
Stem Cells NADPH Oxidase-BM microenvironment 2012
12
REGENERATIVE MEDICINE NADPH Oxidase 2 Regulates Bone Marrow Microenvironment Following Hindlimb Ischemia: Role in Reparative Mobilization of Progenitor Cells NORIFUMI URAO, a,b RONALD D. MCKINNEY, a,b TOHRU FUKAI, b,c,d MASUKO USHIO-FUKAI a,b a Department of Pharmacology, Center for Lung and Vascular Biology; c Department of Medicine; d Department of Pharmacology; and b Center for Cardiovascular Research, University of Illinois at Chicago, Chicago, Illinois, USA Key Words. Hematopoietic stem cell • Mobilization • Reactive oxygen species • Ischemia • Hypoxia ABSTRACT Bone marrow (BM) microenvironment, which is regulated by hypoxia and proteolytic enzymes, is crucial for stem/ progenitor cell function and mobilization involved in post- natal neovascularization. We demonstrated that NADPH oxidase 2 (Nox2)-derived reactive oxygen species (ROS) are involved in postischemic mobilization of BM cells and revascularization. However, role of Nox2 in regulating BM microenvironment in response to ischemic injury remains unknown. Here, we show that hindlimb ischemia of mice increases ROS production in both the endosteal and cen- tral region of BM tissue in situ, which is almost completely abolished in Nox2 knockout (KO) mice. This Nox2-de- pendent ROS production is mainly derived from Gr-1 1 myeloid cells in BM. In vivo injection of hypoxyprobe reveals that endosteum at the BM is hypoxic with high expression of hypoxia-inducible factor-1a in basal state. Following hindlimb ischemia, hypoxic areas and HIF-1a expression are expanded throughout the BM, which is inhibited in Nox2 KO mice. This ischemia-induced altera- tion of Nox2-dependent BM microenvironment is associ- ated with an increase in vascular endothelial growth factor expression and Akt phosphorylation in BM tissue, thereby promoting Lin 2 progenitor cell survival and expansion, leading to their mobilization from BM. Fur- thermore, hindlimb ischemia increases proteolytic enzymes membrane type 1-matrix metalloproteinase (MMP) expression and MMP-9 activity in BM, which is inhibited in Nox2 KO mice. In summary, Nox2-dependent increase in ROS plays a critical role in regulating hypoxia expan- sion and proteolytic activities in BM microenvironment in response to tissue ischemia. This in turn promotes progen- itor cell expansion and reparative mobilization from BM, leading to postischemic neovascularization and tissue repair. STEM CELLS 2012;30:923–934 Disclosure of potential conflicts of interest is found at the end of this article. INTRODUCTION Bone marrow (BM)-derived stem and progenitor cells have a critical role in maintaining vascular health and facilitating vasculogenesis/angiogenesis [1]. The BM microenvironment, also termed the stem cell niche, provides essential cues that regulate stem and progenitor cell functions and their mobiliza- tion into the circulation involved in postnatal neovasculariza- tion [2]. Hypoxia and proteolytic enzymes are key extrinsic factors that affect BM microenvironment regulating stem and progenitor cell function and their reparative mobilization. Stem/progenitor cells are originally shown to be located in two different BM microenvironments, including the BM inter- face (endosteal niche) that provides low oxygen environment for maintaining hematopoietic stem cells quiescence and the vascular niche that provides oxygenated environment for cell proliferation and differentiation [3–5]. Ischemic injury increases cytokines and growth factors such as vascular endo- thelial growth factor (VEGF) in BM and circulation, which in turn activates matrix metalloproteinase (MMP)-9 and releases soluble Kit ligand in the BM [6]. MMPs including MMP-9 that is secreted mainly by neutrophils in BM [7] and MT (membrane type)1-MMP [8] that is anchored on the cell sur- face play a significant role in stem/progenitor cell mobiliza- tion and angiogenesis. Thus, understanding mechanisms by which ischemic injury regulates BM microenvironment and stem/progenitor cell function is essential for developing novel therapeutic strategies for treatment of various ischemic diseases. Although excess amounts of reactive oxygen species (ROS) such as superoxide anion (O 2 ) and hydrogen peroxide (H 2 O 2 ) are detrimental to cells, ROS at appropriate level function as signaling molecules to mediate normal cell Author contributions: N.U.: conception and design, financial support, collection and/or assembly of data, data analysis and interpretation, manuscript writing, and final approval of manuscript; R.D.M.: administrative support, collection of data, provision of study material, and final approval of manuscript; T.F.: financial support, data analysis and interpretation, and final approval of manuscript; M.U.: financial support, conception and design, data analysis and interpretation, manuscript writing, and final approval of manuscript. Correspondence: Masuko Ushio-Fukai, Ph.D., Department of Pharmacology, Center for Lung and Vascular Biology, Center for Cardiovascular Research, University of Illinois at Chicago, 835 S. Wolcott, M/C868, E403 MSB, Chicago, Illinois 60612, USA. Telephone: 312-996-7665; Fax: 312-996-1225; e-mail: [email protected] Received August 16, 2011; accepted for publication January 9, 2012; first published online in STEM CELLS EXPRESS January 30, 2012. V C AlphaMed Press 1066-5099/2012/$30.00/0 doi: 10.1002/stem.1048 STEM CELLS 2012;30:923–934 www.StemCells.com
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Stem Cells NADPH Oxidase-BM microenvironment 2012

REGENERATIVE MEDICINE
NADPH Oxidase 2 Regulates Bone Marrow Microenvironment
Following Hindlimb Ischemia: Role in Reparative Mobilization
of Progenitor Cells
NORIFUMI URAO,a,b RONALD D. MCKINNEY,a,b TOHRU FUKAI,b,c,d MASUKO USHIO-FUKAIa,b
aDepartment of Pharmacology, Center for Lung and Vascular Biology; cDepartment of Medicine; dDepartment of
Pharmacology; and bCenter for Cardiovascular Research, University of Illinois at Chicago, Chicago, Illinois, USA
Key Words. Hematopoietic stem cell • Mobilization • Reactive oxygen species • Ischemia • Hypoxia
ABSTRACT
Bone marrow (BM) microenvironment, which is regulatedby hypoxia and proteolytic enzymes, is crucial for stem/progenitor cell function and mobilization involved in post-
natal neovascularization. We demonstrated that NADPHoxidase 2 (Nox2)-derived reactive oxygen species (ROS)are involved in postischemic mobilization of BM cells and
revascularization. However, role of Nox2 in regulating BMmicroenvironment in response to ischemic injury remains
unknown. Here, we show that hindlimb ischemia of miceincreases ROS production in both the endosteal and cen-tral region of BM tissue in situ, which is almost completely
abolished in Nox2 knockout (KO) mice. This Nox2-de-pendent ROS production is mainly derived from Gr-11
myeloid cells in BM. In vivo injection of hypoxyprobereveals that endosteum at the BM is hypoxic with highexpression of hypoxia-inducible factor-1a in basal state.
Following hindlimb ischemia, hypoxic areas and HIF-1a
expression are expanded throughout the BM, which isinhibited in Nox2 KO mice. This ischemia-induced altera-tion of Nox2-dependent BM microenvironment is associ-
ated with an increase in vascular endothelial growthfactor expression and Akt phosphorylation in BM tissue,thereby promoting Lin2 progenitor cell survival and
expansion, leading to their mobilization from BM. Fur-thermore, hindlimb ischemia increases proteolytic enzymes
membrane type 1-matrix metalloproteinase (MMP)expression and MMP-9 activity in BM, which is inhibitedin Nox2 KO mice. In summary, Nox2-dependent increase
in ROS plays a critical role in regulating hypoxia expan-sion and proteolytic activities in BM microenvironment in
response to tissue ischemia. This in turn promotes progen-itor cell expansion and reparative mobilization from BM,leading to postischemic neovascularization and tissue
repair. STEM CELLS 2012;30:923–934
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Bone marrow (BM)-derived stem and progenitor cells have acritical role in maintaining vascular health and facilitatingvasculogenesis/angiogenesis [1]. The BM microenvironment,also termed the stem cell niche, provides essential cues thatregulate stem and progenitor cell functions and their mobiliza-tion into the circulation involved in postnatal neovasculariza-tion [2]. Hypoxia and proteolytic enzymes are key extrinsicfactors that affect BM microenvironment regulating stem andprogenitor cell function and their reparative mobilization.Stem/progenitor cells are originally shown to be located intwo different BM microenvironments, including the BM inter-face (endosteal niche) that provides low oxygen environmentfor maintaining hematopoietic stem cells quiescence and thevascular niche that provides oxygenated environment for cell
proliferation and differentiation [3–5]. Ischemic injuryincreases cytokines and growth factors such as vascular endo-thelial growth factor (VEGF) in BM and circulation, which inturn activates matrix metalloproteinase (MMP)-9 and releasessoluble Kit ligand in the BM [6]. MMPs including MMP-9that is secreted mainly by neutrophils in BM [7] and MT(membrane type)1-MMP [8] that is anchored on the cell sur-face play a significant role in stem/progenitor cell mobiliza-tion and angiogenesis. Thus, understanding mechanisms bywhich ischemic injury regulates BM microenvironment andstem/progenitor cell function is essential for developing noveltherapeutic strategies for treatment of various ischemicdiseases.
Although excess amounts of reactive oxygen species(ROS) such as superoxide anion (O2
��) and hydrogen peroxide(H2O2) are detrimental to cells, ROS at appropriate levelfunction as signaling molecules to mediate normal cell
Author contributions: N.U.: conception and design, financial support, collection and/or assembly of data, data analysis and interpretation,manuscript writing, and final approval of manuscript; R.D.M.: administrative support, collection of data, provision of study material, andfinal approval of manuscript; T.F.: financial support, data analysis and interpretation, and final approval of manuscript; M.U.: financialsupport, conception and design, data analysis and interpretation, manuscript writing, and final approval of manuscript.
Correspondence: Masuko Ushio-Fukai, Ph.D., Department of Pharmacology, Center for Lung and Vascular Biology, Center forCardiovascular Research, University of Illinois at Chicago, 835 S. Wolcott, M/C868, E403 MSB, Chicago, Illinois 60612, USA. Telephone:312-996-7665; Fax: 312-996-1225; e-mail: [email protected] Received August 16, 2011; accepted for publication January 9, 2012; firstpublished online in STEM CELLS EXPRESS January 30, 2012. VC AlphaMed Press 1066-5099/2012/$30.00/0 doi: 10.1002/stem.1048
STEM CELLS 2012;30:923–934 www.StemCells.com

growth, migration, differentiation, apoptosis, and senescence[9]. Evidence suggests that redox status and ROS affect stem/progenitor cell function including cell survival and prolifera-tion [10–13] as well as interaction between stem cells andtheir niches [14]. Self-renewal and differentiation of hemato-poietic stem cells are regulated by intracellular ROS andniche microenvironment [15]. One of the major sources ofROS involved in signaling is NADPH oxidase (Nox), whichis expressed in phagocytes and nonphagocytic cells [9]. Dif-ferentiated myeloid cells such as neutrophils and monocytes/macrophages generate a large amount of O2
�� extracellularlyvia activation of Nox2 (also known as gp91phox). Nox iso-forms are also expressed in stem/progenitor cells [16], andNox-derived ROS are involved in differentiation, prolifera-tion, senescence, or apoptosis [12, 17, 18]. We previouslydemonstrated that hindlimb ischemia increases Nox2-depend-ent ROS production in isolated BM-derived mononuclear cells(BM-MNCs) and that postischemic neovascularization andmobilization of BM cells (BMCs) are impaired in Nox2�/�
mice [19]. However, the mechanisms by which Nox2 regulateprogenitor cell function and mobilization and the role ofNox2 in BM microenvironment remains unknown.
In this study, using a hindlimb ischemia model of Nox2�/�
mice and in vivo injection of O2� reactive dye, we provide
the direct evidence that ROS production is markedlyincreased in entire BM, in a Nox2-dependent manner, fol-lowing ischemic injury in vivo. We also show that Nox2-derived ROS play an important role in hindlimb ischemia-induced expansion of hypoxia, hypoxia-inducible factor-1a(HIF-1a)/VEGF expression, as well as Akt and proteolyticenzyme (MT1-MMP and MMP-9) activation in BM in situ.Furthermore, these ROS-mediated, hypoxic BM microenvir-onment alterations induced by ischemic injury regulate pro-genitor cell survival and expansion, thereby promoting theirmobilization from BM. Our study will provide new insightsinto role of Nox in redox regulation of BM microenviron-ment during postnatal neovascularization and tissue repair inresponse to injury.
MATERIALS AND METHODS
Ethics Statement for Animal Study
Study protocols were approved by the Animal Care and Insti-tutional Biosafety Committee of University of Illinois at Chi-cago (A09-066), and the experiments were performed accord-ing to the Guide for the Care and Use of Laboratory Animalsof the NIH.
Mouse Hindlimb Ischemia Model
Nox2�/� mice and their age- (range, 8–12 weeks) and sex-matched wild-type (WT) mice (C57BL/6J) were purchasedfrom Jackson Laboratories (Bar Harbor, ME, http://www.jax.org) and were fed regular chow and water containing antibiot-ics, such as sulfamethoxazole and trimethoprim (Hi-Tech Phar-macal, Amityville, NY) under specific pathogen-free conditionsat University of Illinois at Chicago. Mice were subjected to uni-lateral hindlimb surgery under anesthesia with ketamine (100mg/kg) and xylazine (10 mg/kg). To induce hindlimb ischemia,left femoral arteries were ligated and removed from the branchpoint for profunda femoris to the proximal portion of the saphe-nous artery. All arterial branches between the ligations wereobliterated using an electrical coagulator. Mice with majorbleeding or signs of infections during observation were eutha-nized. To examine the BM, we used only long bones of contra-lateral legs that were not subjected to hindlimb surgery to
exclude the direct effect of ischemia in the BM. Samples werecollected immediately after euthanasia.
O2�2 Detection in Mice
Dihydroethidium (DHE) (Invitrogen, Grand Island, NY, http://www.invitrogen.com) was prepared as a 1 mg/ml solution in1% dimethyl sulfoxide and administered at 10 mg/kg b.wt. byi.v. injection. Mice were killed and immediately perfusion-fixed with 4% paraformaldehyde 1 hour after DHE injections.Frozen sections of the BM were prepared by the direct flush-ing of BM cavities with O.C.T. compound that allowed it tokeep the structure of BM central regions and were observedby confocal microscope (Zeiss, Thornwood, NY, https://www.micro-shop.zeiss.com) with excitation at 510–550 nmand emission at >585 nm to detect oxidized ethidium. Insome experiments, frozen sections were stained with rat-originanti-CD45 or anti-Gr-1 (RB6-8C5) (eBioscience, San Diego,CA, http://www.ebioscience.com) and with DyLight 488-con-jugated anti-rat IgG angibodies (Jackson ImmunoResearchLaboratories, West Grove, PA, http://www.jacksonimmuno.com). To quantify fluorescence on captured images, a paralle-logram with size of approximately 50 � 500 lm was drawnfrom the endosteum into the BM and defined as the endostealregion. The same size of parallelogram was drawn in the mid-dle of BM as the central region. The mean fluorescence inten-sity of the regions and background signals in the area with nocell were measured on ImageJ software (NIH). The fluores-cent signal was calculated as the mean fluorescence intensitysubtracted by the background signals in each sections.
O2�2 Detection Using Isoluminol Enhanced
Chemiluminescence Assay
O2�� production in isolated Gr1þ cells was measured using iso-
luminol enhanced chemiluminescent method in 6-mm diameterwells of 96-well, flat-bottom, white tissue culture plates (E&KScientific, Campbell, CA). Freshly isolated BM Gr-1þ cellswere resuspended in Krebs-HEPES buffer. The buffer with 100lM isoluminol and 40 U/ml horseradish peroxidase (HRP) wasincubated at room temperature for 5 minutes. An aliquot (200ll) of the cells (1 � 106 cells) was added into the well andassayed for chemiluminescence (CL) at 37�C in a Wallac 1420Multilabel Counter (PerkinElmer Life Sciences, Boston, MA,http://www.perkinelmer.com). The CL counts per second wascontinually recorded for 30 minutes. To isolate Gr-1þ cellsfrom total BM cells, immunomagnetic cell sorting was per-formed using EasySep (Stem Cell Technologies, Vancouver,BC, http://www.stemcell.com) with biotinylated anti-Gr-1 anti-bodies (RB6-8C5; eBioscience). The measurement was startedwithin 1 hour after sacrificing the mice.
Hypoxia Detection by Pimonidazole
Mice were injected 60 mg/kg pimonidazole hydrochloride (Hypo-xyprobe) (Chemicon, Temecula, CA, http://www.chemicon.com)into peritoneal cavity 3 hours prior to tissue harvest. Prepa-ration of paraffin sections of decalcified femurs and immu-nolabeling of pimonidazole were performed according tothe literature [20]. Briefly, deparaffinized sections were sub-jected to antigen retrieval in 10 mM citrate buffer (pH 6.0,at 90�C for 20 minutes), incubated in 50 mM glycine inphosphate buffered saline (PBS) (pH 3.5) for 5 minutes, andpermeablized in 0.3% Triton X-100 for 15 minutes. Afterblocking, sections were incubated with mouse fluoresceinisothiocyanate (FITC)-labeled anti-pimonidazole antibody(NPI, Burlington, MA) and labeled by anti-mouse IgG con-jugated to HRP (Invitrogen). The signal was amplified usingTSATM biotin system (PerkinElmer, Waltham, MA) and la-beled by Alexa488-conjugated streptavidin (Invitrogen)
924 Role of NADPH Oxidase in Bone Marrow Microenvironment

followed by mounting with Vectashield mounting mediumwith 40,6-diamidino-2-phenylindole (Vector Laboratories, Burlin-game, CA, http://www.vectorlabs.com). Images were capturedby a Zeiss LSM 510 Meta confocal microscope with appropriatelaser and filter settings and processed by LSM510 software(Zeiss). The acquired images were measured, and the thresholdwas determined based on the histograms. The number of fluores-cence positive cells was counted using ImageJ software (NIH).
Immunostaining
Paraffin sections of decalcified femurs were labeled with anti-HIF-1a (Novus Biologicals, Littleton, CO), anti-VEGF, anti-MMP14 (MT1-MMP) (Abcam, Cambridge, MA, http://www.abcam.com), anti-phospho-Akt (Ser473) (Cell SignalingTechnology, Danvers, MA, http://www.cellsignal.com), orrabbit IgG at matched concentration. Donkey biotinylatedanti-rabbit IgG (Vector Laboratories) was used as secondaryantibody followed by TSA biotin system (PerkinElmer) forsignal enhancement, Alexa488-conjugated streptavidin (Invi-trogen) for fluorescence, or 3,30-diaminobenzidine (VectorLaboratories) for immunohistochemistry. Images for fluores-cence were captured by a confocal microscopy and processedby LSM510 software (Zeiss). For immunohistochemistory, anAxio Scope with AxioVision software (Zeiss) was used. Theacquired images were measured, and the threshold was deter-mined based on the histograms. The number of fluorescencepositive cells was counted or % of fluorescence positive areawas analyzed using ImageJ software (NIH).
Flow Cytometry
After euthanasia, legs were cut and attached muscles werestripped quickly. The femurs and tibiae were flushed with ice-cold Dulbecco’s phosphate-buffered saline supplemented with2% FCS (DPBS/FCS). Single cell suspension was obtained bygently passing through 23-gauge needles. Unfractionated cellswere stained with Lineage cocktail antibodies (Gr-1, CD11b,Ter119, B220, and CD3e) (eBioscience) and MMP14 (MT1-MMP) (Abcam). The cells were then washed, resuspended in 2lg/ml propidium iodide in DPBS/2% FCS, and analyzed by aflow cytometer (DAKO ADP Cyan) in Research ResourcesCenter in University of Illinois at Chicago. Summit (DAKONorth America, Carpinteria, CA) or FlowJo 7.6 (Tree Star, Ash-land, OR, http://www.treestar.com/) software was used for pop-ulation analysis. For apoptosis detection, FITC Annexin V Apo-ptosis Detection Kit (BD Biosciences, San Diego, CA, http://www.bdbiosciences. com) was used, as described in the manu-facturer’s instructions. Peripheral blood was drawn from retro-orbital cavities under anesthesia and was separated to form thebuffy coat. The number of circulating leukocyte or total BMcells was counted manually in the cell suspension from thebuffy coat or tibiae, respectively, using hemocytometer.
Flow Cytometric Cell Separation
Gr-1þ, Lin�, or cKitþ/Lin� cells were sorted by MoFlo fluo-rescence-activated cell sorter (DAKO Cytomation, Carpinte-ria, CA) located in the research resource center at Universityof Illinois at Chicago from BM-MNCs separated by densitygradient with Histopaque1083 (Sigma-Aldrich, St. Louis, MO,http://www.sigmaaldrich.com). Labeling was performed withPacific blue-conjugated anti-Gr-1 (RB6-8C5), biotinylatedLineage cocktail antibodies, and allophycocyanin (APC)-con-jugated anti-cKit (2B8) antibodies (eBioscience). Streptavidin-Alexa488 was used for Lineage detection.
Real-Time Polymerase Chain Reaction Analysis
Total RNA was prepared from cells using TRI Reagent (Mo-lecular Research Center, Cincinnati, OH) according to manu-
facturer’s protocol. Reverse transcription was carried outusing high-capacity cDNA reverse transcription kit (AppliedBiosystems, Carlsbad, CA, http://www.appliedbiosystems.com) or SuperScript VILO cDNA Synthesis Kit (Invitrogen).The real-time polymerase chain reactions (PCRs) were run onan ABI Prism 7000 Sequence Detection System (Applied Bio-systems) in the SYBR Green PCR kit and the QuantiTectPrimer Assay (Qiagen, Valencia, CA, http://www1.qiagen.com) for specific genes. Expression of genes was normalizedand expressed as fold changes relative to b-actin unless other-wise indicated.
Colony-Forming Unit Assay
Total BM cells (104 per well) were plated in six-well plateswith 1.5 ml of methylcellulose-based medium (Methocult(M3434), Stem Cell Technologies) and colonies (>50 cells)were scored at day 12. Colony-forming unit (CFU) was nor-malized by total BM cell number counted manually usinghemocytometer.
Gelatin Zymography
BM plasma was collected by flushing femurs with a total of0.5 ml PBS. After spinning, supernatants were analyzed formetalloproteinase activity by gelatinolytic zymography, asdescribed [21].
Statistical Analyses
Comparison between groups was analyzed by unpaired Stu-dent’s two-tailed t test (two groups) or analysis of variance(ANOVA) for experiments with more than two subgroups fol-lowed by Bonferroni post hoc analysis. p < .05 was consid-ered as statistically significant.
RESULTS
Nox2 Is Involved in Hindlimb Ischemia-InducedIncrease in O2
�2Production in the Endosteal and
Central BM In Situ
To determine the role of Nox2 in BM microenvironment dur-ing postnatal neovascularization, we examined the spatial dis-tribution of O2
�� in the BM tissue in situ in response to hind-limb ischemia in WT mice and Nox2�/� mice, which impairpostischemic angiogenesis [19, 22]. To eliminate a posthar-vesting phenomenon that may influence the measurement ofO2�� in the BM, we injected O2
�� detecting dye, DHE, into themice before sacrifice [23]. Figure 1 shows that hindlimbischemia significantly increased DHE staining in the centralBM and to lesser extent in the endosteal region defined aswithin 50 lm from the bone surface in WT mice, which werealmost completely inhibited in Nox2�/� mice. Colocalizationanalysis for DHE positive signals with hematopoietic makerCD45, myeloid maker Gr-1 in BM tissues shows that the ma-jority of ROS producing cells in BM after hindlimb ischemiawere differentiated myeloid cells (Fig. 2A). Hind limb ische-mia also significantly increased Nox2 mRNA in both BM-MNCs and cKitþLin� BM progenitor cells (Fig. 2B). Theextent of Nox2 gene expression in Gr-1þ cells was muchhigher than that of cKitþLin� cells (4.38-fold). Since wefound that Gr-1þ cells are major ROS producing cells in BMtissue in response to hindlimb ischemia, we measured ROSproduction in isolated Gr-1þ BM cells from WT and Nox2�/�
mice using isoluminol assay. Figure 2C shows that hindlimbischemia significantly increased ROS production in WT Gr-
Urao, McKinney, Fukai et al. 925
www.StemCells.com

1þ cells, which was almost completely abolished in Nox2�/�
Gr-1þ cells with significant decrease in basal ROS levels.
Hindlimb Ischemia Promotes Hypoxia Expansionand Increases HIF-1a and VEGF ExpressionThroughout the BM, in a Nox2-Dependent Manner
Since oxygen is consumed to produce O2�� by activation of
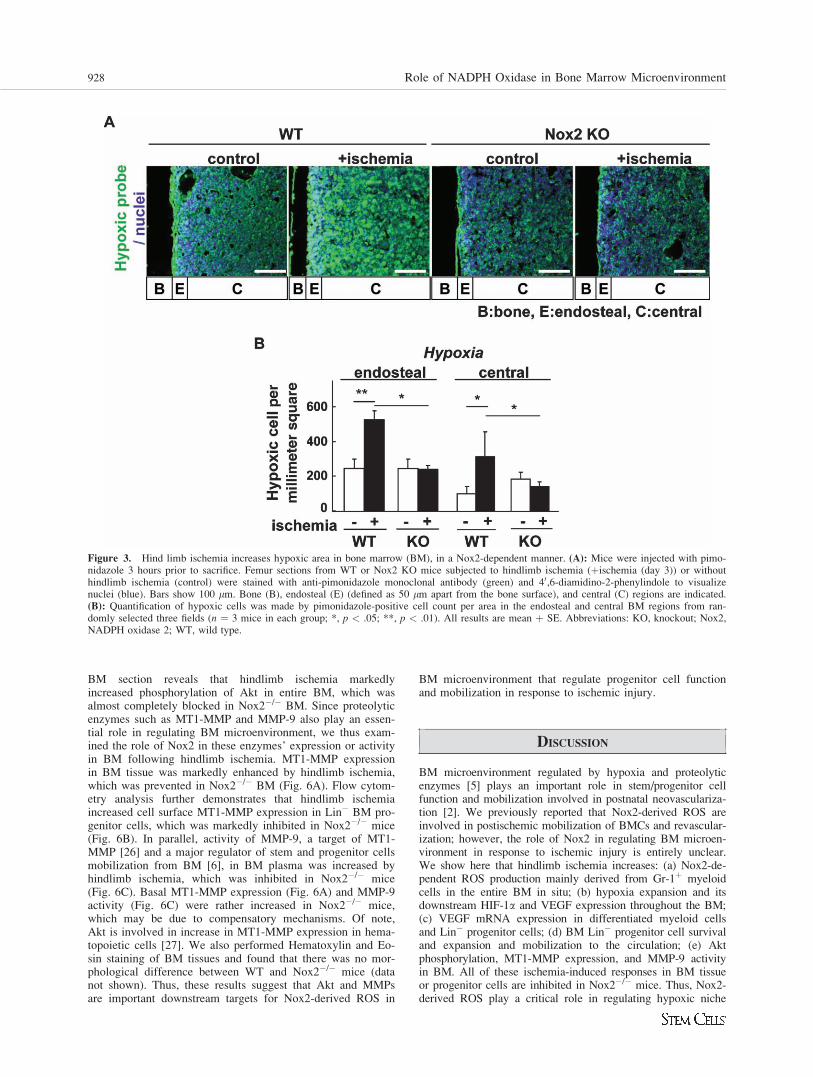
Nox, we next examined the role of Nox2 in the spatial distribu-tion of hypoxia in BM in response to hindlimb ischemia. Forthis purpose, we injected a hypoxic bioprobe, pimonidazole,which detects low O2 tension less than 1.3% [20]. Figure 3shows that without hindlimb ischemia, a majorityof pimonidazole-positive cells was preferentially incorporatedin the hypoxic endosteal area adjacent to long bonesurface while its staining is lower in the central BM area.In response to hindlimb ischemia, the hypoxic area stained bypimonidazole was expanded throughout the BM, which wasalmost completely abolished in Nox2�/� mice (Fig. 3).
Since HIF-1a is a downstream target of hypoxia, weexamined the distribution and expression of HIF-1a in theBM after hindlimb ischemia. Immunofluorescence analysis ofBM tissue revealed that HIF-1a protein was highly expressed
at the endosteum in basal state (Fig. 4A, 4C). Hind limb is-chemia increased expression of HIF-1a in both endosteal andcentral BM, which was almost completely inhibited inNox2�/� mice (Fig. 4A, 4C). These results suggest that Nox2activation may contribute to expansion of hypoxic BM micro-environment and increase in HIF-1a expression in response totissue ischemia.
We then examined the expression of VEGF protein, oneof the key HIF-a target genes inducing stem cell mobilization,using immunostaining in BM tissue. We found that VEGFexpression was increased and expanded in whole BM follow-ing hindlimb ischemia, which was almost completely inhib-ited in Nox2�/� mice (Fig. 4B, 4D). Furthermore, VEGFmRNA expression in both BM-MNC and Lin� progenitorcells was significantly upregulated following hindlimb ische-mia, which was abolished in Nox2�/� mice (Fig. 4E). Basalexpression of VEGF protein and mRNA were rather increasedin Nox2�/� BM tissue, BM-MNC, and progenitor cells pre-sumably due to compensatory mechanism. These results sug-gest that there is causal relationship between Nox2-ROS andhypoxia/HIF1-a/VEGF pathway in BM microenvironment inresponse to hindlimb ischemia.
Figure 1. Nox2 is involved in O2�� production in bone marrow (BM) in situ in response to hindlimb ischemia. (A): Representative images of
superoxide (O2��) production in BM of femur from WT or Nox2 KO mice subjected to hindlimb ischemia on the contralateral leg (þischemia
(day 3)) or without hindlimb ischemia (control). To detect O2�� in situ, DHE was injected 60 minutes before tissue harvest. Long bone surface is
shown by dotted lines. Images were taken by confocal microscopy with �20 objective lens. Bars show 100 lm. Bone (B), approximate endosteal(E) (defined as 50 lm apart from the bone surface), and central (C) BM regions are indicated. (B): A graph represents the mean fluorescence in-tensity of DHE subtracted by background signals in indicated regions of BM from randomly selected three areas from each mouse, and three in-dependent experiments were performed. All data shown are mean þ SE and *, p < .01. Abbreviations: DHE, dihydroethidium; KO, knockout;Nox2, NADPH oxidase 2; ROS, reactive oxygen species; WT, wild type.
926 Role of NADPH Oxidase in Bone Marrow Microenvironment
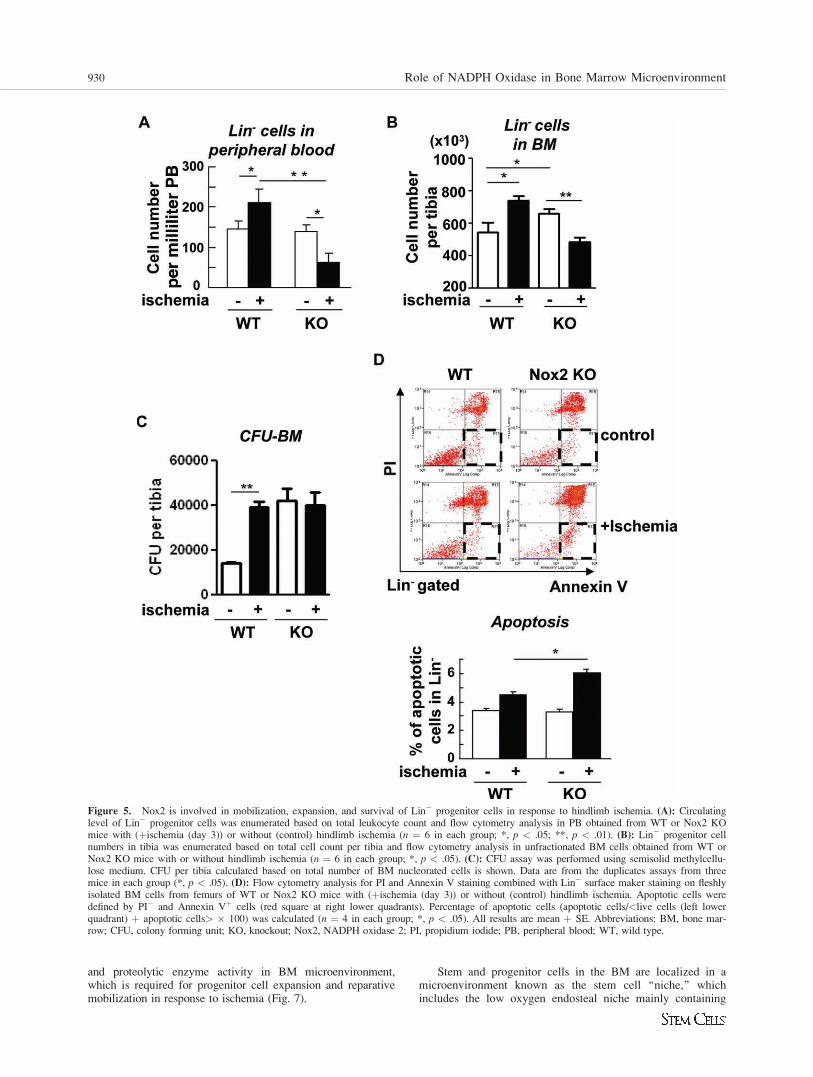
Nox2 Is Involved in Promoting Cell Expansion andSurvival Following Hindlimb Ischemia, TherebyStimulating Mobilization of Lin2 Progenitor Cells
Since hypoxia and HIF-1a in BM microenvironment may reg-ulate stem/progenitor cell function and mobilization [20], wenext examined the role of Nox2 in Lin� progenitor cell mobi-lization after hindlimb ischemia. Figure 5A shows that thenumber of circulating Lin� progenitor cells in peripheralblood was increased following hindlimb ischemia, which wassignificantly inhibited in Nox2�/� mice (Fig. 5A). Under thiscondition, the pool size of Lin� progenitor cells in the BM,as measured by flow cytometry analysis (Fig. 5B) and CFUassay (Fig. 5C), was significantly increased following hind-limb ischemia. Although the basal progenitor pool was signifi-cantly higher in Nox2�/� BM cells, ischemia-inducedenhancement of progenitor cell expansion was markedlyinhibited (Fig. 5B, 5C). Thus, Nox2-mediated increase in
hypoxic BM microenvironment may promote progenitor cellexpansion, thereby stimulating their mobilization from BM af-ter ischemic injury. To address the underlying mechanism, weexamined the role of Nox2 in cell survival and proliferationof BM Lin� progenitor cells. Flow cytometry analysis withAnnexin V and propidium iodide (PI) staining shows that ap-optotic cells (Annexin Vþ/PI�) were significantly increased inNox2�/� mice following hindlimb ischemia, as compared tothose in WT mice (Fig. 5D).
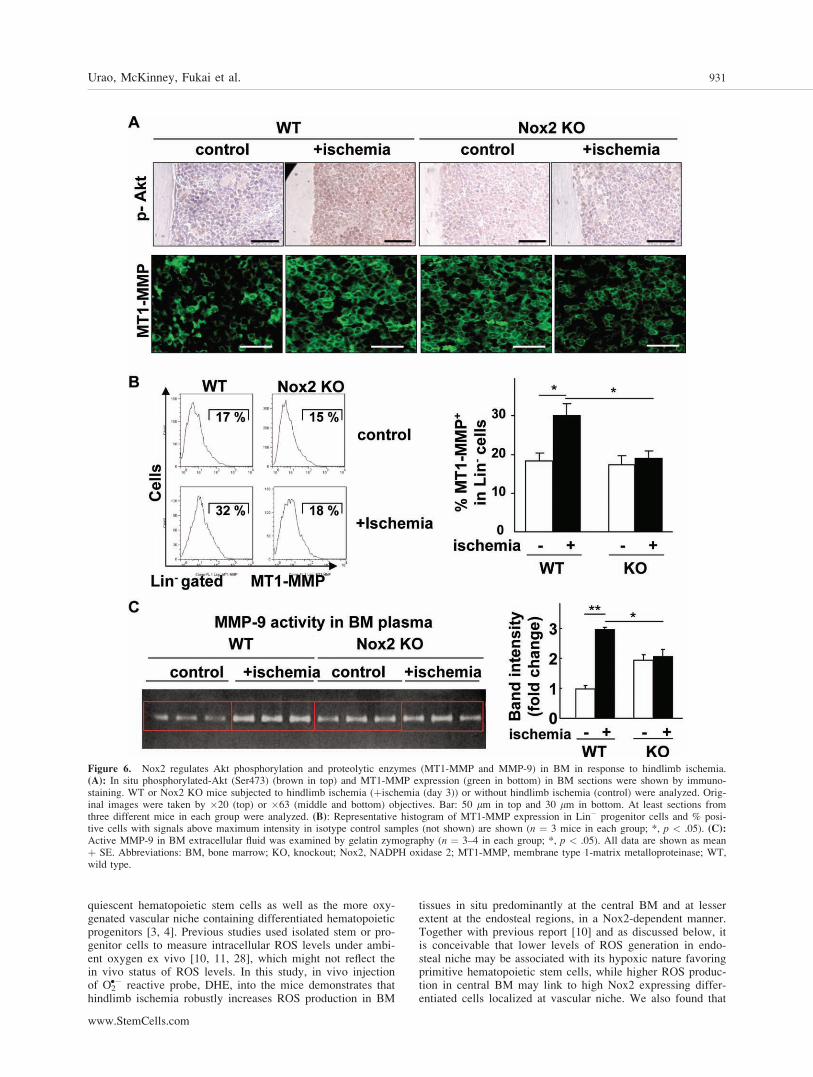
Nox2 Is Involved in Akt Phosphorylation andMMPs Activation in BM Microenvironment inResponse to Hindlimb Ischemia
Since Akt is shown to be involved in cell survival and HIF-1a induction [24, 25], we examined the role of Nox2 in Aktactivation in BM tissue following hindlimb ischemia. Asshown in Figure 6A, immunostaining with p-Akt antibody on
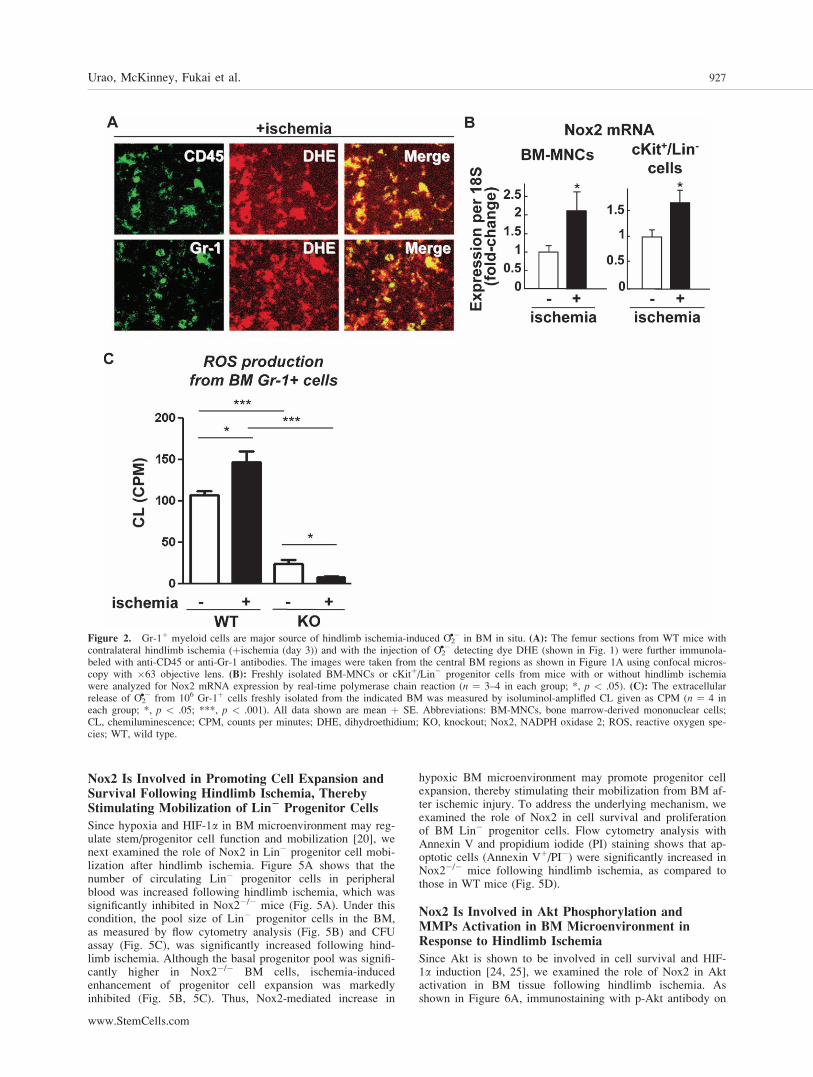
Figure 2. Gr-1þ myeloid cells are major source of hindlimb ischemia-induced O2�� in BM in situ. (A): The femur sections from WT mice with
contralateral hindlimb ischemia (þischemia (day 3)) and with the injection of O2�� detecting dye DHE (shown in Fig. 1) were further immunola-
beled with anti-CD45 or anti-Gr-1 antibodies. The images were taken from the central BM regions as shown in Figure 1A using confocal micros-copy with �63 objective lens. (B): Freshly isolated BM-MNCs or cKitþ/Lin� progenitor cells from mice with or without hindlimb ischemiawere analyzed for Nox2 mRNA expression by real-time polymerase chain reaction (n ¼ 3–4 in each group; *, p < .05). (C): The extracellularrelease of O2
�� from 106 Gr-1þ cells freshly isolated from the indicated BM was measured by isoluminol-amplifled CL given as CPM (n ¼ 4 ineach group; *, p < .05; ***, p < .001). All data shown are mean þ SE. Abbreviations: BM-MNCs, bone marrow-derived mononuclear cells;CL, chemiluminescence; CPM, counts per minutes; DHE, dihydroethidium; KO, knockout; Nox2, NADPH oxidase 2; ROS, reactive oxygen spe-cies; WT, wild type.
Urao, McKinney, Fukai et al. 927
www.StemCells.com
BM section reveals that hindlimb ischemia markedlyincreased phosphorylation of Akt in entire BM, which wasalmost completely blocked in Nox2�/� BM. Since proteolyticenzymes such as MT1-MMP and MMP-9 also play an essen-tial role in regulating BM microenvironment, we thus exam-ined the role of Nox2 in these enzymes’ expression or activityin BM following hindlimb ischemia. MT1-MMP expressionin BM tissue was markedly enhanced by hindlimb ischemia,which was prevented in Nox2�/� BM (Fig. 6A). Flow cytom-etry analysis further demonstrates that hindlimb ischemiaincreased cell surface MT1-MMP expression in Lin� BM pro-genitor cells, which was markedly inhibited in Nox2�/� mice(Fig. 6B). In parallel, activity of MMP-9, a target of MT1-MMP [26] and a major regulator of stem and progenitor cellsmobilization from BM [6], in BM plasma was increased byhindlimb ischemia, which was inhibited in Nox2�/� mice(Fig. 6C). Basal MT1-MMP expression (Fig. 6A) and MMP-9activity (Fig. 6C) were rather increased in Nox2�/� mice,which may be due to compensatory mechanisms. Of note,Akt is involved in increase in MT1-MMP expression in hema-topoietic cells [27]. We also performed Hematoxylin and Eo-sin staining of BM tissues and found that there was no mor-phological difference between WT and Nox2�/� mice (datanot shown). Thus, these results suggest that Akt and MMPsare important downstream targets for Nox2-derived ROS in
BM microenvironment that regulate progenitor cell functionand mobilization in response to ischemic injury.
DISCUSSION
BM microenvironment regulated by hypoxia and proteolyticenzymes [5] plays an important role in stem/progenitor cellfunction and mobilization involved in postnatal neovasculariza-tion [2]. We previously reported that Nox2-derived ROS areinvolved in postischemic mobilization of BMCs and revascular-ization; however, the role of Nox2 in regulating BM microen-vironment in response to ischemic injury is entirely unclear.We show here that hindlimb ischemia increases: (a) Nox2-de-pendent ROS production mainly derived from Gr-1þ myeloidcells in the entire BM in situ; (b) hypoxia expansion and itsdownstream HIF-1a and VEGF expression throughout the BM;(c) VEGF mRNA expression in differentiated myeloid cellsand Lin� progenitor cells; (d) BM Lin� progenitor cell survivaland expansion and mobilization to the circulation; (e) Aktphosphorylation, MT1-MMP expression, and MMP-9 activityin BM. All of these ischemia-induced responses in BM tissueor progenitor cells are inhibited in Nox2�/� mice. Thus, Nox2-derived ROS play a critical role in regulating hypoxic niche
Figure 3. Hind limb ischemia increases hypoxic area in bone marrow (BM), in a Nox2-dependent manner. (A): Mice were injected with pimo-nidazole 3 hours prior to sacrifice. Femur sections from WT or Nox2 KO mice subjected to hindlimb ischemia (þischemia (day 3)) or withouthindlimb ischemia (control) were stained with anti-pimonidazole monoclonal antibody (green) and 40,6-diamidino-2-phenylindole to visualizenuclei (blue). Bars show 100 lm. Bone (B), endosteal (E) (defined as 50 lm apart from the bone surface), and central (C) regions are indicated.(B): Quantification of hypoxic cells was made by pimonidazole-positive cell count per area in the endosteal and central BM regions from ran-domly selected three fields (n ¼ 3 mice in each group; *, p < .05; **, p < .01). All results are mean þ SE. Abbreviations: KO, knockout; Nox2,NADPH oxidase 2; WT, wild type.
928 Role of NADPH Oxidase in Bone Marrow Microenvironment
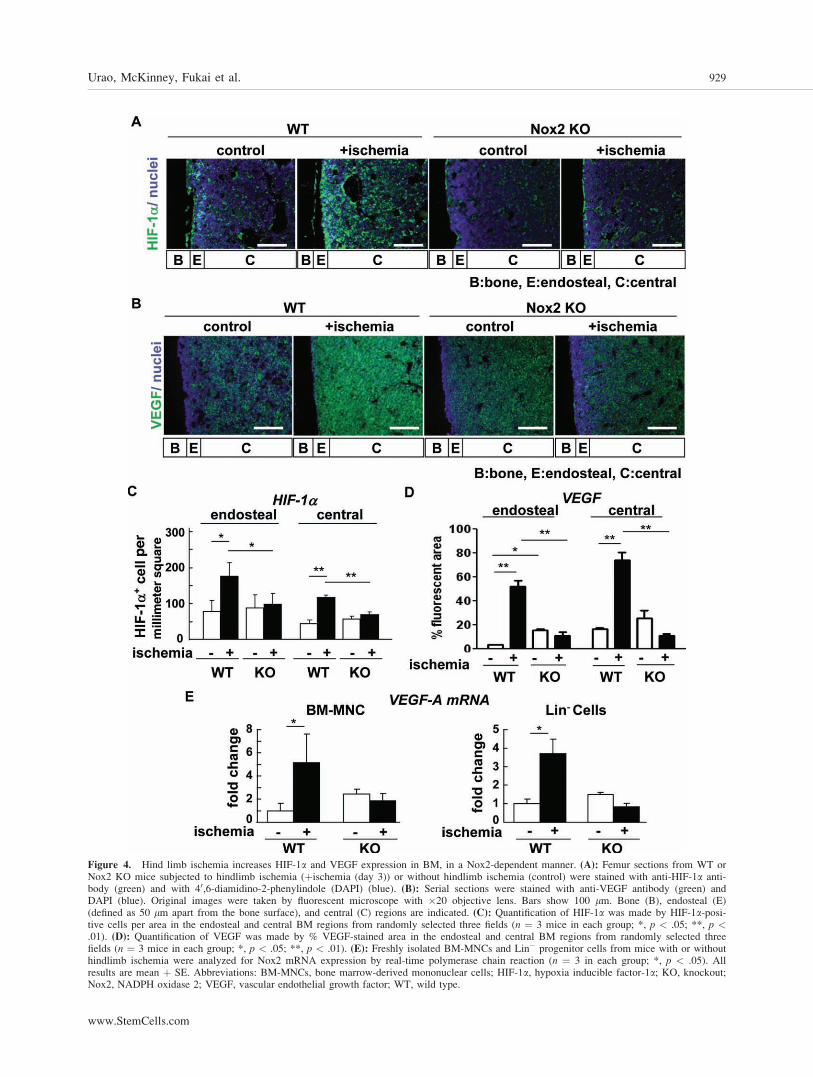
Figure 4. Hind limb ischemia increases HIF-1a and VEGF expression in BM, in a Nox2-dependent manner. (A): Femur sections from WT orNox2 KO mice subjected to hindlimb ischemia (þischemia (day 3)) or without hindlimb ischemia (control) were stained with anti-HIF-1a anti-body (green) and with 40,6-diamidino-2-phenylindole (DAPI) (blue). (B): Serial sections were stained with anti-VEGF antibody (green) andDAPI (blue). Original images were taken by fluorescent microscope with �20 objective lens. Bars show 100 lm. Bone (B), endosteal (E)(defined as 50 lm apart from the bone surface), and central (C) regions are indicated. (C): Quantification of HIF-1a was made by HIF-1a-posi-tive cells per area in the endosteal and central BM regions from randomly selected three fields (n ¼ 3 mice in each group; *, p < .05; **, p <.01). (D): Quantification of VEGF was made by % VEGF-stained area in the endosteal and central BM regions from randomly selected threefields (n ¼ 3 mice in each group; *, p < .05; **, p < .01). (E): Freshly isolated BM-MNCs and Lin� progenitor cells from mice with or withouthindlimb ischemia were analyzed for Nox2 mRNA expression by real-time polymerase chain reaction (n ¼ 3 in each group; *, p < .05). Allresults are mean þ SE. Abbreviations: BM-MNCs, bone marrow-derived mononuclear cells; HIF-1a, hypoxia inducible factor-1a; KO, knockout;Nox2, NADPH oxidase 2; VEGF, vascular endothelial growth factor; WT, wild type.
Urao, McKinney, Fukai et al. 929
www.StemCells.com
and proteolytic enzyme activity in BM microenvironment,which is required for progenitor cell expansion and reparativemobilization in response to ischemia (Fig. 7).
Stem and progenitor cells in the BM are localized in amicroenvironment known as the stem cell ‘‘niche,’’ whichincludes the low oxygen endosteal niche mainly containing
Figure 5. Nox2 is involved in mobilization, expansion, and survival of Lin� progenitor cells in response to hindlimb ischemia. (A): Circulatinglevel of Lin� progenitor cells was enumerated based on total leukocyte count and flow cytometry analysis in PB obtained from WT or Nox2 KOmice with (þischemia (day 3)) or without (control) hindlimb ischemia (n ¼ 6 in each group; *, p < .05; **, p < .01). (B): Lin� progenitor cellnumbers in tibia was enumerated based on total cell count per tibia and flow cytometry analysis in unfractionated BM cells obtained from WT orNox2 KO mice with or without hindlimb ischemia (n ¼ 6 in each group; *, p < .05). (C): CFU assay was performed using semisolid methylcellu-lose medium. CFU per tibia calculated based on total number of BM nucleorated cells is shown. Data are from the duplicates assays from threemice in each group (*, p < .05). (D): Flow cytometry analysis for PI and Annexin V staining combined with Lin� surface maker staining on fleshlyisolated BM cells from femurs of WT or Nox2 KO mice with (þischemia (day 3)) or without (control) hindlimb ischemia. Apoptotic cells weredefined by PI� and Annexin Vþ cells (red square at right lower quadrants). Percentage of apoptotic cells (apoptotic cells/<live cells (left lowerquadrant) þ apoptotic cells> � 100) was calculated (n ¼ 4 in each group; *, p < .05). All results are mean þ SE. Abbreviations: BM, bone mar-row; CFU, colony forming unit; KO, knockout; Nox2, NADPH oxidase 2; PI, propidium iodide; PB, peripheral blood; WT, wild type.
930 Role of NADPH Oxidase in Bone Marrow Microenvironment
quiescent hematopoietic stem cells as well as the more oxy-genated vascular niche containing differentiated hematopoieticprogenitors [3, 4]. Previous studies used isolated stem or pro-genitor cells to measure intracellular ROS levels under ambi-ent oxygen ex vivo [10, 11, 28], which might not reflect thein vivo status of ROS levels. In this study, in vivo injectionof O2
�� reactive probe, DHE, into the mice demonstrates thathindlimb ischemia robustly increases ROS production in BM
tissues in situ predominantly at the central BM and at lesserextent at the endosteal regions, in a Nox2-dependent manner.Together with previous report [10] and as discussed below, itis conceivable that lower levels of ROS generation in endo-steal niche may be associated with its hypoxic nature favoringprimitive hematopoietic stem cells, while higher ROS produc-tion in central BM may link to high Nox2 expressing differ-entiated cells localized at vascular niche. We also found that
Figure 6. Nox2 regulates Akt phosphorylation and proteolytic enzymes (MT1-MMP and MMP-9) in BM in response to hindlimb ischemia.(A): In situ phosphorylated-Akt (Ser473) (brown in top) and MT1-MMP expression (green in bottom) in BM sections were shown by immuno-staining. WT or Nox2 KO mice subjected to hindlimb ischemia (þischemia (day 3)) or without hindlimb ischemia (control) were analyzed. Orig-inal images were taken by �20 (top) or �63 (middle and bottom) objectives. Bar: 50 lm in top and 30 lm in bottom. At least sections fromthree different mice in each group were analyzed. (B): Representative histogram of MT1-MMP expression in Lin� progenitor cells and % posi-tive cells with signals above maximum intensity in isotype control samples (not shown) are shown (n ¼ 3 mice in each group; *, p < .05). (C):Active MMP-9 in BM extracellular fluid was examined by gelatin zymography (n ¼ 3–4 in each group; *, p < .05). All data are shown as meanþ SE. Abbreviations: BM, bone marrow; KO, knockout; Nox2, NADPH oxidase 2; MT1-MMP, membrane type 1-matrix metalloproteinase; WT,wild type.
Urao, McKinney, Fukai et al. 931
www.StemCells.com

ischemic injury-induced ROS in BM is mainly derived fromdifferentiated myeloid cells including Gr-1þ cells and thatNox2 gene expression in Gr-1þ cells is approximately 4.4-fold higher than that in cKitþLin� cells. Given the diffusiblenature of ROS, these results suggest that hindlimb ischemiaincreases ROS mainly from Gr-1þ cells in BM in a Nox2-de-pendent manner, which may create oxidative microenviron-ment to regulate BM stem cell niche.
Hypoxic microenvironment is an important determinantfor stem/progenitor function in the BM [29, 30]. Since oxygenis a source for O2
�� induced by Nox activation, we investi-gated the role of Nox2 in the spatial distribution of hypoxiain the BM following hindlimb ischemia. This study using invivo injection of hypoxic bioprobe pimonidazole, whichcross-links to protein adducts at oxygen tension below 10 mmHg (less than 1.3% O2) [20], reveals that the endosteum atthe bone BM interface is more hypoxic than the central BMin mice in the basal state. This is consistent with previousreports that the endosteal niche in steady state is hypoxic,while the central BM is more oxygenated presumably due tothe higher density of endothelial sinuses where oxygenatedblood circulates. Hypoxic endosteal niche maintains hemato-poietic stem cells in quiescent state, which requires low oxy-gen tension and ROS level, while vascular niche with highoxygen tension contain more proliferative and differentiatedstem/progenitor cells [3]. In this study, hindlimb ischemiaexpands low oxygen (hypoxic) area throughout the BM, in aNox2-dependent manner. In line with our results, granulocytecolony stimulating factor (G-CSF) or cyclophosphamide(CY), which stimulates mobilization of stem/progenitor cells,has been shown to increase hypoxia in the BM [20]. Of note,G-CSF- and CY-induced stem cell mobilization was shown tobe blunted in Nox2�/� mice [31]. In this study, the mecha-nism by which hindlimb ischemia increases hypoxia throughNox2 in contralateral legs remains unclear. It is possible thatischemic injury may increase cytokines, growth factors, neu-
rotransmitters, and metabolites such as lactates, ROS them-selves, and inflammatory cells including Gr-1þ cells that pro-duce large amounts of ROS in circulation, which mayregulate contralateral BM microenvironment. The reason whyBM becomes hypoxia by activation of Nox2 may be presum-ably due to the increase in oxygen consumption. Piccoli et al.[32] reported that the half of the oxygen consumption in he-matopoietic stem and progenitor cells is dependent on Nox. Inaddition, the consumption of respiratory burst by Nox in dif-ferentiated myeloid cells has been shown to contributeto local hypoxia [33]. Thus, Nox2 seems to be responsiblefor increased oxygen consumption and hypoxia in progenitorcells and Gr-1þ cells in mobilizing or inflammatory conditionafter hindlimb ischemia. Contribution of ROS-mediated vaso-constriction in the BM vasculature in reducing oxygen supplyalso cannot be excluded. Taken together, it is likely thathind limb ischemia increases Nox2-dependent ROS-produc-ing oxidative microenvironment, which in turn creates ahypoxic niche throughout the BM by increasing oxygenconsumption.
To address the consequence of increasing hypoxia in BMafter hindlimb ischemia, we examined the expression of HIF-1a, a key downstream target of hypoxia involved in cell sur-vival, angiogenesis [34], and hematopoietic stem cell function[15]. Immunofluorescence analysis of BM tissue sectionsshows that hindlimb ischemia increases expression of HIF-1ain entire BM tissue, in a Nox2-dependent manner. Consistentwith our finding about the role of Nox in HIF-1a level, it hasbeen shown that lipopolysaccharide increases HIF-1a expres-sion through activation of Nox in myeloid cells [35]. Intermit-tent hypoxia activates HIF-1a through Nox2, which isrequired for hypoxia-induced ROS generation [36], suggestingthat positive feed-forward mechanism among Nox2, hypoxia,and HIF-1a may exist. Of note, HIF-1a is also regulated byROS or kinase in a hypoxia-independent manner. It isreported that Nox-derived ROS promote HIF-1a stabilization
Figure 7. Nox2 regulates BM microenvironment involved in progenitor cell function and mobilization in response to tissue ischemic injury.Following ischemic injury, ROS production is increased in entire BM in a Nox2-dependent manner, which is required for increasing hypoxicniche and its downstream HIF-1a and VEGF expression throughout the BM. Ischemic injury-induced Nox2-derived ROS also increase Akt phos-phorylation and its downstream MT1-MMP (also known as MMP14) expression and MMP-9 activity in BM. It is possible that ROS and Aktpathway are involved in HIF-1a expression through hypoxia-independent mechanism. These Nox2-dependent alterations of BM microenvironmentpromote progenitor cell survival and expansion, thereby promoting their mobilization, leading to reparative neovascularization and tissue repair.Abbreviations:BM; bone marrow, HIF-1a; hypoxia inducible factor-1a; MT1-MMP, membrane type1-matrix metalloproteinase; Nox2, NADPHoxidase 2; ROS; reactive oxygen species; VEGF, vascular endothelial growth factor;
932 Role of NADPH Oxidase in Bone Marrow Microenvironment

in circulating hematopoietic stem cells by inhibiting von Hip-pel-Lindau under normoxic conditions [37]. HIF-1a is alsoregulated by Akt as discussed below. In this study, we alsofound that hindlimb ischemia increases VEGF expression, adownstream target of HIF-1a involved in stem/progenitor cellmobilization and reparative neovascularization [38] in wholeBM tissue, BM-MNCs, and Lin� progenitor cells, in a Nox2-dependent manner. Diffusible staining of VEGF may be dueto the possibility that VEGF is produced and secreted fromvarious cell types including endothelial cells, progenitor cells,and differentiated myeloid cells in BM following ischemicinjury. Most recently, Rehn et al. [39] reported that hypoxicniche regulates hematopoietic stem cell function via the hy-poxia-VEGF axis. Thus, these results suggest that Nox2-derived ROS play an important role for providing redox-sensi-tive BM microenvironment at least in part by increasingthe hypoxia/HIF-1a/VEGF pathway in response to tissueischemia.
ROS at physiological levels act as signaling molecules tomediate various biological responses including stem/progeni-tor cell function. Hypoxia and HIF-1a influence stem/progeni-tor cells expansion by regulating cell survival and prolifera-tion [15]. Stem cells can survive better under lower oxygencondition [15]. In this study, we show that Nox2 is requiredfor ischemia-induced Lin� progenitor cell expansion in BM,as measured by Lin� progenitor cell pool using flow cytome-try analysis and CFU assay. This is at least in part throughincreasing progenitor cell survival, thereby promoting theirmobilization to circulation after ischemic injury. Of note,Nox2�/� mice had more basal Lin� progenitor cell pool(higher CFU) in unfractionated BMCs, which is consistentwith previous report [31]. This may be in part through thecompensatory increase in basal expression of VEGF inNox2�/� BM (Fig. 4). We found that BM transplantation ofNox2�/� BMCs was less efficient than that from WT mice(unpublished observations), indicating that Nox2�/� hemato-poietic stem cells have lower reconstitution capacity after BMtransplantation. This may be in part due to homing defect ofNox2�/� BMCs to BM, since we observed that SDF-1-induced BMCs migration mediated through Akt is inhibited inNox2�/� mice [19]. Akt is shown to be activated by hypoxiaor ROS [40] and involved cell survival and HIF-1a expressionand activity [24, 25, 41] as well as MT1-MMP expression inhematopoietic stem cells [27]. This study shows that hindlimbischemia markedly increases phosphorylation of Akt in theBM, which is almost completely blocked in Nox2�/� BM.Thus, Akt is an important downstream target for Nox2-derived ROS in BM microenvironment regulating stem andprogenitor function and mobilization in response to ischemicinjury. It is also possible that Nox2-mediated increase in hy-poxia/HIF-1a/VEGF pathway may also indirectly activate Aktin the BM. This may represent a novel mechanism by whichNox2-derived ROS modulate stem/progenitor function by reg-ulating BM microenvironment.
To further define the mechanism underlying Nox2-de-pendent mobilization of stem/progenitor cells from BM, wealso examined the major proteolytic enzymes’ expression andactivities in the BM in WT and Nox2�/� mice. Both MT1-MMP and MMP-9 play an important role in stem/progenitorcell migration or mobilization through inhibiting adhesiveinteractions necessary to retain stem/progenitor cells withinthe BM [6]. This study demonstrates that hindlimb ischemiaincreases MT1-MMP expression in BM tissues and Lin� pro-genitor cells, in a Nox2-dependent manner. This is consistent
with previous report that G-CSF increases MT1-MMP expres-sion and decreases its endogenous inhibitor, reversion-induc-ing cysteine-rich protein with Kazal motifs (RECK), expres-sion in human CD34þ progenitor cells through Akt-dependentpathway, thereby promoting their mobilization [27]. We alsofound that hindlimb ischemia decreases RECK expression inBM tissue, in a Nox2-dependent manner (unpublished obser-vations). Furthermore, this study shows that hindlimb ische-mia increases MMP-9 activities in BM plasma, which isinhibited in Nox2�/� mice. Consistent with our data, low-dose irradiation that potentially induces ROS promotes pro-genitor mobilization through MMP-9 and VEGF production[42]. Basal MT1-MMP expression in BM tissue, but not thatin Lin� cells, as well as MMP-9 activity in BM plasma areincreased in Nox2�/� mice, which is associated with compen-satory increase in VEGF expression in BM, as describedabove. Of note, these steady state basal increases in proteo-lytic enzymes seem not to affect the number of circulatingBM progenitors in Nox2�/� mice, while hindlimb ischemia-induced responses are blunted in these mice (Fig. 6A). Thusischemic injury-induced acute increase in MT1-MMP andMMP-9 may play more important role in mobilizing progeni-tor cells to circulation. Our results are consistent with thenotion that Nox2-derived ROS play an important role in regu-lating proteolytic activity in BM in response to hindlimb is-chemia, which may facilitate the stem/progenitor cellmobilization.
CONCLUSION
In summary, this study demonstrates that ischemic injuryincreases ROS mainly derived from Gr-1þ myeloid cellsthrough Nox2 activation in entire BM, which increaseshypoxic niche and its downstream HIF-1a and VEGF expres-sion as well as Akt and proteolytic enzyme activity in BM tis-sue. These Nox2-dependent alterations of BM microenviron-ment promote Lin� progenitor cell survival and expansion,thereby promoting their mobilization to circulation, leading toreparative neovascularization and tissue repair (Fig. 7). Thisstudy should provide insights into redox regulation of BMmicroenvironment as a potential therapeutic strategy for treat-ment of various ischemic diseases [2, 13, 43-45].
ACKNOWLEDGMENTS
We thank Dr. Nihal Kaplan for technical assistance and helpfulcomments and discussions. This work was supported by fundsfrom NIH R01 Heart and Lung (HL)077524, HL077524-S1 (toM.U.-F.), and HL070187 (to T.F.); American Heart Association(AHA) Grant-In-Aid 0755805Z (to M.U.-F.), AHA NationalCenter Research Program (NCRP) Innovative Research Grant0970336N (to M.U.-F), and AHA Post-doctoral Fellowship09POST2250151 (to N.U.).
DISCLOSURE OF POTENTIAL
CONFLICTS OF INTEREST
The authors indicate no potential conflicts of interest.
Urao, McKinney, Fukai et al. 933
www.StemCells.com

REFERENCES
1 Werner N, Kosiol S, Schiegl T et al. Circulating endothelial progeni-tor cells and cardiovascular outcomes. N Engl J Med 2005;353:999–1007.
2 Dimmeler S. Regulation of bone marrow-derived vascular progenitorcell mobilization and maintenance. Arterioscler Thromb Vasc Biol2010;30:1088–1093.
3 Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches.Nat Rev Immunol 2006;6:93–106.
4 Yin T, Li L. The stem cell niches in bone. J Clin Invest 2006;116:1195–1201.
5 Kiel MJ, Morrison SJ. Uncertainty in the niches that maintain haema-topoietic stem cells. Nat Rev Immunol 2008;8:290–301.
6 Rafii S, Avecilla S, Shmelkov S et al. Angiogenic factors reconstitutehematopoiesis by recruiting stem cells from bone marrow microenvir-onment. Ann N Y Acad Sci 2003;996:49–60.
7 Page-McCaw A, Ewald AJ, Werb Z. Matrix metalloproteinases andthe regulation of tissue remodelling. Nat Rev Mol Cell Biol 2007;8:221–233.
8 Itoh Y, Seiki M. MT1-MMP: A potent modifier of pericellular micro-environment. J Cell Physiol 2006;206:1–8.
9 Ushio-Fukai M. Redox signaling in angiogenesis: Role of NADPH ox-idase. Cardiovasc Res 2006;71:226–235.
10 Jang YY, Sharkis SJ. A low level of reactive oxygen species selectsfor primitive hematopoietic stem cells that may reside in the low-oxy-genic niche. Blood 2007;110:3056–3063.
11 Tothova Z, Kollipara R, Huntly BJ et al. FoxOs are critical mediatorsof hematopoietic stem cell resistance to physiologic oxidative stress.Cell 2007;128:325–339.
12 Sardina JL, Lopez-Ruano G, Sanchez-Sanchez B et al. Reactive oxy-gen species: Are they important for haematopoiesis? Crit Rev OncolHematol 2011 [Epub ahead of Print].
13 Haneline LS. Redox regulation of stem and progenitor cells. AntioxidRedox Signal 2008;10:1849–1852.
14 Hosokawa K, Arai F, Yoshihara H et al. Function of oxidative stressin the regulation of hematopoietic stem cell–niche interaction. Bio-chem Biophys Res Commun 2007;363:578–583.
15 Suda T, Takubo K, Semenza GL. Metabolic regulation of hematopoi-etic stem cells in the hypoxic niche. Cell Stem Cell 2011;9:298–310.
16 Piccoli C, D’Aprile A, Ripoli M et al. Bone-marrow derived hemato-poietic stem/progenitor cells express multiple isoforms of NADPH ox-idase and produce constitutively reactive oxygen species. BiochemBiophys Res Commun 2007;353:965–972.
17 Fan J, Cai H, Tan WS. Role of the plasma membrane ROS-generatingNADPH oxidase in CD34þ progenitor cells preservation by hypoxia.J Biotechnol 2007;130:455–462.
18 Ushio-Fukai M, Urao N. Novel role of NADPH oxidase in angiogene-sis and stem/progenitor cell function. Antioxid Redox Signal 2009;11:2517–2533.
19 Urao N, Inomata H, Razvi M et al. Role of nox2-based NADPH oxi-dase in bone marrow and progenitor cell function involved in neovas-cularization induced by hindlimb ischemia. Circ Res 2008;103:212–220.
20 Levesque JP, Winkler IG, Hendy J et al. Hematopoietic progenitorcell mobilization results in hypoxia with increased hypoxia-inducibletranscription factor-1 alpha and vascular endothelial growth factor Ain bone marrow. Stem Cells 2007;25:1954–1965.
21 Galis ZS, Asanuma K, Godin D et al. N-Acetyl-cysteine decreases thematrix-degrading capacity of macrophage-derived foam cells: New tar-get for antioxidant therapy? Circulation 1998;97:2445–2453.
22 Tojo T, Ushio-Fukai M, Yamaoka-Tojo M et al. Role of gp91phox(Nox2)-containing NAD(P)H oxidase in angiogenesis in response tohindlimb ischemia. Circulation 2005;111:2347–2355.
23 Brennan AM, Suh SW, Won SJ et al. NADPH oxidase is the primarysource of superoxide induced by NMDA receptor activation. Nat Neu-rosci 2009;12:857–863.
24 Gao N, Ding M, Zheng JZ et al. Vanadate-induced expression of hy-poxia-inducible factor 1a and vascular endothelial growth factorthrough phosphatidylinositol 3-kinase/Akt pathway and reactive oxy-gen species. J Biol Chem 2002;277:31963–31971.
25 Choi YK, Kim CK, Lee H et al. Carbon monoxide promotes VEGFexpression by increasing HIF-1a protein level via two distinct mecha-nisms, translational activation and stabilization of HIF-1a protein. JBiol Chem 2010;285:32116–32125.
26 Han YP, Yan C, Zhou L et al. A matrix metalloproteinase-9 activationcascade by hepatic stellate cells in trans-differentiation in the three-dimensional extracellular matrix. J Biol Chem 2007;282:12928–12939.
27 Vagima Y, Avigdor A, Goichberg P et al. MT1-MMP and RECK areinvolved in human CD34þ progenitor cell retention, egress, and mobi-lization. J Clin Invest 2009;119:492–503.
28 Ito K, Hirao A, Arai F et al. Regulation of oxidative stress by ATM isrequired for self-renewal of haematopoietic stem cells. Nature 2004;431:997–1002.
29 Eliasson P, Jonsson JI. The hematopoietic stem cell niche: Low in ox-ygen but a nice place to be. J Cell Physiol 2010;222:17–22.
30 Mohyeldin A, Garzon-Muvdi T, Quinones-Hinojosa A. Oxygen instem cell biology: A critical component of the stem cell niche. CellStem Cell 2010;7:150–161.
31 van Os R, Robinson SN, Drukteinis D et al. Respiratory burst of neu-trophils is not required for stem cell mobilization in mice. Br J Hae-matol 2000;111:695–699.
32 Piccoli C, Ria R, Scrima R et al. Characterization of mitochondrialand extra-mitochondrial oxygen consuming reactions in human hema-topoietic stem cells. Novel evidence of the occurrence of NAD(P)Hoxidase activity J Biol Chem 2005;280:26467–26476.
33 Edlund J, Fasching A, Liss P et al. The roles of NADPH-oxidase andnNOS for the increased oxidative stress and the oxygen consumptionin the diabetic kidney. Diabetes Metab Res Rev 2010;26:349–356.
34 Semenza GL. Hypoxia-inducible factor 1 (HIF-1) pathway. Sci STKE2007;2007:cm8.
35 Nishi K, Oda T, Takabuchi S et al. LPS induces hypoxia-induciblefactor 1 activation in macrophage-differentiated cells in a reactive ox-ygen species-dependent manner. Antioxid Redox Signal 2008;10:983–995.
36 Yuan G, Khan SA, Luo W et al. Hypoxia-inducible factor 1 mediatesincreased expression of NADPH oxidase-2 in response to intermittenthypoxia. J Cell Physiol 2011;226:2925–2933.
37 Piccoli C, D’Aprile A, Ripoli M et al. The hypoxia-inducible factor isstabilized in circulating hematopoietic stem cells under normoxic con-ditions. FEBS Lett 2007;581:3111–3119.
38 Rey S, Semenza GL. Hypoxia-inducible factor-1-dependent mecha-nisms of vascularization and vascular remodelling. Cardiovasc Res2011;86:236–242.
39 Rehn M, Olsson A, Reckzeh K et al. Hypoxic induction of vascularendothelial growth factor regulates murine hematopoietic stem cellfunction in the low-oxygenic niche. Blood 2011;118:1534–1543.
40 Beitner-Johnson D, Rust RT, Hsieh TC et al. Hypoxia activates Aktand induces phosphorylation of GSK-3 in PC12 cells. Cell Signal2001;13:23–27.
41 Dayan F, Bilton RL, Laferriere J et al. Activation of HIF-1alpha inexponentially growing cells via hypoxic stimulation is independent ofthe Akt/mTOR pathway. J Cell Physiol 2009;218:167–174.
42 Heissig B, Rafii S, Akiyama H et al. Low-dose irradiation promotestissue revascularization through VEGF release from mast cells andMMP-9-mediated progenitor cell mobilization. J Exp Med 2005;202:739–750.
43 Krankel N, Spinetti G, Amadesi S et al. Targeting stem cell nichesand trafficking for cardiovascular therapy. Pharmacol Ther 2011;129:62–81.
44 Grek CL, Townsend DM, Tew KD. The impact of redox and thiol sta-tus on the bone marrow: Pharmacological intervention strategies. Phar-macol Ther 2010;129:172–184.
45 Kobayashi CI, Suda T. Regulation of reactive oxygen species in stemcells and cancer stem cells. J Cell Physiol 2012;227:421–430.
934 Role of NADPH Oxidase in Bone Marrow Microenvironment




















