
The roles of the cerebellum and basal ganglia in timing and error prediction
11
The roles of the cerebellum and basal ganglia in timing and error prediction Jean-Claude Dreher and Jordan Grafman Cognitive Neuroscience Section, National Institute of Neurological Disorder and Stroke, Building 10, Room 5C205, MSC 1440 Bethesda, Maryland 20892–1440, USA Keywords: attention, fMRI, predictability, striatum, switching Abstract Recent evidence that the cerebellum and the basal ganglia are activated during the performance of cognitive and attention tasks challenges the prevailing view of their primary function in motor control. The specific roles of the basal ganglia and the cerebellum in cognition, however, have been difficult to identify. At least three functional hypotheses regarding their roles have been proposed. The first hypothesis suggests that their main function is to switch attentional set. The second hypothesis states that they provide error signals regarding stimuli or rewards. The third hypothesis is that they operate as an internal timing system, providing a precise representation of temporal information. Using functional magnetic resonance imaging, we tested these three hypotheses using a task-switching experiment with a 2 3 2 factorial design varying timing (random relative to fixed) and task order (unpredictable relative to predictable). This design allowed us to test whether switching between tasks, timing irregularity and/or task order unpredictability activate the basal ganglia and/or the cerebellum. We show that the cerebellum is primarily activated with timing irregularity while the anterior striatum is activated with task order unpredictability, supporting their distinctive roles in two forms of readjustment. Task order unpredictability alone, independent of reward delivery, is sufficient to induce striatal activation. In addition, activation of the cerebellum and basal ganglia were not specific to switching attention because these regions were both activated during switching between tasks and during the simultaneous maintenance of two tasks without switching between them. Introduction The basal ganglia and the cerebellum are known to be involved in motor control because of the motor deficits resulting from their dysfunction. This motor function has been supplemented in recent years by numerous studies showing that the basal ganglia and the cerebellum are also involved in a wide range of nonmotor, cognitive tasks (Leiner et al., 1991; Ivry, 1996; Middleton & Strick, 2000). Although several studies have contrasted the roles of the cerebellum and of the basal ganglia in motor control and learning (Jueptner & Weiller, 1998; Doya, 2000), it has been difficult to distinguish their specific roles in cognitive performance. Three main hypotheses have been offered. The first hypothesis is that the cerebellum and basal ganglia mediate switching attentional set (Akshoomoff & Courchesne, 1992; Owen et al., 1993; Hayes et al., 1998; Le et al., 1998). Indeed, both cerebellar lesions patients (Courchesne & Allen, 1997) and parkinsonian patients showed impaired performance in switching attention set or switching between tasks (Gotham et al., 1988; Eslinger & Grattan, 1993; Owen et al., 1993; Hayes et al., 1998; Cools et al., 2001b). FMRI studies also support the view that the lateral cerebellar hemisphere is activated in shifting attention (Le et al., 1998), but no specific basal ganglia activation was found for switch as compared to repeat trials during task switching (Dove et al., 2000; Sohn et al., 2000). The second hypothesis is that the cerebellum and the basal ganglia code prediction errors (Fiez et al., 1992; Kawato & Gomi, 1992; Schultz & Dickinson, 2000; Ivry & Fiez, 2001). Evidence that the cerebellum codes prediction errors was first proposed for the motor domain, based on the fact that the cerebellum modifies its output with unexpected sensory input (Marr, 1969; Albus, 1971; Kawato & Gomi, 1992). This error-correction function has been extended to the cognitive domain, the cerebellum providing a more general error- correction role, in modifying internal thought (Bracke-Tolkmitt et al., 1989; Canavan et al., 1994; Courchesne & Allen, 1997; Hikosaka et al., 1999; Tamada et al., 1999; Doya, 2000; Imamizu et al., 2000; Blakemore et al., 2001; Ivry & Fiez, 2001). Evidence that the basal ganglia code an error prediction is mainly based on the fact that monkey’s striatum codes a prediction error of reward delivery (Schultz, 2000; Schultz & Dickinson, 2000). A recent fMRI study in humans confirms this hypothesis, the ventral striatum being activated by reward unpredictability (Berns et al., 2001). However, it is unclear whether striatal activity necessarily requires reward delivery or whether the crucial factor is unpredictability itself. The third hypothesis is that the basal ganglia and the cerebellum operate as an internal timing system, providing the precise representation of temporal representation across various tasks. Cerebellar patients have difficulty with perceptual tasks requiring precise timing (Ivry, 1996, 1997) and Parkinson’s disease patients often underestimate time intervals (Pastor et al., 1992; Harrington et al., 1998). Direct evidence that the cerebellar neocortex is essential for timing the interval between stimulus and response in eyeblink conditioning has been provided by extensive neurophysiological and lesion studies in the rabbit. Correspondence: Jordan Grafman, PhD, as above. E-mail: [email protected] Received 3 May 2002, revised 28 July 2002, accepted 6 August 2002 doi:10.1046/j.1460-9568.2002.02212.x European Journal of Neuroscience, Vol. 16, pp. 1609–1619, 2002 ª Federation of European Neuroscience Societies
Transcript of The roles of the cerebellum and basal ganglia in timing and error prediction

The roles of the cerebellum and basal ganglia in timingand error prediction
Jean-Claude Dreher and Jordan GrafmanCognitive Neuroscience Section, National Institute of Neurological Disorder and Stroke, Building 10, Room 5C205,
MSC 1440 Bethesda, Maryland 20892±1440, USA
Keywords: attention, fMRI, predictability, striatum, switching
Abstract
Recent evidence that the cerebellum and the basal ganglia are activated during the performance of cognitive and attention tasks
challenges the prevailing view of their primary function in motor control. The speci®c roles of the basal ganglia and thecerebellum in cognition, however, have been dif®cult to identify. At least three functional hypotheses regarding their roles have
been proposed. The ®rst hypothesis suggests that their main function is to switch attentional set. The second hypothesis states
that they provide error signals regarding stimuli or rewards. The third hypothesis is that they operate as an internal timing system,providing a precise representation of temporal information. Using functional magnetic resonance imaging, we tested these three
hypotheses using a task-switching experiment with a 2 3 2 factorial design varying timing (random relative to ®xed) and task
order (unpredictable relative to predictable). This design allowed us to test whether switching between tasks, timing irregularity
and/or task order unpredictability activate the basal ganglia and/or the cerebellum. We show that the cerebellum is primarilyactivated with timing irregularity while the anterior striatum is activated with task order unpredictability, supporting their distinctive
roles in two forms of readjustment. Task order unpredictability alone, independent of reward delivery, is suf®cient to induce
striatal activation. In addition, activation of the cerebellum and basal ganglia were not speci®c to switching attention becausethese regions were both activated during switching between tasks and during the simultaneous maintenance of two tasks without
switching between them.
Introduction
The basal ganglia and the cerebellum are known to be involved in
motor control because of the motor de®cits resulting from their
dysfunction. This motor function has been supplemented in recent
years by numerous studies showing that the basal ganglia and the
cerebellum are also involved in a wide range of nonmotor, cognitive
tasks (Leiner et al., 1991; Ivry, 1996; Middleton & Strick, 2000).
Although several studies have contrasted the roles of the cerebellum
and of the basal ganglia in motor control and learning (Jueptner &
Weiller, 1998; Doya, 2000), it has been dif®cult to distinguish their
speci®c roles in cognitive performance. Three main hypotheses have
been offered.
The ®rst hypothesis is that the cerebellum and basal ganglia
mediate switching attentional set (Akshoomoff & Courchesne, 1992;
Owen et al., 1993; Hayes et al., 1998; Le et al., 1998). Indeed, both
cerebellar lesions patients (Courchesne & Allen, 1997) and
parkinsonian patients showed impaired performance in switching
attention set or switching between tasks (Gotham et al., 1988;
Eslinger & Grattan, 1993; Owen et al., 1993; Hayes et al., 1998;
Cools et al., 2001b). FMRI studies also support the view that the
lateral cerebellar hemisphere is activated in shifting attention (Le
et al., 1998), but no speci®c basal ganglia activation was found for
switch as compared to repeat trials during task switching (Dove et al.,
2000; Sohn et al., 2000).
The second hypothesis is that the cerebellum and the basal ganglia
code prediction errors (Fiez et al., 1992; Kawato & Gomi, 1992;
Schultz & Dickinson, 2000; Ivry & Fiez, 2001). Evidence that the
cerebellum codes prediction errors was ®rst proposed for the motor
domain, based on the fact that the cerebellum modi®es its output with
unexpected sensory input (Marr, 1969; Albus, 1971; Kawato & Gomi,
1992). This error-correction function has been extended to the
cognitive domain, the cerebellum providing a more general error-
correction role, in modifying internal thought (Bracke-Tolkmitt et al.,
1989; Canavan et al., 1994; Courchesne & Allen, 1997; Hikosaka
et al., 1999; Tamada et al., 1999; Doya, 2000; Imamizu et al., 2000;
Blakemore et al., 2001; Ivry & Fiez, 2001). Evidence that the basal
ganglia code an error prediction is mainly based on the fact that
monkey's striatum codes a prediction error of reward delivery
(Schultz, 2000; Schultz & Dickinson, 2000). A recent fMRI study in
humans con®rms this hypothesis, the ventral striatum being activated
by reward unpredictability (Berns et al., 2001). However, it is unclear
whether striatal activity necessarily requires reward delivery or
whether the crucial factor is unpredictability itself.
The third hypothesis is that the basal ganglia and the cerebellum
operate as an internal timing system, providing the precise representation
of temporal representation across various tasks. Cerebellar patients have
dif®culty with perceptual tasks requiring precise timing (Ivry, 1996,
1997) and Parkinson's disease patients often underestimate time
intervals (Pastor et al., 1992; Harrington et al., 1998). Direct evidence
that the cerebellar neocortex is essential for timing the interval between
stimulus and response in eyeblink conditioning has been provided by
extensive neurophysiological and lesion studies in the rabbit.
Correspondence: Jordan Grafman, PhD, as above.E-mail: [email protected]
Received 3 May 2002, revised 28 July 2002, accepted 6 August 2002
doi:10.1046/j.1460-9568.2002.02212.x
European Journal of Neuroscience, Vol. 16, pp. 1609±1619, 2002 ã Federation of European Neuroscience Societies

Although cerebellar function is also associated with associative
learning and sensorimotor coordination (Marr, 1969; Albus, 1971;
Thach, 1998; Miall et al., 2001), we concentrate on testing the three
theories mentioned above because they concern both the basal
ganglia and the cerebellum. The goal of this study was thus to test
whether activation of the cerebellum and basal ganglia are speci®c to
switching attention between tasks, to error prediction or to timing
irregularity. To address these questions, we conducted a task-
switching fMRI experiment using a 2 3 2 factorial design varying
timing (®xed relative to random) and task order (predictable relative
to unpredictable).
Materials and methods
Subjects
Eight healthy subjects (mean age 25 years; range 20±31) with at least
a high school education were recruited following procedures
approved by the Institutional Review Board. All subjects were native
speakers of English and strongly right-handed, as measured by the
Edinburgh handedness inventory (mean score 90.5). None of the
subjects presented with an abnormal neurological history. One or two
days before the MR session, subjects participated in a behavioural
testing session during which they were trained to perform each of the
tasks and were required to attain an overall accuracy score > 90% to
participate in the fMRI experiment.
Stimuli and task conditions
Subjects responded to visually presented letters (vowels or conson-
ants, either in lower or upper case, in red or in green) by pressing
response buttons with their right or left hand (Fig. 1). There were
eight conditions, each cued by a distinct visual instruction, consisting
of two conditions used for baseline (Task A, vowel-consonant
discrimination; Task B, case discrimination), four tasks switching
conditions (obtained by crossing the task order and timing factors)
and two conditions used for control (Union task, A or B) (see full
description below). The tasks were administered in six scanning runs.
Each condition was included once in each run as a block of 24 trials
for a total of 192 trials/run. Each run was pseudo-randomly ordered
so that each condition appeared at different serial positions within a
run and two conditions never appeared twice in immediate succession
to prevent confounding order effects. Control tasks or tasks used for
baseline alternated with switch conditions.
Sustained attention tasks (Baseline)
In the `vowel±consonant' condition, subjects had to press the right
button if the letter was a vowel and the left if the letter was a
consonant. In the `case discrimination' condition, subjects had to
press the right button if the letter was in upper case and the left if the
letter was in lower case. In both of these conditions, the colour of the
letters was irrelevant. The baseline was composed of the mean of
these two discrimination tasks [(A + B)/2]. In both of these two
conditions, the colour of the stimuli (appearing every 2.5 s) alternated
every three letters.
Switching tasks
In the `switching' conditions, subjects had to take into account the
colour of the letters to know which task to perform. If the letter was
red, subjects had to perform the vowel±consonant condition (i.e. to
respond with the right button if the letter was a vowel and with the
left if it was a consonant). Conversely, if the letter was in green,
subjects had to perform the case discrimination task (i.e. to respond
with the right button if the letter was in upper case and with the left if
it was in lower case). There were four different switch conditions:
`®xed predictable', `random predictable', `®xed unpredictable',
`random unpredictable'). In the ®xed conditions the stimuli appeared
every 2.5 s whereas, in the random condition, the timing between two
stimuli was pseudo-randomized (2.5 s 6260, 6390, 6510 ms). In
the predictable condition, a switch between tasks occurred every three
stimuli and was pseudo-randomized in the unpredictable condition.
Across each condition, the number of red/green letters (12 of each),
of switches (11) and of left/right responses (12 of each) was
consistently maintained.
Control condition
The mean of the following two conditions was used to control for the
active maintenance of the two task sets without switching between
them. In both of these conditions, subjects had to press the right
button if the letter was a vowel or was in upper case (and if both were
true) and the left button otherwise (Union condition). In the ®rst
control condition stimuli appeared every 2.5 s while, in the second
control, the timing between two stimuli was pseudo-random every
2.5 s 6 260, 390 or 510 ms. In both of these control conditions, the
colour of the letter was irrelevant and changed pseudo-randomly.
Stimulus duration was 500 ms in all conditions.
This design allowed us to test the three hypotheses mentioned in
the introduction. If the cerebellum and/or the basal ganglia are
speci®c to switching attention between tasks, they should be more
activated by task switching than by maintaining two task sets without
switching between them. Furthermore, if the cerebellum and/or the
basal ganglia are speci®c to timing adjustment, they should be
activated by the main effect of timing irregularity (random > ®xed
timing conditions) because the uncertainty of the temporal relation-
ship between events is maximal in the random condition.
Behaviourally, a timing operation may be revealed either by slower
reaction times (RTs) in the random compared to the ®xed timing
conditions or by a reduction in RTs with an increasing time lag in the
random timing condition. The latter possibility is likely to occur if
subjects use the probabilistic information conveyed by the passage of
time to predict the likelihood of stimulus presentation (i.e. the more
time the subjects have, the more probable the stimulus occurrence). In
both cases, the main effect of timing irregularity (random > ®xed
timing) should reveal brain regions that attempt but fail to predict the
timing of stimulus presentation [at least for short interstimulus
intervals (ISIs)]. Conversely, the main effect of task order unpredict-
ability (unpredictable > predictable task order) should reveal brain
regions activated in error prediction, because more adjustment to the
current task is needed when task order is unpredictable than when it is
fully predictable. Indeed, subjects may constantly try to anticipate the
following task. Thus, when task order is unpredictable, discordance
between subject's expectation and the effective task to perform will
result in error correction. In contrast, when task order is predictable,
routine preparation for the whole sequence of tasks is possible and no
error signal needs to be emitted.
Apparatus
Stimuli were generated by a PC computer using the Expe6 software
package (ftp://ftp.lscp.ehess.fr/pub/expe6/) and projected onto a
screen at the subjects' feet. Subjects viewed the stimuli through a
mirror attached to the head coil. Subjects' behavioural responses were
recorded when they pressed one of two MRI-compatible response
buttons held in each hand.
1610 J.-C. Dreher and J. Grafman
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

Data analysis
Behavioural analysis
RT and percentage of errors were analysed with a MANOVA including
the two factors of Task Order (predictable vs. unpredictable) and
Timing (®xed vs. random).
Imaging data
High-resolution structural images were obtained using a standard 1.5
GE whole-body signa scanner with an RF head coil. For each subject,
six time series of 180 whole-brain images (four ®rst images removed)
were obtained with a gradient-echo, echo-planar scanning sequence
(TR 3 s, TE 40 ms, ¯ip angle 90°; FOV 24 cm, acquisition matrix
64 3 64, 22 axial slices, thickness 6 mm). Each run was pseudo-
randomly ordered according to a latin square design so that each
condition appeared only once at different serial positions within a run
and that baseline/control and switch conditions alternated. The order
of runs was also counterbalanced across subjects. Using SPM96 with
modi®ed memory-mapping procedures, for each subject, the series of
functional images for the six runs was realigned, normalized to the
Montreal Neurological Institute template, smoothed with a Gaussian
®lter (10-mm FWHM kernel), and ®nally scaled across scans. Then,
the data from all subjects were pooled and statistical parametric maps
were computed from local MR signals using linear multiple
regression with conditions, modelled as two temporal basis functions,
and runs as covariates (Friston et al., 1991). Only regions formed by
more than four adjacent active voxels (voxel size 4 mm3) were
reported (extend threshold P < 0.5). In accordance with our hypo-
theses, only cerebellar and subcortical regions are reported (Z > 3.09,
i.e. P < 0.001, uncorrected for multiple comparisons).
Mean values are quoted 6 SEM.
Results
Behavioural data
Behavioural results are depicted in Fig. 2. There was a signi®cant
response time cost (RT difference between switch and no switch
trials) in all task-switching conditions [F1,7 = 32.2; P < 0.001] (main
effect of Switch). Furthermore, performance was signi®cantly worse
when the order of the tasks was unpredictable than when it was
predictable (RT = 932.2 6 130.1 vs. 818 6 142.9 ms, F1,7 = 14.4,
P < 0.01; 4.7 6 3.1% of errors vs. 3.9 6 3.4%, F1,7 = 5.7, P < 0.05;
main effect of Task Order). Finally, no main effect of Timing was
found: ®xed timing, compared to random timing, did not signi®cantly
reduced the error rate (3.8 6 3.4% vs. 4.8 6 3.3%, F1,7 = 2.0,
P = 0.19) nor the RTs (865.6 6 138.1 vs. 884.6 6 123.8 ms,
F1,7 = 2.2, P = 0.18). No interaction reached statistical signi®cance.
The mean RTs of the individual tasks used for baseline and control
were: vowel/consonant, 717.3 6 100.1 ms; case discrimination,
656.5 6 89.8 ms; ®rst union, 778 6 115.2 ms; second union,
747.1 6 79.2 ms.
It is possible that the different time lags between stimuli in the
random timing conditions induce different types of behaviour (e.g.
subjects may be better prepared with long than with short ISIs) which
may not be observed by directly comparing the ®xed to the random
timing conditions. In order to better understand the contribution of the
various time lags in random timing, we thus performed an additional
three-factors repeated-measures ANOVA only for the two random-
timing switching conditions, which included task order (Predictable
vs. Unpredictable), time lag (®ve intervals of 408 ms from 1480 to
3520 ms) and switch (no switch vs. switch trials). As before, we
found a main effect of Switch (F1,7 = 15.3, P < 0.01) and a main
effect of Task Order (F1,7 = 9.5, P < 0.05). There was a trend toward
a reduction of RTs with time lag (F4,28 = 2.2, P = 0.09); this trend
became signi®cant (F4,56 = 3.2, P < 0.05) in another analysis adding
seven pilot subjects. No interactions were found between these
factors. Error rates did not show any signi®cant main effect or
interactions.
To ensure that the control condition effectively induced the
subjects to maintain two tasks in memory without switching between
them, we directly compared the control and the switching conditions.
RTs were signi®cantly slower in the task-switching conditions
averaged together (mean 875.0 6 129.9 ms) than in the control
condition (mean 763.2 6 96.1 ms; F1,7 = 34.8; P < 0.001), indicat-
ing that the switching conditions demanded additional cognitive
processes over those needed for the control task. Furthermore,
subjects did maintain two task-sets simultaneously in memory in the
control condition because, if subjects coded the control condition as a
single stimulus±response mapping, they should have directly associ-
ated the lower case consonants to a left motor response and other
letters to a right button press. However, RTs were not faster for lower
case consonants than for other letters (vowels and upper case
consonants) (F1,7 = 0.95; P = 0.4).
Imaging data
Tables 1 and 2 present results from all the brain regions activated in
the different conditions. In the following, we focus on subcortical and
cerebellar activation to test our hypotheses. First, we investigated
which cerebellar and subcortical regions were signi®cantly activated
in all switching conditions averaged together relative to baseline
(Z > 3.09, i.e. P < 0.001, uncorrected). Activation was found in the
cerebellar hemispheres (lobules VI, VIIIA, VIIIB, Crus I, in
Schmahmann's nomenclature; Schmahmann et al., 1999), the vermis
(lobules VII, VIII) (Fig. 3), the thalamus and the caudate nucleus
(Fig. 4). We plotted the activation of each task-switching condition
separately relative to baseline at the peak of activation of each of
these brain regions (Z > 3.09, i.e. P < 0.001, uncorrected) (Figs 3
and 4, bottom).
In order to control for the higher working memory load necessary
to keep both task sets active during the task-switching conditions, we
also examined the voxels commonly activated by each task-switching
condition relative to the control condition, which required subjects to
maintain two task-sets in memory without switching between them.
This was done by selecting the voxels activated in all task-switching
conditions averaged together relative to baseline (Z > 3.09,
P < 0.001, uncorrected for multiple comparisons) which were also
activated in each individual task-switching condition compared
separately to baseline (Z > 2.33, P < 0.01, uncorrected). None of
the brain regions mentioned above (caudate nucleus, thalamus and
cerebellum) were signi®cantly activated in this comparison (Z < 1.7,
P > 0.05, uncorrected). Furthermore, all these brain regions were
signi®cantly activated in the control condition relative to baseline
(Z > 3.09, i.e. P < 0.001, uncorrected), indicating that they were not
speci®cally activated during task switching (graph bars in Figs 3
and 4).
The main effect of task-order unpredictability (subtraction of the
two task-switching conditions with predictable order from the two
with unpredictable task order) activated the anterior putamen
bilaterally (Z > 3.09, i.e. P < 0.001, uncorrected) (Table 1, Fig. 5).
No signi®cant activation was found in the cerebellum.
The main effect of timing irregularity (subtraction of the two task-
switching conditions with ®xed timing from those with random
timing) was to activate the right posterior cerebellar hemispheres (VI/
Cerebellar and striatal functions in timing and error prediction 1611
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

VIIB/Crus I) (Schmahmann et al., 1999) and the dentate nucleus
(Z > 3.09, i.e. P < 0.001, uncorrected) (Table 1, Fig. 6). No signi®-
cant activation was found in the basal ganglia.
Brain regions showing interactions between the timing and task-
order factors (Z > 3.09, i.e. P < 0.001, uncorrected) activated the
right head of the caudate nucleus and the posterior putamen
bilaterally (Table 1, Fig. 7). No interactions were found in the
cerebellum.
Finally, we investigated whether the combination of task order and
timing unpredictability activated speci®c subcortical regions relative
to baseline (Z > 3.09, i.e. P < 0.001, uncorrected). We found that the
task-switching condition with unpredictable task order and random
timing speci®cally activated the substantia nigra (Fig. 8).
Functional connectivity analysis
In order to better understand what the basal ganglia/cerebellum do
with task order/timing information, we performed a functional
connectivity analysis on our data. This analysis allows us to point
out which brain regions correlate with the cerebellum and the basal
ganglia in each task-switching condition. For each subject, we
measured the mean blood oxygenation level-dependent (BOLD)
TABLE 1. Foci of activations in the different statistical contrasts
All task-switching conditionsaveraged together vs. baseline
Main effect ofirregular timing
Main effect ofunpredictable task order
Anatomical structure (Brodmann's area) x y z Z-value x y z Z-value x y z Z-value
CerebellumLeft cerebellar hemisphere ±28 ±68 ±40 6.72
±44 ±64 ±28 6.68Right cerebellar hemisphere 32 ±64 ±32 3.71 28
20±60±40
±32±44
4.965.56
Vermis (VIIB/VIIIB) 0 ±68 ±32 5.80Basal ganglia
Left Caudate nucleus ±16 ±16 24 5.15Right Caudate nucleus 16 4 20 5.13Left Putamen ±20 4 0 3.66Right Putamen 20
1612
168
±4
0±4±8
4.604.513.83
ThalamusLeft thalamus (vl) ±12 ±16 8 4.78Right thalamus (dm) 28 ±12 8 4.79
FrontalL sFG (BA 6) ±28 4 56 7.56L mFG (BA 8/9/44) ±40 20 32 7.18 ±44 16 32 5.79R mFG (BA 8/9/46) 48 16 40 7.77
60 28 32 6.47R iFG (BA 44/45) 36 24 20 6.21L Somato-motor area (BA 4) ±28 ±16 60 5.16R Somato-motor area (BA 4) 20 ±16 68 6.21Parietal
L IPS (BA 7/40) ±36 ±56 52 8.36 ±32 ±56 48 6.10R IPS (BA 7/40) 36 ±60 44 8.05 40 ±48 48 5.74
Temporal gyrusL iTG (BA 20) ±52 ±28 ±12 6.64R iTG (BA 37) 52 ±56 ±4 7.54 52 ±56 ±4 7.54
InsulaL INS ±20 24 ±4 5.59R INS 28 28 4 5.37L m occip. gyrus (BA 18/19) ±28 ±88 4 6.19R m occip. gyrus (BA 18/19) 28 ±84 ±8 5.38
All areas were signi®cant at P < 0.001 (uncorrected for multiple comparisons); x, y, and z are standardized stereotaxic coordinates of Talairach and Tournoux.Abbreviations: L, left; R, right. sFG, superior frontal gyrus; mFG, middle frontal gyrus; IPS, intra-parietal sulcus; INS, insula; iTG, inferior temporal gyrus; sTG,superior temporal gyrus; Somatosens. cx, somatosensory cortex; occip. gyrus, occipital gyrus; vl, ventro-lateral, dm, dorso-medial.
TABLE 2. Areas revealing interactions between task order and timing
Talairach coordinates
Anatomical structures x y z Z-value
Frontal cortexRight PM/Motor cx (BA 4/6) 52 ±8 28 6.5Left Motor cx (BA 4) ±44 ±12 32 6.1Right mFG (BA 9/46) 40 28 28 6.5Left mFG (BA 9/46) ±36 28 24 5.8Left fronto-polar cx (BA 10) ±28 48 8 5.2Pre-SMA ±4 12 56 4.9
Cingulate cortexAnterior cingulate (BA 24) 4 28 16 5.4
Basal gangliaRight Caudate nucleus 16 12 4 3.79Left Putamen ±24 ±8 ±4 4.08Right Putamen 20 ±4 4 3.68
Temporal gyrusRight mTG (BA 19/37) 40 ±72 4 5.7Left mTG (BA 37) ±52 ±36 ±8 4.8
BA, Brodmann's area; mFG, medial frontal gyrus; mTG, middle temporalgyrus; PM, pre-motor cortex; x, y, z, Talairach coordinates. All areas weresigni®cant at P < 0.001 (uncorrected for multiple comparisons).
1612 J.-C. Dreher and J. Grafman
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619
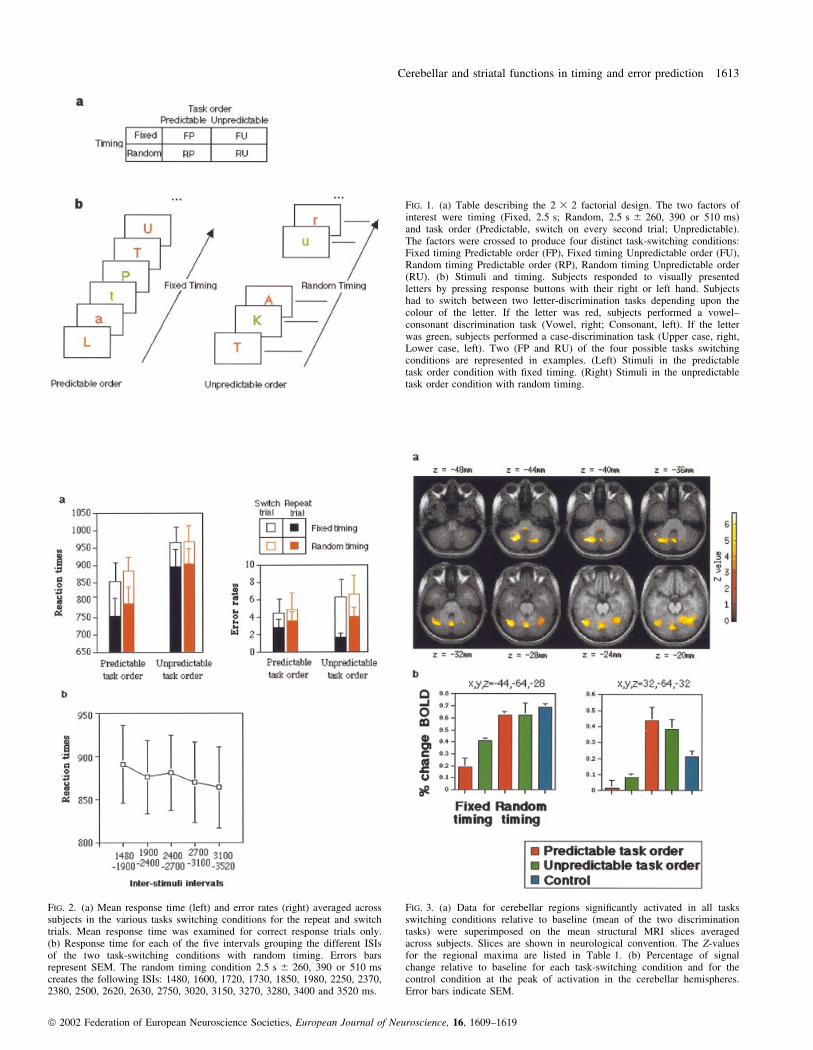
FIG. 1. (a) Table describing the 2 3 2 factorial design. The two factors ofinterest were timing (Fixed, 2.5 s; Random, 2.5 s 6 260, 390 or 510 ms)and task order (Predictable, switch on every second trial; Unpredictable).The factors were crossed to produce four distinct task-switching conditions:Fixed timing Predictable order (FP), Fixed timing Unpredictable order (FU),Random timing Predictable order (RP), Random timing Unpredictable order(RU). (b) Stimuli and timing. Subjects responded to visually presentedletters by pressing response buttons with their right or left hand. Subjectshad to switch between two letter-discrimination tasks depending upon thecolour of the letter. If the letter was red, subjects performed a vowel±consonant discrimination task (Vowel, right; Consonant, left). If the letterwas green, subjects performed a case-discrimination task (Upper case, right,Lower case, left). Two (FP and RU) of the four possible tasks switchingconditions are represented in examples. (Left) Stimuli in the predictabletask order condition with ®xed timing. (Right) Stimuli in the unpredictabletask order condition with random timing.
FIG. 2. (a) Mean response time (left) and error rates (right) averaged acrosssubjects in the various tasks switching conditions for the repeat and switchtrials. Mean response time was examined for correct response trials only.(b) Response time for each of the ®ve intervals grouping the different ISIsof the two task-switching conditions with random timing. Errors barsrepresent SEM. The random timing condition 2.5 s 6 260, 390 or 510 mscreates the following ISIs: 1480, 1600, 1720, 1730, 1850, 1980, 2250, 2370,2380, 2500, 2620, 2630, 2750, 3020, 3150, 3270, 3280, 3400 and 3520 ms.
FIG. 3. (a) Data for cerebellar regions signi®cantly activated in all tasksswitching conditions relative to baseline (mean of the two discriminationtasks) were superimposed on the mean structural MRI slices averagedacross subjects. Slices are shown in neurological convention. The Z-valuesfor the regional maxima are listed in Table 1. (b) Percentage of signalchange relative to baseline for each task-switching condition and for thecontrol condition at the peak of activation in the cerebellar hemispheres.Error bars indicate SEM.
Cerebellar and striatal functions in timing and error prediction 1613
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

signal in each task-switching condition relative to baseline at the peak
of activation of the following brain regions activated by all task-
switching conditions relative to baseline (Table 1): the dorsolateral
prefrontal cortex (DLPFC) (x,y,z = 48,16,40; x,y,z = ±28,4,56), the
intraparietal sulcus (IPS) region (x,y,z = ±36,56,52; x,y,z = 36,60,44),
the pre-supplementary motor area (pre-SMA) (x,y,z = ±4,16,52), the
caudate nucleus (x,y,z = ±16,16,24; x,y,z = 16,4,20) and the cerebel-
lum (x,y,z = ±28,68,40; x,y,z = 32,64,32; x,y,z = 0,68,32). In each
task-switching condition, we then performed correlations between
each of these brain regions. We focused on brain regions correlating
with the caudate nucleus in the unpredictable task order conditions
and with brain regions correlating with the cerebellum in the random
timing conditions (Fig. 9). Figure 9a represents the pattern of
activation in the different task-switching conditions relative to
baseline (Z > 3.09, i.e. P < 0.001, uncorrected) and Fig. 9b (top)
represents the matrix of interregional correlations (R2). In order to
clarify the results, we also collapsed together the data from left and
right hemispheres in these brain networks (Fig. 9b, bottom).
When task order was unpredictable (and timing was ®xed or
random), the caudate activation positively correlated with activation
of the DLPFC and the cerebellum. In addition, when both task order
was unpredictable and timing was random, the caudate activation
FIG. 4. (Top) Data for striatal and thalamic regions signi®cantly activated inall tasks switching conditions relative to baseline were superimposed on themean structural MRI slices averaged across subjects. Slices are shown inneurological convention. The Z-values for the regional maxima are listed inTable 1. (Bottom) Percentage of signal change relative to baseline for eachtask-switching condition and for the control condition in the caudatenucleus and thalamus. Error bars indicate SEM.
FIG. 5. (Left) Main effect of task order unpredictability superimposed on anormalized coronal MRI slice of one subjects. (Right) Percentage of signalchange relative to baseline for each task-switching condition in the rightanterior putamen. Error bars indicate SEM.
FIG. 6. (Top) Main effect of timing irregularity superimposed on the meanstructural MRI slices averaged across subjects. The Z-values for the regionalmaxima are listed in Table 1. (Bottom) Percentage of signal change relativeto baseline for each task-switching condition in the right cerebellarhemisphere. Error bars indicate SEM.
FIG. 7. Interactions between task order and timing superimposed onnormalized structural MRI slices averaged across subjects. Percentage ofsignal change relative to baseline for each task-switching condition isshown at the peaks of striatal activation. Error bars indicate SEM.
1614 J.-C. Dreher and J. Grafman
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

correlated with the pre-SMA and the IPS. Thus, the caudate nucleus
communicates information relative to task order unpredictability to a
number of cortical regions.
When timing was random (and task order was predictable or
unpredictable), the cerebellum activation positively correlated with
the DLPFC, the IPS and the caudate activation. In addition, when
both timing and task order were unpredictable, the cerebellum
activation positively correlated with the pre-SMA activation.
Discussion
Our results show that both the cerebellum and the striatum were
activated bilaterally by all tasks switching conditions relative to
baseline (Figs 3 and 4). These activations cannot be attributed to
motor function because identical motor responses were required for
the task-switching and baseline conditions. Activations of the
cerebellum and basal ganglia do not support the switching attention
hypothesis because these brain regions were also activated by a
control condition requiring maintenance of two task-sets without
switching between them (Figs 3 and 4). Rather, our results show that
the cerebellum and the basal ganglia are differently sensitive to the
unpredictability of task order and timing during the processing of
sequences of tasks. The anterior striatum was activated by the main
effect of task order unpredictability (Fig. 5). In contrast, the right
posterior cerebellar hemisphere and dentate nucleus were activated
by the main effect of timing irregularity (Fig. 6).
Brain regions activated with task order unpredictability or timing
irregularity may, in part, be due to a form of oddball response.
However, it should be kept in mind that in classical oddball
paradigms both the timing and the type of stimulus (target or
distractor) are unpredictable (and thus confounded) while our study
dissociated the two components of timing and task order unpredict-
ability. Event-related potential (ERP) studies of the oddball paradigm
have revealed that rare target stimuli generate the P3b event-related
potential (Clark et al., 2000). Moreover, oddball paradigms that have
investigated the neural responses to infrequent stimuli in the auditory
or visual domains found a large degree of spatial overlap primarily in
the bilateral cerebellum and the frontal and parietal areas, as well as
in the supramarginal gyrus, frontal operculum and insular cortex
bilaterally (Linden et al., 1999; Clark et al., 2000; Stevens et al.,
2000). Although we limit our discussion to the cerebellum and the
basal ganglia in the present paper, we have previously described and
discussed the roles of distinct prefrontal regions and of the
intraparietal cortex with task order predictability/unpredictability
and timing regularity (Dreher et al., 2002).
Our results that the cerebellum and basal ganglia are not speci®c
for switching attention are in accordance with the fact that cerebellar
patients are not impaired on attention tasks that require rapid visual
orienting between spatial positions (Dimitrov et al., 1996; Helmuth
et al., 1997; Yamaguchi et al., 1998). Although early studies reported
attentional de®cits in cerebellar patients (Courchesne et al., 1994;
Courchesne & Allen, 1997), it has recently been proposed that these
de®cits might be secondary to the coordination of motor responses
(Ravizza & Ivry, 2001; Bischoff-Grethe et al., 2002). The observation
that the basal ganglia are not speci®c for switching between tasks is
also provided by several event-related fMRI studies directly
comparing switch to repeat trials (Kimberg et al., 2000; Sohn et al.,
2000). Furthermore, early-stage medicated Parkinson's disease
patients performed normally in switching attention between tasks in
which verbal responses were given (Rogers et al., 1998). Although
patients with Parkinson's disease have also been reported to have
impaired performance in switching attention set (Taylor et al., 1986;
Gotham et al., 1988; Eslinger & Grattan, 1993; Owen et al., 1993;
Hayes et al., 1998) or switching between tasks (Cools et al., 2001b),
this impairment may re¯ect a dopaminergic de®cit (Hayes et al.,
1998; Cools et al., 2001a,b), a dif®culty to instantiate a new
attentional set (Owen et al., 1993), or may be related to the disrupted
forms of motor output (Robertson & Flowers, 1990). The dopami-
nergic de®cit interpretation seems especially pertinent because
several studies reported improved performance in Parkinson's disease
patients after dopaminergic medication (Hayes et al., 1998; Cools
et al., 2001a).
When compared to baseline, all task-switching conditions averaged
together not only activated the cerebellum and the striatum but also
recruited a bilateral prefronto-parietal network (Table 1, Fig. 9a).
This network is typically reported in functional neuroimaging
experiments using attention tasks, working memory tasks and dual-
tasks (Cohen et al., 1997; Nobre et al., 1997; Adcock et al., 2000;
Bunge et al., 2000), indicating that this network is not speci®c to task
switching.
Task order unpredictability activates the anterior striatum
Increased response times on both the switch and repeat trials of the
unpredictable task order conditions show that unpredictability was
related to the overall structure of the sequence of tasks. This con®rms
a recent behavioural study (Sohn & Carlson, 2000). The anterior
putamen activation found with task order unpredictability (Fig. 5) is
in accordance with the observation that striatal neurons detect
unpredicted reward events, irrespective of the speci®c behavioural
situation in which such events occur (Ravel et al., 2001). A previous
fMRI study has shown that unpredictability of reward delivery in
humans activates the ventral striatum (Berns et al., 2001) but could
not specify whether this activation was speci®c to the unpredictability
of the reward delivery or to the unpredictability of the sequence itself.
Our results suggest that reward delivery is unnecessary for activation
of the anterior striatum and that unpredictability of task order alone is
suf®cient to induce this activation. However, the role of dopamine
cannot be totally excluded in our study, even though no reward was
delivered. Indeed, the discharges of midbrain dopamine neurons have
properties similar to the reward prediction error of temporal
difference reinforcement learning models (Schultz & Dickinson,
2000; Schultz, 2000). Dopamine neuron activity serves as an
effective reinforcement signal for learning sensorimotor associations
in the striatum. It is thus possible that the dopamine signal is required
with unpredictable sequences, especially with combined unpredict-
ability of task order and timing. Con®rming this view, substantia
nigra activation was found when comparing the switching condition
with unpredictable task order and random timing to the baseline
(Fig. 8).
When considering other brain regions activated with task order
unpredictability, we found a large network including the right inferior
frontal gyrus (BA 45), the left middle frontal gyrus (BA 9), the
intraparietal cortex bilaterally and the inferior temporal cortex (BA
20 and BA 37) (Table 1, and see Dreher et al., 2002). This con®rms
that the basal ganglia is not the only brain region coding the error
signals (Schultz & Dickinson, 2000; Schultz, 2000). Our results are
also consistent with recent studies showing that decision-making in
the presence of uncertainty involves a fronto-parietal network (Paulus
et al., 2001; Huettel et al., 2002) and that the dorsolateral prefrontal
cortex is associated with the adjustment of inferential learning on the
basis of unpredictability during a causal associative learning task
(Fletcher et al., 2001). At the outset, when all associations were
unpredictable, DLPFC activation was maximal. This response
Cerebellar and striatal functions in timing and error prediction 1615
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

FIG. 9. (a) Pattern of brain activation in the different task-switching conditions relative to baseline (Z > 3.09, i.e. P < 0.001, uncorrected). (b) Results of thefunctional connectivity analysis. (Top) Matrix of interregional correlations in the different task-switching conditions. The correlation coef®cients R2 arerepresented as colour gradations. (Bottom) To summarize the results in a lateral view of the brain, data from the left and right hemispheres were collapsedtogether. The connection between two brain regions indicate correlation coef®cients R2 > 0.5 when collapsing together the two hemispheres across the brainregions found in all task-switching conditions relative to baseline. (c) Hypothetical processes performed by the components of the bilateral prefronto-parietalnetwork. The cerebellum is assumed to compute timing adjustment and the basal ganglia to react to task order unpredictability.
FIG. 8. (Left) Substantia nigra activation found in the task-switchingcondition with unpredictable task order and random timing relative tobaseline superimposed on a normalized axial MRI slice averaged acrosssubjects. (Right) Percentage of signal change relative to baseline for eachtask-switching condition. Error bars indicate SEM.
1616 J.-C. Dreher and J. Grafman
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

attenuated with learning but, subsequently, activation in the DLPFC
was evoked by surprise violations of the learned association.
Cerebellar hemisphere activation with timing adjustment
Our behavioural analysis of the random-timing task-switching
conditions suggests that a timing operation was elicited by the ISI
manipulation. Indeed, there was an important trend toward a
reduction of RTs with time lag, suggesting that subjects could use
the probabilistic information conveyed by the passage of time to
predict the likelihood of stimulus presentation (i.e. the more time the
subjects had, the more probable the stimulus occurrence). A similar
result has recently been reported (Meiran et al., 2000). Therefore, the
increased cerebellar activation found in random relative to ®xed
timing may be attributed to the fact that the cerebellum tries to predict
the timing of stimulus presentation, yet fails to do it for short ISIs
(resulting in a prediction error because the actual outcome at short ISI
differs from the predicted timing outcome).
However, we can't rule out the possibility that, even if the trials
were blocked by duration of ISI, subjects would take longer to
perform the tasks at short ISIs than at long ones. Thus, it is possible
that the longer RT for short ISIs may be caused by a refractory period
in task performance rather than a lack of time necessary to make a
prediction about stimulus onset (Meiran et al., 2000). At the same
time, subjects may anticipate (predict) when random events will
happen and look for a pattern in random timing onset.
There is a large body of evidence that is consistent with the
hypothesis that the posterior part of the cerebellum is related to
timing adjustment (Ivry et al., 1988; Ivry, 1997). Bilateral lesions of
the lateral part of the posterior lobe of the cerebellum induce de®cits
in monitoring and reproducing timing (Ivry et al., 1988; Nichelli
et al., 1996; Ivry, 1997). Studies of eyeblink conditioning also
showed that the posterior cerebellum plays a critical role in precise
timing adjustment (Yeo & Hardiman, 1992; Gruart & Yeo, 1995).
Similarly, the lateral cerebellar cortex and cerebellar vermis
contribute to a supramodal (auditory and visual) production of a
timed motor response, particularly when it is novel or complex
(Penhune et al., 1998). Although these studies support the cerebellar
timing hypothesis, they provide little insight into what particular
regions within the cerebellar hemispheres are most critical for timing
processes. Our results show that the right cerebellar hemisphere
(lobules VI/VII/Crus I) may be particularly suited to perform this
function. Moreover, the posterior cerebellar activation which we
found is consistent with a division of the cerebellum made according
to a rostro-caudal axis. The posterior lobe, especially HVI±HVIIa,
has been linked to such higher order functions as attention and
working memory (Allen et al., 1997; Desmond et al., 1997), while the
anterior lobe represents movement execution. The absence of anterior
cerebellar hemisphere activation in our study is explained by the fact
that the same motor responses were needed in all tasks, and were
subtracted in the contrast.
The timing adjustment hypothesis may be considered as a
particular case of a more general error correction process.
However, our results show the distinct nature of the signal processed
by the cerebellum and the basal ganglia: the cerebellum adjusts to
timing information while the striatum adjusts to task order predict-
ability (Fig. 9c). This distinction is relative for the striatum because
the head of the caudate nucleus and bilateral putamen showed an
interaction between timing and task order (Fig. 7).
It should also be noted that the cerebellum does not work in
isolation, despite the fact that no other brain region than the
cerebellum was activated with random relative to ®xed timing
(Fig. 9b). Indeed, our functional connectivity analysis showed that
when timing was random (and task order was predictable or
unpredictable), the cerebellar activation positively correlated with
the DLPFC, the IPS and the caudate activation. Thus, the cerebellum
is likely to share timing information with these brain regions, which is
consistent with their known anatomical connectivity (Middleton &
Strick, 2000; Clower et al., 2001; Middleton & Strick, 2001).
An alternative view of the discrimination of temporal intervals, the
basis of prediction in the random timing condition, is to consider it a
®ne-scale discrimination of sensory information (Gao et al., 1996). If
so, then cerebellar activity could, in part, re¯ect the sensory
processing necessary to predict time onset. Finally, it may be noted
that although the cerebellar neural circuitry involved in storage of
memories for learned motor responses (e.g. eyelid conditioning) is
now well known (Medina et al., 2002), the exact mechanisms
underlying the cerebellar timing function remain unclear. A recent
study indicated that the conditioned response expression and timing
are dissociable and involve different inhibitory inputs (Bao et al.,
2002).
Comparing the neural basis of sequences of tasks andsequences of movements
Several studies support distinct roles for the basal ganglia and the
cerebellum in learning motor sequences (Jueptner & Weiller, 1998;
Doya, 2000). Doya and collaborators suggested that the cerebellum is
involved in supervised learning and the basal ganglia in reinforce-
ment learning. Jueptner proposed a double dissociation between
selection of movements, which requires the basal ganglia but not the
cerebellum, and sensory information processing, which involves the
cerebellum but not the basal ganglia. Unlike these studies comparing
the roles of the basal ganglia and the cerebellum in motor learning,
our study is the ®rst to directly compare the roles of the basal ganglia
and of the cerebellum in processing sequences of cognitive tasks. The
main difference between motor sequences and cognitive sequences is
that, in motor sequences, one act leads to the next in a chain-like
fashion. In contrast, during sequences of tasks, decisions (e.g.
between two motor responses) need to be taken following rules of the
type: `if the letter is red, is it a vowel'? Unlike previous studies
comparing the roles of the basal ganglia and the cerebellum in motor
learning, our results suggest a distinction in the pattern of brain
activation between unpredictability of task order and timing irregu-
larity.
The question of whether the factors of order and timing involve the
same neural basis for sequences of movements and for sequences of
tasks is still unanswered. A 2 3 2 factorial design varying task order
and timing predictability has previously been used for sequences of
simple ®nger movements cued by the colour of two stimuli (Sakai
et al., 2000). It is, however, dif®cult to directly compare our results
with those of this study because no analysis of the main effects of
stimulus order and timing irregularity was performed. This study
reported bilateral posterior cerebellar hemisphere activation when the
timing was random and no cerebellar activation with unpredictability
of the order of the movement (`response selection'). In addition, no
basal ganglia activation was found with response selection uncer-
tainty (Sakai et al., 2000). However, the role of the basal ganglia in
unpredictable sequences of movements requires further investigation.
Indeed, at the beginning of learning a motor sequence the anterior
striatum is activated (Grafton et al., 1992; Petersen et al., 1998;
Hikosaka et al., 1999). This activation decreased when motor
sequence become more automatic while activation of the posterior
striatum increased. Similarly in our study, when task order cannot be
learned (unpredictable order), the anterior striatum was activated
while a decrease of activation was found when task order was
Cerebellar and striatal functions in timing and error prediction 1617
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

overlearned predictable task order. This suggests that both motor and
task sequences may require the anterior striatum when they are
unpredictable. Further studies directly comparing motor and cogni-
tive sequences are needed to solve this issue (Koechlin et al., 2002)
To conclude, our study provides new evidence that the cerebellum
and striatum are involved not only in the control of movements but
also in the sequencing of cognitive tasks. The cerebellum is primarily
activated by timing irregularity while the basal ganglia show a more
complex pattern of activation, the anterior striatum being activated by
task order unpredictability while the head of the caudate nucleus and
the posterior putamen responding to an interaction between timing
and task order. Furthermore, task order unpredictability alone,
independently of reward delivery, is suf®cient to induce striatal
activation.
Acknowledgement
This research was supported by a postdoctoral fellowship from the FYSSENfoundation to J.-C.D.
Abbreviations
DLPFC, dorsolateral prefrontal cortex; fMRI, functional magnetic resonanceimaging; IPS, intraparietal sulcus; ISI, interstimulus interval; pre-SMA, pre-supplementary motor area; RT, reaction time.
References
Adcock, R.A., Constable, R.T., Gore, J.C. & Goldman-Rakic, P.S. (2000)Functional neuroanatomy of executive processes involved in dual-taskperformance. Proc. Natl Acad. Sci. USA, 97, 3567±3572.
Akshoomoff, N.A. & Courchesne, E. (1992) A new role for the cerebellum incognitive operations. Behav. Neurosci., 106, 731±738.
Albus, J.S. (1971) The theory of cerebellar function. Mathemat. Biosciences,10, 25±61.
Allen, G., Buxton, R.B., Wong, E.C. & Courchesne, E. (1997) Attentionalactivation of the cerebellum independent of motor involvement. Science,275, 1940±1943.
Bao, S., Chen, L., Kim, J.J. & Thompson, R.F. (2002) Cerebellar corticalinhibition and classical eyeblink conditioning. Proc. Natl Acad. Sci. USA,99, 1592±1597.
Berns, G.S., McClure, S.M., Pagnoni, G. & Montague, P.R. (2001)Predictability modulates human brain response to reward. J. Neurosci.,21, 2793±2798.
Bischoff-Grethe, A., Ivry, R.B. & Grafton, S.T. (2002) Cerebellar involvementin response reassignment rather than attention. J. Neurosci., 22, 546±553.
Blakemore, S.J., Frith, C.D. & Wolpert, D.M. (2001) The cerebellum isinvolved in predicting the sensory consequences of action. Neuroreport, 12,1879±1884.
Bracke-Tolkmitt, R., Linden, A.G.M., Canavan, B., Rockstroh, E., Scholz, K.,Wessel, H.-C. & Diener, H.C. (1989) The cerebellum contributes to mentalskills. Behav. Neurosci., 103, 442±446.
Bunge, S.A., Klingberg, T., Jacobsen, R.B. & Gabrieli, J.D. (2000) A resourcemodel of the neural basis of executive working memory. Proc. Natl Acad.Sci. USA, 97, 3573±3578.
Canavan, A.G., Sprengelmeyer, R., Diener, H.C. & Homberg, V. (1994)Conditional associative learning is impaired in cerebellar disease in humans.Behav. Neurosci., 108, 475±485.
Clark, V.P., Fannon, S., Lai, S., Benson, R. & Bauer, L. (2000) Responses torare visual target and distractor stimuli using event-related fMRI. J.Neurophysiol., 83, 3133±3139.
Clower, D.M., West, R.A., Lynch, J.C. & Strick, P.L. (2001) The inferiorparietal lobule is the target of output from the superior colliculus,hippocampus, and cerebellum. J. Neurosci., 21, 6283±6291.
Cohen, J.D., Perlstein, W.M., Braver, T.S., Nystrom, L.E., Noll, D.C., Jonides,J. & Smith, E.E. (1997) Temporal dynamics of brain activation during aworking memory task [see comments]. Nature, 386, 604±608.
Cools, R., Barker, R.A., Sahakian, B.J. & Robbins, T.W. (2001a) Enhanced or
impaired cognitive function in Parkinson's disease as a function ofdopaminergic medication and task demands. Cereb. Cortex, 11, 1136±1143.
Cools, R., Barker, R.A., Sahakian, B.J. & Robbins, T.W. (2001b) Mechanismsof cognitive set ¯exibility in Parkinson's disease. Brain, 124, 2503±2512.
Courchesne, E. & Allen, G. (1997) Prediction and preparation, fundamentalfunctions of the cerebellum. Learn. Mem., 4, 1±35.
Courchesne, E., Townsend, J., Akshoomoff, N.A., Saitoh, O., Yeung-Courchesne, R., Lincoln, A.J., James, H.E., Haas, R.H., Schreibman, L. &Lau, L. (1994) Impairment in shifting attention in autistic and cerebellarpatients. Behav. Neurosci., 108, 848±865.
Desmond, J.E., Gabrieli, J.D., Wagner, A.D., Ginier, B.L. & Glover, G.H.(1997) Lobular patterns of cerebellar activation in verbal working-memoryand ®nger-tapping tasks as revealed by functional MRI. J. Neurosci., 17,9675±9685.
Dimitrov, M., Grafman, J., Kosseff, P., Wachs, J., Alway, D., Higgins, J.,Litvan, I., Lou, J.S. & Hallett, M. (1996) Preserved cognitive processes incerebellar degeneration. Behav. Brain Res., 79, 131±135.
Dove, A., Pollmann, S., Schubert, T., Wiggins, C.J. & von Cramon, D.Y.(2000) Prefrontal cortex activation in task switching: an event-related fMRIstudy. Brain Res. Cogn. Brain Res., 9, 103±109.
Doya, K. (2000) Complementary roles of basal ganglia and cerebellum inlearning and motor control. Curr. Opin. Neurobiol., 10, 732±739.
Dreher, J.C., Koechlin, E., Ali, S.O. & Grafman, J. (2002) The roles of timingand task order during task switching. Neuroimage, 17, 95±109.
Eslinger, P.J. & Grattan, L.M. (1993) Frontal lobe and frontal-striatalsubstrates for different forms of human cognitive ¯exibility.Neuropsychologia, 31, 17±28.
Fiez, J.A., Petersen, S.E., Cheney, M.K. & Raichle, M.E. (1992) Impairednon-motor learning and error detection associated with cerebellar damage.A single case study. Brain, 115, 155±178.
Fletcher, P.C., Anderson, J.M., Shanks, D.R., Honey, R., Carpenter, T.A.,Donovan, T., Papadakis, N. & Bullmore, E.T. (2001) Responses of humanfrontal cortex to surprising events are predicted by formal associativelearning theory. Nature Neurosci., 4, 1043±1048.
Friston, K.J., Frith, C.D., Liddle, P.F. & Frackowiak, R.S. (1991) Comparingfunctional (PET) images: the assessment of signi®cant change. J. Cereb.Blood Flow Metab., 11, 690±699.
Gao, J.H., Parsons, L.M., Bower, J.M., Xiong, J., Li, J. & Fox, P.T. (1996)Cerebellum implicated in sensory acquisition and discrimination rather thanmotor control. Science, 272, 545±547.
Gotham, A.M., Brown, R.G. & Marsden, C.D. (1988) `Frontal' cognitivefunction in patients with Parkinson's disease `on' and `off' levodopa. Brain,111, 299±321.
Grafton, S.T., Mazziotta, J.C., Woods, R.P. & Phelps, M.E. (1992) Humanfunctional anatomy of visually guided ®nger movements. Brain, 115, 565±587.
Gruart, A. & Yeo, C.H. (1995) Cerebellar cortex and eyeblink conditioning:bilateral regulation of conditioned responses. Exp. Brain Res., 104, 431±448.
Harrington, D.L., Haaland, K.Y. & Knight, R.T. (1998) Cortical networksunderlying mechanisms of time perception. J. Neurosci., 18, 1085±1095.
Hayes, A.E., Davidson, M.C., Keele, S.W. & Rafal, R.D. (1998) Toward afunctional analysis of the basal ganglia. J. Cogn. Neurosci., 10, 178±198.
Helmuth, L.L., Ivry, R.B. & Shimizu, N. (1997) Preserved performance bycerebellar patients on tests of word generation, discrimination learning, andattention. Learn. Mem., 3, 456±474.
Hikosaka, O., Nakahara, H., Rand, M.K., Sakai, K., Lu, X., Nakamura, K.,Miyachi, S. & Doya, K. (1999) Parallel neural networks for learningsequential procedures. Trends Neurosci., 22, 464±471.
Huettel, S.A., Mack, P.B. & McCarthy, G. (2002) Perceiving patterns inrandom series: dynamic processing of sequence in prefrontal cortex. NatureNeurosci., 5, 485±490.
Imamizu, H., Miyauchi, S., Tamada, T., Sasaki, Y., Takino, R., Putz, B.,Yoshioka, T. & Kawato, M. (2000) Human cerebellar activity re¯ecting anacquired internal model of a new tool. Nature, 403, 192±195.
Ivry, R. (1996) The representation of temporal information in perception andmotor control. Curr. Opin. Neurobiol., 6, 851±857.
Ivry, R. (1997) Cerebellar timing systems. Int. Rev. Neurobiol., 41, 555±573.Ivry, R. & Fiez, J.A. (2001) Cerebellar contributions to cognition and imagery.
In Gazzaniga, M.S. (ed.), The New Cognitive Neuroscience. MIT Press,Cambridge, MA, pp 999±1011.
Ivry, R.B., Keele, S.W. & Diener, H.C. (1988) Dissociation of the lateral andmedial cerebellum in movement timing and movement execution. Exp.Brain Res., 73, 167±180.
Jueptner, M. & Weiller, C. (1998) A review of differences between basal
1618 J.-C. Dreher and J. Grafman
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619

ganglia and cerebellar control of movements as revealed by functionalimaging studies. Brain, 121, 1437±1449.
Kawato, M. & Gomi, H. (1992) A computational model of four regions of thecerebellum based on feedback-error learning. Biol. Cybern, 68, 95±103.
Kimberg, D.Y., Aguirre, G.K. & D'Esposito, M. (2000) Modulation of task-related neural activity in task-switching: an fMRI study (1). Brain Res.Cogn. Brain Res., 10, 189±196.
Koechlin, E., Danek, A., Burnod, Y. & Grafman, J. (2002) Medial prefrontaland subcortical mechanisms underlying the acquisition of motor andcognitive action sequences in humans. Neuron, 35, 371±381.
Le, T.H., Pardo, J.V. & Hu, X. (1998) 4 T-fMRI study of nonspatial shifting ofselective attention: cerebellar and parietal contributions. J. Neurophysiol.,79, 1535±1548.
Leiner, H.C., Leiner, A.L. & Dow, R.S. (1991) The human cerebro-cerebellarsystem: its computing, cognitive, and language skills. Behav. Brain Res., 44,113±128.
Linden, D.E., Prvulovic, D., Formisano, E., Vollinger, M., Zanella, F.E.,Goebel, R. & Dierks, T. (1999) The functional neuroanatomy of targetdetection: an fMRI study of visual and auditory oddball tasks. Cereb.Cortex, 9, 815±823.
Marr, D. (1969) A theory of cerebellar cortex. J. Physiol. (Lond.), 202, 437±470.
Medina, J.F., Christopher Repa, J., Mauk, M.D. & LeDoux, J.E. (2002)Parallels between cerebellum- and amygdala-dependent conditioning.Nature Rev. Neurosci., 3, 122±131.
Meiran, N., Chorev, Z. & Sapir, A. (2000) Component processes in taskswitching. Cognit. Psychol., 41, 211±253.
Miall, R.C., Reckess, G.Z. & Imamizu, H. (2001) The cerebellum coordinateseye and hand tracking movements. Nature Neurosci., 4, 638±644.
Middleton, F.A. & Strick, P.L. (2000) Basal ganglia and cerebellar loops:motor and cognitive circuits. Brain Res. Brain Res. Rev., 31, 236±250.
Middleton, F.A. & Strick, P.L. (2001) Cerebellar projections to the prefrontalcortex of the primate. J. Neurosci., 21, 700±712.
Nichelli, P., Alway, D. & Grafman, J. (1996) Perceptual timing in cerebellardegeneration. Neuropsychologia, 34, 863±871.
Nobre, A.C., Sebestyen, G.N., Gitelman, D.R., Mesulam, M.M., Frackowiak,R.S. & Frith, C.D. (1997) Functional localization of the system forvisuospatial attention using positron emission tomography. Brain, 120, 515±533.
Owen, A.M., Roberts, A.C., Hodges, J.R., Summers, B.A., Polkey, C.E. &Robbins, T.W. (1993) Contrasting mechanisms of impaired attentional set-shifting in patients with frontal lobe damage or Parkinson's disease. Brain,116, 1159±1175.
Pastor, M.A., Artieda, J., Jahanshahi, M. & Obeso, J.A. (1992) Timeestimation and reproduction is abnormal in Parkinson's disease. Brain, 115,211±225.
Paulus, M.P., Hozack, N., Zauscher, B., McDowell, J.E., Frank, L., Brown,G.G. & Braff, D.L. (2001) Prefrontal, parietal, and temporal cortexnetworks underlie decision-making in the presence of uncertainty.Neuroimage, 13, 91±100.
Penhune, V.B., Zattore, R.J. & Evans, A.C. (1998) Cerebellar contributions to
motor timing: a PET study of auditory and visual rhythm reproduction. J.Cogn. Neurosci., 10, 752±765.
Petersen, S.E., van Mier, H., Fiez, J.A. & Raichle, M.E. (1998) The effects ofpractice on the functional anatomy of task performance. Proc. Natl Acad.Sci. USA, 95, 853±860.
Ravel, S., Sardo, P., Legallet, E. & Apicella, P. (2001) Rewardunpredictability inside and outside of a task context as a determinant ofthe responses of tonically active neurons in the monkey striatum. J.Neurosci., 21, 5730±5739.
Ravizza, S.M. & Ivry, R.B. (2001) Comparison of the basal ganglia andcerebellum in shifting attention. J. Cogn. Neurosci., 13, 285±297.
Robertson, C. & Flowers, K.A. (1990) Motor set in Parkinson's disease. J.Neurol. Neurosurg. Psychiatry, 53, 583±592.
Rogers, R.D., Sahakian, B.J., Hodges, J.R., Polkey, C.E., Kennard, C. &Robbins, T.W. (1998) Dissociating executive mechanisms of task controlfollowing frontal lobe damage and Parkinson's disease. Brain, 121, 815±842.
Sakai, K., Hikosaka, O., Takino, R., Miyauchi, S., Nielsen, M. & Tamada, T.(2000) What and when: parallel and convergent processing in motorControl. J. Neurosci., 20, 2691±2700.
Schmahmann, J.D., Doyon, J., McDonald, D., Holmes, C., Lavoie, K.,Hurwitz, A.S., Kabani, N., Toga, A., Evans, A.C. & Petrides, M. (1999)Three-dimensional MRI atlas of the human cerebellum in proportionalstereotaxic space. Neuroimage, 10, 233±260.
Schultz, W. (2000) Multiple reward signals in the brain. Nature Rev.Neurosci., 1, 199±207.
Schultz, W. & Dickinson, A. (2000) Neuronal coding of prediction errors.Annu. Rev. Neurosci., 23, 473±500.
Sohn, M.H. & Carlson, R.A. (2000) Effects of repetition and foreknowledge intask-set recon®guration. J. Exp. Psychol. Learn. Mem. Cogn., 26, 1445±1460.
Sohn, M.H., Ursu, S., Anderson, J.R., Stenger, V.A. & Carter, C.S. (2000) Therole of prefrontal cortex and posterior parietal cortex in task switching.Proc. Natl Acad. Sci. USA, 97, 13448±13453.
Stevens, A.A., Skudlarski, P., Gatenby, J.C. & Gore, J.C. (2000) Event-relatedfMRI of auditory and visual oddball tasks. Magn. Reson. Imaging, 18, 495±502.
Tamada, T., Miyauchi, S., Imamizu, H., Yoshioka, T. & Kawato, M. (1999)Cerebro-cerebellar functional connectivity revealed by the laterality indexin tool-use learning. Neuroreport, 10, 325±331.
Taylor, A.E., Saint-Cyr, J.A. & Lang, A.E. (1986) Frontal lobe dysfunction inParkinson's disease. The cortical focus of neostriatal out¯ow. Brain, 109,845±883.
Thach, W.T. (1998) A role for the cerebellum in learning movementcoordination. Neurobiol. Learn. Mem., 70, 177±188.
Yamaguchi, S., Tsuchiya, H. & Kobayashi, S. (1998) Visuospatial attentionshift and motor responses in cerebellar disorders. J. Cogn. Neurosci., 10,95±107.
Yeo, C.H. & Hardiman, M.J. (1992) Cerebellar cortex and eyeblinkconditioning: a reexamination. Exp Brain Res., 88, 623±638.
Cerebellar and striatal functions in timing and error prediction 1619
ã 2002 Federation of European Neuroscience Societies, European Journal of Neuroscience, 16, 1609±1619




















