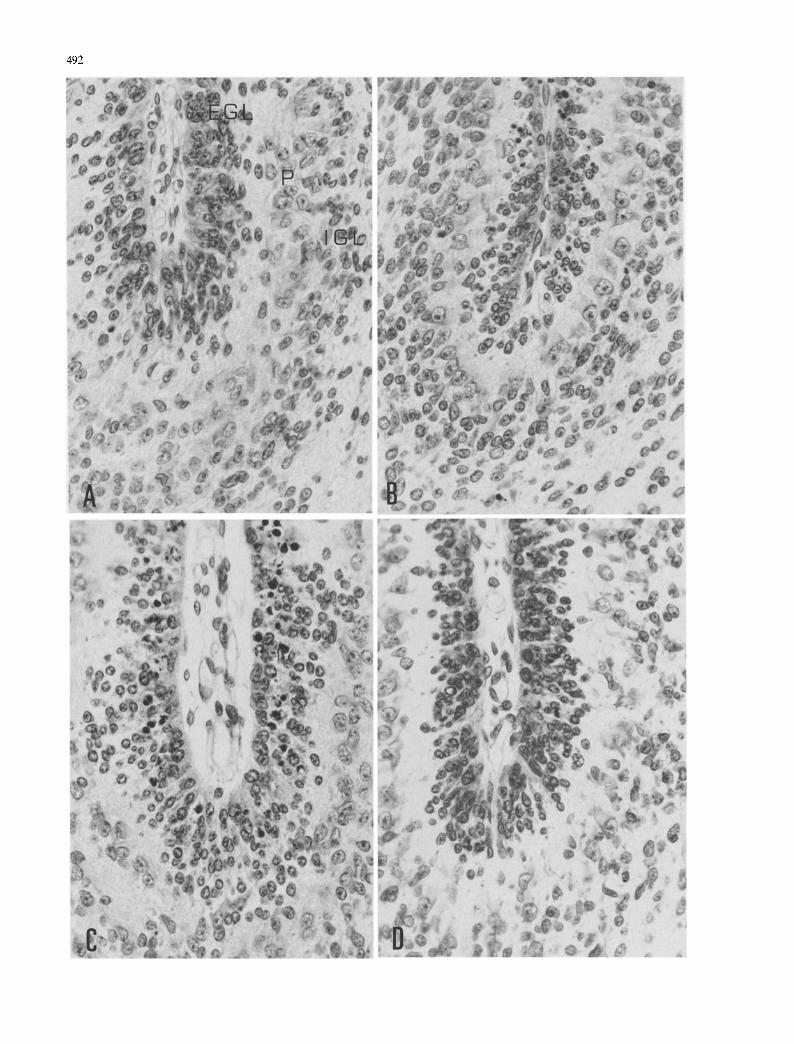
492
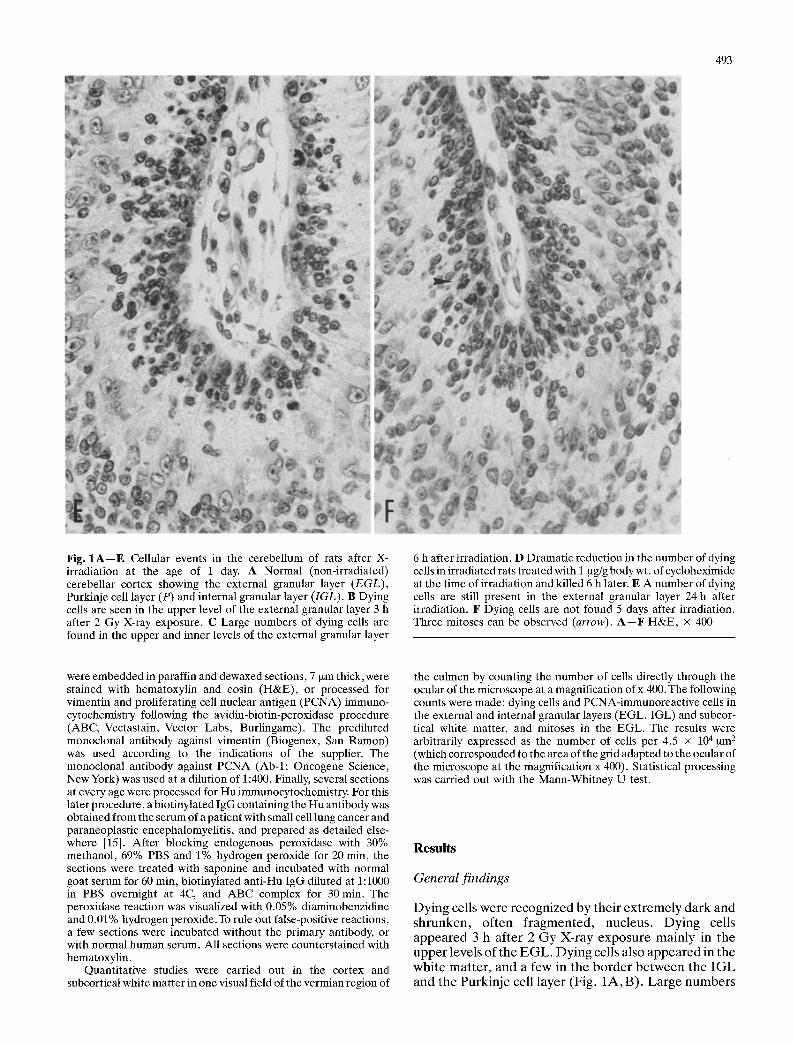
493
Fig. 1A- -E Cellular events in the cerebellum of rats after X- irradiation at the age of 1 day. A Normal (non-irradiated) cerebellar cortex showing the external granular layer (EGL), Purkinje cell layer (P) and internal granular layer (IGL). B Dying cells are seen in the upper level of the external granular layer 3 h after 2 Gy X-ray exposure. C Large numbers of dying cells are found in the upper and inner levels of the external granular layer
6 h after irradiation. D Dramatic reduction in the number of dying cells in irradiated rats treated with i btg/g body wt. of cycloheximide at the time of irradiation and killed 6 h later. E A number of dying cells are still present in the external granular layer 24 h after irradiation. F Dying cells are not found 5 days after irradiation. Three mitoses can be observed (arrow). A--F H&E, x 400
were embedded in paraffin and dewaxed sections, 7 ~m thick,were stained with hematoxylin and eosin (H&E), or processed for vimentin and proliferating cell nuclear antigen (PCNA) immuno- cytochemistry following the avidin-biotin-peroxidase procedure (ABC, Vectastain, Vector Labs, Burlingame). The prediluted monoclonal antibody against vimentin (Biogenex, San Ramon) was used according to the indications of the supplier. The monoclonal antibody against PCNA (Ab-1; Oncogene Science, New York) was used at a dilution of 1:400. Finally, several sections at every age were processed for Hu immunocytochemistry. For this later procedure, a biotinylated IgG containing the Hu antibody was obtained from the serum of a patient with small cell lung cancer and paraneoplastic encephalomyelitis, and prepared as detailed else- where [15]. After blocking endogenous peroxidase with 30% methanol, 69% PBS and 1% hydrogen peroxide for 20 min, the sections were treated with saponine and incubated with normal goat serum for 60 min, biotinylated anti-Hu IgG diluted at 1:1000 in PBS overnight at 4C, and ABC complex for 30 min. The peroxidase reaction was visualized with 0.05% diaminobenzidine and 0.01% hydrogen peroxide. To rule out false-positive reactions, a few sections were incubated without the primary antibody, or with normal human serum. All sections were counterstained with hematoxylin.
Quantitative studies were carried out in the cortex and subcortical white matter in one visual field of the vermian region of
the culmen by counting the number of cells directly through the ocular of the microscope at a magnification of x 400. The following counts were made: dying cells and PCNA-immunoreactive cells in the external and internal granular layers (EGL, IGL) and subcor- tical white matter, and mitoses in the EGL. The results were arbitrarily expressed as the number of cells per 4.5 • 104 ~*m 2 (which corresponded to the area of the grid adapted to the ocular of the microscope at the magnification x 400). Statistical processing was carried out with the Mann-Whitney U test.
Results
General findings
D yi ng cells were recognized by the i r ex t remely dark and sh r unke n , o f ten f r a gme n t e d , nucleus . D y ing cells appeared 3 h af ter 2 Gy X-ray exposure ma in ly in the uppe r levels of the E G L . D y i ng cells also appea red in the whi te mat ter , and a few in the bo rde r b e t w e e n the I G L and the Purk in j e cell layer (Fig. 1A, B). Large n u m b e r s

494
of dying cells were found 6 h after irradiation in rats aged 1 and 3 days. Dying cells were observed in the upper and inner levels of the external granular layer (Fig. 1C) and in the white matter. A few dying cells were seen in the IGL. The number of dying cells was largely reduced 24 h after irradiation (Fig. 1E). Dying cells were rare 48 h after 2 Gy X-ray exposure (Fig. 1F). Mitoses were usually encountered in the EGL throughout develop- ment. However, the number of mitotic cells was reduced during the 1st h after X-ray exposure. Some mitotic cells reappeared 48 h later; normal values of mitoses were reached 5 days after irradiation (Fig. 1F).
The number of dying cells was dramatically reduced in irradiated animals treated with 1 btg/g body weight of cycloheximide at the time of irradiation and killed 6 h later (Fig. 1D).
Identification of dying cells
In the normal developing cerebellum the Hu antibody recognized immature cells in the inner level of the EGL, and cells of the Purkinje and IGL as well. Immature cells in the upper level of the EGL remained unlabelled by the serum containing the anti-Hu antibody (Fig. 2A). At 3 h after irradiation, Hu-immunoreactive cells in the inner level were preserved (Fig. 2B), but 6 h after irradiation apoptotic cells extended to the inner, Hu- immunoreactive, level of the external cellular layer (Fig. 2C). At 5 days after irradiation Hu immunocyto- chemistry did not reveal differences between control and irradiated rats (Fig. 2D).
Vimentin-immunoreactive cells were Bergmann glia located at the border between the Purkinje cell layer and the IGL, and glial cells in the white matter (Fig. 3). Degenerating Bergmann glial cells, which were recog- nized by their globulose immunoreactive soma and shortened and fragmented apical processes, were not observed at 3 h (Fig. 3B), but were noted 6 h after irradiation (Fig. 3C). Degenerating Bergmann glial cells were no longer seen 48 h after irradiation (Fig. 3D). A few vimentin-immunoreactive apoptotic cells were observed in the cerebellar white matter 6 h after X-ray exposure (data not shown).
Quantitative studies
The number of dying cells, mitoses and PCNA-immu- noreactive cells in the cerebellar cortex and white matter in control rats and irradiated animals at P1 and P3, killed at different intervals post-irradiation, are shown in Table 1.
A significant increase in the number of dying cells in the EGL was seen from 3 to 48 h after irradiation in rats irradiated at P1, and from 3 to 24 h in rats irradiated at P3. Significantly increased numbers of dying cells were also observed in the IGL and cerebellar white matter after irradiation in both groups of rats, although the effects were greater in rats irradiated at P1 than in P3-irradiated rats. The injection of cycloheximide at a dose 1 btg/g body wt. dramatically reduced the number of dying cells 6 h after irradiation (Table 1).
As seen in Table 1, a significant decrease in the number of mitoses in EGL occurred from 30 min to 48 h (in rats irradiated at P1) or 24 h (in rats irradiated at P3). Later on, the number of mitoses was similar in irradiated animals and controls.
The number of PCNA-immunoreactive cells in rats irradiated either at P1 or P3 was significantly reduced in EGL until 48 h after irradiation, but significant reduc- tions in the number of PCNA-immunoreactive cells in IGL and white matter were only observed in rats irradiated at P1.
Discussion
The present results support previous observations [20, 26] proposing that X-ray-induced cell death in the developing cerebellum resembles apoptosis and de- pends on protein synthesis, as indirectly suggested by cycloheximide studies. Similar findings were observed in the irradiated neocortex and hippocampus during devel- opment [13, 14]. Taken together these data point to the likelihood that X-ray-induced cell death in the develop- ing brain is probably related to the activation of death genes or to the inhibition of survival proteins [14]. Since recent reports in the developing cerebral cortex have shown a relationship between naturally occurring cell death and transient c-los expression [16], studies are
PCNA-immunoreactive cells after irradiation
In the normal developing cerebellum, the anti-PCNA antibody recognized large numbers of cells in the EGL (mainly in the upper, but also in the inner level), and small numbers in the IGL, the border between this layer and the Purkinje cell layer, and white matter (Fig. 4A). Although a few fragmented PCNA-immunoreactive nuclei were seen 3 h after irradiation (Fig. 4B), immu- noreactive fragmented nuclei and globules were mainly found in the EGL 6 h after irradiation (Fig. 4C). The number of PCNA-immunoreactive cells was largely decreased 24 and 48 h after irradiation (Fig. 4D).
Fig. 2A--D. Hu-immunoreactive cells in the cerebellum of rats after irradiation at the age of 1 day. A In the normal (non- irradiated) cerebellum, immunoreactive cells are in the inner level (b) of the external granular layer (EGL), Purkinje cell layer (P) and internal granular layer (IGL). Hu-immunoreaetive ceils are not found in the upper level (a) of EGL. B At 3 h after irradiation, dying cells are observed in the upper (a), but not in the inner level (b) of EGL. C Dying cells are found throughout the EGL (levels a and b) 6 h after irradiation. D A normal pattern of Hu immuno- reactivity is found 5 days after X-ray exposure. Hu-immunoreac- tive cells are found in the inner (b), but not in the upper level (a) of the external granular layer. Dying cells are absent but a few mitoses can be observed (arrows). A--D Hematoxylin counterstaining, • 4O0
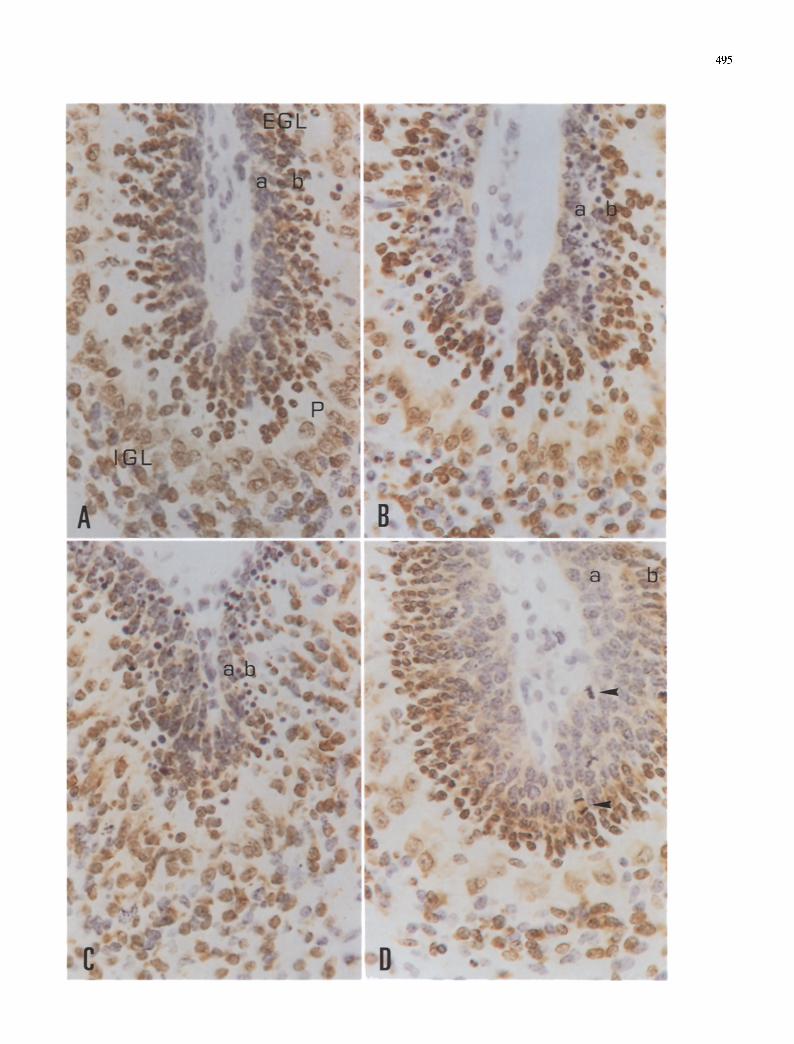
495

496
Fig. 3A--D. Vimentin-immunoreactive cells in the cerebellum of rats irradiated at the age of 3 days. A Normal (non-irradiated) cerebellum showing immunoreactive Bergmann glia with the cellular bodies located at the border of the internal granular layer (IGL) and the Purkinje cell layer (P), and the apical processes reaching the external granular layer (EGL). B Dying cells are
found in the external granular layer (asterisk), but Bergmann glia appear normal 3 h after irradiation. C Degenerating Bergmann glial cells (arrows) are observed 6 h after irradiation. D Normal appearance of Bergmann glial cells 48 h after X-rays exposure. A- -D Hematoxylin counterstaining, • 400
497
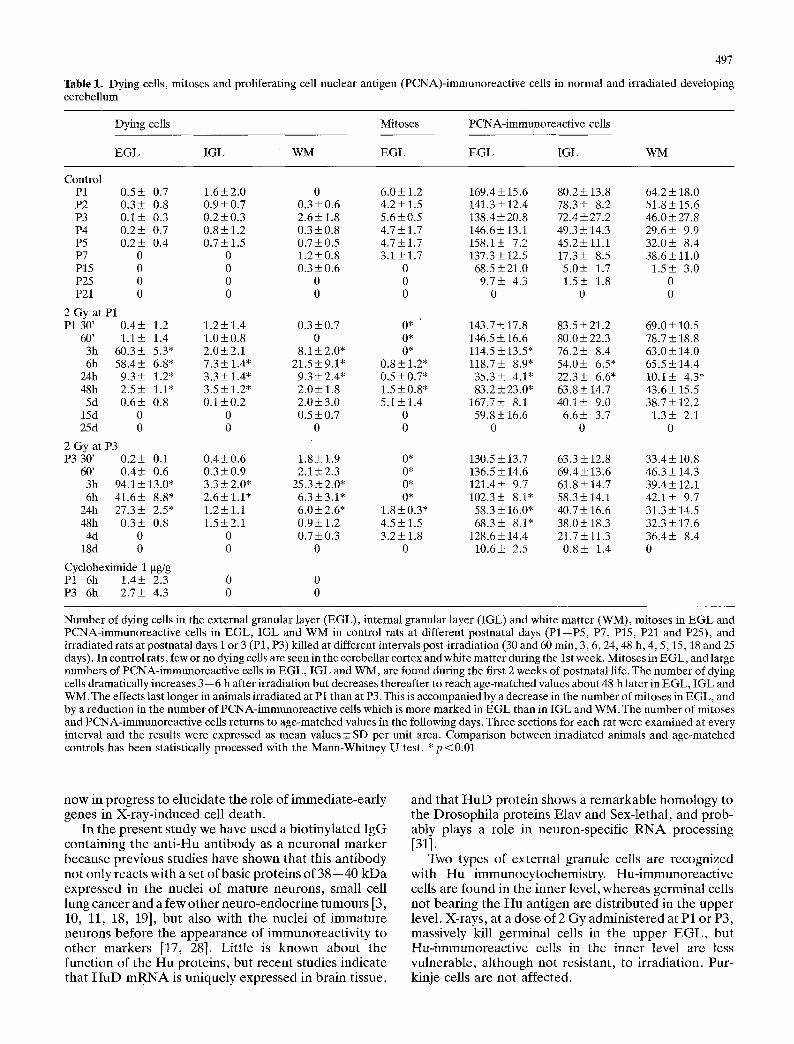
]Fable 1. Dying cells, mitoses and proliferating cell nuclear antigen (PCNA)-immunoreactive cells in normal and irradiated developing cerebellum
Dying cells Mitoses PCNA-immunoreactive cells
EGL IGL WM EGL EGL IGL WM
Control P1 0.5+_ 0.7 1.6+-2.0 0 6.0+-1.2 169.4+15.6 80.2+-13.8 64.2+-18.0 P2 0.3+ 0.8 0.9+-0.7 0.3+-0.6 4.2+-1.5 141.3+-12.4 78.3+- 8.2 51.8+-15.6 P3 0.1+ 0.3 0.2+-0.3 2.6+-1.8 5.6+-0.5 138.4+-20.8 72.4+-27.2 46.0+_ 27.8 P4 0.2+ 0.7 0.8+1.2 0.3+-0.8 4.7+-1.7 146.6+13.1 49.3+14.3 29.6+- 9.9 P5 0.2+- 0.4 0.7+-1.5 0.7+-0.5 4.7+-1.7 158.1+ 7.2 45.2_+11.1 32.0+- 8.4 P7 0 0 1.2+-0.8 3.1_+1.7 137.3+-12.5 17.3+- 8.5 38.6+11.0 P15 0 0 0.3+0.6 0 68.5+-21.0 5.0+- 1.7 1.5+ 3.0 P25 0 0 0 0 9.7_+ 4.3 1.5+ 1.8 0 P21 0 0 0 0 0 0 0
2 Gy at P1 P1 30' 0.4+ 1.2 1.2+1.4 0.3+-0.7 0* 143.7+-17.8 83.5+-21.2 69.0+-10.5
60' 1.1+ 1.4 1.0+0.8 0 0* 146.5+-16.6 80.0+-22.3 78.7+18.8 3h 60.3+ 5.3* 2.0+-2.1 8.1+-2.0" 0* 114.5+-13.5" 76.2+- 8.4 63.0+-14.0 6h 58.4+- 6.8* 7.3+-1.4" 21.5+-9.1" 0.8+-1.2" 118.7+- 8.9* 54.0+- 6.5* 65.5+14.4
24h 9.3+- 1.2" 3.3+-1.4" 9.3+-2.4* 0.5+-0.7* 35.3+ 4.1" 22.3+ 6.6* 10.1+- 4.3* 48h 2,5+ 1.1" 3.5+-1.2" 2.0+-1.8 1.5+0.8" 83.2_+23.0* 63.8+14.7 43.6+-15.5 5d 0.6+ 0.8 0.1+0.2 2.0+-3.0 5.1_+1.4 167.7_+ 8.1 40.1+- 9.0 38.7+-12.2
15d 0 0 0.5+0.7 0 59.8+16.6 6.6+- 3.7 1.3+- 2.1 25d 0 0 0 0 0 0 0
2 Gy at P3 P3 30' 0.2_+ 0.1 0.4_+0.6 1.8+1.9 0* 130.5+-13.7 63.3+-12.8 33.4+-10.8
60' 0.4+ 0.6 0.3+0.9 2.1+2.3 0* 136.5+14.6 69.4+-13.6 46.3+-14.3 3h 94.1_+13.0" 3.3_+2.0* 25.3+2.0* 0* 121.4+- 9.7 61.8+-14.7 39.4+12.1 6h 41.6+ 8,8* 2.6+-1.1" 6.3+-3.1" 0* 102.3+- 8.1" 58.3+14.1 42.1+ 9.7
24h 27.3+ 2.5* 1.2+1.1 6.0+2.6* 1.8+0.3" 58.3+16.0" 40.7+-16.6 31.3+14.5 48h 0.3+ 0.8 1.5+2.1 0.9+1.2 4.5+-1.5 68.3+- 8.1" 38.0+_18.3 32.3+-17.6 4d 0 0 0.7+-0.3 3.2+1.8 128.6_+14.4 21.7+-11.3 36.4+ 8.4
18d 0 0 0 0 10.6_+ 2.5 0.8+- 1.4 0
Cycloheximide 1 btg/g P1 6h 1.4+- 2.3 0 0 P3 6h 2.7_+ 4.3 0 0
Number of dying cells in the external granular layer (EGL), internal granular layer (IGL) and white matter (WM), mitoses in EGL and PCNA-immunoreactive cells in EGL, IGL and WM in control rats at different postnatal days (P1--P5, P7, P15, P21 and P25), and irradiated rats at postnatal days i or 3 (P1, P3) killed at different intervals post-irradiation (30 and 60 rain, 3, 6, 24, 48 h, 4, 5, 15, 18 and 25 days). In control rats, few or no dying cells are seen in the cerebellar cortex and white matter during the 1st week. Mitoses in EGL, and large numbers of PCNA-immunoreactive cells in EGL, IGL and WM, are found during the first 2 weeks of postnatal life. The number of dying cells dramatically increases 3--6 h after irradiation but decreases thereafter to reach age-matched values about 48 h later in EGL, IGL and WM. The effects last longer in animals irradiated at P1 than at P3. This is accompanied by a decrease in the number of mitoses in EGL, and by a reduction in the number of PCNA-immunoreactive cells which is more marked in EGL than in IGL and WM. The number of mitoses and PCNA-immunoreactive cells returns to age-matched values in the following days. Three sections for each rat were examined at every interval and the results were expressed as mean values + SD per unit area. Comparison between irradiated animals and age-matched controls has been statistically processed with the Mann-Whitney U test. *p <0.01
now in progress to e luc ida te the role of immed ia t e - ea r ly genes in X- ray- induced cell dea th .
I n the p resen t s tudy we have used a b io t i ny l a t ed I g G con ta in ing the a n t i - H u a n t i b o d y as a n e u r o n a l m a r k e r because previous s tudies have shown that this an t i body no t on ly reacts wi th a set of basic p ro te ins of 38- - 40 k D a expressed in the nucle i of m a t u r e n e u r o n s , small cell lung cancer and a few o the r n e u r o - e n d o c r i n e t u m o u r s [3, 10, 11, 18, 19], bu t also wi th the nucle i of i m m a t u r e n e u r o n s before the appea rance of i m m u n o r e a c t i v i t y to o the r markers [17, 28]. Li t t le is k n o w n abou t the func t ion of the H u pro te ins , bu t r ecen t s tudies indica te tha t H u D m R N A is u n i q u e l y expressed in b ra in t issue,
and that H u D p ro t e in shows a r e m a r k a b l e homology to the Drosoph i l a pro te ins Elav and Sex-lethal , and prob- ably plays a role in neuron-spec i f ic R N A processing [311.
Two types of ex te rna l g ranu le cells are recognized with H u immunocy tochemis t ry . H u - i m m u n o r e a c t i v e cells are f ound in the i n n e r level , whereas ge rmina l cells no t bea r ing the H u an t igen are d i s t r ibu ted in the uppe r level. X-rays, at a dose of 2 Gy admin i s t e red at P1 or P3, massively kill ge rmina l cells in the uppe r E G L , bu t H u - i m m u n o r e a c t i v e cells in the i n n e r level are less vu lne rab le , a l though no t res is tant , to i r rad ia t ion . Pur- kinje cells are no t affected.
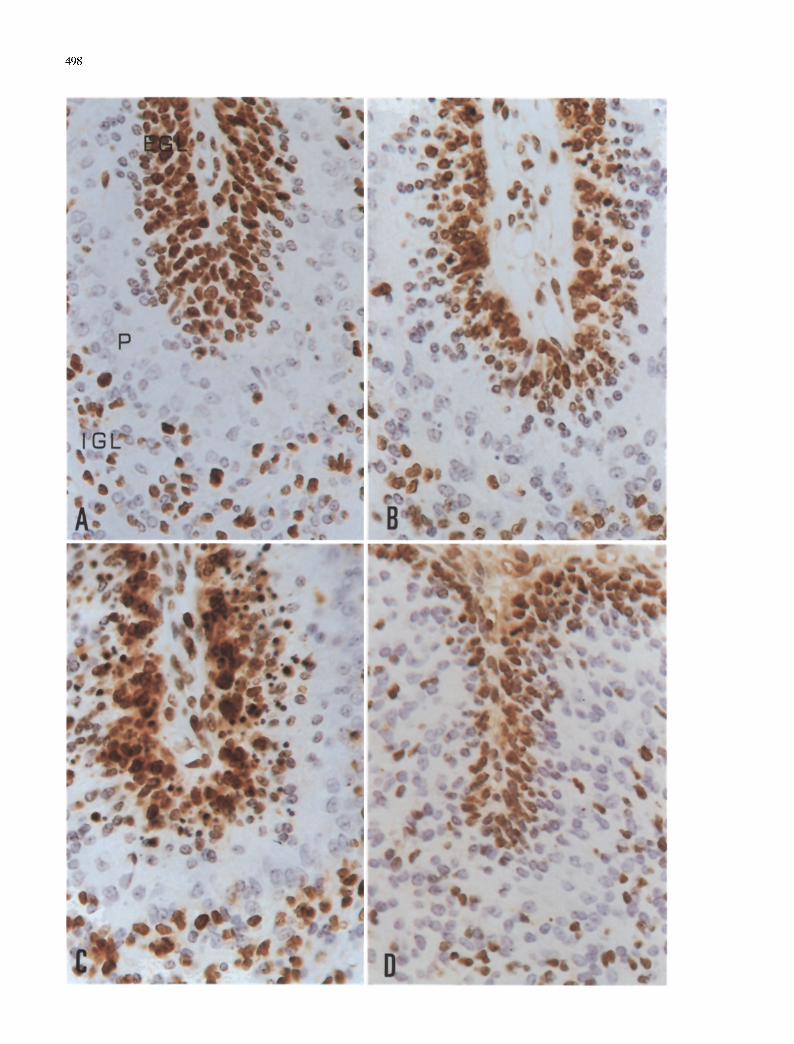
498

Vimentin-immunoreactive cells at P1 and P3 are Bergmann glia and scattered astrocytes in the subcorti- cal cerebellar white matter. These cells are vulnerable to X-rays, as dying cells of both types are found after irradiation. Therefore, the present immunocytochemi- cal findings indicate that radiosensitive populations in the developing cerebellum are germinal cells in the upper level of the EGL, immature neurons in the inner level of the external germinal layer, Bergmann glia, astrocytes in the white matter, and non-typified imma- ture cells (probably glial cell precursors) in the white matter.
Early studies, based on tritiated thymidine autora- diography, suggested that radiosensitive cells during the development of the forebrain and retina fall into two classes: (1) prophase cells just about to divide, and (2) early postmitotic primitive cells that are beginning to differentiate [23]. In the present study we have focused on the number of mitoses and PCNA-immunoreact ive cells to learn about radiosensitive cells in the developing cerebellum.
Mitotic cells are mainly found in the EGL, but a few mitoses are also encountered in the IGL. The number of mitoses decreases as early as 30 min after irradiation and does not begin to increase until 24 h later, thus indicating that X-rays block the M phase of the cell cycle.
PCNA/cyclin is a nuclear polypeptide with a molecu- lar mass of about 36 kDa which is expressed in prolifer- ating cells in the late G1 and S phase of the cell cycle, and it plays a role as an auxiliary protein of D N A polymerase 6 in D N A synthesis [4--9, 27, 29, 30, 32, 33]. In the developing cerebellar cortex, PCNA-immu- noreactive cells are mainly observed in the E G L (par- ticularly in the upper level), but also in the IGL and subcortical white matter. Dying PCNA-immunoreact ive cells are found 6 h after irradiation, and the number of PCNA-immunoreact ive cells is largely reduced 24 and 48 h later in the E G L and IGL, and, to a lesser extent, in the white matter. This suggests that most cells in the late G1 and S phases of the cell cycle are radiosensitive populations.
Reconsti tution of the cerebellum occurs not earlier than 48 h after irradiation. Five days after X-ray expo- sure the number of PCNA-immunoreact ive cells in the E G L and IGL reach age-matched values. The number of PCNA-immunoreact ive cells in the white matter, which is only decreased in animals irradiated at P1, is normal-
<
Fig. 4A--D. Proliferating on nuclear antigen (PCNA)-immuno- reactive cells in the cerebellum of the rat irradiated at the age of 3 days. A In the normal (non-irradiated) cerebellum, PCNA- immuno-reactive cells are found in the upper level and most of the inner level of the external granular layer (EGL). Immunoreactive cells are seen scattered in the internal granular layer (IGL) and at the border of the Purkinje cell layer (P). B At 3 h after irradiation, a few fragmented PCNA-immunoreactive nuclei are found in EGL. C At 6 h after irradiation, large numbers of immunoreactive fragmented nuclei are observed throughout the external granular layer. D Reduced numbers of PCNA-immunoreactive cells are seen 24 h after irradiation. A--D Hematoxylin counterstaining, 40O
499
ized at the same time. Cerebella irradiated at P1 or P3 do not differ f rom controls at the age of 1 month. There- fore, the present findings indicate that reconstitution of the irradiated cerebellum depends not only on the proliferation of surviving cells in the EGL, but on the division of ceils in the IGL and white matter as well.
Acknowledgements. We wish to thank C. Cin6s from the Radio- protection Department for her help in dosimetry and irradiation, and T. Yohannan for editorial assistance.
References
1. Altman J, Anderson WJ (1972) Experimental reorganization of the cerebellar cortex. I. Morphological effects of elimina- tion of all microneurons with prolonged x-irradiation started at birth. J Comp Neurol 146:355--406
2. Altman J, Anderson WJ, Wright KA (1969) Reconstitution of the external granular layer of the cerebellar cortex in infant rats after low-level X-irradiation. Anat Rec 163:453--472
3. Anderson NE, Roseblum MK, Graus F, Wiley RG, Posner JB (1988) Autoantibodies in paraneoplastic syndromes associated with small-cell lung cancer. Neurology 38:1391--1398
4. Bravo R (1986) Sylrthesis of the nuclear protein cyclin (PCNA) and its relationship with DNA replication. Exp Cell Res 163:287--293
5. Bravo R, Cells JE (1980) A search for differential polypeptide synthesis throughout the cell cycle of HeLa cells. J Cell Biol 84:795-- 802
6. Bravo R, Macdonald-Bravo H (1984) Induction of the nuclear protein cyclin in quiescent mouse 3T3 cells stimulated by serum and growth factors: correlation with DNA synthesis. EMBO J 3:3177--3181
7. Bravo R, Macdonald-Bravo H (1987) Existence of two popu- lations of cyclin/proliferating cell nuclear antigen during the cell cycle: association with DNA replication sites. J Cell Biol 105:1549-- 1554
8. Bravo R, Frank R, Blundell PA, Macdonald-Bravo H (1987) Cyclin/PCNA is the auxiliary protein of DNA polymerase 6. Nature 326:515--517
9. Celis JE, Madsen E Celis A, Nielsen HV, Gesser B (1987) Cyclin (PCNA, auxiliary protein of DNA polymerase 6) is a central component of the pathway(s) leading to DNA replica- tion and cell division. FEBS Lett 220:1--7
10. Dalmau J, Furneaux HM, Gralla RJ, Kris MG, Posner JB (1990) Detection of the anti-Hu antibody in the serum of patients with small cell lung cancer: a quantitative Western blot analysis. Ann Neurol 27:544--552
11. Dalmau J, Graus E Rosenblum MK, Posner JB (1992) Anti-Hu-associated paraneoplastic encephalomyelitis/sensory neuronopathy. A clinical study of 71 patients. Medicine 71:59--72
12. Das GD (1977) Experimental analysis of embryogenesis of cerebellum in rat. II. Morphogenetic malformations following X-ray irradiation on day 18 of gestation. J Comp Neurol 176:435--452
13. Ferrer I (1992) The effect of cycloheximide on natural and X-ray-induced cell death in the developing cerebral cortex. Brain Res 588:351.--357
14. Ferrer I, Serrano T, Alcfintara S, Tortosa A, Graus F (1993) X-ray-induced cell death in the developing hippocampal cortex involves neurons and requires protein synthesis. J Neuropathot Exp Neurol 52:370-378
15. Furneaux HM, Rosenblum MK, Dalmau J, Wong E,Woodruff P, Graus F, Posner JB (1990) Selective expression of Purkinje cell antigens in tumor tissue from patients with paraneoplastic cerebellar degeneration. N Engl J Med 322:1844--1851

500
16. Gonzalez-Martin C, de Diego I, Crespo D, Fair6n A (1992) Transient c-fos expression accompanies naturally occurring cell death in the developing interhemispheric cortex of the rat. Dev Brain Res 68:83--95
17. Graus F, Ferrer I (1990) Analysis of a neuronal antigen (Hu) expression in the developing rat brain detected by autoanti- bodies from patients with paraneoplastic encephalomyelitis. Neurosci Lett 112:14--18
18. Graus E Cordon-Cardo C, Posner J (1985) Neuronal anti- nuclear antibody in sensory neuronopathy from lung cancer. Neurology 35:538--543
19. Graus E Elkon KB, Cordon-Cardo C, Posner JB (1986) Sensory neuronopathy and small cell lung cancer. Anti- neuronal antibody that also reacts with the tumor. Am J Med 80:45--52
20. Harmon BV, Allan DJ (1988) X-ray-induced cell death by apoptosis in the immature rat cerebellum. Scanning Microsc 2:561 --568
21. Hicks SP (1958) Radiation as an experimental tool in mam- malian developmental neurology. Physiol Rev 38:337--356
22. Hicks SP, D'Amato CJ (1966) Effects of ionizing radiation on mammalian development. In: Woollam DHM (ed) Advances in teratology. Logos Press, London, pp 196--250
23. Hicks SP, D'Amato CJ, Coy MA, O'Brien ED, Thruston JM, Foftes DL (1961) Migrating cells in the developing nervous system studied by their radiosensitivity and tritiated thymidine uptake. In: Fundamental aspects of Radiosensitivity; Brook- haven Symposium Biology no. 14. Upton, New York, pp 246--261
24. Inouye M (1979) Cerebellar malformations in prenatally X-irradiated rats: quantitative analysis and detailed descrip- tion. Teratology 20:353--364
25. Inouye M, Kameyama Y (1983) Cell death in the developing rat cerebellum following X-irradiation of 3 to 100 rad: a quantitative study. J Radiat Res (Tokyo) 24:259--269
26. Inouye M, Tamaru M, Kameyama Y (1992) Effects of cyclo- heximide and actinomycin D on radiation-induced apoptotic cell death in the developing mouse cerebellum. Int J Rad Biol 61:669--674
27. Kurki P, Ogata K, Tan EM (1988) Monoclonal antibodies to proliferating cell nuclear antigen (PCNA)/cyclin as probes for proliferating cells by immunofluorescence microscopy and flow cytometry. J Immunol Methods 109:49--59
28. Marusich MF, Weston JA (1992) Identification of early neu- rogenic cells in the neural cell lineage. Dev Biol 149:295--306
29. Matthews MB, Bernstein RM, Franza BR, Garrels JI (1984) Identity of the proliferating cell nuclear antigen and cyclin. Nature 309:374--376
30. Ogata K, Kurki P, Celis JE, Nakamura RM, Tan EM (1987) Monoclonal antibodies to a nuclear protein (PCNA/cyclin) associated with DNA replication. Exp Cell Res 168:475--486
31. Szabo A, Dalmau J, Manley G, Rosenfeld M,Wong E, Henson J, Posner JB, Furneaux HM (1991) HuD, a paraneoplastic encephalomyelitis antigen, contains RNA-binding domains and is homologous to Elav and Sex-lethal. Cell 67:325--333
32. Takasaki J, Deu JS, Tan EM (1981) A nuclear antigen associated with cell proliferation and blast transformation. Its distribution in synchronized cells. J Exp Med 156:1899-- 1909
33. Takasaki Y, Fishwild D, Tan EM (1984) Characterization of proliferating cell nuclear antigen recognized by autoantibodies in lupus sera. J Exp Med 159:981--992




















