
p53, p21WAF1/CIP1, and MDM2 Involvement in the Proliferation and Apoptosis in an In Vitro Model of...
10
Catherine Demoliou-Mason J.-Kuang Hsieh, Dimitris Kletsas, Gerard Clunn, Alun D. Hughes, Michael Schachter and Vitro Model of Conditionally Immortalized Human Vascular Smooth Muscle Cells , and MDM2 Involvement in the Proliferation and Apoptosis in an In WAF1/CIP1 p53, p21 Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2000 American Heart Association, Inc. All rights reserved. Greenville Avenue, Dallas, TX 75231 is published by the American Heart Association, 7272 Arteriosclerosis, Thrombosis, and Vascular Biology doi: 10.1161/01.ATV.20.4.973 2000;20:973-981 Arterioscler Thromb Vasc Biol. http://atvb.ahajournals.org/content/20/4/973 World Wide Web at: The online version of this article, along with updated information and services, is located on the http://atvb.ahajournals.org//subscriptions/ at: is online Arteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions: http://www.lww.com/reprints Information about reprints can be found online at: Reprints: document. Question and Answer Permissions and Rights page under Services. Further information about this process is available in the which permission is being requested is located, click Request Permissions in the middle column of the Web Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for can be obtained via RightsLink, a service of the Arteriosclerosis, Thrombosis, and Vascular Biology in Requests for permissions to reproduce figures, tables, or portions of articles originally published Permissions: by guest on May 2, 2014 http://atvb.ahajournals.org/ Downloaded from by guest on May 2, 2014 http://atvb.ahajournals.org/ Downloaded from
Transcript of p53, p21WAF1/CIP1, and MDM2 Involvement in the Proliferation and Apoptosis in an In Vitro Model of...

Catherine Demoliou-MasonJ.-Kuang Hsieh, Dimitris Kletsas, Gerard Clunn, Alun D. Hughes, Michael Schachter andVitro Model of Conditionally Immortalized Human Vascular Smooth Muscle Cells
, and MDM2 Involvement in the Proliferation and Apoptosis in an InWAF1/CIP1p53, p21
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2000 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.20.4.9732000;20:973-981Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/20/4/973World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

p53, p21WAF1/CIP1, and MDM2 Involvement in theProliferation and Apoptosis in an In Vitro Model of
Conditionally Immortalized Human Vascular SmoothMuscle Cells
J.-Kuang Hsieh,* Dimitris Kletsas,* Gerard Clunn, Alun D. Hughes,Michael Schachter, Catherine Demoliou-Mason*
Abstract—Using an in vitro model of a conditionally immortalized cell line, we have investigated how human vascularsmooth muscle cells (VSMCs) are affected by the expression of simian virus 40 (SV40) large T antigen (LT antigen),which binds to cell cycle regulators such as the tumor suppressor protein p53. Cells were obtained after infection ofsaphenous vein-derived VSMCs with a nonreplicative retroviral vector containing a temperature-sensitive mutant ofSV40 LT antigen and were shown to have maintained some characteristics and responses of VSMCs. Underpermissive-temperature conditions (36°C), the increased rate of cell proliferation was shown to be associated withexpression of LT antigen and with LT antigen binding to and inactivation of p53. p53 inactivation failed to blockapoptosis induced by serum withdrawal or UV irradiation. Downregulation of LT antigen expression at a nonpermissivetemperature (39°C) was shown to be associated with growth arrest, increased expression of the cell cycle inhibitorp21WAF1/CIP1, increased murine double minute-2 promoter activity, and differential expression of murine double minute-2gene products, suggesting that p53-induced transcription/transactivation may be involved in VSMC cycle control but notnecessarily in apoptosis. The established SMC line HVTs-SM1 may be a useful model for study of the processesinvolved in myointimal hyperplasia and cellular aging, as well as for the study of cell cycle control in general.(Arterioscler Thromb Vasc Biol. 2000;20:973–981.)
Key Words: vascular smooth musclen SV-40n p53n MDM2 n p21WAF1/CIP1
V ascular smooth muscle cell (VSMC) proliferation haslong been considered to be a key event in remodeling of
the vascular wall. Remodelling occurs during development,with vascular injury in diseases like atherosclerosis, and invascular restenosis after invasive intervention.1,2 More re-cently, the precise role of cell proliferation in atherogenesishas received more critical attention,2 and other processes suchas contraction, cell migration, and apoptosis have also beenproposed to be involved. The mechanisms regulating cellcycle control in VSMCs remain largely unknown.2 PrimaryVSMCs especially those of human origin, are difficult tostudy in vitro: their availability is limited, they grow slowly,and they have a limited life span. Furthermore, cell responsesmay vary between cell lines depending on the donor, disease,or tissue.2,3 Recently, there has been an interest in in vitromodels of established VSMC lines obtained from animal orhuman primary cells by transfection with simian virus 40(SV40) large T (LT)-antigen or other viral oncogenes.4–7
However, although there are some reports of immortalizedcell lines derived from animal cells, human cells in generalhave been more difficult to immortalize.
We describe herein some of the cellular characteristics ofan SMC line derived from human saphenous vein, which wasestablished by infection with a temperature-sensitive mutantSV40 LT antigen. Rapid proliferation of cells was associatedwith LT antigen expression and LT antigen–dependent inac-tivation of G1/S checkpoint control via inactivation of cellcycle regulators like p53.8 Downregulation of LT antigenexpression under nonpermissive temperature conditions wasshown to be associated with growth arrest, the appearance ofmorphological characteristics typical of senescent cells, andupregulation of the expression of growth regulator(s) down-stream of p53, such as p21WAF1/CIP1.9 Transformed cellsshowed high levels of apoptosis, which increased after serumwithdrawal or DNA damage by UV irradiation. Because thiscell line maintains some characteristics of human VSMCs it
Received March 11, 1999; revision accepted October 13, 1999.From the Department of Clinical Pharmacology (G.C., A.D.H., M.S., C.D.-M.), National Heart and Lung Institute, Imperial College of Science,
Technology, and Medicine, St. Mary’s Hospital, London, UK; the Ludwig Institute (J.-K.H.), St. Mary’s Hospital, London, UK; and the Institute ofBiology (D.K.), NCSR “Demokritos,” Athens, Greece.
*J.-K.H., D.K., and C.D.-M. all contributed equally to this work.Correspondence to Dr A.D. Hughes, Clinical Pharmacology, National Heart and Lung Institute, Imperial College of Science, Technology, and
Medicine, St. Mary’s Hospital, London W2 1NY, UK. E-mail [email protected]© 2000 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol.is available at http://www.atvbaha.org
973 by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

may provide a useful in vitro model to study the regulation ofphenotype and to investigate changes in cell cycle controlinvolved in cellular aging, cell death, and other processescontributing to vascular disease.
MethodsReagents, Antibodies, and PlasmidsAll culture media were from Gibco Life Technologies/BRL. [methyl-3H]Thymidine (740 GBq/mmol) was from ICN Flow. Unfractionatedporcine heparin (Paynes & Byrne) was a gift from Dr B. Mulloy,National Institute of Biological Standards and Control, SouthMimms, UK. Platelet-derived growth factor-BB, epidermal growthfactor, and transforming growth factor-b were from Life Technolo-gies. All other reagents used, unless otherwise specified, were fromSigma Chemical Co. Primary and secondary antibodies were fromDAKO A/S. The p53 (non–cross-reactive) human-specific monoclo-nal antibody (mAb) DO-1,10 the p21WAF1/CIP1-specific mAbpAb53G817,11 the murine double minute-2 (MDM2)-luciferase (luc)reporter plasmid, and the independent reporter plasmid cytomegalo-virus (CMV)–b-galactosidase were kindly donated by Dr X. Lu(Ludwig Institute for Cancer Research, London, UK) and the SV40LT antigen–specific mAbs 419 and 42312 by Dr T. Kamalati(Haddow Laboratories, Institute for Cancer Research, Sutton, UK).The mAb SM-E713 for smooth muscle myosin heavy-chains type 1and 2 was a gift from Prof P. Pauletto (Clinica Medica I, Padova,Italy).
Cell Culture Conditions: Transfection andIsolation of Conditionally ImmortalizedHuman VSMCsHuman SMCs were isolated from explants derived from normalsaphenous vein from patients undergoing coronary artery bypasssurgery as previously described.3 Cells that migrated from theseexplants were subcultured at a split ratio of 1;2 in Dulbecco’smodified Eagle’s medium (DMEM) containing 15% fetal calf serum(FCS). Equal numbers of cells from 12 separate lines derived fromhuman saphenous vein–derived SMCs with similar proliferativeresponses were pooled at passage 2, subcultured, used at passage 3or 4, and are referred to in this study as normal or untransformedVSMCs.
Actively growing primary VSMCs obtained from a single middle-aged patient were infected at passage 4 with conditioned mediumfrom the amphotropic packaging PA317 cell line by using polybrene(8 mg/mL). The PA317 cell line produces the replication-defectivehelper virus–free vector pZipSV40-U19-5, which has been con-structed by inserting at the singleBamHI restriction site of the vectorpZipNeoSV(X) a full-length SV40 LT antigen carrying both thetemperature-sensitive A58 and U19 mutations.14 The temperature-sensitive A58 mutation confers temperature sensitivity for LTantigen expression, and the U19 mutation disables replication of theretroviral vector.14,15 Seven days after infection, the cells were putunder G418 (500mg/mL) selection. The infection was carried out byDr M. J. O’Hare at Haddow Laboratories, Institute of CancerResearch, Sutton, Surrey, UK. Transformed cells that survived G418selection were subcultured in DMEM containing 10% FCS and 200mg/mL G418. Infection, selection, and maintenance of cells werecarried out at the permissive temperature of 36°C. After'100population doublings, the cells were subcloned by serial dilutionbecause there was no apparent crisis. The various clones obtainedhad a similar morphology, but only 20% survived subculturing for anadditional 10 passages. One clone, designated herein as HVTs-SM1,has been in continuous culture for.2 years, exceeding 200 popu-lation doublings.
Plasmid Transfection and luc AssayHVTs-SM1 cells plated in 3-cm-diameter dishes (104/dish) in 10%FCS–DMEM were transfected 24 hours later with 1 to 2mg ofMDM2-luc reporter plasmid by using the calcium phosphate precip-
itation method.9 An equivalent amount of CMV–b-galactosidaseplasmid was cotransfected for normalization of transfection effi-ciency. The transfected cells were incubated at 36°C or 39°C, and atthe indicated time points they were washed 3 times with PBS, lysedin 100mL of lysis buffer (Promega UK Ltd), and assayed for reporteractivity of luc andb-galactosidase.16
DNA Synthesis and Cell ProliferationNormal and HVTs-SM1 cells were plated at 36°C in 96-well platesat 104 cells/well in NCTC-109 medium containing 10% FCS (10%FCS–NCTC-109). Twenty-four hours later, the cells were washed 3times with PBS and growth-arrested (72 hours) in serum-freemedium. DNA synthesis induced by the various stimulants wasmeasured 30 hours after stimulation by using 5mCi [methyl-3H]thymidine that was added 6 hours before harvesting the DNA, aspreviously described.17 Cell growth rates were measured by plating53104 cells/cm2 in 6-well plates in 10% FCS–NCTC-109 medium.Twenty-four hours later, the cells were growth-arrested in serum-freeNCTC-109 medium for 72 hours. Then the medium was replacedwith serum-free medium or 10% FCS–containing medium, in thepresence or absence of 150mg/mL heparin, and the cells wereincubated at 36°C or 39°C with medium changes every 3 days. At theindicated time points, cells were trypsinized and counted with aCoulter counter. In addition, in some studies, cells synthesizing DNAwere also identified after dual labeling with bromodeoxyuridine andHoechst 33258 dye. In brief, cells plated on glass coverslips atdifferent densities in 24-well plates overnight were growth-arrestedas above and labeled with 50mmol/L bromodeoxyuridine added infresh serum-free medium for the last 24 hours. The cells were fixedin freshly prepared 4% paraformaldehyde/PBS (10 minutes), perme-abilized with 0.2% Triton X-100/PBS (5 minutes), treated with 2NHCl (10 minutes), incubated with fluorescein-conjugated anti-bromodeoxyuridine mAb/PBS (30 minutes), and then stained with 1mg/mL Hoechst 33258/PBS (30 minutes) in the dark at roomtemperature. Cells were washed 3 times with PBS at each step.Hoechst 33258– and bromodeoxyuridine-positive nuclei werecounted on a Zeiss fluorescence microscope with a 203 objective; afield containing'200 cells was used for quantification purposes.
Immunofluorescence MicroscopyCells were plated on glass coverslips and were grown in 10%FCS–DMEM. They were washed with PBS, fixed in ice-cold ethanolor a methanol/acetone (1;1, vol/vol) solution (10 minutes), washed5 times with PBS, and air dried. Then they were incubated withprimary antibody (diluted 1;50 or 1;100 in 10% serum) overnight at4°C, washed 3 times for 10 minutes with PBS and incubated withFITC-conjugated anti-mouse or anti-rabbit IgG (1;50) in PBS at37°C (1 hour). After being washed, the cells were stained withHoechst 33258/PBS (1 ng/ml, 10 minutes) and observed under afluorescence microscope.
Immunoprecipitation and ImmunoblottingSubconfluent cultures of normal or HVTs-SM1 cells grown at 36°Cwere growth-arrested in serum-free NCTC-109 medium, which wasreplaced after 7 hours with either fresh serum-free or 10% FCSmedium. After incubation at 36°C or 39°C for the indicated periods,the cells were washed 3 times with ice-cold PBS and lysed with 100mL/106 cells of lysis buffer. Proteins in cell lysates were resolved byelectrophoresis on 10% or 12.5% SDS-polyacrylamide gels, fol-lowed by Western blotting on nitrocellulose membranes (Schleicherand Schuell). In brief, the membranes were blocked with 10% nonfatmilk in 10 mmol/L Tris-HCl, pH 7.4, 150 mmol/L NaCl, and 0.05%Tween-20 (TTBS) buffer; washed with TTBS, and incubated withthe primary antibody at room temperature for 1 hour or at 4°Covernight. Then they were incubated (1 to 2 hours) with horseradishperoxidase–conjugated secondary antibody (1;1000) in TTBS atroom temperature. Immunoreactive bands were visualized on KodakX-OMAT AR film by chemiluminescence (ECL kit) according to themanufacturer’s (Amersham) instructions.
974 Arterioscler Thromb Vasc Biol. April 2000
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

For immunoprecipitation, cell lysates (1.0mg) were preclearedwith protein G–Sepharose beads (10mL) and mixed with 5 to 10mgof purified antibody or 1 to 2 mL of ascites at 4°C overnight.Antibody complexes were precipitated with 10mL of proteinG–Sepharose beads at 4°C (1 to 2 hours). After the beads werewashed 3 times with lysis buffer and twice with the same bufferwithout NP-40, bound immunocomplexes were extracted by boiling(3 minutes) in 20mL of 23-concentrated SDS–polyacrylamide gelelectrophoresis sample buffer, electrophoresed, and immunoblotted.
UV Irradiation and Cell ApoptosisNormal and HVTs-SM1 cells were irradiated at 50 mJ/cm2 in anXL-1000 UV cross-linker (Spectronics Corp) and then incubated at36°C in 10% FCS–DMEM. At the indicated time points they weretrypsinized and counted. In parallel, the detached and adherent cellsfrom each of 2 additional wells were collected for flow cytometry,washed with PBS, fixed in 70% ice-cold ethanol, and centrifuged at2000 rpm (10 minutes 4°C). Cells and apoptotic bodies in each pelletwere resuspended in PBS containing 50mg/mL propidium iodideand 12mg/mL RNase A and incubated in the dark (30 minutes, roomtemperature). Flow cytometry was performed in a FACSORT(Becton-Dickinson), and fluorescence signals were plotted by usingCell Quest software.
Statistics and Data AnalysisAll data are presented as mean6SD of n observations. Statisticalcomparisons were made with a Student’st test; a value ofP,0.05was considered significant.
ResultsConditionally Immortalized HumanVein–Derived SMCsNormal VSMCs (passaged at a 1;3 split ratio) stoppeddividing after 10 to 15 passages and became senescent. Thetransformed HVTs-SM1 cells, however, have been in contin-uous culture at the permissive temperature for.200 popula-tion doublings. The transformed cells had lost the elongated,spindlelike shape of normal VSMCs and were smaller with amorphology ranging from polygonal epithelioid-like tofibroblast-like (Figure 1A versus 1B). When HVTs-SM1 cellswere transferred to 39°C, after'5 to 7 days, they acquiredmorphological characteristics more similar to those of normalcells (Figure 1C), except that they were enlarged and flat-tened, a characteristic of in vitro senescent cells.18,19
We investigated whether the transformed HVTs-SM1 cellsretained cytoskeletal proteins typical of SMCs. The majorityof transformed cells stained positively for smooth musclea-actin. The decorated filaments appeared to be more cyto-plasmic and irregular in the transformed cells, whereas innormal cells they traversed the cell along its long axis (Figure2: A1, A2) There were no observable differences in thestrength of smooth musclea-actin staining and filamentdistribution when either normal or transformed VSMCswere incubated at 39°C for 24 to 48 hours. Staining forvimentin was positive in both cell types, but comparedwith normal cells, the filaments of the transformed cellswere more sparse, less organized, and appeared spreadfrom the perinuclear bundles to the periphery of the cell(Figure 2: B1, B2). Antibody staining for von Willebrandfactor and desmin was negative for both normal andtransformed cells. The transformed cells showed weak andmainly perinuclear rodlike staining for smooth musclemyosin heavy chain (Figure 2, C2), whereas in normal
VSMCs, the staining was observed in organized filamentsalong the long axis of the cell (Figure 2, C1).
Expression of LT Antigen and Cell CycleRegulatory Molecules in HVTs-SM1 CellsPrevious reports on cells immortalized with SV40 haveidentified that the expression of a functional LT antigen isrequired for maintenance of the immortalized phenotype.15
Those studies also established that SV40 LT antigen binds towild-type p53 and extends its half-life.8,20 We thereforeinvestigated whether HVTs-SM1 cells expressed LT antigenconditionally and whether LT antigen expression had aneffect on p53 protein levels. Indirect immunofluorescencewith anti–LT antigen or with anti–human-specific wild-typep53 antibodies in cells grown at 36°C showed positivenuclear staining for both antigens in all cells (Figure 3). LTantigen and p53 nuclear immunofluorescence staining werereduced to almost background levels when cells were trans-ferred to 39°C (Figure 3: A2, B2), and normal cells grownunder similar conditions showed no nuclear staining with
Figure 1. Comparison of cell morphologies of normal VSMCsand transformed cells (HVTs-SM1). Phase-contrast microscopyof (A) normal confluent and (B) transformed confluent cells cul-tured at 36°C and (C) transformed confluent cells cultured at39°C. Magnification 3100.
Hsieh et al Conditionally Immortalized hVSMCs 975
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

either antibody. Similarly, immunoblotting studies showedelevated levels of LT antigen and p53 in cells grown at 36°Cbut not in cells cultured at 39°C (Figure 4). Levels of LTantigen and p53 were unaffected by the presence or absenceof 10% FCS, and p53 was found to coimmunoprecipitate withLT antigen (Figure 4). It should be noted that the HVTs-SM1cells had to be cultured for 5 to 7 days at 39°C before theprotein levels of LT antigen and p53 were reduced below thedetection limits of the antibodies used.
We also examined the transcription of molecules that areknown to be regulated by p53, such as p21 and MDM2,because it had been documented that LT antigen binding top53 may adversely affect its function.20 The specific mAbto the cell cycle inhibitor p21 detected a 21-kDa proteinband when the transformed cells were cultured at 39°Conly (Figure 4). Taking into account the observedtemperature-dependent LT antigen and p53 protein expres-sion profiles and the coimmunoprecipitation of LT antigen
Figure 2. Immunofluorescence stainingof smooth muscle cytoskeletal proteins.Normal SMCs (column 1) and trans-formed HVTs-SM1 cells (column 2) cul-tured at 36°C were assessed for smoothmuscle–specific a-actin (A), vimentin (B),and smooth muscle myosin heavy chain(C). Magnification 3400.
Figure 3. LT antigen and p53 immuno-fluorescence staining. TransformedHVTs-SM1 cells cultured at 36°C (A1,B1) or 39°C (A2, B2) were stained withLT antigen mAb (A) or with p53 mAb (B).Magnification 3400.
976 Arterioscler Thromb Vasc Biol. April 2000
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from
with p53, these results are in keeping with the proposal thatp53-induced p21 expression may be inhibited by LTantigen binding to p53.9
To investigate the effect of LT antigen expression on thetranscriptional regulation of MDM2 by p53, HVTs-SM1 cells were transfected with an MDM2-luc reporter plasmid. The
luc activity profile obtained showed a 4-fold increase inMDM2-promoter activity when the cells were cultured at39°C rather than at 36°C (Figure 5). Furthermore, Westernblot analysis of HVTs-SM1 lysates from cells cultured at the2 temperatures indicated that MDM2 protein levels were alsoreduced in HVTs-SM1 cells cultured at permissive tempera-ture (Figure 5, insert).
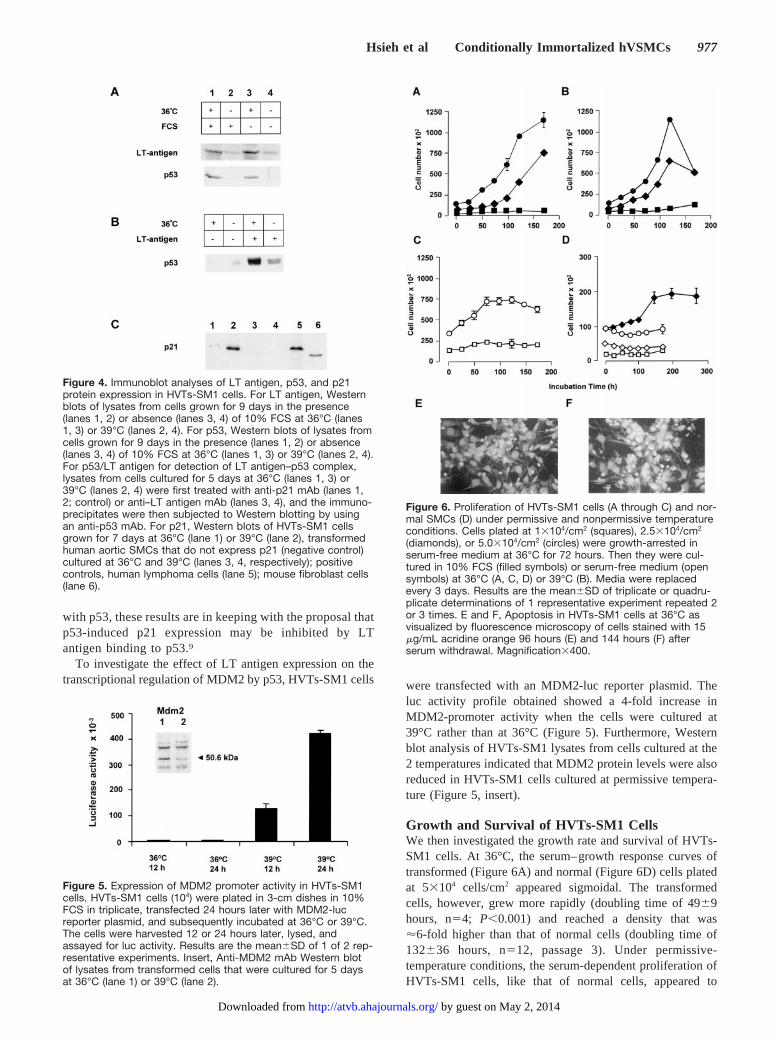
Growth and Survival of HVTs-SM1 CellsWe then investigated the growth rate and survival of HVTs-SM1 cells. At 36°C, the serum–growth response curves oftransformed (Figure 6A) and normal (Figure 6D) cells platedat 53104 cells/cm2 appeared sigmoidal. The transformedcells, however, grew more rapidly (doubling time of 4969hours, n54; P,0.001) and reached a density that was'6-fold higher than that of normal cells (doubling time of132636 hours, n512, passage 3). Under permissive-temperature conditions, the serum-dependent proliferation ofHVTs-SM1 cells, like that of normal cells, appeared to
Figure 4. Immunoblot analyses of LT antigen, p53, and p21protein expression in HVTs-SM1 cells. For LT antigen, Westernblots of lysates from cells grown for 9 days in the presence(lanes 1, 2) or absence (lanes 3, 4) of 10% FCS at 36°C (lanes1, 3) or 39°C (lanes 2, 4). For p53, Western blots of lysates fromcells grown for 9 days in the presence (lanes 1, 2) or absence(lanes 3, 4) of 10% FCS at 36°C (lanes 1, 3) or 39°C (lanes 2, 4).For p53/LT antigen for detection of LT antigen–p53 complex,lysates from cells cultured for 5 days at 36°C (lanes 1, 3) or39°C (lanes 2, 4) were first treated with anti-p21 mAb (lanes 1,2; control) or anti–LT antigen mAb (lanes 3, 4), and the immuno-precipitates were then subjected to Western blotting by usingan anti-p53 mAb. For p21, Western blots of HVTs-SM1 cellsgrown for 7 days at 36°C (lane 1) or 39°C (lane 2), transformedhuman aortic SMCs that do not express p21 (negative control)cultured at 36°C and 39°C (lanes 3, 4, respectively); positivecontrols, human lymphoma cells (lane 5); mouse fibroblast cells(lane 6).
Figure 5. Expression of MDM2 promoter activity in HVTs-SM1cells. HVTs-SM1 cells (104) were plated in 3-cm dishes in 10%FCS in triplicate, transfected 24 hours later with MDM2-lucreporter plasmid, and subsequently incubated at 36°C or 39°C.The cells were harvested 12 or 24 hours later, lysed, andassayed for luc activity. Results are the mean6SD of 1 of 2 rep-resentative experiments. Insert, Anti-MDM2 mAb Western blotof lysates from transformed cells that were cultured for 5 daysat 36°C (lane 1) or 39°C (lane 2).
Figure 6. Proliferation of HVTs-SM1 cells (A through C) and nor-mal SMCs (D) under permissive and nonpermissive temperatureconditions. Cells plated at 13104/cm2 (squares), 2.53104/cm2
(diamonds), or 5.03104/cm2 (circles) were growth-arrested inserum-free medium at 36°C for 72 hours. Then they were cul-tured in 10% FCS (filled symbols) or serum-free medium (opensymbols) at 36°C (A, C, D) or 39°C (B). Media were replacedevery 3 days. Results are the mean6SD of triplicate or quadru-plicate determinations of 1 representative experiment repeated 2or 3 times. E and F, Apoptosis in HVTs-SM1 cells at 36°C asvisualized by fluorescence microscopy of cells stained with 15mg/mL acridine orange 96 hours (E) and 144 hours (F) afterserum withdrawal. Magnification3400.
Hsieh et al Conditionally Immortalized hVSMCs 977
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

decrease through contact inhibition (Figure 6A versus Figure6D). Anti-bromodeoxyuridine antibody labeling and acridineorange staining showed that cell division as well as cell deathby apoptosis was taking place in HVTs-SM1 cells concur-rently during this period (data not shown). After a longincubation period (3 to 4 months), normal VSMCs continuedto proliferate very slowly and form a densely packed multi-layer, whereas the transformed cells did not form such amultilayer.
Unlike the situation with nontransformed cells, serumwithdrawal under permissive conditions did not cause theHVTs-SM1 cells to growth-arrest (.90% of cells werebromodeoxyuridine-positive 24 hours later). However, thecells were unable to sustain continuous proliferation, theirnumbers declined (Figure 6C), and acridine orange stainingdemonstrated many apoptotic cells during this period (Figure6E and 6F). In contrast, serum withdrawal from normalSMCs induced only a small loss of cells within the first 24 to48 hours. The remaining cells appeared to survive in agrowth-arrested state for at least 1 week (Figure 6D).
The temperature change to 39°C appeared to have no effecton the serum-dependent proliferation of HVTs-SM1 cells inthe first few days. However, cell number declined 5 to 7 dayslater (Figure 6B), and acridine orange staining indicatedincreased apoptosis. The fate of surviving cells depended ontheir density. If the cells did not become very sparse after theperiod of rapid cell death, they continued to proliferate veryslowly and remained viable for at least 3 to 4 months. Flowcytometry indicated that the majority of cells accumulated atthe G1 as well as G2 phases of the cell cycle (the Table),inagreement with previous reports.15 With time, there was asignificant increase in cell size and the appearance of perinu-clear vacuoles, and these cells resembled in vitro senescent oratherosclerotic cells.18,19 Provided that the temperature waslowered to 36°C before the cells became very sparse, theywere able to resume very quickly as vigorous a growth inresponse to serum as that of cells that had not been exposedto the nonpermissive temperature.
We also investigated whether HVTs-SM1 cells retainedresponses to smooth muscle growth factors and heparin. Asshown in Figure 7A, under permissive-temperature condi-tions, induction of HVTs-SM1 cells by 10% FCS had a largepositive effect on thymidine incorporation, whereas inductionwith platelet-derived growth factor and epidermal growth
factor had only a small positive effect; transforming growthfactor-b had no apparent effect. Heparin (150mg/mL) inhib-ited thymidine incorporation induced by serum by'50% andthat induced by platelet-derived growth factor and epidermalgrowth factor to a lesser extent (Figure 7A). Heparin inhibi-tion of serum-induced DNA synthesis was broadly compara-
Flow-Cytometric Analyses of UV-Irradiated Normal VSMCs and HVTs-SM1 Cells
Time After UV Irradiation
Normal VSMCs, % HVTs-SM1 Cells, %
G1 S G2 G1 S G2
Zero time (control) 70 1 18 65 11 17
6 Hours 71 3 18 47 9 10
12 Hours 68 4 20 37 12 14
24 Hours 67 6 19 29 8 29
Flow-cytometric determinations of the time-dependent frequency distribution of cell phases (G1, S,G2) of UV-irradiated normal VSMCs and HVTs-SM1 cells are presented. Subconfluent cells platedovernight in 6-well plates in 10% FCS/NCTC-109 at 36°C were UV irradiated at 50 mJ/cm2 andharvested either immediately or after incubation in fresh 10% FCS/NCTC-109 medium for theindicated time periods. Harvested cells were fixed in 70% ethanol and stained with propidium iodidebefore analysis. Results are from 1 of 2 representative determinations.
Figure 7. DNA synthesis and proliferation of HVTs-SM1 cellsinduced by serum and growth factors in the presence andabsence of heparin. A, Cells plated at 53104 in 96-well plates at36°C were growth-arrested for 48 to 72 hours in serum-freemedium and then induced with either 10% FCS or 5 ng/mLgrowth factor in medium only, in the absence (unfilled bars) orpresence (filled bars) of 150 mg/mL heparin. B, Time-dependentproliferation of cells plated at 53104 in 35-mm-diameter plates.Cells were incubated with 10% FCS (circles) or 0.5% FCS(squares) in the absence (unfilled symbols) or presence (filledsymbols) of heparin (150 mg/mL). Results represent themean6SD of quadruplicate determinations of 1 of 2 representa-tive experiments.
978 Arterioscler Thromb Vasc Biol. April 2000
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

ble to what we previously observed in normal VSMCs,3 andno further effect was seen after an increase in heparinconcentration up to 5-fold. Serum-induced cell proliferationwas inhibited by 25% to 30% when cells were cultured in thepresence of 150mg/mL heparin for 7 days (Figure 7B). Whencells were cultured in the presence of 0.5% serum, addition ofheparin (150mg/mL) resulted in a small net reduction in cellnumber (Figure 7B versus Figure 6C).
LT Antigen Transformation Increases SMCSensitivity to UV IrradiationFlow cytometry of UV-irradiated (50 mJ/cm2) normal SMCscultured at 36°C, showed no significant time-dependentincrease in the percentage of apoptotic cell bodies (control, 0time, 10%; 24 hours after irradiation, 7%), suggesting that thecells were resistant to UV irradiation. The majority of cellsremained in the G1 phase of the cell cycle, with only a smallproportion of cells transversing the S phase (the Table), inkeeping with the slow proliferation rate of normal SMCs. Incontrast, the transformed cells showed a 6-fold increase in thepercentage of apoptotic bodies as early as 6 hours afterirradiation (control, 0 time, 6%; 6 hours after irradiation,34%). This was accompanied by a time-dependent increase indegraded DNA and a decrease in the percentage of cells in theG1 phase. UV irradiation had no significant effect on theproportion of cells traversing the S phase. However, 24 hoursafter irradiation, there was an increase in the proportion ofcells in the G2 phase (the Table). Compared with nonirradi-ated HVTs-SM1 cells, there was an'40% to 50% reductionin cell numbers 48 to 50 hours after irradiation; this wasaccompanied by a period of growth arrest and then recovery(Figure 8). Early apoptosis of UV-irradiated HVTs-SM1 cellswas also confirmed by detection of DNA fragmentation 4hours after irradiation (data not shown).
DiscussionIn vitro aging and senescence limit the life span of primaryhuman VSMCs.18,19 This limitation and the interindividualand intraindividual variabilities of cellular responses togrowth factors/inhibitors3 emphasize the need for established
cell lines. However, in contrast to rodent cells, only a fewsuch human cell lines have been established by viral infec-tion.21 In a previous study that used SV40 infection of humanVSMCs, the maximum life span reported was'60 popula-tion doublings.22 Using a temperature-sensitive replication-defective SV40 vector, we have been able to establish animmortal (life span .200 population doublings) humanVSMC line that we have named HVTs-SM1. These cells areconditionally immortalized because their proliferative pheno-type is expressed only under the permissive temperature.Under nonpermissive-temperature conditions, the cellsgrowth-arrest and eventually lose viability in a density-dependent manner, implying the presence of an autoregula-tory mechanism for survival. The transformed cells aresmaller and have lost the typical spindle morphology thatcharacterizes normal VSMCs. Nevertheless, the cells con-tinue to express smooth muscle–specific cytoskeletal pro-teins, including myosin heavy chain anda-actin. We havealso shown at permissive temperatures that HVTs-SM1 cellsretain, to some extent, their response to SMC growth factors,like platelet-derived growth factor-BB, and to growth inhib-itors like heparin.3,23 However, DNA synthesis in response toplatelet-derived growth factor-BB was 2- to 3-fold lower thanin normal cells, possibly as a result of the LT antigen–induceddownregulation of platelet-derived growth factor receptorexpression.24
SMCs play an important role in the pathogenesis ofatherosclerosis, vascular graft occlusion, and restenosis afterangioplasty. Although the precise role of smooth muscleproliferation and apoptosis in the development of thesediseases, particularly atherosclerosis, is debated,2,25 it is likelythat these processes contribute, at least at some stage, to theirpathogenesis. HVTs-SM1 cells may be a useful model tostudy the role of growth regulators in normal and pathologicalVSMC proliferation, cell death, and other aspects of cellcycle control.
SV40 LT antigen is known to have transforming andtumorigenic potential, because it is able to target and inacti-vate several molecules involved in the control of cell cycleprogression, including the tumor suppressor molecules Rband p53.20 p53 mediates transcription of the cyclin-dependentkinase inhibitor p21WAF1, and this is a key event in cell growtharrest and cell apoptosis due to DNA damage.9 p21 regulatesthe activity of Rb by inhibiting its phosphorylation, thesubsequent release of the E2F family members, and finallythe cell’s entry into the S phase.26 p53 is also known tointeract with the tumor progression factor MDM2,27,28whichpromotes p53 degradation and can inhibit p53 transactivationand transrepression.29,30 Interestingly, both p53 and MDM2have been reported to be expressed in SMCs and macro-phages in human atherosclerotic tissue,31 and high levels ofp53 in association with CMV infection were seen in 1 studyof coronary restenosis.32 Our results indicate that the rapidand continuous proliferation of HVTs-SM1 cells underpermissive-temperature conditions is associated with theexpression of SV40 LT antigen and with LT antigen bindingand inactivation of p53. This conclusion was based on theevidence that the expression of p21WAF1 protein and of MDM2promoter activity was inhibited under these conditions. Be-
Figure 8. Proliferation of UV-irradiated HVTs-SM1 cells.UV-irradiated (open symbols) and nonirradiated (filled symbols)cells cultured in 10% FCS/NCTC at 36°C for the indicated timepoints were harvested and counted. Results are the mean6SDof triplicate determinations of 1 of 2 experiments.
Hsieh et al Conditionally Immortalized hVSMCs 979
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

cause Rb activity is regulated by p21-dependent phosphory-lation, deregulation of p21 expression would also be expectedto affect Rb activity. In addition, as previously reported,direct binding of LT antigen to Rb may release E2F tran-scription factor(s) and thus, contribute further to the deregu-lation of cell growth arrest and apoptosis.33 Deregulation ofcell cycle control at the level of p53 and of growth regulatorsthat depend on p53 activity in HVTs-SM1 cells is furthersuggested by the fact that when LT antigen is no longerfunctional owing to a temperature shift to 39°C, the cells wereable to growth-arrest. Under these conditions, the cellsexpress high levels of p21, as is observed in senescent cells invitro.34 Furthermore, cells remained viable, could be rescued,and resumed proliferation even several weeks later afterreturn to the permissive temperature.
Apoptosis of VSMCs has been observed in vasculatureremodeling during development and may be another impor-tant mechanism in the modulation of cellularity in vasculardisease.35 We observed that proliferation of HVTs-SM1 cellsat the permissive temperature was accompanied by apoptosis.Compared with normal cells, apoptosis of transformed cellswas increased by serum withdrawal, suggesting that the cellsare more dependent on serum factors for survival than forproliferation. It has been proposed recently that p53 is apotent inducer of apoptosis when Rb is inhibited in smoothmuscle.22 However, the data from our system in which bothRb and p53 are inactivated by SV40 LT antigen (at thepermissive temperature) imply the presence of a p53-independent pathway for apoptosis. This inference is sup-ported by the conclusions of a recent study, which examinedthe effect of p53 inactivation on atherogenesis in apoE-knockout mice, an animal model for atherosclerosis.36 How-ever, the cell death observed after cell transfer to thenonpermissive temperature may be attributable to the slowloss of SV40 LT antigen expression. UV irradiation inducedrapid apoptosis in HVTs-SM1 cells grown at the permissivetemperature, whereas normal cells did not undergo detectableapoptosis after UV irradiation. The reason for this differenceis unclear but may be a result of their slow proliferation rate,which allows DNA repair to take place. This questionrequires further study, and examination of the effects of morepathophysiologically relevant mediators such as oxidizedlipoproteins, oxysterols, and nitric oxide may be of particularinterest. Apoptosis after DNA damage by UV irradiation hasalso been associated with increased p53 activity,37 but fromthe results of the present study, it appears that p53-inducedtranscriptional activity is not required for UV-induced apo-ptosis. Overall, 1 possible hypothesis is that in HVTs-SM1cells, E2F1 overexpression (due to inactivation of Rb by LTantigen) can induce apoptosis independently of p53, aspreviously shown in Saos-2 cells,27 which are null for p53 andlack a functional Rb. Further studies will be required toconfirm this hypothesis.
In conclusion, we have shown that the established HVTs-SM1 cell line can be used as an in vitro model to studyspecific perturbations of cell proliferation. This model ofhuman VSMCs offers opportunities to establish a betterunderstanding of the processes and molecules involved in the
aging and senescence of the vasculature and in the control ofthe cell cycle in human VSMCs.
AcknowledgmentsThis work was supported by the Thrombosis Research Trust (UK),the British Heart Foundation (UK), the British Council, the UKHome Office, and the Greek-Britain Scientific and TechnologicalCollaboration. D. Kletsas was supported with an FEBS fellowship.We are grateful to M. J. O’Hare for the transfection of cells and K.Gallagher for assistance with tissue culture. We also thank H.Pratsinis and members of the Clinical Pharmacology Department,Imperial College, for their helpful comments and assistance.
References1. Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s.
Nature. 1993;362:801–809.2. Schwartz SM, Murry CE. Proliferation and the monoclonal origin of
atherosclerotic lesions.Ann Rev Med. 1998;49:437–460.3. Munro E, Chan P, Patel M, Betteridge L, Gallagher K, Schachter M,
Sever P, Wolfe J. Consistent responses of the human vascular smoothmuscle cell in culture: implications for restenosis.J Vasc Surg. 1994;20:482–487.
4. Reilly CF. Rat vascular smooth muscle cells immortalised with SV40large T antigen possess defined smooth muscle cell characteristicsincluding growth inhibition by heparin.J Cell Physiol. 1990;142:342–351.
5. Legrand A, Greenspan P, Nagpal ML, Nachtigal SA, Nachtigal M. Char-acterisation of human vascular smooth muscle cells transformed by theearly genetic region of SV40 virus.Am J Pathol. 1991;139:629–640.
6. Jahn L, Sadoshima J, Greene A, Parker C, Morgan KG, Izumo S. Con-ditional differentiation of heart- and smooth muscle-derived cells trans-formed by a temperature-sensitive mutant of SV40 T antigen.J Cell Sci.1996;109:397–407.
7. Ehler E, Jat PS, Noble MD, Citi S, Draeger A. Vascular smooth musclecells of H-2Kb-tsA58 transgenic mice: characterization of cell lines withdistinct properties.Circulation. 1995;95:3289–3296.
8. Levine AJ. p53, the cellular gatekeeper for growth and division.Cell.1997;88:323–331.
9. El-Deiry WS, Harper JW, O’Connor PM, Velculescu VE, Canman CE,Jackman J, Pietenpol JA, Burrell M, Hill ME, Wang Y, Wiman KG,Mercer WE, Kastan MB, Kohn KW, Elledge SJ, Kinzler KW, Vogelstein.WAF1/CIP1 is induced in p-53-mediated arrest and apoptosis.CancerRes. 1994;54:1169–1174.
10. Vojtesek B, Bartek J, Midgley CA, Lane DP. An immunochemicalanalysis of the human nuclear phosphoprotein p53: new monoclonalantibodies and epitope mapping using recombinant p53.J ImmunolMethods. 1992;151:237–244.
11. Fredersdorf S, Burns J, Milne AM, Packham G, Fallis L, Gillett CE,Royds JA, Peston D, Hall PA, Hanby AM, Barnes DM, Shousha S,O’Hare MJ, Lu X. High level expression of p27(kip1) and cyclin D1 insome human breast cancer cells: inverse correlation between theexpression of p27(kip1) and degree of malignancy in human breast andcolorectal cancers.Proc Natl Acad Sci USA. 1997;94:6380–6385.
12. Harlow E, Crawford LV, Pim DC, Williamson NM. Monoclonal anti-bodies specific for simian virus 40 tumor antigens.J Virol. 1981;39:861–869.
13. Borrione AC, Zanellato AM, Scannapieco G, Pauletto P, Sartore S.Myosin heavy-chain isoforms in adult and developing rabbit vascularsmooth muscle.Eur J Biochem. 1989;183:413–417.
14. Stamps AC, Davies SC, Burman J, O’Hare MJ. Analysis of proviralintegration in human mammary epithelial cell lines immortalized byretroviral infection with a temperature-sensitive SV40 T-antigen con-struct.Int J Cancer. 1994;57:865–874.
15. Jat PS, Sharp PA. Cell lines established by a temperature-sensitive SimianVirus 40 Large-T-Antigen gene are growth restricted at the nonper-missive temperature.Mol Cell Biol. 1989;9:1672–1681.
16. Fuchs B, O’Connor D, Fallis L, Sheidtmann KH, Lu X. p53 phosphory-lation mutants retain transcription activity.Oncogene. 1995;10:789–793.
17. Patel MK, Betteridge LJ, Hughes AD, Clunn GF, Schachter M, Shaw RJ,Sever PS: Effects of angiotensin II on the expression of the early growthresponse gene c-fos and DNA synthesis in human vascular smoothmuscle cells.J Hypertens. 1996;14:341–347.
980 Arterioscler Thromb Vasc Biol. April 2000
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from

18. Cristofalo VJ, Pignolo RJ. Replicative senescence of humanfibroblast-like cells in culture.Physiol Rev. 1993;73:617–638.
19. Dice JF. Cellular and molecular mechanisms of aging.Physiol Rev.1993;73:149–159.
20. Manfredi JJ, Prives C. The transforming activity of simian virus 40 largetumor antigen.Biochim Biophys Acta. 1994;1198:65–83.
21. Shay JW, Van-Der-Haegen BA, Ying Y, Wright WE. The frequency ofimmortalization of human fibroblasts and mammary epithelial cells trans-fected with SV40 large T-antigen.Exp Cell Res. 1993;209:45W52.
22. Bennett MR, MacDonald K, Chan S-W, Boyle JL, Weissberg PL. Coop-erative interactions between RB and p53 regulate cell proliferation, cellsenescence, and apoptosis in human vascular smooth muscle cells fromatherosclerotic plaques.Circ Res. 1998;82:704–712.
23. Chan P, Mill S, Mulloy B, Kakkar V, Demoliou-Mason C. Heparininhibition of human vascular smooth muscle cell hyperplasia.Int Angiol.1992;11:261–267.
24. Wang JL, Nister M, Bongcamrudloff E, Ponten J, Westermark B. Sup-pression of platelet-derived growth factora- andb-receptor mRNA levelsin human fibroblasts by SV40 T/T antigen.J Cell Physiol. 1996;166:12–21.
25. Mann JM, Kaski JC, Pereira WI, Arie S, Ramirez JA, Pileggi F. Histo-logical patterns of atherosclerotic plaques in unstable angina patients varyaccording to clinical presentation.Heart. 1998;80:19–22.
26. Reed SI, Bailly E, Vjekosiav D, Hengst L, Resnitzky D, Slingerland J. GIcontrol in mammalian cells.J Cell Sci. 1994;18:69–73.
27. Chen CY, Oliner JD, Zhan Q, Fornace AJ, Vogelstein B, Kastan MB.Interactions between p53 and MDM2 in a mammalian cell cyclecheckpoint pathway.Proc Natl Acad Sci USA. 1994;91:2684–2688.
28. Barak Y, Juven T, Haffner R, Oren M. MDM2 expression is induced bywild type p53 activity.EMBO J. 1993;12:461–468.
29. Momand J, Zambetti GP, Oslon DC, George D, Levine AJ. The MDM-2oncogene product forms a complex with p53 protein and inhibits p53-mediated transactivation.Cell. 1992;69:1237–1245.
30. Kubbutat MH, Jones SN, Vousden KH. Regulation of p53 stability byMDM2. Nature. 1997;387:299–303.
31. Ihling C, Haendeler J, Menzel G, Hess RD, Fraedrich G, Schaefer HE,Zeiher AM. Co-expression of p53 and MDM2 in human atherosclerosis:implications for the regulation of cellularity of atherosclerotic lesions.J Pathol. 1998;185:303–312.
32. Speir E, Modali R, Huang ES, Leon MB, Shawl F, Finkel T, Epstein SE.Potential role of human cytomegalovirus and p53 interaction in coronaryrestenosis.Science. 1994;265:391–394.
33. Hsieh J-K, Fredersdorf S, Kouzarides T, Martin K, Lu X. E2F1-inducedapoptosis requires DNA binding but not transactivation and is inhibitedby the retinoblastoma protein through direct interaction.Genes Dev.1997;11:1840–1852.
34. Stein GH, Dulic V. Origins of G1 arrest in senescent human fibroblasts.BioEssays. 1995;17:537–543.
35. Isner JM, Kearney M, Bortman S, Passeri J. Apoptosis in human athero-sclerosis and restenosis.Circulation. 1995;91:2703–2711.
36. Guevara NV, Kim HS, Antonova EI, Chan L. The absence of p53accelerates atherosclerosis by increasing cell proliferation in vivo.NatMed. 1999;5:335–339.
37. Lane D, Lu X, Hupp T, Hall PA. The role of the p53 protein in theapoptotic response.Phil Trans R Soc Lond B Biol Sci. 1994;345:277–280.
Hsieh et al Conditionally Immortalized hVSMCs 981
by guest on May 2, 2014http://atvb.ahajournals.org/Downloaded from




















