
Involvement of right piriform cortex in olfactory familiarity judgments
10
Involvement of right piriform cortex in olfactory familiarity judgments Jane Plailly, a, * Moustafa Bensafi, a Mathilde Pachot-Clouard, b Chantal Delon-Martin, b David A. Kareken, c Catherine Rouby, a Christoph Segebarth, b and Jean-P. Royet a,d a Neurosciences et Syste `mes Sensoriels, Universite ´ Claude Bernard Lyon 1, UMR CNRS 5020, IFR 19, Institut Fe ´de ´ratif des Neurosciences de Lyon, 50 Avenue Tony Garnier, 69366 Lyon Cedex 07, France b Unite ´ mixte INSERM/Universite ´ Joseph Fourier U594, LRC-CEA, Ho ˆpital Michallon, 38043 Grenoble, France c Neuropsychology Section, Department of Neurology, Indiana University School of Medicine, Indianapolis, IN 46202, USA d CERMEP, 69003 Lyon, France Received 16 April 2004; revised 12 October 2004; accepted 26 October 2004 Available online 9 December 2004 Previous studies have shown activation of right orbitofrontal cortex during judgments of odor familiarity. In the present study, we sought to extend our knowledge about the neural circuits involved in such a task by exploring the involvement of the right prefrontal areas and limbic/ primary olfactory structures. Fourteen right-handed male subjects were tested using fMRI with a single functional run of two olfactory conditions (odor detection and familiarity judgments). Each condition included three epochs. During the familiarity condition, subjects rated whether odors were familiar or unfamiliar. During the detection condition, participants decided if odors were present. When contrasting the familiarity with the detection conditions, activated areas were found mainly in the right piriform cortex (PC) and hippocampus, the left inferior frontal gyrus and amygdala, and bilaterally in the mid-fusiform gyrus. Further analyses demonstrated that the right PC was more strongly activated than the left PC. This result supports the notion that the right PC is preferentially involved in judgments of odor familiarity. D 2004 Elsevier Inc. All rights reserved. Keywords: Olfaction; Familiarity judgment; Recognition memory; Piriform cortex; fMRI Introduction Hemispheric asymmetry is well-established for high-level brain functions such as language and spatial attention (e.g., Broca, 1863; Weintraub and Mesulam, 1987). Hemispheric predominance also exists in sensory functions such as hand somatosensory represen- tation (Soros et al., 1999) and temporal and spectral auditory resolution (Zatorre et al., 2002). Studies in olfaction lead to similar conclusions. Early cerebral imaging studies showed functional lateralization of olfactory processes in the right hemisphere, especially in orbitofrontal cortex (OFC) (Zatorre et al., 1992); most subsequent studies confirm this result (Dade et al., 1998; Sobel et al., 1998; Yousem et al., 1997). However, Zald and colleagues reported stronger activation in the left OFC and amygdala for very aversive odors, pointing to these areas as being important in the emotional processing of olfactory information (Zald and Pardo, 1997; Zald et al., 1998). Using positron emission tomography (PET), Royet et al. (1999, 2001) found that judgments of odor familiarity preferentially activated right OFC, whereas hedonic judgments principally activated left OFC. Beyond OFC, recent cerebral imaging data have extended these observations to piriform cortex (PC). In a functional magnetic resonance imaging (fMRI) study, we showed that left piriform– amygdala region activation was associated with subjects’ ratings of the odorants’ emotional intensities (Royet et al., 2003), a result consistent with Gottfried et al.’s (2002) and Anderson et al.’s (2003) findings. Convergent results from Dade et al.’s (2002) lesion and PET studies further show that the piriform region mediates olfactory long-term recognition memory, giving support to the notion that this area may be more than primary sensory cortex (e.g., Schoenbaum and Eichenbaum, 1995). Specifically, Dade et al. (2002) found that the extent of piriform activity corresponded with different cognitive demands, in which PC activity followed a continuum between mnemonic encoding (no significant activity), to short-term recognition (weak bilateral activity), to long-term recognition (strong bilateral activity). These authors further suggested that piriform activity could be related to odor familiarity, which would require that subjects compare odors with previously stored olfactory representations, and thus represent a type of long-term olfactory reference memory. In the present study, we explored that question by specifically asking whether PC is involved in the processing of odor familiarity, and if so, whether familiarity-evoked activations are lateralized, as previously suggested (Royet et al., 1999, 2001). We studied 1053-8119/$ - see front matter D 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.neuroimage.2004.10.028 * Corresponding author. Fax: +33 4 37 28 76 01. E-mail address: [email protected] (J. Plailly). Available online on ScienceDirect (www.sciencedirect.com). www.elsevier.com/locate/ynimg NeuroImage 24 (2005) 1032 – 1041
-
Upload
ujf-grenoble -
Category
Documents
-
view
0 -
download
0
Transcript of Involvement of right piriform cortex in olfactory familiarity judgments

www.elsevier.com/locate/ynimg
NeuroImage 24 (2005) 1032–1041
Involvement of right piriform cortex in olfactory
familiarity judgments
Jane Plailly,a,* Moustafa Bensafi,a Mathilde Pachot-Clouard,b Chantal Delon-Martin,b
David A. Kareken,c Catherine Rouby,a Christoph Segebarth,b and Jean-P. Royeta,d
aNeurosciences et Systemes Sensoriels, Universite Claude Bernard Lyon 1, UMR CNRS 5020, IFR 19, Institut Federatif des Neurosciences de Lyon,
50 Avenue Tony Garnier, 69366 Lyon Cedex 07, FrancebUnite mixte INSERM/Universite Joseph Fourier U594, LRC-CEA, Hopital Michallon, 38043 Grenoble, FrancecNeuropsychology Section, Department of Neurology, Indiana University School of Medicine, Indianapolis, IN 46202, USAdCERMEP, 69003 Lyon, France
Received 16 April 2004; revised 12 October 2004; accepted 26 October 2004
Available online 9 December 2004
Previous studies have shown activation of right orbitofrontal cortex
during judgments of odor familiarity. In the present study, we sought to
extend our knowledge about the neural circuits involved in such a task
by exploring the involvement of the right prefrontal areas and limbic/
primary olfactory structures. Fourteen right-handed male subjects
were tested using fMRI with a single functional run of two olfactory
conditions (odor detection and familiarity judgments). Each condition
included three epochs. During the familiarity condition, subjects rated
whether odors were familiar or unfamiliar. During the detection
condition, participants decided if odors were present. When contrasting
the familiarity with the detection conditions, activated areas were found
mainly in the right piriform cortex (PC) and hippocampus, the left
inferior frontal gyrus and amygdala, and bilaterally in the mid-fusiform
gyrus. Further analyses demonstrated that the right PC was more
strongly activated than the left PC. This result supports the notion that
the right PC is preferentially involved in judgments of odor familiarity.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Olfaction; Familiarity judgment; Recognition memory; Piriform
cortex; fMRI
Introduction
Hemispheric asymmetry is well-established for high-level brain
functions such as language and spatial attention (e.g., Broca, 1863;
Weintraub and Mesulam, 1987). Hemispheric predominance also
exists in sensory functions such as hand somatosensory represen-
tation (Soros et al., 1999) and temporal and spectral auditory
resolution (Zatorre et al., 2002). Studies in olfaction lead to similar
conclusions. Early cerebral imaging studies showed functional
1053-8119/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.neuroimage.2004.10.028
* Corresponding author. Fax: +33 4 37 28 76 01.
E-mail address: [email protected] (J. Plailly).
Available online on ScienceDirect (www.sciencedirect.com).
lateralization of olfactory processes in the right hemisphere,
especially in orbitofrontal cortex (OFC) (Zatorre et al., 1992);
most subsequent studies confirm this result (Dade et al., 1998;
Sobel et al., 1998; Yousem et al., 1997). However, Zald and
colleagues reported stronger activation in the left OFC and
amygdala for very aversive odors, pointing to these areas as being
important in the emotional processing of olfactory information
(Zald and Pardo, 1997; Zald et al., 1998). Using positron emission
tomography (PET), Royet et al. (1999, 2001) found that judgments
of odor familiarity preferentially activated right OFC, whereas
hedonic judgments principally activated left OFC.
Beyond OFC, recent cerebral imaging data have extended these
observations to piriform cortex (PC). In a functional magnetic
resonance imaging (fMRI) study, we showed that left piriform–
amygdala region activation was associated with subjects’ ratings of
the odorants’ emotional intensities (Royet et al., 2003), a result
consistent with Gottfried et al.’s (2002) and Anderson et al.’s
(2003) findings. Convergent results from Dade et al.’s (2002)
lesion and PET studies further show that the piriform region
mediates olfactory long-term recognition memory, giving support
to the notion that this area may be more than primary sensory
cortex (e.g., Schoenbaum and Eichenbaum, 1995). Specifically,
Dade et al. (2002) found that the extent of piriform activity
corresponded with different cognitive demands, in which PC
activity followed a continuum between mnemonic encoding (no
significant activity), to short-term recognition (weak bilateral
activity), to long-term recognition (strong bilateral activity). These
authors further suggested that piriform activity could be related to
odor familiarity, which would require that subjects compare odors
with previously stored olfactory representations, and thus represent
a type of long-term olfactory reference memory.
In the present study, we explored that question by specifically
asking whether PC is involved in the processing of odor familiarity,
and if so, whether familiarity-evoked activations are lateralized, as
previously suggested (Royet et al., 1999, 2001). We studied

J. Plailly et al. / NeuroImage 24 (2005) 1032–1041 1033
familiarity judgments and their relation to PC activation with fMRI
using a classical block paradigm design. Familiar and unfamiliar
odors were presented in a same epoch allowing subjects to rate
familiarity in a binary fashion by pressing one of two buttons. A
control condition was employed in which subjects had to judge the
presence or absence of an odor. The contrast between both
conditions allowed us to identify the areas specifically involved
in the familiarity judgment of odors.
Materials and methods
Subjects
Fourteen healthy right-handed men (18–32 years old) partici-
pated in the study. Participation required a medical screening.
Exclusion criteria were rhinal disorders (colds, active allergies,
history of nasal-sinus surgery, or asthma), neurologic disease,
ferrous implants (e.g., pacemakers, cochlear implants), or claus-
trophobia. Participants scored at least 87% correct in a forced-
choice suprathreshold detection test, and had a breathing cycle
mean duration of 3.88 s (F0.72). All participants provided written
informed consent as approved by the local Institutional Review
Board, and according to French regulations on biomedical experi-
ments on healthy volunteers.
Odorous stimuli
One hundred and eight stimuli were used, 27 of which were
employed before imaging sessions and 81 of which were used
Table 1
List of odorants selected for the Da, Db, Dc, Fa, Fb, and Fc epochs
Da Db Dc
1 Plum Sage Acacia
2 Turpentine
3
4 Tarragon Orange
5 Parsley Acetophenone
6 Guaiacol
7 Jonquil Camphor
8 Neroli
9 Musk
10 Carrot Pine needle
11 EBA
12 1,4-Dichlorobutane Butanol
13 Eglantine
14
15 Cherry 2-Bromopheno
16 Acetol Liqueur wine Tangerine
17 2-Octanol
18 Liquorice Blackcurrant
Unfamiliar odors
Mean (SD) 4.00 (0.72) 4.11 (0.87) 4.29 (0.92)
Range 3.14–5.11 3.19–5.16 3.09–5.17
Familiar odors
Mean (SD)
Range
Note. EBA, ethyl benzoyl acetate; PPA, phenyl propionaldehyde; SD, standard d
scale, with 1 representing low familiarity, and 10 representing high familiarity (s
during scanning. For fMRI, 54 odorants were used for the
familiarity (F) condition and 27 odorants for the detection (D)
condition (Table 1). For F conditions, three sets (Fa, Fb, Fc)
contained nine familiar and nine unfamiliar odorants selected so as
to provide high- and low-familiarity scores from data as derived
from previous work (Royet et al., 1999). Analysis of variance
(ANOVA) indicated that familiarity scores were significantly
higher for familiar than for unfamiliar odorants [F(1,48) =
129.173, P b 0.0001]. For D conditions, three sets (Da, Db, Dc)
contained nine odorants with low familiarity and nine bottles with
odorless air. Odors with low familiarity were selected to avoid
implicit familiarity judgments. For training, three sets of nine low
familiarity odorants and nine bottles with odorless air were used. In
each set, the presentation order of stimuli was pseudorandomized,
but identical for all subjects. Odorants were diluted to a concen-
tration of 10% using mineral oil (Sigma Aldrich, France). For
presentation, 5 ml of this solution was absorbed into compressed
polypropylene filaments inside of a 100 ml white polyethylene
squeeze-bottles equipped with a dropper (Osi, France).
Stimulating and recording materials
Odors were presented using an airflow olfactometer, which
allowed synchronizing stimulation with breathing. The stimulation
equipment was essentially the one used in a previous PET study
(Royet et al., 1999), but adapted so as to avoid interference with
the static magnetic field of the scanner (Royet et al., 2003). Briefly,
the apparatus was split into two modules: the electronic part of the
olfactometer positioned outside the magnet room (shielded with a
Faraday cage), and the nonferrous (DuraluminR) air-dilution
Fa Fb Fc
Apricot Raspberry Citronella
Bergamot Orange Tetralin trans-2-Hexenal
Diethyl ether Mint Cypress
Pine Honey Geranium
Bornyl acetate Patchouli Vienna bread
Caramel Jasmine Anise
Grass Basil Iris
Strawberry Toluene Incense
Pepper Green lily Lavender
Oyster Banana Gingerbread
Gardenia Vanilla Apple
Celery Lily Garlic
Hazelnut Bitter almond PPA
Tar Caprylic aldehyde Rose
l Methyl acetate Passion fruit Thyme
Biscuit Eucalyptus Lime
Tobacco Coconut 1-Octen-3-ol
3-Methyl anisol Clove Camomile
4.56 (0.28) 4.38 (0.39) 4.37 (0.23)
4.03–4.91 3.41–4.69 3.89–4.61
6.53 (0.75) 6.28 (0.67) 6.24 (0.23)
5.13–7.27 4.89–7.24 4.96–6.91
eviation. Italic, familiar odorants. Odorants were rated on a 10-point rating
ee Royet et al., 1999).

J. Plailly et al. / NeuroImage 24 (2005) 1032–10411034
injection head placed within the stray-field of the magnet.
Compressed air (10 l/min) was pumped into the olfactometer,
and delivered continuously through a standard anesthesia mask. A
detailed description was recently given (Vigouroux et al., in press).
The ventral breathing rhythm of the subject was recordedwith the
aid of a foot bellows in polyvinyl chloride (Herga Electric Limited,
Suffolk, UK) held on the stomach with a weaved cotton belt.
Movements of the abdominal wall produced variations in the internal
volume of the foot bellows. The flow was transformed into an
electrical signal that was amplified, and then successively trans-
mitted to the acquisition system and to a headphone via a voltage-to-
frequency converter. The experimenter could therefore listen to the
progressive frequency variations that accompanied respiration (high
frequency during inspiration and low frequency during expiration).
This method made respiration easily detected for the experimenter
and allowed easy timing of the stimulus delivery. During scanning,
the subject was instructed to avoid sniffing or blocking his breathing,
but instead to breathe regularly, thus allowing the experimenter to
anticipate the beginning of an inspiration phase. One stimulus was
then injected into the olfactometer by squeezing one bottle into the
injection head, so that the odor (or odorless air) was carried to the
subject’s anesthesia mask.
Subjects rated familiarity or judged odor presence by pressing
one of two buttons. The response signal was then transmitted
outside the radiofrequency shielded room by fiber optics to analog-
to-digital converters powered by nickel–cadmium batteries.
Behavioral data were recorded on line (100 Hz sampling rate)
using a NEC PC computer equipped with a digital acquisition
board DAQCard-500 (National Instruments, USA). LabView 5.0
software (National Instruments) was used to acquire, store, and
read data. Data analysis was performed with the WinDaq Wave-
form Browser 1.91 software (DataQ Instruments, USA).
Experimental procedure
A single functional run was presented in blocks that consisted
of two olfactory conditions (F and D) alternating with odorless rest
(R) epochs (Fig. 1). Each epoch lasted 60 s. Both F and D
conditions were presented three times each, either three F followed
by three D conditions or vice versa. Within the same condition, the
presentation order of the three sets (a, b, c) was counterbalanced
across subjects according to a balanced experimental (Latin square)
design. For olfactory conditions, subjects were asked to judge
whether or not they smelled an odor (D condition) or whether the
odor was familiar or unfamiliar (F condition). Subjects were then
asked to make a dyesT or dnoT rating using the two buttons with
Fig. 1. Experimental procedure showing the functional run including 12
epochs of 60 s each. Two olfactory conditions were performed: one
detection condition with three epochs (Da, Db, Dc) and one familiarity
condition with three epochs (Fa, Fb, Fc). Example of an epoch (Da) for
which 15 stimuli (from S1 to S15) were delivered. R, rest.
their dominant hand. For half of the subjects, dyesT and dnoTresponses were obtained with the index and the middle fingers,
respectively. For the other half of the subjects, the meaning of the
two key-press buttons was reversed. For the R condition, no
stimulation was provided and the subjects were instructed not to
use the key-press buttons.
General instructions were provided to subjects before the
functional run. During the run, and 3 s before each experimental
condition (F, D or R), subjects were instructed orally by means of
specific keywords (dfamiliarity,T ddetection,T and drestT) which task
was to be performed next. Subjects wore earplugs to protect
hearing from excessive scanner noise and kept their eyes closed
during scanning. The day before the fMRI examination, subjects
were trained outside the MR facility to breathe regularly, to detect
odorants without sniffing during normal inspiration, and to give the
most rapid possible response (odor vs. no odor) using the buttons.
Imaging parameters
Functional MR imaging was performed on a 1.5-T MR imager
(Philips NT). Twenty-five adjacent 5-mm-thick axial slices were
imaged. The imaging volume covered the whole brain and was
oriented parallel to the bicommissural plane. The image planes
were positioned from scout images acquired in the sagittal plane. A
3D three-shot PRESTO MR imaging sequence (Liu et al., 1993)
was used with the following parameters: TR = 26 ms, TE = 38 ms,
flip angle = 148, field-of-view = 256 � 205 mm2, imaging matrix =
64 � 51 (voxel size of 4 � 4 � 5 mm3). This sequence is less
prone to magnetic susceptibility artifacts than the usual echo planar
imaging (EPI) sequence (Van Gelderen et al., 1995), particularly in
the OFC and mesial temporal region (Zald and Pardo, 2000).
During the functional run, the volume of interest was scanned 144
times successively. The signal was averaged three times, leading to
an acquisition time per volume of 5 s. A high-resolution anatomical
3D T1-weighted MR scan was acquired before the functional run.
Data processing and statistical analyses
Functional images were analyzed using SMP99 (Wellcome
Department of Cognitive Neurology, London, UK). Image process-
ing included interscan realignment, spatial normalization to stereo-
tactic space as defined by the Montreal Neurological Institute
(MNI) reference brain template, and image smoothing with a three-
dimensional Gaussian kernel (FWMH: 8 � 8 � 10 mm3) to
overcome residual anatomical variability during group analysis,
increase the signal-to-noise ratio and conform to Random Field
Theory assumptions underlying the statistical analysis (Friston et
al., 1995a). A boxcar reference function was convolved with
SPM99’s dcanonical hemodynamicT response function. A high-pass
filter (cutoff frequency of 1/720 Hz) was used to eliminate
instrumental and physiological very low frequency signal fluctua-
tions. Global differences in BOLD signal were covaried out from all
voxels, and comparisons across conditions were effected with t
tests. The statistical significance of signal differences was assessed
through Z scores in an omnibus sense, using an uncorrected height
threshold (P b 0.001). Only clusters of more than 10 adjacent
activated voxels were taken into account as a significant hemody-
namic response (Z N 3.20 at voxel level). Duvernoy’s (1991) and
Mai et al.’s (1997) anatomic atlases were used to localize and
describe activated anatomic regions as the often used Talairach’s
atlas (Talairach and Tournoux, 1988) describes subcortical and
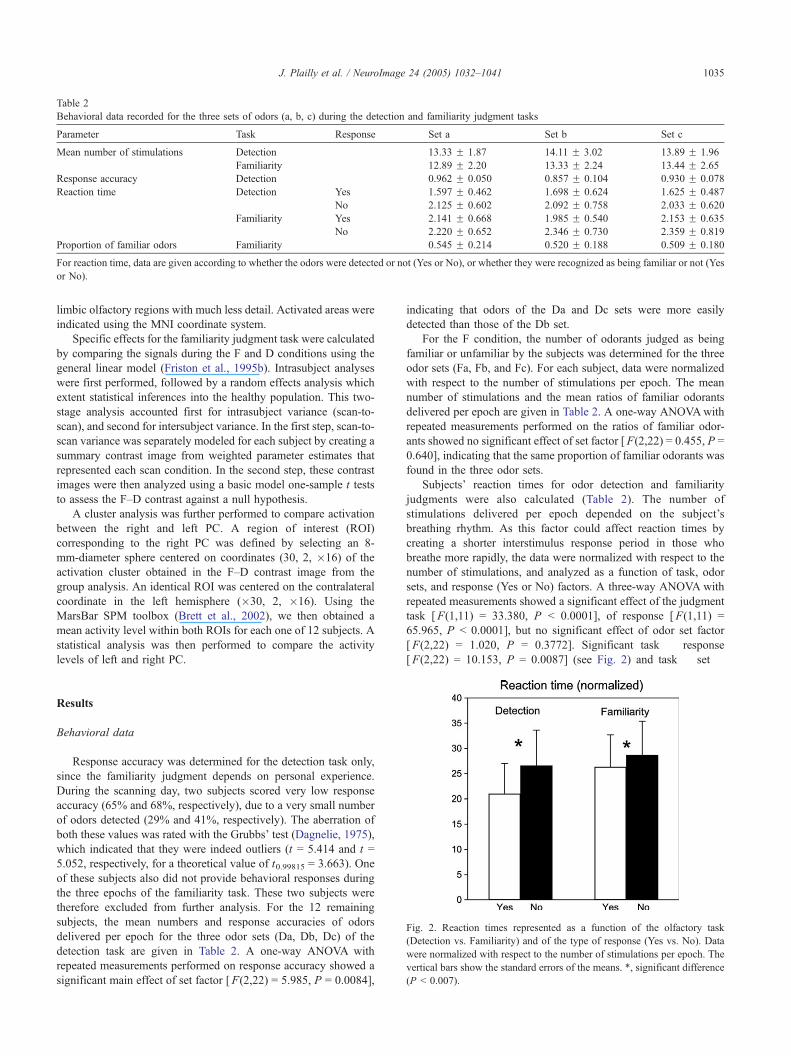
Table 2
Behavioral data recorded for the three sets of odors (a, b, c) during the detection and familiarity judgment tasks
Parameter Task Response Set a Set b Set c
Mean number of stimulations Detection 13.33 F 1.87 14.11 F 3.02 13.89 F 1.96
Familiarity 12.89 F 2.20 13.33 F 2.24 13.44 F 2.65
Response accuracy Detection 0.962 F 0.050 0.857 F 0.104 0.930 F 0.078
Reaction time Detection Yes 1.597 F 0.462 1.698 F 0.624 1.625 F 0.487
No 2.125 F 0.602 2.092 F 0.758 2.033 F 0.620
Familiarity Yes 2.141 F 0.668 1.985 F 0.540 2.153 F 0.635
No 2.220 F 0.652 2.346 F 0.730 2.359 F 0.819
Proportion of familiar odors Familiarity 0.545 F 0.214 0.520 F 0.188 0.509 F 0.180
For reaction time, data are given according to whether the odors were detected or not (Yes or No), or whether they were recognized as being familiar or not (Yes
or No).
J. Plailly et al. / NeuroImage 24 (2005) 1032–1041 1035
limbic olfactory regions with much less detail. Activated areas were
indicated using the MNI coordinate system.
Specific effects for the familiarity judgment task were calculated
by comparing the signals during the F and D conditions using the
general linear model (Friston et al., 1995b). Intrasubject analyses
were first performed, followed by a random effects analysis which
extent statistical inferences into the healthy population. This two-
stage analysis accounted first for intrasubject variance (scan-to-
scan), and second for intersubject variance. In the first step, scan-to-
scan variance was separately modeled for each subject by creating a
summary contrast image from weighted parameter estimates that
represented each scan condition. In the second step, these contrast
images were then analyzed using a basic model one-sample t tests
to assess the F–D contrast against a null hypothesis.
A cluster analysis was further performed to compare activation
between the right and left PC. A region of interest (ROI)
corresponding to the right PC was defined by selecting an 8-
mm-diameter sphere centered on coordinates (30, 2, �16) of the
activation cluster obtained in the F–D contrast image from the
group analysis. An identical ROI was centered on the contralateral
coordinate in the left hemisphere (�30, 2, �16). Using the
MarsBar SPM toolbox (Brett et al., 2002), we then obtained a
mean activity level within both ROIs for each one of 12 subjects. A
statistical analysis was then performed to compare the activity
levels of left and right PC.
Fig. 2. Reaction times represented as a function of the olfactory task
(Detection vs. Familiarity) and of the type of response (Yes vs. No). Data
were normalized with respect to the number of stimulations per epoch. The
vertical bars show the standard errors of the means. *, significant difference
(P b 0.007).
Results
Behavioral data
Response accuracy was determined for the detection task only,
since the familiarity judgment depends on personal experience.
During the scanning day, two subjects scored very low response
accuracy (65% and 68%, respectively), due to a very small number
of odors detected (29% and 41%, respectively). The aberration of
both these values was rated with the Grubbs’ test (Dagnelie, 1975),
which indicated that they were indeed outliers (t = 5.414 and t =
5.052, respectively, for a theoretical value of t0.99815 = 3.663). One
of these subjects also did not provide behavioral responses during
the three epochs of the familiarity task. These two subjects were
therefore excluded from further analysis. For the 12 remaining
subjects, the mean numbers and response accuracies of odors
delivered per epoch for the three odor sets (Da, Db, Dc) of the
detection task are given in Table 2. A one-way ANOVA with
repeated measurements performed on response accuracy showed a
significant main effect of set factor [F(2,22) = 5.985, P = 0.0084],
indicating that odors of the Da and Dc sets were more easily
detected than those of the Db set.
For the F condition, the number of odorants judged as being
familiar or unfamiliar by the subjects was determined for the three
odor sets (Fa, Fb, and Fc). For each subject, data were normalized
with respect to the number of stimulations per epoch. The mean
number of stimulations and the mean ratios of familiar odorants
delivered per epoch are given in Table 2. A one-way ANOVAwith
repeated measurements performed on the ratios of familiar odor-
ants showed no significant effect of set factor [F(2,22) = 0.455, P =
0.640], indicating that the same proportion of familiar odorants was
found in the three odor sets.
Subjects’ reaction times for odor detection and familiarity
judgments were also calculated (Table 2). The number of
stimulations delivered per epoch depended on the subject’s
breathing rhythm. As this factor could affect reaction times by
creating a shorter interstimulus response period in those who
breathe more rapidly, the data were normalized with respect to the
number of stimulations, and analyzed as a function of task, odor
sets, and response (Yes or No) factors. A three-way ANOVA with
repeated measurements showed a significant effect of the judgment
task [F(1,11) = 33.380, P b 0.0001], of response [F(1,11) =
65.965, P b 0.0001], but no significant effect of odor set factor
[F(2,22) = 1.020, P = 0.3772]. Significant task � response
[F(2,22) = 10.153, P = 0.0087] (see Fig. 2) and task � set �

Table 3
Areas activated in the F–D contrast
Brain region L/R k Z value MNI coordinates
x y z
Amygdala L 54 4.29 �24 �6 �12
Cingulate gyrus L 67 4.21 �8 16 40
Mid-fusiform gyrus R 432 3.93 40 �40 �16
Amygdala R 3.85 20 �16 �10
Hippocampal
region (CA3)
R 3.84 22 �20 �8
Temporal
piriform cortex
R 3.75 30 2 �16
Parahippocampal
gyrus
R 3.61 42 �16 �10
Posterior Insula R 3.29 46 �10 �6
Superior occipital
gyrus
L 62 3.79 �16 �100 14
Inferior frontal gyrus,
pars orbitalis
L 13 3.27 �50 28 2
Note. F, familiarity; D, detection; L, left; R, right; k, size of the cluster in
number of connected voxels; x, y, z, MNI coordinates in mm of the
maximum in the Montreal Neurological Institute Brain template; CA,
Cornu Ammonis.
J. Plailly et al. / NeuroImage 24 (2005) 1032–10411036
response [F(2,22) = 3.833, P = 0.0373] interactions were noted.
Mean comparisons indicated that reaction times in the D condition
were longer when no odor was detected (P b 0.0001), longer in the
F condition when odors were perceived as more unfamiliar (P b
0.0070), and longer with unfamiliar odors during familiarity
judgments (as compared to detection judgments; P b 0.0001).
Breathing data
Breathing changes could be expected as a function of epoch, as
during F-epochs odors were delivered at each inspiration, and
during D epochs odors were delivered on 50% of inspirations.
Inspiratory airflow measures were therefore analyzed as a function
of task (Detection and Familiarity), three epochs (a, b, and c), and
subject responses (Yes and No) (Fig. 3). A three-way ANOVAwith
repeated measurements showed no significant effect of task
[F(1,22) = 2.169, P = 0.169], epochs [F(2,22) = 0.415, P =
0.665], and response [F(1,22) = 1.409, P = 0.260] factors.
fMRI data
F–D contrast
Familiarity-specific responses (F–D) were present in the right
temporal piriform area (30, 2, �16; Z = 3.75), spanning the
cortico-amygdaloid transition area, the preamygdalar claustrum,
the periamygdalar area, and the lateral amygdaloid nucleus (22,
�16, �10; Z = 3.85; Table 3 and Fig. 4A). PC activation was not
detected in the left hemisphere (using the same statistical
significance threshold), but strong activation was present in the
amygdala (�24, �6, �12; Z = 4.29) spanning approximate areas
for the basomedial, basolateral, central, lateral and medial
amygdaloid nuclei, and the anterior amygdaloid area. Familiarity-
related activation in the right PC extended into the right mid-
fusiform gyrus (40, �40, �16; Z = 3.93) and hippocampal region
(Fig. 4B; 22, �22, �8; Z = 3.84). The familiarity judgment task
also activated the left cingulate gyrus (�8, 16, 40; Z = 4.21) and
the left inferior frontal gyrus in its opercular part (�50, 28, 2; Z =
3.27), as depicted in Fig. 4C. Finally, we noted significant
activation in the left occipital gyrus (�16, �100, 14; Z = 3.79)
and the right middle frontal gyrus (44, 20, �18; Z = 3.40).
Fig. 3. Inspiratory airflow as a function of the olfactory task (Detection vs.
Familiarity) and of the type of response (Yes vs. No). Data were normalized
with respect to the number of stimulations per epoch. The vertical bars
show the standard errors of the means.
Comparison of activations between the right and left PC
Mean activity levels were measured in the right and left PC
using ROIs. A one-way ANOVA with repeated measurements on
these data showed significantly higher right than left PC activation
[F(1,11) = 4.846, P = 0.0499].
Discussion
The aim of this fMRI study was to determine the cerebral
regions that mediate judgments of odor familiarity. With such an
approach, we identified odor-evoked neural responses in several
olfactory and limbic regions, including piriform cortex, amygdala,
hippocampus, the pars orbitalis of the inferior frontal gyrus, and
the mid-fusiform gyrus.
Activation of the mesial temporal region
Olfactory-related activation of mesial temporal regions remains
inconsistent across studies. Whereas some authors have reported
activation in PC in PET studies (e.g., Kareken et al., 2001, 2003,
2004; Savic et al., 2000; Small et al., 1997; Zatorre et al., 1992),
we did not find any mesial temporal activation in our previous
studies (Royet et al., 1999, 2001). Piriform cortex activation has
also been inconsistently detected across fMRI studies (e.g., Sobel
et al., 1998; Yousem et al., 1999). A well-known problem with the
EPI pulse sequence usually applied in fMRI is magnetic
susceptibility artefact, which induces signal loss in these regions
(Zald and Pardo, 2000). The PRESTO sequence used in the current
study appears to reduce these artifacts and to provide stronger
signal in the regions affected by susceptibility differences.
Habituation may also contribute to signal loss in these ventral
regions, particularly with blocked designs and the use of only one
or two odorants (Poellinger et al., 2001; Sobel et al., 2000). Since
we used a different odorant on each breathing cycle (from 12 to 20
different odorants per 60-s epoch, depending on the duration of
breathing cycle of the subject), we minimized both self-adaptation

Fig. 4. Localization of task-specific activations in the F–D contrasts. (A) Piriform cortex; (B) hippocampus; (C) inferior frontal gyrus. Neural responses are
overlaid on coronal and axial sections from a subject’s normalized T1-weighted brain image. Clusters were thresholded at t = 3.10.
J. Plailly et al. / NeuroImage 24 (2005) 1032–1041 1037
(Engen, 1982) and sensory habituation (Demonet et al., 1993). As
in our previous fMRI study (Royet et al., 2003), the present data
suggest that the pulse sequence and stimulation paradigm provided
sufficient sensitivity to detect activation in primary olfactory areas.
A PC activation that is stronger in the F than D condition could
be explained by different factors. A possible confound lies first of
all in the higher proportion of odorous stimulations delivered in
the F than D condition. We decided for this particular design
because we considered it was important to keep the subjects’
attention focused during the detection task (in which they had to
respond dyesT and dnoT equivalently). At the same time, if we had
presented blanks during the familiarity task, this might have
confused the subjects and this would have introduced a third type
of event (no odor in addition to the familiar and the unfamiliar
odors). Further experiments are therefore needed to rule out this
specific confound. It would be of interest, in particular, to design
an event-related fMRI study in which familiar and unfamiliar
odors would be distinguished as discrete events in both conditions.
Unfamiliar odors of the F and D conditions could then be
contrasted and any PC activation could then be specifically
associated with the F task. It should nevertheless be noted that the
piriform cortex may become activated in retrieving odor associ-
ations alone, without any direct chemosensory stimulation
(Gottfried et al., 2004). This strongly suggests that the task
performed by subjects may be a decisive factor in activating the
piriform cortex.
A second source of potential confound may lie in the
different respiratory patterns across conditions, as it has

J. Plailly et al. / NeuroImage 24 (2005) 1032–10411038
previously been shown that sniffing alone can induce piriform
activation (Sobel et al., 1998; but also see Kareken et al., 2004
for contradictory results), and that odor imagery alone can lead
to breathing differences (Bensafi et al., 2003). In the present
study, however, inspiratory airflow was not higher in the F
condition than in the D condition. Finally, a third source of
potential confound is identified in the differences in odor
familiarity, since odors selected a priori as being familiar were
only delivered in the F condition. Nevertheless, the same number
of a priori unfamiliar odors were presented in both conditions.
We then noted that reaction times were much longer when the
unfamiliar odors were presented in the F condition, and even
longer than reaction times of familiar odors presented in this
same condition. This suggests that subjects performed different
judgments across conditions, and that activation differences in
PC could be better explained by the type of task than by
perceptual differences in odor familiarity. This could also explain
why Savic and Berglund (2004) did not find differential
activation in PC when subjects passively smelled familiar or
unfamiliar odorants. It nevertheless stands to reason, however,
that the task (when not explicitly required by the experiment)
should be facilitated by the intrinsic properties of the odors
themselves: That is, highly familiar odors would more easily
facilitate familiarity judgments, and highly pleasant or unpleasant
odors would more easily encourage emotional judgments.
Involvement of piriform cortex in recognition memory
Recognition memory is known to involve two different
processes: familiarity and recollection (Bogacz et al., 2001;
Mandler, 1980; Rajaram, 1998). According to the ddual processtheoryT, processes underlying familiarity are perceptual in nature,
and those subserving recollection include the retrieval of con-
textual information. Lehrner et al. (1999) demonstrated that these
two forms of recognition memory processes also exist in olfaction.
In other words, familiarity judgments are made on the basis of
feelings devoid of specific information about the encoding
episode, and thus relate to implicit memory. By contrast,
recollection is more directly tied to specific events, and thus
relates to explicit memory. To illustrate these concepts, Bogacz
et al. (2001) note that, b. . .it is not uncommon to be able to
recognize that a person is familiar to us even though we cannot
immediately recollect anything more about the person or our
previous encounters with themQ (Bogacz et al., 2001). Since
familiarity judgments are inherent in recognition memory, our
results relate to previous findings in humans indicating that PC is
involved in long-term odor recognition memory (Dade et al.,
2002). More recently, Gottfried et al. (2004) further reported that
PC responds to non-olfactory stimuli with which odors were
previously associated.
Further experiments are needed to compare activation
produced by different memory processes. Familiarity and
recollection could involve slightly different neural networks,
but the present experimental design cannot distinguish between
them. Our findings nevertheless cohere with a large body of
research using animal models, and lend support to the theory that
the piriform cortex is involved in learning- and memory-related
processes (e.g., Datiche et al., 2001; Schoenbaum and Eichen-
baum, 1995). For instance, synaptic potentiation has been shown
to occur in rat PC in vitro (e.g., Jung et al., 1990; Saar et al.,
2002) and in vivo at the conclusion of learning (Litaudon et al.,
1997; Roman et al., 1993). These findings are thus consistent
with models demonstrating that the primary olfactory cortex is a
parallel-distributed architecture characteristic of associative mem-
ory systems (e.g., Bower, 1991; Haberly, 2001; Haberly and
Bower, 1989).
Lateralization of familiarity judgment process
The current study shows that activation of the right PC was
stronger than that of the left PC. This is in line with the results of a
large body of other research. In a monorhinal odor recognition
task, Savic et al. (2000) noted significant right, but not left,
piriform activity. Although Dade et al. (2002) did not explicitly
report hemispheric asymmetry for long-term olfactory memory,
their results distinctly indicated strong activation in right OFC, and
more activation in right than left PC. Interestingly, Gottfried et al.’s
study of cross-modal visual–olfactory associations also showed
unilateral right PC activation, and in this case without direct
olfactory stimulation. Using behavioral measures, Broman et al.
(2001) finally observed that odors presented to the right nostril
were rated as more familiar than odors presented to the left nostril.
They also reported that episodic recognition via the right nostril
tended to nominally have more dknowT responses and fewer
drememberT responses than did odors recognized via the left nostril,which is in keeping with the right-nostril advantage for familiarity.
Taken together, these findings are consistent with the notion that
left temporal lobe structures mediate processing of distinctiveness
(i.e., what clearly distinguishes one percept from another), whereas
right temporal lobe structures subserve processes underlying
perceptual fluency (i.e., what involves perceptual analysis of the
surface features of an item; Blaxton and Theodore, 1997; Rajaram,
1998). Such a perceptual analysis of surface features is especially
observed for odors which are intrinsically difficult to name
(Lawless and Engen, 1977).
Findings in brain-damaged patients and results from neuro-
imaging studies converge with these data. For instance, epilepsy
patients with left temporal lobe lesions who were asked to
recognize previously seen abstract designs provided more dknowTthan drememberT responses, whereas right temporal lesioned
patients showed the opposite pattern (Blaxton and Theodore,
1997). Along the same line, Henson et al. (1999) explored word
recognition with fMRI and showed a dissociation whereby a
dknowT judgment induced right frontal activation, and a
drememberT judgment induced left frontal activation.
In conclusion, the present data, which are consistent with our
previous work in PET, indicate a preferential involvement of the
right hemisphere in familiarity judgments (Royet et al., 1999,
2001). An intriguing result in the present study is the lack of
activation in right OFC. When specifically examining activation
resulting from familiarity versus rest, and from detection versus
rest, we nevertheless saw activation in the right OFC for both
contrasts (44, 32, �16; Z = 3.62 and 46, 30, �12; Z = 2.79,
respectively). This could explain the lack of activation when we
compared images in the familiarity versus detection contrast.
Participation of the hippocampal region, inferior frontal and
mid-fusiform gyri in modality-independent mnemonic and semantic
processing
Since only a few authors have previously reported hippocampal
activations in olfaction studies (e.g., Kareken et al., 2003; Suzuki

J. Plailly et al. / NeuroImage 24 (2005) 1032–1041 1039
et al., 2001), the right hippocampal activation during the familiarity
judgment task was not anticipated in the present study. Inconsistent
activation of the hippocampal formation is not specific to olfaction,
and has been characteristic of other studies of memory (Andreasen
et al., 1995; Shallice et al., 1994; Tulving et al., 1996). Our data are
nevertheless consistent with a recent finding showing hippocampal
activation during the retrieval of olfactory episodic memories
(Gottfried et al., 2004).
Lesion studies recently examined whether the brain structures
that comprise the medial temporal lobe memory system (i.e., the
hippocampal and parahippocampal regions) differ in how they
support recollective and familiarity components (Manns et al.,
2003; Yonelinas et al., 2002). Our data do not permit distinguish-
ing these aspects of memory, as the subjects may well have had
consciously evoked memories from the odorants. They are,
however, consistent with the idea that both regions probably
contribute to olfactory recognition memory.
Activation of the left inferior frontal gyrus, in the pars
orbitalis, during familiarity judgments further supports the
hypothesis that this region is involved in the selection and
integration of semantic information in a modality-independent
manner (Homae et al., 2002; Kareken et al., 2003). In a recent
study, Savic and Berglund (2004) found that left frontal and right
parahippocampal region activation positively correlated with
familiarity ratings, showing the engagement of semantic circuits
during passive smelling of familiar odorants. Along the same line,
the mid-fusiform gyrus activation in the current study might lead
to similar interpretations, since it has further been associated with
visual, tactile and auditory recognition and categorization of
objects (Adams and Janata, 2002; Joseph and Gathers, 2003;
Stoeckel et al., 2003). Its involvement in olfactory object
recognition therefore reinforces the idea of the polymodal nature
of this area (Adams and Janata, 2002) and its role in semantic
processing (Price, 2000; Wagner et al., 1998).
Conclusion
Complementing previous PET studies that demonstrate right
OFC involvement in odor familiarity judgments, the present fMRI
study shows that right PC is also activated during this task, an
activation that may be related to olfactory recognition memory. In
previous fMRI and PET studies, we demonstrated that a neural
network in the left hemisphere, involving the OFC and primary
olfactory areas, mediated olfactory hedonic perception (Royet et al.,
2000, 2003). It thus appears that odor processing activates a large
neural network involving both hemispheres. Nevertheless, this
network possesses hemispheric predominance depending on the
type of olfactory task performed (see Royet and Plailly, 2004, for
review). The present data provide further evidence that the right
hippocampal region, left inferior frontal gyrus and mid-fusiform
gyrus take part in recognition memory, likely cross-modally to
assist in gathering relevant associations to enable identification of
olfactory percepts.
Acknowledgments
We thank the technical team (M. Vigouroux, B. Bertrand, and
V. Farget) for designing and building the stimulation and recording
materials and J.P. Lomberget and M.B. Sanglerat for medical
examinations of subjects participating in the study. We are grateful
to the companies Givaudan, International Flavors and Fragrances,
Lenoir, Davenne, and Perlarom for supplying the odorants used in
this study. This work was supported by research grants from the
dRegion Rhone-AlpesT and the dGIS Sciences de la Cognition,T thedCentre National de la Recherche Scientifique,T and the dUniversiteClaude-Bernard de Lyon.T
References
Adams, R.B., Janata, P., 2002. Comparison of neural circuits underlying
auditory and visual object categorization. NeuroImage 16, 361–377.
Anderson, A.K., Christoff, K., Stappen, I., Panitz, D., Ghahremani, D.G.,
Glover, G., Gabrieli, J.D., Sobel, N., 2003. Dissociated neural
representations of intensity and valence in human olfaction. Nat.
Neurosci. 6, 196–202.
Andreasen, N.C., O’Leary, D.S., Cizadlo, T., Arndt, S., Rezai, K., Watkins,
G.L., Ponto, L.L., Hichwa, R.D., 1995. Remembering the past: two
facets of episodic memory explored with positron emission tomography.
Am. J. Psychiatry 152, 1576–1585.
Bensafi, M., Porter, J., Pouliot, S., Mainland, J., Johnson, B., Zelano, C.,
Young, N., Bremner, E., Aframian, D., Khan, R., Sobel, N., 2003.
Olfactomotor activity during imagery mimics that during perception.
Nat. Neurosci. 6, 1142–1144.
Blaxton, T.A., Theodore, W.H., 1997. The role of the temporal lobes in
recognizing visuospatial materials: remembering versus knowing. Brain
Cogn. 35, 5–25.
Bogacz, R., Brown, M.W., Giraud-Carrier, C., 2001. Model of familia-
rity discrimination in the perirhinal cortex. J. Comput. Neurosci. 10,
5–23.
Bower, J.M., 1991. Piriform cortex and olfactory recognition. In: Davis,
J.D., Eichenbaum, H. (Eds.), Olfaction: a Model System for Computa-
tional Neuroscience. MIT Press, Cambridge, MA, pp. 266–285.
Brett, M., Anton, J.L., Valabregue, R., Poline, J.B., 2002. Region of interest
analysis using an SPM toolbox [abstract]. The 8th International
Conference on Functional Mapping of the Human Brain, June 2–6,
2002, Sendai, Japan. Available on CD-ROM in NeuroImage 16 (2).
Broca, P., 1863. Localisation des fonctions cerebrales. Siege de la faculte du
langage articule. Bull. Soc. Anthropol. Paris 4, 200–204.
Broman, D.A., Olsson, M.J., Nordin, S., 2001. Lateralization of olfactory
cognitive functions: effects of rhinal side of stimulation. Chem. Senses
26, 1187–1192.
Dade, L.A., Jones-Gotman, M., Zatorre, R.J., Evans, A.C., 1998. Human
brain function during odor encoding and recognition. A PET activation
study. Ann. N. Y. Acad. Sci. 855, 572–574.
Dade, L.A., Zatorre, R.J., Jones-Gotman, M., 2002. Olfactory learning:
convergent findings from lesion and brain imaging studies in humans.
Brain 125, 86–101.
Dagnelie, P., 1975. Theorie et Methodes Statistiques, Les Presses
Agronomiques de Gembloux, A.S.B.L 2.
Datiche, F., Roullet, F., Cattarelli, M., 2001. Expression of Fos in the
piriform cortex after acquisition of olfactory learning: an immunohis-
tochemical study in the rat. Brain Res. Bull. 55, 95–99.
Demonet, J.F., Wise, R., Frackowiak, R.S.J., 1993. Les fonctions
linguistiques explorees en tomographie par emission de positons.
Med./Sci. 9, 934–942.
Duvernoy, H.M., 1991. The Human Brain-Surface, Three Dimensional
Sectional Anatomy and MRI. Springer, Wien.
Engen, T., 1982. The Perception of Odors. Academic Press, New York.
Friston, K.J., Ashburner, J., Frith, C.D., Poline, J.B., Healther, J.D.,
Frackowiak, R.S.J., 1995a. Spatial registration and normalisation of
images. Hum. Brain Mapp. 3, 165–189.
Friston, K.J., Holmes, A.P., Worsley, K.J., Poline, J.B., Frith, C.D.,
Frackowiak, R.S.J., 1995b. Statistical parametric maps in functional
imaging: a general linear approach. Hum. Brain Mapp. 2, 189–210.

J. Plailly et al. / NeuroImage 24 (2005) 1032–10411040
Gottfried, J.A., Deichmann, R., Winston, J.S., Dolan, R.J., 2002. Functional
heterogeneity in human olfactory cortex: an event-related functional
magnetic resonance imaging study. J. Neurosci. 22, 10819–10828.
Gottfried, J.A., Smith, A.P.R., Rugg, M.D., Dolan, R.J., 2004. Remem-
brance of odors past: human olfactory cortex in cross-modal recognition
memory. Neuron 42, 687–695.
Haberly, L.B., 2001. Parallel-distributed processing in olfactory cortex: new
insights from morphological and physiological analysis of neuronal
circuitry. Chem. Senses 26, 551–576.
Haberly, L.B., Bower, J.M., 1989. Olfactory cortex: model circuit for study
of associative memory. TINS 12, 258–264.
Henson, R.N.A., Rugg, M.D., Shallice, T., Josephs, O., Dolan, R.J.,
1999. Recollection and familiarity in recognition memory: an event-
related functional magnetic resonance imaging study. J. Neurosci. 19,
3962–3972.
Homae, F., Hashimoto, R., Nakajima, K., Miyashita, Y., Sakai, K.L., 2002.
From perception to sentence comprehension: the convergence of
auditory and visual information of language in the left inferior frontal
cortex. NeuroImage 16, 883–900.
Joseph, J.E., Gathers, A.D., 2003. Effects of structural similarity on neural
substrates for object recognition. Cogn. Affect. Behav. Neurosci. 3,
1–16.
Jung, M.W., Larson, J., Lynch, G., 1990. Long-term potentiation of
monosynaptic EPSPs in rat piriform cortex in vitro. Synapse 6,
279–283.
Kareken, D.A., Doty, R.L., Moberg, P.J., Mosnik, D., Hsing Chen, S.,
Farlow, M.R., Hutchins, G.D., 2001. Olfactory-evoked regional
cerebral blood flow in Alzheimer’s disease. Neuropsychology 15,
18–29.
Kareken, D.A., Mosnik, D.M., Doty, R.L., Dzemidzic, M., Hutchins,
G.D., 2003. Functional anatomy of human odor sensation, discrim-
ination, and identification in health and aging. Neuropsychology 17,
482–495.
Kareken, D.A., Sabri, M., Radnovich, A.J., Claus, E., Foresman, B.,
Hector, D., Hutchins, G.D., 2004. Functional anatomy of human odor
sensation, discrimination, and identification in health and aging.
NeuroImage 22, 456–465.
Lawless, H., Engen, T., 1977. Associations to odors: interference,
mnemonics, and verbal labeling. J. Exp. Psychol. Hum. Learn. Mem.
3, 52–59.
Lehrner, J.P., Walla, P., Laska, M., Deecke, L., 1999. Different forms of
human odor memory: a developmental study. Neurosci. Lett. 272,
17–20.
Litaudon, P., Mouly, A.M., Sullivan, R., Gervais, R., Cattarelli, M., 1997.
Learning-induced changes in rat piriform cortex activity mapped using
multisite recording with voltage sensitive dye. Eur. J. Neurosci. 9,
1593–1602.
Liu, G., Sobering, G., Duyn, J., Moonen, C., 1993. A functional MRI
technique combining principles of echo-shifting with a train of
observations (PRESTO). Magn. Reson. Med. 30, 764–768.
Mai, J.K., Assheuer, J., Paxinos, G., 1997. Atlas of the Human Brain.
Academic Press, San Diego.
Mandler, G., 1980. Recognizing: the judgment of previous occurrence.
Psychol. Rev. 87, 252–271.
Manns, J.R., Hopkins, R.O., Reed, J.M., Kitchener, E.G., Squire, L.R.,
2003. Recognition memory and the human hippocampus. Neuron 37,
171–180.
Poellinger, A., Thomas, R., Lio, P., Lee, A., Makris, N., Rose, B.R.,
Kwong, K.K., 2001. Activation and habituation in olfaction—An fMRI
study. NeuroImage 13, 547–560.
Price, C.J., 2000. The anatomy of language: contributions from functional
neuroimaging. J. Anat. 3, 335–359.
Rajaram, S., 1998. The effects of conceptual salience and perceptual
distinctiveness on conscious recollection. Psychon. Bull. Rev. 5, 71–78.
Roman, F.S., Chaillan, F.A., Soumireu-Mourat, B., 1993. Long-term
potentiation in rat piriform cortex following discrimination learning.
Brain Res. 601, 265–272.
Royet, J.P., Koenig, O., Gregoire, M.C., Cinotti, L., Lavenne, F., Le Bars, D.,
Costes, N., Vigouroux, M., Farget, V., Sicard, G., Holley, A., Mauguiere,
F., Comar, D., Froment, J.C., 1999. Functional anatomy of perceptual
and semantic processing for odours. J. Cogn. Neurosci. 11, 94–109.
Royet, J.P., Zald, D., Versace, R., Costes, N., Lavenne, F., Koenig, O.,
Gervais, R., 2000. Emotional responses to pleasant and unpleasant
olfactory, visual, and auditory stimuli: a positron emission tomography
study. J. Neurosci. 20, 7752–7759.
Royet, J.P., Hudry, J., Zald, D.H., Godinot, D., Gregoire, M.C., Lavenne, F.,
Costes, N., Holley, A., 2001. Functional neuroanatomy of different
olfactory judgments. NeuroImage 13, 506–519.
Royet, J.P., Plailly, J., Delon-Martin, C., Kareken, D.A., Segebarth, C.,
2003. Functional anatomy of the emotional responses to odors:
Influence of hedonic valence, hedonic judgment, handedness, and
gender. NeuroImage 20, 713–728.
Royet, J.P., Plailly, J., 2004. Lateralization of olfactory processes. Review.
Chem. Senses 29, 731–745.
Saar, D., Grossman, Y., Barkai, E., 2002. Learning-induced enhancement of
postsynaptic potentials in pyramidal neurons. J. Neurophysiol. 87,
2358–2363.
Savic, I., Berglundd, H., 2004. Passive perception of odors and semantic
circuits. Hum. Brain Mapp. 21, 271–278.
Savic, I., Gulyas, B., Larsson, M., Roland, P., 2000. Olfactory functions are
mediated by parallel and hierarchical processing. Neuron 26, 735–745.
Schoenbaum, G., Eichenbaum, H., 1995. Information coding in the
rodent prefrontal cortex. I. Single-neuron activity in orbitofrontal
cortex compared with that in pyriform cortex. J. Neurophysiol. 74,
733–750.
Shallice, T., Fletcher, P., Frith, C.D., Grasby, P., Frackowiak, R.S., Dolan,
R.J., 1994. Brain regions associated with acquisition and retrieval of
verbal episodic memory. Nature 368, 633–635.
Small, D.N., Jones-Gotman, M., Zatorre, R.J., Petrides, M., Evan, A.C.,
1997. Flavor processing: more than the sum of its parts. NeuroReport 8,
3913–3917.
Sobel, N., Prabhakaran, V., Desmond, J.E., Glover, G.H., Goode, R.L.,
Sullivan, E.V., Gabrieli, J.D.E., 1998. Sniffing and smelling: separate
subsystems in the human olfactory cortex. Nature 392, 282–286.
Sobel, N., Prabhakaran, V., Zhao, Z., Desmond, J.E., Glover, G.H.,
Sullivan, E.V., Gabrieli, J.D.E., 2000. Time-course of odorant-induced
activation in the human primary olfactory cortex. J. Neurophysiol. 82,
537–551.
Soros, P., Knecht, S., Imai, T., Gurtler, S., Lutkenhoner, B., Ringelstein,
E.B., Henningsen, H., 1999. Cortical asymmetries of the human
somatosensory hand representation in right-and left-handers. Neurosci.
Lett. 271, 89–92.
Stoeckel, M.C., Weder, B., Binkofski, F., Buccino, G., Shah, N.J., Seitz,
R.J., 2003. A fronto-parietal circuit for tactile object discrimination: an
event-related fMRI study. NeuroImage 19, 1103–1114.
Suzuki, Y., Critchley, H.D., Suckling, J., Fukuda, R., Williams, S.C.R.,
Andrew, C., Howard, R., Ouldred, E., Bryant, C., Chir, B., Swift, C.G.,
Jackson, S.H.D., 2001. Functional magnetic resonance imaging of
odour identification: the effect of aging. J. Gerontol. 56, 756–760.
Tulving, E., Markowitsch, H.J., Craik, F.I.M., Habib, R., Houle, S., 1996.
Novelty and familiarity activations in PET studies of memory encoding
and retrieval. Cereb. Cortex 6, 71–79.
Van Gelderen, P., Ramsey, N.F., Liu, G., Duyn, J.H., Frank, J.A.,
Weinberger, D.R., Moonen, C.T., 1995. Three-dimensional functional
magnetic resonance imaging of human brain on a clinical 1.5-T scanner.
Proc. Natl. Acad. Sci. U. S. A. 92, 6906–6910.
Vigouroux, M., Bertrand, B., Farget, V., Plailly, J., Royet, J.P., 2004. A
stimulation method using odors suitable for PET and fMRI studies with
recording of physiological and behavioral signals. J. Neurosci. Methods
(in press).
Wagner, A.D., Schacter, D.L., Rotte, M., Koutstaal, W., Maril, A., Dale,
A.M., Rosen, B.R., Buckner, R.L., 1998. Building memories: remem-
bering and forgetting of verbal experiences as predicted by brain
activity. Science 281, 1188–1191.

J. Plailly et al. / NeuroImage 24 (2005) 1032–1041 1041
Weintraub, S., Mesulam, M.M., 1987. Right cerebral dominance in spatial
attention. Further evidence based on ipsilateral neglect. Arch. Neurol.
44, 621–625.
Yonelinas, A.P., Kroll, N.E., Quamme, J.R., Lazzara, M.M., Sauve, M.J.,
Widaman, K.F., Knight, R.T., 2002. Effects of extensive temporal lobe
damage or mild hypoxia on recollection and familiarity. Nat. Neurosci.
5, 1236–1241.
Yousem, D.M., Williams, S.C.R., Howard, R.O., Andrew, C., Simmons, A.,
Allin, M., Geckle, R.J., Suskind, D., Bullmore, E.T., Brammer, M.J.,
Doty, R.L., 1997. Functional MR imaging during odour stimulation:
preliminary data. Neuroradiology 204, 833–838.
Yousem, D.M., Maldjian, J.A., Siddiqi, F., Hummel, T., Alsop, D.C.,
Geckle, R.J., Bilker, W.B., Doty, R.L., 1999. Gender effects on odor-
stimulated functional magnetic resonance imaging. Brain Res. 818,
480–487.
Zald, D.H., Pardo, J.V., 1997. Emotion, olfaction, and the human amygdala:
amygdala activation during aversive olfactory stimulation. Proc. Natl.
Acad. Sci. U. S. A. 94, 4119–4124.
Zald, D.H., Pardo, J.V., 2000. Functional neuroimaging of the olfactory
system in humans. Int. J. Psychophysiol. 36, 165–181.
Zald, D.H., Donndelinger, M.J., Pardo, J.V., 1998. Elucidating dynamic
brain interactions with across-subjects correlational analyses of positron
emission tomographic data: the functional connectivity of the amygdala
and orbitofrontal cortex during olfactory tasks. J. Cereb. Blood Flow
Metab. 18, 896–905.
Zatorre, R.J., Jones-Gotman, M., Evans, A.C., Meyer, E., 1992. Functional
localization and lateralization of human olfactory cortex. Nature 360,
339–340.
Zatorre, R.J., Belin, P., Penhune, V.B., 2002. Structure and function of
auditory cortex: music and speech. Trends Cogn. Sci. 6, 37–46.














![[2019] CCJ 9 (AJ) - Election Judgments](https://static.fdokumen.com/doc/165x107/63223e3628c445989105af86/2019-ccj-9-aj-election-judgments.jpg)





