
Tumorigenic Heterogeneity in Cancer Stem Cells Evolved from Long-term Cultures of...
11
2005;65:3126-3135. Cancer Res Jorge S. Burns, Basem M. Abdallah, Per Guldberg, et al. Mesenchymal Stem Cells Human from Long-term Cultures of Telomerase-Immortalized Tumorigenic Heterogeneity in Cancer Stem Cells Evolved Updated version http://cancerres.aacrjournals.org/content/65/8/3126 Access the most recent version of this article at: Cited Articles http://cancerres.aacrjournals.org/content/65/8/3126.full.html#ref-list-1 This article cites by 67 articles, 19 of which you can access for free at: Citing articles http://cancerres.aacrjournals.org/content/65/8/3126.full.html#related-urls This article has been cited by 5 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on February 7, 2014. © 2005 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on February 7, 2014. © 2005 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Tumorigenic Heterogeneity in Cancer Stem Cells Evolved from Long-term Cultures of...

2005;65:3126-3135. Cancer Res Jorge S. Burns, Basem M. Abdallah, Per Guldberg, et al. Mesenchymal Stem Cells
Humanfrom Long-term Cultures of Telomerase-Immortalized Tumorigenic Heterogeneity in Cancer Stem Cells Evolved
Updated version
http://cancerres.aacrjournals.org/content/65/8/3126
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/65/8/3126.full.html#ref-list-1
This article cites by 67 articles, 19 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/65/8/3126.full.html#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Tumorigenic Heterogeneity in Cancer Stem Cells Evolved from
Long-term Cultures of Telomerase-Immortalized Human
Mesenchymal Stem Cells
Jorge S. Burns,1Basem M. Abdallah,
1Per Guldberg,
3Jørgen Rygaard,
4
Henrik D. Schrøder,2and Moustapha Kassem
1
1Department of Endocrinology and Metabolism and 2Institute of Pathology, Odense University Hospital, Odense, Denmark; 3Institute ofCancer Biology, Danish Cancer Society; and 4Bartholin Instituttet, Kommunehospitalet, Copenhagen, Denmark
Abstract
Long-term cultures of telomerase-transduced adult humanmesenchymal stem cells (hMSC) may evolve spontaneousgenetic changes leading to tumorigenicity in immunodefi-cient mice (e.g., hMSC-TERT20). We wished to clarify whetherthis unusual phenotype reflected a rare but dominantsubpopulation or if the stem cell origin allowed most cellsto behave as cancer stem cells. Cultures of the hMSC-TERT20strain at population doubling 440 were highly clonogenic(94%). From 110 single-cell clones expanded by 20 populationdoublings, 6 underwent detailed comparison. Like theparental population, each clone had ���1.2 days doublingtime with loss of contact inhibition. All retained 1,25-(OH)2vitamin D3–induced expression of osteoblastic markers:collagen type I, alkaline phosphatase, and osteocalcin. Allshared INK4a/ARF gene locus deletion and epigeneticsilencing of the DBCCR1 tumor suppressor gene. Despitein vitro commonality, only four of six clones shared thegrowth kinetics and 100% tumorigenicity of the parentalpopulation. In contrast, one clone consistently formed latenttumors and the other established tumors with only 30%penetrance. Changing the in vitro microenvironment tomimic in vivo growth aspects revealed concordant clonalheterogeneity. Latent tumor growth correlated with extra-cellular matrix entrapment of multicellular spheroids andhigh procollagen type III expression. Poor tumorigenicitycorrelated with in vitro serum dependence and high p27Kip1
expression. Aggressive tumorigenicity correlated with goodviability plus capillary morphogenesis on serum starvationand high cyclin D1 expression. Thus, hMSC-TERT20 clonesrepresent cancer stem cells with hierarchical tumorigenicity,providing new models to explore the stem cell hypothesis forcancer. (Cancer Res 2005; 65(8): 3126-35)
Introduction
Bone marrow–derived adult human mesenchymal stem cells(hMSC) can generate specialized mesodermal bone, cartilage,muscle, and adipose tissue, and this multipotency may extend topluripotency in specific circumstances (1). However, hMSCs lacktelomerase activity in vitro (2), with a limited proliferative life spanthat decreases with age (3). Overcoming this limitation, retroviral
vector-mediated telomerase reverse transcriptase expression inprimary hMSCs (hMSC-TERT20) greatly extends their in vitroproliferative life span and also improves their osteogenicdifferentiation in vivo (2, 4). Similarly, endogenous telomeraseactivity has been considered to be largely responsible for theextended proliferative capacity of cultured embryonic stem cells(5). However, enhanced telomerase expression is often a feature ofmany tumors, and unlike adult stem cells, embryonic stem cellscan spontaneously form teratomas. Initial studies addressingwhether telomerase-immortalized human somatic cells developedcancer-associated changes concluded they did not (6, 7). Nonethe-less, this has been challenged by spontaneous alterations in c-mycproto-oncogene expression (8) and/or other genetic/epigeneticchanges in long-term cultures (9–11) that can lead to in vitrotransformation to contact inhibition and anchorage independence(12, 13). That a highly expressed telomerase gene may have‘‘extracurricular’’ activity beyond maintaining telomere length(14, 15) is consistent with evidence that it enhances cellproliferation (16). However, only recently have spontaneouschanges in a telomerase-immortalized human cell strain sufficedfor tumorigenesis (17). To clarify whether this merely reflected arare but dominant subpopulation, the tumorigenic hMSC-TERT20strain was single cell cloned by limiting dilution. From 110examples, 6 expanded clones were compared with the parentalpopulation with respect to morphology, telomerase expression,previously described genetic and epigenetic changes, osteoblasticdifferentiation potential, saturation density, contact inhibition,tumorigenicity, and expression of prognostic markers. Initially, wefound a discrepancy between common in vitro characteristics andclone-specific differences in tumorigenicity.However, monolayer culture does not explore all aspects that
may govern tumorigenicity. These include (a) host-tissue inter-actions that nourish and support expansion of the tumorpopulation and (b) growth in three-dimensional clusters. Bloodsupply is essential for nourishing tumor growth, and rapidlygrowing solid tumors are often characterized by areas of poor flowand hypoxia with a metabolic environment that compromisesgrowth until there is rescue by vascularization (18). A tumor cell’sresistance to nutrient starvation may influence tumorigenicity, andexogenous growth factor dependence can discriminate tumorigenicfrom nontumorigenic human cancer sublines (19). Thus, we testedgrowth and survival of the hMSC-TERT20 clones in serum-deprivedconditions.With respect to cellular growth in three-dimensional clusters,
tissue architecture can inhibit tumor growth with build-up of solidstress restricting changes in cell geometry needed for proliferation(20). Tumor-derived matrix metalloproteinases (MMP) can breakdown and relax the extracellular matrix (ECM) of surrounding
Requests for reprints: Jorge S. Burns, Laboratory for Molecular Endocrinology,KMEB, Department of Endocrinology and Metabolism, Odense University Hospital,Medical Biotechnology Center, Winsløwparken 25, DK-5000 Odense C, Denmark.Phone: 45-6550-4081; Fax: 45-6550-3950; E-mail: [email protected].
I2005 American Association for Cancer Research.
Cancer Res 2005; 65: (8). April 15, 2005 3126 www.aacrjournals.org
Research Article
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

tissue, aiding early as well as late stages of tumor growth (21).Recent reviews of this ‘‘tumor jailbreak’’ phenotype (22) highlightedmembrane-bound MMP-1 (MT1-MMP) as the principle regulator ofpericellular proteolysis, allowing tumor cells to breach host tissuebarriers (23, 24). Changing the in vitro microenvironment of tumorcell lines to growth as multicellular spheroids (MCS) may revealdifferences in proliferation rate not observed in monolayer culture(25). Thus, we extended our in vitro analysis to include three-dimensional culture of MCS. These altered microenvironmentsrevealed clone-specific differences that correlated with thesignificant differences in tumorigenic phenotype.
Materials and Methods
Cell culture and derivation of clones. The hMSC-TERT20 strain was
initiated by increasing the split ratio of continually passaged hMSC-TERT20
cells to 1:20 (17). hMSC-TERT20 parental cells and derived clones weregrown in phenol red–free MEM (Gibco Invitrogen Co., Tastrup, Denmark)
supplemented with 10% fetal bovine serum (FBS; Gibco Invitrogen Co.,
batch tested) and antibiotics. Cells were maintained in a humidified
incubator at 37jC and 5% CO2 and detached when near confluent byincubation with 0.25% trypsin in 0.1% EDTA (Gibco Invitrogen Co.). Seeding
density (1:20) was 1,250 cells/cm2 in T80 flasks; vessels were from Nunc
(Roskilde, Denmark) unless stated otherwise. For limiting dilution cloning,
hMSC-TERT20 population doubling (PD) 440 cells were seeded in 100 ALmedium at 0.3 per well in four flat-bottomed 96-well plates. Optical
microscopy, within 12 hours of seeding, identified wells with only one cell.
Subsequent monitoring confirmed growth of only a single colony per well,
and medium was changed weekly. Clones at �60% confluence werepassaged into 24-well plates and refed twice weekly with 1 mL medium.
Clonogenicity was determined by the ability to grow to at least PD 20 (>106
cells) when subsequently passaged in T25 flasks. Six arbitrarily selectedclones showing morphologic uniformity were expanded for detailed
characterization. The human fibrosarcoma cell line HT-1080 (26) was
grown under the same conditions as hMSC-TERT20 cells. Telomerase-
immortalized microvascular endothelial (TIME) cells (27), a gift from Prof.McMahon (University of California at San Francisco, San Francisco, CA),
were grown in supplemented ECBM-MV2 medium (PromoCell GmbH,
Heidelberg, Germany).
Telomerase activity: real-time quantitative Telomeric RepeatAmplification Protocol assay. The SYBR Green Real-time Quantitative
Telomeric Repeat Amplification Protocol (TRAP) assay was a modified
version of the method by Wege et al. (28). Briefly, cell pellet extracts wereprepared using a 1� CHAPS lysis method (29). Real-time PCR amplification
of serial dilutions (1,000, 100, 10, and 1 cell equivalents, including heat-
inactivated samples) ensured assay specificity and linearity.
Proliferation rate. Cells from subconfluent cultures were detached bytrypsin and counted using a hemocytometer. PD was calculated using the
formula: PD = log [(n cells in) / (n cells out)] / log 2. PD times were
calculated from the average of three consecutive passages.
Saturation density and contact inhibition. Cells were seeded at 0.53 �105/cm2 in 12-well plates and grown to confluence. Triplicate wells were
harvested using trypsin and cells counted in a hemocytometer to determine
saturation density. Parallel cultures were fed every 3 days and counted at10 days postconfluence. Results represent the average of three separate
experiments.
Flow cytometry. Cells harvested using trypsin with 0.1% EDTA were
centrifuged at 200 � g for 5 minutes at 4jC. Cell pellets resuspended in ice-cold PBS� were fixed in ice-cold 96% ethanol. The nuclei were released
using a 0.04% pepsin solution (Sigma-Aldrich A/S, Brøndby, Denmark) in
0.1 mol/L HCl, and RNA was removed with 10 Ag/mL RNase (Qiagen, VWR
International A/S, Albertslund, Denmark) in PBS/TB [PBS + 0.5% (v/v)Tween 20 (Merck, VWR International A/S) + 0.1% (w/v) bovine serum
albumin]. The nuclei were spun down and resuspended in 50 Ag/mL
propidium iodide (Sigma-Aldrich A/S) in PBS/TB and incubated overnight
at 4jC. Samples were filtered on a 50 Am nylon mesh before measuring
10,000 to 20,000 nuclei on a FACScan flow cytometer (BD Biosciences,Brøndby, Denmark) using a 488 nm excitation argon laser with >695 fluo-
rescence emission.
Mutation and deletion analysis: denaturing gradient gel electro-phoresis. Genomic DNA from 106 cells was isolated using a Purescript DNAIsolation Kit (Gentra Systems Inc., Biotech Line AS, Slangerup, Denmark).
TP53 gene mutations were explored using a combination of PCR and
denaturing gradient gel electrophoresis. The 12 sets of oligonucleotide
primer sequences and conditions for analyzing the entire coding region andall splice junctions were as described (30). For deletion analysis of the
INK4a/ARF locus, primers and PCR conditions were as described (31).
Epigenetic analysis: methylation-specific PCR. The methylation status
of the DBCCR1 promoter CpG island (Genbank accession no. AF027734)
was examined by methylation-specific PCR (32). Genomic DNA was treated
with sodium bisulfite (33), and primer sequences and conditions for
methylation-specific PCR analysis of the DBCCR1 gene promoter were as
described (34). Amplification products were resolved in a 2% agarose gel.
Anchorage-independent growth (soft agar colony assay). Soft agar
assays were as described (6) with minor modifications. Briefly, 2 � 104 cells
were plated in triplicate 35 mm wells of six-well plates in 2 mL medium
with 0.36% low gelling temperature agarose (Sigma-Aldrich A/S) plus 10%
FBS on a 4 mL base layer of 0.9% agarose in medium. Cells were fed
additional medium, 10% FBS (200 AL) weekly. After 2 to 6 weeks, colonies
>40 cells were counted under an optical microscope. HT-1080 cells were a
positive control.
Osteoblastic differentiation studies. Cells seeded at 4.2 � 104/cm2 insix-well plates were grown in standard medium for 24 hours. At 50% to
60% confluence, medium was replaced with control medium (MEM with
10% FBS) or medium with osteoblastic inductors, 1,25-(OH)2 vitamin D3,L-ascorbic acid and h-glycerophosphate as described (35) for 4 days. Total
RNA was isolated with TRIzol (Invitrogen A/S, Tastrup, Denmark) according
to manufacturer’s instructions. Expression of collagen type 1a, alkaline
phosphatase, and osteocalcin relative to h-actin mRNA expression wasanalyzed from two confluent wells per sample by real-time PCR. Methods
determining the integrity and purity of total RNA, conditions for first-strand
cDNA synthesis and real-time PCR, and software for generating relative
expression values were as described (35).Xenograft tumorigenicity. Immunodeficient mice (NMRI nu/nu) were
maintained in pathogen-free conditions. Cells (5 � 106) at passage 6 were
mixed with Matrigel 1:1 before implantation (100 AL) to facilitate
establishment (6). Perpendicular diameters of emerging tumors were
measured with digital calipers every 2 days to estimate tumor volume
until the xenografts reached 1 to 1.5 cm3 or termination of the study.
HT-1080 cells were a positive control. Tissue samples fixed in 4%
formaldehyde-0.075 mol/L NaPO4 (pH 7), dehydrated, embedded in paraffin,
and sectioned at 4 Am were processed for histologic and immunohisto-
chemical evaluation. H&E staining determined histologic structure.Serum dependence. Cells were seeded at 2 � 105 per well in 24-well
plates in medium (10% FBS). After overnight attachment, the wells were
washed two times with PBS� before applying low-serum (0.1% FBS) or
serum-free medium. Proliferation and viability was monitored for 10 days byharvesting triplicate wells and counting with a hemocytometer using trypan
blue dye exclusion to define viable cells.
Spheroid culture and immunocytochemistry. MCS cultures formedafter seeding 106 cells on ultralow adhesion tissue culture dishes (Corning,Biotech Line A/S, Slangerup, Denmark). DNA synthesis and proliferativeindex were determined by 5-bromo-2V-deoxyuridine (BrdUrd) uptake andKi-67 expression, respectively. Cell spheroid cultures labeled with 100 Amol/LBrdUrd for 90 minutes were fixed overnight in 0.5% w/v paraformaldehyde,PBS (pH 7.2; for BrdUrd staining), or 4% formaldehyde ( for Ki-67 staining),washed three times in 9 mg/mL isotonic saline, and pelleted (280 � g ,8 minutes) in polystyrene conical tubes. The final pellet was aspirated drybefore preparing a cell block using Shandon Cytoblock (Thermo ElectronCorporation, Pittsburgh, PA) according to manufacturer’s instructions. Cellblocks were embedded in paraffin and sectioned (4 Am) using conventionalhistopathologic methods. Immunocytochemistry for BrdUrd used thechromogen diaminobenzidine according to manufacturer’s instructions
Tumorigenic Heterogeneity in Cloned Cancer Stem Cells
www.aacrjournals.org 3127 Cancer Res 2005; 65: (8). April 15, 2005
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

(Roche A/S, Hvidovre, Denmark). pKi-67 antigen was detected using MIB-1primary antibody (DAKOCytomation, Glostrup, Denmark). Procollagen type
III (PIIINP) was detected using a polyclonal rabbit anti-human PIIINP
antibody (36), a gift from J. Risteli (University of Oulu, Oulu, Finland). Cyclin
D1 in MCS sections was detected using a murine monoclonal antibody(clone DCS-6, Novocastra Laboratories Ltd., Newcastle upon Tyne,
United Kingdom) and in tumor sections using a rabbit monoclonal
antibody (ref. 37; clone SP4, Lab Vision Corp., AH Diagnostics, Aarhus,
Denmark). p16INK4a expression in MCS was examined using a murinemonoclonal antibody (clone E6H4, DAKOCytomation). p27Kip1 in MCS was
detected using a murine monoclonal antibody (ref. 38; clone SX53G8, a gift
from X. Lu, Ludwig Institute for Cancer Research UCL, London, United
Kingdom). In tumor sections, p27Kip1 was detected using a rabbit polyclonalIgG targeting the amino terminus (clone sc-527, Santa Cruz Biotechnology,
Inc., AH Diagnostics, Aarhus, Denmark). pRB1 was detected using a mouse
monoclonal antibody (NCL-RB1, Novocastra Laboratories Ltd, Newcastleupon Tyne, United Kingdom). Detection employed Immunoperoxidase and
Envision Plus according to manufacturer’s instructions (DAKOCytomation,
Glostrup, Denmark).
Results
hMSC-TERT20 clones share common in vitro monolayercharacteristics. The hMSC-TERT20 population was single cellcloned 186 PDs after first showing tumorigenic potential (Fig. 1A).Eight 96-well plates seeded at limiting dilution yielded 117 wellscontaining single cells that showed high clonogenicity (94%); 110clones expanded by 20 PDs to over a million cells. Most expandableclones (82%) matched the parental population morphologically;cuboidal cells formed a dense cobblestone pattern when confluent.Some (12%) had a flattened squamous morphology, a minority (4%)contained cells with a fusiform morphology, and two clones grewexceptionally slowly [doubling time (DT), f15 days]. From themajority of clones showing uniform and consistent morphology, sixwere arbitrarily selected for comparison.Telomerase activity was normalized to the parental hMSC-
TERT20 population and all six clones showed levels of activityequivalent to that found in the HT-1080 positive control (Table 1).The DT of each clone in monolayer culture was very similar to
that of the parental hMSC-TERT20 population (DT, 1.2 days) aswas the corresponding proliferation-associated Ki-67 labelingindex (LI; Table 1).The clone with highest saturation density, hMSC-TERT20-BD6, had
tightly packed cuboidal cells with a density of 36.8 � 104 cells/cm2 atconfluence with continued growth forming a multilayered culture.All the clones lacked contact inhibition (Table 1).The DNA content profile for all the clones indicated a diploid
phenotype (Table 1) in contrast to the tetraploid fibrosarcoma cellline HT-1080. This agreed with previous chromosomal analysis ofthe hMSC-TERT20 parental population, consistently showing anormal 46 chromosome XY phenotype (17).In contrast to early-passage hMSC-TERT20 cells (PD 85), all
clones showed genetic deletion at the INK4a/ARF locus andhypermethylation of the DBCCR1 gene promoter (Fig. 1B) like theparental population, hMSC-TERT20 PD 417 (17). Despite exploringthe entire coding region and all splice junctions, no TP53 genealteration was detected in the parental hMSC-TERT20 populationor its clones (data not shown).Growth in soft agar revealed heterogeneity among the clones.
Four clones grew poorly in soft agar or not at all, whereas hMSC-TERT20-BD11 and -DB9 resembled the positive hMSC-TERT20parental cells and HT-1080 control (Table 1).All clones stimulated by osteogenic medium expressed osteo-
blastic differentiation markers, although the basal levels and degree
of induction varied between the clones (Fig. 2A). Clones hMSC-TERT20-BB3 and -DB9 had the lowest Col1A1 expression underbasal conditions, but in osteogenic medium it was induced 10- to25-fold. Low basal alkaline phosphatase expression meant thathMSC-TERT20-DB9 also had a relatively high induction of thismarker (Fig. 2B). However, when normalized to h-actin levels, theabsolute level of alkaline phosphatase expression (2 arbitrary units)was much lower than that of clones expressing the highest levels:
Figure 1. hMSC-TERT20-derived clones retain common genetic and epigeneticchanges. A, growth curve for hTERT infected human mesenchymal cellspassaged at 1:20 (hMSC-TERT20). Days in culture refer to the cumulative periodsince infection with the hTERT retroviral vector. Time points when tumorigenicitywas first detected in nude mice and single-cell clones were initiated bylimiting dilution. B, detection of INK4a/ARF deletion in hMSC-TERT20 clones.C, detection of DBCCR1 hypermethylation in hMSC-TERT20 clones bymethylation-specific PCR. DNA treated with sodium bisulfite was PCRamplified with primer pairs specific for unmethylated (U ) and methylated(M ) DBCCR1 alleles. Sss I-methylated DNA provided a positive control formethylated DBCCR1 alleles.
Cancer Research
Cancer Res 2005; 65: (8). April 15, 2005 3128 www.aacrjournals.org
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

hMSC-TERT20-BC8 (20 arbitrary units) and hMSC-TERT20-BD11(40 arbitrary units). Osteocalcin was only detected in osteogenicmedium (Fig. 2C).Clonal heterogeneity in tumor formation. Despite common
DTs and genetic/epigenetic changes, the clones had heterogeneoustumor phenotypes (Fig. 3A) that did not correlate with soft agarcolony formation assays. Four clones (hMSC-TERT20-BB3, -BD6,-DB9, and -BD11) readily formed tumors, and hMSC-TERT20-BB3and -BD11 formed tumors first. Histomorphologically, hMSC-TERT20-BD11 tumors had more dispersed cells with a spindle-shaped morphology, whereas hMSC-TERT20-BB3, -BB6, -CE8, and-DB9 tumors had densely packed cuboidal cells. Although hMSC-TERT20-BB3 and -BD11 had the fastest tumor volume growth(Fig. 3A), this reflected different phenotypes. hMSC-TERT20-BB3had a cell density of 45 nuclei/100 Am2 and a Ki-67 LI of 61.3%,whereas hMSC-TERT20-BD11 had a cell density of only 26 nuclei/100 Am2 but a Ki-67 LI of 42.7%. Microscopic analysis of the lung,liver, kidney, spleen, or lymph nodes did not show any metastases.Consistently, hMSC-TERT20-CE8 only formed tumors after a
dormant period of 24 days; then, the latent tumors grew withkinetics similar to the other clones (Fig. 3A).In first round analysis, hMSC-TERT20-BC8 (passage 6) formed
small nodules at 10 days, in three of five sites, but they receded withno macroscopically visible tumors 18 days postimplantation.Different tumorigenic clones on the opposite flank of the hostforced termination of the hMSC-TERT20-BC8 experiment at21 days. Testing nontumorigenicity more stringently, five furtherhMSC-TERT20-BC8 (passage 9) implantations were analyzed for anextended period. The three tumors from this second round hadatypical growth kinetics (Fig. 3B). Histologic analysis of the tumorthat appeared at 1 week with a volume plateau at 0.5 cm3 revealeda multilobed tumor with interlobular heterogeneity, suggestingselective expansion of only a subset of implanted cells. Cellscultured from the tumor latent until day 97 morphologicallyresembled those of the original hMSC-TERT20-BC8 clone (data not
shown). Ultimately, the hMSC-TERT20-BC8 clone might best bedescribed as poorly tumorigenic.Poor tumor formation correlated with serum dependence
in vitro. When grown in low-serum (0.1% FBS), hMSC-TERT20-BC8 cells were the most susceptible to death (Table 1). By day 9,viability declined to only 16% with the majority dead floating cells(Fig. 4B). Similar serum dependency was seen in 18% of the 110expandable clones. In contrast, hMSC-TERT20-BD11 cells not onlyresisted death (90% viable on day 9) but, compared with theparental population (Fig. 4A), showed enhanced morphologicchange to a reticular network of cords of cells termed capillarymorphogenesis, a phenotype typical of endothelial cells (refs. 27, 39;Fig. 4C). Similar capillary morphogenesis on serum withdrawal wasseen in 16% of the 110 expandable clones. Within 4 days, thecapillary morphogenesis of hMSC-TERT20-BC8 had deteriorated,whereas that of hMSC-TERT20-BD11 was reinforced with morecells forming thicker cords. hMSC-TERT20-BD11 cells also showedcapillary morphogenesis and high viability under serum-freeconditions; changes in cell shape were visible within 2 hours; andintercellular organization to a reticular network initiated within 6hours and was uniform across the whole culture by 12 hours.Confluence was not necessary for morphologic change, but cord-like structures formed more quickly and distinctly in regions ofhigher cell density. Immunocytochemical analysis at 3 days showed14% of the hMSC-TERT20-BD11 cells expressed the marker CD31with equivalent intensity to control endothelial cells (Fig. 4F).hMSC-TERT20-BD11 cells in 10% FBS medium were predominantlynegative for CD31 (Fig. 4E). As expected for mesenchymal stromalcells and in common with endothelial cells, hMSC-TERT20-BD11cells stained strongly for CD105 (endoglin; Fig. 4H and I).
Clonal heterogeneity in three-dimensional spheroid cul-tures correlated with in vivo phenotype. We explored whethertumor-specific heterogeneity (e.g., hMSC-TERT20-CE8 latency)might be attributable to three-dimensional growth. Cells grownon ultralow adhesion plastic soon formed multicellular aggregates.
Table 1. Phenotype of hMSC-TERT20 and clones
Cells Relative
TRAP
activity*
DNA
contentcDT (d) Saturation
density,
10�4
cells/cm2
Contact
inhibited
Soft agar
colonies
(%)
Monolayer
Ki-67+
(%)
Tumors
(n/n)bViability
(%) in
0.1%
FBS
Spheroid Tumor
Ki-67+
(%)BrdUrdLI (%)
Ki-67+
(%)
hMSC-
TERT20
1.0 Diploid 1.20 F 0.0 26.3 F 1.6 No 8.6 F 2.3 ND 10/10 ND ND ND ND
-BB3 0.99 Diploid 1.06 F 0.08 23.2 F 2.0 No 0 66.3 F 4.6 5/5 39 F 4.6 25.9 F 0.8 60.4 F 3.2 61.3 F 6.9
-BC8 0.99 Diploid 1.32 F 0.03 21.5 F 1.8 No 0 71.1 F 8.5 3/10 16 F 1.5 12.4 F 2.7 46.5 F 5.1 43.6 F 3.5-BD6 0.83 Diploid 1.21 F 0.11 36.8 F 1.8 No 3 F 0.5 69.6 F 3.5 5/5 37 F 6.1 17.5 F 1.6 57.1 F 7.1 43.5 F 2.2
-BD11 0.95 Diploid 1.25 F 0.14 21.5 F 1.9 No 16 F 2.9 68.4 F 3.4 5/5 90 F 6.4 21.5 F 2.8 56.7 F 6.3 42.7 F 6.0
-CE8 1.0 Diploid 1.19 F 0.14 14.7 F 0.5 No 0 58.1 F 4.3 5/5 59 F 4.5 3.6 F 3.4 16.2 F 1.7 43.6 F 22.3
-DB9 1.0 Diploid 1.10 F 0.08 24.2 F 2.2 No 12 F 2.9 76.3 F 7.1 5/5 91 F 3.0 21.3 F 7.3 56.7 F 3.2 40.7 F 2.6HT-1080 1.1 Tetraploid ND ND No 8.4 F 2.7 ND 5/5 ND ND ND ND
NOTE: Phenotypic characterization of human bone marrow stromal cell strain hMSC-TERT20, its derived clones, and HT-1080 fibrosarcoma cell line
(positive control).
*Relative TRAP activity data were normalized to the hMSC-TERT20 parental population.cQuantification of DNA content was determined by FACScan flow cytometry based on propidium iodide staining.bhMSC-TERT20 clones were implanted mixed (1:1, v/v) with Matrigel s.c. in immunodeficient mice (NMRI). ND, not determined. LI (%), Labelling Index
of the percentage of BrdUrd-positive cells synthesizing DNA.
Tumorigenic Heterogeneity in Cloned Cancer Stem Cells
www.aacrjournals.org 3129 Cancer Res 2005; 65: (8). April 15, 2005
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

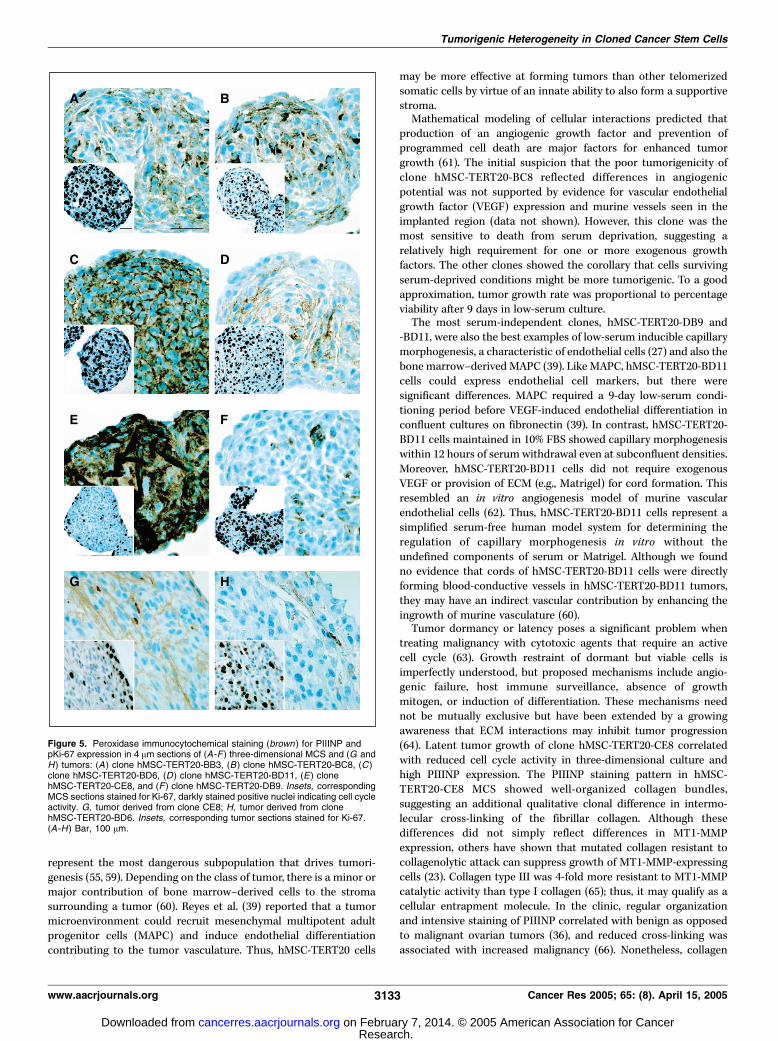
Most clones formed overnight aggregates 100 to 200 Am indiameter consisting of a few hundred cells, but hMSC-TERT20-CE8formed smaller spheroids <50 Am in diameter with <50 cells.Spheroid cultures were grown in suspension for 2 weeks beforeanalysis. The cellular density of the solid cell spheroids mimickedthat observed in the tumor sections (e.g., hMSC-TERT20-BD11 hadthe widest internuclear distances; Fig. 5D). Moreover, hMSC-TERT20-CE8 had a unique three-dimensional culture phenotype ofminimal DNA synthesis (BrdUrd labeling virtually undetectable)confirmed by Ki-67 immunocytochemistry showing low cell cycleactivity (Table 1; Fig. 5E , inset). Unlike the widely distributed pKi-67-positive cells seen in most clones, the staining pattern of hMSC-TERT20-CE8 sections showed preferential staining of cells in thespheroid periphery (Fig. 5A-F, inset).The fibroproliferative response adjacent to malignant tumors is
often characterized by increased expression of type I and IIIprocollagens. Thus, we explored whether the hMSC-TERT20 clones
shared these tumor-stroma characteristics. Coincident with reducedgrowth, hMSC-TERT20-CE8 spheroids had intense and organizedstaining for PIIINP (Fig. 5E). Staining the tumors for PIIINP andKi-67 showed that the in vivo phenotype corresponded with thespheroid cultures. In hMSC-TERT20-CE8 tumors, regions of strongerPIIINP staining had reciprocally less pKi-67 staining that waspreferentially distributed to the tumor periphery (Fig. 5G). Thisstaining pattern was not observed in tumors from the other clones(e.g., hMSC-TERT20-BD6; Fig. 5H). These results were consistentwith previous reports that inhibition of three-dimensional cellgrowth could be generated by solid stress imposed by a meshwork ofECM molecules (23). MT1-MMP was highlighted as the primarymetalloproteinase conferring a growth advantage to tumors inthree-dimensional cultures by pericellular proteolysis of the ECM.Thus, MT1-MMP expression was analyzed by semiquantitative real-time PCR. However, all clones expressed similar MT1-MMP levels,with hMSC-TERT20-CE8 having the second highest level (Fig. 2D).
Figure 2. Real-time PCR gene expressionanalysis of induced osteoblasticdifferentiation markers and relativeexpression of genes encoding ECMproteins and MMP. A-C, expression ofosteoblastic markers collagen 1a, alkalinephosphatase, and osteocalcin comparingcell clones with the parental populationgrown in basal medium (white columns ) orbasal medium + osteogenic growth factors(black columns ). D, relative expression ofMT1-MMP. E, relative expression ofcollagen type III. Expression of targetgenes was normalized for h-actin.Columns, mean of at least twoindependent experiments. *, P < 0.002(paired t test).
Cancer Research
Cancer Res 2005; 65: (8). April 15, 2005 3130 www.aacrjournals.org
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Nonetheless, real-time PCR analysis of monolayer cultures showedthat collagen type III mRNA expression correlated with the spheroidimmunocytochemistry (Fig. 2E).Tumorigenicity correlated with expression of prognostic
markers. We wished to determine whether the parental cell strainshowed heterogeneity for markers correlated previously to cancerin other tumor systems and whether these markers then associatedwith the tumorigenic potential of its clones. Given INK4a/ARF locusalteration, we analyzed members of its complex signaling networkin the tumors and MCS of hMSC-TERT20 cells compared with themost aggressive (hMSC-TERT20-BD11) and least tumorigenic(hMSC-TERT20-BC8) clones. As anticipated from genetic observa-tions, p16INK4a was not detected (data not shown). However, theretinoblastoma protein (pRB1) was highly expressed in all cases(Fig. 4J-L ; LI >90% positive nuclei). In hMSC-TERT20 tumors, cyclinD1 (LI 61%) and p27Kip1 (LI 49%) showed heterogeneous expressionand this was also seen in sections from the cultured MCS (Fig. 4Mand P). In contrast, clone hMSC-TERT20-BD11 tumors showed highcyclin D1 expression (LI 84%), and despite a similar LI, the p27Kip1
nuclear staining intensity was often less than that seen in theparental population (Fig. 4R). Clone hMSC-TERT20-BC8 showed amarkedly elevated p27Kip1 expression (LI 83%; Fig. 4Q).
Discussion
Telomerase may be considered an immortalizing agent of choice,with enhanced chromosome stability compared with other popularimmortalizing agents (e.g., SV40 large T antigen; ref. 40). Manylaboratories have ‘‘telomerized’’ a wide range of different humancell types (41, 42). Nonetheless, varying consequences of telomeraseexpression may reflect different technical approaches. For example,recent reports of similar genetic alterations in long-term cultures oftelomerase-immortalized cells (11, 13) used a coselectable marker.Neomycin selection can influence gene expression and metabolismand may have a role in generating genetic alterations (43, 44).However, our use of a nonselectable vector confirms that geneticalterations in long-term cultures of telomerase-immortalized cellsare not simply artifacts from use of selectable markers. Othercontributory factors, including vector integration site, levels ofectopic gene expression, culture conditions, including platingdensity, and inherent qualities of different cell types, will inevitablyvary (45). Therefore, the outcome of ectopic telomerase expressionin long-term cultures is likely to remain unpredictable.The tumorigenic phenotype of hMSC-TERT20 cells arose from
the fundamental force of natural selection for cells that grow morerapidly in vitro . The spontaneous genetic and epigenetic changes weobserved, deletion at the INK4a/ARF locus and epigenetic silencingby methylation of the tumor suppressor gene DBCCR1 , correspondto similar changes frequently found in human tumors andosteosarcoma-derived cell lines (46–48). The bmi-1 pathway,important for stem cell self-renewal, targets dual inactivation ofp16INK4a and p14ARF (49). Thus, p16INK4a and p14ARF pathways arelikely to govern key fundamental aspects of cell growth regulationcommon to tumors and stem cells, with susceptibility forspontaneous alteration in both in vitro and in vivo microenviron-ments. Our finding no TP53 gene alteration supports data that thisis no longer necessary after inactivation of the p53 pathway via ARFloss (50, 51). Although its role may be permissive rather than directlyoncogenic (52), telomerase is likely to be very influential by being aninitiating event in our tumorigenesis model. The tumorigenichMSC-TERT20 population and its clones had a diploid karyotypeand DNA content. When escape from a senescence-induced G1
arrest is mediated by p53 loss in a telomerase-null background (53)or p16INK4a inactivation is an early event (54), aberrant growth isoften accompanied by telomere attrition, genomic instability, anddevelopment of aneuploid karyotypes. Studies exploring theprognostic significance of telomerase, p16INK4a, and/or p53 alter-ations will need to consider the order of events.Tumor interactions with surrounding host tissue were not
components of the in vitro selection pressure that led to evolutionof the hMSC-TERT20 population. Moreover, induction of tumorvasculature can be a nonautonomous event, also favoring growth ofneighboring cells. Large differences in the tumorigenic potential ofindividual cells within a tumor has led to the classification of a rarebut dominant subpopulation, known as cancer stem cells (55).Thus, it was highly relevant to determine whether tumorigenicitywas the outcome for (a) most cells in the hMSC-TERT20population, (b) a dominant subset of autonomous cells, or (c)interdependent interactions between different subsets of nonau-tonomous cells. Consistent with the hMSC-TERT20 populationrepresenting a cancer stem cell model, all of the single-cell clonestested were tumorigenic to a greater or lesser extent. Althoughsome cells did not survive the single-cell cloning procedure (6%),cloning per se did not necessarily introduce a bias favoring theoutgrowth of more aggressive cells. The parental population was
Figure 3. Growth kinetics for hMSC-TERT20 clone tumors. A, growth curves forfirst round implantations: hMSC-TERT20-BB3 (5), hMSC-TERT20-BC8 (n),hMSC-TERT20-BD6 (.), hMSC-TERT20-BD11 (E), hMSC-TERT20-CE8 (o),hMSC-TERT20-DB9 (4). B, growth curves for individual hMSC-TERT20-BC8tumors from second round implantations: BC8-NMT1 (o), BC8-NMT2 (E),BC8-NMT3 (5).
Tumorigenic Heterogeneity in Cloned Cancer Stem Cells
www.aacrjournals.org 3131 Cancer Res 2005; 65: (8). April 15, 2005
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

highly tumorigenic and clonogenic from the outset, and two of thesix clones formed less aggressive tumors. Therefore, in mesenchy-mal stem cells, telomerase activation, subsequent immortalization,and long-term culture can induce a cellular autonomy capable offorming rapidly growing tumors.The original observation that late-passage hMSC-TERT20 cells
formed tumors was unexpected, contrasting with the boneformation hitherto characteristic of hydroxyapatite/tricalcium phos-phate implantation experiments with early-passage hMSC-TERT20cells (2). However, tumorigenicity did not totally prevent induction ofosteoblast differentiation markers when cells were treated withosteogenic medium. This need not be surprising because humanosteosarcoma cell lines show induction of differentiation markers
(56) and hTERT expression plus p16INK4a inactivation did notabrogate differentiation pathways in keratinocytes (57). Nonetheless,no single clone had optimal induction of all differentiation markers.The relationship between osteoblast proliferation and differentiationinvolves complex hormone modulation of gene expression depen-dent on the maturation state of the osteoblast (58). Although somedifferentiation pathways were clearly retained in the clones,complete differentiation to bone was compromised.It may be more than coincidental that adult mesenchymal stem
cells provided the first example of spontaneous tumorigenicity intelomerized human cells (17). Parallels between tumor cells andstem cells fuel hypotheses that tumors may originate from thetransformation of stem cells, and cancer stem cells may ultimately
Figure 4. Contrasting phenotypes of the parental populationand poorly tumorigenic versus highly tumorigenic clones. A,phase-contrast photomicrograph showing capillarymorphogenesis of hMSC-TERT20 cells after 9 days in 0.1%FBS medium. B, clone hMSC-TERT20-BC8 under sameconditions showed mostly floating dead cells. C, clonehMSC-TERT20-BD11 in 0.1% FBS showed enhanced capillarymorphogenesis. D -F, peroxidase immunocytochemical staining(brown) for endothelial marker CD31 in (D ) TIME endothelialcells (positive control), (E ) clone hMSC-TERT20-BD11 cells in10% FBS medium, and (F ) clone hMSC-TERT20-BD11 cells in0.1% FBS medium. G -I, peroxidase immunocytochemicalstaining (brown ) for CD105 in (G) TIME cells (positive control),(H ) clone hMSC-TERT20-BD11 cells in 10% FBS, and (I )clone hMSC-TERT20-BD11 cells in 0.1%FBS. J -R, peroxidaseimmunocytochemical staining (brown ) for prognostic markers(J -L ) pRB1, (M -O) cyclin D1, and (P -R) p27Kip1 in tumor sectionsderived from (J , M , and P) hMSC-TERT20, (K , N , and Q )clone hMSC-TERT20-BC8, and (L , O , and R ) clonehMSC-TERT20-BD11. Insets, immunocytochemistry forcorresponding MCS sections, peroxidase-immunostained(brown , positive) nuclei contrasted with counterstained(blue , negative) nuclei. (A-R ) Bar, 100 Am.
Cancer Research
Cancer Res 2005; 65: (8). April 15, 2005 3132 www.aacrjournals.org
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
represent the most dangerous subpopulation that drives tumori-genesis (55, 59). Depending on the class of tumor, there is a minor ormajor contribution of bone marrow–derived cells to the stromasurrounding a tumor (60). Reyes et al. (39) reported that a tumormicroenvironment could recruit mesenchymal multipotent adultprogenitor cells (MAPC) and induce endothelial differentiationcontributing to the tumor vasculature. Thus, hMSC-TERT20 cells
may be more effective at forming tumors than other telomerizedsomatic cells by virtue of an innate ability to also form a supportivestroma.Mathematical modeling of cellular interactions predicted that
production of an angiogenic growth factor and prevention ofprogrammed cell death are major factors for enhanced tumorgrowth (61). The initial suspicion that the poor tumorigenicity ofclone hMSC-TERT20-BC8 reflected differences in angiogenicpotential was not supported by evidence for vascular endothelialgrowth factor (VEGF) expression and murine vessels seen in theimplanted region (data not shown). However, this clone was themost sensitive to death from serum deprivation, suggesting arelatively high requirement for one or more exogenous growthfactors. The other clones showed the corollary that cells survivingserum-deprived conditions might be more tumorigenic. To a goodapproximation, tumor growth rate was proportional to percentageviability after 9 days in low-serum culture.The most serum-independent clones, hMSC-TERT20-DB9 and
-BD11, were also the best examples of low-serum inducible capillarymorphogenesis, a characteristic of endothelial cells (27) and also thebone marrow–derived MAPC (39). Like MAPC, hMSC-TERT20-BD11cells could express endothelial cell markers, but there weresignificant differences. MAPC required a 9-day low-serum condi-tioning period before VEGF-induced endothelial differentiation inconfluent cultures on fibronectin (39). In contrast, hMSC-TERT20-BD11 cells maintained in 10% FBS showed capillary morphogenesiswithin 12 hours of serum withdrawal even at subconfluent densities.Moreover, hMSC-TERT20-BD11 cells did not require exogenousVEGF or provision of ECM (e.g., Matrigel) for cord formation. Thisresembled an in vitro angiogenesis model of murine vascularendothelial cells (62). Thus, hMSC-TERT20-BD11 cells represent asimplified serum-free human model system for determining theregulation of capillary morphogenesis in vitro without theundefined components of serum or Matrigel. Although we foundno evidence that cords of hMSC-TERT20-BD11 cells were directlyforming blood-conductive vessels in hMSC-TERT20-BD11 tumors,they may have an indirect vascular contribution by enhancing theingrowth of murine vasculature (60).Tumor dormancy or latency poses a significant problem when
treating malignancy with cytotoxic agents that require an activecell cycle (63). Growth restraint of dormant but viable cells isimperfectly understood, but proposed mechanisms include angio-genic failure, host immune surveillance, absence of growthmitogen, or induction of differentiation. These mechanisms neednot be mutually exclusive but have been extended by a growingawareness that ECM interactions may inhibit tumor progression(64). Latent tumor growth of clone hMSC-TERT20-CE8 correlatedwith reduced cell cycle activity in three-dimensional culture andhigh PIIINP expression. The PIIINP staining pattern in hMSC-TERT20-CE8 MCS showed well-organized collagen bundles,suggesting an additional qualitative clonal difference in intermo-lecular cross-linking of the fibrillar collagen. Although thesedifferences did not simply reflect differences in MT1-MMPexpression, others have shown that mutated collagen resistant tocollagenolytic attack can suppress growth of MT1-MMP-expressingcells (23). Collagen type III was 4-fold more resistant to MT1-MMPcatalytic activity than type I collagen (65); thus, it may qualify as acellular entrapment molecule. In the clinic, regular organizationand intensive staining of PIIINP correlated with benign as opposedto malignant ovarian tumors (36), and reduced cross-linking wasassociated with increased malignancy (66). Nonetheless, collagen
Figure 5. Peroxidase immunocytochemical staining (brown ) for PIIINP andpKi-67 expression in 4 Am sections of (A-F ) three-dimensional MCS and (G andH) tumors: (A ) clone hMSC-TERT20-BB3, (B) clone hMSC-TERT20-BC8, (C )clone hMSC-TERT20-BD6, (D ) clone hMSC-TERT20-BD11, (E) clonehMSC-TERT20-CE8, and (F ) clone hMSC-TERT20-DB9. Insets, correspondingMCS sections stained for Ki-67, darkly stained positive nuclei indicating cell cycleactivity. G, tumor derived from clone CE8; H, tumor derived from clonehMSC-TERT20-BD6. Insets, corresponding tumor sections stained for Ki-67.(A-H ) Bar, 100 Am.
Tumorigenic Heterogeneity in Cloned Cancer Stem Cells
www.aacrjournals.org 3133 Cancer Res 2005; 65: (8). April 15, 2005
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

type III need not be the only ECM molecule responsible for thehMSC-TERT20-CE8 phenotype. The ECM protein fibulin-1D couldmarkedly extend the tumor latency of a human fibrosarcoma cellline (67). It remains possible that PIIINP overexpression is anindirect indicator of altered matrix deposition rather than theprinciple matrix molecule governing tumor dormancy.Further proteins clinically correlated to tumorigenicity were
heterogeneously expressed in the parental hMSC-TERT20 strain andmore specifically associated with the tumorigenic potential of theleast and most aggressive clones. The uniformly high pRB1expression most likely reflected absence of p16INK4a expression(68). Patients with tumors that have low p27Kip1 expression usuallyhave an inferior prognosis to those with high expression (69), and wefound highest p27Kip1 expression in the poorly tumorigenic clone.Overexpression of cyclin D1 is known to correlate with the earlyonset of cancer and tumor progression, and we observed highestcyclin D1 expression in the most aggressive clone hMSC-TERT20-BD11. Notably, increased cyclin D1 expression need not necessarilycorrelate with increased DNA synthesis, and additional roles forcyclin D1 include regulation of cell adhesion and motility (70).
The hope for a much wider application of stem cells intherapeutic intervention, possibly enhanced with gene therapy, ismodulated by caution regarding tumorigenic potential. The qualitycontrol of therapeutic stem cells will be improved with a morecomprehensive understanding of possible risk factors. hMSC-TERT20 cells represent a useful cell system for defining spontane-ous tumorigenic events along the path from stem cell to cancerstem cell.
Acknowledgments
Received 7/22/2004; revised 11/30/2004; accepted 1/12/2005.Grant support: Danish Medical Research Council, Danish Center for Stem Cell
Research, Karen Elise Jensen’s Fond, Novo Nordisk Foundation, Gross M. Brogaard ogHustru Foundation, Jacob Madsen og Hustru Foundation, and Danish LægeforeningResearch Foundation; Alfred Benzon’s Foundation World Laboratory fellowship andresearch fellowship (B.M. Abdallah).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Inger Andersen, Lone Christiansen, and Tina K.L. Nielsen for excellenttechnical assistance; Birgitte Elsnab and Graham Leslie for help with fluorescence-activated cell sorting analysis (Odense University Hospital); and Margit Baeksted andMorten Schou (Bartholin Instituttet) for expertise in animal husbandry.
References1. Jiang Y, Jahagirdar BN, Reinhardt RL, et al. Pluripo-tency of mesenchymal stem cells derived from adultmarrow. Nature 2002;418:41–9.
2. Simonsen JL, Rosada C, Serakinci N, et al. Telomeraseexpression extends the proliferative life-span andmaintains the osteogenic potential of human bonemarrow stromal cells. Nat Biotechnol 2002;20:592–6.
3. Stenderup K, Justesen J, Clausen C, Kassem M. Aging isassociated with decreased maximal life span andaccelerated senescence of bone marrow stromal cells.Bone 2003;33:919–26.
4. Shi S, Gronthos S, Chen S, et al. Bone formation byhuman postnatal bone marrow stromal stem cells isenhanced by telomerase expression. Nat Biotechnol2002;20:587–91.
5. Xu J, Yang X. Telomerase activity in early bovineembryos derived from parthenogenetic activation andnuclear transfer. Biol Reprod 2001;64:770–4.
6. Jiang XR, Jimenez G, Chang E, et al. Telomeraseexpression in human somatic cells does not inducechanges associated with a transformed phenotype. NatGenet 1999;21:111–4.
7. Morales CP, Holt SE, Ouellette M, et al. Absence ofcancer-associated changes in human fibroblasts im-mortalized with telomerase. Nat Genet 1999;21:115–8.
8. Wang J, Hannon GJ, Beach DH. Risky immortalizationby telomerase. Nature 2000;405:755–6.
9. Farwell DG, Shera KA, Koop JI, et al. Genetic andepigenetic changes in human epithelial cells immortal-ized by telomerase. Am J Pathol 2000;156:1537–47.
10. Lindvall C, Hou M, Komurasaki T, et al. Molec-ular characterization of human telomerase reversetranscriptase-immortalized human fibroblasts bygene expression profiling: activation of the epiregu-lin gene. Cancer Res 2003;63:1743–7.
11. Noble JR, Zhong ZH, Neumann AA, Melki JR, Clark SJ,Reddel RR. Alterations in the p16(INK4a) and p53tumor suppressor genes of hTERT-immortalized humanfibroblasts. Oncogene 2004;23:3116–21.
12. Mondello C, Chiesa M, Rebuzzini P, et al. Karyotypeinstability and anchorage-independent growth intelomerase-immortalized fibroblasts from two cente-narian individuals. Biochem Biophys Res Commun2003;308:914–21.
13. Milyavsky M, Shats I, Erez N, et al. Prolonged cultureof telomerase-immortalized human fibroblasts leads toa premalignant phenotype. Cancer Res 2003;63:7147–57.
14. Stewart SA, Hahn WC, O’Connor BF, et al. Telomer-ase contributes to tumorigenesis by a telomere length-
independent mechanism. Proc Natl Acad Sci U S A2002;99:12606–11.
15. Chang S, DePinho RA. Telomerase extracurricularactivities. Proc Natl Acad Sci U S A 2002;99:12520–2.
16. Smith LL, Coller HA, Roberts JM. Telomerasemodulates expression of growth-controlling genes andenhances cell proliferation. Nat Cell Biol 2003;5:474–9.
17. Serakinci N, Guldberg P, Burns JS, et al. Adult humanmesenchymal stem cell as a target for neoplastictransformation. Oncogene 2004;23:5095–8.
18. Dvorak HF. Rous-Whipple Award Lecture. Howtumors make bad blood vessels and stroma. Am JPathol 2003;162:1747–57.
19. Madsen MW, Moyret C, Theillet C, Briand P. Growthfactor requirement, oncogene expression and TP53mutations of a tumorigenic and a non-tumorigenicsubline of the human breast carcinoma cell line, HMT-3909. Eur J Cancer 1995;31A:362–7.
20. Chen CS, Mrksich M, Huang S, Whitesides GM,Ingber DE. Geometric control of cell life and death.Science 1997;276:1425–8.
21. Egeblad M, Werb Z. New functions for the matrixmetalloproteinases in cancer progression. Nat RevCancer 2002;2:161–74.
22. Yamada KM. Cell biology: tumour jailbreak. Nature2003;424:889–90.
23. Hotary KB, Allen ED, Brooks PC, Datta NS, Long MW,Weiss SJ. Membrane type I matrix metalloproteinaseusurps tumor growth control imposed by the three-dimensional extracellular matrix. Cell 2003;114:33–45.
24. Seiki M, Yana I. Roles of pericellular proteolysis bymembrane type-1 matrix metalloproteinase in cancerinvasion and angiogenesis. Cancer Sci 2003;94:569–74.
25. Hedlund TE, Duke RC, Miller GJ. Three-dimensionalspheroid cultures of human prostate cancer cell lines.Prostate 1999;41:154–65.
26. Rasheed S, Nelson-Rees WA, Toth EM, Arnstein P,Gardner MB. Characterization of a newly derived humansarcoma cell line (HT-1080). Cancer 1974;33:1027–33.
27. Venetsanakos E, Mirza A, Fanton C, Romanov SR,Tlsty T, McMahon M. Induction of tubulogenesis intelomerase-immortalized human microvascular endo-thelial cells by glioblastoma cells. Exp Cell Res 2002;273:21–33.
28. Wege H, Chui MS, Le HT, Tran JM, Zern MA. SYBRGreen real-time Telomeric Repeat Amplification Proto-col for the rapid quantification of telomerase activity.Nucleic Acids Res 2003;31:E3.
29. Kim NW, Piatyszek MA, Prowse KR, et al. Specificassociation of human telomerase activity with immortalcells and cancer. Science 1994;266:2011–5.
30. Guldberg P, Nedergaard T, Nielsen HJ, Olsen AC,Ahrenkiel V, Zeuthen J. Single-step DGGE-basedmutation scanning of the p53 gene: application togenetic diagnosis of colorectal cancer. Hum Mutat1997;9:348–55.
31. Gronbaek K, de Nully Brown P, Moller MB, et al.Concurrent disruption of p16INK4a and the ARF-p53pathway predicts poor prognosis in aggressive non-Hodgkin’s lymphoma. Leukemia 2000;14:1727–35.
32. Herman JG, Graff JR, Myohanen S, Nelkin BD, BaylinSB. Methylation-specific PCR: a novel PCR assay formethylation status of CpG islands. Proc Natl Acad SciU S A 1996;93:9821–6.
33. Clark SJ, Harrison J, Paul CL, Frommer M. Highsensitivity mapping of methylated cytosines. NucleicAcids Res 1994;22:2990–7.
34. Gao S, Worm J, Guldberg P, et al. Loss ofheterozygosity at 9q33 and hypermethylation of theDBCCR1 gene in oral squamous cell carcinoma. Br JCancer 2004;91:760–4.
35. Abdallah BM, Jensen CH, Gutierrez G, Leslie RG,Jensen TG, Kassem M. Regulation of human skeletalstem cells differentiation by Dlk1/Pref-1. J Bone MinerRes 2004;19:841–52.
36. Zhu GG, Stenback F, Risteli L, Risteli J, Kauppila A.Organization of type III collagen in benign andmalignant ovarian tumors. An immunohistochemicalstudy. Cancer 1993;72:1679–84.
37. Cheuk W, Wong KO, Wong CS, Chan JK. Consistentimmunostaining for cyclin D1 can be achieved on aroutine basis using a newly available rabbit monoclonalantibody. Am J Surg Pathol 2004;28:801–7.
38. Fredersdorf S, Burns J, Milne AM, et al. High levelexpression of p27(kip1) and cyclin D1 in some humanbreast cancer cells: inverse correlation between theexpression of p27(kip1) and degree of malignancy inhuman breast and colorectal cancers. Proc Natl AcadSci U S A 1997;94:6380–5.
39. Reyes M, Dudek A, Jahagirdar B, Koodie L, Marker PH,Verfaillie CM. Origin of endothelial progenitors inhuman postnatal bone marrow. J Clin Invest 2002;109:337–46.
40. Cotsiki M, Lock RL, Cheng Y, et al. Simian virus 40large T antigen targets the spindle assembly check-point protein Bub1. Proc Natl Acad Sci U S A 2004;101:947–52.
41. Bodnar AG, Ouellette M, Frolkis M, et al. Extension oflife-span by introduction of telomerase into normalhuman cells. Science 1998;279:349–52.
42. Counter CM, Hahn WC, Wei W, et al. Dissociationamong in vitro telomerase activity, telomere maintenance,
Cancer Research
Cancer Res 2005; 65: (8). April 15, 2005 3134 www.aacrjournals.org
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

and cellular immortalization. Proc Natl Acad Sci U S A1998;95:14723–8.
43. Valera A, Perales JC, Hatzoglou M, Bosch F.Expression of the neomycin-resistance (neo) geneinduces alterations in gene expression and metabolism.Hum Gene Ther 1994;5:449–56.
44. Sanchez-Carbayo M, Belbin TJ, Scotlandi K, et al.Expression profiling of osteosarcoma cells transfectedwith MDR1 and NEO genes: regulation of cell adhesion,apoptosis, and tumor suppression-related genes. LabInvest 2003;83:507–17.
45. Keith WN. From stem cells to cancer: balancingimmortality and neoplasia. Oncogene 2004;23:5092–4.
46. Martin JA, Forest E, Block JA, et al. Malignanttransformation in human chondrosarcoma cells sup-ported by telomerase activation and tumor suppressorinactivation. Cell Growth Differ 2002;13:397–407.
47. Nishiyama H, Gill JH, Pitt E, Kennedy W, KnowlesMA. Negative regulation of G(1)/S transition by thecandidate bladder tumour suppressor gene DBCCR1.Oncogene 2001;20:2956–64.
48. Park YB, Park MJ, Kimura K, Shimizu K, Lee SH,Yokota J. Alterations in the INK4a/ARF locus and theireffects on the growth of human osteosarcoma cell lines.Cancer Genet Cytogenet 2002;133:105–11.
49. Pardal R, Clarke MF, Morrison SJ. Applying theprinciples of stem-cell biology to cancer. Nat RevCancer 2003;3:895–902.
50. Cobbers JM, Wolter M, Reifenberger J, et al. Frequentinactivation of CDKN2A and rare mutation of TP53 inPCNSL. Brain Pathol 1998;8:263–76.
51. Sharp R, Recio JA, Jhappan C, et al. Synergismbetween INK4a/ARF inactivation and aberrant HGF/SFsignaling in rhabdomyosarcomagenesis. Nat Med 2002;8:1276–80.
52. Harley CB. Telomerase is not an oncogene. Oncogene2002;21:494–502.
53. Artandi SE, Chang S, Lee SL, et al. Telomeredysfunction promotes non-reciprocal translocationsand epithelial cancers in mice. Nature 2000;406:641–5.
54. Wong DJ, Paulson TG, Prevo LJ, et al. p16(INK4a)lesions are common, early abnormalities that undergoclonal expansion in Barrett’s metaplastic epithelium.Cancer Res 2001;61:8284–9.
55. Reya T, Morrison SJ, Clarke MF, Weissman IL. Stemcells, cancer, and cancer stem cells. Nature 2001;414:105–11.
56. Siggelkow H, Schenck M, Rohde M, et al. Prolongedculture of HOS 58 human osteosarcoma cells with1,25-(OH)2-D3, TGF-h, and dexamethasone revealsphysiological regulation of alkaline phosphatase, dis-sociated osteocalcin gene expression, and proteinsynthesis and lack of mineralization. J Cell Biochem2002;85:279–94.
57. Dickson MA, Hahn WC, Ino Y, et al. Humankeratinocytes that express hTERT and also bypass ap16(INK4a)-enforced mechanism that limits life spanbecome immortal yet retain normal growth anddifferentiation characteristics. Mol Cell Biol 2000;20:1436–47.
58. Lian JB, Stein GS. Concepts of osteoblast growth anddifferentiation: basis for modulation of bone celldevelopment and tissue formation. Crit Rev Oral BiolMed 1992;3:269–305.
59. Marx J. Cancer research. Mutant stem cells may seedcancer. Science 2003;301:1308–10.
60. Ribatti D, Vacca A, Dammacco F. New non-angiogenesis dependent pathways for tumour growth.Eur J Cancer 2003;39:1835–41.
61. Tomlinson IP, Bodmer WF. Modelling the conse-quences of interactions between tumour cells. Br JCancer 1997;75:157–60.
62. Chen CS, Toda K, Fujii K, Imamura S. Furthercharacterization of a new in vitro angiogenesis modelunder serum free culture conditions; suppression ofendothelial cell differentiation by serum. J Dermatol Sci1998;16:208–15.
63. Hart IR. Perspective: tumour spread—the problemsof latency. J Pathol 1999;187:91–4.
64. Bissell MJ, Radisky D. Putting tumours in context.Nat Rev Cancer 2001;1:46–54.
65. Ohuchi E, Imai K, Fujii Y, Sato H, Seiki M, Okada Y.Membrane type 1 matrix metalloproteinase digestsinterstitial collagens and other extracellular matrixmacromolecules. J Biol Chem 1997;272:2446–51.
66. Kauppila S, Bode MK, Stenback F, Risteli L, Risteli J.Cross-linked telopeptides of type I and III collagens inmalignant ovarian tumours in vivo . Br J Cancer 1999;81:654–61.
67. Qing J, Maher VM, Tran H, Argraves WS, Dunstan RW,McCormick JJ. Suppression of anchorage-independentgrowth and Matrigel invasion and delayed tumor forma-tion by elevated expression of fibulin-1D in humanfibrosarcoma-derived cell lines. Oncogene 1997;15:2159–68.
68. Chatterjee SJ, George B, Goebell PJ, et al. Hyper-phosphorylation of pRb: a mechanism for RB tumoursuppressor pathway inactivation in bladder cancer.J Pathol 2004;203:762–70.
69. Blain SW, Scher HI, Cordon-Cardo C, Koff A. p27 as atarget for cancer therapeutics. Cancer Cell 2003;3:111–5.
70. Fu M, Wang C, Li Z, Sakamaki T, Pestell RG.Minireview: Cyclin D1: normal and abnormal functions.Endocrinology 2004;145:5439–47.
Tumorigenic Heterogeneity in Cloned Cancer Stem Cells
www.aacrjournals.org 3135 Cancer Res 2005; 65: (8). April 15, 2005
Research. on February 7, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from




















