
Functional neuroimaging of the olfactory system in humans
17
Ž . International Journal of Psychophysiology 36 2000 165]181 Functional neuroimaging of the olfactory system in humans David H. Zald a,b, U , Jose V. Pardo a,b ´ a Cogniti ¤ e Neuroimaging Unit, Psychiatry Ser ¤ ice, Veterans Affairs Medical Center, 1 Veterans Dri ¤ e, Minneapolis, MN 55417, USA b Di ¤ ision of Neuroscience Research, Department of Psychiatry, Uni ¤ ersity of Minnesota, Minneapolis, MN 55455, USA Received 4 July 1999; received in revised form 5 October 1999; accepted 12 October 1999 Abstract Ž . Ž . Positron emission tomography PET and functional magnetic resonance imaging fMRI have begun to provide unique information regarding the neural underpinnings of olfactory functioning in humans. We review the relative strengths and weaknesses of PET and fMRI techniques for studying olfaction. We then review PET and fMRI studies relating to the olfactory functions of the pyriform cortex, orbitofrontal cortex, amygdala and the entorhinalrhippocampal region. A pixelwise correlational analysis of PET data is also presented in order to clarify the relationship between blood flow in the medial temporal lobes and psychoperceptual variables. Q 2000 Elsevier Science B.V. All rights reserved. Keywords: Amygdala; Entorhinal cortex; Functional MRI; Gyrus rectus; Olfaction; Orbitofrontal; Positron emission tomography; Pyriform cortex 1. Introduction Over the past decade, positron emission tomog- Ž . raphy PET and functional magnetic resonance U Corresponding author. Tel.: q1-612-725-2000, ext. 2473; fax: q1-612-725-2249. Ž . E-mail address: [email protected] D.H. Zald Ž . imaging fMRI have begun to detail the functio- nal neuroanatomy of the olfactory system. Al- though the number of PET and fMRI studies of Ž olfaction remains extremely small especially . compared to studies of other sensory modalities , these studies have nevertheless begun to provide unique insights into the neural correlates of ol- factory processing in humans. Here, we describe the major strengths and weaknesses of PET and 0167-8760r00r$ - see front matter Q 2000 Elsevier Science B.V. All rights reserved. Ž . PII: S 0 1 6 7 - 8 7 6 0 99 00110-5
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Functional neuroimaging of the olfactory system in humans

Ž .International Journal of Psychophysiology 36 2000 165]181
Functional neuroimaging of the olfactory system inhumans
David H. Zalda,b,U, Jose V. Pardoa,b´a Cogniti e Neuroimaging Unit, Psychiatry Ser ice, Veterans Affairs Medical Center, 1 Veterans Dri e, Minneapolis,
MN 55417, USAbDi ision of Neuroscience Research, Department of Psychiatry, Uni ersity of Minnesota, Minneapolis, MN 55455, USA
Received 4 July 1999; received in revised form 5 October 1999; accepted 12 October 1999
Abstract
Ž . Ž .Positron emission tomography PET and functional magnetic resonance imaging fMRI have begun to provideunique information regarding the neural underpinnings of olfactory functioning in humans. We review the relativestrengths and weaknesses of PET and fMRI techniques for studying olfaction. We then review PET and fMRIstudies relating to the olfactory functions of the pyriform cortex, orbitofrontal cortex, amygdala and theentorhinalrhippocampal region. A pixelwise correlational analysis of PET data is also presented in order to clarifythe relationship between blood flow in the medial temporal lobes and psychoperceptual variables. Q 2000 ElsevierScience B.V. All rights reserved.
Keywords: Amygdala; Entorhinal cortex; Functional MRI; Gyrus rectus; Olfaction; Orbitofrontal; Positron emission tomography;Pyriform cortex
1. Introduction
Over the past decade, positron emission tomog-Ž .raphy PET and functional magnetic resonance
U Corresponding author. Tel.: q1-612-725-2000, ext. 2473;fax: q1-612-725-2249.
Ž .E-mail address: [email protected] D.H. Zald
Ž .imaging fMRI have begun to detail the functio-nal neuroanatomy of the olfactory system. Al-though the number of PET and fMRI studies of
Žolfaction remains extremely small especially.compared to studies of other sensory modalities ,
these studies have nevertheless begun to provideunique insights into the neural correlates of ol-factory processing in humans. Here, we describethe major strengths and weaknesses of PET and
0167-8760r00r$ - see front matter Q 2000 Elsevier Science B.V. All rights reserved.Ž .PII: S 0 1 6 7 - 8 7 6 0 9 9 0 0 1 1 0 - 5

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181166
fMRI approaches to the study of olfaction, anddetail some of the insights produced by thesemethods. In particular, we focus on data regard-ing the olfactory related functions of the pyriform
Ž .cortex, orbitofrontal cortex OFC and the amyg-dala.
2. Methodological issues
2.1. PET
PET studies of olfaction typically utilize thebolus H 15O technique for assaying changes in2
Ž .regional cerebral blood flow rCBF . BecauserCBF increases in conjunction with neural activ-ity, rCBF provides an index of dynamic changes inneural activity. PET techniques possess severalstrengths. First, they allow assessment of neuralactivity throughout most, if not all, of the brainsimultaneously. Second, PET imaging providesexcellent visualization of activity within the ven-tral temporal and frontal lobes, where primary
Žand secondary olfactory cortices reside these ar-eas are difficult to measure with most other psy-
.chophysiological techniques . Current PETcameras typically possess a mapping resolutioncapable of localizing a single focus to within a
Ž .couple of millimeters Fox et al., 1986 . However,Žspatial blurring which is applied in the data
analysis phase of most PET studies in order toimprove the signal-to-noise ratio and statistical
.properties of the data prevents the distinction ofseparate foci less than 10]12 mm apart. Thedistance at which two separate foci can be distin-guished is referred to as the Full-Width at Half-
Ž .Maximum FWHM . The FWHM of PET fre-quently makes it difficult to distinguish foci fallingwithin proximal areas of the brain.1 For instance,if a focus falls within the pyriform cortex andanother focus falls in the neighboring periamyg-dalaloid cortex, the two foci may blur together,making it difficult to distinguish the two separateareas of activation. This may present a particular
1Some researchers use even higher levels of blurring, pro-ducing FWHM of 20 mm.
problem in the olfactory domain, since severaldistinguishable olfactory regions lie in adjacentareas of the medial temporal lobe.
Three additional limitations of PET warrantconsideration. The first issue relates to temporalresolution. Most bolus H 15O PET studies collect2data over 60]90-s scan periods, with the peaksensitivity occurring in the first 30]40 s of thescan. This large temporal window precludes stud-ies of single discrete odorant exposures. Instead,subjects must receive either a prolonged exposureor repeated discrete exposures during the scanperiod. With prolonged or repeated exposures tothe same odorant, the long temporal window mayprevent detection of areas that only show activity
Žat the beginning of odorant exposure i.e. areas.sensitive to rapid adaptation or habituation . This
is a major limitation given the well-documentedprocesses of adaptation in olfactory perceptionŽKoster and de Wik, 1991; Cometto-Muniz and¨ ˜
.Cain, 1995 . These problems are reduced withexposure to multiple odorants. However, studieswith multiple odorants can prove problematic be-cause of inconsistencies in the psychoperceptualcharacteristics of different odorants within a sin-gle scan period. They also do not totally eliminateadaptation, since cross-adaptation can occur suchthat exposure to one odorant leads to a decrease
Žin the response or a higher threshold of detec-. Žtion to a second odorant Koster and de Wik,¨
.1991; Cometto-Muniz and Cain, 1995 . On the˜other hand, the relatively long duration of scansreduces the importance of accurately synchroniz-ing the timing of stimulus presentations. Thus,
Ž .whereas, evoked response potential ERP , mag-Ž .netoencephalogram MEG , and some fMRI
studies require synchronized olfactometric stimu-lus presentation, less precise stimulus presenta-tion techniques can be effectively utilized withPET.
A second limitation of PET studies relates tothe exposure to radioactive isotopes. Although,the doses are low and highly controlled, the sub-ject’s exposure must be limited to a discrete num-ber of scans. In most PET centers, subjects partic-ipate in 8]16 scans in a study. By repeating two
Ž .conditions a stimulation and a control conditionmultiple times, it becomes possible to perform

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 167
single-subject analyses with PET. However, mostinvestigators design studies with many differentstimulation and control conditions, thus limitingthe ability to repeat conditions multiple times andprecluding the ability to perform single-subjectanalyses. Because of this, most PET studies con-centrate on group data analyses, and only occa-sionally provide information about the variabilityof responses. A final pragmatic limitation relatesto the inaccessibility of PET. The number of PETcenters remains quite limited. Furthermore, theprocedures can be relatively costly due to theneed to produce radionuclides on site, and themedical personnel necessary to inject the ra-dionuclides.
2.2. fMRI
fMRI measures changes in blood oxygen levelŽ .dependent BOLD signal to assess the functional
status of different brain regions. The BOLD sig-nal reflects changes in the relative proportion ofoxyhemoglobin and deoxyhemoglobin that occurswhen a brain region increases its blood flow dur-
Žing increased neural activity Ogawa et al., 1990,.1992 . fMRI possesses several key advantages over
PET. First, it is highly accessible and much moreaffordable than PET imaging. Second, since itdoes not involve radiation exposure, subjects canbe scanned repetitively, allowing clear-cut single-subject analyses. Third, its spatial resolution often
Žexceeds that of PET although in many studies,the filtering, inter-subject variability and imageacquisition parameters produce resolutions thatare only slightly better than that produced bycurrent PET cameras, and issues related to signalmislocalization due to draining vein effectsnegates some of the advantages gained by higher
.resolution . The temporal resolution also providesa great improvement over PET. In studies utiliz-ing alternating stimulation blocks )20 s each,the improvements in temporal resolution providelittle benefit relative to PET. However, the highertemporal resolution becomes a tremendous aid instudies measuring responses to single trials and instudies directly probing the time course of activa-tions. Such studies have already begun to provideinformation that would not have been available
Žwith PET for example see Sobel et al., 1998a,b;.Sobel et al., in press .
fMRI techniques also possess a number of dis-advantages relative to PET. The most significantof these disadvantages relates to the susceptibilityof ventral frontal and temporal brain regions toartifacts. Such artifacts arise due to the differentmagnetic susceptibility of brain tissue and the
Ž .underlying bone and air Frahm et al., 1988 .Depending upon the specific imaging parameters,the field inhomogeneity in these regions can pro-duce signal dropout, geometric distortions, alias-
Žing, blurring and smearing Yudilevich and Stark,.1987; Farzaneh et al., 1990; Ojemann et al., 1997 .
These problems are so severe in the orbitofrontalŽ .cortex OFC that the area is virtually unmeasur-
Ž .able in many fMRI studies Ojemann et al., 1997 .Pulsatile and gross movement artifacts can alsomake it difficult to image this region. In fact,pulsatile and other subject movement can causesignal changes that exceed those produced by
Ž .actual activation Glover and Lee, 1995 . Pulsatileartifacts pose particular problems for olfactorystudies due to their link to respiration. The fail-ure of some fMRI studies to observe activity inregions such as the OFC likely reflects theseproblems. That is not to say that these areascannot be imaged with fMRI. A number of tech-niques appear to alleviate these problems. Forinstance, local shimming and post-processing canreduce some of the artifacts associated with field
Ž . 2inhomogeneities Morrell and Spielman, 1997 .Alignment of scans parallel to the orbital plane
Ž .and utilization of thin slices Young et al., 1988can also reduce these problems by limiting thedegree to which slices include areas with dramati-cally different field properties. Other artifacts,such as those associated with pulsatile movement,can be alleviated by using special image acquisi-tion parameters, such as spiral pulse sequencesŽ .Glover and Lee, 1995; Noll et al., 1995 . Thus,while application of fMRI techniques to the study
2 Local shimming allows more optimal viewing of the infe-rior frontal lobe, but typically requires the sacrifice of datafrom other areas of the brain.

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181168
of olfaction present a number of unique chal-lenges, these problems can be overcome withcareful attention to imaging methodology.
3. Imaging studies of olfaction
3.1. Primary olfactory cortex
Ž .The primary olfactory cortex POC is locatedwithin the pyriform region at the junction of the
Ž .temporal and frontal lobes Price, 1991 . Thepyriform cortex wraps around this junction, ex-tending into an anterior section of the temporallobe’s medial wall and a posterior section of the
Ž .ventral frontal lobe. Zatorre et al. 1992 were thefirst researchers to demonstrate odorant-inducedactivity in the pyriform cortex of humans. Sub-jects smelled a range of odorants relative to acontrol condition in which they were told to
Žbreathe as if trying to smell but knew that noodorants would be presented in the control condi-
.tion . rCBF increases emerged near the inferiorjunction of the frontal and temporal lobes bilater-ally. Similar bilateral activity arose in the area ofthe pyriform cortex when the same subjects re-
Ž .ceived monorhinal right nostril stimulation withŽ .odorants Zatorre and Jones-Gotman, in press .
This bilateral response likely reflects the recipro-cal connections between the two pyriform corticesthat course through the anterior commissureŽ .Haberly and Price, 1978; Luskin and Price, 1983 .The apparent bilateral activation induced bymonorhinal stimulation warrants caution wheninterpreting studies that attempt to make stricthemispheric distinctions based on right vs. leftnostril stimulation in healthy individuals. OtherPET studies confirm the ability of odorants toincrease activity in, or near, the pyriform cortexŽ .Small et al., 1997; Dade et al., 1998 . Neverthe-less, this activity has not been completely consis-
Ž .tent. For instance, Zald and Pardo 1997a onlysaw small non-significant increases in this regionwhen comparing odor exposure to a no-odor con-dition. Several fMRI studies have also failed toobserve substantial pyriform activations during
Ž .olfaction Yousem et al., 1997; Sobel et al., 1998a .More provocatively, a recent PET study by Dade
Ž .et al. 1998 suggests that subtle cognitive fea-tures may influence the extent to which pyriformactivity emerges in subtraction analyses. Theseauthors observed pyriform activation when sub-jects recognized previously presented odorantsŽ .relative to a no-odor baseline , yet the samesubjects had failed to demonstrate rCBF in-creases in the pyriform cortex when memorizingŽ .encoding the odors during an earlier scan. Sinceboth conditions involved the same odorants, theseresults suggest that novelty, odor recognition orother cognitive aspects of the task influence thedegree of pyriform activity. These findings aresurprising when contrasted with other primarysensory regions which show large and consistentresponses to sensory stimulation that remainrobust regardless of subtle changes in task condi-tions. Moreover, secondary olfactory cortices ap-pear capable of showing strong rCBF increases,
Ževen in cases where the POC does not Zaldand Pardo, 1997a; Royet et al., unpublished
.manuscript .Three major factors may contribute to the in-
consistent ability to observe pyriform activity. Thefirst factor reflects a simple methodological prob-lem, related to the technical difficulties involved
Žin imaging this region i.e. inhomogeneity and.sensitivity to movement artifacts in fMRI studies .
However, this factor alone, cannot explain theinconsistency, since negative results have emergedfrom PET studies and fMRI studies using ap-propriate methodology.
A second factor, may relate to the timing ofresponses in the POC. For instance, if cells in thisarea show transient or rapidly adapting responses,the time course of many fMRI and PET studieswould blur together periods with increased signaland periods with no increased signal. Such tem-poral blurring would thus dilute any signal in-creases. By examining the time course of fMRI
Ž .responses, Sobel et al. in press have providedevidence that the pyriform cortex shows transientresponses to odorants which occur at the start ofodorant exposure but return to baseline duringprolonged exposures. For instance, these authorsnoted a significant increase in pyriform BOLD
Žsignal during an initial 40-s block of five odorant.exposures, separated by 8 s , with only non-

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 169
significant increases on later blocks. Furthermore,signal increases appeared greatest during the veryinitial odorant trial, with transient increases alsooccurring at each switch between odorant andnon-odorant conditions. This pattern of transientand rapidly habituating responses is consistentwith electrophysiological studies of POC activity
Ž .in animals Wilson, 1998 . These findings indicatethat POC activity neither corresponds to the timecourse of olfactory exposure or the time course ofolfactory perception.
The third source of difficulty in observing pyri-form responses to odorants may relate to thesensitivity of pyriform cortex to the mechanicalmovement of nasal hairs caused by sniffing. ThePOC has traditionally been conceptualized as aunimodal sensory region dedicated exclusively toolfaction. However, using Fourier frequency
Ž .transformations of fMRI data, Sobel et al. 1998ademonstrated that the pyriform cortex shows ac-tivity that occurs synchronous with sniffing orairflow through the nose. Especially at high levels
Ž .of airflow as occurs during sniffing , this activityappears large enough to obscure responses to
Ž .odorants. Indeed, Sobel et al. 1998a found onlylimited additional pyriform activity when compar-ing odorant and no-odorant conditions, and the
Ž .volume of pyriform cortex i.e. number of voxelsshowing olfactory responses appeared small rela-
Žtive to those showing sniff-related activity Sobel.et al., in press . This raises the possibility that the
pyriform activity observed in PET studies of ol-faction in part reflects differences in breathingbetween conditions. While most studies of olfac-tion attempt to match breathing patterns acrossconditions, few have actively controlled or docu-mented that airflow through the nostrils was in-deed equivalent across conditions. The finding of
Ž .Sobel et al. 1998a demands the careful controlof sniffing and nasal air flow during olfactorytasks. The ability of sniffing to induce activity inthe POC may help prime olfactory processing.Alternatively, sniff-induced activity may reflectPOC processing of intranasal somatosensory in-formation. This latter possibility remains unex-plored. If true, it would contradict the commonlyheld assumption that somatosensory informationfrom the nose is only transmitted through the
trigeminal nerve’s ramifications in the primarysomatosensory cortex. Taken together with PETdata on the effects of different cognitive condi-tions on pyriform processing, such findings mayrequire major modifications in the conceptualiza-tion of the POC.
3.1.1. Orbitofrontal cortexThe OFC forms the ventral surface of the
frontal lobe. The OFC represents a cytoarchitec-turally, and functionally heterogeneous regionthat evolved as part of a series of concentric rings
Žaround the POC Carmichael and Price, 1994;.Zald and Kim, 1996a,b . The POC and the ante-
rior olfactory nucleus project extensively to theposterior-medial OFC and the agranular regionalong the border between the extreme posterior
Ž .OFC and inferior insula Carmichael et al., 1994 .A lighter projection also reaches a more central
Ž .portion of the OFC Carmichael et al., 1994 .Other OFC regions receive olfactory informationsecondary to the posterior OFC as well as receiv-ing substantial olfactory projections via themediodorsal nucleus of the thalamus, pars magno-
Žcellularis Yarita et al., 1980; Ruschen et al.,1987; Ray and Price, 1993; Carmichael and Price,
.1996In monkeys, the OFC has been described as a
secondary cortical association area for the olfac-Ž .tory system Tanabe et al., 1975a,b; Takagi, 1986 .
Neurophysiological studies indicate that cells inthe OFC show highly selective responses to olfac-
Žtory stimuli Tanabe et al., 1975a; Takagi, 1986;.Critchley and Rolls, 1996 . Indeed, many, if not
the majority, of olfactory responsive cells in themonkey OFC show far greater stimulus selectivitythan cells in the pyriform cortex and olfactorybulb. Consistent with this pattern of stimulusselectivity, lesions to the OFC in humans induceimpairments in olfactory discrimination and iden-
Žtification Jones-Gotman and Zatorre, 1988; Za-.torre and Jones-Gotman, 1991 .
The OFC consistently demonstrates increasedŽrCBF in PET studies of olfaction Zatorre et al.,
1992; Small et al., 1997; Zald and Pardo, 1997a;Royet et al., 1999; Royet et al., unpublished
.manuscript; Zatorre and Jones-Gotman, in press .OFC activity has also been observed during olfac-

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181170
Ž .See facing page for legends .

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 171
Žtory perception in fMRI studies Levy et al., 1997;.Yousem et al., 1997; Sobel et al., 1998a , although
Ž .only Sobel et al. 1998a provide detailed informa-tion about the specific localization of foci. Sur-prisingly, OFC responses actually occur withgreater consistency than pyriform responses inthese studies.
Analysis of patients with OFC lesions indicatea right OFC dominance for certain types of olfac-tory processing. Specifically, right OFC lesionsgenerally produce greater deficits in olfactory
Žprocessing than left OFC lesions Zatorre andJones-Gotman, 1991; Jones-Gotman and Zatorre,
.1993 , although significant olfactory identificationdeficits arise following OFC lesions in either
Ž .hemisphere Jones-Gotman and Zatorre, 1988 .This pattern of right OFC dominance is alsoevident in the neuroimaging literature. Both PETand fMRI studies of olfactory stimulation report
Ž .more frequent or stronger right OFC activationsŽthan left OFC activations Zatorre et al., 1992;
Small et al., 1997; Yousem et al., 1997; Sobel etal., 1998a; Francis et al., 1999; Royet et al., 1999;
.Dade et al., 1998 . The major exception to thispattern is a study of aversive olfaction in whichsignificant left OFC foci emerged, while onlynon-significant activations localized to the right
Ž .OFC Zald and Pardo, 1997a . Although thesource of this difference remains undetermined,it is difficult to dismiss it as simply reflecting
Ž .hedonic pleasantnessrunpleasantness factors,since the same subjects demonstrated a similarpattern of lateralization during exposure to pleas-
Žant odorants Zald and Pardo, unpublished obser-.vations .
A second issue relates to the specific locations
of olfactory related foci within the OFC. Electro-physiological studies in non-human primates sug-gest the presence of separate lateral and central
Žolfactory zones in the posterior OFC Tanabe et.al., 1975b; Yarita et al., 1980; Takagi, 1986 . Be-
Ž .cause most PET studies and some fMRI studiesprovide localizing information within a common
Žcoordinate system i.e. Talairach coordinates; Ta-.lairach and Tournoux, 1988 it becomes possible
to provide information regarding the consistencyof localization across neuroimaging studies. Re-gardless of hemisphere, a relatively central focusis clearly identifiable in most of these studieswwhich falls between 17 and 30 mm lateral to themidline, and 28]45 mm rostral to the anterior
Ž .xcommissure see Fig. 1a . Anatomically, thisplaces most of the peaks in the intermediate orposterior orbital gyrus, although in the righthemisphere this area may extend laterally into
Ž .the lateral orbital gyrus Sobel et al., 1998a .Cytoarchitecturally, this central olfactory area ap-pears most consistent with area 13, although insome cases the peak appears near the boundaryof areas 13 and 11.3
A lateral olfactory responsive area also emerges
3The Brodmann system for labelling the OFC fails tocapture the cytoarchitectural heterogeneity of the OFC. Brod-
Žmann labels most of the OFC as area 11 with area 47 forming. Ž .the inferior frontal gyrus pars orbitalis Brodmann, 1914 .
Ž . Ž .However, Beck 1949 along with Petrides and Pandya 1994 ,identify a posterior medial OFC region which they both labelas area 13 and which appears structurally homologous to
Ž .Walker’s area 13 in the monkey Walker, 1940 . The centralolfactory responsive focus in humans falls in, or near, theborder of this area.
Ž . Ž .Fig. 1. a The central olfactory region in the left OFC. The image derives from Zald and Pardo 1997a and displays an area ofŽsignificant rCBF increase during exposure to unpleasant stimuli from the University of Pennsylvania Smell Identification Test Doty
.et al., 1984 relative to a no-odor control condition. The PET data were thresholded to show only significant activation and thenŽ . Ž .superimposed upon transverse slice zsy16 of a structural MRI. b The left lateral olfactory region in the same subjects during
Ž .exposure to aversive sulfides. The peak of this focus falls slightly inferior to this slice zsy9 during the aversive sulfides conditionbut in other studies we frequently observe activity at this level. The more posterior foci in this slide correspond to the superior endof the amygdalar and insular foci displayed in Fig. 3.Fig. 2. Connections between the olfactory system and the amygdala. Dashed lines show connections that have either beeninconsistently reported in primates, or have only been clearly demonstrated in rodents andror cats. Abbre¨iations: aCo } anteriorcortical nucleus; Endp } endopyriform nucleus; Med } medial nucleus; Nlot } nucleus of the lateral olfactory tract; PAC }
periamygdaloid cortex; and pCo } posterior cortical nucleus.

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181172
Žin some olfactory studies Zald and Pardo, 1997a;.Royet et al., 1999 . This focus falls in the poste-
Žrior inferior frontal gyrus pars orbitalis; Brod-.mann area 47 , and may correspond to the lateral
posterior OFC region reported in monkey elec-Ž .trophysiological studies see Fig. 1b . It appears
Ž .too far lateral 38]46 mm lateral to the midlineto represent the central olfactory region. To date,this focus has only been clearly distinguished inthe left hemisphere. Imaging this area is compli-cated due to its location approximately 1 cmabove the ocular muscles. This problem is exacer-bated in studies with low spatial resolution, be-cause activity induced by muscle movements mayblur together with activity within the lateral olfac-tory region. However, the use of higher spatialresolutions, and inspection of co-registered struc-tural MRIs can typically alleviate these concerns.
Olfactory-related activity has also been re-ported in an extreme medial sector of the ventral
Žfrontal lobe Small et al., 1997; Royet et al.,.1999 . This activity falls within the gyrus rectus or
the medial orbital gyrus, 8]14 mm from the mid-line. This likely reflects a distinct area from thecentral olfactory region, because it lies in ananatomically and cytoarchitectually different re-
Žgion than the central OFC region Carmichael.and Price, 1994 . This area receives substantial
projections from the olfactory system in non-Ž .human primates Carmichael et al., 1994 , making
it a potentially distinct secondary olfactory pro-cessing subregion.
The frequency with which OFC responsesemerge in functional neuroimaging studies of ol-faction is striking when compared to the far lessconsistent observations of POC activation. Thisstands in contrast to other sensory systems inwhich secondary regions do not typically showincreased activity in the absence of an observablesignal increase within primary sensory cortex. Thedifference in OFC and POC responsivity toodorants may largely reflect a differential timecourse of activation. Whereas, the POC may showhabituating and transient responses to odorants,OFC regions may display a more prolonged pe-
Ž .riod of activation. Indeed, Sobel et al. in pressindicate that portions of the OFC show prolongedresponses to odorants that do not rapidly habitu-
ate. This would help explain the greater ease ofobserving OFC activation in studies applying blockdesigns. These findings highlight the importanceof incorporating temporal analysis in fMRI stud-ies of olfaction. Future studies may delineatethose areas whose activity follows a time coursecorresponding to the perception of odorants, asopposed to those linked to the time course ofperipheral stimulation. Future studies will hope-fully also determine if all OFC subregions pro-cessing olfactory stimuli display prolonged re-sponses, or if some OFC areas show a moretransient pattern of activity.
PET studies suggest that the different olfactoryresponsive subregions of the OFC may subservedifferent functions. Specifically, the right centralOFC appears to play a critical role in recognitionmemory. In two separate PET studies, the rightcentral OFC emerged when subjects were askedto judge the familiarity of odorants relative tocontrol conditions in which the subjects made
Ž .other types of judgments Royet et al., 1999 . Inanother study, right central OFC activity emergedwhen subjects were asked to recognize previouslypresented odorants, but not when they were asked
Ž .to encode odorants Dade et al., 1998 . Thesefindings converge with previous neuropsychologi-cal lesion studies which indicate a critical involve-
Žment of the right OFC in odor memory Jones-.Gotman and Zatorre, 1988, 1993 .
In contrast, the left lateral OFC region mayplay a more central role in hedonic judgments.This area emerges when subjects make judge-ments about the edibility of hedonic quality of
Žodorants Royet et al., 1999, unpublished.manuscript and during exposure to odorants with
Žstrong hedonic characteristics Zald and Pardo,.1997a . A study of functional connectivity during
olfactory tasks also suggests laterality differencesbetween the right and left central lateral OFC.Specifically, the left and right OFC appear func-tionally coupled at rest and when trying to detectan odor, but this functional coupling breaks downwhen subjects are exposed to an aversive odorant.This breakdown in functional connectivity sug-gests that the two regions perform different func-
Ž .tions under this condition Zald et al., 1998a .

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 173
Like the pyriform cortex, there is an area of theOFC with responses that correlate with sniffingŽ .Sobel et al., 1998a . The area showing sniff-induced activity for the most part lies in theextreme posterior or medial sections of the infe-rior frontal lobe and appears to be distinct fromthe central and lateral olfactory responsive areasdiscussed above.
3.2. Amygdala
Of all the senses, olfaction possesses the mostintimate relation with the amygdala. The POCabuts the superficial nuclei of the anterior amyg-dala, and some researchers actually include these
Žnuclei as part of the POC Carmichael et al.,.1994; Swanson and Petrovich, 1998 . This close
proximity can create problems in neuroimagingstudies, in some cases making it difficult to de-termine whether a focus lies within posterior sec-tions of the pyriform cortex or anterior sectionsof the amygdala. However, in cases where thefocus falls several millimeters anterior or poste-rior to this border, localization can be determinedwith greater certainty. The shape and anterior]posterior extent of the activation can also facili-tate this determination.
The amygdala receives a complex array of di-rect projections from the POC and the lateralolfactory tract as well as indirect projections fromthe periamygaloid cortex and endopyriform nu-
Žcleus Turner et al., 1978, 1980; Ottersen, 1982;Price, 1991; Amaral et al., 1992; Carmichael et
.al., 1994 . Fig. 2 displays a schematic of some ofthe projections from the POC and lateral olfac-tory tract to the amygdala.
In lower mammals, the amygdala appears es-sential for the recognition of a range of biologi-cally and emotionally significant odorants. To testwhether emotionally salient odorants activate thehuman amygdala, we exposed healthy female sub-jects to a highly unpleasant combination of sul-
Žfides while rCBF was assayed with PET Zald and.Pardo, 1997a . Relative to a no-odor control con-
dition, the aversive sulfides produced large-mag-nitude rCBF increases in the amygdalae bilater-ally. In the right hemisphere, the focus corre-sponded closely to the Talairach coordinates of
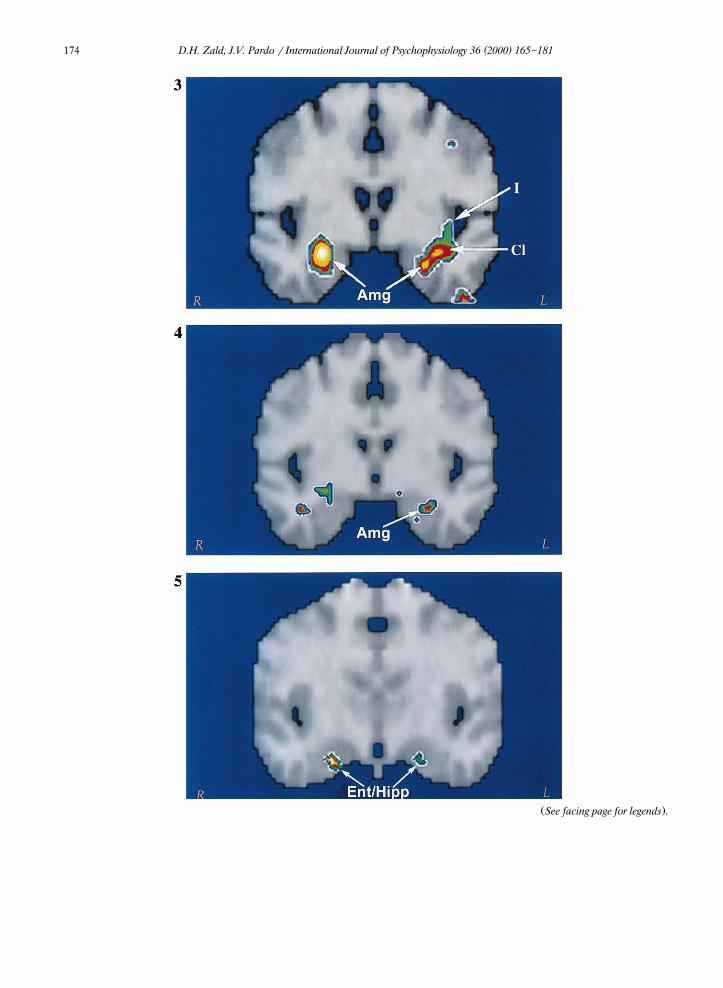
the amygdala. In the left hemisphere, the focusencompassed the amygdala, but also extendedlaterally into the neighboring insula-claustrum re-gion. Indeed, the peak focus mapped just lateralto the amygdala. This could indicate that morethan one focus falls in this region, one of whichfalls in the amygdala. Alternatively, there couldbe a single focus that maps lateral to the amyg-dala. In the following paragraph, we report are-analysis of these data which demonstrate thatmultiple foci map to the left anterior temporallobe, one of which maps directly to the amygdala.
As mentioned above, the spatial resolution ofPET often prevents the distinction of proximal
Ž .foci. In the Zald and Pardo 1997a study, thedata were filtered with a 9-mm three-dimensionalGaussian filter producing a FWHM of 12 mm. Toexamine whether more than one foci localized tothe left anterior temporal insular region, we re-analyzed the data using a smaller 6.75-mm filterwhich produced an FWHM of approximately 9.5mm. This new analysis reveals three discrete fociin this region. The largest of these foci clearly
Žfalls within the amygdala xsy26, ysy1, zsy18; Z-scores3.5; denoted with an ‘Amg’ in
. 4Fig. 3 . A second, equally strong focus appearsslightly lateral to the Talairach coordinates of the
Žamygdala xsy30, ysy1, zsy14; Z-scores.3.5 in the vicinity of the ventral temporal claus-
Ž .trum denoted with a ‘Cl’ in Fig. 2 . Finally, athird focus of lower magnitude localizes to the
Žinferior insula xsy39, ysy6, zsy7; Z-.scores2.9 .
The above re-analysis indicates that bilateralfoci emerge in the amygdalae during exposure toaversive odorants. Indeed, this finding has sincebeen replicated using fMRI with both normal
Žcontrols and social phobics Birbaumer et al.,.1998 . In contrast, similarly strong amygdala acti-
vations do not emerge during exposure to pleas-Žant odorants Zatorre et al., 1992; Small et al.,
.1997; Zald and Pardo, 1997a,b .
4 Ž .x represents mm from the midline qsright ; y repre-Ž .sents mm from the anterior commissure qsanterior ; and z
Žrepresents mm from the intercommissural plane qs.superior .
( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181174
Ž .See facing page for legends .

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 175
Ž .Region of interest ROI analyses of both PETŽ . ŽZald and Pardo, 1997a and fMRI Prabhakaran
.et al., 1999 data suggest that the magnitude orŽvolume of activation within the amygdalae or left
.amygdala correlates with the level of perceivedaversiveness. ROI analyses are limited by theoften arbitrary nature of the size and placementof the ROIs. For instance, a large ROI centeredon the anterior amygdala may blur correlated anduncorrelated activity together, hence obscuring asignificant association. In contrast, a small ROImay fail to capture a significant correlation, byfailing to encompass the correlated region. Analternative approach to detecting relationsbetween rCBF and psychoperceptual variablesutilizes a pixelwise correlational analysis. Such ananalysis examines the correlation between activitywithin each pixel in a brain region and psychoper-ceptual variables of interest. This produces a mapof the location and volume of brain regions whoseactivity varies in relation to a given psychopercep-tual variable. To examine the specific location ofcorrelational foci within the anterior temporallobe, we re-analyzed the aversive sulfide data
Ž .from Zald and Pardo 1997b using this correla-tional technique. We accomplished this by ex-amining the pixel-by-pixel correlation betweensubjects’ psychoperceptual ratings and rCBF
Žchange from the aversive sulfide } no-odor con-
5We utilized the re-analyzed data set, with the higherŽ .spatial resolution FWHMs9.5 mm , in order to maximize
visualization of the amygdala relative to proximal structures.
.trast for the 12 subjects who participated in theaversive sulfide experiment.5
Pixelwise correlational analysis of the aversiveodorant study confirmed that a discrete areawithin the left amygdala correlated significantly
Žwith ratings of aversiveness xsy28, ysy4,.zsy16; rs0.67, P-0.01 , such that larger
rCBF increases were associated with more ex-Ž .treme ratings of aversiveness see Fig. 4 . Ratings
of intensity also show correlations with left amyg-Ždalar rCBF during aversive olfaction Zald and
.Pardo, 1997a . This likely reflects the tight associ-ation between ratings of aversiveness and inten-
Žsity for unpleasant stimuli see Royet et al., 1999,.for example . However, intensity alone cannot
explain amygdalar activation. First, the relationwith aversiveness emerges even though there waslittle variance in intensity ratings across subjects.Second, intense concentrations of pleasant
Žodorants do not induce amygdalar activation Zald.and Pardo, 1997b . Third, amygdalar rCBF does
not correlate with the perceived intensity ofŽpleasant odorants see analysis described in Sec-
.tion 3.3 .The neuroimaging literature on the amygdala’s
response to aversive odorants converges with clin-ical evidence indicating that seizures or electricalstimulation of the human amygdala frequentlyproduces unpleasant olfactory hallucinations, butrarely produces pleasant olfactory hallucinationsŽ .Chitanodh, 1966; Andy, 1967; Andy et al., 1975 .It also agrees with neuroimaging studies whichshow a preferential activation of the amygdala
Žduring exposure to aversive tastes Zald et al.,
Fig. 3. Amygdalar and insular activation during olfactory stimulation with aversive sulfides. The figure displays a coronal sliceŽ . Ž .through the amygdala ysy4 . The rCBF data derive from a high resolution re-analysis of Zald and Pardo 1997b . Abbre¨iations:
Amgsamygdala; Clsclaustrum; and Is insula.Fig. 4. Pixelwise correlation between the change in rCBF and ratings of aversiveness during exposure to aversive sulfides. The figure
Ž .displays a coronal slice through the amygdala ysy4 . Areas in red represent a correlation with aversiveness of r)0.60. A peakcorrelational focus falls within the left amygdala in approximately the same location as the peak focus in the high resolutionre-analysis. The search region was limited to the medial temporal lobe in order to limit the number of comparisons. Correlationalfoci outside of this region are not displayed.Fig. 5. Pixelwise correlation between ratings of intensity and change in rCBF during exposure to pleasant olfactory stimuli. Theimage displays a coronal slice through ysy10. Areas in yellow represent a correlation with a magnitude of r)0.70. The largest
Ž .correlations localize to the hippocampus and anterior entorhinal cortex abbreviated EntrHipp . These foci occupy multiplecoronal slices, with the peak focus in the left hemisphere occurring in the entorhinal cortex several millimeters anterior to this slice.The search region was limited to the medial temporal lobe in order to limit the number of comparisons. Correlational foci outsideof this region are not displayed.

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181176
. Ž1998b and unpleasant visual stimuli Irwin et al.,.1996; Lane et al., 1997; Taylor et al., 1998 rela-
tive to pleasant stimuli from the same sensorymodality. It may be speculated that over evolu-tion the amygdala has taken on a more negativevalence-specific role in sensory processing, orevolved such that aversive stimuli activate a moresizable network of amygdaloid nuclei than simi-larly valenced pleasant stimuli.
3.3. Entorhinal cortex and hippocampus
The POC projects directly to the entorhinalcortex, which in turn provides the major routethrough which olfactory information reaches the
Žhippocampus Price, 1991; Carmichael et al.,.1994 . It has sometimes been speculated that this
projection might play a role in some of the uniqueŽaspects of olfactory memory Herz and Engen,
.1996 . Although neuroimaging studies of olfactionhave generally failed to observe strong hippocam-pal responses to odorants, a few lines of evidencesuggest that, in certain circumstances, exposure toodorants induce changes in hippocampal or en-
Ž .torhinal rCBF. Small et al. 1997 reported a lefthippocampal focus during olfactory exposure rel-ative to a no-odor baseline, although the focuswas somewhat superior to the standard Talairachcoordinates for the hippocampus. In our ownlaboratory, we frequently observe small rCBF in-creases in the left entorhinal cortex or neigh-
Ž .boring fusiform gyrus Z-score)2.5 during ol-factory conditions relative to no-odor control con-ditions, but these usually fail to reach rigorous
Žstatistical criteria Zald and Pardo, unpublished.observations .
At the level of individual subjects, wide varia-tions occur in the extent to which odorants acti-vate the entorhinalrhippocampal region. To seeif these differences relate to individual differ-ences in psychoperceptual features, we performeda pixelwise correlational analyses of rCBF withinthe medial temporal lobe on a group of 14 healthy
Žright-handed female subjects mean age s 27.years, ranges19]49 years exposed to pleasant
wodorants fruit, spice or floral odorants presentedeither using the University of Pennsylvania Smell
Ž .Identification Test UPSIT; Doty et al., 1984 or
xon a saturated cotton-tip wand . During a givenscan, subjects received four odorants, each pre-sented twice in sequence, with an 8-s stimulusduration and a 1-s interstimulus interval. Odorantswere presented birhinally by waving the stimulusback and forth approximately 2 inches in front ofthe subject’s nose. Imaging techniques were iden-
Ž .tical to Zald and Pardo 1997a , except that imageswere only blurred to an FWHM of 9.5 mm inorder to better delineate structures within themedial temporal lobe. In the odorant conditions,as well as during a no-odor control condition,subjects received instructions to breathe throughtheir nose and see if they could smell an odor.Five subjects were scanned with both types of
Ž .odorants i.e. UPSIT and cotton wand producinga total of 19 rCBF change images. A pixelwisecorrelation between ratings of pleasantness andrCBF change revealed no significant relationbetween changes in rCBF and ratings of pleasant-ness in the hippocampal]entorhinal region. How-ever, rCBF in the entorhinalrhippocampal re-gions showed a strong relation with ratings ofperceived intensity. Indeed, this region showed ahigher magnitude and a larger volume of corre-lated pixels than any other area of the brain. Inthe right hemisphere, the peak mapped to the
Žextreme anterior end of the hippocampus xs21,ysy10, zsy17; rs0.70, P-0.001; see Fig..5 . A similar correlational focus emerged in the
left hemisphere, but the peak of this focus lies inŽthe anterior entorhinal cortex xsy24, ysy3,
.zsy27; rs0.66, P-0.005 . In both of theseareas, odorants rated as mild were associated withrCBF decreases relative to the no-odor controlcondition, whereas odorants rated as intense wereassociated with large rCBF increases. While suchan analysis clearly requires replication due to thesmall sample size, it nevertheless, raises the possi-bility of a gating process through which intense
Ž .odorants induce or gain entry into hippocampalprocesses, whereas more mild stimuli do not.
3.4. Other brain regions
Neuroimaging studies also indicate that olfac-tory responses often occur outside of traditionallydefined olfactory areas. Such responses localize to

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 177
Žthe insularrclaustrum region Zatorre et al., 1992;Small et al., 1997; Fulbright et al., 1998; Francis
.et al., 1999 ; several frontal regions outside of theŽOFC Levy et al. 1997; Fulbright et al., 1998;
. ŽRoyet et al., 1999 ; the cingulate Levy et al.,.1997; Yousem et al., 1997; Fulbright et al., 1998 ;
Žand the cerebellum Small et al., 1997; Sobel et.al., 1998b . However, with the exception of por-
wtions of the insularrclaustrum portions of whichreceive direct projections from the olfactory sys-
Ž .xtem Carmichael et al., 1994 , the olfactory func-tions of these regions remain poorly defined.There exists little in the way of animal experi-mental or human clinical literature to implicatethese regions in olfaction. However, most neuro-logical and neuropsychological exams contain nomore than a cursory examination of gross olfac-tory functions. Thus, subtle olfactory processingdeficits may occur following lesions to these ar-eas, which go undetected due to the frequentfailure to rigorously assess olfactory functions.
4. Caveats and conclusions
The current review demonstrates the ability offunctional neuroimaging techniques to providenovel insights into the neural substrates of olfac-tory processing. For instance, these studies indi-cate that the pyriform cortex shows activity syn-chronized to sniffing, while its responses toodorants appear transient and subject to rapidhabituation. This research also indicates that thehuman OFC possesses more than one olfactoryresponsive region, and that these regions responddifferently depending upon the odorants pre-sented or the specific demands of the task. Neu-roimaging studies also provide evidence that theamygdala preferentially responds to highly aver-sive odorants relative to more neutral or pleasantodorants. Nevertheless, the study of olfaction withneuroimaging techniques remains in its infancy.Whereas ERP studies often deal with subtle is-sues related to olfaction, neuroimaging studieshave only just begun to delve into the complexi-ties of olfactory processing. For instance, only oneneuroimaging study to date has dealt with the
interaction between the olfactory and trigeminalŽ . 6nerves Yousem et al., 1997 .
Many methodological issues also require atten-tion. Indeed, even rudimentary questions such asthe effect of different control conditions remainlargely unaddressed. Most of the studies reviewedin this paper employ a subtraction methodologyin which activation in a control condition is di-rectly subtracted from a stimulation condition.Thus, activations refer to increases in rCBF orBOLD signal relative to the control condition. Inthe ideal world the control condition matches thestimulation condition in all regards except oneŽe.g. matching all factors except the presence of
.an odorant . This has been difficult to fully imple-ment in most neuroimaging studies of olfactionbecause humans actively exert control over olfac-tory exposure by altering their respiration andattention to olfaction. In many studies, subjectsare given different instructions in the control andstimulation conditions, which means that factorssuch as attention to olfaction are not matched foracross conditions. Although such factors may onthe surface seem minor, they may substantiallyinfluence the results of subtraction analyses. Pre-liminary analyses of data from our own laboratorysuggest that attending to olfaction alone does notsignificantly increase rCBF within primary andsecondary olfactory regions relative to resting orbreathing through the nose without trying to smellŽ .Zald and Pardo, unpublished observations .However, different control conditions may alteractivity within the OFC enough to change therelative magnitude of olfactory-induced foci withindifferent regions of the OFC. In a similar vein,
Ž .the data of Sobel et al. 1998a,b make it evidentthat differences in sniffing between conditionscan produce large changes in the activity of someolfactory regions. Use of olfactometric techniques
6 These authors indicate that odorants with strong trigemi-nal components produce a more widespread pattern of activa-tion than relatively pure odorants. Interestingly, the OFCappears less responsive during exposure to mixedolfactoryrtrigeminal stimuli than during exposure to pureodorants.

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181178
may alleviate some of these issues, especially withthe introduction of olfactometers that are com-
Žpatible with the demands of fMRI Sobel et al.,.1997; Lorig et al., 1999 . Another major issue
relates to gender. Some investigators have exclu-Ž .sively studied men Royet et al., 1999 , while
other researchers have exclusively studied womenŽ .Zald and Pardo, 1997a , and still other investiga-tors have examined a combination of men and
Žwomen e.g. Zatorre et al., 1992; Small et al.,.1997 . Given that males and females show numer-
Žous differences in olfactory perception Doty et.al., 1985; Velle, 1987 , such gender differences
require systematic investigation. Indeed, YousemŽ .et al. 1999 , have recently provided evidence that
females demonstrate more robust cortical activa-tions during exposure to odorants than men ex-posed to the same odorants.
Despite some of the methodological issuesraised above, neuroimaging techniques are likelyto provide increasingly detailed information aboutthe olfactory system of both normal controls andpatients with olfactory dysfunction. A particularlyripe area for investigation relates to the delin-eation of the different functions subserved by thevarious brain regions already identified by neu-roimaging studies. Humans can perform manyolfactory functions, including odor detection, dis-crimination, classification, recognition and encod-ing. Humans can also form unimodal and cross-modal associations with odorants and make anumber of judgments about odorants such asfamiliarity, intensity and pleasantness. However,only recently have neuroimaging researchers be-
Žgun to attend to such issues see Royet et al.,.1999; Dade et al., 1998, for example . It also
remains unknown to what extent human neuralresponses to olfactory stimuli can be modified bycognitive factors such as expectation or motivatio-nal factors such as hunger or satiety.
The use of functional neuroimaging as a diag-nostic tool also appears ripe for exploration. A
Ž .preliminary study by Levy et al. 1998a demon-strated significantly lower BOLD responses toodorants in anosmics relative normals, but therelationship between specific types of olfactorydysfunction and responses in olfactory structuresremains unexplored. The measurement of neural
responses to odorants may also prove useful as abiological index of treatment response in olfac-
Ž .tory disorders. For instance, Levy et al. 1998breported significant increases in fMRI responsesto odorants in hyposmics treated with theo-phylline. Functional neuroimaging assessmentsmay also provide useful diagnostic information indisorders such as Alzheimer’s dementia, in whicholfactory deficits develop early in the course ofthe disorder. There already exists evidence thatbehavioral measures of olfactory identification canprovide useful diagnostic information about
ŽAlzheimer’s dementia Doty, 1991; Morgan et al.,.1995 . The simultaneous measurement of brain
activity during olfactory tasks may further in-crease the diagnostic utility of such assessmentsby providing information about the specific neuralcorrelates of such deficits. Neuroimaging of re-sponses to odorants may also provide useful theo-retical and diagnostic information about psychi-atric and neurologic disorders that involve limbicregions that process olfactory stimuli. For in-stance, such probes may facilitate our under-standing of anxiety disorders that are frequently
Žspeculated to involve amygdalar dysfunction see.Birbaumer et al., 1998, for example .
As the availability of neuroimaging techniquesincreases, we will likely witness a dramatic in-crease in neuroimaging research on olfaction.While the olfactory system presents a number ofunique methodological problems, most of the toolsfor conducting methodologically sound studies ofolfaction already exist. Given the demonstratedability of both PET and fMRI to address many ofthe issues raised in this review, these imagingtechniques appear poised to provide a monumen-tal increase in our understanding of the neuralsubstrates of human olfaction.
Acknowledgements
This research was supported by the Depart-ment of Veterans Affairs; the Minnesota MedicalFoundation; NARSAD; the Minnesota Obesity
ŽCenter; the University of Minnesota Grants-in-.Aid of Research, Artistry, and Scholarship ; and a
ŽNational Research Service Award to Dr Zald 1

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 179
.F32 MH11641-01A1 . We thank Joel Lee for as-sistance with the pixelwise correlational analyses,Darren Dutcher for assistance with Fig. 2 andDorothy Mattson for helpful comments on anearlier draft of this manuscript.
References
Andy, O.J., 1967. The amygdala and hippocampus in olfactoryaura. Electroencephalogr. Clin. Neurophysiol. 23, 292.
Andy, O.J., Jurko, M.F., Hughes, J.R., 1975. The amygdala inrelation to olfaction. Confin. Neurol. 37, 215]222.
Amaral, D.G., Price, J.L., Pitkanen, A., Carmichael, S.T., 1992.Anatomical organization of the primate amygdaloid com-
Ž .plex. In: Aggleton, J.P. Ed. , Neurobiological Aspects ofEmotion, Memory, and Mental Dysfunction. Wiley-Liss,New York, pp. 1]66.
Beck, E., 1949. A cytoarchitectural investigation into theboundaries of cortical areas 13 and 14 in the human brain.J. Anat. 83, 147]157.
Birbaumer, N., Grodd, W., Diedrich, O. et al., 1998. fMRIreveals amygdala activation to human faces in socialphobics. Neuroreport 9, 223]1226.
Brodmann, K., 1914. Physiologie des gehrins. Neue DeutscheChirugie 2, 85]426.
Carmichael, S.T., Price, J.L., 1994. Architectonic subdivisionof the orbital and medial prefrontal cortex in the macaquemonkey. J. Comp. Neurol. 346, 366]402.
Carmichael, S.T., Price, J.L., 1996. Connectional networkswithin the orbital and medial prefrontal cortex of macaquemonkeys. J. Comp. Neurol. 346, 179]207.
Carmichael, S.T., Clugnet, M.C., Price, J.L., 1994. Centralolfactory connections in the macaque monkey. J. Comp.Neurol. 346, 403]434.
Chitanodh, H., 1966. Stereotaxic amygdolectomy in the treat-ment of olfactory seizures and psychiatric disorders witholfactory hallucinations. Confin. Neurol. 27, 181]196.
Cometto-Muniz, J.E., Cain, W.S., 1995. Olfactory adaptation.˜Ž .In: Doty, R.L. Ed. , Handbook of Olfaction and Gustation.
Marcel Dekker, New York, pp. 257]282.Critchley, H.D., Rolls, E.T., 1996. Olfactory neuronal respon-
ses in the primate orbitofrontal cortex: analysis in anolfactory discrimination task. J. Neurophysiol. 75,1659]1672.
Dade, L.A., Jones-Gotman, M., Zattore, R.J., Evans, A.C.,1998. Human brain function during odor encoding andrecognition: a PET activation study. Ann. N.Y. Acad. Sci.855, 572]574.
Doty, R.L., 1991. Olfactory dysfunction in neurodegenerativeŽ .disorders. In: Gretchell, T.V. Ed. , Smell and Taste in
Health and Disease. Raven, New York, pp. 735]751.Doty, R.L., Shaman, P., Dann, M., 1984. Development of the
University of Pennsylvania Smell Identification Test: astandardized microencapsulated test of olfactory function.Physiol. Behav. 32, 489]502.
Doty, R., Applebaum, S., Zusho, H., Settle, R.G., 1985. Sex
differences in odor identification ability: a cross-culturalanalysis. Neuropsychologia 23, 667]682.
Farzaneh, F., Riederer, S.J., Pelc, N.J., 1990. Analysis of T2limitations and off-resonance effects on spatial resolutionand artifacts in echo-planar imaging. Magn. Reson. Med.14, 123]139.
Fox, P.T., Mintun, M.A., Raichle, M.E., Miezin, F.M., Allman,J.M., Van Essen, D.C., 1986. Mapping human visual cortexwith positron emission tomography. Nature 323, 806]809.
Frahm, J., Merbolt, K.-D., Hanicke, W., 1988. Direct FLASH¨MR imaging of magnetic field inhomogeneities by gradientcompensation. Magn. Reson. Med. 6, 474]480.
Francis, S., Rolls, E.T., Bowtell, R. et al., 1999. The represen-tation of pleasant touch in relationship with taste andolfactory areas. Neuroreport 10, 453]459.
Fulbright, R.K., Skudlarski, P., Lacadie, C.M. et al., 1998.Functional MR imaging of regional brain responses topleasant and unpleasant odors. Am. J. Neuroradiol. 19,1721]1726.
Glover, G.H., Lee, A.T., 1995. Motion artifacts in fMRI:comparison of 2DFT with PR and spiral scan methods.Magn. Reson. Med. 33, 624]635.
Haberly, L.B., Price, J.L., 1978. Association and commissuralfiber systems of the olfactory cortex of the rat. II. J. Comp.Neurol. 181, 781]818.
Herz, R.S., Engen, T., 1996. Odor memory: review and analy-sis. Psychon. Bull. Rev. 3, 300]313.
Irwin, W., Davidson, R.J., Lowe, M.J., Mock, B.J., Sorenson,J.A., Turski, P.A., 1996. Human amygdala activation de-tected with echo-planar functional magnetic resonanceimaging. Neuroreport 7, 1765]1769.
Jones-Gotman, M., Zatorre, R.J., 1988. Olfactory identifica-tion deficits in patients with focal cerebral excision. Neu-ropsychologia 26, 387]400.
Jones-Gotman, M., Zatorre, R.J., 1993. Odor recognition me-mory in humans: role of right temporal and orbitofrontalregions. Brain Cogn. 22, 182]198.
Koster, E.P., de Wik, R.A., 1991. Olfactory adaption. In:¨Ž .Laing, D.G., Doty, R.L., Breiphol, W. Eds. , The Human
Sense of Smell. Springer-Verlag, Berlin, pp. 199]215.Lane, R.D., Reiman, E., Bradley, M.M., Lang, P.J., Ahern,
G.L., Davidson, R.J., 1997. Neuroanatomical correlates ofpleasant and unpleasant emotion. Neuropsychologia 35,1437]1444.
Levy, L.M., Henkin, R.I., Hutter, A., Lin, C.S., Martins, D.,Schellinger, D., 1997. Functional MRI of human olfaction.Comput. Assist. Tomogr. 21, 849]856.
Levy, L.M., Henkin, R.I., Hutter, A., Lin, C.S., Schellinger, D.,1998a. Mapping brain activation to odorants in patientswith smell loss by functional MRI. J. Comput. Assist.Tomogr. 22, 96]103.
Levy, L.M., Henkin, R.I., Lin, C.S., Hutter, A., Schellinger, D.,1998b. Increased brain activation in response to odors inpatients with hyposmia after theophylline treatment de-monstrated by fMRI. J. Comput. Assist. Tomogr. 22,760]770.
Lorig, T.G., Elmes, D.L., Zald, D.H., Pardo, J.V., 1999. A

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181180
computer-controlled olfactometer for fMRI and electro-physiological studies of olfaction. Behav. Res. Methods:Instrum. Comput. 31, 370]375.
Luskin, M.B., Price, J.L., 1983. The topographic organizationof associational fibers of the olfactory system in the rat,including centrifugal fibers to the olfactory bulb. J. Comp.Neurol. 216, 264]291.
Morgan, C.D., Nordin, S., Murphy, C., 1995. Odor identifica-tion as an early marker for Alzheimer’s disease: impact oflexical functioning and detection sensitivity. J. Clin. Exp.Neuropsychol. 17, 793]803.
Morrell, G., Spielman, D., 1997. Dynamic shimming formulti-slice magnetic resonance imaging. Magn. Reson. Med.38, 477]483.
Noll, D.C., Cohen, J.D., Meyer, C.H., Schneider, W., 1995.Spiral K-space MR imaging of cortical activation. J. Magn.Reson. Imaging 5, 49]56.
Ogawa, S., Lee, T.M., Kay, A.R., Tank, D.W., 1990. Brainmagnetic resonance imaging with contrast dependent onblood oxygenation. Proc. Natl. Acad. Sci. U.S.A. 87,9868]9872.
Ogawa, S., Tank, D.W., Menon, R. et al., 1992. Intrinsic signalchanges accompanying sensory stimulation: functional brainmapping with magnetic resonance imaging. Proc. Natl.Acad. Sci. U.S.A. 89, 5951]5955.
Ojemann, J.G., Akbudak, E., Snyder, A.Z., McKinstry, R.C.,Raichle, M.E., Conturo, T.E., 1997. Anatomic localizationand quantitative analysis of gradient refocussed echo-planarfMRI susceptibility artifacts. Neuroimage 6, 156]167.
Ottersen, O.P., 1982. Connections of the amygdala of the rat.IV. Corticoamygdaloid and intraamyglaloid connections asstudied with axonal transport of horseradish peroxidase. J.Comp. Neurol. 205, 30]48.
Petrides, M., Pandya, D.N., 1994. Comparative architectonicanalysis of the human and the macaque frontal cortex. In:
Ž .Boller, F., Grafman, J. Eds. , Handbook of Neuropsy-Ž .chology Vol. 9 . Elsevier, Amsterdam, pp. 17]58.
Prabhakaran, V., Sobel, N., Desmond, J.E., Glover, G.H.,Sullivan, E.V., Gabrieli, J.D.E., 1999. Hedonic value ofodors is reflected in the level of activation in the amygdala.
Ž .Neuroimage, 5:S198 abstract .Ž .Price, J.L., 1991. The olfactory system. In: Paxinos, G. Ed. ,
The Human Nervous System. Academic Press, New York,pp. 979]998.
Ray, J.P., Price, J.L., 1993. The organization of projectionsfrom the mediodorsal nucleus of the thalamus to orbitaland medial prefrontal cortex in macaque monkeys. J. Comp.Neurol. 337, 1]31.
Royet, J.P., Koenig, O., Gregoire, M.C. et al., 1999. Functionalanatomy of perceptual and semantic processing for odors.J. Cogn. Neurosci. 11, 94]109.
Royet, J.P., Hudry, J., Zald, D.H., et al. Functional neuroana-tomy of different olfactory judgements. Unpublishedmanuscript.
Ruschen, F.T., Amaral, D.G., Price, J.L., 1987. The afferentinput to the magnocellular division of the mediodorsal
thalamic nucleus in the monkey, Macaca fascicularis. J.Comp. Neurol. 256, 175]210.
Small, D.M., Jones-Gotman, M., Zatorre, R.J., Petrides, M.,Evans, A.C., 1997. Flavor processing: more than the sum ofits parts. Neuroreport 8, 3913]3917.
Sobel, N., Prabhakaran, V., Desmond, J.E., Glover, G.H.,Sullivan, E.V., Gabrieli, J.D.E., 1997. A method for functio-nal magnetic resonance imaging of olfaction. J. Neurosci.Methods 78, 115]123.
Sobel, N., Prabhakaran, V., Desmond, J.E. et al., 1998a. Snif-fing and smelling } separate subsystems in the humanolfactory cortex. Nature 392, 282]286.
Sobel, N., Prabhakaran, V., Hartley, C.A. et al., 1998b. Odorantinduced and sniff-induced activation in the cerebellum inthe human. J. Neurosci. 18, 8990]9001.
Ž .Sobel, N., Prabhakaran, V., Zhao, Z. et al. in press . Timecourse of odorant-induced activation in the human primaryolfactory cortex. J. Neurophysiol.
Swanson, L.W., Petrovich, G.D., 1998. What is the amygdala.TINS 21, 323]331.
Takagi, S.F., 1986. Studies on the olfactory nervous system ofthe Old World monkey. Prog. Neurobiol. 27, 195]250.
Talairach, J., Tournoux, P., 1988. Co-planar Stereotaxic Atlasof the Human Brain. Thieme, New York.
Tanabe, T., Iino, M., Takagi, S.F., 1975a. Discrimination ofodors in olfactory bulb, pyriform-amygdaloid areas, andorbitofrontal cortex of the monkey. J. Neurophysiol. 38,1284]1296.
Tanabe, T., Yarita, H., Iino, M., Ooshima, Y., Takagi, S.F.,1975b. An olfactory projection area in orbitofrontal cortexof the monkey. J. Neurophysiol. 38, 1269]1283.
Taylor, S.F., Liberzon, I., Fig, L.M., Decker, L., Minoshima, S.,Koeppe, R.A., 1998. The effect of emotional content onvisual recognition memory: A PET activation study. Neu-roimage 8, 188]197.
Turner, B.H., Gupta, K.C., Mishkin, M., 1978. The locus andcytoarchitecture of the projection area of the olfactory bulbin Macaca mulatta. J. Comp. Neurol. 177, 381]396.
Turner, B.H., Mishkin, M., Knapp, M., 1980. Organization ofthe amygdalopetal projections from modality-specific corti-cal association areas in the monkey. J. Comp. Neurol. 191,515]543.
Velle, W., 1987. Sex differences in sensory functions. Perspect.Biol. Med. 30, 490]522.
Walker, A.E., 1940. A cytoarchitectural study of the prefrontalarea of the macaque monkey. J. Comp. Neurol. 73, 59]86.
Wilson, D.A., 1998. Synaptic correlates of odor habituation inthe rat anterior pyriform cortex. J. Neurophysiol. 80,998]1001.
Yarita, H., Iino, M., Tanabe, T., Kogure, S., Takagi, S.F., 1980.A transthalamic olfactory pathway to orbitofrontal cortexin the monkey. J. Neurophysiol. 45, 69]85.
Young, I.R., Cox, I.J., Bryant, D.J., Bydder, G.M., 1988. Thebenefits of increasing spatial resolution as a means ofreducing artifacts due to field inhomogeneities. Magn.Reson. Imaging 6, 585]590.

( )D.H. Zald, J.V. Pardo r International Journal of Psychophysiology 36 2000 165]181 181
Yousem, D.M., Williams, S.C.R., Howard, R.O. et al., 1997.Functional MR imaging during odor stimulation -pre-liminary data. Radiology 204, 833]838.
Yousem, D.M., Maldjian, J.A., Siddiqi, F. et al., 1999. Gendereffects on odor-stimulated functional magnetic resonanceimaging. Brain Res. 818, 480]487.
Yudilevich, E., Stark, H., 1987. Spiral sampling in magneticresonance imaging } the effect of inhomogeneities. IEEETrans. Med. Imaging MI-6, 337]345.
Zald, D.H., Kim, S.W., 1996a. Anatomy and function of theorbital frontal cortex, I: anatomy, neurocircuitry, andobsessive]compulsive disorder. J. Neuropsychiatry Clin.Neurosci. 8, 125]138.
Zald, D.H., Kim, S.W., 1996b. Anatomy and function of theorbital frontal cortex, II: function and relevance to obses-sive]compulsive disorder. J. Neuropsychiatry Clin. Neu-rosci. 8, 249]261.
Zald, D.H., Pardo, J.V., 1997a. The amygdala, olfaction andemotion: amygdala activation during aversive olfaction.Proc. Natl. Acad. Sci. U.S.A. 94, 4119]4124.
Zald, D.H., Pardo, J.V., 1997. Amygdala activation during
aversive olfaction. J. Neuropsychiatry Clin. Neurosci., 9,Ž .134 abstract .
Zald, D.H., Donndelinger, M.J., Pardo, J.V., 1998a. Eluci-dataing dynamic brain interactions with across-subjects cor-relational analyses of PET data: the functional connectivityof the amygdala and orbitofrontal cortex during olfactorytasks. J. Cereb. Blood Flow Metab. 18, 896]905.
Zald, D.H., Lee, J.T., Fluegel, K.W., Pardo, J.V., 1998b. Aver-sive gustatory stimulation activates limbic circuits in hu-mans. Brain 121, 1143]1154.
Zatorre, R.J., Jones-Gotman, M., 1991. Human olfactory dis-crimination after unilateral frontal or temporal lobectomy.Brain 114, 71]84.
Ž .Zatorre, R.J., Jones-Gotman, M. in press . Functional imag-ing of the chemical senses. In: Toga, A.W., Mazziotta, J.C.Ž .Eds. , Brain Mapping: The Applications. Academic Press,San Diego.
Zatorre, R.J., Gotman, M.J., Evans, A.C., Meyer, E., 1992.Functional localization and lateralization of human olfac-tory cortex. Nature 360, 339]340.




















