
Olfactory Toxicity of Methimazole: Dose-Response and StructureActivity Studies and Characterization...
11
http://tpx.sagepub.com/ Toxicologic Pathology http://tpx.sagepub.com/content/23/4/477 The online version of this article can be found at: DOI: 10.1177/019262339502300404 1995 23: 477 Toxicol Pathol Mary Beth Genter, Nora J. Deamer, Bonita L. Blake, Deborah S. Wesley and Patricia E. Levi of Flavin-Containing Monooxygenase Activity in the Long-Evans Rat Olfactory Mucosa Olfactory Toxicity of Methimazole: Dose-Response and Structure-Activity Studies and Characterization Published by: http://www.sagepublications.com On behalf of: Society of Toxicologic Pathology can be found at: Toxicologic Pathology Additional services and information for http://tpx.sagepub.com/cgi/alerts Email Alerts: http://tpx.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: http://tpx.sagepub.com/content/23/4/477.refs.html Citations: by guest on July 17, 2011 tpx.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Olfactory Toxicity of Methimazole: Dose-Response and StructureActivity Studies and Characterization...

http://tpx.sagepub.com/Toxicologic Pathology
http://tpx.sagepub.com/content/23/4/477The online version of this article can be found at:
DOI: 10.1177/019262339502300404
1995 23: 477Toxicol PatholMary Beth Genter, Nora J. Deamer, Bonita L. Blake, Deborah S. Wesley and Patricia E. Levi
of Flavin-Containing Monooxygenase Activity in the Long-Evans Rat Olfactory MucosaOlfactory Toxicity of Methimazole: Dose-Response and Structure-Activity Studies and Characterization
Published by:
http://www.sagepublications.com
On behalf of:
Society of Toxicologic Pathology
can be found at:Toxicologic PathologyAdditional services and information for
http://tpx.sagepub.com/cgi/alertsEmail Alerts:
http://tpx.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://tpx.sagepub.com/content/23/4/477.refs.htmlCitations:
by guest on July 17, 2011tpx.sagepub.comDownloaded from

477
Olfactory Toxicity of Methimazole:Dose-Response and Structure-Activity Studies and
Characterization of Flavin-ContainingMonooxygenase Activity in the
Long-Evans Rat Olfactory Mucosa*1MARY BETH GENTER, NORA J. DEAMER, BONITA L. BLAKE,
DEBORAH S. WESLEY, AND PATRICIA E. LEVI
Department of Toxicology, North Carolina State University, Raleigh, North Carolina 27695-7633
* Address correspondence to: Dr. Mary Beth Genter, Depart-ment of Toxicology, North Carolina State University, Raleigh,North Carolina 27695-7633.
1 Supported by NIEHS grants ES-07046 and ES-00044 as wellas by a Research Starter Award from the Pharmaceutical Manu-facturers Association (M.B.G.). Portions of this work were pre-sented at the Nasal Toxicity and Dosimetry of Inhaled Xeno-biotics: Implications for Human Health Meeting, Durham, NorthCarolina, September 1993.
ABSTRACT
Methimazole is a compound administered to humans for the treatment of hyperthyroidism and is usedexperimentally as a model substrate for the flavin-containing monooxygenase (FMO) system. Previous resultsfrom this laboratory demonstrated that methimazole is an olfactory system toxicant, causing nearly completedestruction of the olfactory epithelium in the male Long-Evans rat following a single ip dose of 300 mg/kg.The present studies were undertaken to determine the dose-response relationship for methimazole-inducedolfactory mucosal damage and to determine whether or not similar damage occurs as a result of oral ad-ministration, mimicking the relevant route of human exposure. We also investigated the mechanism ofolfactory toxicity of methimazole by means of a structure-activity study and began the characterization ofthe form(s) of FMO present in the olfactory mucosa of the male Long-Evans rat. Dose-response analysisdemonstrated that methimazole causes olfactory mucosal damage at doses of 25 mg/kg ip and greater. Theresults of gavage studies showed that a single oral dose of 50 mg/kg also caused olfactory mucosal damage.Two structurally related compounds, methylimidazole and methylpyrrole, were not olfactory toxicants,suggesting that a reactive intermediate generated in the course of metabolizing methimazole to an S-oxideis the olfactory toxic species. Microsomal incubation studies revealed the presence ofmethimazole S-oxidationactivity in olfactory mucosal microsomes at levels comparable to those in liver. An anti-mouse liver FMOantibody reacted on Western blots with olfactory mucosal microsomes. These findings demonstrate a dose-response for the olfactory toxicity of methimazole and suggest that characterization of human olfactorymucosal FMO activity may be necessary to assess the potential for human risk associated with therapeuticexposure to methimazole.
Keywords. Flavin-containing monooxygenase (FMO); β,β’-iminodipropionitrile (IDPN); methimazole;methylpyrrole; methylimidazole; olfactory mucosa
INTRODUCTION
Methimazole is used in the treatment of hyper-thyroid conditions in humans at doses of 0.2-2 mg/kg/day po (6, 17, 31 ). The drug belongs to the thiour-
eylene class of antithyroid drugs, specifically, thethioamides (16). Methimazole is readily absorbedfrom the gastrointestinal tract and has a relativelyshort half-life in the blood (4-6 hr), but a relativelylong duration (40 hr) of action (6). Methimazole isexcreted primarily in the urine, but the presence ofa glucuronide derivative in the bile suggests ente-rohepatic circulation, as well (5). The mode of actionof methimazole is poorly understood, but it is be-lieved that the thioamide drugs inhibit the incor-poration of oxidized iodine into tyrosine residuesin thyroglobulin by binding to and inactivating thy-roid peroxidase activity (5. 6). The S-oxidation ofmethimazole is carried out by the flavin-containing
by guest on July 17, 2011tpx.sagepub.comDownloaded from

478
FIG. 1. - Structures of methimazole, MZ, and MP.
monooxygenase (FMO) system, with no apparentcontribution from the cytochrome P-450 enzymes( 19, 24, 38) and, as such, is used as a model substratefor measuring FMO-mediated metabolism in vitro.Because of the high affinity of methimazole for FMO,it can be used as a competitive inhibitor in FMOmetabolism studies (19).
#,#’-iminodipropionitrile (IDPN) is among a
growing list of compounds that have been found tocause degeneration of the olfactory mucosa of thenasal cavity following systemic (i.e., noninhalation)exposure (12). The secondary amine nitrogen ofIDPN is reportedly an FMO substrate, and theN-hydroxylated form of IDPN is hypothesized tobe the neurotoxic metabolite (23). Therefore, weused methimazole (300 mg/kg ip) in order to try tocompetitively inhibit the FMO-mediated metabo-lism of IDPN and thereby decrease the olfactorytoxicity of IDPN. Instead of achieving this goal, wediscovered that methimazole itself is an olfactorymucosal toxicant, causing complete destruction ofthe olfactory epithelium in the male Long-Evans rat(14).Methimazole (Fig. 1) is ultimately metabolized to
N-methylimidazole and sulfite. In the metabolic
process, it is believed that FMO enzymes oxidizemethimazole to intermediate metabolites ofsulfenic
(R-SOH) and sulfinic (R-S02H) acids (19, 24, 38);several toxic effects associated with methimazole areattributed to these intermediate metabolites (19, 24).Previous reports have suggested that glutathionerapidly reduces the sulfenic acid metabolite ( 19, 24).The studies reported here were undertaken to de-
termine the dose-response relationship for the ol-factory toxicity of methimazole and to determinewhether or not the same endpoint was observedfollowing oral administration (to mimic the routeof human exposure). We also attempted to deter-mine whether or not the FMO-mediated S-oxida-tion was responsible for the olfactory toxicity ofmethimazole by means of a structure-activity studyand initiated the characterization of the form(s) ofFMO present in the olfactory mucosa of the maleLong-Evans rat.
METHODS
Animals
Male Long-Evans rats (8-10 wk of age) were pur-chased from Charles Rivers Laboratories (Raleigh,NC) and maintained under conditions consistentwith National Institutes of Health guidelines (34).Rats were housed 2-3/cage in standard plastic cages,6.5 x 9.5 x 18.5 in. Rats were maintained on a 12:12-hr photoperiod and had free access to food (Pro-lab Animal Diet, Agway, Syracuse, NY) and tapwater. Temperature and relative humidity weremaintained in the range of 19-23°C and 20-60%,respectively. Studies were approved prior to incep-tion by the North Carolina State University Insti-tutional Animal Care and Use Committee, and an-imal numbers were minimized to the extent possi-ble.
Reagents. Reagents used were of the highestquality commercially available. Methimazole anddimethyl sulfoxide (DMSO) were obtained fromSigma Chemical Co. (St. Louis, MO). Methylpyrrole(MP) and methylimidazole (MZ) were purchasedfrom Aldrich Chemical Co. (Milwaukee, WI). Rab-bit anti-mouse liver FMO antiserum was elicited inrabbits from FMO purified from mouse liver as pre-viously described (29, 35).Methimazole ip Dose-Response Study. Groups
of 2 rats each received 0 (DMSO vehicle), 2, 25, 50,100, 150, or 300 mg/kg methimazole ip. Rats weresacrificed 32 hr later by C02 asphyxiation. Nasalcavities were prepared for histopathology as de-scribed later.Methimazole Gavage Dose-Response Study.
Groups of 3 rats each were used in the gavage study,except for the absolute and vehicle (com oil) controlgroups, which had 2 rats each. Rats received a singledose of 0, 2, 5, 10, 25, or 50 mg/kg of methimazoleand were sacrificed 32 hr later by C02 asphyxiation.Nasal cavities were prepared for histopathology asdescribed later.Methimazole Structure-Activity Relationship
Study. Both MZ and MP (Fig. 1) were initiallyadministered ip in corn oil at 0.88 mmol/kg (equiv-alent to the 100 mg/kg dose of methimazole as al-ready mentioned; n = 2/group). When these animalssurvived with no apparent adverse effects, a secondgroup of rats received 2.63 mmol/kg of either MZor MP (equivalent to the 300 mg/kg dose of meth-imazole ; n = 3/group). All rats treated with meth-imazole analogs were sacrificed 24 hr later and nasalcavities were prepared for histopathology as de-scribed next.
Histopathology. For all the preceding studies,heads were removed and prepared for nasal cavityhistopathology as previously described (12). Briefly,
by guest on July 17, 2011tpx.sagepub.comDownloaded from

479
nasal cavities were flushed with 10% neutral-buf-fered formalin and immersion-fixed for 24 hr in thesame fixative. Tissues were then transferred to 8 Lof 10% formic acid containing Rexyn 101 ion ex-change resin (Fisher Scientific) for 7 days. Afterwashing in running tap water for 12 hr, the decal-cified tissues were sectioned as previously described(37), dehydrated through graded alcohols, and em-bedded in paraffin using standard techniques. TheLevel 2 section (containing predominantly respi-ratory epithelium, with squamous epithelium liningthe floor of the nasal cavity and olfactory epitheliumlining the dorsal meatus of the nasal cavity) and theLevel 3 section (taken through the ethmoid turbi-nates at the approximate level of the septal windowand lined predominantly by olfactory epithelium)were examined by light microscopy. Five-Am sec-tions were stained with hematoxylin and eosin (H&E)for evaluation.Morphometry of ip Dose-Response Lesions. The
fraction of lesioned olfactory mucosa in Level 3nasal cavity sections (37) from the rats in the ipdose-response study was quantitated with the aidof a Zeiss videoplan. The basement membrane un-derlying damaged and intact regions of the olfactorymucosa was traced directly from the H&E-stainedsections. The fraction of the olfactory epitheliumdamaged by methimazole administration was ex-pressed as the length of basement membrane un-derlying damaged epithelium divided by the totallength of the basement membrane underlying theolfactory epithelium in that nasal cavity section.Calibration was achieved with a 2-mm stage mi-crometer.
Microsome PreparationControl rat and mouse olfactory mucosal and liv-
er microsomes prepared in conjunction with con-current studies in this laboratory ( 11 ) were used forthe present in vitro studies. Microsomes were storedin a 250-mM sucrose buffer at -80°C until use.Protein concentrations were determined using thebicinchoninic acid assay (32).FMO Activity. FMO activity was determined by
monitoring the rate of methimazole oxidation at412 nm at 37°C using an Aminco DW-2a double-beam spectrophotometer (9). Nasal and liver mi-crosomes were incubated at pH 8.4 at a final con-centration of 0.025 mg/ml in a reaction mixturecomposed of 0.1 M tricine, 1.0 mM EDTA, 0.06mM 5,5’-dithiobis(2-nitrobenzoate), 0.02 mM di-thiotheitol, and 0.1 mM NADPH; 1.0 mM methi-mazole was added to begin the reaction. The ex-tinction coefficient of 28,200 M-1 cm- (19) wasused to calculate nmol of product formed/min/mgprotein.
Western Blot Analysis. Liver and nasal micro-somal proteins were separated on a 4-15% poly-acrylamide minigel and transferred to nitrocelluloseas previously described (11). Rabbit anti-mouse liv-er FMO antiserum was applied at a 1:1,000 dilution,and antibody binding was visualized using a Vec-tastain kit (Vector Laboratories. Burlingame, CA),with H202 and diaminobenzidine tetrahydrochlo-ride as the substrate and chromagen, respectively.
Statistical Methodology. Analysis of variance andt-test were performed on the morphometric datausing JMP In software (SAS Institute, Cary, NC).The desired level of significance (p < 0.05) was cor-rected for multiple t-test comparisons (Bonferronicorrection), resulting in use of p < 0.01 for deter-
mining statistically significant differences amongtreatment groups.
RESULTS
HistopathologyLesion Characterization. Olfactory mucosal
damage was the endpoint of interest in these studies,and olfactory mucosal degeneration resulted fromboth the oral and ip routes of exposure. The lesion,regardless of its distribution within the nasal cavity,was characterized by complete sloughing of the neu-ronal and sustentacular cell components of the ep-ithelium in the affected areas, with the basementmembrane and basal cells remaining intact (Fig. 2).The subepithelial Bowman’s glands were also af-fected, characterized by degeneration of the acinarportion of both the superficial and deep Bowman’sglands. No lesions were observed in the subepithe-lial nerve bundles (Fig. 2).
Dose-Response Analysis. The susceptibility of theLong-Evans rat to the olfactory toxic effects ofmeth-imazole varied with the route of exposure. The &dquo;noobserved adverse effect level&dquo; for olfactory mucosaldamage was 2 mg/kg by ip administration (Table I).In contrast, in the gavage study, olfactory mucosaldamage was only noted at 50 mg/kg, the highestdose administered (Table I).
Morphorrtetry. Morphometric analysis of themethimazole-induced (ip) lesions demonstrated thatthe extent of olfactory mucosal damage in responseto different doses of methimazole varied with thedose administered (Fig. 3). Administration of a sin-gle 300-mg/kg ip dose resulted in nearly 100% ol-factory mucosal loss in Level 3 nasal cavity sections.The fraction of the mucosa damaged diminished asdose decreased (Fig. 4), with the lesion affecting pri-marily the epithelium lining of the dorsal medialmeatus of the nasal cavity in rats receiving the lowerdoses of methimazole. As shown in Fig. 4. the low-dose lesion affected the dorsal septum and the me-
by guest on July 17, 2011tpx.sagepub.comDownloaded from

480
FIG. 2.-Light micrograph showing control (a) and methimazole-induced olfactory mucosal damage (b). a) Structuralcomponents of normal olfactory mucosa are indicated: DB = deep Bowman’s glands; N = neuronal layer; NB = olfactorynerve bundles; S = sustentacular cell layer. SB = acinar cells of superficial Bowman’s glands; Basal cells are indicatedby arrowheads. H&E. 40 x. b) Sustentacular cells and olfactory neurons are completely sloughed off 24 hr after admin-istration of methimazole, while basal cells (arrowheads) and the basement membrane remain intact. The subepithelialareas typically occupied by Bowman’s glands appears vacuolate (*). This micrograph is typical of the damage seen,regardless of the location of the lesion within the nasal cavity or the dose of methimazole that was administered.
by guest on July 17, 2011tpx.sagepub.comDownloaded from
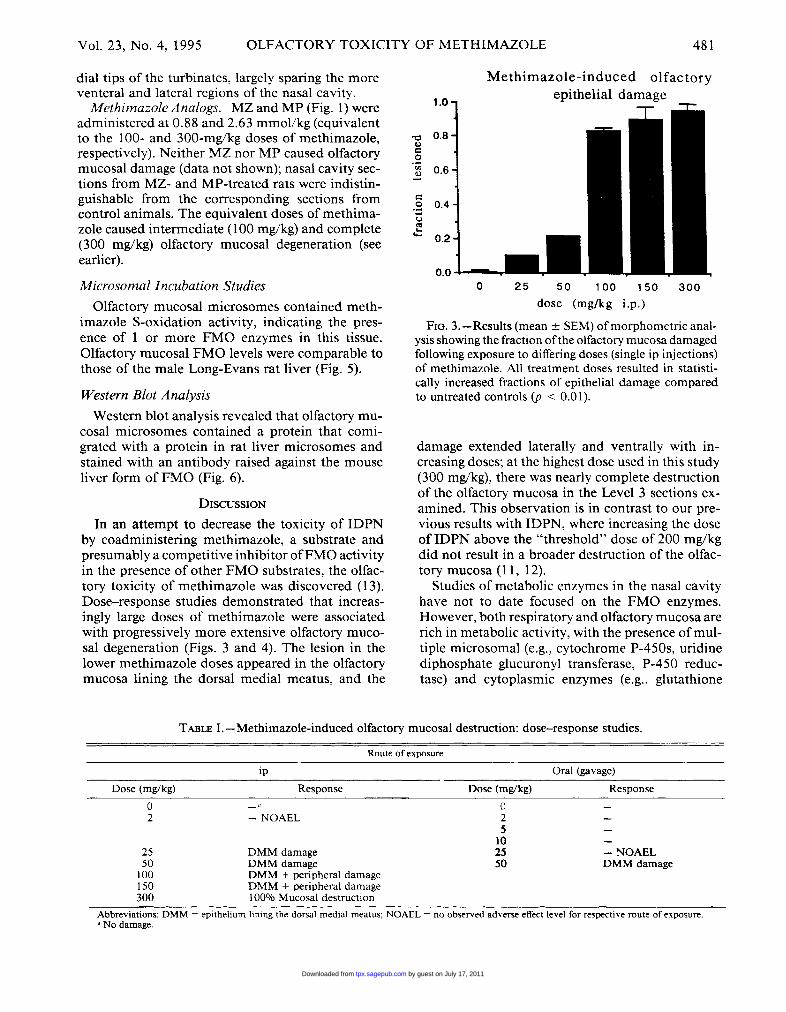
481
dial tips of the turbinates, largely sparing the moreventeral and lateral regions of the nasal cavity.MethimazoleAnalogs. MZ and MP (Fig. 1) were
administered at 0.88 and 2.63 mmol/kg (equivalentto the 100- and 300-mg/kg doses of methimazole,respectively). Neither MZ nor MP caused olfactorymucosal damage (data not shown); nasal cavity sec-tions from MZ- and MP-treated rats were indistin-
guishable from the corresponding sections fromcontrol animals. The equivalent doses of methima-zole caused intermediate (100 mg/kg) and complete(300 mg/kg) olfactory mucosal degeneration (seeearlier).
Microsomal Incubation Studies
Olfactory mucosal microsomes contained meth-imazole S-oxidation activity, indicating the pres-ence of 1 or more FMO enzymes in this tissue.
Olfactory mucosal FMO levels were comparable tothose of the male Long-Evans rat liver (Fig. 5).
Western Blot AnalysisWestern blot analysis revealed that olfactory mu-
cosal microsomes contained a protein that comi-grated with a protein in rat liver microsomes andstained with an antibody raised against the mouseliver form of FMO (Fig. 6).
DISCUSSION
In an attempt to decrease the toxicity of IDPNby coadministering methimazole, a substrate andpresumably a competitive inhibitor of FMO activityin the presence of other FMO substrates, the olfac-tory toxicity of methimazole was discovered (13).Dose-response studies demonstrated that increas-ingly large doses of methimazole were associatedwith progressively more extensive olfactory muco-sal degeneration (Figs. 3 and 4). The lesion in thelower methimazole doses appeared in the olfactorymucosa lining the dorsal medial meatus, and the
FIG. 3.-l2esults (mean ± SEM) of morphometric anal-ysis showing the fraction of the olfactory mucosa damagedfollowing exposure to differing doses (single ip injections)of methimazole. All treatment doses resulted in statisti-
cally increased fractions of epithelial damage comparedto untreated controls (p < 0.01).
damage extended laterally and ventrally with in-creasing doses; at the highest dose used in this study(300 mg/kg), there was nearly complete destructionof the olfactory mucosa in the Level 3 sections ex-amined. This observation is in contrast to our pre-vious results with IDPN, where increasing the doseof IDPN above the &dquo;threshold&dquo; dose of 200 mg/kgdid not result in a broader destruction of the olfac-
tory mucosa (11, 12).Studies of metabolic enzymes in the nasal cavity
have not to date focused on the FMO enzymes.However, both respiratory and olfactory mucosa arerich in metabolic activity, with the presence of mul-tiple microsomal (e.g., cytochrome P-450s, uridinediphosphate glucuronyl transferase, P-450 reduc-tase) and cytoplasmic enzymes (e.g., glutathione
TABLE L-Methimazole-induced olfactory mucosal destruction: dose-response studies.
Abbreviations: DMM = epithelium lining the dorsal medial meatus; NOAEL = no observed adverse effect level for respective route of exposure.a No damage.
by guest on July 17, 2011tpx.sagepub.comDownloaded from

FIG. 4.-Low-power light micrographs of H&E-stained transverse sections from male Long-Evans rat nasal cavitiesafter receiving a single ip dose of methimazole: 300 mg/kg (a), 100 mg/kg (b), 25 mg/kg (c), and 0 mg/kg (d). Note thatthe extent of the olfactory mucosal damage decreases as the dose of methimazole is reduced. Whereas nearly all of theolfactory mucosa is destroyed in Level 3 sections in rats receiving 300 mg/kg (note the absence of any intact epithelium
by guest on July 17, 2011tpx.sagepub.comDownloaded from

in this micrograph) (a), a dose of 25 mg/kg results in a more limited lesion (c). affecting primarily the mucosa lining thedorsal medial airways (*). The nasal septum (S) is indicated. Boundaries of olfactory mucosal damage are delineated byarrows in (b) and (c). H&E. x 8.
by guest on July 17, 2011tpx.sagepub.comDownloaded from
484
FIG. 5.-FMO activity in male Long-Evans rat liverand olfactory mucosal microsomes. Data represent themean value of duplicate incubations (± SD).
S-transferases, rhodanese, esterases, dehydrogen-ases) well documented (7, 25). Lesion developmentin the nasal cavity has been attributable to multiplefactors, often acting in conjunction with each other.For example, airflow patterns, chemical reactivity,and solubility are responsible for the deposition andthe ultimate development of lesions in the anteriornasal cavity as a result of inhalation exposure tosuch compounds as chlorine gas, formaldehyde, andglutaraldehyde (15, 22, 36). It is also recognized thata growing number of compounds cause olfactorymucosal damage following noninhalation routes ofexposure. Carbon tetrachloride caused olfactorysensory cell loss following gavage administration(11). The herbicide active ingredient dichlobenil (2,6-dichlorobenzonitrile) causes olfactory mucosaldamage and increased olfactory bulb glial fibrillaryacidic protein levels following dermal exposure(8). Still others (e.g., 3,3’-iminodipropionitrile,3-methylindole, 2,6-dichlorobenzonitrile, nitrosa-mines, bromobenzene, acetaminophen) cause olfac-tory mucosal damage following iv or ip injection (2,4, 10, 12, 25). While the role of blood flow in re-gional deposition of such compounds as the mech-anism underlying the site specificity of lesion de-velopment is currently under investigation (18), ithas been observed that several metabolic enzymeshave distinct patterns of localization in the mucosa
lining the main routes of airflow through the nasalcavity; therefore, the hypothesis that such com-pounds cause olfactory mucosal damage as a resultof site-specific binding and bioactivation has beenproposed (3, 1 l, 13).The FMO enzymes are microsomal enzymes that
are primarily responsible for oxidative metabolismof phosphorous, nitrogen, and sulfur atoms; meth-
FIG. 6. - Western blot showing FMO protein in rat liverand olfactory mucosal (OM) microsomes (3 gg protein/lane). Arrows (Lane 1 ) indicate position of prestained mo-lecular weight markers (49.5 and 80 kDa). Lane 2: rat OMmicrosomes; Lane 3: rat OM microsomes + rat liver mi-crosomes. Note the presence of an approximately 69.8-kDa protein in Lane 2, which stains strongly with the anti-liver FMO antibody and which appears to comigrate witha rat liver FMO protein (Lane 3).
imazole has long been known to be a high-affinitysubstrate for the FMO enzymes (19). Five forms ofFMO enzymes have been characterized and havebeen identified in multiple tissues including lung,liver, kidney, and brain ( 1, 20, 21 ). Previous studieshave demonstrated the presence of FMO in rabbit
olfactory mucosal microsomes (27, 30). Here wedemonstrate that the male Long-Evans rat olfactorymucosa likewise contains both FMO protein andmetabolic activity; the isozyme(s) of FMO respon-sible for the activity appears, based on Western blotanalysis, to be one of the forms of FMO also foundin the rodent liver. Based on the current nomencla-ture for the different isoforms of FMO, the formthat appears to be present based on the Western blotresults herein is Form 1 (20, 21 ).Methimazole is believed to be metabolized pri-
marily by FMO-mediated sulfoxidation (24), andMZ is a primary metabolite (19). Sulfenic and sul-finic acids are believed to be reactive intermediatemetabolites produced in the process of convertingmethimazole to MZ; we hypothesize that these re-active intermediates are responsible for the toxicityseen in the olfactory mucosa. This hypothesis isstrengthened by the fact that neither MP nor MZresulted in olfactory toxicity. Previous work withmethimazole S-oxidation reactions in vitro revealedthat reactive intermediates are produced, and thesecan be eliminated by the addition of glutathione tothe reaction mixture (19, 24, 28).
In summary, methimazole causes degeneration ofthe rat olfactory in a dose-related manner. The low-est dose of methimazole associated with olfactorymucosal degeneration in these studies was 25 mg/kg (by ip injection). The risk of olfactory mucosaldegeneration in humans taking this compound ther-apeutically can be estimated only after establishingthe pharmacokinetics of the compound in humans
by guest on July 17, 2011tpx.sagepub.comDownloaded from

485
and determining the FMO content of the humanolfactory mucosa.
ACKNOWLEDGMENT
We thank Elizabeth Gross Bermudez (ChemicalIndustry Institute of Toxicology, RTP, NC) for as-sistance with morphometric analyses.
REFERENCES .
1. Blake BL, Philpot RM, Levi PE, and Hodgson E(1995). Identification of novel FMO4 expression inrabbit brain. Biochem. Biophys. Res. Comm. (in press).
2. Brandt I, Brittebo EB, Feil VJ, and Bakke JE (1990).Irreversible binding and toxicity of the herbicidedichlobenil (2,6-dichlorobenzonitrile) in the olfac-tory mucosa of mice. Toxicol. Appl. Pharmacol. 103:491-501.
3. Brittebo EB (1987). Metabolic activation of phenac-etin in rat nasal mucosa: Dose-dependent binding tothe glands of Bowman. Cancer Res. 47: 1449-1456.
4. Brittebo EB, Eriksson C, and Brandt I (1990). Acti-vation and toxicity of bromobenzene in nasal tissuesin mice. Arch. Toxicol. 64: 54-60.
5. Connors JM (1990). Thyroid and antithyroid drugs.In: Modern Pharmacology, CR Craig and RE Stitzel(eds). Little, Brown and Co., Boston, pp. 910-923.
6. Cooper DS, Bode HH, Nath B, Saxe V, Maloof F,and Ridgway EC (1984). Methimazole pharmacologyin man: Studies using a newly developed radioim-munoassay for methimazole. J. Clin. Endocrinol.Metab. 58: 473-479.
7. Dahl AR and Hadley WH (1991). Nasal cavity en-zymes involved in xenobiotic metabolism: Effects onthe toxicity of inhalants. CRC Crit. Rev. Toxicol. 21:345-372.
8. Deamer NJ, O’Callaghan JP, and Genter MB (1994).Olfactory toxicity resulting from dermal applicationof 2,6-dichlorobenzonitrile (dichlobenil) in the C57BImouse. Neurotoxicology 15: 287-294.
9. Dixit A and Roche TE (1984). Spectrophotometricassay of the flavin-containing monooxygenase andchanges in its activity in female mouse liver withnutritional and diurnal conditions. Arch. Biochem.Biophys. 233: 50-63.
10. Emeigh Hart SG, Cartun RW, Wyand DS, KhairallahEA, and Cohen SD (1995). Immunohistochemicallocalization of acetaminophen in target tissues of theCD-1 mouse: Correspondence of covalent bindingwith toxicity. Fundam. Appl. Toxicol. 24: 260-274.
11. Genter MB, Deamer NJ, Cao Y, and Levi PE (1994).Effects of P450 inhibition and induction on the ol-
factory toxicity of β,β’-iminodipropionitrile (IDPN)in the rat. J. Biochem. Toxicol. 9: 31-39.
12. Genter MB, Llorens J, O’Callaghan JP, Peele DB,Morgan KT, and Crofton KM (1992). Olfactory tox-icity of β,β’-iminodipropionitrile (IDPN) in the rat.J. Pharmacol. Exp. Therap. 263: 1432-1439.
13. Genter MB, Owens DM. and Deamer NJ (1995). Dis-tribution of microsomal epoxide hydrolase and glu-tathione S-transferase in the rat olfactory mucosa:Relevance to distribution of lesions caused by sys-temically-administered olfactory toxicants. Chem.Senses (in press).
14. Genter MB, Zainal TA, and Deamer NJ (1994). Ol-factory toxicity of methimazole: Results of intraper-itoneal and oral dosing studies. Inh. Toxicol. 6(suppl.):433-437.
15. Gross EA, Mellick PW, Kari FW, Miller FJ, and Mor-gan KT (1994). Histopathology and cell replicationresponses in the respiratory tract of rats and miceexposed by inhalation to glutaraldehyde for up to 13weeks. Fund. Appl. Toxicol. 23: 348-362.
16. Haynes RC and Maurad F (1985). Thyroid and an-tithyroid drugs. In: The Pharmacological Basis ofTherapeutics, 7th ed., AG Gilman, LS Goodman, TWRall, and F Murad (eds). Macmillan Publishing, NewYork, pp. 1408-1411.
17. Jansson R, Dahlberg PA, Johansson H, and Lind-strom B (1983). Intrathyroidal concentrations ofmethimazole in patients with Graves’ disease. J. Clin.Endocrinol. Metab. 57: 129-132.
18. Joyner DR, Fleishman A, Gross EA, and Morgan KT(1995). Use of glycol methacrylate sections for de-termination of regional distribution of intravascular-ly-administered microspheres for study of regionalnasal blood perfusion in the rat. Presented at theannual meeting of the North Carolina Society of Tox-icology, Research Triangle Park, NC, February 1995.
19. Kedderis GL and Rickert DE (1985). Loss of rat livermicrosomal cytochrome P-450 during methimazolemetabolism: Role of flavin-containing monooxygen-ase. Drug Metab. Disposition 13: 58-61.
20. Lawton MP, Cashman JR, Cresteil T, Dolphin CT,Elfarra AA, Hines RN, Hodgson E, Kimura T, OzolsJ, Phillips IR, Philpot RM, Poulson LL, Rettie AE,Shephard EA, Williams DE, and Ziegler DM (1994).A nomenclature for the mammalian flavin-contain-
ing monooxygenase gene family based on amino acidsequence identities. Arch. Biochem. Biophys. 308: 254-257.
21. Lawton MP and Philpot RM (1993). Molecular ge-netics of the flavin-dependent monooxygenases.Pharmacogenetics 3: 40-44.
22. Monticello TM, Miller FJ, and Morgan KT (1991).Regional increases in rat nasal epithelial cell prolif-eration following acute and subchronic inhalation offormaldehyde. Toxicol. Appl. Pharmacol. 111: 409-421.
23. Morandi A, Gambetti P, Arora PK, and Sayre LM(1987). Mechanism of neurotoxic action of β,β’-im-inodipropionitrile (IDPN): N-hydroxylation enhanc-es neurotoxic potency. Brain Res. 437: 69-76.
24. Poulsen LL, Hyslop RM. and Ziegler DM (1979).S-oxidation of N-substituted thioureas catalyzed bythe pig liver microsomal FAD-containing monooxy-genase. Arch. Biochem. Biophys. 198: 78-88.
25. Rangga-Tabbu C and Sleight SD (1992). Sequentialstudy in rats of nasal lesions induced by N-nitroso-
by guest on July 17, 2011tpx.sagepub.comDownloaded from

486
dimethylamine and N-nitorsopyrrolidine. Fundam.Appl. Toxicol. 19: 147-156.
26. Reed CJ (1993). Drug metabolism in the nasal cavity:Relevance to toxicology. Drug Metab. Rev. 25: 173-205.
27. Sabourin PJ and Dahl AR (1985). Distribution oftheFAD-containing monooxygenase in respiratory tracttissues. In: Annual Report LMF-114, MA Medinskyand BA Muggenburg (eds). National Technical In-formation Service, Springfield, Virginia, pp. 156.
28. Sabourin PJ and Hodgson E (1984). Characterizationof the purified microsomal FAD-containing mono-oxygenase from mouse and pig liver. Chem. Biol.Interact. 51: 125-139.
29. Sabourin PJ, Smyser BP, and Hodgson E (1984). Pu-rification of the flavin-containing monooxygenasefrom mouse and pig liver microsomes. Int. J. Bio-chem. 16: 713-720.
30. Shehin-Johnson SE, Williams DE, Larsen-SuS, Stres-ser DM, and Hines RN (1995). Tissue-specific ex-pression of flavin-containing monooxygenase (FMO)forms 1 and 2 in the rabbit. J. Pharmacol. Exp. Ther-ap. 272: 1293-1299.
31. Shiroozu A, Okamura K, Ikenoue H, Sato K, Na-kashima T, Yoshinari M, Fujishima M, and Yoshi-zumi T (1986). Treatment of hyperthyroidism witha small single daily dose of methimazole. J. Clin.Endocrinol. Metab. 63: 125-128.
32. Smith PK, Krohn RI, Hermanson GT, Mallia AK,
Gartner FH, Provenzano MD, Fujimoto EK, GoekeNM, Olson BJ, and Klenk DC (1985). Measurementof protein using bicinchoninic acid. Anal. Biochem.150: 76-85.
33. Turk MAM, Flory W, and Henk WG (1986). Chem-ical modulation of 3-methylindole toxicosis in mice:Effect on bronchiolar and olfactory mucosal injury.Vet. Pathol. 23: 563-570.
34. U.S. Department of Health and Human Services(1985). Guide for the Care and Use of LaboratoryAnimals. Animal Resources Program, Division ofResearch Resources, National Institutes of Health,Bethesda, MD.
35. Venkatesh K, Levi PE, and Hodgson E (1991). Theflavin-containing monooxygenase of mouse kidney:A comparison with the liver enzyme. Biochem. Phar-macol. 42: 1211-1420.
36. Wolf DC, Morgan KT, Gross EA, Barrow C, MossOR, James AR, and Popp JA (1995). Two-year in-halation exposure of female and male B6C3F1 miceand F344 rats to chlorine gas induces lesions confinedto the nose. Fund. Appl. Toxicol. 24: 111-131.
37. Young JT (1981). Histopathologic examination of therat nasal cavity. Fund. Appl. Toxicol. 1 : 309-312.
38. Ziegler DM (1993). Recent studies on the structureand function of multisubstrate flavin-containingmonooxygenases. Annu. Rev. Pharmacol. Toxicol. 33:179-199.
by guest on July 17, 2011tpx.sagepub.comDownloaded from




















