
Do trout swim better than eels? Challenges for estimating ...

Marine Biology (2006) 149: 1567–1576 DOI 10.1007/s00227-006-0308-2
RESEARCH ARTICLE
Heather M. Lamson · Jen-Chieh Shiao · Yoshiyuki Iizuka Wann-Nian Tzeng · David K. Cairns
Movement patterns of American eels (Anguilla rostrata) between salt- and freshwater in a coastal watershed, based on otolith microchemistry
Received: 5 August 2005 / Accepted: 15 March 2006 / Published online: 8 April 2006© Springer-Verlag 2006
Abstract Otolith strontium:calcium ratios were used totrace lifetime movements of American eels (Anguillarostrata) captured in salt-water bays and adjoining fresh-water ponds in Prince Edward Island, Canada. Eels wereclassiWed into migratory contingents based on theirmovement patterns. A pond with a pool-and-weir salmo-nid Wshway and a pond drained by a low-gradient chan-nel contained eels that had entered freshwater at all ages,but a pond with a 2.2 m vertical spillway contained onlyeels that had entered freshwater in the elver year. Salt-water residents were the dominant migratory contingentin salt-water bays (85% of 39), which overturns the para-digm of obligate catadromy for this species. Freshwaterresidency was the sole pattern found in the pond with thevertical spillway (100% of 12) and the majority contin-gent in the pond with the low-gradient channel (54% of24). Inter-habitat shifting was the dominant migratorycontingent in eels sampled from the pond with the pool-and-weir Wshway (85% of 20). Resident eels were estab-lished in salt- and freshwater habitats by the year after
their arrival in continental waters. Eels that shiftedbetween habitats increased their rate of inter-habitatshifting with age. The high degree of plasticity in habitatuse found in this study is consistent with worldwideAnguillid patterns as revealed by Sr:Ca.
Introduction
Anguillid eels are viewed as textbook catadromous spe-cies, spawning in the open sea, migrating to freshwaterhabitats to rear, and returning to the ocean to completetheir life cycle. There is, however, great variability in con-tinental habitat use, and anguillid eels in continentalwaters may be found in habitats ranging from full-strength salt-water to freshwater (Morrison et al. 2003;Daverat et al. 2004).
Analysis of strontium:calcium ratios of eel otolithshas greatly expanded our knowledge of eel movementpatterns, showing that some eels settle and remain in salt,brackish, or freshwater during their continental lives,while others shift among these habitats (Morrison et al.2003; Tzeng et al. 1997, 2000; Tsukamoto and Arai2001). In European (Anguilla anguilla) and Japanese(Anguilla japonica) eels, Sr:Ca data have revealed thatsome eels spend their entire lives in salt-water (Tsukam-oto et al. 1998; Arai et al. 2003). These species can thusno longer be regarded as obligate catadromous Wsh, butinstead use catadromy as a facultative life history option.
Sr:Ca studies on American eels (Anguilla rostrata) atthree locations have shown a variety of inter-habitatmovement patterns. However, none of these investiga-tions has sampled eels in full-strength salt-water. Ameri-can eels are commonly found in continental salt-waterduring their yellow (growth) phase, but it has not beendemonstrated that American eels can complete their lifecycles in the marine environment.
Secor (1999) noted that estuarine and diadromousWshes commonly exhibit varying movement patternsamong salt, brackish, and freshwater. He termed thesegroups “migratory contingents.” Tosi et al. (1988) and
Communicated by R.J. Thompson, St. John´s
H. M. LamsonBiology Department, University of New Brunswick, Fredericton, NB E3B 6E1, Canada
J. C. ShiaoInstitute of Zoology, Academia Sinica, Nankang, Taipei 11529, Taiwan, ROC
Y. IizukaInstitute of Earth Sciences, Academia Sinica, Nankang, Taipei, 11529, Taiwan, ROC
W. N. TzengInstitute of Fisheries Sciences, National Taiwan University, Taipei 10617, Taiwan, ROC
D. K. Cairns (&)Department of Fisheries and Oceans, Box 1236, Charlottetown, PE C1A 7M8, CanadaE-mail: [email protected].: +1-902-5667825Fax: +1-902-5667948

1568
Édeline and Élie (2004) found that glass eels sampledfrom marine waters showed distinct salinity preferencesin laboratory trials, some choosing salt-water while oth-ers preferred fresh. This implies that membership inmigratory contingents may be determined before arrivalat the coast. Eels which attempt to ascend rivers mayencounter natural or artiWcial obstacles. Eels under10 cm long are able to creep up wet vertical surfaces(Legault 1988). Hence dams or waterfalls where watertrickles down vertical walls may impose an age-depen-dent barrier to migration, with upstream movement pos-sible only for young eels (Cairns et al. 2004).
We used the Sr:Ca technique to investigate the ontog-eny, frequency, and directions of American eel move-ments between salt- and freshwater in a small watershedin eastern Canada. To permit the examination of eVectsof obstacle type on movement patterns, we chose a studyarea consisting of salt-water bays and adjacent freshwa-ter ponds which were formed by dams of three diVerenttypes: earthen dam with a vertical spillway, concrete damwith a salmonid Wsh ladder, and earthen dam with a low-gradient outlet channel. Patterns of movement betweensalinity zones, as indicated by Sr:Ca proWles, are used totest the following predictions:
1. Ponds formed by dams with vertical water drops (sal-monid Wsh ladder, vertical spillway) will contain onlyeels that entered at a small size, but the pond drainedby a low-gradient channel will contain eels thatentered at all sizes.
2. Eels will show a variety of migratory contingents,including residence in salt-water, residence in freshwa-ter, and movements between these habitats.
3. Some eels will show salt-water residence only, indicat-ing an exclusively marine life cycle.
4. Eels choose their migratory contingents upon arrivalin continental waters, so that colonization of salt-water and of adjacent freshwater ponds occurs simul-taneously or nearly so.
Methods
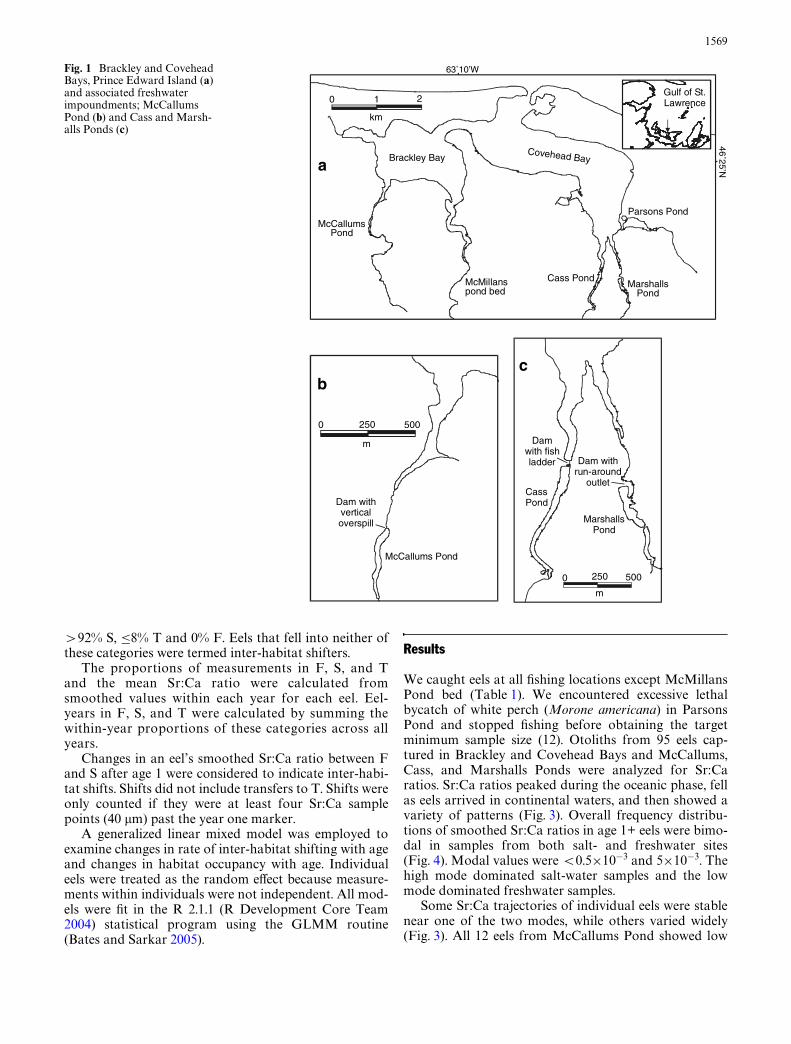
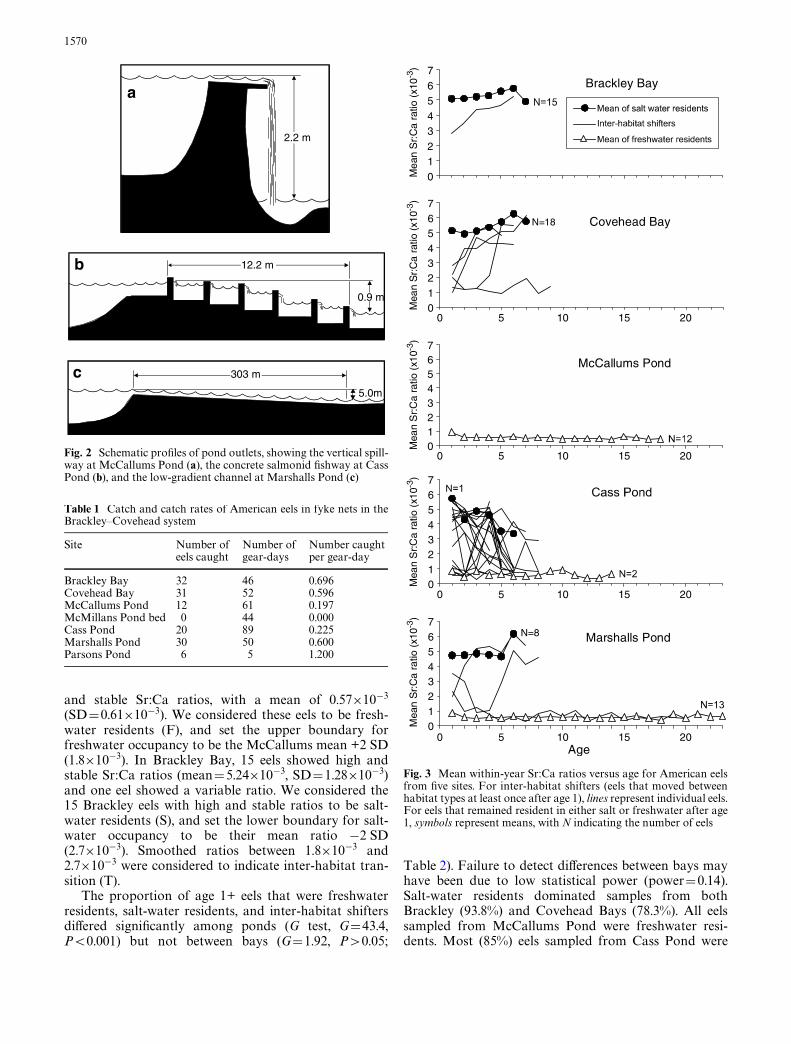
This study was conducted on the north shore of PrinceEdward Island in Brackley and Covehead Bays and asso-ciated ponds (Fig. 1). Both bays have full-strength salt-water (>28 ppt) and their combined watersheds total81 km2. Four streams entering these bays are blocked bydams at head of tide, forming freshwater impoundments.Water exits McCallums Pond to Brackley Bay by fallingvertically 2.2 m from a wooden spillway set in an earthendam (Fig. 2). Cass Pond on Covehead Bay has a 5-cham-ber pool-and-weir salmonid Wshway through whichwater drops 0.9 m over a horizontal distance of 12.2 m.Chambers are 1.8 m wide, 1.8–4.3 m long, and 0.7–1.3 mdeep. Water also leaves Cass Pond over a vertical con-crete spillway 5 m wide. Marshalls Pond drains intoCovehead Bay by a low-gradient channel with a rockybottom that falls 5.0 m over a horizontal distance of
303 m (1.7% slope). Parsons Pond connects to CoveheadBay via a culvert with wooden baZes on its Xoor that areintended to aid Wsh movement. The sole unimpoundedstream in the Brackley–Covehead system runs throughthe bed of the former McMillans Pond, whose damwashed out in 1996.
Eels were Wshed by fyke net in Brackley and Cove-head Bays, associated ponds, and the stream runningthrough McMillans Pond bed in May–November 2003.Eels were anaesthetized with clove oil, measured for totallength, weighed, and frozen until the otoliths wereremoved.
One otolith per eel was subject to microchemical anal-ysis. Otoliths were embedded in epoWx resin, then groundand polished until the primordium was exposed. Theotoliths were then carbon coated under a high vacuumevaporator prior to analysis with an electron probe mic-roanalyzer. Using a JEOL JXA-8900R system equippedwith wavelength dispersive X-ray spectrometers, Sr andCa concentrations were measured at 10 �m intervalsfrom the primordium to the otolith edge. Beam condi-tions were an acceleration voltage of 15 kV, a current of3 nA and a 5£4 �m rectangular scanning beam. A syn-thetic aragonite (CaCO3) and strontiantite (SrCO3NMNH R10065) were used for standard calibrations. Srconcentrations were measured for 80 s at Sr L� peakpositions and 20 s at both the lower and upper sides ofthe baseline. Ca was measured for 20 s at the Ca K� peakand for 10 s at both sides of the baseline. After Sr:Caratio analysis the otolith was polished to remove the car-bon layer and etched for 1–2 min with 5% EDTA toreveal annular rings for age determination.
Sr:Ca reference levels were established to identifyoccupancy of fresh, salt, and inter-habitat transitionwaters, on the basis of eels from the present study thatoccupied only one habitat type after age 1 (see Results).For each specimen, a Wve-point running mean was usedto smooth short-term Xuctuations in Sr:Ca ratios thatare likely due to otolith surface Xaws or analytic artifacts(Kotake et al. 2003). Smoothed Sr:Ca ratios that wereless than the mean +2 SD of smoothed measurements ofage 1+ eels from McCallums Pond were considered toindicate freshwater occupancy (F). Smoothed Sr:Caratios that were greater than the mean ¡2 SD ofsmoothed measurements of 15 salt-water resident age 1+eels from Brackley Bay were considered to indicate salt-water occupancy (S). Sr:Ca ratios intermediate betweenthese reference levels were considered to indicate transi-tion between habitat types (T).
Eels were assigned to one of three migratory contin-gents (freshwater residents, salt-water residents, andinter-habitat shifters) on the basis of habitat occupancypatterns. Criteria for identifying residents allowed forbrief excursions from the habitat of residence, or slightdelays in the arrival of young eels in freshwaters. An eelwas classed as a freshwater resident if its combinedsmoothed Sr:Ca ratios after age 1 were >97% F, or>92% F, ·8% T and 0% S. Salt-water residents includedeels whose post-age 1 Sr:Ca ratios were >97% S, or
1569
>92% S, ·8% T and 0% F. Eels that fell into neither ofthese categories were termed inter-habitat shifters.
The proportions of measurements in F, S, and Tand the mean Sr:Ca ratio were calculated fromsmoothed values within each year for each eel. Eel-years in F, S, and T were calculated by summing thewithin-year proportions of these categories across allyears.
Changes in an eel’s smoothed Sr:Ca ratio between Fand S after age 1 were considered to indicate inter-habi-tat shifts. Shifts did not include transfers to T. Shifts wereonly counted if they were at least four Sr:Ca samplepoints (40 �m) past the year one marker.
A generalized linear mixed model was employed toexamine changes in rate of inter-habitat shifting with ageand changes in habitat occupancy with age. Individualeels were treated as the random eVect because measure-ments within individuals were not independent. All mod-els were Wt in the R 2.1.1 (R Development Core Team2004) statistical program using the GLMM routine(Bates and Sarkar 2005).
Results
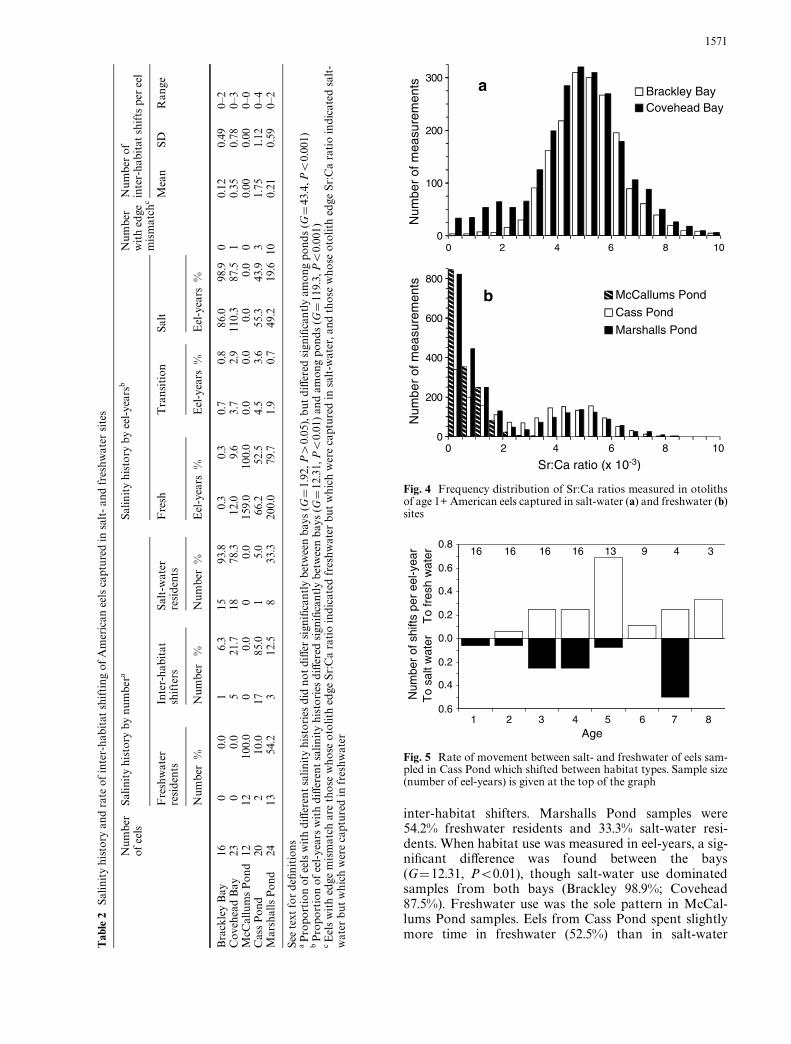
We caught eels at all Wshing locations except McMillansPond bed (Table 1). We encountered excessive lethalbycatch of white perch (Morone americana) in ParsonsPond and stopped Wshing before obtaining the targetminimum sample size (12). Otoliths from 95 eels cap-tured in Brackley and Covehead Bays and McCallums,Cass, and Marshalls Ponds were analyzed for Sr:Caratios. Sr:Ca ratios peaked during the oceanic phase, fellas eels arrived in continental waters, and then showed avariety of patterns (Fig. 3). Overall frequency distribu-tions of smoothed Sr:Ca ratios in age 1+ eels were bimo-dal in samples from both salt- and freshwater sites(Fig. 4). Modal values were <0.5£10¡3 and 5£10¡3. Thehigh mode dominated salt-water samples and the lowmode dominated freshwater samples.
Some Sr:Ca trajectories of individual eels were stablenear one of the two modes, while others varied widely(Fig. 3). All 12 eels from McCallums Pond showed low
Fig. 1 Brackley and Covehead Bays, Prince Edward Island (a) and associated freshwater impoundments; McCallums Pond (b) and Cass and Marsh-alls Ponds (c)
0 250 500
m
c
CassPond
Damwith fishladder Dam with
run-aroundoutlet
MarshallsPond
250 500
m
0
Dam withverticaloverspill
McCallums Pond
Brackley BayCovehead Bay
Parsons Pond
MarshallsPond
Cass PondMcMillanspond bed
McCallumsPond
Gulf of St.Lawrence0 1 2
km
a
b
63˚10’W46˚25’N
1570
and stable Sr:Ca ratios, with a mean of 0.57£10¡3
(SD=0.61£10¡3). We considered these eels to be fresh-water residents (F), and set the upper boundary forfreshwater occupancy to be the McCallums mean +2 SD(1.8£10¡3). In Brackley Bay, 15 eels showed high andstable Sr:Ca ratios (mean=5.24£10¡3, SD=1.28£10¡3)and one eel showed a variable ratio. We considered the15 Brackley eels with high and stable ratios to be salt-water residents (S), and set the lower boundary for salt-water occupancy to be their mean ratio ¡2 SD(2.7£10¡3). Smoothed ratios between 1.8£10¡3 and2.7£10¡3 were considered to indicate inter-habitat tran-sition (T).
The proportion of age 1+ eels that were freshwaterresidents, salt-water residents, and inter-habitat shiftersdiVered signiWcantly among ponds (G test, G=43.4,P<0.001) but not between bays (G=1.92, P>0.05;
Table 2). Failure to detect diVerences between bays mayhave been due to low statistical power (power=0.14).Salt-water residents dominated samples from bothBrackley (93.8%) and Covehead Bays (78.3%). All eelssampled from McCallums Pond were freshwater resi-dents. Most (85%) eels sampled from Cass Pond were
Fig. 2 Schematic proWles of pond outlets, showing the vertical spill-way at McCallums Pond (a), the concrete salmonid Wshway at CassPond (b), and the low-gradient channel at Marshalls Pond (c)
a
b
c 303 m
5.0m
12.2 m
0.9 m
2.2 m
Table 1 Catch and catch rates of American eels in fyke nets in theBrackley–Covehead system
Site Number of eels caught
Number of gear-days
Number caught per gear-day
Brackley Bay 32 46 0.696Covehead Bay 31 52 0.596McCallums Pond 12 61 0.197McMillans Pond bed 0 44 0.000Cass Pond 20 89 0.225Marshalls Pond 30 50 0.600Parsons Pond 6 5 1.200
Fig. 3 Mean within-year Sr:Ca ratios versus age for American eelsfrom Wve sites. For inter-habitat shifters (eels that moved betweenhabitat types at least once after age 1), lines represent individual eels.For eels that remained resident in either salt or freshwater after age1, symbols represent means, with N indicating the number of eels
01234567
Covehead Bay
Brackley Bay
01234567
N=18
McCallums Pond
01234567
0
0
1
1
2
2
3
3
4
4
5
5
6
6
7
7
Marshalls Pond
Age0
0
0
0
5
5
5
5
1
1
1
1
0
0
0
0
1
1
1
1
5
5
5
5
2
2
2
2
0
0
0
0
N=8
Mea
n S
r:C
a ra
tio (
x10
)-3
Mea
n S
r:C
a ra
tio (x
10)
-3M
ean
Sr:
Ca
ratio
(x1
0)
-3M
ean
Sr:C
a ra
tio (
x10
)-3
Mea
n S
r:C
a ra
tio (
x10
)-3
1571
inter-habitat shifters. Marshalls Pond samples were54.2% freshwater residents and 33.3% salt-water resi-dents. When habitat use was measured in eel-years, a sig-niWcant diVerence was found between the bays(G=12.31, P<0.01), though salt-water use dominatedsamples from both bays (Brackley 98.9%; Covehead87.5%). Freshwater use was the sole pattern in McCal-lums Pond samples. Eels from Cass Pond spent slightlymore time in freshwater (52.5%) than in salt-water
Tab
le2
Salin
ity
hist
ory
and
rate
of i
nter
-hab
itat
shi
ftin
g of
Am
eric
an e
els
capt
ured
in s
alt-
and
fres
hwat
er s
ites
See
text
for
deW
niti
ons
a P
ropo
rtio
n of
eel
s w
ith
diV
eren
t sal
init
y hi
stor
ies
did
not d
iVer
sig
niW
cant
ly b
etw
een
bays
(G
=1.
92, P
>0.
05),
but d
iVer
ed s
igniW
cant
ly a
mon
g po
nds
(G=
43.4
, P<
0.00
1)b
Pro
port
ion
of e
el-y
ears
wit
h diV
eren
t sa
linit
y hi
stor
ies
diV
ered
sig
niW
cant
ly b
etw
een
bays
(G=
12.3
1, P
<0.
01)
and
amon
g po
nds
(G=
119.
3, P
<0.
001)
c E
els
wit
h ed
ge m
ism
atch
are
tho
se w
hose
oto
lith
edge
Sr:
Ca
rati
o in
dica
ted
fres
hwat
er b
ut w
hich
wer
e ca
ptur
ed in
sal
t-w
ater
, and
tho
se w
hose
oto
lith
edge
Sr:
Ca
rati
o in
dica
ted
salt
-w
ater
but
whi
ch w
ere
capt
ured
in f
resh
wat
er
Num
ber
of e
els
Salin
ity
hist
ory
by n
umbe
raSa
linit
y hi
stor
y by
eel
-yea
rsb
Num
ber
wit
h ed
ge
mis
mat
chc
Num
ber
of
inte
r-ha
bita
t shi
fts
per
eel
Fre
shw
ater
re
side
nts
Inte
r-ha
bita
t sh
ifte
rsSa
lt-w
ater
re
side
nts
Fre
shT
rans
itio
nSa
ltM
ean
SDR
ange
Num
ber
%N
umbe
r%
Num
ber
%E
el-y
ears
%E
el-y
ears
%E
el-y
ears
%
Bra
ckle
y B
ay16
00.
01
6.3
1593
.80.
30.
30.
70.
886
.098
.90
0.12
0.49
0–2
Cov
ehea
d B
ay23
00.
05
21.7
1878
.312
.09.
63.
72.
911
0.3
87.5
10.
350.
780–
3M
cCal
lum
s P
ond
1212
100.
00
0.0
00.
015
9.0
100.
00.
00.
00.
00.
00
0.00
0.00
0–0
Cas
s P
ond
202
10.0
1785
.01
5.0
66.2
52.5
4.5
3.6
55.3
43.9
31.
751.
120–
4M
arsh
alls
Pon
d24
1354
.23
12.5
833
.320
0.0
79.7
1.9
0.7
49.2
19.6
100.
210.
590–
2
Fig. 4 Frequency distribution of Sr:Ca ratios measured in otolithsof age 1+ American eels captured in salt-water (a) and freshwater (b)sites
0
100
200
300
0 2 4 6 8 10
Brackley BayCovehead Bay
0
200
400
600
800
0 2 4 6 8 10
Sr:Ca ratio (x 10 )-3
McCallums Pond
Cass Pond
Marshalls Pond
Num
ber
of m
easu
rem
ents
Num
ber
of m
easu
rem
ents
b
a
Fig. 5 Rate of movement between salt- and freshwater of eels sam-pled in Cass Pond which shifted between habitat types. Sample size(number of eel-years) is given at the top of the graph
0.6
0.4
0.2
0.0
0.2
0.4
0.6
0.8
1
16 1616
432
16
5
13
6
9
7
4
8
3
Age
Num
ber
of s
hifts
per
eel
-yea
rT
o sa
lt w
ater
To
fres
h w
ater
1572
(43.9%) and Marshalls Pond eels spent the majority(79.7%) of their time in freshwater. Proportion of timespent in the various habitats diVered signiWcantly amongponds (G=119.3, P<0.001).
Eels from all sites except McCallums Pond showedinter-habitat shifts (Table 2). Mean rate of shifting didnot diVer between samples in the bays (Brackley 0.13shifts per eel, Covehead, 0.35 shifts per eel; ANOVA,F=1.02, P>0.05). Rate of shifting diVered among pondsamples (ANOVA, F=28.2, P<0.0001), with highestrates found in Cass Pond (1.75 shifts per eel).
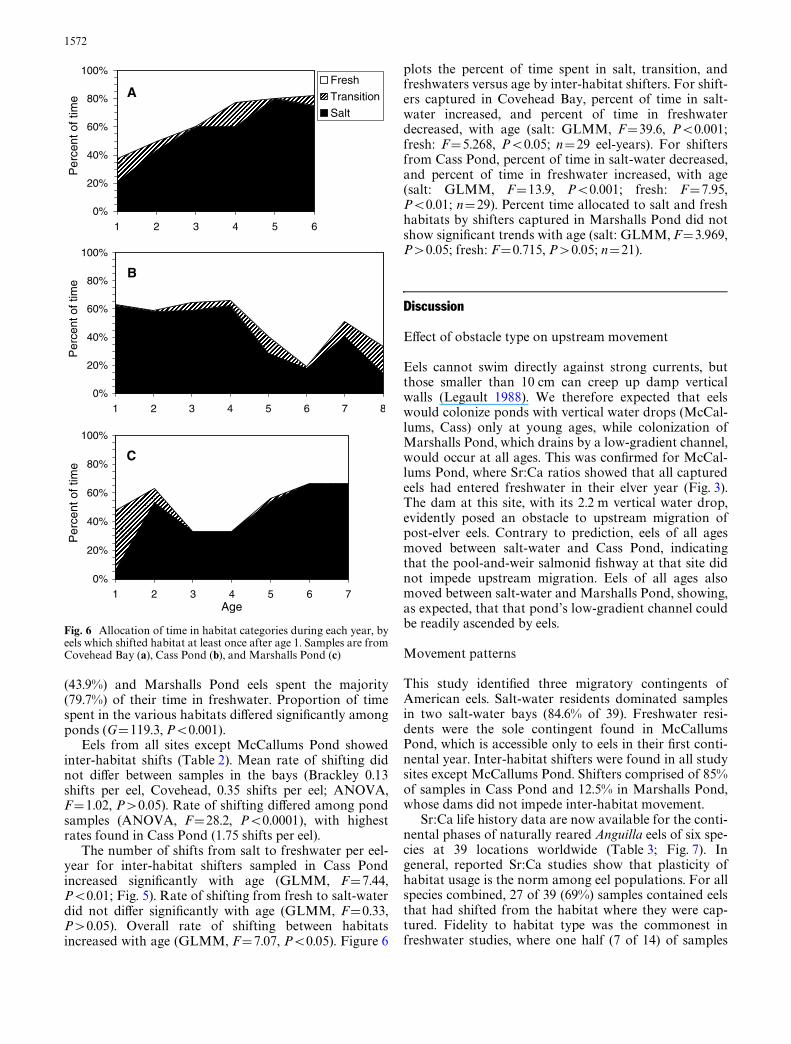
The number of shifts from salt to freshwater per eel-year for inter-habitat shifters sampled in Cass Pondincreased signiWcantly with age (GLMM, F=7.44,P<0.01; Fig. 5). Rate of shifting from fresh to salt-waterdid not diVer signiWcantly with age (GLMM, F=0.33,P>0.05). Overall rate of shifting between habitatsincreased with age (GLMM, F=7.07, P<0.05). Figure 6
plots the percent of time spent in salt, transition, andfreshwaters versus age by inter-habitat shifters. For shift-ers captured in Covehead Bay, percent of time in salt-water increased, and percent of time in freshwaterdecreased, with age (salt: GLMM, F=39.6, P<0.001;fresh: F=5.268, P<0.05; n=29 eel-years). For shiftersfrom Cass Pond, percent of time in salt-water decreased,and percent of time in freshwater increased, with age(salt: GLMM, F=13.9, P<0.001; fresh: F=7.95,P<0.01; n=29). Percent time allocated to salt and freshhabitats by shifters captured in Marshalls Pond did notshow signiWcant trends with age (salt: GLMM, F=3.969,P>0.05; fresh: F=0.715, P>0.05; n=21).
Discussion
EVect of obstacle type on upstream movement
Eels cannot swim directly against strong currents, butthose smaller than 10 cm can creep up damp verticalwalls (Legault 1988). We therefore expected that eelswould colonize ponds with vertical water drops (McCal-lums, Cass) only at young ages, while colonization ofMarshalls Pond, which drains by a low-gradient channel,would occur at all ages. This was conWrmed for McCal-lums Pond, where Sr:Ca ratios showed that all capturedeels had entered freshwater in their elver year (Fig. 3).The dam at this site, with its 2.2 m vertical water drop,evidently posed an obstacle to upstream migration ofpost-elver eels. Contrary to prediction, eels of all agesmoved between salt-water and Cass Pond, indicatingthat the pool-and-weir salmonid Wshway at that site didnot impede upstream migration. Eels of all ages alsomoved between salt-water and Marshalls Pond, showing,as expected, that that pond’s low-gradient channel couldbe readily ascended by eels.
Movement patterns
This study identiWed three migratory contingents ofAmerican eels. Salt-water residents dominated samplesin two salt-water bays (84.6% of 39). Freshwater resi-dents were the sole contingent found in McCallumsPond, which is accessible only to eels in their Wrst conti-nental year. Inter-habitat shifters were found in all studysites except McCallums Pond. Shifters comprised of 85%of samples in Cass Pond and 12.5% in Marshalls Pond,whose dams did not impede inter-habitat movement.
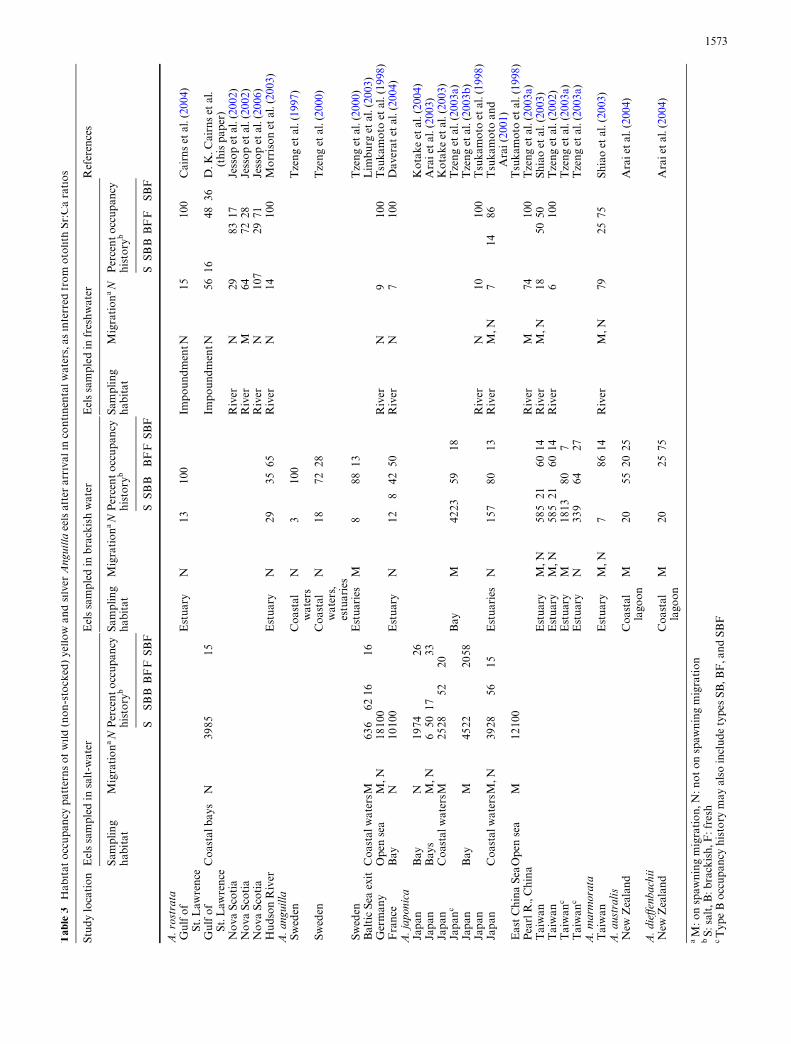
Sr:Ca life history data are now available for the conti-nental phases of naturally reared Anguilla eels of six spe-cies at 39 locations worldwide (Table 3; Fig. 7). Ingeneral, reported Sr:Ca studies show that plasticity ofhabitat usage is the norm among eel populations. For allspecies combined, 27 of 39 (69%) samples contained eelsthat had shifted from the habitat where they were cap-tured. Fidelity to habitat type was the commonest infreshwater studies, where one half (7 of 14) of samples
Fig. 6 Allocation of time in habitat categories during each year, byeels which shifted habitat at least once after age 1. Samples are fromCovehead Bay (a), Cass Pond (b), and Marshalls Pond (c)
0%
20%
40%
60%
80%
100%
1 2 3 4 5 6 7 8
Per
cent
of t
ime
B
0%
20%
40%
60%
80%
100%
1 2 3 4 5 6
Per
cent
of t
ime
Fresh
Transition
Salt
A
0%
20%
40%
60%
80%
100%
1 2 3 4 5 6 7Age
Per
cent
of t
ime
C
1573
Tab
le3
Hab
itat
occ
upan
cy p
atte
rns
of w
ild (n
on-s
tock
ed)
yello
w a
nd s
ilver
Ang
uilla
eel
s af
ter
arri
val i
n co
ntin
enta
l wat
ers,
as
infe
rred
fro
m o
tolit
h Sr
:Ca
rati
os
a M
: on
spaw
ning
mig
rati
on, N
: not
on
spaw
ning
mig
rati
onb
S: s
alt,
B: b
rack
ish,
F: f
resh
c T
ype
B o
ccup
ancy
his
tory
may
als
o in
clud
e ty
pes
SB, B
F, a
nd S
BF
Stud
y lo
cati
onE
els
sam
pled
in s
alt-
wat
erE
els
sam
pled
in b
rack
ish
wat
erE
els
sam
pled
in fr
eshw
ater
Ref
eren
ces
Sam
plin
g ha
bita
tM
igra
tion
aN
Per
cent
occ
upan
cy
hist
oryb
Sam
plin
g ha
bita
tM
igra
tion
aN
Per
cent
occ
upan
cy
hist
oryb
Sam
plin
g ha
bita
tM
igra
tion
aN
Per
cent
occ
upan
cy
hist
oryb
SSB
BB
FF
SBF
SSB
BB
FF
SBF
SSB
BB
FF
SBF
A. r
ostr
ata
Gul
f of
St
. Law
renc
eE
stua
ryN
1310
0Im
poun
dmen
tN15
100
Cai
rns
etal
. (20
04)
Gul
f of
St
. Law
renc
eC
oast
al b
ays
N39
8515
Impo
undm
entN
5616
4836
D. K
. Cai
rns
etal
. (t
his
pape
r)N
ova
Scot
iaR
iver
N29
8317
Jess
op e
tal
. (20
02)
Nov
a Sc
otia
Riv
erM
6472
28Je
ssop
et
al. (
2002
)N
ova
Scot
iaR
iver
N10
729
71Je
ssop
et
al. (
2006
)H
udso
n R
iver
Est
uary
N29
3565
Riv
erN
1410
0M
orri
son
etal
. (20
03)
A. a
ngui
llaSw
eden
Coa
stal
w
ater
sN
310
0T
zeng
et
al. (
1997
)
Swed
enC
oast
al
wat
ers,
es
tuar
iesN
1872
28T
zeng
et
al. (
2000
)
Swed
enE
stua
ries
M8
8813
Tze
ng e
tal
. (20
00)
Bal
tic
Sea
exit
Coa
stal
wat
ersM
636
6216
16L
imbu
rg e
tal
. (20
03)
Ger
man
yO
pen
sea
M, N
1810
0R
iver
N9
100
Tsu
kam
oto
etal
. (19
98)
Fra
nce
Bay
N10
100
Est
uary
N12
842
50R
iver
N7
100
Dav
erat
et
al. (
2004
)A
. jap
onic
aJa
pan
Bay
N19
7426
Kot
ake
etal
. (20
04)
Japa
nB
ays
M, N
650
1733
Ara
i eta
l. (2
003)
Japa
nC
oast
al w
ater
sM25
2852
20K
otak
e et
al. (
2003
)Ja
panc
Bay
M42
2359
18T
zeng
et
al. (
2003
a)Ja
pan
Bay
M45
2220
58T
zeng
et
al. (
2003
b)Ja
pan
Riv
erN
1010
0T
suka
mot
o et
al. (
1998
)Ja
pan
Coa
stal
wat
ersM
, N39
2856
15E
stua
ries
N15
780
13R
iver
M, N
714
86T
suka
mot
o an
d A
rai (
2001
)E
ast
Chi
na S
eaO
pen
sea
M12
100
Tsu
kam
oto
etal
. (19
98)
Pea
rl R
., C
hina
Riv
erM
7410
0T
zeng
et
al. (
2003
a)T
aiw
anE
stua
ryM
, N58
521
6014
Riv
erM
, N18
5050
Shia
o et
al. (
2003
)T
aiw
anE
stua
ryM
, N58
521
6014
Riv
er6
100
Tze
ng e
tal
. (20
02)
Tai
wan
cE
stua
ryM
1813
807
Tze
ng e
tal
. (20
03a)
Tai
wan
cE
stua
ryN
339
6427
Tze
ng e
tal
. (20
03a)
A. m
arm
orat
aT
aiw
anE
stua
ryM
, N7
8614
Riv
erM
, N79
2575
Shia
o et
al. (
2003
)A
. aus
tral
isN
ew Z
eala
ndC
oast
al
lago
onM
2055
2025
Ara
i eta
l. (2
004)
A. d
ieV
enba
chii
New
Zea
land
Coa
stal
la
goon
M20
2575
Ara
i eta
l. (2
004)

1574
showed freshwater residency only. The exclusivity of thiscontingent at many locations may be due to long dis-tances to other salinity zones (Hudson River, USA: Mor-rison et al. 2003; Pearl River, China: Tzeng et al. 2003a),or to dams which impede upstream movements of eels atpost-elver ages (Cairns et al. 2004; D. K. Cairns et al.,this paper). Most samples of eels from brackish and salt-water contained eels that had used other habitats (brack-ish: 13 of 15, 87%; salt: 7 of 10, 70%). Movement patternsshown by American eels in the Brackley–Covehead sys-tem reXect the highly plasticity of Anguillid habitat useworldwide. Although available Sr:Ca studies are suY-cient to portray the broad strokes of Anguilla movementpatterns, it must be emphasized that uncertainty aboutrepresentativity of study sites and limited sample sizespreclude robust statistical conclusions.
Sr:Ca studies have demonstrated that some Europeanand Japanese eels never leave salt-water (Tsukamotoet al. 1998; Arai et al. 2003). This study is the Wrst to mea-sure Sr:Ca ratios of American eels sampled in salt-water.Our Wnding that most eels from Brackley and CoveheadBays had an exclusive salt-water Sr:Ca proWle indicatesthat American eels can likewise complete their life cyclein the sea. The catadromy paradigm for the American eelis thus overturned. Like European and Japanese eels,American eels must now be considered as species where
catadromy is a facultative life history option. The highrepresentation of exclusive salt-water residency in eelsfrom bay samples (84.6%) also suggests that non-catadr-omy may be an important and common pattern forAmerican eels.
Tosi et al. (1988) and Édeline and Élie (2004) foundthat European glass eels have distinct individual salinitypreferences. This implies that young eels separate intomigratory contingents upon arrival on the coast, withsalt-seeking eels remaining in marine waters while fresh-seekers ascend into freshwaters. The freshwater ponds inthe Brackley–Covehead system are adjacent to salt-waterbays, and all are within 5 km of the open sea. Given asalinity preference that is activated on arrival in coastalwaters and an upstream ascent rate of 0.6 km/day (Whiteand Knights 1997), we hypothesized that there would belittle time lag between the colonization of salt-water byeels destined to be salt residents, and colonization offreshwaters by eels destined to be fresh residents. Consis-tent with expectation, salt- and freshwater residents wereestablished in their respective habitats by age 1 (Fig. 3).Early movement to settlement areas may have an adap-tive basis. Because of their superior climbing ability,young eels can overcome vertical barriers to upstreammigration (such as the dam at McCallums Pond) andreach habitats that are inaccessible at older ages.
Fig. 7 Percentage of sampled Anguilla eels which showed movements out of the habitat type where they were captured, based on Sr:Ca anal-ysis. The habitat type given in each panel is the habitat where captured. All species includes A. marmorata, A. australis, and A. dieVenbachii.See Table 3 for data sources
Percent of eels which showed movement out of the habitat type where they were captured
A. rostrata,salt water
0
1
0 1-33 34-66 >67
Num
ber
ofst
udie
s A.rostrata,brackish
water
0
1
0 1-33 34-66 >67
A. rostrata,fresh water
0
1
2
0 1-33 34-66 >67
A. rostrata,all habitats
0
1
2
3
0 1-33 34-66 >67
A. anguilla,salt water
0
1
2
0 1-33 34-66 >67
Num
ber
ofst
udie
s
A. anguilla,brackish
water
0
1
2
0 1-33 34-66 >67
A. anguilla,fresh water
0
1
2
0 1-33 34-66 >67
A. anguilla,all habitats
0
1
2
3
4
5
0 1-33 34-66 >67
A. japonica,salt water
0
1
2
3
0 1-33 34-66 >67
Num
ber
ofst
udie
s A.japonica,brackish
water
0
1
2
0 1-33 34-66 >67
A. japonica,fresh water
0
1
2
3
0 1-33 34-66 >67
A. japonica, all habitats
0
1
2
3
4
5
0 1-33 34-66 >67
All
speciesspecies,
,salt
water
0
1
2
3
4
0 1-33 34-66 >67
Num
ber
ofst
udie
s AllAll species,
brackishwater
0
1
2
3
4
5
0 1-33 34-66 >67
fresh water
0
2
4
6
0 1-33 34-66 >67
All species,all habitats
0
246
81012
0 1-33 34-66 >67

1575
Eels may choose among salinity zones, and they mayalso choose between sedentary and mobile lifestyles(Feunteun et al. 2003). Some eels in the Brackley–Cove-head system shifted between habitats only once and thenremained in the new habitat. This could be interpreted assearching for suitable habitat, and then settling therewhen they Wnd it (Fig. 3). However, others shifted contin-uously, suggesting that nomadic behavior is an inherentproperty of some eels.
Implications for conservation
Prince Edward Island has over 800 artiWcial impound-ments (MacFarlane 1999), and most streams areblocked by one or several dams. Our results suggestthat eels can readily reach ponds formed by damsequipped with pool-and-weir salmonid Wshways or low-gradient channels. These passage facilities thus permitthe natural spatial behavior that is characteristic of thehabitat-shifting eel contingent. In contrast, small damswith vertical spillways and no passage facilities mayblock the normal behavior of habitat-shifting eels bylimiting upstream passage to young ages. This mayhave negative eVects on populations in systems wheresuch dams occur.
Recent declines in eel abundance indices haveprompted eVorts to devise management schemes thatwould assure adequate escapement to the spawninggrounds (Richkus and Whalen 2000; ICES 2003). Suchschemes must recognize contributions of escaping silvereels from unWshed, as well as Wshed areas. In much ofeastern North America eel exploitation is restricted tocoastal and estuarine waters. Our results suggest thatWshing in marine waters may aVect eel populations innearby freshwaters, due to movement between the twohabitats. Neither marine populations nor those of adja-cent freshwater are discrete. Population models that esti-mate escapement of silver eels from salt, brackish, andfreshwater habitats must account for these movements.
The relative importance of marine and freshwaters tothe eel’s life cycle, and patterns of migration betweenthese habitats, are vital issues in eel biology and conser-vation. The Sr:Ca technique will Wnd wide employ as thekey to their understanding.
Acknowledgments This study received support from the NationalScience Council, ROC (NSC 91¡2313-B-002-291 and 92-2313-B-002-057). We thank Angus McLennan, Corey Muttart, ValérieTremblay, Noella McDonald, and Robbie Moore for assistance inthe laboratory and Weld, and Mark Grimmett for measuring chemi-cal concentrations in water samples. Tillmann Benfey and AllenCurry provided valuable advice at all stages and improved the man-uscript with their comments.
References
Arai T, Kotake A, Ohji M, Miller MJ (2003) Occurrence of sea eelsof Anguilla japonica along the Sanriku Coast of Japan. IchthyolRes 50:78–81
Arai T, Kotake A, Lokman PM, Miller MJ, Tsukamoto K (2004)Evidence of diVerent habitat use by New Zealand freshwater eelsAnguilla australis and A. dieVenbachii, as revealed by otolithmicrochemistry. Mar Ecol Prog Ser 266:213-225
Bates D, Sarkar D (2005) Linear mixed eVects models using S4 mod-els. LME 4 package for R. Available at http://www.micro-arrays.unife.it/CRAN/doc/packages/lme4.pdf
Cairns DK, Shiao JC, Iizuka Y, Tzeng WN, MacPherson CD (2004)Movement patterns of American eels in an impounded water-course, as indicated by otolith microchemistry. N Am J FishManage 24:452–458
Daverat F, Élie P, LaHaye M (2004) Première caractérisationdes histoires de vie des anguilles (Anguilla anguilla) occupantla zone aval du bassin versant Gironde-Garonne-Dordogne:apport d’une méthode de microchimie. Cybium 28(Suppl1):83-90
Édeline É, Élie P (2004) Is salinity choice related to growth in juve-nile eel Anguilla anguilla? Cybium 28(Suppl 1):77-82
Feunteun E, LaVaille P, Robinet T, Briand C, Baisez A, Olivier J-M,Acou A (2003) A review of upstream migration and movementsin inland waters by anguillid eels: towards a general theory. In:Aida K, Tsukamoto K, Yamauchi K (eds) Eel biology. Springer,Berlin Heidelberg New York, pp 191-213
ICES (2003) Report of the thirteenth session of the joint EIFAC/ICES working group on eels. EIFAC occasional paper no. 36
Jessop BM, Shiao JC, Iizuka Y, Tzeng WN (2002) Migratorybehaviour and habitat use by American eels Anguilla rostrataas revealed by otolith microchemistry. Mar Ecol Prog Ser233:217–229
Jessop BM, Shiao JC, Iizuka Y, Tzeng WN (2006) Migration betweenfreshwater and estuary of juvenile American eels Anguilla rostrataas revealed by otolith microchemistry. Mar Ecol Prog Ser (in press)
Kotake A, Arai T, Ozawa T, Nojima S, Miller MJ, Tsukamoto K(2003) Variation in migratory history of Japanese eels, An-guilla japonica, collected in coastal waters of the Amakusa Is-lands, Japan, inferred from otolith Sr/Ca ratios. Mar Biol142:849-854
Kotake A, Arai T, Ohji M, Yamane S, Miyazaki N, Tsukamoto K(2004) Application of otolith microchemistry to estimate themigratory history of Japanese eel Anguilla japonica on the San-riku coast of Japan. J Appl Icthyol 20:150–153
Legault A (1988) Le franchissement des barrages par l’escalade del’anguille: étude en Sèvre Niortaise. Bull Fr Pêche Piscic 308:1–10
Limburg KE, Wickstrom H, Svedang H, Elfman M, Kristiansson P(2003) Do stocked freshwater eels migrate? Evidence from theBaltic suggests “yes”. Am Fish Soc Symp 33:275-284
MacFarlane RE (1999) An evaluation of the potential impacts ofsome Prince Edward Island impoundments on salmonid habitat.MSc Thesis, Acadia University, Wolfville, Nova Scotia
Morrison WE, Secor DH, Piccoli PM (2003) Estuarine habitat useby Hudson River American eels as determined by otolith stron-tium:calcium ratios. Am Fish Soc Symp 33:87-99
R Development Core Team (2004) R: a language and environmentfor statistical computing. R Foundation for Statistical Comput-ing, Vienna, Austria. ISBN 3-900051-00-3. Available at http://www.R-project.org
Richkus WA, Whalen K (2000) Evidence for a decline in the abun-dance of the American eel, Anguilla rostrata (LeSueur), in NorthAmerica since the early 1980s. Dana 12:83–97
Secor DH (1999) Specifying divergent migrations in the concept ofstock: the contingent hypothesis. Fish Res 43:13–34
Shiao JC, Iizuka Y, Chang CW, Tzeng WN (2003) Disparities inhabitat use and migratory behavior between tropical eel Anguillamarmorata and temperate eel A. japonica in four Taiwanese riv-ers. Mar Ecol Prog Ser 261:233–242
Tosi L, Sala L, Sola C, Spampanato A, Tongiorgi P (1988) Experi-mental analysis of the thermal and salinity preferences of glass-eels, Anguilla anguilla (L), before and during the upstream migra-tion. J Fish Biol 33:721-733
Tsukamoto K, Arai T (2001) Facultative catadromy of the eel An-guilla japonica between fresh water and seawater habitats. MarEcol Prog Ser 220:265–276

1576
Tsukamoto K, Nakai I, Tesch WV (1998) Do all freshwater eels mi-grate? Nature 396:635-636
Tzeng WN, Severin KP, Wickstrom H (1997) Use of otolith micro-chemistry to investigate the environmental history of Europeaneel Anguilla anguilla. Mar Ecol Prog Ser 149:73–81
Tzeng WN, Wang CH, Wickstrom H, Reizenstein M (2000) Occur-rence of the semi-catadromous European eel Anguilla anguilla inthe Baltic Sea. Mar Biol 137:93–98
Tzeng WN, Shiao JC, Iizuka Y (2002) Use of otolith Sr:Ca ratios tostudy the riverine migratory behaviors of Japanese eel Anguillajaponica. Mar Ecol Prog Ser 245:213-221
Tzeng WN, Iizuka Y, Shiao JC, Yamada Y, Oka HP (2003a) Identi-Wcation and growth rates comparison of divergent migratorycontingents of Japanese eel (Anguilla japonica). Aquaculture216:77-86
Tzeng WN, Shiao JC, Yamada Y, Oka HP (2003b) Life history pat-terns of Japanese eel Anguilla japonica in Mikawa Bay, Japan.Am Fish Soc Symp 33:285-293
White EM, Knights B (1997) Dynamics of upstream migration ofthe European eel, Anguilla anguilla (L), in the Rivers Severn andAvon, England, with special reference to the eVects of man-madebarriers. Fish Manag Ecol 4:311-324

Mar Biol (2006) 149:1587–1588 DOI 10.1007/s00227-006-0418-x
123
ERRATUM
Movement patterns of American eels (Anguilla rostrata) between salt- and freshwater in a coastal watershed, based on otolith microchemistry
Heather M. Lamson · Jen-Chieh Shiao · Yoshiyuki Iizuka · Wann-Nian Tzeng · David K. Cairns
Published online: 26 June 2006© Springer-Verlag 2006
Mar Biol (2006) DOI 10.1007/s00227-006-0308-2
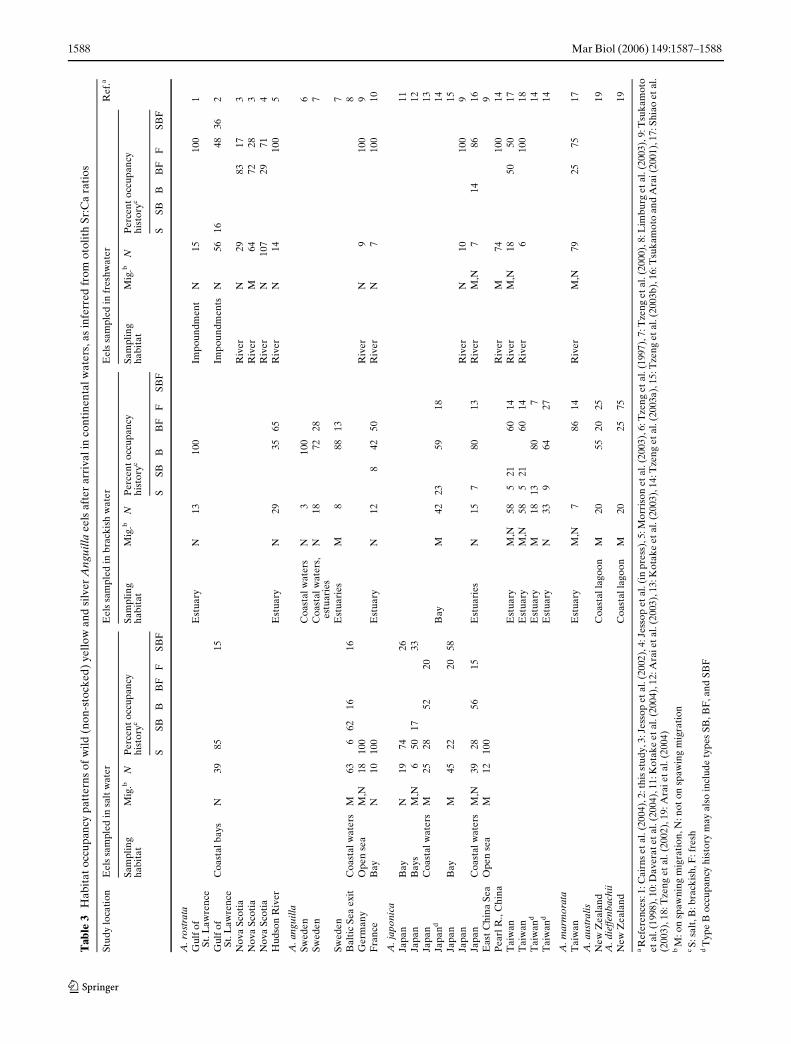
Unfortunately Table 3 contained errors. The correctTable 3 is shown below.
The online version of the original article can be found at http://dx.doi.org/10.1007/s00227-006-0308-2.
H. M. LamsonBiology Department, University of New Brunswick, Fredericton, NB, Canada E3B 6E1
J.-C. ShiaoInstitute of Zoology, Academia Sinica, Nankang, Taipei 11529, Taiwan, ROC
Y. IizukaInstitute of Earth Sciences, Academia Sinica, Nankang, Taipei 11529, Taiwan, ROC
W.-N. TzengInstitute of Fisheries Sciences, National Taiwan University, Taipei 10617, Taiwan, ROC
D. K. Cairns (&)Department of Fisheries and Oceans, Box 1236, Charlottetown, PE, Canada C1A 7M8 e-mail: [email protected]
1588 Mar Biol (2006) 149:1587–1588
123
Tab
le3
Hab
itat
occ
upan
cy p
atte
rns
of w
ild (
non-
stoc
ked)
yel
low
and
silv
er A
ngui
lla e
els
afte
r ar
riva
l in
cont
inen
tal w
ater
s, a
s in
ferr
ed f
rom
oto
lith
Sr:C
a ra
tios
a R
efer
ence
s: 1
: Cai
rns
etal
. (20
04),
2: t
his
stud
y, 3
: Jes
sop
etal
. (20
02),
4: J
esso
p et
al. (
in p
ress
), 5
: Mor
riso
n et
al. (
2003
), 6
: Tze
ng e
tal
. (19
97),
7: T
zeng
eta
l. (2
000)
, 8: L
imbu
rg e
tal
. (20
03),
9: T
suka
mot
oet
al. (
1998
), 1
0: D
aver
at e
tal
. (20
04),
11:
Kot
ake
etal
. (20
04),
12:
Ara
i eta
l. (2
003)
, 13:
Kot
ake
etal
. (20
03),
14:
Tze
ng e
tal
. (20
03a)
, 15:
Tze
ng e
tal
. (20
03b)
, 16:
Tsu
kam
oto
and
Ara
i (20
01),
17:
Shi
ao e
tal
.(2
003)
, 18:
Tze
ng e
tal
. (20
02),
19:
Ara
i eta
l. (2
004)
b M
: on
spaw
ning
mig
rati
on, N
: not
on
spaw
ing
mig
rati
onc S:
sal
t, B
: bra
ckis
h, F
: fre
shd
Typ
e B
occ
upan
cy h
isto
ry m
ay a
lso
incl
ude
type
s SB
, BF
, and
SB
F
Stud
y lo
cati
onE
els
sam
pled
in s
alt w
ater
Eel
s sa
mpl
ed in
bra
ckis
h w
ater
Eel
s sa
mpl
ed in
fre
shw
ater
Ref
.a
Sam
plin
g ha
bita
tM
ig.b
NP
erce
nt o
ccup
ancy
hi
stor
ycSa
mpl
ing
habi
tat
Mig
.bN
Per
cent
occ
upan
cy
hist
oryc
Sam
plin
g ha
bita
tM
ig.b
NP
erce
nt o
ccup
ancy
hi
stor
yc
SSB
BB
FF
SBF
SSB
BB
FF
SBF
SSB
BB
FF
SBF
A. r
ostr
ata
Gul
f of
St. L
awre
nce
Est
uary
N13
100
Impo
undm
ent
N15
100
1
Gul
f of
St. L
awre
nce
Coa
stal
bay
sN
3985
15Im
poun
dmen
tsN
5616
4836
2
Nov
a Sc
otia
Riv
erN
2983
173
Nov
a Sc
otia
Riv
erM
6472
283
Nov
a Sc
otia
Riv
erN
107
2971
4H
udso
n R
iver
Est
uary
N29
3565
Riv
erN
1410
05
A. a
ngui
llaSw
eden
Coa
stal
wat
ers
N3
100
6Sw
eden
Coa
stal
wat
ers,
es
tuar
ies
N18
7228
7
Swed
enE
stua
ries
M8
8813
7B
alti
c Se
a ex
itC
oast
al w
ater
sM
636
6216
168
Ger
man
yO
pen
sea
M,N
1810
0R
iver
N9
100
9F
ranc
eB
ayN
1010
0E
stua
ryN
128
4250
Riv
erN
710
010
A. j
apon
ica
Japa
nB
ayN
1974
2611
Japa
nB
ays
M,N
650
1733
12Ja
pan
Coa
stal
wat
ers
M25
2852
2013
Japa
ndB
ayM
4223
5918
14Ja
pan
Bay
M45
2220
5815
Japa
nR
iver
N10
100
9Ja
pan
Coa
stal
wat
ers
M,N
3928
5615
Est
uari
esN
157
8013
Riv
erM
,N7
1486
16E
ast
Chi
na S
eaO
pen
sea
M12
100
9P
earl
R.,
Chi
naR
iver
M74
100
14T
aiw
anE
stua
ryM
,N58
521
6014
Riv
erM
,N18
5050
17T
aiw
anE
stua
ryM
,N58
521
6014
Riv
er6
100
18T
aiw
and
Est
uary
M18
1380
714
Tai
wan
dE
stua
ryN
339
6427
14
A. m
arm
orat
aT
aiw
anE
stua
ryM
,N7
8614
Riv
erM
,N79
2575
17A
. aus
tral
isN
ew Z
eala
ndC
oast
al la
goon
M20
5520
2519
A. d
ieV
enba
chii
New
Zea
land
Coa
stal
lago
onM
2025
7519
Copyright © 2022 FDOKUMEN
















![Anguilla anguilla L. Biochemical and Genotoxic Responses to Benzo[ a]pyrene](https://static.fdokumen.com/doc/165x107/631d4597f26ecf94330a787a/anguilla-anguilla-l-biochemical-and-genotoxic-responses-to-benzo-apyrene.jpg)



