
An EBV membrane protein expressed in immortalized lymphocytes transforms established rodent cells
10
Cell, Vol. 43, 831-840, December 1985 (Part 2), Copyright 0 1985 by MIT 0092-8674/85/l 30831-l 0 $02.0010 An EBV Membrane Protein Expressed in Immortalized Lymphocytes Transforms Established Rodent Cells David Wang, David Liebowitz, and Elliott Kieff Kovler Viral Oncology Laboratories The University of Chicago 910 East 58th Street Chicago, Illinois 60637 Summary Epstein-Barr virus expresses a cytoplasmic and plas- ma membrane protein (LMP) in latently infected growth transformed lymphocytes. The gene specifying LMP has now been expressed in NIH3T3 and Rat-l cells. Ex- pression of the gene in these cells resulted in altered cell morphology and some resistance to the growth in- hibiting effect of medium containing low serum. In Rat-l cells, LMP expression often led to loss of contact inhibition and anchorage-independent growth in soft agar. Rat-l cells expressing LMP were uniformly tu- morigenic in nude mice. Thus, LMP is a transforming gene which is likely to account for many aspects of EBV induced cell transformations. This is the first demonstration of a transforming gene in Epstein-Barr virus, a ubiquitous human pathogen associated with neoplasia. Introduction Epstein-Barr Virus (EBV) is believed to cause Burkitt Lym- phoma, nasopharyngeal carcinoma, and lymphomas in immune deficient patients (for reviews see Kieff et al., 1985, and Dambaugh et al., 1986). In vitro, virus infection of human lymphocytes rapidly and efficiently induces sus- tained lymphocyte proliferation (Henle et al., 1967; Pope et al., 1968). Latently infected proliferating lymphocytes will form tumors in the brain of adult nude mice, and EBV induces lymphomas in marmosets (Giovanella et al., 1979; Miller et al., 1977). Latently infected proliferating lympho- cytes and cells of EBV associated malignancies contain complete EBV genomes as an episome or integrated into cell DNA (zur Hausen et al., 1970; Kawai et al., 1973; Nonoyama and Pagano, 1972; Lindahl et al., 1976; Matsuo et al., 1984). In such cells the latent viral genome charac- teristically expresses only four genes that encode proteins (for review see Dambaugh et al., 1986). These four genes encode three nuclear proteins and one membrane protein (Hennessy and Kieff, 1983; 1985; Hennessy et al., 1983; 1984; 1985; Fennewald et al., 1984; Dambaugh et al., 1984). The role of each of these genes in induction and maintenance of cell proliferation is unknown. Since mem- brane proteins encoded by “tumor” viruses play an impor- tant role in cell transformation (for review see Bishop, 1985) we have investigated the role of the EBV membrane protein in this process. The EBV latent infection membrane protein (LMP) is translated from an RNA transcribed from the right end of the virus genome (Figure 1; Fennewald et al., 1984; Hen- nessy et al., 1984). The transcript is 2.6 kb and is spliced to a 2.4 kb mRNA (Fennewald et al., 1984). The RNA has a 1.3 kb open reading frame which begins about 60 nucleotides after the cap site and crosses two introns (Fennewald et al., 1984). From the nucleotide sequence of the open reading frame, LMP was predicted to have six hydrophobic transmembrane domains and a 200 amino acid acidic carboxy terminus. Antibody to the acidic car- boxy domain expressed in bacteria from an EBV DNA- /acZ fusion reacts with a 60 kd protein in the cytoplasmic and plasma membranes of latently infected lymphocytes (Fennewald et al., 1984; Hennessy et al., 1984). A mono- clonal antibody, S12, directed against the bacterial fu- sion protein was used to confirm that LMP is distributed throughout the infected lymphocyte cytoplasmic mem- branes, including the plasma membrane (Mann et al., 1985; Liebowitz et al., 1986). Only a short-reverse turn domain is exposed on the outer cell surface, and the protein patches in lymphocyte plasma membranes (Lie- bowitz et al., 1986). The observation that the LMP patches in the plasma membrane also suggested that it could be part of a complex that might play a direct role in virus in- duction of cell proliferation. To further explore the relation- ship of LMP with the induction of cell proliferation, the ef- fect of the gene on the growth of continuous rodent cell lines has been assayed. Although previous attempts to identify a transforming gene in EBV infected continuous lymphoblastoid cell lines or Burkitt tumors were unsuc- cessful (Diamond et al., 1983), our results indicate that the LMP gene transforms established rodent cell lines. Results Expression of the LMP in Murine Cells The first approach to expressing LMP in murine cells was to insert an EBV DNA fragment that included 5.6 kb 5’ to the LMP mRNA cap site and 0.3 kb 3’ to its polyadenyla- tion site into the positive selection vector pSV2-neo (Fig- ure 1). Twenty-eight clones of G418 resistant 3T3 cells from three different transfections were selected, but by immu- noblot and immunofluorescence analysis contained no detectable LMP Two further constructs (Figure 1) were made. One con- struct, pZip neo LM delta, consisted of the LMP gene with- out its polyadenylation site recombined into the cloning site of pZip(neo) SV(X)l, a vector which provides MuLV transcriptional regulation, capping site, 5’ leader with cis- acting site for packaging into MuLV, and 3’ polyadenyla- tion signal (Cepko et al., 1984). The second construct, pSV2gptMTLM, consisted of the human metallothionine promoter and capping site (Karin and Richards, 1982), the LMP gene with its own polyadenylation site, and the vector pSV2gpt. The pZip(neo) SV(X)l and pSV2gpt vectors con- tain genes that permit positive selection of transfected cells in G418 or mycophenolic acid, respectively. In the next series of experiments, pZip neo LM delta was transfected into psi am 22b, an NIH3T3 cell line express-
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of An EBV membrane protein expressed in immortalized lymphocytes transforms established rodent cells

Cell, Vol. 43, 831-840, December 1985 (Part 2), Copyright 0 1985 by MIT 0092-8674/85/l 30831-l 0 $02.0010
An EBV Membrane Protein Expressed in Immortalized Lymphocytes Transforms Established Rodent Cells
David Wang, David Liebowitz, and Elliott Kieff Kovler Viral Oncology Laboratories The University of Chicago 910 East 58th Street Chicago, Illinois 60637
Summary
Epstein-Barr virus expresses a cytoplasmic and plas- ma membrane protein (LMP) in latently infected growth transformed lymphocytes. The gene specifying LMP has now been expressed in NIH3T3 and Rat-l cells. Ex- pression of the gene in these cells resulted in altered cell morphology and some resistance to the growth in- hibiting effect of medium containing low serum. In Rat-l cells, LMP expression often led to loss of contact inhibition and anchorage-independent growth in soft agar. Rat-l cells expressing LMP were uniformly tu- morigenic in nude mice. Thus, LMP is a transforming gene which is likely to account for many aspects of EBV induced cell transformations. This is the first demonstration of a transforming gene in Epstein-Barr virus, a ubiquitous human pathogen associated with neoplasia.
Introduction
Epstein-Barr Virus (EBV) is believed to cause Burkitt Lym- phoma, nasopharyngeal carcinoma, and lymphomas in immune deficient patients (for reviews see Kieff et al., 1985, and Dambaugh et al., 1986). In vitro, virus infection of human lymphocytes rapidly and efficiently induces sus- tained lymphocyte proliferation (Henle et al., 1967; Pope et al., 1968). Latently infected proliferating lymphocytes will form tumors in the brain of adult nude mice, and EBV induces lymphomas in marmosets (Giovanella et al., 1979; Miller et al., 1977). Latently infected proliferating lympho- cytes and cells of EBV associated malignancies contain complete EBV genomes as an episome or integrated into cell DNA (zur Hausen et al., 1970; Kawai et al., 1973; Nonoyama and Pagano, 1972; Lindahl et al., 1976; Matsuo et al., 1984). In such cells the latent viral genome charac- teristically expresses only four genes that encode proteins (for review see Dambaugh et al., 1986). These four genes encode three nuclear proteins and one membrane protein (Hennessy and Kieff, 1983; 1985; Hennessy et al., 1983; 1984; 1985; Fennewald et al., 1984; Dambaugh et al., 1984). The role of each of these genes in induction and maintenance of cell proliferation is unknown. Since mem- brane proteins encoded by “tumor” viruses play an impor- tant role in cell transformation (for review see Bishop, 1985) we have investigated the role of the EBV membrane protein in this process.
The EBV latent infection membrane protein (LMP) is
translated from an RNA transcribed from the right end of the virus genome (Figure 1; Fennewald et al., 1984; Hen-
nessy et al., 1984). The transcript is 2.6 kb and is spliced to a 2.4 kb mRNA (Fennewald et al., 1984). The RNA has a 1.3 kb open reading frame which begins about 60 nucleotides after the cap site and crosses two introns (Fennewald et al., 1984). From the nucleotide sequence of the open reading frame, LMP was predicted to have six hydrophobic transmembrane domains and a 200 amino acid acidic carboxy terminus. Antibody to the acidic car- boxy domain expressed in bacteria from an EBV DNA- /acZ fusion reacts with a 60 kd protein in the cytoplasmic and plasma membranes of latently infected lymphocytes (Fennewald et al., 1984; Hennessy et al., 1984). A mono- clonal antibody, S12, directed against the bacterial fu- sion protein was used to confirm that LMP is distributed throughout the infected lymphocyte cytoplasmic mem- branes, including the plasma membrane (Mann et al., 1985; Liebowitz et al., 1986). Only a short-reverse turn domain is exposed on the outer cell surface, and the protein patches in lymphocyte plasma membranes (Lie- bowitz et al., 1986). The observation that the LMP patches in the plasma membrane also suggested that it could be part of a complex that might play a direct role in virus in- duction of cell proliferation. To further explore the relation- ship of LMP with the induction of cell proliferation, the ef- fect of the gene on the growth of continuous rodent cell lines has been assayed. Although previous attempts to identify a transforming gene in EBV infected continuous lymphoblastoid cell lines or Burkitt tumors were unsuc- cessful (Diamond et al., 1983), our results indicate that the LMP gene transforms established rodent cell lines.
Results
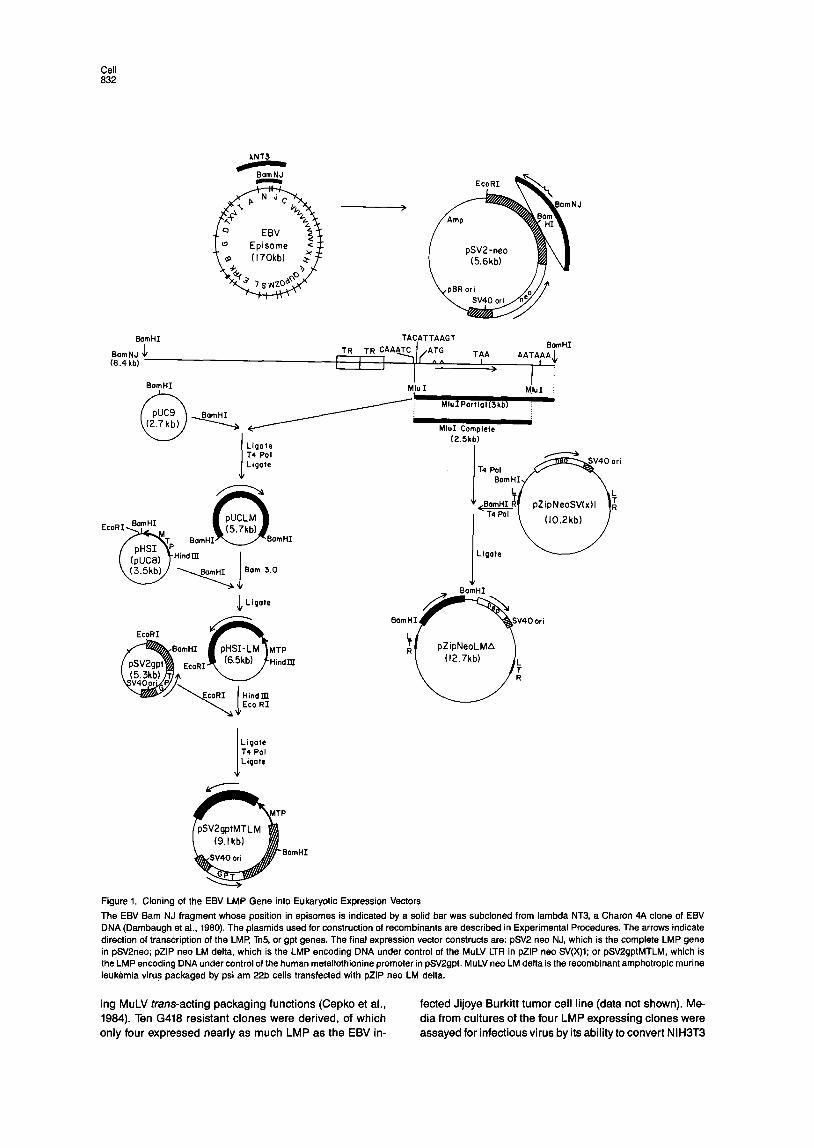
Expression of the LMP in Murine Cells The first approach to expressing LMP in murine cells was to insert an EBV DNA fragment that included 5.6 kb 5’ to the LMP mRNA cap site and 0.3 kb 3’ to its polyadenyla- tion site into the positive selection vector pSV2-neo (Fig- ure 1). Twenty-eight clones of G418 resistant 3T3 cells from three different transfections were selected, but by immu- noblot and immunofluorescence analysis contained no detectable LMP
Two further constructs (Figure 1) were made. One con- struct, pZip neo LM delta, consisted of the LMP gene with- out its polyadenylation site recombined into the cloning site of pZip(neo) SV(X)l, a vector which provides MuLV transcriptional regulation, capping site, 5’ leader with cis- acting site for packaging into MuLV, and 3’ polyadenyla- tion signal (Cepko et al., 1984). The second construct, pSV2gptMTLM, consisted of the human metallothionine promoter and capping site (Karin and Richards, 1982), the LMP gene with its own polyadenylation site, and the vector pSV2gpt. The pZip(neo) SV(X)l and pSV2gpt vectors con- tain genes that permit positive selection of transfected cells in G418 or mycophenolic acid, respectively.
In the next series of experiments, pZip neo LM delta was transfected into psi am 22b, an NIH3T3 cell line express-
Cell 032
ANT3
Q EBV i 0 Episome z 8 (170kb) 2
BarnHI
J BamNJ (8.4 kbl
TACATTAAGT TR TR CAAATC ATG
BomHI
I ,lI ,.,* TAP, AATAAAJ J >
Ml”1 I : My :
MlufPartial(3kb)
Ligate T4 Pal Ligate
MluI Complete (2.5kb)
I Ligate T4 Pal Ligate
Barn”1
Figure 1. Cloning of the EBV LMP Gene into Eukaryotic Expression Vectors
The EBV Barn NJ fragment whose position in episomes is indicated by a solid bar was subcloned from lambda NT3, a Charon 4A clone of EBV DNA (Dambaugh et al., 1980). The plasmids used for construction of recombinants are described in Experimental Procedures. The arrows indicate direction of transcription of the LMP, Tn5, or gpt genes. The final expression vector constructs are: pSV2 neo NJ, which is the complete LMP gene in pSV2neo; pZlP neo LM delta, which is the LMP encoding DNA under control of the MuLV LTR in pZlP neo SV(X)l; or pSVZgptMTLM, which is the LMP encoding DNA under control of the human metallothionine promoter in pSV2gpt. MuLV neo LM delta is the recombinant amphotropic murine leuk(tmia virus packaged by psi am 22b cells transfected with pZlP neo LM delta.
ing MuLV trans-acting packaging functions (Cepko et al., fected Jijoye Burkitt tumor cell line (data not shown). Me- 1984). Ten G418 resistant clones were derived, of which dia from cultures of the four LMP expressing clones were only four expressed nearly as much LMP as the EBV in- assayed for infectiousvirus by its ability to convert NIH3T3
Epstein-Barr Virus Transformation 833
A. lClom 3T3-LM n-
Ey B. RAT-I CLONES 3,c.j ,
Ei ” 2c-J ~ncu-z~~
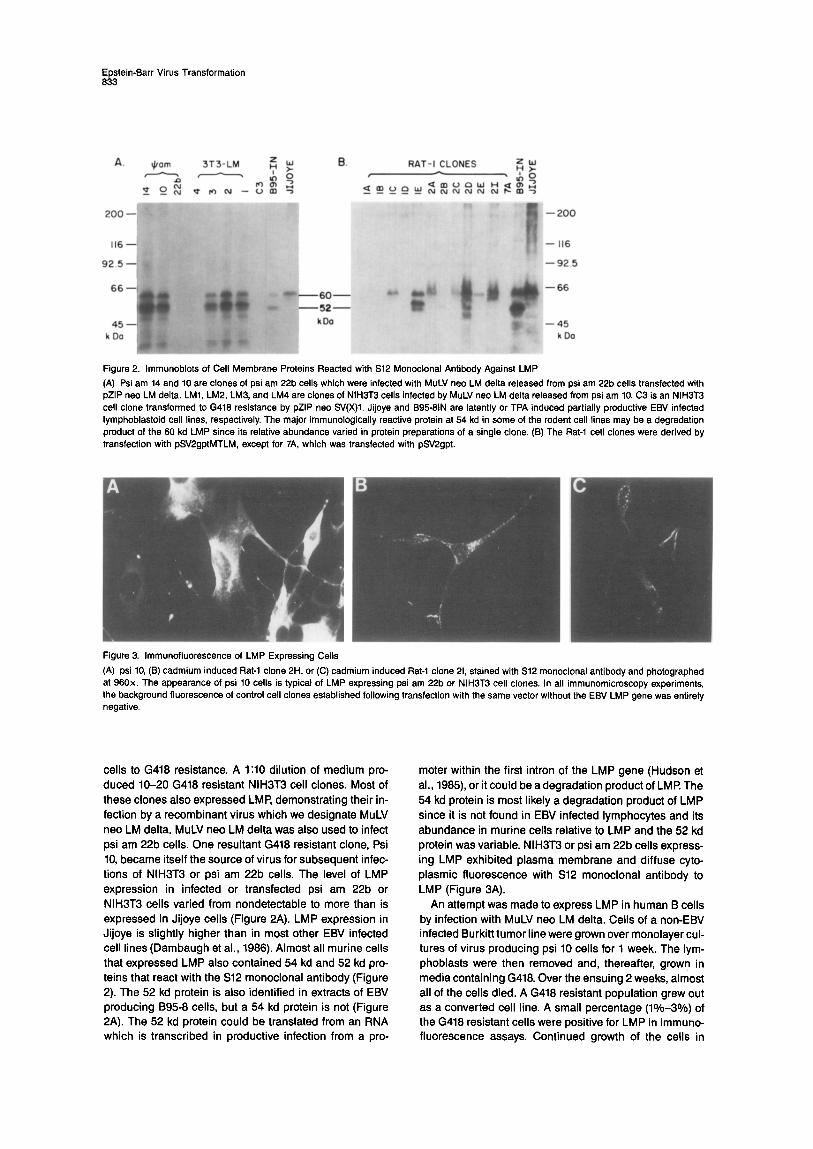
Figure 2. lmmunoblots of Cell Membrane Proteins Reacted with S12 Monoclonal Antibody Against LMP
(A) Psi am 14 and IO are clones of psi am 22b cells which were infected with MuLV neo LM delta released from psi am 22b cells transfected with pZlP neo LM delta. LMl, LM2, LM3, and LM4 are clones of NIH3T3 cells infected by MuLV neo LM delta released from psi am 10. C3 is an NIH3T3 cell clone transformed to G416 resistance by pZlP neo SV(X)l. Jijoye and B95-6lN are latently or TPA induced partially productive EBV infected lymphoblastoid cell lines, respectively. The major immunologically reactive protein at 54 kd in some of the rodent cell lines may be a degradation product of the 60 kd LMP since its relative abundance varied in protein preparations of a single clone. (B) The Rat-l cell clones were derived by transfection with pSV2gptMTLM, except for 7A, which was transfected with pSV2gpt.
Figure 3. lmmunofluorescence of LMP Expressing Cells
(A) psi 10, (B) cadmium induced Rat-l clone 2H, or(C) cadmium induced Rat-l clone 21, stained with S12 monoclonal antibody and photographed at 960x. The appearance of psi IO cells is typical of LMP expressing psi am 22b or NIH3T3 cell clones. In all immunomicroscopy experiments, the background fluorescence of control cell clones established following transfection with the same vector without the EBV LMP gene was entirely negative
cells to G418 resistance. A 1:lO dilution of medium pro- duced lo-20 G418 resistant NIH3T3 cell clones. Most of these clones also expressed LMP, demonstrating their in- fection by a recombinant virus which we designate MuLV neo LM delta. MuLV neo LM delta was also used to infect psi am 22b cells. One resultant G418 resistant clone, Psi 10, became itself the source of virus for subsequent infec- tions of NIH3T3 or psi am 22b cells. The level of LMP expression in infected or transfected psi am 22b or NIH3T3 cells varied from nondetectable to more than is expressed in Jijoye cells (Figure 2A). LMP expression in Jijoye is slightly higher than in most other EBV infected cell lines (Dambaugh et al., 1986). Almost all murine cells that expressed LMP also contained 54 kd and 52 kd pro- teins that react with the S12 monoclonal antibody (Figure 2). The 52 kd protein is also identified in extracts of EBV producing 895-8 cells, but a 54 kd protein is not (Figure 2A). The 52 kd protein could be translated from an RNA which is transcribed in productive infection from a pro-
moter within the first intron of the LMP gene (Hudson et al., 1985) or it could be a degradation product of LMP The 54 kd protein is most likely a degradation product of LMP since it is not found in EBV infected lymphocytes and its abundance in murine cells relative to LMP and the 52 kd protein was variable. NIH3T3 or psi am 22b cells express- ing LMP exhibited plasma membrane and diffuse cyto- plasmic fluorescence with S12 monoclonal antibody to LMP (Figure 3A).
An attempt was made to express LMP in human B cells by infection with MuLV neo LM delta. Cells of a non-EBV infected Burkitt tumor line were grown over monolayer cul- tures of virus producing psi 10 cells for 1 week. The lym- phoblasts were then removed and, thereafter, grown in media containing G418. Over the ensuing 2 weeks, almost all of the cells died. A 6418 resistant population grew out as a converted cell line. A small percentage (1%-30/o) of the G418 resistant cells were positive for LMP in immuno- fluorescence assays. Continued growth of the cells in

Cell 034
3T3 3T3C3 3T3LMI 3T3LM2 3T3LM3 3T3LM4
Figure 4. Dependence of Cell Growth on Serum
NIH3T3 cell clones LMl, LM2, LM3, LM4, and C3 (origins and properties shown in Figure 2) were seeded at the same starting density into medium with 10% serum. After 1 day, cells were switched to medium with lo%, l%, or 0.5% serum in which they were maintained for 2 weeks. The cells were photographed with phase contrast at 50x to show fields that reflect typical cell morphology during the 2 week culture interval and the density of the respective cultures on day 14.
medium containing G418 or recloning by limiting dilution in medium containing G418 resulted in cultures with a similar small fraction of cells expressing LMP
Characteristics of 3T3 Cells Expressing LMP Clones of NIH3T3 ceil that expressed LMP (LMl, LM2, and LM3, see Figure 2A) were longer and thinner than cells not expressing LM!? They grew to a higher cell den- sity than G418 resistant NIH3T3 cells transfected with pZip(neo) SV(X)l. The LMP expressing cells, however, did not heap up (Figure 4). In 1% and 0.5% serum, cells ex- pressing LMP maintained their long thin shape but grew at a reduced rate. Over 14 days in low serum, cells con- tinuously detached from plastic and it took five times as long for the cultures to reach confluence as in 10% serum. Clone LM2, which expressed the most LMP, grew best in low serum and at 14 days reached almost as high a cell density as in 10% serum. Control cells (G418 resistant clones of pZip(neo) SV(X) transfected NIH3T3 cells) or LM4 cells (an MuLV neo LM delta infected clone that did not express detectable amounts of LMP) grew less well than LMP expressing clones in 1% or 0.5% serum, al- though they were equally resistant to G418. In low serum,
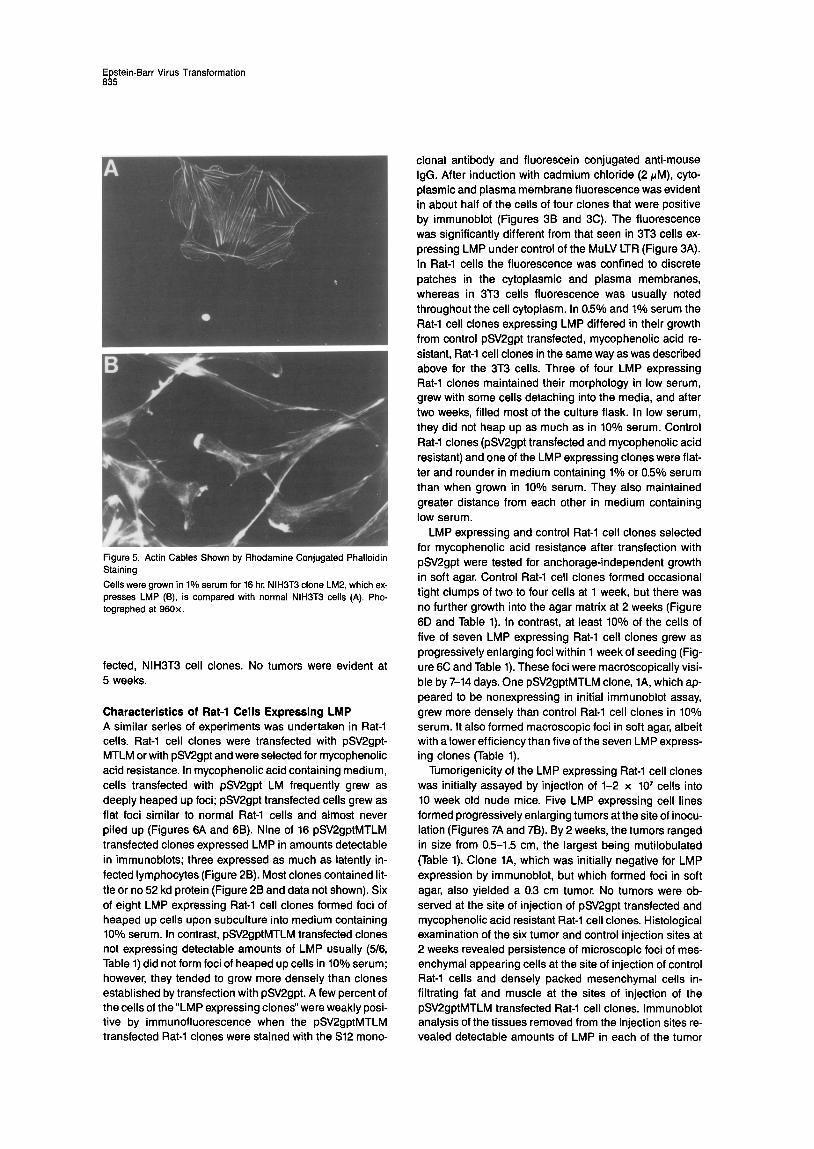
control cells were larger, flatter, and rounder. They re- mained attached to the plastic surface and were well separated from each other even after 14 days (Figure 4). While control cultures developed thick actin cables 12-18 hr after transfer to low serum, cultures of cells expressing LMP did not, indicating a reduced sensitivity to low serum inhibition of cell growth (Figure 5). Neither control nor LMP expressing NIH3T3 cells grew in soft agar, although EJ Ras transfected NIH3T3 cells readily formed macro- scopic colonies in soft agar.
Because we were specifically interested in the proper- ties of the 60 kd LMP clones were derived using pSV2- gptMTLM to ascertain whether this construct would favor the expression of LMP over the 52 kd protein. Most of the resultant clones expressed LMP at levels comparable to the MuLV LTR promoted clones. They contained at least fivefold less or no 52 kd protein, suggesting that this pro- tein is not simply a degradation product of LMP (data not shown). The growth properties of these clones in low se- rum or soft agar were the same as the previously derived MuLV recombinant, LMP expressing, NIH3T3 cell clones. Three week old nude mice were injected with 5 x lo5 cells of each of six LMP expressing, pSV2gptMTLM trans-
Epstein-Barr Virus Transformation 035
Figure 5. Acbn Cables Shown by Rhodamine Conjugated Phalloidin Staining
Cells were grown in 1% serum for 16 hr. NIH3T3 clone LM2, which ex- presses LMP (B), is compared with normal NIH3T3 cells (A). Pho- tographed at 960x.
fected, NIH3T3 cell clones. No tumors were evident at 5 weeks.
Characteristics of Rat-l Cells Expressing LMP A similar series of experiments was undertaken in Rat-l cells. Rat-l cell clones were transfected with pSV2gpt- MTLM or with pSV2gpt and were selected for mycophenolic acid resistance. In mycophenolic acid containing medium, cells transfected with pSV2gpt LM frequently grew as deeply heaped up foci; pSV2gpt transfected cells grew as flat foci similar to normal Rat-l cells and almost never piled up (Figures 6A and 6B). Nine of 16 pSV2gptMTLM transfected clones expressed LMP in amounts detectable in immunoblots; three expressed as much as latently in- fected lymphocytes (Figure 2B). Most clones contained lit- tle or no 52 kd protein (Figure 2B and data not shown). Six of eight LMP expressing Rat-l cell clones formed foci of heaped up cells upon subculture into medium containing 10% serum. In contrast, pSV2gptMTLM transfected clones not expressing detectable amounts of LMP usually (96, Table 1) did not form foci of heaped up cells in 10% serum; however, they tended to grow more densely than clones established by transfection with pSV2gpt. A few percent of the cells of the “LMP expressing clones” were weakly posi- tive by immunofluorescence when the pSV2gptMTLM transfected Rat-l clones were stained with the S12 mono-
clonal antibody and fluorescein conjugated anti-mouse IgG. After induction with cadmium chloride (2 PM), cyto- plasmic and plasma membrane fluorescence was evident in about half of the cells of four clones that were positive by immunoblot (Figures 3B and 3C). The fluorescence was significantly different from that seen in 3T3 cells ex- pressing LMP under control of the MuLV LTR (Figure 3A). In Rat-l cells the fluorescence was confined to discrete patches in the cytoplasmic and plasma membranes, whereas in 3T3 cells fluorescence was usually noted throughout the cell cytoplasm. In 0.5% and 1% serum the Rat-l cell clones expressing LMP differed in their growth from control pSV2gpt transfected, mycophenolic acid re- sistant, Rat-l cell clones in the same way as was described above for the 3T3 cells. Three of four LMP expressing Rat-l clones maintained their morphology in low serum, grew with some cells detaching into the media, and after two weeks, filled most of the culture flask. In low serum, they did not heap up as much as in 100/o serum. Control Rat-l clones (pSV2gpt transfected and mycophenolic acid resistant) and one of the LMP expressing clones were flat- ter and rounder in medium containing 1% or 0.5% serum than when grown in 10% serum. They also maintained greater distance from each other in medium containing low serum.
LMP expressing and control Rat-l cell clones selected for mycophenolic acid resistance after transfection with pSV2gpt were tested for anchorage-independent growth in soft agar. Control Rat-l cell clones formed occasional tight clumps of two to four cells at 1 week, but there was no further growth into the agar matrix at 2 weeks (Figure 6D and Table 1). In contrast, at least 10% of the cells of five of seven LMP expressing Rat-l cell clones grew as progressively enlarging foci within 1 week of seeding (Fig- ure 6C and Table 1). These foci were macroscopically visi- ble by 7-14 days. One pSV2gptMTLM clone, lA, which ap- peared to be nonexpressing in initial immunoblot assay, grew more densely than control Rat-l cell clones in 10% serum. It also formed macroscopic foci in soft agar, albeit with a lower efficiency than five of the seven LMP express- ing clones (Table 1).
Tumorigenicity of the LMP expressing Rat-l cell clones was initially assayed by injection of l-2 x 10’ cells into 10 week old nude mice. Five LMP expressing cell lines formed progressively enlarging tumors at the site of inocu- lation (Figures 7A and 78). By 2 weeks, the tumors ranged in size from 0.5-1.5 cm, the largest being mutilobulated (Table 1). Clone lA, which was initially negative for LMP expression by immunoblot, but which formed foci in soft agar, also yielded a 0.3 cm tumor. No tumors were ob- served at the site of injection of pSV2gpt transfected and mycophenolic acid resistant Rat-l cell clones. Histological examination of the six tumor and control injection sites at 2 weeks revealed persistence of microscopic foci of mes- enchymal appearing cells at the site of injection of control Rat-l cells and densely packed mesenchymal cells in- filtrating fat and muscle at the sites of injection of the pSV2gptMTLM transfected Rat-l cell clones. lmmunoblot analysis of the tissues removed from the injection sites re- vealed detectable amounts of LMP in each of the tumor

Cell 836
Figure 6. Growth Properties of Transformed and Control Rat-l Cells
Comparison of growth in 10% serum (A and B) or in soft agarose (C and D) of Rat-l cell clone 2D which expresses LMP (A, C) versus a Rat-t cell clone 8A transfected with pSV2gpt (B, D). Photographs are with phase contrast at 50x.
Table 1. Characteristics of Rat-l Cell Clones
LMP Expressiona Morphologic Growth in
Tumor Size (cm)
Transformationb Soft Agar c d e
pSV2gptMTLM Clones 1A B C D E G H I 2A B C D E F G H I
pSV2gpt Clones 7A B C 0A B C
+
++
+
+
+ +++ +
+ ++
f
++ ++ *
++ * ++ ++
*
+ 80% + 100%
<5%
50%
60%
10%
0.3’ 0.35 (0.4)
0.5’ 0.2 (1.2)
0.8’
0.8’
0.4 (0.2) 0.5 (0.35)
0.9’ 0.8 (3.2) 0.2 (0.7)
1 .o (1.1) 1.5’ 0.9 (3.3)
0 0 0
o,o 0 0,O.l 0
0
a + + indicates same level as B95-8; + + + indicates higher levels than 895-8; - indicates none detected. b Extent of heaping of cells in 10% serum. f indicates dense growth; + or + + indicate extent of heaping up. c Tumor diameter at 14 days after injecting 2 x 10’ cells. d Tumor diameter at 14 days after injecting lo5 cells. B Volume of tumors shown in column (d) at 5 weeks in ems. r Tumor tissue and cell line established from tumor shown to exoress LMP.

Epstein-Barr Virus Transformation 837
-116
-92.5
-66 -60
-45
Figure 7. Nude Mice Injected with LMP Expressing Rat-l Cell Clones
(A) A mouse injected 2 weeks previously with 2 x IO’ cells of clone 2E on the left and 21 on the right. (6) A mouse injected 5 weeks previously with 10” cells of clone 21 on the back of the neck, clone 2H on the left, and clone 2F on the right. (C) An immunoblot of proteins extracted from the nude mouse tumor generated from clone 2E or from subcutaneous tissue of the control nude mice injected with control Rat-l cells (clone 8A) transfected with pSV2gpt. Similar results were seen with tumor tissue derived from other clones (data not shown).
tissues including the tumor induced by clone lA, whereas control tissues were negative (Figure 7C and data not shown). Cell lines established from each of the tumors also contained detectable amounts of LMP on immunoblot (data not shown). In a second series of experiments, 12 week old nude mice were inoculated with lo5 cells of pSV2gptMTLM transfected clones or with 10’ pSV2gpt transfected Rat-l cell clones. Progressively enlarging solid tumors, ranging from 0.2-l cm in diameter, were evi- dent by 2 weeks at each of the injection sites of LMP ex- pressing cells, while one of the control injection sites had 0.1 cm soft nodule. Almost all of the tumors continued to increase in size over an ensuing 3 weeks of observation. Four out of eight tumors were more than 1 cm3 by 5 weeks. At 5 weeks, the experiment was terminated because of necrosis of the skin overlying the tumors.
Discussion
These data indicate that the EBV LMP gene can transform cells of the established Rat-l line to loss of contact inhibi- tion, anchorage independence, and tumorigenicity in nude mice. LMP also increases the resistance of both NIH3T3 and Rat-l cells to low serum inhibition of cell growth. In its capacity to transform established cells to tumorigenicity, LMP mimics the effect of polyoma virus middle T, adenovirus ElB, SK, or other tyrosine kinase on- cogenes, and ras genes (for review see Bishop, 1985).
Furthermore, EBV LMP is similar to some of these mem- brane “transforming” genes in its affects on cell morphol- ogy, cell growth in low serum, loss of contact inhibition, and anchorage independence (for review see Land et al., 1983). However, the primary amino acid sequence of LMP is not significantly homologous to any of these other pro- teins; nor does LMP have homology to specific active or conserved sites defined in various of these other genes, such as the tyrosine kinase or GTP binding sites (Fen- newald et al., 1984). Because of its lack of homology to these other genes, EBV LMP may affect cell growth by a different mechanism.
Structurally, LMP has six hydrophobic membrane span- ning domains separated by short reverse turns, one of which is exposed on the outer cell surface (Fennewald et al., 1984; Liebowitzet al., 1986). The short amino-terminal and long acidic hydrophilic carboxy-terminal domains of LMP are intracellular. Since so little of LMP is exposed on the outer cell surface, it is unlikely to be a receptor for a growth factor.
The effects of LMP on Rat-l and NIH3T3 cells indicate that LMP is likely to be important in the capacity of EBV to induce and maintain lymphocyte transformation. EBV nuclear protein genes are likely to be necessary comple- ments to LMP in the growth transformation of primary B cells, since EBV isolates deleted for the EBNA2 gene are unable to transform B lymphocytes (Dambaugh et al., 1984; Hennessy et al., 1985). LMP may also interact with

Cell 038
c-myc in the evolution of the African Burkitt tumor cell (for review see Leder, 1983). Nuclear proteins of other onco- genie DNA viruses, c-myc and ~53, have been noted to play important roles in establishing the immortality of pri- mary cells in long term culture (for review see Land et al., 1983; Bishop, 1985; and Jenkins et al., 1984). EBV as- sociated human malignancies like most other malignan- cies probably result from a multistep process. Studies are in progress to consider the interaction of LMP with EBV nuclear and cellular proteins in the processes of primary cell transformation.
Further characterization of the biological and chemical effects of LMP in Rat-l and NIH3T3 cells will probably be relevant to understanding LMP effects in EBV induced cell proliferation. Notably, LMP expression in some of the pSVgptMTLM transfected Rat-l and NIH3T3 cells is simi- lar to that in EBV immortalized lymphocytes. Also, im- munomicroscopy indicates that LMP patches in the plas- ma membrane of Rat-l cells as in the plasma membrane of EBV transformed lymphocytes (Liebowitz et al., 1986). Patching could be important in the biologic effect of LMP since LMP could interact with a growth factor receptor, and growth factors are essential to the continued proliferation of EBV transformed lymphocytes (Gordon et al., 1984; 1985).
The differences in the effects of LMP expression on the properties of Rat-l and NIH3T3 cells are surprising since both are established rodent cell lines. The more pro- nounced effect of LMP on Rat-l cells could be due to for- tuitous complementation with the intrinsically active im- mortalizing gene (or genes) of the Rat-l cell line or it could be due to other differences between Rat-l and NIH3T3 lines. Analysis of the biochemical interactions of LMP in Rat-l and NIH3T3 cells may reveal differences that can be associated with the different biologic effect.
Experimental Procedures
Ceil Culture, Transfections, Selection, and Cloning Louckes, an EBV negative American Burkitt tumor ceil line, Jijoye, an EBV positive African Burkitt tumor cell line, and 184, an in vitro trans- formed latently EBV infected ceil line, were maintained in RPM1 1640 medium supplemented with 10% newborn calf serum (Dambaugh et al., 1966). NIH3T3 cells were obtained from Bob Lowe (Merck, Sharp, and Dhome Research Laboratories, West Point, Pennsylvania) at their eleventh passage and were used for transfection between passages 13 and 15. Rat-l ceils were obtained from Mitchel Villereal (Department of Physiology, The University of Chicago) at their third passage after receipt from Robert Weinberg (Whitehead Foundation, MIT). Rat-l cells were used for transfection between passages three and five in our laboratory. Psi am 22b NIH3T3 cells were obtained from R. Mulligan (Whitehead Institute, MIT) at passage five and were used for transfec- tion at passage eight. NIH3T3 or Rat-l cells were split I:30 twice a week so that they did not reach confluence and were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM, Flow Laboratories) sup- plemented with 10% heat inactivated fetal calf serum (K. C. Biological) and 2 pglmi of gentamicin sulfate. Ceils at 50% confluence in T25 flasks were transfected with l-10 pg of plasmid DNA and 10 rg of salmon sperm DNA by the calcium phosphate technique (Graham and van der Eb, 1973). Twenty-four hours later, cells were split into T75 flasks and placed under selection. Selective media contained 400 &ml of G418 or 15 rglml of mycophenolic acid, 160 pglml xanthine, and 10 pglml hypoxanthine. Characteristically IO-100 clones emerged in each T75 flask. Only six or seven well isolated clones were derived from each flask for further study. Psi am 22b transfected clones were main-
tained under P2 plus biohazard safety conditions. For assay of psi am 22b transfected clones for virus, culture medium from ceils growing in log phase was aspirated, filtered through a 0.45 micron filter (Nalge), and overlayed onto cell cultures 24 hr after seeding. Twenty-four hours later cells were placed under selection. Louckes cells were exposed to virus by growing them in T150 flasks in complete lymphocyte growth medium with 5 bglml polybrene (Sigma Chemical Corp) over mono- layers of psi 10 cells, a clone of virus producing cells. Louckes cells were thereafter maintained in G418 at 1.5 mglml. Cells were cloned by limiting dilution in 0.5 mglml G416.
Piasmid Constructions The EBV DNA Barn HI fragment, from which all constructs were made (see Figure I), is a segment of episomal EBV DNA which extends from the last Barn HI site at the right end of the linear EBV genome to the first Barn HI site at the left end. The fragment was subcloned by T. Dambaugh from lambda NT3, which is a clone of a partial Eco RI digest of episomal EBV DNA in charon 4A (Dambaugh et al., 1980). The strategy for inserting the Barn HI fragment into pSV2-neo vector (Mulli- gan and Berg, 1961; Southern and Berg, 1961; obtained from American Type Culture Collection) and for subsequent cloning into pZip neo SV(X)l (obtained from R. Mulligan, Whitehead institute, MIT) or for recombination with the human metallothionine promoter (obtained from M. Karin, University of Southern California) and insertion into pSV2gpt (obtained from ATCC) are shown in Figure 1. Piasmids were transformed into DHI and identified by colony hybridization and mini preparation to have each of the restriction endonuclease sites and orientations shown in Figure 1. Plasmid DNAs were then prepared in bulk for transformations. With reference to Figure 1, it should be noted that ligation of a T4 DNA polymerase treated Mlu I fragment to a T4 DNA polymerase treated Barn HI fragment results in regeneration of Barn HI sites. initially pSV2gptMTLM was constructed both as shown in Figure 1 or with the metailothionine promoter-LMP segment tran- scribing clockwise. The latter construct rarely yielded mycophenolic acid resistant clones which expressed LMP
Assays for Growth Properties For studies of serum or plasma dependence, 1 x IO5 cells were seeded in a T25 flask (Corning Corp.) in DMEM with 10% fetal calf se- rum for 24 hr. The cultures were then transferred to DMEM with IO%, l%, or 0.5% serum. The cells were observed daily and fed every 4 days until 2 weeks elapsed with fresh medium supplemented with the appro- priate serum concentration.
Anchorage dependence of cell growth was assayed in 0.3% agarose in medium with 10% fetal calf serum. One milliliter of 0.6% agarose (Seaplaque, FMC) in phosphate buffered saline (PBS) was layered onto a 60 mm plastic petri dish (Falcon). 5 x 1O’well trypsinized cells in 3 ml of complete medium were mixed with 3 ml of 0.6% agarose in complete medium and layered over the 0.6% agarose. Cultures were fed with two drops of complete medium twice a week.
Actin cable formation in low serum was assayed by seeding 2 x lo3 cells per well in an eight well chamber slide (Miles) in DMEM with 10% fetal calf serum. After 24 hr the cells were switched to DMEM with 1% fetal calf serum. Sixteen hours later, the slide was rinsed in PBS, fixed at 25OC in 10% formalin in PBS (pH Z5) for 20 min, rinsed with PBS, and treated with 1% NP40 in PBS for 20 min at 25OC. The slides were then incubated with rhodamine conjugated phailoidin for 30 min at 3pC, rinsed in PBS, and observed through a rhodamine filter (Verder- ame et al., 1980).
immunofiuorescence and lmmunobiots For immunofluorescence, cells on slides or lymphocytes in suspension were washed in PBS, dried on slides rapidly at 3pC, fixed in methanol at -2pC for 6 min, washed 2x in PBS with 10% goat serum, in- cubated with S12 monocional antibody ascites fluid diluted 1 :lOO in 5% goat serum and PBS for 1.5 hr at 25OC, washed with PBS, incubated with a 1:lOO dilution of biotin conjugated goat anti-mouse IgG (BRL) for 1 hr at 25“C, and incubated with FITC conjugated strepavidin (BRL) for 30 min at 37oC. Cells were photographed at 960x magnification.
For immunoblot analysis, cells were scraped into cold PBS, cen- trifuged to a pellet, resuspended for 10 min in 10 mM Tris HCI (pH 7.5) with 1 mM PMSF, and homogenized with a tight fitting dounce. Nuclei were removed by centrifugation for 5 min at 4OC and 1000 rpm. Mem-

Epstein-Barr Virus Transformation 839
branes were centrifuged to a pellet at 35,000 rpm for 35 min at 4OC in an SW41 rotor. The membrane pellets were resuspended in 0.5% SDS and equal amounts of protein were run on a 7% SDS-polyacrylamide gel. The gel was transferred to nitrocellulose for immunostaining with S12 monoclonal, rabbit anti-mouse IgG (Cappel) and jz51 protein A (Amersham Corp.).
For immunoblots of nude mouse tumors, a small piece of the tumor, or subcutaneous tissue from the site of injection of control Rat-l cells that did not cause tumors, was placed in 50 to 100 ~1 SDS sample buffer. The samples were subjected to three freeze-thaw cycles and then boiled for 5 min. Supernatants were run on a 7% SDS-polyacryl- amide gel and analyzed as above.
Nude Mouse Injections Freshly trypsinized cells (1-2 x IO7 or 1 x 105) were injected into the subcutaneous tissue of 10 or 12 week old nude mice. Mice were ob- served every other day and sacrificed at 14 days.
Acknowledgments
This investigation was dependent on advice or materials from Timothy Dambaugh (cloning of EBV Barn NJ and early transfection experi- ments), Richard Mulligan (pZip neo SV(X)l and psi am 22b), Michael Karin (clone of human metallothionine promoter and its sequence), Young Whang (assistance in cloning), Thomas Benjamin and Hides- aburo Hanafusa (advice), Ellen Woodland and Hans Schreiber (nude mice), Bob Pollack (rhodamine conjugated phalloiden), Karen Mann and David Thorley-Lawson (S12 monoclonal antibody), and Robert Weinberg and Louis Parada (Rat-l cells and their properties).
This research was supported by grants CA 17281 and CA 19264 from the USPHS National Institutes of Health and ACS MV32J from the American Cancer Society. D. W. is a predoctoral trainee supported by IIE 15822181 from the Institute of International Education, D. L. is a predoctoral trainee supported by grant GM 07281 from the National In- stitutes of Health, and E. K. was an American Cancer Society Faculty Research Awardee.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received September 5, 1985; revised October 10, 1985
References
Bishop, M. J. (1985). Viral Oncogenes. Cell 4.2, 23-38.
Cepko, C. L., Roberts, B. E., and Mulligan, R. C. (1984). Construction and applications of a highly transmissible murine retrovirus shuttle vector. Cell 37, 1053-1062.
Dambaugh, T., Beisel, C., Hummel, M., King, W., Fennewald, S., Cheung, A., Heller, M., RaabTraub, N., and Kieff, E. (1980). Epstein- Barr virus DNA. VII. Molecular cloning and detailed mapping of EBV(B95-8) DNA. Proc. Natl. Acad. Sci. USA 77, 2999-3003.
Dambaugh, T, Hennessy, K., Chamnankit, L., and Kieff, E. (1984). The U2 region of Epstein-Barr virus DNA may encode EBNAP. Proc. Natl. Acad. Sci. USA 81, 7207-7211.
Dambaugh, T., Hennessy, K., Fennewald, S., and Kieff, E. (1986). The EBV genome and its expression in latent infection. In The Epstein-Barr Virus: Recent Advances, M. A. Epstein and B. G. Achong, eds. (Lon- don: William Heinemann Medical Books LTD), in press,
Diamond, A., Cooper, G. M., Ritz, J., and Lane, M.-A., (1983). Identifi- cation and molecular cloning of the human Blym transforming gene ac- tivated in Burkitt’s lymphomas. Nature 305, 112-116.
Fennewald, S., Van Santen, V., and Kieff, E. (1984). Nucleotide se- quence of an mRNA transcribed in latent growth-transforming virus in- fection indicates that it may encode a membrane protein. J. Virol. 57, 411-419.
Giovanella, B., Nilsson, K., Zech, L., Yim, O., Klein, G., and Stehlin, J. S. (1979). Growth of diploid, Epstein-Barr virus-carrying human lym- phoblastoid cell lines heterotransplanted into nude mice under im- munologically privileged conditions. Int. J. Cancer 24, 103-113.
Gordon, J., Levy, S., Melamed, M., England, L., and Hughes-Jones, N. (1984). Immortalized B lymphocytes produce B cell growth factor. Nature 310, 145-147.
Gordon, Q., Arnan, P., Rosen, A., Ernberg, I., Ehlin-Henriksson, B., and Klein, G. (1985). Capacity of B-lymphocyte lines of diverse tumor origin to produce and respond to B-cell growth factors: a progression model for B-cell lymphomagenesis. Int. J. Cancer 35, 251-256.
Graham, F. L., and van der Eb, A. J. (1973). A new technique for the assay for infectivity of human adenovirus DNA. Virology 52, 456-467
Henle, W., Diehl, B., Kohn, G., zur Hausen, H., and Henle, G. (1967). Herpes type virus and chromosome marker in normal leukocytes after growth with irradiated Burkitt cells. Science 757. 10641065.
Hennessy, K., and Kieff, E. (1983). One of two Epstein-Barr virus nu- clear antigens contains a glycine-alanine copolymer domain. Proc. Natl. Acad. Sci. USA 80, 5665-5689.
Hennessy, K., and Kieff, E. (1985). A second nuclear protein is en- coded by Epstein-Barr virus in latent infection. Science 227, 1238-1240.
Hennessy, K., Heller, M., van Santen, V., and Kieff, E. (1983). A simple repeat array in Epstein-Barr virus DNA encodes part of EBNA. Science 220, 1396-1398.
Hennessy, K., Fennewald, S., Hummel, M., Cole, T., and Kieff, E. (1984). A membrane protein encoded by Epstein-Barr virus in latent growth transforming infection. Proc. Natl. Acad. Sci. USA 87, 7207-7211.
Hennessy, K., Fennewald, S., and Kieff, E. (1985). A third nuclear pro- tein in lymphoblasts immortalized by Epstein-Barr virus. Proc. Natl. Acad. Sci. USA 82, 5944-5948.
Hudson, G., Farrell, P., and Barrell, B. (1985). Two related but differen- tially expressed potential membrane proteins encoded by the EcoRl D het region of Epstein-Barr virus 895-8. J. Virol. 53, 528-535.
Jenkins, J., Rudge, K., and Currie, G. (1984). Cellular immortalization by a cDNA clone encoding the transformation-associated phos- phoprotein P53. Nature 372, 651-654.
Karin, M., and Richards, R. I. (1982). Human metallothionine genes- primary structure of the metallothionine-II gene and a related processed gene. Nature 299, 797-802.
Kawai, Y., Nonoyama, M., and Pagano, J. (1973). Reassociation ki- netics for EBV DNA. Non-homology to mammalian DNA and homology to viral DNA in various diseases. J. Virol. 12, 1006-1012.
Kieff, E., Hennessy, K., Dambaugh, T, Matsuo, T., Fennewald, S., Heller, M., Petti, L., and Hummel, M. (1985). Persistence and expres- sion of the Epstein-Barr virus genome in latent infection and growth transformation of lymphocytes. In Epstein-Barr Virus and Associated Diseases, P Levine, G. Pearson, and D. Ablashi, eds. (Boston: Marti- nus Nijhoff Publishing), pp. 221-247.
Land, H., Parada, L. F., and Weinberg, R. A. (1983). Cellular on- cogenes and multistep carcinogenesis. Science 222, 771-778.
Leder, P (1983). Translocation among antibody genes in human can- cer. Science 222, 765-771.
Liebowitz, D., Wang, D., and Kieff, E. (1986). Orientation and patching of the latent infection membrane protein encoded by Epstein-Barr vi- rus. J. Viral., in press,
Lindahl, T., Adams, A., Bjursell, G., Bornkamm, G. W., Kascha-Dierich, G., and Jehn, U. (1976). Covalently closed circular duplex DNA of Epstein-Barr virus in a human lymphoid cell line. J. Mol. Biol. 702, 51 l-530.
Mann, K. t?, Staunton, D., and Thorley-Lawson, D. A. (1985). Epstein- Barr virus encoded protein in the plasma membranes of transformed cells. J. Virol. 55, 7lO-720.
Matsuo, T, Heller, M., Petti, L., O’Shiro, E., and Kieff, E. (1984). The entire Epstein-Barr virus genome integrates into human lymphocyte DNA. Science 226, 1322-1325.
Miller, G., Shope, T., Coope, D., Waters, L., Pagano, J., Bornkamm, G., and Henle, W. (1977). Lymphoma in cotton-top marmosets after inocu- lation with Epstein-Barr virus: tumor incidence, histologic spectrum, antibody responses, demonstration of viral DNA, and characterization of viruses. J. Exp. Med. 745, 948-967

Cell 040
Mulligan, Ft. C., and Berg, P (1981). Factors governing the expression of a bacterial gene in mammalian cells. Mol. Cell. Biol. 7,. 854-864.
Nonoyama, M., and Pagano, J. (1972). Separation of Epstein-Barr virus DNA from large chromosomal DNA in non-virus producing cells. Na- ture New Biol. 238, 169-171.
Pope, J., Horne, M., and Scott, W. (1968). Transformation of fetal hu- man leukocytes in vitro by filtrates of a human leukemic cell line con- taining herpes-like viruses. Int. J. Cancer 3, 857-866.
Southern, P J., and Berg, P (1981). Transformation of mammalian cells to antibiotic resistance with a bacterial gene under control of the SV40 early region promotor. J. Mol. Appl. Genet. 7, 327-341.
Verderame, M., Alcorta, D., Egnor, M., Smith, K., and Pollack, Ft. (1980). Cytoskeletal fraction patterns quantitated with fluorescein isothiocyanate-phalloidin in normal and transformed cells. Proc. Natl. Acad. Sci. USA 77 6624-6628.
zur Hausen, H., Schulte-Holthausen, H., Klein, G., Henle, W., Henle, G., Clifford, P, and Santesson, L. (1970). EBV DNA in biopsies of Bur- kitt’s tumors and anaplastic carcinomas of the nasopharynx. Nature 228, 1056-1058.




















