
Chemical communication in the genus Anguilla: a minireview
19
Chemical communication in the Genus Anguilla: a minireview Mar Huertas 1) , Adelino V.M. Canário & Peter C. Hubbard (Centro de Ciências do Mar, Universidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal) (Accepted: 15 July 2008) Summary Eels of the Genus Anguilla are important fish in both scientific and economic terms. Unfor- tunately, current stocks of the European eel in particular (Anguilla anguilla L.) are becoming increasingly endangered. Chemical communication plays important roles in several key as- pects of fish biology. Due to its complex life-cycle, the eel offers a unique opportunity to study various aspects of olfactory biology and chemical communication almost in isolation, feeding (during freshwater ‘yellow’ stage), adaptation to seawater, migration and, finally, reproduction. The aim of this brief review is to summarise what is known about chemical communication in the eel (during reproduction in particular) and, more importantly, stim- ulate further research. To date, eels have been shown to have high olfactory sensitivity to substances released by conspecifics. Both the bile fluid and skin mucus have been implicated as the routes of release of these odorants but other routes, such as the urine, cannot be ex- cluded. Furthermore, the nature of these odorants depends on both the sex and reproductive status of the donor; exposure to water conditioned by mature con-specifics causes a stim- ulation of sexual maturation in immature eels. These results are consistent with a role for chemical communication in eel reproduction. The European eel is, therefore, proposed as a model species for studies on olfaction and chemical communication in fish because of its ancestral status, its high olfactory sensitivity and its physiological plasticity. However, more research is necessary; specifically, identification of the key odorants involved, their behav- ioural and physiological effects and how the olfactory system may change at various stages of its life-cycle. Keywords: eel, chemical communication, pheromone, reproduction, olfaction. 1) Corresponding author’s e-mail address: [email protected] © Koninklijke Brill NV, Leiden, 2008 Behaviour 145, 1389-1407 Also available online - www.brill.nl/beh
Transcript of Chemical communication in the genus Anguilla: a minireview

Chemical communication in the Genus Anguillaa minireview
Mar Huertas1) Adelino VM Canaacuterio amp Peter C Hubbard(Centro de Ciecircncias do Mar Universidade do Algarve Campus de Gambelas
8005-139 Faro Portugal)
(Accepted 15 July 2008)
Summary
Eels of the Genus Anguilla are important fish in both scientific and economic terms Unfor-tunately current stocks of the European eel in particular (Anguilla anguilla L) are becomingincreasingly endangered Chemical communication plays important roles in several key as-pects of fish biology Due to its complex life-cycle the eel offers a unique opportunity tostudy various aspects of olfactory biology and chemical communication almost in isolationfeeding (during freshwater lsquoyellowrsquo stage) adaptation to seawater migration and finallyreproduction The aim of this brief review is to summarise what is known about chemicalcommunication in the eel (during reproduction in particular) and more importantly stim-ulate further research To date eels have been shown to have high olfactory sensitivity tosubstances released by conspecifics Both the bile fluid and skin mucus have been implicatedas the routes of release of these odorants but other routes such as the urine cannot be ex-cluded Furthermore the nature of these odorants depends on both the sex and reproductivestatus of the donor exposure to water conditioned by mature con-specifics causes a stim-ulation of sexual maturation in immature eels These results are consistent with a role forchemical communication in eel reproduction The European eel is therefore proposed as amodel species for studies on olfaction and chemical communication in fish because of itsancestral status its high olfactory sensitivity and its physiological plasticity However moreresearch is necessary specifically identification of the key odorants involved their behav-ioural and physiological effects and how the olfactory system may change at various stagesof its life-cycle
Keywords eel chemical communication pheromone reproduction olfaction
1) Corresponding authorrsquos e-mail address mhuertasualgpt
copy Koninklijke Brill NV Leiden 2008 Behaviour 145 1389-1407Also available online - wwwbrillnlbeh
1390 Huertas Canaacuterio amp Hubbard
Why is the eel interesting
The increasing interest in Anguillid eels is related to several factors Firstlythe genus Anguilla has a complex life-cycle which includes a worldwidedistribution and a catadromous migration that have been subject of severalecological and genetic studies (Aoyama amp Tsukamoto 1997 Westin 1998)Secondly the eel is a highly appreciated commercial species with a welldeveloped aquaculture industry (Peacuterez et al 2004) Finally many eel speciesare endangered in several cases natural stocks have been reduced to levelsbelow those necessary for a viable population (van Ginneken et al 2007)A concerted effort by governments scientists fisheries and the aquacultureindustry is necessary to prevent the extinction of this once common andwide-spread fish
An example of complex life-cycle the European eel
The European eel (Anguilla anguilla L) is one of the 18 species of the genusAnguilla which in turn belongs to the subdivision Elopomorpha a groupthat appeared early in teleost evolution and the order anguilliformes whichalso includes moray eels (Nelson 2006) The amazing life-cycle of Europeaneels (Figure 1) consists of a migration of adults over a distance of 5000 kmacross the Atlantic Ocean to the Sargasso Sea between Bermuda Island andPuerto Rico and the opposite migration of larvae to continental waters (vanGinneken amp Maes 2005) Adult males start to migrate in September andfemales one or two months later (Usui 1991 Tesch 2003) with a swim-ming velocity of 05 body lengths the journey should take between 6 and 7months (van Ginneken et al 2005) During this journey the adults do not eatbut nevertheless undergo sexual maturation After spawning they die (vanGinneken amp Maes 2005) The resulting leaf-like shape larvae called lep-tocephali (Lecompte-Finiger et al 2004) swim in less than one year (vanGinneken et al 2005) to the European coast aided by Gulf-Steam currentsand on reaching the continental shelf metamorphose into glass eels be-fore entering rivers in western Europe and North Africa (Lecompte-Finiger1994 McCleave et al 1998) around November to December Transparentglass eels undergo a series of metamorphic events until they reach the yel-low stage (Elie et al 1982) and after a variable growth period becomesilver eels (Deelder 1984) in a process more linked with puberty than meta-morphosis (Dufour 2000 Aroua et al 2005) Then after a variable period
Chemical communication in eels 1391
Figure 1 Life cycle of the European eel (Anguilla anguilla) Sexually immature lsquoyellowrsquoeels grow in rivers lakes and estuaries for several years (10ndash30) until the lsquosilverrsquo eel stageThen they migrate across the Atlantic Ocean to the Sargasso see where sexually mature fishmate and spawn sexual maturation occurs lsquoen routersquo but feeding ceases The resulting lep-tocephali larvae then travel to the European coast and when they reach the continental plat-form metamorphose into lsquoglassrsquo eels and enter estuaries (between November and February)and make their way upstream to inland freshwaters Olfaction may be involved in any or all
of these stages
(6 to 30 years) silver adults in the first stages of gametogenesis completethe cycle by their migration to the Sargasso Sea (van Ginneken et al 2005)Many aspects of the eelrsquos life-cycle are still unknown such as where exactlythe spawning location is how do they find this location at what depth dothey swim in the Atlantic Ocean how do the two sexes meet (since malesstart migrating before the females) (Haraldstad et al 1985 Usui 1991) andwhen and how does sexual maturation occur Current tracking and teleme-try studies promise to provide some answers to these questions (McCleave ampArnold 1999 Jellyman amp Tsukamoto 2005) However other answers maybe found by laboratory experiments The role of olfaction in these processesis largely unknown The aim of this review is to briefly outline what is knownand suggest directions that future research should take
Eel aquaculture and current situation of natural stocks
Globally anguillid eels are an important and prized food source (Heinsbroek1991 Usui 1991 Gousset 1992) Since the capture of wild stocks is insuffi-
1392 Huertas Canaacuterio amp Hubbard
cient to satisfy the market eel aquaculture has increased exponentially overthe last 30 years Asian countries are the highest producers and consumers(more than 220 000 metric tonnes per year) whereas European productionconstitutes less than 5 of the worldwide total (about 10 000 metric tonnes(Peacuterez et al 2004 ICES 2005) The main problems associated with eelaquaculture are provision and control of appropriate environmental factors(temperature salinity light) the formulation of diets (to reduce mortalityby cannibalism and increase growth) and the artificial induction of sexualreproduction
In captivity the European eel does not reach sexual maturity without hor-monal manipulation only the Japanese eel Anguilla japonica has been suc-cessfully induced to spawn under artificial conditions (Tanaka et al 2003)Even then mass production of high quality glass eels has not yet becomeroutine (Furuita et al 2006) To induce sexual maturation in Japanese andEuropean eels injections with human chorionic gonadotropin (hCG) in thecase of males or pituitary extract in the case of females are necessary(Ohta et al 1996a 1997 Hao-ran et al 1998 Ijiri et al 1998 Asturianoet al 2002) After pituitary extract treatment females usually require treat-ment with the maturation-inducing steroid 1720β-dihydroxy-4-pregnen-3-one (1720βP) to induce ovulation (Ohta et al 1996b Kagawa et al 1997)
The inability to produce glass eels commercially means that the aquacul-ture industry depends on the capture of wild stocks increasing the fishing ef-fort in the eel stocks Thus together the over-exploitation of natural stocksthe change in oceanographic conditions (global warming) degradation andreduction of suitable freshwater habitats pollution and parasitism have re-sulted in an alarming decrease of wild glass and adult eel population (Briandet al 2003 Dekker 2003 Starkie 2003) As a result the current Europeaneel population stands at about 1 of the level of 30 years ago there areno signs of recovery This is of major concern to eel conservation fisheriesaquaculture and industry As a consequence this species will be included inCITES by 2009 and the ICESEIFAC Eel Working Group (September 2007)proposed that a self-sufficient eel aquaculture industry was vital for the re-covery of natural stocks and research focusing on spawner quality and repro-duction was urgently needed to achieve this (van Ginneken et al 2007)
How can studies of olfaction help
Communication is defined as ldquointegration between a lsquosignallerrsquo who pro-duces a sensory stimulus or signal (a chemical in the case of chemical com-
Chemical communication in eels 1393
munication) and a lsquoreceiverrsquo who perceives the signal and makes a conse-quent behavioral decisionrdquo (Rosenthal amp Lobel 2006) Pheromones are sub-stances released by conspecifics which then provoke an innate behaviouralandor physiological response in the receiver (Stacey amp Sorensen 2002)There is little doubt that pheromonal communication plays an important rolein many aspects of fish biology However there is great variability in thetissues of origin of pheromones their routes and regulation of release theirchemical structure and the nature of the receiversrsquo response(s) which en-compass a wide repertoire of behavioural and physiological effects (Staceyamp Sorensen 2002) For example many studies have shown that pheromoneshelp to co-ordinate reproductive events (Stacey amp Sorensen 2002) are in-volved in inter-specific or intra-specific recognition mate-choice (Rosenthalamp Lobel 2006) act as riverine pheromone plumes in species such as the sealamprey (Polkinghorne et al 2001 Li et al 2002) or Arctic char (Selset ampDoving 1980) or play an important role in the alarm reaction (Win 2000Ide et al 2003 Kristensen amp Closs 2004) However most available infor-mation about fish pheromones is derived from freshwater spawning species(eg salmonids and cyprinids) very little is known about species that spawnin sea water
Several aspects of anguillid eel biology together with their highly devel-oped sense of smell (Silver 1982 Crnjar et al 1992 Sola 1993ab Sola ampTongiorgi 1998) suggest that chemical communication could be involved atkey stages of their life-history eg the migration over the Atlantic Ocean(van Ginneken et al 2005) and complex reproductive cycle (Gousset 1990Pedersen 2003 2004) Furthermore genetic proximity (Albert et al 2006)the same spawning location (Aoyama amp Tsukamoto 1997) and several phys-iological similarities (Tesch 2003) between American and European eelssuggest that even if they are not using exactly the same strategies to com-municate olfaction plays a similar role in both species
However little is known about olfactory sensitivity of the anguillid eelto conspecific or hetero-specific derived stimuli or whether these changeswith the sex or maturational state of the donor or of the potential role thatpheromones might may play in eel biology Further olfaction in the pisciv-orous anguillid eel could be an important sense in interactions with otheraquatic species (such as hunting and territoriality) If olfaction plays a rolein one or several aspects of eel reproduction and behaviour this knowledgecould be included in reproduction protocols for the benefit of the aquaculture
1394 Huertas Canaacuterio amp Hubbard
industry or in strategies for the management or re-stocking of wild popula-tions
Do anguillid eels communicate using chemical cues
Eels have a well developed olfactory rosette (Schulte 1972 Yamamotoamp Ueda 1978 Ostrander et al 2000) (Figure 2) This organ experiencesdramatic changes during sexual maturation (Pankhurst amp Lythgoe 1983Sorensen amp Pankhurst 1988) which may be an indication of the involve-ment of chemical communication andor adaptation to the marine environ-ment As yet few studies have addressed the uses of chemical senses byeels most to date are behavioural studies showing the chemo-attraction (orrepulsion) to a specific odorant (Table 1) The main aim of these studies wasrelated to the detection of chemical cues by means of olfaction or loss oforientation in anosmic individuals (Hain 1975 Barbin 1998 Barbin et al1998) that suggest that olfaction is important in the control of eel life cycleThe substances described as chemo-attractants for glass and yellow eels in-clude water conditioned by conspecifics (Pesaro et al 1981 Sorensen 1986Briand et al 2002) extracts of mucus (Saglio 1982) bile salts (Sola 1993b)and plant- and animal-conditioned water (Sorensen 1986) In these casesodorants could act as chemical cues involved in migration from estuarine tofreshwater environments In some studies of American (Anguilla rostrata)(Creutzberg 1958 Miles 1968 Kleckner 1983 Barbin 1998 Barbin et al1998) and European eels (Westin 1998) the importance of chemical sensesto find specific sites and co-ordinate spawning in the marine environment wasinvestigated Other studies have shown the use of olfaction in prey capture(Carton amp Montgomery 2003) which may be related to the high sensitivityto amino acids in the first stages of growth (Silver 1982 Crnjar et al 1992Sola 1993a Sola amp Tongiorgi 1998)
Recently some studies have suggested a role for chemical signals duringreproduction Culture density in the first stages of glass eel culture affectsgrowth and sex differentiation during the later yellow eel phase (Huertas ampCerda 2006) (Figure 3) How is this effect mediated It was suggested thatsocial interaction is an important factor during growth and sexual differen-tiation and probably this social interaction is mainly mediated by chemicalcues rather than visual or tactile signals (Huertas et al 2006) Consequently
Chemical communication in eels 1395Ta
ble
1C
hem
ical
com
mun
icat
ion
stud
ies
inee
lspe
cies
Spec
ies
Odo
rant
Stag
eE
ffec
tR
efer
ence
Aa
ngui
lla
Wat
erco
nditi
oned
bygl
ass
eel
Gla
ssndashe
lver
Che
mo
attr
actio
nPe
saro
etal
(19
81)
Ar
ostr
atta
Wat
erco
nditi
oned
bym
atur
efe
mal
eM
atur
em
ale
Spaw
ning
beha
vior
Sore
nsen
ampW
inn
(198
4)A
ang
uill
aW
ater
cond
ition
edby
undi
ffer
entia
ted
eel
Elv
erC
hem
oat
trac
tion
(eel
trap
)B
rian
det
al(
2002
)A
ang
uill
aW
ater
cond
ition
edby
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
sex
ual
Hue
rtas
etal
(20
06)
mat
ure
eels
pher
omon
eA
ros
trat
aC
hem
ical
cues
rele
ased
bym
atur
eee
lsM
atur
eC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Kle
ckne
r(1
983)
Aa
ngui
lla
Che
mic
alcu
esin
tidal
wat
ers
Elv
erC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Cre
utzb
erg
(195
8)A
ros
trat
aC
hem
ical
cues
infr
eshw
ater
sG
lass
eel
Che
mo
attr
actio
n(s
igna
lmig
ratio
n)M
iles
(196
8)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tG
lass
eel
Che
mo
attr
actio
nTo
si(1
990)
Ar
ostr
ata
Che
mic
alcu
esin
tidal
wat
ers
Yel
low
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ros
trat
aC
hem
ical
cues
intid
alw
ater
sSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)W
estin
(199
8)A
ros
trat
aR
inse
plan
tsa
nim
als
inan
imat
eob
ject
sG
lass
eel
Che
mo
attr
actio
nSo
rens
en(1
986)
Aa
ngui
lla
Geo
smin
(inl
and
wat
erod
our)
Gla
ssee
lC
hem
oat
trac
tion
Tosi
(199
3)A
ang
uill
aM
ucus
bile
int
estin
alflu
idof
Yel
low
eel
Che
mo
attr
actio
n(o
nly
muc
us)
Sagl
io(1
982)
undi
ffer
entia
ted
eels
Aa
ngui
lla
Bile
and
muc
usof
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
Hue
rtas
etal
(20
07)
mat
ure
eels
Aa
ngui
lla
Bile
salts
and
taur
ine
Gla
ssee
lC
hem
oat
trac
tion
(thr
esho
ld10
minus10M
)So
la(1
993b
)A
ang
uill
aA
min
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
Sola
(199
3a)
Aa
ngui
lla
Non
-pro
tein
amin
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
(thr
esho
ld10
minus9M
)So
laamp
Tong
iron
i(19
98)
Aa
ngui
lla
Am
ino
acid
sY
elow
eel
Olf
acto
ryse
nsiti
vity
Crn
jar
etal
(19
92)
Ar
ostr
ata
Am
ino
acid
sSi
lver
eel
Olf
acto
ryse
nsiti
vity
Silv
er(1
982)
Ar
eind
hard
tii
Food
Silv
eree
lC
hem
oat
trac
tion
Car
ton
etal
(20
03)
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

1390 Huertas Canaacuterio amp Hubbard
Why is the eel interesting
The increasing interest in Anguillid eels is related to several factors Firstlythe genus Anguilla has a complex life-cycle which includes a worldwidedistribution and a catadromous migration that have been subject of severalecological and genetic studies (Aoyama amp Tsukamoto 1997 Westin 1998)Secondly the eel is a highly appreciated commercial species with a welldeveloped aquaculture industry (Peacuterez et al 2004) Finally many eel speciesare endangered in several cases natural stocks have been reduced to levelsbelow those necessary for a viable population (van Ginneken et al 2007)A concerted effort by governments scientists fisheries and the aquacultureindustry is necessary to prevent the extinction of this once common andwide-spread fish
An example of complex life-cycle the European eel
The European eel (Anguilla anguilla L) is one of the 18 species of the genusAnguilla which in turn belongs to the subdivision Elopomorpha a groupthat appeared early in teleost evolution and the order anguilliformes whichalso includes moray eels (Nelson 2006) The amazing life-cycle of Europeaneels (Figure 1) consists of a migration of adults over a distance of 5000 kmacross the Atlantic Ocean to the Sargasso Sea between Bermuda Island andPuerto Rico and the opposite migration of larvae to continental waters (vanGinneken amp Maes 2005) Adult males start to migrate in September andfemales one or two months later (Usui 1991 Tesch 2003) with a swim-ming velocity of 05 body lengths the journey should take between 6 and 7months (van Ginneken et al 2005) During this journey the adults do not eatbut nevertheless undergo sexual maturation After spawning they die (vanGinneken amp Maes 2005) The resulting leaf-like shape larvae called lep-tocephali (Lecompte-Finiger et al 2004) swim in less than one year (vanGinneken et al 2005) to the European coast aided by Gulf-Steam currentsand on reaching the continental shelf metamorphose into glass eels be-fore entering rivers in western Europe and North Africa (Lecompte-Finiger1994 McCleave et al 1998) around November to December Transparentglass eels undergo a series of metamorphic events until they reach the yel-low stage (Elie et al 1982) and after a variable growth period becomesilver eels (Deelder 1984) in a process more linked with puberty than meta-morphosis (Dufour 2000 Aroua et al 2005) Then after a variable period
Chemical communication in eels 1391
Figure 1 Life cycle of the European eel (Anguilla anguilla) Sexually immature lsquoyellowrsquoeels grow in rivers lakes and estuaries for several years (10ndash30) until the lsquosilverrsquo eel stageThen they migrate across the Atlantic Ocean to the Sargasso see where sexually mature fishmate and spawn sexual maturation occurs lsquoen routersquo but feeding ceases The resulting lep-tocephali larvae then travel to the European coast and when they reach the continental plat-form metamorphose into lsquoglassrsquo eels and enter estuaries (between November and February)and make their way upstream to inland freshwaters Olfaction may be involved in any or all
of these stages
(6 to 30 years) silver adults in the first stages of gametogenesis completethe cycle by their migration to the Sargasso Sea (van Ginneken et al 2005)Many aspects of the eelrsquos life-cycle are still unknown such as where exactlythe spawning location is how do they find this location at what depth dothey swim in the Atlantic Ocean how do the two sexes meet (since malesstart migrating before the females) (Haraldstad et al 1985 Usui 1991) andwhen and how does sexual maturation occur Current tracking and teleme-try studies promise to provide some answers to these questions (McCleave ampArnold 1999 Jellyman amp Tsukamoto 2005) However other answers maybe found by laboratory experiments The role of olfaction in these processesis largely unknown The aim of this review is to briefly outline what is knownand suggest directions that future research should take
Eel aquaculture and current situation of natural stocks
Globally anguillid eels are an important and prized food source (Heinsbroek1991 Usui 1991 Gousset 1992) Since the capture of wild stocks is insuffi-
1392 Huertas Canaacuterio amp Hubbard
cient to satisfy the market eel aquaculture has increased exponentially overthe last 30 years Asian countries are the highest producers and consumers(more than 220 000 metric tonnes per year) whereas European productionconstitutes less than 5 of the worldwide total (about 10 000 metric tonnes(Peacuterez et al 2004 ICES 2005) The main problems associated with eelaquaculture are provision and control of appropriate environmental factors(temperature salinity light) the formulation of diets (to reduce mortalityby cannibalism and increase growth) and the artificial induction of sexualreproduction
In captivity the European eel does not reach sexual maturity without hor-monal manipulation only the Japanese eel Anguilla japonica has been suc-cessfully induced to spawn under artificial conditions (Tanaka et al 2003)Even then mass production of high quality glass eels has not yet becomeroutine (Furuita et al 2006) To induce sexual maturation in Japanese andEuropean eels injections with human chorionic gonadotropin (hCG) in thecase of males or pituitary extract in the case of females are necessary(Ohta et al 1996a 1997 Hao-ran et al 1998 Ijiri et al 1998 Asturianoet al 2002) After pituitary extract treatment females usually require treat-ment with the maturation-inducing steroid 1720β-dihydroxy-4-pregnen-3-one (1720βP) to induce ovulation (Ohta et al 1996b Kagawa et al 1997)
The inability to produce glass eels commercially means that the aquacul-ture industry depends on the capture of wild stocks increasing the fishing ef-fort in the eel stocks Thus together the over-exploitation of natural stocksthe change in oceanographic conditions (global warming) degradation andreduction of suitable freshwater habitats pollution and parasitism have re-sulted in an alarming decrease of wild glass and adult eel population (Briandet al 2003 Dekker 2003 Starkie 2003) As a result the current Europeaneel population stands at about 1 of the level of 30 years ago there areno signs of recovery This is of major concern to eel conservation fisheriesaquaculture and industry As a consequence this species will be included inCITES by 2009 and the ICESEIFAC Eel Working Group (September 2007)proposed that a self-sufficient eel aquaculture industry was vital for the re-covery of natural stocks and research focusing on spawner quality and repro-duction was urgently needed to achieve this (van Ginneken et al 2007)
How can studies of olfaction help
Communication is defined as ldquointegration between a lsquosignallerrsquo who pro-duces a sensory stimulus or signal (a chemical in the case of chemical com-
Chemical communication in eels 1393
munication) and a lsquoreceiverrsquo who perceives the signal and makes a conse-quent behavioral decisionrdquo (Rosenthal amp Lobel 2006) Pheromones are sub-stances released by conspecifics which then provoke an innate behaviouralandor physiological response in the receiver (Stacey amp Sorensen 2002)There is little doubt that pheromonal communication plays an important rolein many aspects of fish biology However there is great variability in thetissues of origin of pheromones their routes and regulation of release theirchemical structure and the nature of the receiversrsquo response(s) which en-compass a wide repertoire of behavioural and physiological effects (Staceyamp Sorensen 2002) For example many studies have shown that pheromoneshelp to co-ordinate reproductive events (Stacey amp Sorensen 2002) are in-volved in inter-specific or intra-specific recognition mate-choice (Rosenthalamp Lobel 2006) act as riverine pheromone plumes in species such as the sealamprey (Polkinghorne et al 2001 Li et al 2002) or Arctic char (Selset ampDoving 1980) or play an important role in the alarm reaction (Win 2000Ide et al 2003 Kristensen amp Closs 2004) However most available infor-mation about fish pheromones is derived from freshwater spawning species(eg salmonids and cyprinids) very little is known about species that spawnin sea water
Several aspects of anguillid eel biology together with their highly devel-oped sense of smell (Silver 1982 Crnjar et al 1992 Sola 1993ab Sola ampTongiorgi 1998) suggest that chemical communication could be involved atkey stages of their life-history eg the migration over the Atlantic Ocean(van Ginneken et al 2005) and complex reproductive cycle (Gousset 1990Pedersen 2003 2004) Furthermore genetic proximity (Albert et al 2006)the same spawning location (Aoyama amp Tsukamoto 1997) and several phys-iological similarities (Tesch 2003) between American and European eelssuggest that even if they are not using exactly the same strategies to com-municate olfaction plays a similar role in both species
However little is known about olfactory sensitivity of the anguillid eelto conspecific or hetero-specific derived stimuli or whether these changeswith the sex or maturational state of the donor or of the potential role thatpheromones might may play in eel biology Further olfaction in the pisciv-orous anguillid eel could be an important sense in interactions with otheraquatic species (such as hunting and territoriality) If olfaction plays a rolein one or several aspects of eel reproduction and behaviour this knowledgecould be included in reproduction protocols for the benefit of the aquaculture
1394 Huertas Canaacuterio amp Hubbard
industry or in strategies for the management or re-stocking of wild popula-tions
Do anguillid eels communicate using chemical cues
Eels have a well developed olfactory rosette (Schulte 1972 Yamamotoamp Ueda 1978 Ostrander et al 2000) (Figure 2) This organ experiencesdramatic changes during sexual maturation (Pankhurst amp Lythgoe 1983Sorensen amp Pankhurst 1988) which may be an indication of the involve-ment of chemical communication andor adaptation to the marine environ-ment As yet few studies have addressed the uses of chemical senses byeels most to date are behavioural studies showing the chemo-attraction (orrepulsion) to a specific odorant (Table 1) The main aim of these studies wasrelated to the detection of chemical cues by means of olfaction or loss oforientation in anosmic individuals (Hain 1975 Barbin 1998 Barbin et al1998) that suggest that olfaction is important in the control of eel life cycleThe substances described as chemo-attractants for glass and yellow eels in-clude water conditioned by conspecifics (Pesaro et al 1981 Sorensen 1986Briand et al 2002) extracts of mucus (Saglio 1982) bile salts (Sola 1993b)and plant- and animal-conditioned water (Sorensen 1986) In these casesodorants could act as chemical cues involved in migration from estuarine tofreshwater environments In some studies of American (Anguilla rostrata)(Creutzberg 1958 Miles 1968 Kleckner 1983 Barbin 1998 Barbin et al1998) and European eels (Westin 1998) the importance of chemical sensesto find specific sites and co-ordinate spawning in the marine environment wasinvestigated Other studies have shown the use of olfaction in prey capture(Carton amp Montgomery 2003) which may be related to the high sensitivityto amino acids in the first stages of growth (Silver 1982 Crnjar et al 1992Sola 1993a Sola amp Tongiorgi 1998)
Recently some studies have suggested a role for chemical signals duringreproduction Culture density in the first stages of glass eel culture affectsgrowth and sex differentiation during the later yellow eel phase (Huertas ampCerda 2006) (Figure 3) How is this effect mediated It was suggested thatsocial interaction is an important factor during growth and sexual differen-tiation and probably this social interaction is mainly mediated by chemicalcues rather than visual or tactile signals (Huertas et al 2006) Consequently
Chemical communication in eels 1395Ta
ble
1C
hem
ical
com
mun
icat
ion
stud
ies
inee
lspe
cies
Spec
ies
Odo
rant
Stag
eE
ffec
tR
efer
ence
Aa
ngui
lla
Wat
erco
nditi
oned
bygl
ass
eel
Gla
ssndashe
lver
Che
mo
attr
actio
nPe
saro
etal
(19
81)
Ar
ostr
atta
Wat
erco
nditi
oned
bym
atur
efe
mal
eM
atur
em
ale
Spaw
ning
beha
vior
Sore
nsen
ampW
inn
(198
4)A
ang
uill
aW
ater
cond
ition
edby
undi
ffer
entia
ted
eel
Elv
erC
hem
oat
trac
tion
(eel
trap
)B
rian
det
al(
2002
)A
ang
uill
aW
ater
cond
ition
edby
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
sex
ual
Hue
rtas
etal
(20
06)
mat
ure
eels
pher
omon
eA
ros
trat
aC
hem
ical
cues
rele
ased
bym
atur
eee
lsM
atur
eC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Kle
ckne
r(1
983)
Aa
ngui
lla
Che
mic
alcu
esin
tidal
wat
ers
Elv
erC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Cre
utzb
erg
(195
8)A
ros
trat
aC
hem
ical
cues
infr
eshw
ater
sG
lass
eel
Che
mo
attr
actio
n(s
igna
lmig
ratio
n)M
iles
(196
8)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tG
lass
eel
Che
mo
attr
actio
nTo
si(1
990)
Ar
ostr
ata
Che
mic
alcu
esin
tidal
wat
ers
Yel
low
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ros
trat
aC
hem
ical
cues
intid
alw
ater
sSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)W
estin
(199
8)A
ros
trat
aR
inse
plan
tsa
nim
als
inan
imat
eob
ject
sG
lass
eel
Che
mo
attr
actio
nSo
rens
en(1
986)
Aa
ngui
lla
Geo
smin
(inl
and
wat
erod
our)
Gla
ssee
lC
hem
oat
trac
tion
Tosi
(199
3)A
ang
uill
aM
ucus
bile
int
estin
alflu
idof
Yel
low
eel
Che
mo
attr
actio
n(o
nly
muc
us)
Sagl
io(1
982)
undi
ffer
entia
ted
eels
Aa
ngui
lla
Bile
and
muc
usof
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
Hue
rtas
etal
(20
07)
mat
ure
eels
Aa
ngui
lla
Bile
salts
and
taur
ine
Gla
ssee
lC
hem
oat
trac
tion
(thr
esho
ld10
minus10M
)So
la(1
993b
)A
ang
uill
aA
min
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
Sola
(199
3a)
Aa
ngui
lla
Non
-pro
tein
amin
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
(thr
esho
ld10
minus9M
)So
laamp
Tong
iron
i(19
98)
Aa
ngui
lla
Am
ino
acid
sY
elow
eel
Olf
acto
ryse
nsiti
vity
Crn
jar
etal
(19
92)
Ar
ostr
ata
Am
ino
acid
sSi
lver
eel
Olf
acto
ryse
nsiti
vity
Silv
er(1
982)
Ar
eind
hard
tii
Food
Silv
eree
lC
hem
oat
trac
tion
Car
ton
etal
(20
03)
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

Chemical communication in eels 1391
Figure 1 Life cycle of the European eel (Anguilla anguilla) Sexually immature lsquoyellowrsquoeels grow in rivers lakes and estuaries for several years (10ndash30) until the lsquosilverrsquo eel stageThen they migrate across the Atlantic Ocean to the Sargasso see where sexually mature fishmate and spawn sexual maturation occurs lsquoen routersquo but feeding ceases The resulting lep-tocephali larvae then travel to the European coast and when they reach the continental plat-form metamorphose into lsquoglassrsquo eels and enter estuaries (between November and February)and make their way upstream to inland freshwaters Olfaction may be involved in any or all
of these stages
(6 to 30 years) silver adults in the first stages of gametogenesis completethe cycle by their migration to the Sargasso Sea (van Ginneken et al 2005)Many aspects of the eelrsquos life-cycle are still unknown such as where exactlythe spawning location is how do they find this location at what depth dothey swim in the Atlantic Ocean how do the two sexes meet (since malesstart migrating before the females) (Haraldstad et al 1985 Usui 1991) andwhen and how does sexual maturation occur Current tracking and teleme-try studies promise to provide some answers to these questions (McCleave ampArnold 1999 Jellyman amp Tsukamoto 2005) However other answers maybe found by laboratory experiments The role of olfaction in these processesis largely unknown The aim of this review is to briefly outline what is knownand suggest directions that future research should take
Eel aquaculture and current situation of natural stocks
Globally anguillid eels are an important and prized food source (Heinsbroek1991 Usui 1991 Gousset 1992) Since the capture of wild stocks is insuffi-
1392 Huertas Canaacuterio amp Hubbard
cient to satisfy the market eel aquaculture has increased exponentially overthe last 30 years Asian countries are the highest producers and consumers(more than 220 000 metric tonnes per year) whereas European productionconstitutes less than 5 of the worldwide total (about 10 000 metric tonnes(Peacuterez et al 2004 ICES 2005) The main problems associated with eelaquaculture are provision and control of appropriate environmental factors(temperature salinity light) the formulation of diets (to reduce mortalityby cannibalism and increase growth) and the artificial induction of sexualreproduction
In captivity the European eel does not reach sexual maturity without hor-monal manipulation only the Japanese eel Anguilla japonica has been suc-cessfully induced to spawn under artificial conditions (Tanaka et al 2003)Even then mass production of high quality glass eels has not yet becomeroutine (Furuita et al 2006) To induce sexual maturation in Japanese andEuropean eels injections with human chorionic gonadotropin (hCG) in thecase of males or pituitary extract in the case of females are necessary(Ohta et al 1996a 1997 Hao-ran et al 1998 Ijiri et al 1998 Asturianoet al 2002) After pituitary extract treatment females usually require treat-ment with the maturation-inducing steroid 1720β-dihydroxy-4-pregnen-3-one (1720βP) to induce ovulation (Ohta et al 1996b Kagawa et al 1997)
The inability to produce glass eels commercially means that the aquacul-ture industry depends on the capture of wild stocks increasing the fishing ef-fort in the eel stocks Thus together the over-exploitation of natural stocksthe change in oceanographic conditions (global warming) degradation andreduction of suitable freshwater habitats pollution and parasitism have re-sulted in an alarming decrease of wild glass and adult eel population (Briandet al 2003 Dekker 2003 Starkie 2003) As a result the current Europeaneel population stands at about 1 of the level of 30 years ago there areno signs of recovery This is of major concern to eel conservation fisheriesaquaculture and industry As a consequence this species will be included inCITES by 2009 and the ICESEIFAC Eel Working Group (September 2007)proposed that a self-sufficient eel aquaculture industry was vital for the re-covery of natural stocks and research focusing on spawner quality and repro-duction was urgently needed to achieve this (van Ginneken et al 2007)
How can studies of olfaction help
Communication is defined as ldquointegration between a lsquosignallerrsquo who pro-duces a sensory stimulus or signal (a chemical in the case of chemical com-
Chemical communication in eels 1393
munication) and a lsquoreceiverrsquo who perceives the signal and makes a conse-quent behavioral decisionrdquo (Rosenthal amp Lobel 2006) Pheromones are sub-stances released by conspecifics which then provoke an innate behaviouralandor physiological response in the receiver (Stacey amp Sorensen 2002)There is little doubt that pheromonal communication plays an important rolein many aspects of fish biology However there is great variability in thetissues of origin of pheromones their routes and regulation of release theirchemical structure and the nature of the receiversrsquo response(s) which en-compass a wide repertoire of behavioural and physiological effects (Staceyamp Sorensen 2002) For example many studies have shown that pheromoneshelp to co-ordinate reproductive events (Stacey amp Sorensen 2002) are in-volved in inter-specific or intra-specific recognition mate-choice (Rosenthalamp Lobel 2006) act as riverine pheromone plumes in species such as the sealamprey (Polkinghorne et al 2001 Li et al 2002) or Arctic char (Selset ampDoving 1980) or play an important role in the alarm reaction (Win 2000Ide et al 2003 Kristensen amp Closs 2004) However most available infor-mation about fish pheromones is derived from freshwater spawning species(eg salmonids and cyprinids) very little is known about species that spawnin sea water
Several aspects of anguillid eel biology together with their highly devel-oped sense of smell (Silver 1982 Crnjar et al 1992 Sola 1993ab Sola ampTongiorgi 1998) suggest that chemical communication could be involved atkey stages of their life-history eg the migration over the Atlantic Ocean(van Ginneken et al 2005) and complex reproductive cycle (Gousset 1990Pedersen 2003 2004) Furthermore genetic proximity (Albert et al 2006)the same spawning location (Aoyama amp Tsukamoto 1997) and several phys-iological similarities (Tesch 2003) between American and European eelssuggest that even if they are not using exactly the same strategies to com-municate olfaction plays a similar role in both species
However little is known about olfactory sensitivity of the anguillid eelto conspecific or hetero-specific derived stimuli or whether these changeswith the sex or maturational state of the donor or of the potential role thatpheromones might may play in eel biology Further olfaction in the pisciv-orous anguillid eel could be an important sense in interactions with otheraquatic species (such as hunting and territoriality) If olfaction plays a rolein one or several aspects of eel reproduction and behaviour this knowledgecould be included in reproduction protocols for the benefit of the aquaculture
1394 Huertas Canaacuterio amp Hubbard
industry or in strategies for the management or re-stocking of wild popula-tions
Do anguillid eels communicate using chemical cues
Eels have a well developed olfactory rosette (Schulte 1972 Yamamotoamp Ueda 1978 Ostrander et al 2000) (Figure 2) This organ experiencesdramatic changes during sexual maturation (Pankhurst amp Lythgoe 1983Sorensen amp Pankhurst 1988) which may be an indication of the involve-ment of chemical communication andor adaptation to the marine environ-ment As yet few studies have addressed the uses of chemical senses byeels most to date are behavioural studies showing the chemo-attraction (orrepulsion) to a specific odorant (Table 1) The main aim of these studies wasrelated to the detection of chemical cues by means of olfaction or loss oforientation in anosmic individuals (Hain 1975 Barbin 1998 Barbin et al1998) that suggest that olfaction is important in the control of eel life cycleThe substances described as chemo-attractants for glass and yellow eels in-clude water conditioned by conspecifics (Pesaro et al 1981 Sorensen 1986Briand et al 2002) extracts of mucus (Saglio 1982) bile salts (Sola 1993b)and plant- and animal-conditioned water (Sorensen 1986) In these casesodorants could act as chemical cues involved in migration from estuarine tofreshwater environments In some studies of American (Anguilla rostrata)(Creutzberg 1958 Miles 1968 Kleckner 1983 Barbin 1998 Barbin et al1998) and European eels (Westin 1998) the importance of chemical sensesto find specific sites and co-ordinate spawning in the marine environment wasinvestigated Other studies have shown the use of olfaction in prey capture(Carton amp Montgomery 2003) which may be related to the high sensitivityto amino acids in the first stages of growth (Silver 1982 Crnjar et al 1992Sola 1993a Sola amp Tongiorgi 1998)
Recently some studies have suggested a role for chemical signals duringreproduction Culture density in the first stages of glass eel culture affectsgrowth and sex differentiation during the later yellow eel phase (Huertas ampCerda 2006) (Figure 3) How is this effect mediated It was suggested thatsocial interaction is an important factor during growth and sexual differen-tiation and probably this social interaction is mainly mediated by chemicalcues rather than visual or tactile signals (Huertas et al 2006) Consequently
Chemical communication in eels 1395Ta
ble
1C
hem
ical
com
mun
icat
ion
stud
ies
inee
lspe
cies
Spec
ies
Odo
rant
Stag
eE
ffec
tR
efer
ence
Aa
ngui
lla
Wat
erco
nditi
oned
bygl
ass
eel
Gla
ssndashe
lver
Che
mo
attr
actio
nPe
saro
etal
(19
81)
Ar
ostr
atta
Wat
erco
nditi
oned
bym
atur
efe
mal
eM
atur
em
ale
Spaw
ning
beha
vior
Sore
nsen
ampW
inn
(198
4)A
ang
uill
aW
ater
cond
ition
edby
undi
ffer
entia
ted
eel
Elv
erC
hem
oat
trac
tion
(eel
trap
)B
rian
det
al(
2002
)A
ang
uill
aW
ater
cond
ition
edby
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
sex
ual
Hue
rtas
etal
(20
06)
mat
ure
eels
pher
omon
eA
ros
trat
aC
hem
ical
cues
rele
ased
bym
atur
eee
lsM
atur
eC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Kle
ckne
r(1
983)
Aa
ngui
lla
Che
mic
alcu
esin
tidal
wat
ers
Elv
erC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Cre
utzb
erg
(195
8)A
ros
trat
aC
hem
ical
cues
infr
eshw
ater
sG
lass
eel
Che
mo
attr
actio
n(s
igna
lmig
ratio
n)M
iles
(196
8)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tG
lass
eel
Che
mo
attr
actio
nTo
si(1
990)
Ar
ostr
ata
Che
mic
alcu
esin
tidal
wat
ers
Yel
low
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ros
trat
aC
hem
ical
cues
intid
alw
ater
sSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)W
estin
(199
8)A
ros
trat
aR
inse
plan
tsa
nim
als
inan
imat
eob
ject
sG
lass
eel
Che
mo
attr
actio
nSo
rens
en(1
986)
Aa
ngui
lla
Geo
smin
(inl
and
wat
erod
our)
Gla
ssee
lC
hem
oat
trac
tion
Tosi
(199
3)A
ang
uill
aM
ucus
bile
int
estin
alflu
idof
Yel
low
eel
Che
mo
attr
actio
n(o
nly
muc
us)
Sagl
io(1
982)
undi
ffer
entia
ted
eels
Aa
ngui
lla
Bile
and
muc
usof
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
Hue
rtas
etal
(20
07)
mat
ure
eels
Aa
ngui
lla
Bile
salts
and
taur
ine
Gla
ssee
lC
hem
oat
trac
tion
(thr
esho
ld10
minus10M
)So
la(1
993b
)A
ang
uill
aA
min
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
Sola
(199
3a)
Aa
ngui
lla
Non
-pro
tein
amin
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
(thr
esho
ld10
minus9M
)So
laamp
Tong
iron
i(19
98)
Aa
ngui
lla
Am
ino
acid
sY
elow
eel
Olf
acto
ryse
nsiti
vity
Crn
jar
etal
(19
92)
Ar
ostr
ata
Am
ino
acid
sSi
lver
eel
Olf
acto
ryse
nsiti
vity
Silv
er(1
982)
Ar
eind
hard
tii
Food
Silv
eree
lC
hem
oat
trac
tion
Car
ton
etal
(20
03)
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

1392 Huertas Canaacuterio amp Hubbard
cient to satisfy the market eel aquaculture has increased exponentially overthe last 30 years Asian countries are the highest producers and consumers(more than 220 000 metric tonnes per year) whereas European productionconstitutes less than 5 of the worldwide total (about 10 000 metric tonnes(Peacuterez et al 2004 ICES 2005) The main problems associated with eelaquaculture are provision and control of appropriate environmental factors(temperature salinity light) the formulation of diets (to reduce mortalityby cannibalism and increase growth) and the artificial induction of sexualreproduction
In captivity the European eel does not reach sexual maturity without hor-monal manipulation only the Japanese eel Anguilla japonica has been suc-cessfully induced to spawn under artificial conditions (Tanaka et al 2003)Even then mass production of high quality glass eels has not yet becomeroutine (Furuita et al 2006) To induce sexual maturation in Japanese andEuropean eels injections with human chorionic gonadotropin (hCG) in thecase of males or pituitary extract in the case of females are necessary(Ohta et al 1996a 1997 Hao-ran et al 1998 Ijiri et al 1998 Asturianoet al 2002) After pituitary extract treatment females usually require treat-ment with the maturation-inducing steroid 1720β-dihydroxy-4-pregnen-3-one (1720βP) to induce ovulation (Ohta et al 1996b Kagawa et al 1997)
The inability to produce glass eels commercially means that the aquacul-ture industry depends on the capture of wild stocks increasing the fishing ef-fort in the eel stocks Thus together the over-exploitation of natural stocksthe change in oceanographic conditions (global warming) degradation andreduction of suitable freshwater habitats pollution and parasitism have re-sulted in an alarming decrease of wild glass and adult eel population (Briandet al 2003 Dekker 2003 Starkie 2003) As a result the current Europeaneel population stands at about 1 of the level of 30 years ago there areno signs of recovery This is of major concern to eel conservation fisheriesaquaculture and industry As a consequence this species will be included inCITES by 2009 and the ICESEIFAC Eel Working Group (September 2007)proposed that a self-sufficient eel aquaculture industry was vital for the re-covery of natural stocks and research focusing on spawner quality and repro-duction was urgently needed to achieve this (van Ginneken et al 2007)
How can studies of olfaction help
Communication is defined as ldquointegration between a lsquosignallerrsquo who pro-duces a sensory stimulus or signal (a chemical in the case of chemical com-
Chemical communication in eels 1393
munication) and a lsquoreceiverrsquo who perceives the signal and makes a conse-quent behavioral decisionrdquo (Rosenthal amp Lobel 2006) Pheromones are sub-stances released by conspecifics which then provoke an innate behaviouralandor physiological response in the receiver (Stacey amp Sorensen 2002)There is little doubt that pheromonal communication plays an important rolein many aspects of fish biology However there is great variability in thetissues of origin of pheromones their routes and regulation of release theirchemical structure and the nature of the receiversrsquo response(s) which en-compass a wide repertoire of behavioural and physiological effects (Staceyamp Sorensen 2002) For example many studies have shown that pheromoneshelp to co-ordinate reproductive events (Stacey amp Sorensen 2002) are in-volved in inter-specific or intra-specific recognition mate-choice (Rosenthalamp Lobel 2006) act as riverine pheromone plumes in species such as the sealamprey (Polkinghorne et al 2001 Li et al 2002) or Arctic char (Selset ampDoving 1980) or play an important role in the alarm reaction (Win 2000Ide et al 2003 Kristensen amp Closs 2004) However most available infor-mation about fish pheromones is derived from freshwater spawning species(eg salmonids and cyprinids) very little is known about species that spawnin sea water
Several aspects of anguillid eel biology together with their highly devel-oped sense of smell (Silver 1982 Crnjar et al 1992 Sola 1993ab Sola ampTongiorgi 1998) suggest that chemical communication could be involved atkey stages of their life-history eg the migration over the Atlantic Ocean(van Ginneken et al 2005) and complex reproductive cycle (Gousset 1990Pedersen 2003 2004) Furthermore genetic proximity (Albert et al 2006)the same spawning location (Aoyama amp Tsukamoto 1997) and several phys-iological similarities (Tesch 2003) between American and European eelssuggest that even if they are not using exactly the same strategies to com-municate olfaction plays a similar role in both species
However little is known about olfactory sensitivity of the anguillid eelto conspecific or hetero-specific derived stimuli or whether these changeswith the sex or maturational state of the donor or of the potential role thatpheromones might may play in eel biology Further olfaction in the pisciv-orous anguillid eel could be an important sense in interactions with otheraquatic species (such as hunting and territoriality) If olfaction plays a rolein one or several aspects of eel reproduction and behaviour this knowledgecould be included in reproduction protocols for the benefit of the aquaculture
1394 Huertas Canaacuterio amp Hubbard
industry or in strategies for the management or re-stocking of wild popula-tions
Do anguillid eels communicate using chemical cues
Eels have a well developed olfactory rosette (Schulte 1972 Yamamotoamp Ueda 1978 Ostrander et al 2000) (Figure 2) This organ experiencesdramatic changes during sexual maturation (Pankhurst amp Lythgoe 1983Sorensen amp Pankhurst 1988) which may be an indication of the involve-ment of chemical communication andor adaptation to the marine environ-ment As yet few studies have addressed the uses of chemical senses byeels most to date are behavioural studies showing the chemo-attraction (orrepulsion) to a specific odorant (Table 1) The main aim of these studies wasrelated to the detection of chemical cues by means of olfaction or loss oforientation in anosmic individuals (Hain 1975 Barbin 1998 Barbin et al1998) that suggest that olfaction is important in the control of eel life cycleThe substances described as chemo-attractants for glass and yellow eels in-clude water conditioned by conspecifics (Pesaro et al 1981 Sorensen 1986Briand et al 2002) extracts of mucus (Saglio 1982) bile salts (Sola 1993b)and plant- and animal-conditioned water (Sorensen 1986) In these casesodorants could act as chemical cues involved in migration from estuarine tofreshwater environments In some studies of American (Anguilla rostrata)(Creutzberg 1958 Miles 1968 Kleckner 1983 Barbin 1998 Barbin et al1998) and European eels (Westin 1998) the importance of chemical sensesto find specific sites and co-ordinate spawning in the marine environment wasinvestigated Other studies have shown the use of olfaction in prey capture(Carton amp Montgomery 2003) which may be related to the high sensitivityto amino acids in the first stages of growth (Silver 1982 Crnjar et al 1992Sola 1993a Sola amp Tongiorgi 1998)
Recently some studies have suggested a role for chemical signals duringreproduction Culture density in the first stages of glass eel culture affectsgrowth and sex differentiation during the later yellow eel phase (Huertas ampCerda 2006) (Figure 3) How is this effect mediated It was suggested thatsocial interaction is an important factor during growth and sexual differen-tiation and probably this social interaction is mainly mediated by chemicalcues rather than visual or tactile signals (Huertas et al 2006) Consequently
Chemical communication in eels 1395Ta
ble
1C
hem
ical
com
mun
icat
ion
stud
ies
inee
lspe
cies
Spec
ies
Odo
rant
Stag
eE
ffec
tR
efer
ence
Aa
ngui
lla
Wat
erco
nditi
oned
bygl
ass
eel
Gla
ssndashe
lver
Che
mo
attr
actio
nPe
saro
etal
(19
81)
Ar
ostr
atta
Wat
erco
nditi
oned
bym
atur
efe
mal
eM
atur
em
ale
Spaw
ning
beha
vior
Sore
nsen
ampW
inn
(198
4)A
ang
uill
aW
ater
cond
ition
edby
undi
ffer
entia
ted
eel
Elv
erC
hem
oat
trac
tion
(eel
trap
)B
rian
det
al(
2002
)A
ang
uill
aW
ater
cond
ition
edby
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
sex
ual
Hue
rtas
etal
(20
06)
mat
ure
eels
pher
omon
eA
ros
trat
aC
hem
ical
cues
rele
ased
bym
atur
eee
lsM
atur
eC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Kle
ckne
r(1
983)
Aa
ngui
lla
Che
mic
alcu
esin
tidal
wat
ers
Elv
erC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Cre
utzb
erg
(195
8)A
ros
trat
aC
hem
ical
cues
infr
eshw
ater
sG
lass
eel
Che
mo
attr
actio
n(s
igna
lmig
ratio
n)M
iles
(196
8)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tG
lass
eel
Che
mo
attr
actio
nTo
si(1
990)
Ar
ostr
ata
Che
mic
alcu
esin
tidal
wat
ers
Yel
low
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ros
trat
aC
hem
ical
cues
intid
alw
ater
sSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)W
estin
(199
8)A
ros
trat
aR
inse
plan
tsa
nim
als
inan
imat
eob
ject
sG
lass
eel
Che
mo
attr
actio
nSo
rens
en(1
986)
Aa
ngui
lla
Geo
smin
(inl
and
wat
erod
our)
Gla
ssee
lC
hem
oat
trac
tion
Tosi
(199
3)A
ang
uill
aM
ucus
bile
int
estin
alflu
idof
Yel
low
eel
Che
mo
attr
actio
n(o
nly
muc
us)
Sagl
io(1
982)
undi
ffer
entia
ted
eels
Aa
ngui
lla
Bile
and
muc
usof
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
Hue
rtas
etal
(20
07)
mat
ure
eels
Aa
ngui
lla
Bile
salts
and
taur
ine
Gla
ssee
lC
hem
oat
trac
tion
(thr
esho
ld10
minus10M
)So
la(1
993b
)A
ang
uill
aA
min
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
Sola
(199
3a)
Aa
ngui
lla
Non
-pro
tein
amin
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
(thr
esho
ld10
minus9M
)So
laamp
Tong
iron
i(19
98)
Aa
ngui
lla
Am
ino
acid
sY
elow
eel
Olf
acto
ryse
nsiti
vity
Crn
jar
etal
(19
92)
Ar
ostr
ata
Am
ino
acid
sSi
lver
eel
Olf
acto
ryse
nsiti
vity
Silv
er(1
982)
Ar
eind
hard
tii
Food
Silv
eree
lC
hem
oat
trac
tion
Car
ton
etal
(20
03)
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

Chemical communication in eels 1393
munication) and a lsquoreceiverrsquo who perceives the signal and makes a conse-quent behavioral decisionrdquo (Rosenthal amp Lobel 2006) Pheromones are sub-stances released by conspecifics which then provoke an innate behaviouralandor physiological response in the receiver (Stacey amp Sorensen 2002)There is little doubt that pheromonal communication plays an important rolein many aspects of fish biology However there is great variability in thetissues of origin of pheromones their routes and regulation of release theirchemical structure and the nature of the receiversrsquo response(s) which en-compass a wide repertoire of behavioural and physiological effects (Staceyamp Sorensen 2002) For example many studies have shown that pheromoneshelp to co-ordinate reproductive events (Stacey amp Sorensen 2002) are in-volved in inter-specific or intra-specific recognition mate-choice (Rosenthalamp Lobel 2006) act as riverine pheromone plumes in species such as the sealamprey (Polkinghorne et al 2001 Li et al 2002) or Arctic char (Selset ampDoving 1980) or play an important role in the alarm reaction (Win 2000Ide et al 2003 Kristensen amp Closs 2004) However most available infor-mation about fish pheromones is derived from freshwater spawning species(eg salmonids and cyprinids) very little is known about species that spawnin sea water
Several aspects of anguillid eel biology together with their highly devel-oped sense of smell (Silver 1982 Crnjar et al 1992 Sola 1993ab Sola ampTongiorgi 1998) suggest that chemical communication could be involved atkey stages of their life-history eg the migration over the Atlantic Ocean(van Ginneken et al 2005) and complex reproductive cycle (Gousset 1990Pedersen 2003 2004) Furthermore genetic proximity (Albert et al 2006)the same spawning location (Aoyama amp Tsukamoto 1997) and several phys-iological similarities (Tesch 2003) between American and European eelssuggest that even if they are not using exactly the same strategies to com-municate olfaction plays a similar role in both species
However little is known about olfactory sensitivity of the anguillid eelto conspecific or hetero-specific derived stimuli or whether these changeswith the sex or maturational state of the donor or of the potential role thatpheromones might may play in eel biology Further olfaction in the pisciv-orous anguillid eel could be an important sense in interactions with otheraquatic species (such as hunting and territoriality) If olfaction plays a rolein one or several aspects of eel reproduction and behaviour this knowledgecould be included in reproduction protocols for the benefit of the aquaculture
1394 Huertas Canaacuterio amp Hubbard
industry or in strategies for the management or re-stocking of wild popula-tions
Do anguillid eels communicate using chemical cues
Eels have a well developed olfactory rosette (Schulte 1972 Yamamotoamp Ueda 1978 Ostrander et al 2000) (Figure 2) This organ experiencesdramatic changes during sexual maturation (Pankhurst amp Lythgoe 1983Sorensen amp Pankhurst 1988) which may be an indication of the involve-ment of chemical communication andor adaptation to the marine environ-ment As yet few studies have addressed the uses of chemical senses byeels most to date are behavioural studies showing the chemo-attraction (orrepulsion) to a specific odorant (Table 1) The main aim of these studies wasrelated to the detection of chemical cues by means of olfaction or loss oforientation in anosmic individuals (Hain 1975 Barbin 1998 Barbin et al1998) that suggest that olfaction is important in the control of eel life cycleThe substances described as chemo-attractants for glass and yellow eels in-clude water conditioned by conspecifics (Pesaro et al 1981 Sorensen 1986Briand et al 2002) extracts of mucus (Saglio 1982) bile salts (Sola 1993b)and plant- and animal-conditioned water (Sorensen 1986) In these casesodorants could act as chemical cues involved in migration from estuarine tofreshwater environments In some studies of American (Anguilla rostrata)(Creutzberg 1958 Miles 1968 Kleckner 1983 Barbin 1998 Barbin et al1998) and European eels (Westin 1998) the importance of chemical sensesto find specific sites and co-ordinate spawning in the marine environment wasinvestigated Other studies have shown the use of olfaction in prey capture(Carton amp Montgomery 2003) which may be related to the high sensitivityto amino acids in the first stages of growth (Silver 1982 Crnjar et al 1992Sola 1993a Sola amp Tongiorgi 1998)
Recently some studies have suggested a role for chemical signals duringreproduction Culture density in the first stages of glass eel culture affectsgrowth and sex differentiation during the later yellow eel phase (Huertas ampCerda 2006) (Figure 3) How is this effect mediated It was suggested thatsocial interaction is an important factor during growth and sexual differen-tiation and probably this social interaction is mainly mediated by chemicalcues rather than visual or tactile signals (Huertas et al 2006) Consequently
Chemical communication in eels 1395Ta
ble
1C
hem
ical
com
mun
icat
ion
stud
ies
inee
lspe
cies
Spec
ies
Odo
rant
Stag
eE
ffec
tR
efer
ence
Aa
ngui
lla
Wat
erco
nditi
oned
bygl
ass
eel
Gla
ssndashe
lver
Che
mo
attr
actio
nPe
saro
etal
(19
81)
Ar
ostr
atta
Wat
erco
nditi
oned
bym
atur
efe
mal
eM
atur
em
ale
Spaw
ning
beha
vior
Sore
nsen
ampW
inn
(198
4)A
ang
uill
aW
ater
cond
ition
edby
undi
ffer
entia
ted
eel
Elv
erC
hem
oat
trac
tion
(eel
trap
)B
rian
det
al(
2002
)A
ang
uill
aW
ater
cond
ition
edby
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
sex
ual
Hue
rtas
etal
(20
06)
mat
ure
eels
pher
omon
eA
ros
trat
aC
hem
ical
cues
rele
ased
bym
atur
eee
lsM
atur
eC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Kle
ckne
r(1
983)
Aa
ngui
lla
Che
mic
alcu
esin
tidal
wat
ers
Elv
erC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Cre
utzb
erg
(195
8)A
ros
trat
aC
hem
ical
cues
infr
eshw
ater
sG
lass
eel
Che
mo
attr
actio
n(s
igna
lmig
ratio
n)M
iles
(196
8)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tG
lass
eel
Che
mo
attr
actio
nTo
si(1
990)
Ar
ostr
ata
Che
mic
alcu
esin
tidal
wat
ers
Yel
low
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ros
trat
aC
hem
ical
cues
intid
alw
ater
sSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)W
estin
(199
8)A
ros
trat
aR
inse
plan
tsa
nim
als
inan
imat
eob
ject
sG
lass
eel
Che
mo
attr
actio
nSo
rens
en(1
986)
Aa
ngui
lla
Geo
smin
(inl
and
wat
erod
our)
Gla
ssee
lC
hem
oat
trac
tion
Tosi
(199
3)A
ang
uill
aM
ucus
bile
int
estin
alflu
idof
Yel
low
eel
Che
mo
attr
actio
n(o
nly
muc
us)
Sagl
io(1
982)
undi
ffer
entia
ted
eels
Aa
ngui
lla
Bile
and
muc
usof
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
Hue
rtas
etal
(20
07)
mat
ure
eels
Aa
ngui
lla
Bile
salts
and
taur
ine
Gla
ssee
lC
hem
oat
trac
tion
(thr
esho
ld10
minus10M
)So
la(1
993b
)A
ang
uill
aA
min
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
Sola
(199
3a)
Aa
ngui
lla
Non
-pro
tein
amin
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
(thr
esho
ld10
minus9M
)So
laamp
Tong
iron
i(19
98)
Aa
ngui
lla
Am
ino
acid
sY
elow
eel
Olf
acto
ryse
nsiti
vity
Crn
jar
etal
(19
92)
Ar
ostr
ata
Am
ino
acid
sSi
lver
eel
Olf
acto
ryse
nsiti
vity
Silv
er(1
982)
Ar
eind
hard
tii
Food
Silv
eree
lC
hem
oat
trac
tion
Car
ton
etal
(20
03)
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

1394 Huertas Canaacuterio amp Hubbard
industry or in strategies for the management or re-stocking of wild popula-tions
Do anguillid eels communicate using chemical cues
Eels have a well developed olfactory rosette (Schulte 1972 Yamamotoamp Ueda 1978 Ostrander et al 2000) (Figure 2) This organ experiencesdramatic changes during sexual maturation (Pankhurst amp Lythgoe 1983Sorensen amp Pankhurst 1988) which may be an indication of the involve-ment of chemical communication andor adaptation to the marine environ-ment As yet few studies have addressed the uses of chemical senses byeels most to date are behavioural studies showing the chemo-attraction (orrepulsion) to a specific odorant (Table 1) The main aim of these studies wasrelated to the detection of chemical cues by means of olfaction or loss oforientation in anosmic individuals (Hain 1975 Barbin 1998 Barbin et al1998) that suggest that olfaction is important in the control of eel life cycleThe substances described as chemo-attractants for glass and yellow eels in-clude water conditioned by conspecifics (Pesaro et al 1981 Sorensen 1986Briand et al 2002) extracts of mucus (Saglio 1982) bile salts (Sola 1993b)and plant- and animal-conditioned water (Sorensen 1986) In these casesodorants could act as chemical cues involved in migration from estuarine tofreshwater environments In some studies of American (Anguilla rostrata)(Creutzberg 1958 Miles 1968 Kleckner 1983 Barbin 1998 Barbin et al1998) and European eels (Westin 1998) the importance of chemical sensesto find specific sites and co-ordinate spawning in the marine environment wasinvestigated Other studies have shown the use of olfaction in prey capture(Carton amp Montgomery 2003) which may be related to the high sensitivityto amino acids in the first stages of growth (Silver 1982 Crnjar et al 1992Sola 1993a Sola amp Tongiorgi 1998)
Recently some studies have suggested a role for chemical signals duringreproduction Culture density in the first stages of glass eel culture affectsgrowth and sex differentiation during the later yellow eel phase (Huertas ampCerda 2006) (Figure 3) How is this effect mediated It was suggested thatsocial interaction is an important factor during growth and sexual differen-tiation and probably this social interaction is mainly mediated by chemicalcues rather than visual or tactile signals (Huertas et al 2006) Consequently
Chemical communication in eels 1395Ta
ble
1C
hem
ical
com
mun
icat
ion
stud
ies
inee
lspe
cies
Spec
ies
Odo
rant
Stag
eE
ffec
tR
efer
ence
Aa
ngui
lla
Wat
erco
nditi
oned
bygl
ass
eel
Gla
ssndashe
lver
Che
mo
attr
actio
nPe
saro
etal
(19
81)
Ar
ostr
atta
Wat
erco
nditi
oned
bym
atur
efe
mal
eM
atur
em
ale
Spaw
ning
beha
vior
Sore
nsen
ampW
inn
(198
4)A
ang
uill
aW
ater
cond
ition
edby
undi
ffer
entia
ted
eel
Elv
erC
hem
oat
trac
tion
(eel
trap
)B
rian
det
al(
2002
)A
ang
uill
aW
ater
cond
ition
edby
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
sex
ual
Hue
rtas
etal
(20
06)
mat
ure
eels
pher
omon
eA
ros
trat
aC
hem
ical
cues
rele
ased
bym
atur
eee
lsM
atur
eC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Kle
ckne
r(1
983)
Aa
ngui
lla
Che
mic
alcu
esin
tidal
wat
ers
Elv
erC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Cre
utzb
erg
(195
8)A
ros
trat
aC
hem
ical
cues
infr
eshw
ater
sG
lass
eel
Che
mo
attr
actio
n(s
igna
lmig
ratio
n)M
iles
(196
8)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tG
lass
eel
Che
mo
attr
actio
nTo
si(1
990)
Ar
ostr
ata
Che
mic
alcu
esin
tidal
wat
ers
Yel
low
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ros
trat
aC
hem
ical
cues
intid
alw
ater
sSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)W
estin
(199
8)A
ros
trat
aR
inse
plan
tsa
nim
als
inan
imat
eob
ject
sG
lass
eel
Che
mo
attr
actio
nSo
rens
en(1
986)
Aa
ngui
lla
Geo
smin
(inl
and
wat
erod
our)
Gla
ssee
lC
hem
oat
trac
tion
Tosi
(199
3)A
ang
uill
aM
ucus
bile
int
estin
alflu
idof
Yel
low
eel
Che
mo
attr
actio
n(o
nly
muc
us)
Sagl
io(1
982)
undi
ffer
entia
ted
eels
Aa
ngui
lla
Bile
and
muc
usof
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
Hue
rtas
etal
(20
07)
mat
ure
eels
Aa
ngui
lla
Bile
salts
and
taur
ine
Gla
ssee
lC
hem
oat
trac
tion
(thr
esho
ld10
minus10M
)So
la(1
993b
)A
ang
uill
aA
min
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
Sola
(199
3a)
Aa
ngui
lla
Non
-pro
tein
amin
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
(thr
esho
ld10
minus9M
)So
laamp
Tong
iron
i(19
98)
Aa
ngui
lla
Am
ino
acid
sY
elow
eel
Olf
acto
ryse
nsiti
vity
Crn
jar
etal
(19
92)
Ar
ostr
ata
Am
ino
acid
sSi
lver
eel
Olf
acto
ryse
nsiti
vity
Silv
er(1
982)
Ar
eind
hard
tii
Food
Silv
eree
lC
hem
oat
trac
tion
Car
ton
etal
(20
03)
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171
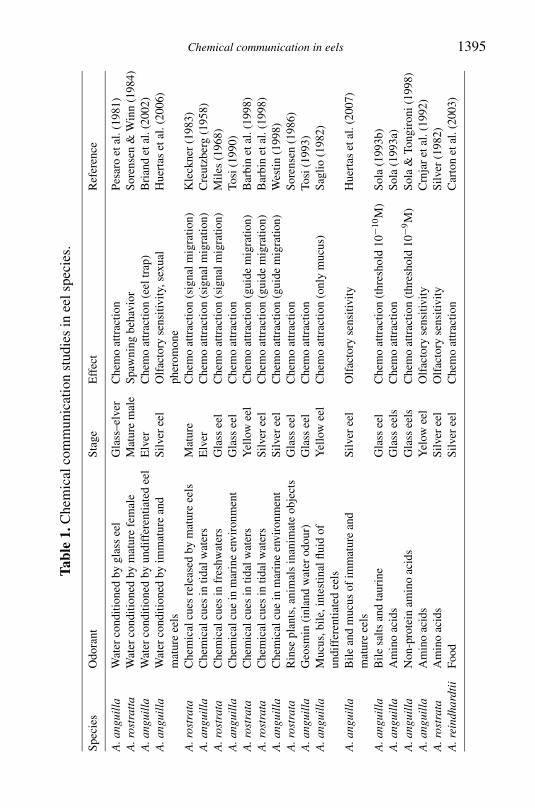
Chemical communication in eels 1395Ta
ble
1C
hem
ical
com
mun
icat
ion
stud
ies
inee
lspe
cies
Spec
ies
Odo
rant
Stag
eE
ffec
tR
efer
ence
Aa
ngui
lla
Wat
erco
nditi
oned
bygl
ass
eel
Gla
ssndashe
lver
Che
mo
attr
actio
nPe
saro
etal
(19
81)
Ar
ostr
atta
Wat
erco
nditi
oned
bym
atur
efe
mal
eM
atur
em
ale
Spaw
ning
beha
vior
Sore
nsen
ampW
inn
(198
4)A
ang
uill
aW
ater
cond
ition
edby
undi
ffer
entia
ted
eel
Elv
erC
hem
oat
trac
tion
(eel
trap
)B
rian
det
al(
2002
)A
ang
uill
aW
ater
cond
ition
edby
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
sex
ual
Hue
rtas
etal
(20
06)
mat
ure
eels
pher
omon
eA
ros
trat
aC
hem
ical
cues
rele
ased
bym
atur
eee
lsM
atur
eC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Kle
ckne
r(1
983)
Aa
ngui
lla
Che
mic
alcu
esin
tidal
wat
ers
Elv
erC
hem
oat
trac
tion
(sig
nalm
igra
tion)
Cre
utzb
erg
(195
8)A
ros
trat
aC
hem
ical
cues
infr
eshw
ater
sG
lass
eel
Che
mo
attr
actio
n(s
igna
lmig
ratio
n)M
iles
(196
8)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tG
lass
eel
Che
mo
attr
actio
nTo
si(1
990)
Ar
ostr
ata
Che
mic
alcu
esin
tidal
wat
ers
Yel
low
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ros
trat
aC
hem
ical
cues
intid
alw
ater
sSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)B
arbi
net
al(
1998
)A
ang
uill
aC
hem
ical
cue
inm
arin
een
viro
nmen
tSi
lver
eel
Che
mo
attr
actio
n(g
uide
mig
ratio
n)W
estin
(199
8)A
ros
trat
aR
inse
plan
tsa
nim
als
inan
imat
eob
ject
sG
lass
eel
Che
mo
attr
actio
nSo
rens
en(1
986)
Aa
ngui
lla
Geo
smin
(inl
and
wat
erod
our)
Gla
ssee
lC
hem
oat
trac
tion
Tosi
(199
3)A
ang
uill
aM
ucus
bile
int
estin
alflu
idof
Yel
low
eel
Che
mo
attr
actio
n(o
nly
muc
us)
Sagl
io(1
982)
undi
ffer
entia
ted
eels
Aa
ngui
lla
Bile
and
muc
usof
imm
atur
ean
dSi
lver
eel
Olf
acto
ryse
nsiti
vity
Hue
rtas
etal
(20
07)
mat
ure
eels
Aa
ngui
lla
Bile
salts
and
taur
ine
Gla
ssee
lC
hem
oat
trac
tion
(thr
esho
ld10
minus10M
)So
la(1
993b
)A
ang
uill
aA
min
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
Sola
(199
3a)
Aa
ngui
lla
Non
-pro
tein
amin
oac
ids
Gla
ssee
lsC
hem
oat
trac
tion
(thr
esho
ld10
minus9M
)So
laamp
Tong
iron
i(19
98)
Aa
ngui
lla
Am
ino
acid
sY
elow
eel
Olf
acto
ryse
nsiti
vity
Crn
jar
etal
(19
92)
Ar
ostr
ata
Am
ino
acid
sSi
lver
eel
Olf
acto
ryse
nsiti
vity
Silv
er(1
982)
Ar
eind
hard
tii
Food
Silv
eree
lC
hem
oat
trac
tion
Car
ton
etal
(20
03)
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171
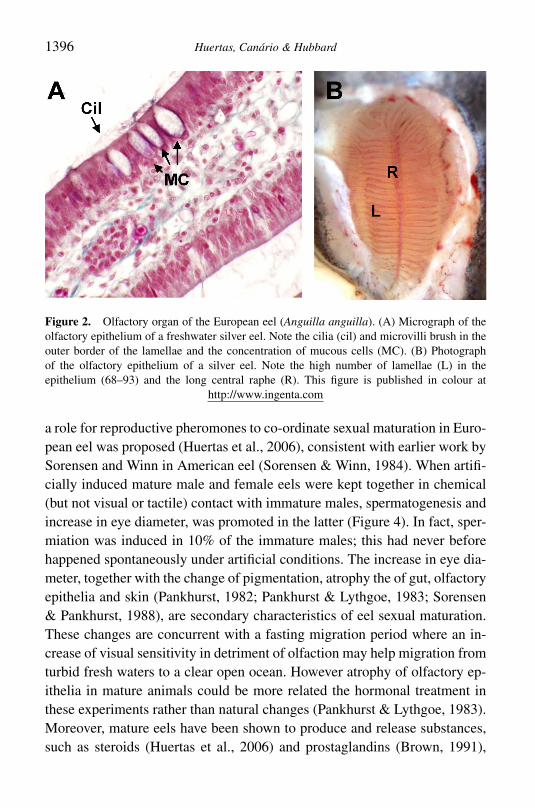
1396 Huertas Canaacuterio amp Hubbard
Figure 2 Olfactory organ of the European eel (Anguilla anguilla) (A) Micrograph of theolfactory epithelium of a freshwater silver eel Note the cilia (cil) and microvilli brush in theouter border of the lamellae and the concentration of mucous cells (MC) (B) Photographof the olfactory epithelium of a silver eel Note the high number of lamellae (L) in theepithelium (68ndash93) and the long central raphe (R) This figure is published in colour at
httpwwwingentacom
a role for reproductive pheromones to co-ordinate sexual maturation in Euro-pean eel was proposed (Huertas et al 2006) consistent with earlier work bySorensen and Winn in American eel (Sorensen amp Winn 1984) When artifi-cially induced mature male and female eels were kept together in chemical(but not visual or tactile) contact with immature males spermatogenesis andincrease in eye diameter was promoted in the latter (Figure 4) In fact sper-miation was induced in 10 of the immature males this had never beforehappened spontaneously under artificial conditions The increase in eye dia-meter together with the change of pigmentation atrophy the of gut olfactoryepithelia and skin (Pankhurst 1982 Pankhurst amp Lythgoe 1983 Sorensenamp Pankhurst 1988) are secondary characteristics of eel sexual maturationThese changes are concurrent with a fasting migration period where an in-crease of visual sensitivity in detriment of olfaction may help migration fromturbid fresh waters to a clear open ocean However atrophy of olfactory ep-ithelia in mature animals could be more related the hormonal treatment inthese experiments rather than natural changes (Pankhurst amp Lythgoe 1983)Moreover mature eels have been shown to produce and release substancessuch as steroids (Huertas et al 2006) and prostaglandins (Brown 1991)
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171
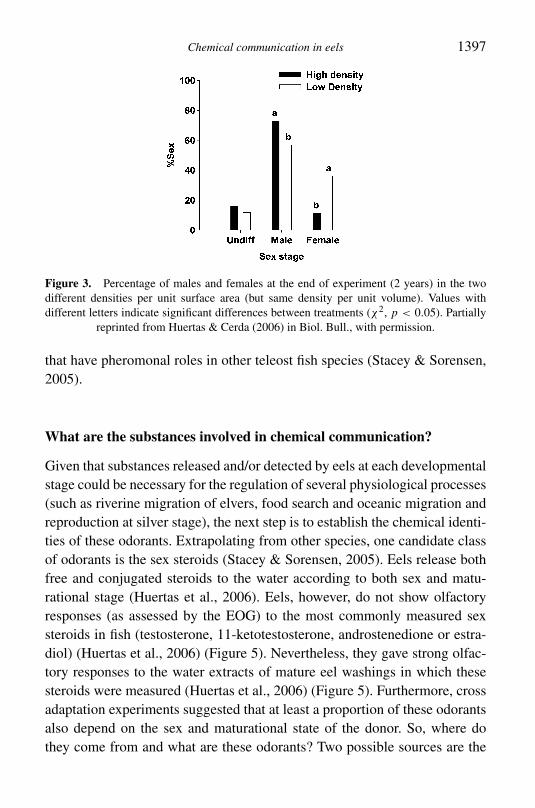
Chemical communication in eels 1397
Figure 3 Percentage of males and females at the end of experiment (2 years) in the twodifferent densities per unit surface area (but same density per unit volume) Values withdifferent letters indicate significant differences between treatments (χ2 p lt 005) Partially
reprinted from Huertas amp Cerda (2006) in Biol Bull with permission
that have pheromonal roles in other teleost fish species (Stacey amp Sorensen2005)
What are the substances involved in chemical communication
Given that substances released andor detected by eels at each developmentalstage could be necessary for the regulation of several physiological processes(such as riverine migration of elvers food search and oceanic migration andreproduction at silver stage) the next step is to establish the chemical identi-ties of these odorants Extrapolating from other species one candidate classof odorants is the sex steroids (Stacey amp Sorensen 2005) Eels release bothfree and conjugated steroids to the water according to both sex and matu-rational stage (Huertas et al 2006) Eels however do not show olfactoryresponses (as assessed by the EOG) to the most commonly measured sexsteroids in fish (testosterone 11-ketotestosterone androstenedione or estra-diol) (Huertas et al 2006) (Figure 5) Nevertheless they gave strong olfac-tory responses to the water extracts of mature eel washings in which thesesteroids were measured (Huertas et al 2006) (Figure 5) Furthermore crossadaptation experiments suggested that at least a proportion of these odorantsalso depend on the sex and maturational state of the donor So where dothey come from and what are these odorants Two possible sources are the
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171
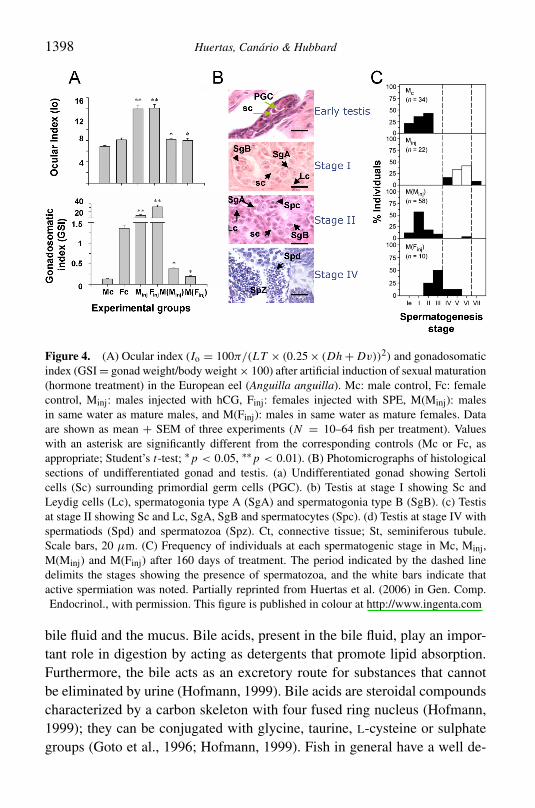
1398 Huertas Canaacuterio amp Hubbard
Figure 4 (A) Ocular index (Io = 100π(LT times (025 times (Dh + Dv))2) and gonadosomaticindex (GSI = gonad weightbody weight times 100) after artificial induction of sexual maturation(hormone treatment) in the European eel (Anguilla anguilla) Mc male control Fc femalecontrol Minj males injected with hCG Finj females injected with SPE M(Minj) malesin same water as mature males and M(Finj) males in same water as mature females Dataare shown as mean + SEM of three experiments (N = 10ndash64 fish per treatment) Valueswith an asterisk are significantly different from the corresponding controls (Mc or Fc asappropriate Studentrsquos t-test lowastp lt 005 lowastlowastp lt 001) (B) Photomicrographs of histologicalsections of undifferentiated gonad and testis (a) Undifferentiated gonad showing Sertolicells (Sc) surrounding primordial germ cells (PGC) (b) Testis at stage I showing Sc andLeydig cells (Lc) spermatogonia type A (SgA) and spermatogonia type B (SgB) (c) Testisat stage II showing Sc and Lc SgA SgB and spermatocytes (Spc) (d) Testis at stage IV withspermatiods (Spd) and spermatozoa (Spz) Ct connective tissue St seminiferous tubuleScale bars 20 μm (C) Frequency of individuals at each spermatogenic stage in Mc MinjM(Minj) and M(Finj) after 160 days of treatment The period indicated by the dashed linedelimits the stages showing the presence of spermatozoa and the white bars indicate thatactive spermiation was noted Partially reprinted from Huertas et al (2006) in Gen CompEndocrinol with permission This figure is published in colour at httpwwwingentacom
bile fluid and the mucus Bile acids present in the bile fluid play an impor-tant role in digestion by acting as detergents that promote lipid absorptionFurthermore the bile acts as an excretory route for substances that cannotbe eliminated by urine (Hofmann 1999) Bile acids are steroidal compoundscharacterized by a carbon skeleton with four fused ring nucleus (Hofmann1999) they can be conjugated with glycine taurine L-cysteine or sulphategroups (Goto et al 1996 Hofmann 1999) Fish in general have a well de-
Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

Chemical communication in eels 1399
veloped olfactory sensitivity to bile acids (Doslashving et al 1980 Hara 1994Michel amp Lubomudrov 1995 Zhang et al 2001 Siefkes amp Li 2004) butwith the exception of the sea lamprey their roles if any in relation to olfac-tion are not yet clear Moreover in modern mammals bile acids are mostlyC24 structures whereas in primitive mammals cartilaginous fish and reptilesC27 forms are often found (Haslewood 1967) Bile acids are normally ex-creted into the environment via the faeces and recent evidence has shownthat in the sea lamprey (Petromyzon marinus L) specific bile acids activelyreleased by mature males through the gills act as powerful attractants for ma-ture females (Li et al 2002) and the adults are attracted to their freshwaterspawning sites by a mixture of steroidal compounds released by conspe-cific larvae (Sorensen et al 2005) Olfactory detection of bile acids in sealamprey is mediated by independent receptors (Li amp Sorensen 1997) Thusdifferent bile acids possibly acting as mixtures could be related to the di-versity in biological responses In the trout conspecific bile fluid has beenshown to have pheromonal activity (Vermeirssen amp Scott 2001) It is possi-ble that bile acids play similar roles in the eel A recent study assessed theolfactory sensitivity of immature European eels to bile fluid from immatureand mature conspecifics of either sex Electro-olfactogram (EOG) studiesshowed that eels have a high olfactory sensitivity to bile fluid (down to di-lutions of 1 107 Figure 6A) Furthermore it was shown that the sensitivityto bile fluid from the eel is dependent not only on the sex but also state ofmaturity of the donor (Huertas et al 2007) Crude fractionation of the bilefluid showed that the active components in the EOG possibly bile acidswere retained by C18 mini-columns and highlighted the bile as the potentialsource of substances for intra-specific sexual communication in Europeaneel (Huertas et al 2007) However it is not yet known whether eels releasedifferent bile acids into the water as a function of physiological stage furtherwork is needed to clarify this
Skin mucus is another possible source of odorants (Stabell amp Selset1980) eels are notoriously slimy fish Indeed conspecific mucus (but notbile fluid) acts as an attractant for yellow eels (Saglio 1982) Furthermoreanecdotal evidence where sexual maturity has been induced in eels by hor-monal treatment suggests that mdash to humans at least mdash mucus from maturemale and female eels smells different and different again from immature fish(Sorensen amp Winn 1984 M Huertas personal observation) High olfactory
1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

1400 Huertas Canaacuterio amp Hubbard
Figure 5 (A) Amplitudes of olfactory responses of immature male European eels to C18solid-phase extracts of water conditioned by control male (water Mc) sexually maturemale (water Minj) and mature female (water Finj) conspecifics by recording of the electro-olfactogram (EOG) Data are shown as means plusmn SEM (N = 6) and are blank-corrected andnormalized to the amplitude of response to 10minus3 M L-asparagine In the middle and bottompanels the periods of spermiation (Sp) and ovulation (Ov) of Minj and Finj respectively areindicated Values with an asterisk denote statistical differences from responses to the waterfrom untreated males (Studentrsquos t test lowastp lt 005 lowastlowastp lt 001 (B) Normalized data of olfac-tory responses to different steroids and PGF2α in sexually immature eels by recording of theEOG The compounds were tested at 10minus6 M and for 1720α-dihydroxypregn-4-en-3-one(1720αP) estradiol-17β (E2) and testosterone (T) and different conjugates were assayedfree (F) sulphated (S) and glucuronidated (G) As a positive control the olfactory responseto L-glutamine (Gln) 10minus3 M is included Data are the means + SEM (N = 2ndash5 eels) andare blank corrected and normalized to the amplitude of response to 10minus3 M L-asparagine
Partially reprinted from Huertas et al (2007) in J Fish Biol with permission
sensitivity to conspecific mucus has been demonstrated in eels with thresh-olds of detection around 1 106 (Huertas et al 2007) Mucus contained ahigher fraction of odorants that were able to pass through C18 solid-phaseextraction cartridges than bile fluid (sex steroids and bile acids are likely tobe trapped whereas amino acids are likely to pass through) However mucusfrom mature eels had a higher proportion of olfactory activity in the eluatethan immature fish Cross-adaptation experiments suggest again that thereare qualitative differences in the odorants contained in both bile and mucus
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171
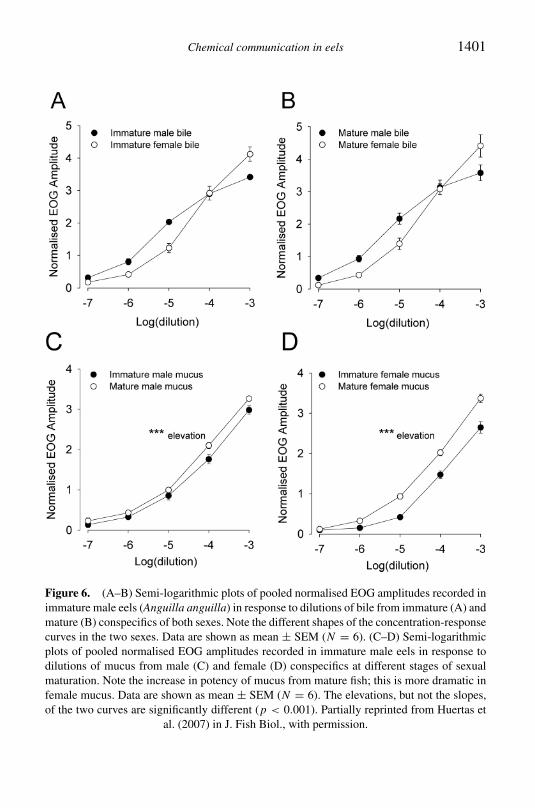
Chemical communication in eels 1401
Figure 6 (AndashB) Semi-logarithmic plots of pooled normalised EOG amplitudes recorded inimmature male eels (Anguilla anguilla) in response to dilutions of bile from immature (A) andmature (B) conspecifics of both sexes Note the different shapes of the concentration-responsecurves in the two sexes Data are shown as mean plusmn SEM (N = 6) (CndashD) Semi-logarithmicplots of pooled normalised EOG amplitudes recorded in immature male eels in response todilutions of mucus from male (C) and female (D) conspecifics at different stages of sexualmaturation Note the increase in potency of mucus from mature fish this is more dramatic infemale mucus Data are shown as mean plusmn SEM (N = 6) The elevations but not the slopesof the two curves are significantly different (p lt 0001) Partially reprinted from Huertas et
al (2007) in J Fish Biol with permission
1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

1402 Huertas Canaacuterio amp Hubbard
depending on both the sex and state of sexual maturation of the donor Theseresults are consistent with a role for chemical communication in the repro-duction of the eel and suggest that both bile and mucus are potential sourcesof the odorants involved However other sources such as the urine cannotbe excluded
Conclusions and future prospects
Several salient features of the European eel recommend it as a model speciesfor studies in fish olfaction and chemical communication Firstly it belongsto an ancient lsquostemrsquo teleost group the Elopomorpha and may therefore helpto understand the evolution of olfaction in fish Secondly the high physiolog-ical plasticity of eels permits a comparison of the olfactory processes underseveral conditions for example marine vs freshwater individuals sexuallymature vs immature growing vs fasting It is known that eels release sub-stances that are detected by conspecifics wherein they trigger physiologicaland possibly behavioural responses This suggests that eels are using chem-ical communication to co-ordinate important events such as reproductionmigration and nutrition Immature male eels have high olfactory sensitiv-ity to conspecific bile and mucus both being possible vehicles for releaseof substances involved in chemical communication The chemical nature ofthese odorants is unknown but bile acids remain possible candidates It isalso clear that eels can detect bile acids from hetero-specifics these odorantscould also be involved in interaction between species (eg predator or preydetection)
Much work remains to be done before the precise roles of chemical com-munication and the identity of the odorants involved are clearly definedfor eels Little is known for example about differences in olfactory sen-sitivity between the sexes or whether sexual maturation andor adaptationto seawater prior to migration affect the olfactory system Further studiesneed to be directed to the chemical identification of the odorants involvedAlso the cellular and molecular mechanisms of odorant detection and signaltransduction are still unknown how do these change with sexual maturationor adaptation to seawater during migration Finally the physiological andbehavioural responses related to chemical communication are poorly under-stood do females for example use olfactory cues to find the males in the
Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

Chemical communication in eels 1403
vast expanse of the Sargasso Sea What is the relevance of olfaction duringmigration compared to other senses such as vision and sensitivity to mag-netic fields There is a sense of urgency for this kind of studies which couldhelp protect a species which complex biology has been a hot topic of debatefor the last three decades If such studies are not undertaken soon then theopportunity may be lost Permanently
Acknowledgements
The work in the authorsrsquo laboratory was funded by the Portuguese National ScienceFoundation and European Social Funds projects POCTIBSE388152001 and POCIBIA-BDE554632004 and fellowship SFRHBPD263392005 to MH
References
Albert V Jonsson B amp Bernatchez L (2006) Natural hybrids in Atlantic eels (Anguillaanguilla A-rostrata) evidence for successful reproduction and fluctuating abundancein space and time mdash Mol Ecol 15 1903-1916
Aoyama J amp Tsukamoto K (1997) Evolution of the freshwater eels mdash Naturwis-senschaften 84 17-21
Aroua S Schmitz M Baloche S Vidal B Rousseau K amp Dufour S (2005) Endocrineevidence that silvering a secondary metamorphosis in the eel is a pubertal rather thana metamorphic event mdash Neuroendocrinology 82 221-232
Asturiano JF Peacuterez L Olivares L Tomaacutes A Martinez S Falco S amp Jover M (2002)I Congreso Iberoamericano Virtual de Acuicultura
Barbin GP (1998) The role of olfaction in homing and estuarine migratory behavior ofyellow-phase American eels mdash Can J Fish Aquat Sci 55 564-575
Barbin GP Parker SJ amp McCleave JD (1998) Olfactory cues play a critical role in theestuarine migration of silver-phase American eels mdash Environ Biol Fish 53 283-291
Briand C Fatin D Fontenelle G amp Feunteun E (2003) Estuarine and fluvial recruitmentof the European glass eel Anguilla anguilla in an exploited Atlantic estuary mdash FishManag Ecol 10 377-384
Briand C Fatin D amp Legault A (2002) Role of eel odour on the efficiency of an eelAnguilla anguilla ladder and trap mdash Environ Biol Fish 65 473-477
Brown JA Gray CJ Hattersley G amp Robinson J (1991) Prostaglandins in the kidneyurinary bladder and gills of the rainbow trout and European eel adapted to fresh waterand seawater mdash Gen Comp Endocrinol 84 328-335
Carton AG amp Montgomery JC (2003) Evidence of a rheotactic component in the odoursearch behaviour of freshwater eels mdash J Fish Biol 62 501-516
Creutzberg F (1958) Use of tidal streams by migrating elvers (Anguilla-Vulgaris-Turt) mdashNature 181 857-858
Crnjar R Scalera G Bigiani A Barbarossa IT Magherini PC amp Pietra P (1992)Olfactory sensitivity to amino acids in the juvenile stages of the European eel Anguillaanguilla (L) mdash J Fish Biol 40 567-576
1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

1404 Huertas Canaacuterio amp Hubbard
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus1758) mdash In FAO Fish Synop p 73
Dekker W (2003) Did lack of spawners cause the collapse of the European eel Anguillaanguilla mdash Fish Manag Ecol 10 365-376
Doslashving KB Selset R amp Thommensen G (1980) Olfactory sensitivity to bile acids insalmonid fishes mdash Acta Physiol Scand 108 123-131
Dufour S (2000) Puberty in teleosts new insights into the role of peripheral signals in thestimulation of pituitary gonadotropins mdash Reprod Physiol Fish 82 61-74
Elie P Lecomte-Finiger R Cantrelle I amp Charlon N (1982) Deacutefinition des limites desdiffeacuterents stades pigmentaires durant la phase civelle drsquoAnguilla anguilla L (Poissonteacuteleacuteosteacuteen anguilliforme) mdash Vie Milieu 32 149-157
Furuita H Unuma T Nomura K Tanaka H Okuzawa K Sugita T amp Yamamoto T(2006) Lipid and fatty acid composition of eggs producing larvae with high survivalrate in the Japanese eel mdash J Fish Biol 69 1178-1189
van Ginneken V Antonissen E Muumlller UK Booms R Eding E Verreth J amp van denThillart G (2005) Eel migration to the Sargasso remarkably high swimming efficiencyand low energy costs mdash J Exp Biol 208 1329-1335
van Ginneken V Durif C Dufour S Sbaihi M Boot R Noorlander K Doornbos JMurk AJ amp van den Thillart G (2007) Endocrine profiles during silvering of theEuropean eel (Anguilla anguilla L) living in saltwater mdash Anim Biol 57 453-465
van Ginneken V amp Maes GE (2005) The European eel (Anguilla anguilla Linnaeus) itslifecycle evolution and reproduction a literature review mdash Rev Fish Biol Fish 15367-398
Goto T Ui T Une M Kuramoto T Kihira K amp Hoshita T (1996) Bile salt compositionand distribution of the D-Cysteinolic acid conjugated bile salts in fish mdash Fish Sci 62606-609
Gousset B (1990) European eel (Anguilla anguilla L) farming technologies in Europe andin Japan application of a comparative analysis mdash Aquaculture 87 209-235
Gousset B (1992) Eel culture in Japan mdash Bull Inst Oceacuteanogr (Monaco) (No Speacutec) 101-128
Hain JHW (1975) Behavior of migratory eels Anguilla-Rostrata in response to currentsalinity and lunar period mdash Helgol Wiss Meeresunters 27 211-233
Hara TJ (1994) Olfaction and gustation in fish mdash an overview mdash Acta Physiol Scand152 207-217
Haraldstad O Vollestad LA amp Jonsson B (1985) Descent of European silver eels An-guilla anguilla L in a Norwegian Watercourse mdash J Fish Biol 26 37-41
Haslewood GAD (1967) Bile salt evolution mdash J Lipid Res 8 535-550Heinsbroek L (1991) A review of eel culture in Japan and Europe mdash Aquacult Fish
Manage 22 57-72Hofmann AF (1999) Bile acids the good the bad and the ugly mdash News Physiol Sci 14
24-29Huertas M amp Cerda J (2006) Stocking density at early developmental stages affects growth
and sex ratio in the European eel (Anguilla anguilla) mdash Biol Bull 211 286-296Huertas M Hubbard PC Canario AVM amp Cerda J (2007) Olfactory sensitivity to
conspecific bile fluid and skin mucus in the European eel Anguilla anguilla (L) mdash JFish Biol 70 1907-1920
Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

Chemical communication in eels 1405
Huertas M Scott AP Hubbard PC Canario AVM amp Cerda J (2006) Sexually matureEuropean eels (Anguilla anguilla L) stimulate gonadal development of neighbouringmales possible involvement of chemical communication mdash Gen Comp Endocrinol147 304-313
ICES (2005) International Council for the Exploration of the Sea Report of the ICESEIFACWorking Group on Eels mdash In ICES CM 2005ACFM 01 Galway p 71
Ide LM Urbinati EC amp Hoffmann A (2003) The role of olfaction in the behaviouraland physiological responses to conspecific skin extract in Brycon cephalus mdash J FishBiol 63 332-343
Ijiri S Kayaba T Takeda N Tachiki H Adachi S amp Yamauchi K (1998) Pretreatmentreproductive stage and oocyte development induced by salmon pituitary homogenate inthe Japanese eel Anguilla japonica mdash Fish-Sci 64 531-537
Jellyman D amp Tsukamoto K (2005) Swimming depths of offshore migrating longfin eelsAnguilla dieffenbachii mdash Mar Ecol Prog Ser 286 261-267
Kagawa H Tanaka H Otha H Okuzawa K amp Iinuma N (1997) Induced ovulation byinjection of 1720β-dihydroxy-4-pregen-3-one in the artificially matured Japanese eelwith special reference to ovulation time mdash Fish Sci 63 365-367
Kleckner R (1983) Spawning of American eel Anguilla rostrata relative to thermal frontsin the Sargasso Sea mdash Environ Biol Fishes 9 289-293
Kristensen EA amp Closs GP (2004) Anti-predator response of naiumlve and experiencedcommon bully to chemical alarm cues mdash J Fish Biol 64 643-652
Lecompte-Finiger R (1994) Les stades larvaires (leptoceacutephales et anguilles) de lrsquoanguilleeuropeacuteenne migrations et meacutetamorphose mdash Annee Biol 33 1-17
Lecompte-Finiger R Maunier C amp Khafif M (2004) Les larves leptoceacutephales ces meacute-connues mdash Cybium 28 83-95
Li W amp Sorensen PW (1997) Highly independent olfactory receptor sites for naturallyoccurring bile acids in the sea lamprey Petromyzon marinus mdash J Comp Physiol ASens Neural Behav Physiol 180 429-438
Li WM Scott AP Siefkes MJ Yan HG Liu Q Yun SS amp Gage DA (2002)Bile acid secreted by male sea lamprey that acts as a sex pheromone mdash Science 296138-141
Lin H-R Xie G Zhang L-H Wang X-D amp Chen L-X (1998) Artificial induction ofgonadal maturation and ovulation in the Japanese eel (Anguilla japonica T et S) mdashBull Fr Peche Piscic 349 163-176
McCleave JD amp Arnold GP (1999) Movements of yellow- and silver-phase Europeaneels (Anguilla anguilla L) tracked in the western North Sea mdash ICES J Marine Sci 56510-536
McCleave JD Brickley PJ OrsquoBrien KM Kistner DA Wong MW Gallagher M ampWatson SM (1998) Do leptocephali of the European eel swim to reach continentalwaters Status of the question mdash J Mar Biol Ass UK 78 285-306
Michel WC amp Lubomudrov LM (1995) Specificity and sensitivity of the olfactory organof the zebrafish Danio rerio mdash J Comp Physiol A Sens Neural Behav Physiol 177191-199
Miles SG (1968) Rheotaxis of elvers of American eel (Anguilla rostrata) in laboratory towater from different streams in Nova Scotia mdash J Fish Res Board Can 25 1591-
Nelson JS (2006) Fishes of the world 4th edn mdash Wiley Hoboken NJ
1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

1406 Huertas Canaacuterio amp Hubbard
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose K (1996a) Milt production in theJapanese eel Anguilla japonica induced by repeated injections of human gonadotropinmdash Fish Sci 62 44-49
Ohta H Kagawa H Tanaka H Okuzawa K amp Hirose KU (1996b) Changes in fertil-ization and hatching rates with time after ovulation induced by 17 20β-dihydroxy-4-pregnen-3-one in the Japanese eel Anguilla japonica mdash Aquaculture 139 291-301
Ohta H Kagawa H Tanaka H Okuzawa K Iinuma N amp Hirose K (1997) Artificialinduction of maturation and fertilization in the Japanese eel Anguilla japonica mdash FishPhysiol Biochem 17 163-169
Ostrander G Bullock G amp Bunton T (2000) The laboratory fish mdash Elsevier San DiegoCA
Pankhurst NW (1982) Relation of visual changes to the onset of sexual maturation in theEuropean eel Anguilla anguilla (L) mdash J Fish Biol 21 127-140
Pankhurst NW amp Lythgoe J (1983) Changes in vision and olfaction during sexual matu-ration in the European eel Anguilla anguilla (L) mdash J Fish Biol 23 229-240
Pedersen BH (2003) Induced sexual maturation of the European eel Anguilla anguilla andfertilization of the eggs mdash Aquaculture 224 323-338
Pedersen BH (2004) Fertilization of eggs rate of embryonic development and hatchingfollowing induced maturation of the European eel Anguilla anguilla mdash Aquaculture237 461-473
Peacuterez L Barrera R Asturiano JF amp Jover M (2004) Produccioacuten de anguilas pasadopresente y futuro mdash Revista Aquatic 20 51-78
Pesaro M Balsamo M Gandolfi G amp Tongiorgi P (1981) Discrimination among dif-ferent kinds of water in juvenile eels Anguilla anguilla (L) mdash Monit Zool Ital 15183-191
Polkinghorne CN Olson JM Gallaher DG amp Sorensen PW (2001) Larval sea lampreyrelease two unique bile acids to the water at a rate sufficient to produce detectableriverine pheromone plumes mdash Fish Physiol Biochem 24 15-30
Rosenthal GG amp Lobel PS (2006) Communication mdash In Behaviour and physiology offish (Sloman KA Wilson RW amp Balshine S eds) Elsevier San Diego CA p39-78
Saglio P (1982) Pieacutegeage drsquoanguilles (Anguilla anguilla L) dans le milieu naturel au moyendrsquoextraits biologiques drsquoorigine intraspeacutecifique Mise en eacutevidence de lrsquoattractiviteacutepheacuteromonale du mucus eacutepidemique mdash Acta Ecol 3 223-231
Schulte E (1972) Studies of the regio olfactoria of the eel Anguilla anguilla L I Finestructure of the olfactory epithelium mdash Z ZellforSch Mikrosk Anat 125 210-228
Selset R amp Doving KB (1980) Behaviour of mature anadromous char (Salmo alpinus L)towards odorants produced by smolts of their own population mdash Acta Physiol Scand2 113-122
Siefkes M amp Li W (2004) Electrophysiological evidence for detection and discriminationof pheromonal bile acids by the olfactory epithelium of female sea lampreys (Petromy-zon marinus) mdash J Comp Physiol A Sens Neural Behav Physiol 190 193-199
Silver W (1982) Electrophysiological responses from the peripheral olfactory system of theAmerican eel Anguilla rostrata mdash J Comp Physiol A Sens Neural Behav Physiol148 379-388
Sola C (1993a) Behavioural responses of glass eels (Anguilla anguilla) towards aminoacids mdash J Fish Biol 42 683-691
Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171

Chemical communication in eels 1407
Sola C (1993b) Bile salts and taurine as chemical stimuli for glass eels Anguilla anguillaa behavioural study mdash Environ Biol Fish 37 197-204
Sola C amp Tongiorgi P (1998) Behavioural responses of glass eels of Anguilla anguilla tonon-protein amino acids mdash J Fish Biol 53 1253-1262
Sorensen PW (1986) Origins of the freshwater attractant(s) of migrating elvers of theAmerican eel Anguilla rostrata mdash Environ Biol Fish 17 185-200
Sorensen PW Fine JM Dvornikovs V Jeffrey CS Shao F Wang JZ Vrieze LAAnderson KR amp Hoye TR (2005) Mixture of new sulphated steroids functions as amigratory pheromone in the sea lamprey mdash Nature Chem Biol 1 324-328
Sorensen PW amp Pankhurst NW (1988) Histological changes in the gonad skin intestineand olfactory epithelium of artificially-matured male American eels Anguilla rostrata(LeSueur) mdash J Fish Biol 32 297-307
Sorensen PW amp Winn HE (1984) The induction of maturation and ovulation in Americaneels Anguilla rostrata (LeSueur) and the relevance of chemical and visual cues to malespawning behaviour mdash J Fish Biol 25 261-268
Stabell OB amp Selset R (1980) Comparison of mucus collecting methods in fish olfactionmdash Acta Physiol Scand 108 91-96
Stacey N amp Sorensen P (2002) Hormonal pheromones in fish mdash In Non mammalianhormone-behaviour systems (Pfaff DW Harnold AP Etgen AM Fahrbach SEamp Rubin RT eds) HancourtAcademic Press London p 375-434
Stacey N amp Sorensen P (2005) Reproductive pheromones mdash In Behaviour and physi-ology of fish (Sloman KA Wilson RW amp Balshine S eds) Academic Press SanDiego CA p 359-412
Starkie A (2003) Management issues relating to the European eel Anguilla anguilla mdashFish Manag Ecol 10 361-364
Tanaka H Kagawa H Ohta H Unuma T amp Nomura K (2003) The first production ofglass eel in captivity fish reproductive physiology facilitates great progress in aquacul-ture mdash Fish Physiol Biochem 28 493-497
Tesch F-W (2003) The eel mdash Blackwell Science OxfordUsui A (1991) Eel culture 2nd edn mdash Blackwell Science LondonVermeirssen ELM amp Scott AP (2001) Male priming pheromone is present in bile as well
as urine of female rainbow trout mdash J Fish Biol 58 1039-1045Westin L (1998) The spawning migration of European silver eel (Anguilla anguilla L) with
particular reference to stocked eel in the Baltic mdash Fish Res 38 257-270Win T (2000) Isolation and structural identification of an alarm pheromone from the gi-
ant danio Danio malabaricus (Cyprinidae Ostiophysi Pisces) mdash Carl-von-OssietzkyUniversitaumlt Oldenburg
Yamamoto M amp Ueda K (1978) Comparative morphology of fish olfactory epithelium 4Anguilliformes and Myctophiformes mdash Bull Jpn Soc Sci Fish 44 1207-1212
Zhang C Brown S amp Hara T (2001) Biochemical and physiological evidence that bileacids produced and released by lake char (Salvelinus namaycush) function as chemicalsignals mdash J Comp Physiol B Biochem Syst Environ Physiol 171 161-171













![Anguilla anguilla L. Biochemical and Genotoxic Responses to Benzo[ a]pyrene](https://static.fdokumen.com/doc/165x107/631d4597f26ecf94330a787a/anguilla-anguilla-l-biochemical-and-genotoxic-responses-to-benzo-apyrene.jpg)






