
Enzymatic and nonenzymatic antioxidants as an adaptation to phagocyte-induced damage in Anguilla...
13
Ecotoxicology and Environmental Safety 57 (2004) 290–302 Enzymatic and nonenzymatic antioxidants as an adaptation to phagocyte-induced damage in Anguilla anguilla L. following in situ harbor water exposure Iqbal Ahmad, M. Pacheco, and M.A. Santos Animal Physiology/Ecotoxicology Sector, Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal Received 1 August 2002; received in revised form 18 March 2003; accepted 28 April 2003 Abstract Anguilla anguilla L. were caged for 8 and 48 h in harbor water of Aveiro Lagoon, Portugal. Respiratory burst activity (RBA) of peritoneal, head kidney, and gill phagocytes was measured. Lipid peroxidation (LPO) was estimated in gill, kidney, and liver. Liver ethoxyresorufin-O-deethylase (EROD) activity, cytochrome P450 (Cyt P450) content, and bile metabolites were assayed. Various antioxidant enzymes, viz., glutathione peroxidase, catalase, and glutathione S-transferase and nonenzymatic antioxidant, viz., total reduced glutathione were also studied. Harbor water xenobiotics induced a significant RBA increase in gill after 8 h; whereas in peritoneum and head kidney it increased after 48 h exposure. These responses were adversely associated with tissue-specific peroxidative damage since significant LPO increase was observed in gill (8 and 48 h), kidney (48 h), and liver (48 h). The tissue most affected was gill. Moreover, liver EROD activity, Cyt P450 content and bile metabolites remain unaltered after 8 h; in contrast, 48 h exposure showed significant EROD activity decrease and pyrene-type bile metabolites increase. Decreased EROD activity may be associated with concomitant liver damage, as increased LPO was observed after 48 h. Furthermore, the tissue-specific damage corresponded to the differences in the antioxidant potentials of the tissues, since the initial exposure period caused a significant increase in liver antioxidant activities, whereas gill and kidney showed a significant decrease, demonstrating that liver is highly adaptive to oxidative damage. However, at 48 h exposure gill, kidney, and liver showed a suppressive antioxidant effect, probably due to PAHs, since a significant induction at PAH-type bile metabolites has been seen. Our results demonstrate that phagocyte activation and associated peroxidative damage are concomitantly corroborated with enzymatic and nonenzymatic antioxidant activity differences. In addition, hepatic antioxidant induction after short-term exposure may serve as a potent biomarker for water pollutants in fish. r 2003 Elsevier Inc. All rights reserved. Keywords: Anguilla anguilla L.; Phagocytes; Respiratory burst activity; Harbor water contamination; Peroxidative damage; Antioxidants 1. Introduction An increasing variety of industrial, agricultural and other chemicals are entering the aquatic environment and being taken up into the tissues of invertebrates and vertebrates (Ahmad et al., 2000; Khan et al., 1995; Walker and Livingstone, 1992; Walsh and O’Halloran, 1988). The uptake of these contaminants can occur from sediments, suspended particulate materials, water col- umn and food sources. The major route of input depends on the particular dietary and ecological life styles of the organisms (Livingstone, 1991, 1998; Van Veld, 1990). Such a diverse array of chemicals can have many different mechanisms of toxicity and several mechanisms may exist for a single contaminant, each contributing to various degrees of the final overall deleterious effects (Livingstone, 1991; Livingstone et al., 2000; Walker et al., 1996). The cytotoxic reactive oxygen species (ROS) are continuously produced by contami- nant-stimulated phagocytes (Ahmad et al., 1998; Fatima et al., 2000) and excessive ROS have shown their role in the development of local tissue damage (Ahmad et al., 2003; Fatima et al., 2000). However, there is a lack of information on the defense mechanisms that neutralize the impact of ROS on fish. The exclusive emphasis of previous research work has been centered on ROS deleterious capacity for biological macromolecules ARTICLE IN PRESS Corresponding author. Fax: +351-234426408. E-mail address: [email protected] (M.A. Santos). 0147-6513/03/$ - see front matter r 2003 Elsevier Inc. All rights reserved. doi:10.1016/S0147-6513(03)00080-0
Transcript of Enzymatic and nonenzymatic antioxidants as an adaptation to phagocyte-induced damage in Anguilla...

Ecotoxicology and Environmental Safety 57 (2004) 290–302
ARTICLE IN PRESS
�Corresp
E-mail a
0147-6513/0
doi:10.1016
Enzymatic and nonenzymatic antioxidants as an adaptationto phagocyte-induced damage in Anguilla anguilla L. following
in situ harbor water exposure
Iqbal Ahmad, M. Pacheco, and M.A. Santos�
Animal Physiology/Ecotoxicology Sector, Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal
Received 1 August 2002; received in revised form 18 March 2003; accepted 28 April 2003
Abstract
Anguilla anguilla L. were caged for 8 and 48 h in harbor water of Aveiro Lagoon, Portugal. Respiratory burst activity (RBA) of
peritoneal, head kidney, and gill phagocytes was measured. Lipid peroxidation (LPO) was estimated in gill, kidney, and liver. Liver
ethoxyresorufin-O-deethylase (EROD) activity, cytochrome P450 (Cyt P450) content, and bile metabolites were assayed. Various
antioxidant enzymes, viz., glutathione peroxidase, catalase, and glutathione S-transferase and nonenzymatic antioxidant, viz., total
reduced glutathione were also studied. Harbor water xenobiotics induced a significant RBA increase in gill after 8 h; whereas in
peritoneum and head kidney it increased after 48 h exposure. These responses were adversely associated with tissue-specific
peroxidative damage since significant LPO increase was observed in gill (8 and 48 h), kidney (48 h), and liver (48 h). The tissue most
affected was gill. Moreover, liver EROD activity, Cyt P450 content and bile metabolites remain unaltered after 8 h; in contrast, 48 h
exposure showed significant EROD activity decrease and pyrene-type bile metabolites increase. Decreased EROD activity may be
associated with concomitant liver damage, as increased LPO was observed after 48 h. Furthermore, the tissue-specific damage
corresponded to the differences in the antioxidant potentials of the tissues, since the initial exposure period caused a significant
increase in liver antioxidant activities, whereas gill and kidney showed a significant decrease, demonstrating that liver is highly
adaptive to oxidative damage. However, at 48 h exposure gill, kidney, and liver showed a suppressive antioxidant effect, probably
due to PAHs, since a significant induction at PAH-type bile metabolites has been seen. Our results demonstrate that phagocyte
activation and associated peroxidative damage are concomitantly corroborated with enzymatic and nonenzymatic antioxidant
activity differences. In addition, hepatic antioxidant induction after short-term exposure may serve as a potent biomarker for water
pollutants in fish.
r 2003 Elsevier Inc. All rights reserved.
Keywords: Anguilla anguilla L.; Phagocytes; Respiratory burst activity; Harbor water contamination; Peroxidative damage; Antioxidants
1. Introduction
An increasing variety of industrial, agricultural andother chemicals are entering the aquatic environmentand being taken up into the tissues of invertebrates andvertebrates (Ahmad et al., 2000; Khan et al., 1995;Walker and Livingstone, 1992; Walsh and O’Halloran,1988). The uptake of these contaminants can occur fromsediments, suspended particulate materials, water col-umn and food sources. The major route of inputdepends on the particular dietary and ecological lifestyles of the organisms (Livingstone, 1991, 1998; Van
onding author. Fax: +351-234426408.
ddress: [email protected] (M.A. Santos).
3/$ - see front matter r 2003 Elsevier Inc. All rights reserved.
/S0147-6513(03)00080-0
Veld, 1990). Such a diverse array of chemicals can havemany different mechanisms of toxicity and severalmechanisms may exist for a single contaminant, eachcontributing to various degrees of the final overalldeleterious effects (Livingstone, 1991; Livingstone et al.,2000; Walker et al., 1996). The cytotoxic reactive oxygenspecies (ROS) are continuously produced by contami-nant-stimulated phagocytes (Ahmad et al., 1998; Fatimaet al., 2000) and excessive ROS have shown their role inthe development of local tissue damage (Ahmad et al.,2003; Fatima et al., 2000). However, there is a lack ofinformation on the defense mechanisms that neutralizethe impact of ROS on fish. The exclusive emphasis ofprevious research work has been centered on ROSdeleterious capacity for biological macromolecules

ARTICLE IN PRESSI. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302 291
(Girotti, 1990; Stadtman, 1991) and local tissues, wherethey home in (Ahmad et al., 2003; Fatima et al., 2000).The importance of radical reactions in normal biologicalprocesses and in the mechanisms of toxicity of manyforeign compounds has given rise to a research thrustinto cell pro-oxidant and antioxidant processes (Halli-well and Gutteridge, 1999; Kehrer, 1993). The level ofantioxidants against ROS overproduction and itsassociation with local tissue damage can be interpretedas a toxicity measure in aquatic organisms exposed topollution (Ahmad et al., 2000; Di Giulio et al., 1989,1995; Filho, 1996; Lackner, 1998; Lemaire and Living-stone, 1993; Lopez-Torres et al., 1993; Winston andDi Giulio, 1991).Previously, it has been shown that overactivation of
fish phagocytes was associated with the release of ROS,contributing to the overall oxidative stress in fish tissuesinflicted by the constituents of paper mill effluent and bysublethal naphthalene exposure in laboratory conditions(Ahmad et al., 1998, 2003; Fatima et al., 2000).Additionally, we have also observed that this differentiallipid peroxidation (LPO) response was due to differentantioxidant levels in tissues of exposed animals (Ahmadet al., 2000). As several laboratory studies have beendone in order to observe the role of circulating fishphagocytes, we were interested to observe this approachunder natural environmental conditions on the AveiroLagoon harbor water. Our main aim in the presentstudy was to understand the relationship betweendifferent biological responses, viz., overactivated pha-gocytes vs peroxidative damage, its association to theantioxidant enzymes and nonenzymatic antioxidantdefense molecules and liver biotransformation capacityon natural Aveiro Lagoon harbor water. In this work wehave studied phagocytic respiratory burst activity(RBA) for ROS production; tissue oxidative stressestimators as gill, kidney, and liver LPO; liver biotrans-formation indicators such as ethoxyresorufin O-deethy-
Fig. 1. Representation of A. anguilla sampling area and exposure cag
lase (EROD) activity and cytochrome P450 (Cyt P450)content; bile metabolites; antioxidant enzymes: glu-tathione peroxidase (GPX), catalase (CAT), and glu-tathione S-transferase (GST), and nonenzymaticantioxidant such as glutathione (GSH) in gill, kidney,and liver of fish.
2. Materials and methods
2.1. Chemicals
1-chloro-2,4-dinitrobenzene (CDNB), dimethyl sulf-oxide (DMSO), 2,5-dithio-bis-tetranitrobenzoic acid(DTNB), Hank’s balanced salt solution (HBSS), histo-paque-1077, nitroblue tetrazolium (NBT), Roswell ParkMemorial Institute-1640 (RPMI-1640), sigmacote, 2-thiobarbituric acid (TBA), trichloroacetic acid (TCA),resorufin, and dithionite were purchased from SigmaUSA, NADPH and 7-ethoxyresorufin were from Roche,and other routine chemicals and reagents (analyticalgrade) were purchased from local sources.
2.2. Animals
European eel, Anguilla anguilla L., of length26.1273.2 cm and weight 33.6711.3 g, were capturedat the Aveiro Lagoon (Fig. 1) and were acclimatized for7 days under standard conditions (APHA, 1998) in 80-Lartificial sea water aquaria (Sera Meersaltz, Heinsberg,Germany). Water oxygen saturation was maintainedby aeration. Water temperature was maintained at2071.4�C.
2.3. Environmental contaminated water exposure
A. anguilla were caged into a harbor area of the AveiroLagoon (Fig. 1) for 8 and 48h, after acclimatization,
ing site (harbor water) on the Aveiro Lagoon, Aveiro, Portugal.

ARTICLE IN PRESSI. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302292
without any food supply. During acclimatization thewater salinity was maintained according to the harborwater salinity. The caging site is located in an offwardfishing harbor. Two groups of 10 fish each weretranslocated from the laboratory to the exposure sitein a recipient without water (30min) placed, in 80 dm3
net cages, and plunged in the harbor water near thesediments. For control, two more groups of 10 fisheach were maintained in the laboratory at 20�C in asimilar net cage and submitted to the equivalentprevious dry period (30min). The experiment wascarried out in September, at approximately 20�C watertemperature.
2.4. Experiment
The objective of this study was to evaluate the roleof various antioxidant enzymes and nonenzymaticantioxidants against circulating fish phagocytes inducedtoxicity in the natural conditions of Aveiro Lagoonharbor water. These objectives were based on theinformation reported in our previous laboratory studyin A. anguilla concerning naphthalene exposure (Ahmadet al., 2003). A. anguilla showed measurable significantaffects at 8 and 48 h exposure to naphthalene (Ahmadet al., 2003). Keeping in view these preliminaryobservations the field study was conducted by plungingcaged A. anguilla into harbor water for 8 and 48 h.
2.4.1. Isolation of phagocytes from peritoneum cavity,
gill and head kidney
Phagocytes were isolated using our previously de-scribed method (Ahmad et al., 1998). Briefly, the fishblood was drained by severing the caudal vein to avoidthe contamination of peritoneal and gill exudates by redblood cells. An abdominal incision was made and theperitoneal cavity and gill were thoroughly rinsed withcold HBSS in siliconized test tubes, transferred intoseparate unsiliconized Petri dishes, and incubated for 1 hfor adherence. These dishes’ supernatant was decantedand cells were detached with the help of a cell scraper infresh cold HBSS. Detached cells were centrifuged insiliconized tubes at 2500 r.p.m. for 10min (� 3) and thefinal cell pellet was resuspended in 1mL of RPMI-1640medium. Cells were counted using a hemocytometer.Differential counts were performed to assess thephagocyte populations in the cell suspensions. Fishhead kidney (pronephros), a hemopoietic and lymphoidtissue, was dissected out of the fish and minced using acell dissociation sieve tissue grinder kit (Sigma). Thecells were separated from tissue in RPMI-1640. Thesuspension was layered onto Histopaque-1077 andcentrifuged at 1800 r.p.m. for 30min. The upper celllayer was collected, washed twice with RPMI-1640, andused for experiments.
2.4.2. RBA assay
The oxidative burst phenomenon was studied toobserve the activation pattern of phagocytes in differentfish groups. RBA of peritoneal cavity, gill and headkidney fish phagocytes was measured by the method ofFujiki and Yano (1997), modified by Fatima et al.(2000). Briefly, an aliquot (0.1mL) of phagocytesuspension was mixed with 0.5mL HBSS-P containing0.1% NBT and 25� 106 cells/mL yeast cells (heattreated at 100�C for 1 h). The mixture was incubatedat 35�C for 2 h with occasional shaking. The suspensionwas centrifuged at 3000 r.p.m. for 15min after incuba-tion. The cell pellet was washed with 70% methanol,air-dried at 60�C for 15min, and solubilized with 0.5mLof 2M KOH by sonication in an ultrasonic processor(U 200 S control, Germany) for 5min. The sonicatedmaterial was extracted with 0.5mL DMSO for 5min.The NBT reduction potential of phagocytes wasmeasured in the supernatant after centrifugation at4500 r.p.m. for 5min as absorbance at 630 nm andexpressed as A630 nm per 106 cells.
2.4.3. Estimation of LPO in fish tissues
LPO was estimated in gill, kidney, and liver of treatedand control fish. Gill were excise out of the sacrificedfish, gill rackers were removed, and gill lamellae werehomogenized in a Potter–Elvehjem homogenizer. TheLPO activity was determined by the procedure of Utleyet al. (1967) modified and adapted by Fatima et al.(2000). Briefly, the tissues were homogenized in chilled0.15M potassium chloride solution. Thereafter 1.0mLof each homogenate was dispensed in a glass tube andincubated at 37�C in a metabolic shaker for 60min. A1.0mL aliquot of the same tissue homogenate wasincubated at 0�C, which served as a control reaction set.After 1 h incubation, 1. 0mL of 10% chilled TCA and1.0mL of 0.67% TBA were added to each sample andmixed well. The aliquots of each vial were thencentrifuged at 3000 r.p.m. for 10min. The supernatantfrom each tube was decanted into separate tubes andplaced in a boiling water bath. The test tubes wereremoved and cooled at room temperature after 10minboiling. The absorbance of each aliquot was measuredat 535 nm. The rate of LPO was expressed as nanomolesof thiobarbituric acid reactive substance (TBARS)formed per hour per milligram of protein using a molarextinction coefficient of 1.56� 105M�1 cm�1.
2.4.4. Liver EROD assay
Microsomes were obtained according to the methodsof Lange et al. (1993) and Monod and Vindimian(1991), adapted by Pacheco and Santos (1998). ERODactivity was measured as described by Burke and Mayer(1974). The reaction was carried out in a fluorometercuvette containing 1mL 0.5 mM ethoxyresorufin (0.1MTris-HCl buffer, pH 7.4, containing 0.15M KCl and

ARTICLE IN PRESSI. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302 293
20% glycerol) and 25 mL of microsomal suspension at25�C with constant agitation and initiated by adding10 mL of NADPH (10mM). The progressive increase influorescence resulting from the resorufin formation wasmeasured for 3min (excitation wavelength 530 nm,emission wavelength 585 nm) with a Jasco FP-777spectrofluorometer. Results were expressed as pmol/min/mg of microsomal protein.
2.4.5. Cyt P450 assay
Liver microsomal Cyt P450 content was determinedusing the dithionite reduced carbon monoxide differencespectrum between 450 and 490 nm, as previouslydescribed by Hermens et al. (1990).
2.4.6. Fixed wavelength fluorescence (FF) of bile
FF of bile was performed according to the methods ofAas et al. (1998), Lin et al. (1996), and Gagnon andHoldway (2000). FF readings discriminate between2-, 4-, and 5-ring PAH metabolites, i.e., naphthalene-,pyrene-, and benzo(a)pyrene-type metabolites by usingthe wavelengths pairs 290/335, 341/383, and 380/430 nm, respectively. Bile samples were diluted with 20volumes of water to give an initial bile solution that wasassayed for protein content. This solution was furtherdiluted in methanol 50% to 1:400 for FF measurement.A 5-nm slit width was used for both excitation andemission. Bile metabolites are reported on the basis ofmg protein.
2.4.7. Preparation of post-mitochondrial supernantant
(PMS)
PMS was obtained according to Ahmad et al. (2000).Liver, kidney, and gill of the sacrificed fish were quicklyremoved, cleaned of extraneous material, and immedi-ately perfused with ice-cold saline. The tissues werehomogenized in chilled phosphate buffer (0.1M, pH 7.4)containing KCl (1.17%), using a Potter–Elvehjemhomogenizer. The homogenate was filtered throughmuslin cloth and centrifuged at 800 g for 5min at 4�Cto separate the nuclear debris. The supernatantwas centrifuged in a refrigerated ultracentrifuge (OptimaTL, Beckman) at 10,000 r.p.m. for 30min at 4�C toobtain the PMS, which was used for biochemicalanalysis.
2.4.8. GPX activity
GPX activity was assayed according to the methoddescribed by Mohandas et al. (1984) as modified byAthar and Iqbal (1998). The assay mixture consisted of1.44mL phosphate buffer (0.05M, pH 7.0), 0.1mLEDTA (1mM), 0.1mL sodium azide (1mM), 0.05mLGSH reductase (1 IU/mL), 0.1mL GSH (1mM),0.1mL NADPH (0.2mM), 0.01mL H2O2 (0.025mM),and 0.1mL of PMS (10%) in a total volume of 2mL.Oxidation of NADPH was recorded spectrophotome-
trically (Jasco UV/VIS, V-530) at 340 nm at roomtemperature. The enzyme activity was calculated asnanomoles NADPH oxidized/min/mg of protein, usinga molar extinction coefficient of 6.22� 103M�1 cm�1.
2.4.9. CAT activity
CAT activity was assayed by the method of Claiborne(1985) as described by Giri et al. (1996). Briefly, theassay mixture consisted of 1.95mL phosphate buffer(0.05M, pH 7.0), 1mL hydrogen peroxide (0.019M),and 0.05mL 10% PMS in a final volume of 3mL.Change in absorbance was recorded at 240 nm. CATactivity was calculated in terms of nanomoles H2O2
consumed/min/mg protein.
2.4.10. GST activity
GST activity was determined by the method of Habiget al. (1974) with some modification (Raisuddin et al.,1994). The reaction mixture consisted of 1.65mLsodium phosphate buffer (0.1M, pH 6.5), 0.1mLreduced GSH (1mM), 0.05mL CDNB (1mM), and0.2mL PMS (10%) in a total volume of 2mL. Thechange in absorbance was recorded at 340 nm andenzyme activity calculated as nanomoles CDNB con-jugate formed/min/mg protein using a molar extinctioncoefficient of 9.6� 103M�1 cm�1.
2.4.11. Estimation of GSH
GSH was determined in PMS by the method ofJollow et al. (1974) as described by Iqbal et al. (1999).PMS (1mL) was precipitated with 1mL of sulfosalicylicacid (4.0%). The samples were kept at 4�C for 1 h andthen subjected to centrifugation at 3000 r.p.m. for15min at 4�C. The assay mixture contained 0.5mL offiltered aliquot, 2.3mL of sodium phosphate buffer(0.1M, pH 7.4), and 0.2mL DTNB (stock=100mM in0.1M sodium phosphate buffer, pH 7.4) in a totalvolume of 3mL. The reaction product optical density(OD) was read immediately at 412 nm on a spectro-photometer.
2.4.12. Protein measurement
Total protein content was determined according tothe Biuret method (Gornall et al., 1949), using bovineserum albumin (E. Merck-Darmstadt, Germany) as astandard.
2.5. Statistical analysis
The means significance difference between controland exposed animals was determined using the two-tailed Student’s t-test. The significance level was set atPo0.05.

ARTICLE IN PRESS
0
0.25
0.5
0.75
1 peritoneal phagocytes
*
0
0.25
0.5
0.75
1 gill phagocytes
**
0
0.25
0.5
0.75
1
OD
at
630
nm
/106 c
ells
OD
at
630
nm
/106 c
ells
OD
at
630
nm
/106 c
ells
8 hrs 48 hrs
Control Exposed
head kidney phagocytes
**
Fig. 2. RBA in peritoneum, gill and head kidney phagocytes of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values
are expressed as OD at 630 nm/106 cells (mean7SE, n=5/exposure). The significance levels observed are �Po0.05 and ��Po0.01 when compared
with their respective control values.
I. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302294
3. Results
3.1. Effect on RBA of fish phagocytes
Fig. 2 shows the RBA as measured by the NBTreduction of fish phagocytes isolated from peritoneum,gill, and head kidney after 8 and 48 h exposure tocontaminated harbor water. No significant RBA induc-tion was observed after 8 h exposure, whereas 48 hexposure showed significant (Po0.05) RBA induction inperitoneal exudate phagocytes. In gill-adhered phago-cytes, the induction was significantly (Po0.01) pro-nounced at 8 h exposure. A similar significant (Po0.01)RBA induction was observed in head kidney phagocytesafter 48 h exposure; in contrast, 8 h exposure showed nosignificant change in RBA when compared with therespective controls.
3.2. Effect on LPO in fish tissues
Contaminated harbor water exposure increased LPOactivity significantly (Po0.001, Po0.05) in gill after 8and 48 h exposure (respectively) (Fig. 3). A significantly(Po0.01) increased LPO was also found in kidney andliver after 48 h exposure to harbor water. However, 8 hexposure showed no significant change in LPO activityin either kidney or liver.
3.3. Effect on liver EROD activity
Results in Fig. 4 reveal that harbor water is able toinduce a significant (Po0.05) EROD activity decrease inliver after 48 h exposure and a nonsignificant decreasein its activity after 8 h exposure.
3.4. Effect on liver Cyt P450 content
This biomarker exhibited no statistically significantalteration in liver after 8 or 48 h exposure to harborwater (Fig. 5), despite the slight decrease observed inexposed fish.
3.5. Effect on fixed wavelength fluorescence (FF) of bile
Exposed fish exhibit a significant (Po0.05) increase inpyrene-type bile metabolites after 48 h exposure (Fig. 6).Additionally, benzo(a)pyrene-type metabolites increasedconsiderably after 8 and 48 h exposure, despite theabsence of any statistical significance.
3.6. Effect on enzymatic antioxidants in fish tissues
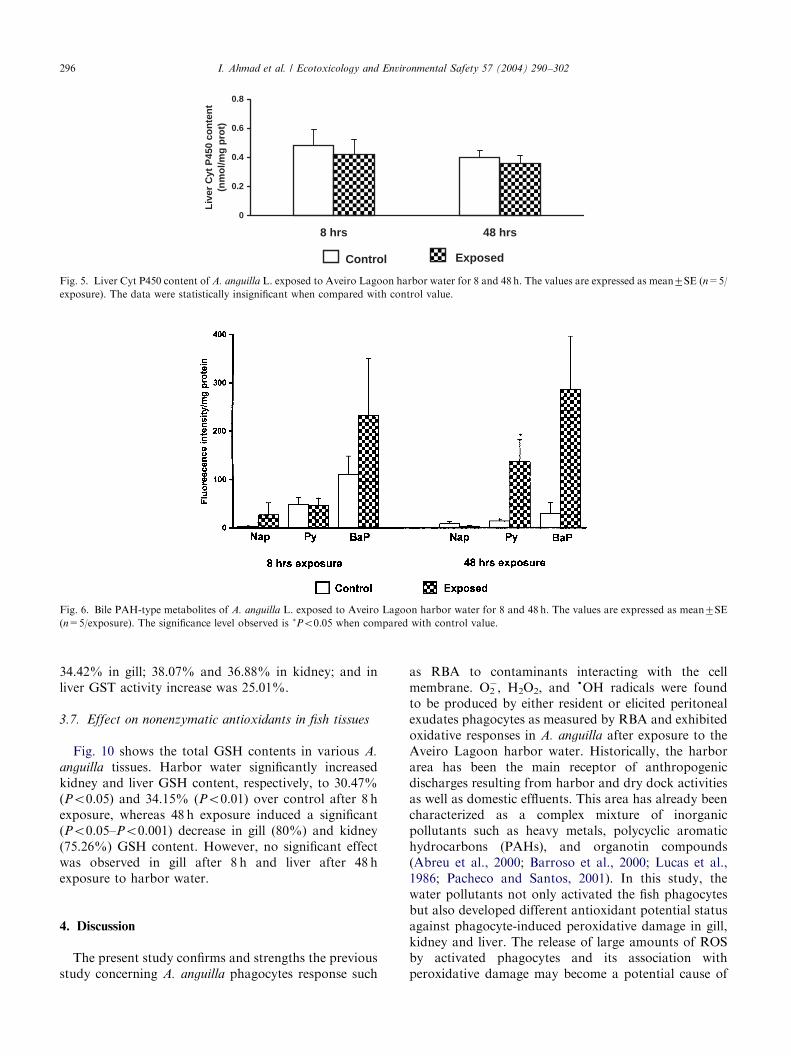
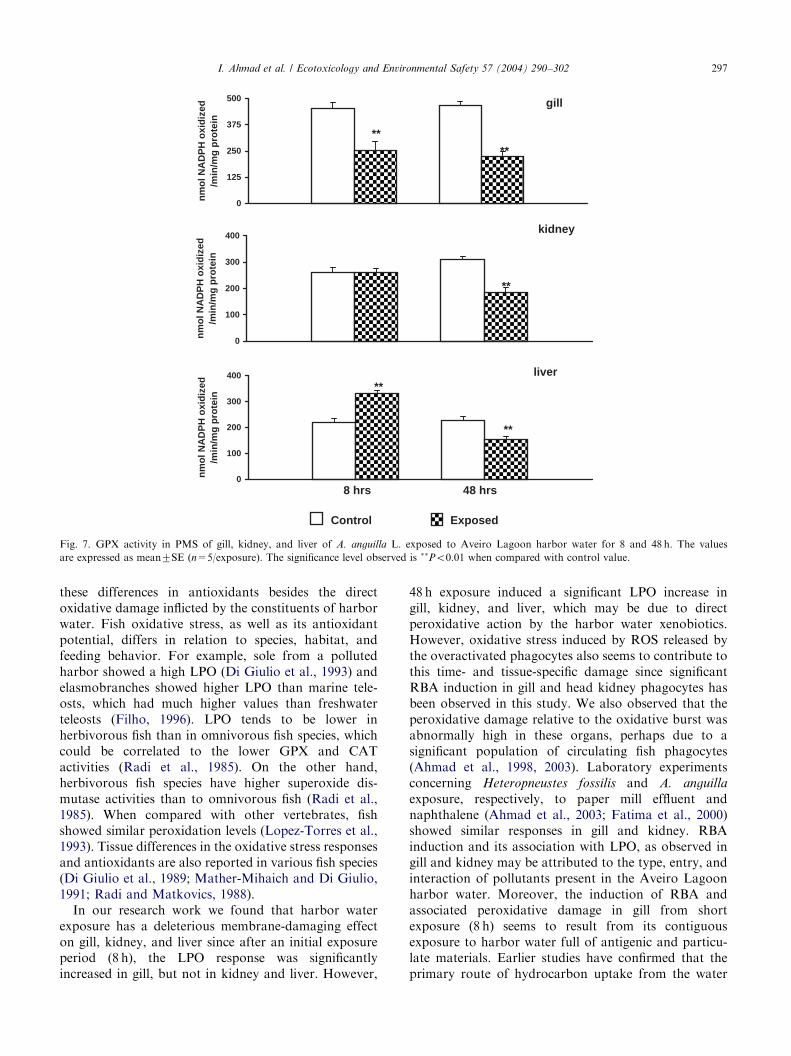
Harbor water exposure showed a significant (Po0.01)decrease in gill GPX activity after 8 and 48 h of exposure(Fig. 7). The decrease was 44.19% and 51.55%,respectively. A significant (Po0.01) decrease in GPX

ARTICLE IN PRESS
0
0.25
0.5
0.75
1
nm
ol T
BA
RS
fo
rmed
/hr
/mg
of
pro
tein
gill
*
***
0
0.25
0.5
0.75
1
nm
ol T
BA
RS
fo
rmed
/hr
/mg
of
pro
tein
kidney
**
0
0.25
0.5
0.75
nm
ol T
BA
RS
fo
rmed
/hr
/mg
of
pro
tein
8 hrs 48 hrs
Control Exposed
liver
**
Fig. 3. LPO values in gill, kidney and liver of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values are expressed
as nanomoles of TBARS released during 1 h of incubation (mean7SE, n=5/exposure). The significance levels observed are �Po0.05, ��Po0.01,
and ���Po0.001 when compared with their respective control values.
0
4
8
12
8 hrs 48 hrs
Liv
er E
RO
D A
ctiv
ity
(pm
ol/m
in/m
g p
rot)
*
Control Exposed
Fig. 4. Liver EROD activity of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values are expressed as mean7SE
(n=5/exposure). The significance level observed is �Po0.05 when compared with control value.
I. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302 295
activity (40.58%) was recorded in kidney after 48 hexposure (Fig. 7). Liver showed a significant (Po0.01)increase (150.10%) in GPX activity after 8 h exposure,in contrast a significant (Po0.01) decrease (32.1%) in itsactivity was observed at 48 h exposure when comparedwith control groups.Gill and kidney show a significant (Po0.05 and
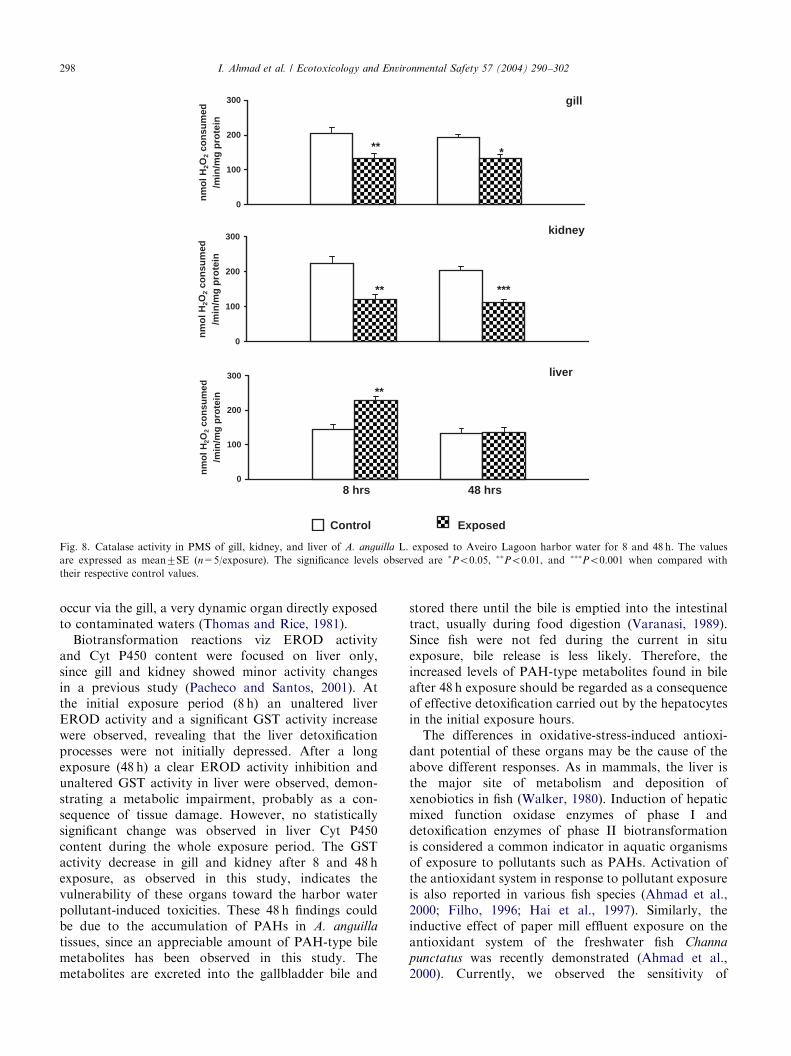
Po0.01) decrease in CAT activity after 8 and 48 hexposure (Fig. 8). The decrease in gill was 35.45% and
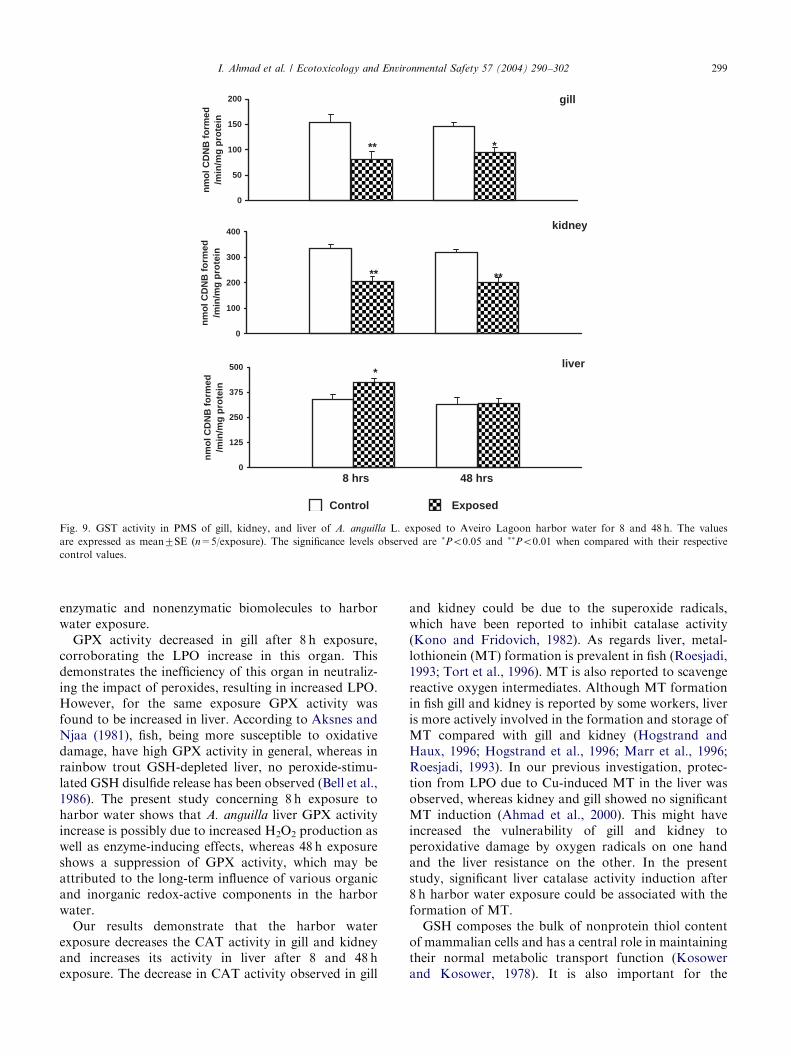
31.22%; in kidney it was 46.73% and 45.33%. However,CAT activity was significantly (Po0.01) increased inliver (58%) after 8 h exposure.GST activity was significantly (Po0.05, Po0.01)
decreased in gill and kidney after 8 and 48 h exposure,whereas liver showed a significant (Po0.05) increase inits activity only after 8 h exposure. After 48 h exposureno significant change in liver GST activity was observed(Fig. 9). The decrease in GST activity was 47.5% and
ARTICLE IN PRESS
0
0.2
0.4
0.6
0.8
8 hrs 48 hrs
Liv
er C
yt P
450
con
ten
t(n
mo
l/mg
pro
t)
Control Exposed
Fig. 5. Liver Cyt P450 content of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values are expressed as mean7SE (n=5/
exposure). The data were statistically insignificant when compared with control value.
Fig. 6. Bile PAH-type metabolites of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values are expressed as mean7SE
(n=5/exposure). The significance level observed is �Po0.05 when compared with control value.
I. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302296
34.42% in gill; 38.07% and 36.88% in kidney; and inliver GST activity increase was 25.01%.
3.7. Effect on nonenzymatic antioxidants in fish tissues
Fig. 10 shows the total GSH contents in various A.
anguilla tissues. Harbor water significantly increasedkidney and liver GSH content, respectively, to 30.47%(Po0.05) and 34.15% (Po0.01) over control after 8 hexposure, whereas 48 h exposure induced a significant(Po0.05–Po0.001) decrease in gill (80%) and kidney(75.26%) GSH content. However, no significant effectwas observed in gill after 8 h and liver after 48 hexposure to harbor water.
4. Discussion
The present study confirms and strengths the previousstudy concerning A. anguilla phagocytes response such
as RBA to contaminants interacting with the cellmembrane. O2
�, H2O2, anddOH radicals were found
to be produced by either resident or elicited peritonealexudates phagocytes as measured by RBA and exhibitedoxidative responses in A. anguilla after exposure to theAveiro Lagoon harbor water. Historically, the harborarea has been the main receptor of anthropogenicdischarges resulting from harbor and dry dock activitiesas well as domestic effluents. This area has already beencharacterized as a complex mixture of inorganicpollutants such as heavy metals, polycyclic aromatichydrocarbons (PAHs), and organotin compounds(Abreu et al., 2000; Barroso et al., 2000; Lucas et al.,1986; Pacheco and Santos, 2001). In this study, thewater pollutants not only activated the fish phagocytesbut also developed different antioxidant potential statusagainst phagocyte-induced peroxidative damage in gill,kidney and liver. The release of large amounts of ROSby activated phagocytes and its association withperoxidative damage may become a potential cause of
ARTICLE IN PRESS
0
125
250
375
500
nm
ol N
AD
PH
oxi
diz
ed/m
in/m
g p
rote
in
gill
**
**
0
100
200
300
400
nm
ol N
AD
PH
oxi
diz
ed/m
in/m
g p
rote
in
kidney
**
1
0
100
200
300
400
nm
ol N
AD
PH
oxi
diz
ed/m
in/m
g p
rote
in
8 hrs 48 hrs
Control Exposed
liver**
**
Fig. 7. GPX activity in PMS of gill, kidney, and liver of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values
are expressed as mean7SE (n=5/exposure). The significance level observed is ��Po0.01 when compared with control value.
I. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302 297
these differences in antioxidants besides the directoxidative damage inflicted by the constituents of harborwater. Fish oxidative stress, as well as its antioxidantpotential, differs in relation to species, habitat, andfeeding behavior. For example, sole from a pollutedharbor showed a high LPO (Di Giulio et al., 1993) andelasmobranches showed higher LPO than marine tele-osts, which had much higher values than freshwaterteleosts (Filho, 1996). LPO tends to be lower inherbivorous fish than in omnivorous fish species, whichcould be correlated to the lower GPX and CATactivities (Radi et al., 1985). On the other hand,herbivorous fish species have higher superoxide dis-mutase activities than to omnivorous fish (Radi et al.,1985). When compared with other vertebrates, fishshowed similar peroxidation levels (Lopez-Torres et al.,1993). Tissue differences in the oxidative stress responsesand antioxidants are also reported in various fish species(Di Giulio et al., 1989; Mather-Mihaich and Di Giulio,1991; Radi and Matkovics, 1988).In our research work we found that harbor water
exposure has a deleterious membrane-damaging effecton gill, kidney, and liver since after an initial exposureperiod (8 h), the LPO response was significantlyincreased in gill, but not in kidney and liver. However,
48 h exposure induced a significant LPO increase ingill, kidney, and liver, which may be due to directperoxidative action by the harbor water xenobiotics.However, oxidative stress induced by ROS released bythe overactivated phagocytes also seems to contribute tothis time- and tissue-specific damage since significantRBA induction in gill and head kidney phagocytes hasbeen observed in this study. We also observed that theperoxidative damage relative to the oxidative burst wasabnormally high in these organs, perhaps due to asignificant population of circulating fish phagocytes(Ahmad et al., 1998, 2003). Laboratory experimentsconcerning Heteropneustes fossilis and A. anguilla
exposure, respectively, to paper mill effluent andnaphthalene (Ahmad et al., 2003; Fatima et al., 2000)showed similar responses in gill and kidney. RBAinduction and its association with LPO, as observed ingill and kidney may be attributed to the type, entry, andinteraction of pollutants present in the Aveiro Lagoonharbor water. Moreover, the induction of RBA andassociated peroxidative damage in gill from shortexposure (8 h) seems to result from its contiguousexposure to harbor water full of antigenic and particu-late materials. Earlier studies have confirmed that theprimary route of hydrocarbon uptake from the water
ARTICLE IN PRESS
0
100
200
300
nm
ol H
2O2
con
sum
ed/m
in/m
g p
rote
in
gill
* **
0
100
200
300
nm
ol H
2O2
con
sum
ed/m
in/m
g p
rote
in
kidney
** ***
0
100
200
300
nm
ol H
2O2
con
sum
ed/m
in/m
g p
rote
in
8 hrs 48 hrs
Control Exposed
liver
**
Fig. 8. Catalase activity in PMS of gill, kidney, and liver of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values
are expressed as mean7SE (n=5/exposure). The significance levels observed are �Po0.05, ��Po0.01, and ���Po0.001 when compared with
their respective control values.
I. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302298
occur via the gill, a very dynamic organ directly exposedto contaminated waters (Thomas and Rice, 1981).Biotransformation reactions viz EROD activity
and Cyt P450 content were focused on liver only,since gill and kidney showed minor activity changesin a previous study (Pacheco and Santos, 2001). Atthe initial exposure period (8 h) an unaltered liverEROD activity and a significant GST activity increasewere observed, revealing that the liver detoxificationprocesses were not initially depressed. After a longexposure (48 h) a clear EROD activity inhibition andunaltered GST activity in liver were observed, demon-strating a metabolic impairment, probably as a con-sequence of tissue damage. However, no statisticallysignificant change was observed in liver Cyt P450content during the whole exposure period. The GSTactivity decrease in gill and kidney after 8 and 48 hexposure, as observed in this study, indicates thevulnerability of these organs toward the harbor waterpollutant-induced toxicities. These 48 h findings couldbe due to the accumulation of PAHs in A. anguilla
tissues, since an appreciable amount of PAH-type bilemetabolites has been observed in this study. Themetabolites are excreted into the gallbladder bile and
stored there until the bile is emptied into the intestinaltract, usually during food digestion (Varanasi, 1989).Since fish were not fed during the current in situexposure, bile release is less likely. Therefore, theincreased levels of PAH-type metabolites found in bileafter 48 h exposure should be regarded as a consequenceof effective detoxification carried out by the hepatocytesin the initial exposure hours.The differences in oxidative-stress-induced antioxi-
dant potential of these organs may be the cause of theabove different responses. As in mammals, the liver isthe major site of metabolism and deposition ofxenobiotics in fish (Walker, 1980). Induction of hepaticmixed function oxidase enzymes of phase I anddetoxification enzymes of phase II biotransformationis considered a common indicator in aquatic organismsof exposure to pollutants such as PAHs. Activation ofthe antioxidant system in response to pollutant exposureis also reported in various fish species (Ahmad et al.,2000; Filho, 1996; Hai et al., 1997). Similarly, theinductive effect of paper mill effluent exposure on theantioxidant system of the freshwater fish Channa
punctatus was recently demonstrated (Ahmad et al.,2000). Currently, we observed the sensitivity of
ARTICLE IN PRESS
0
50
100
150
200
nm
ol C
DN
B f
orm
ed/m
in/m
g p
rote
in
gill
* **
0
100
200
300
400
nm
ol C
DN
B f
orm
ed/m
in/m
g p
rote
inkidney
** **
0
125
250
375
500
nm
ol C
DN
B f
orm
ed/m
in/m
g p
rote
in
8 hrs 48 hrs
Control Exposed
liver *
Fig. 9. GST activity in PMS of gill, kidney, and liver of A. anguilla L. exposed to Aveiro Lagoon harbor water for 8 and 48 h. The values
are expressed as mean7SE (n=5/exposure). The significance levels observed are �Po0.05 and ��Po0.01 when compared with their respective
control values.
I. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302 299
enzymatic and nonenzymatic biomolecules to harborwater exposure.GPX activity decreased in gill after 8 h exposure,
corroborating the LPO increase in this organ. Thisdemonstrates the inefficiency of this organ in neutraliz-ing the impact of peroxides, resulting in increased LPO.However, for the same exposure GPX activity wasfound to be increased in liver. According to Aksnes andNjaa (1981), fish, being more susceptible to oxidativedamage, have high GPX activity in general, whereas inrainbow trout GSH-depleted liver, no peroxide-stimu-lated GSH disulfide release has been observed (Bell et al.,1986). The present study concerning 8 h exposure toharbor water shows that A. anguilla liver GPX activityincrease is possibly due to increased H2O2 production aswell as enzyme-inducing effects, whereas 48 h exposureshows a suppression of GPX activity, which may beattributed to the long-term influence of various organicand inorganic redox-active components in the harborwater.Our results demonstrate that the harbor water
exposure decreases the CAT activity in gill and kidneyand increases its activity in liver after 8 and 48 hexposure. The decrease in CAT activity observed in gill
and kidney could be due to the superoxide radicals,which have been reported to inhibit catalase activity(Kono and Fridovich, 1982). As regards liver, metal-lothionein (MT) formation is prevalent in fish (Roesjadi,1993; Tort et al., 1996). MT is also reported to scavengereactive oxygen intermediates. Although MT formationin fish gill and kidney is reported by some workers, liveris more actively involved in the formation and storage ofMT compared with gill and kidney (Hogstrand andHaux, 1996; Hogstrand et al., 1996; Marr et al., 1996;Roesjadi, 1993). In our previous investigation, protec-tion from LPO due to Cu-induced MT in the liver wasobserved, whereas kidney and gill showed no significantMT induction (Ahmad et al., 2000). This might haveincreased the vulnerability of gill and kidney toperoxidative damage by oxygen radicals on one handand the liver resistance on the other. In the presentstudy, significant liver catalase activity induction after8 h harbor water exposure could be associated with theformation of MT.GSH composes the bulk of nonprotein thiol content
of mammalian cells and has a central role in maintainingtheir normal metabolic transport function (Kosowerand Kosower, 1978). It is also important for the

ARTICLE IN PRESS
0
0.01
0.02
0.03
0.04
nm
ol/g
tis
sue
gill
*
0
0.025
0.05
0.075
nm
ol/g
tis
sue
kidney
***
*
0
0.05
0.1
0.15
nm
ol/g
tis
sue
8 hrs 48 hrs
Control Exposed
liver**
Fig. 10. Effect of Aveiro Lagoon harbor water on reduced glutathione content in gill, kidney, and liver of A. anguilla L. exposed to Aveiro Lagoon
harbor water for 8 and 48 h. The values are expressed as mean7SE (n=5/exposure). The significance levels observed are �Po0.05, ��Po0.01,
and ���Po0.001 when compared with their respective control values.
I. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302300
detoxification and excretion of xenobiotics and forprotecting cells against their toxic actions (Otto andMoon, 1995; Regoli and Principato, 1995). We foundan initial increase in A. anguilla liver and kidney GSHlevels exposed to harbor water. However, the liverlevel decreased to the control level after 48 h exposure,whereas it significantly decreased in kidney and gill. Thisbiphasic response is considered an adaptation toa detoxification activity increase (Buchner et al., 1993;Chatterjee and Bhattacharya, 1984). Moreover, theprotective effect of GSH was attributed to its ability tostablize the pollutants by scavenging the free radicalsand thereby blocking LPO development (Meister andAnderson, 1983). The initial elevated level of GSHappears to be effective in providing protection toorganisms since no significant increase in LPO wasobserved in the liver and kidney after 8 h exposure.Increased liver and kidney GSH level serves as anindicator of degree and duration of fish exposure tooxidant pollutants. The protective and adaptive role ofGSH against oxidative stress-induced toxicity is wellestablished in aquatic animals (Otto and Moon, 1995;Regoli and Principato, 1995).
This investigation establishes that pollution-inducedphagocyte overactivation may not only affects thenormal immune function but also affect other organsstructurally and functionally. The release of ROS byoveractivation of fish phagocytes after exposure toharbor water corroborates the tissue-specific peroxida-tive damage in gill, kidney, and liver after 8 h. The tissuemost affected was gill. This finding also establishesthe development of tissue-specific adaptive responses toneutralize oxidative stress. The organ most highlyadaptive to oxidative damage was liver, compared togill and kidney. Furthermore, the tissue-specific anti-oxidant molecules inhibition/induction proves to be apotent cause of phagocyte-induced peroxidative damageand may be helpful in assessing the oxidative damagerisk associated with the water quality.
Acknowledgments
The financial support from the FCT (Government ofPortugal) provided through Contract S.F.R.H/B.P.D/

ARTICLE IN PRESSI. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302 301
3603/2000 and by the Aveiro University ResearchInstitute/CESAM is gratefully acknowledged.
References
Aas, E., Beyer, J., Goksoyr, A., 1998. PAH in fish bile detected by
fixed wavelength fluorescence. Mar. Environ. Res. 46, 225–228.
Abreu, S.N., Pereira, E., Vale, C., Duarte, A.C., 2000. Accumulation
of mercury in sea bass from a contaminated lagoon (Ria de Aveiro,
Portugal). Mar. Pollut. Bull. 40, 293–297.
Ahmad, I., Fatima, M., Athar, M., Khan, N.Z., Raisuddin, S., 1998.
Responses of circulating fish phagocytes to paper mill effluent
exposure. Bull. Environ. Contam. Toxicol. 61, 746–753.
Ahmad, I., Hamid, T., Fatima, M., Chand, H.S., Jain, S.K., Athar,
M., Raisuddin, S., 2000. Induction of hepatic antioxidants in
freshwater catfish (Channa punctatus Bloch.) is a biomarker of
paper mill effluent exposure. Biochim. Biophys. Acta 1523, 37–48.
Ahmad, I., Pacheco, M., Santos, M.A., 2003. Naphthalene-induced
differential tissue damage association with circulating fish phago-
cytes induction. Ecotoxicol. Environ. Saf. 54, 7–15.
Aksnes, A., Njaa, L.R., 1981. Catalase, glutathione peroxidase and
superoxide dismutase in different fish species. Comp. Biochem.
Physiol. B69, 893–896.
APHA, 1998. Standard methods for the examination of water and
wastewater in. In: Clesceri, L.S., Greenberg, A.E., Eaton, A.D.
(Eds.), Animal Public Health Association, APHA, 20th Edition.
American Public Health Association, Washington, DC, 1998.
Athar, M., Iqbal, M., 1998. Ferric nitrilotriacetate promotes N-
diethylnitrosamine-induced renal tumorigenesis in the rat: implica-
tions for the involvement of oxidative stress. Carcinogenesis 19,
1133–1139.
Barroso, C.M., Moreira, M.H., Gibbs, P.E., 2000. Comparison of
imposex and intersex development in four prosobranch species for
TBT monitoring of a southern European estuarine system (Ria de
Aveiro, NW Portugal). Mar. Ecol. Prog. Ser. 201, 221–232.
Bell, J.G., Andron, J.W., Cowey, C.B., 1986. Effect of selenium
deficiency on hydroperoxide-stimulated release of glutathione
from isolated perfused liver of rainbow trout (Salmo gairdneri).
Br. J. Nutr. 56, 421–428.
Buchner, F., Hofer, R., Krumschnabel, G., Doblander, C., 1993.
Disturbances in the pro-oxidant–antioxidant balance in the liver of
bullhead (Cottus gobio L.) exposed to treated paper mill effluents.
Chemosphere 27, 1329–1338.
Burke, M.D., Mayer, R.T., 1974. Ethoxyresorufin: direct fluorimetric
assay of microsomal-O-deethylation which is preferentially indu-
cible by 3-methylcholanthrene. Drug. Metab. Dispos. 2, 583–588.
Chatterjee, S., Bhattacharya, S., 1984. Detoxification of industrial
pollutants by the glutathione S-transferase system in the liver of
Anabas testudineus (Bloch.). Toxicol. Lett. 22, 187–198.
Claiborne, A., 1985. Catalase activity. In: Greenwald, R.A. (Ed.),
CRC Handbook of Methods in Oxygen Radical Research. CRC
Press, Boca Raton, FL, pp. 283–284.
Di Giulio, R.T., Washburn, P.C., Wenning, R.J., Winston, G.W.,
Jewell, C.S., 1989. Biochemical responses in aquatic animals: a
review of determinants of oxidative stress. Environ. Toxicol. Chem.
8, 1103–1123.
Di Giulio, R.T., Habig, C., Gallagher, E.P., 1993. Effect of black rock
harbour sediments on indices of biotransformation, oxidative stress
and DNA integrity in channel catfish. Aquat. Toxicol. 26, 1–22.
Di Giulio, R.T., Benson, W.H., Sanders, B.M., Veld, P.A., 1995.
Biochemical mechanisms: metabolisms, adaptation and toxicity.
In: Rand, G. (Ed.), Fundamentals of Aquatic Toxicology. Effects,
Environmental Fate and Risk Assessment. Taylor and Francis,
London.
Fatima, M., Ahmad, I., Sayeed, I., Athar, M., Raisuddin, S., 2000.
Pollutant-induced overactivation of phagocytes is concomitantly
associated with peroxidative damage in fish tissues. Aquat. Toxicol.
49, 243–250.
Filho, D.W., 1996. Fish antioxidant defenses a comparative approach.
Br. J. Med. Biol. Res. 29, 1735–1742.
Fujiki, K., Yano, T., 1997. Effect of sodium alginate on the non-
specific defense system of the common carp (Cyprinus carpio L.).
Fish Shellfish Immunol. 7, 417–427.
Gagnon, M.M., Holdway, D.A., 2000. EROD induction and biliary
metabolite excretion following exposure to the water accommo-
dated fraction of crude oil and to chemically dispersed crude oil.
Arch. Environ. Contam. Toxicol. 38, 70–77.
Giri, U., Iqbal, M., Athar, M., 1996. Porphyrine-mediated photo-
sensitization has a weak tumor promoting effect in mouse skin:
possible role of in situ generated reactive oxygen species.
Carcinogenesis 17, 2023–2028.
Girotti, A.W., 1990. Photodynamic lipid peroxidation in biological
system. Photochem. Photobiol. 51, 497–509.
Gornall, A.C., Bardawill, C.J., David, M.M., 1949. Determination
of serum proteins by means of the biuret reaction. J. Biol. Chem.
177, 751–766.
Habig, W.H., Pabst, M.J., Jokoby, W.B., 1974. Glutathione S-
transferase. The first enzymatic step in mercapturic acid formation.
J. Biol. Chem. 249, 7130–7139.
Hai, D.Q., Varga, S.I., Matkovics, B., 1997. Organophosphate effects
on antioxidant system of carp (Cyprinus carpio) and catfish
(Ictalurus nebulosus). Comp. Biochem. Physiol. C117, 83–88.
Halliwell, B., Gutteridge, J.M.C., 1999. Free Radicals in Biology
and Medicine. Oxford University Press, Oxford.
Hermens, J.L.P., Bradbury, S.P., Broderius, S.J., 1990. Influence of
cytochrome P450 mixed function oxidase induction on the acute
toxicity to rainbow trout (Salmo gairdneri) of primary aromatic
amines. Ecotoxicol. Environ. Saf. 20, 156–166.
Hogstrand, C., Haux, C., 1996. Naturally high level of zinc and
metallothionein in liver of several species of the squirrel fish family
from Queenland, Australia. Mar. Biol. 125, 23–31.
Hogstrand, C., Galvez, F., Wood, C.M., 1996. Toxicity, silver
accumulation and metallothionein induction in rainbow trout
during exposure to different silver salts. Environ. Toxicol. Chem.
15, 1102–1108.
Iqbal, M., Giri, U., Giri, D.K., Alam, M.S., Athar, M., 1999. Age-
dependent renal accumulation of 4-hydroxy-2 nonenal (HNE)-
modified proteins following parenteral administration of ferric
nitrilotriacetate commensurates with its differential toxicity:
implications for the involvement of HNE–protein adducts in
oxidative stress and carcinogenesis. Arch. Biochem. Biophys. 365,
101–112.
Jollow, D.W., Mitchell, J.R., Zampagilone, N., Gilete, J.R., 1974.
Bromobenzene-induced liver necrosis: protective role of glu-
tathione and evidence for 3, 4–bromobenzeneoxide as the
hepatotoxic intermediate. Pharmacology 11, 151–169.
Kehrer, J., 1993. Free radicals as mediators of tissue injury and
disease. Crit. Rev. Toxicol. 23, 21–48.
Khan, M.A.Q., Al-Ghais, S.M., Al-mari, S., 1995. Petroleum
hydrocarbons in fish from the Arabian Gulf. Arch. Environ.
Contam. Toxicol. 29, 517–522.
Kono, Y., Fridovich, I., 1982. Superoxide radical inhibit catalase.
J. Biol. Chem. 257, 5751–5754.
Kosower, N.S., Kosower, E.N., 1978. The glutathione status of cells.
Int. Rev. Cytol. 54, 109–159.
Lackner, R., 1998. Oxidative stress in fish by environmental pollutants.
In: Braunbeck, T.D., Hinton, E., Streit, B. (Eds.), Fish Ecotox-
icology. Birkhauser, Basel, pp. 203–224.
Lange, U., Danischewski, D., Siebers, D., 1993. In: Braumbeck, T.,
Hanke, W., Segner, H. (Eds.), Regional Variability and Sexual

ARTICLE IN PRESSI. Ahmad et al. / Ecotoxicology and Environmental Safety 57 (2004) 290–302302
Differences in Ethoxyresorufin-O-deethylase Activities and Cyto-
chrome P450 Concentration in the Liver of Mature Dab (Limanda
limanda L.) in the German Bight. VCH, Verlag Chemie, New York.
Lemaire, P., Livingstone, D.R., 1993. Prooxidant/antioxidant
processes and organic xenobiotic interactions in marine organisms,
in particular the flounder Platichthys flesus and mussel Mytilus
edulis. Tr. Comp. Biochem. Physiol. 1, 1119–1150.
Lin, E.L.C., Cormier, S.M., Torsella, J.A., 1996. Fish biliary polycyclic
aromatic hydrocarbon metabolites estimated by fixed-wavelength
fluorescence: comparison with HPLC-fluorescent detection.
Ecotoxicol. Environ. Saf. 35, 16–23.
Livingstone, D.R., 1991. Organic xenobiotic metabolism in marine
invertebrates. In: Gilles, R. (Ed.), Advances in Comparative and
Environmental Physiology. Springer, Berlin, pp. 45–185.
Livingstone, D.R., 1998. The fate of organic xenobiotics in aquatic
ecosystem: quantitative and qualitative differences in biotransfor-
mation by invertebrates and fish. Comp. Biochem. Physiol. A120,
43–49.
Livingstone, D.R., Chipman, J.K., Lowe, D.M., Minier, C., Mitch-
elmore, C.L., Moore, M.N., Peters, L.D., Pipe, R.K., 2000.
Development of biomarkers to detect the effects of organic
pollution on aquatic invertebrates: recent molecular, genotoxic,
cellular and immunological studies on the common mussel (Mytilus
edulis L.) and other mytilids. Int. J. Pollut. 13, 56–91.
Lopez-Torres, M., Perez-Campo, R., Cadenas, S., Rojas, C., Bajra, C.,
1993. A comparative study of free radicals in vertebrate-II. Non-
enzymatic antioxidants and oxidative stress. Comp. Biochem.
Physiol. B105, 757–763.
Lucas, M., Caldeira, F.M.T., Hall, A., Duarte, A.C., Lima, C., 1986.
Distribution of mercury in the sediments and fishes of the lagoon
of Aveiro, Portugal. Water Sci. Technol. 18, 141–148.
Marr, J.C.A., Lipton, J., Casela, D., Hansen, J.A., Bergman, H.L.,
Mayer, J.S., Hogstrand, C., 1996. Relationship between copper
exposure duration, tissue copper concentration and rainbow trout
growth. Aquat. Toxicol. 36, 17–30.
Mather-Mihaich, E., Di Giulio, R.T., 1991. Oxidant, mixed function
oxidase and peroximal responses in channel catfish exposed to
bleached Kraft mill effluent. Arch. Environ. Contam. Toxicol. 20,
391–397.
Meister, A., Anderson, M.E., 1983. Glutathione. Ann. Rev. Biochem.
52, 711–760.
Mohandas, J., Marshall, J.J., Duggins, G.G., Horvath, J.S., Tiller, D.,
1984. Differential distribution of glutathione and glutathione
related enzymes in rabbit kidney. Possible implications in analgesic
neuropathy. Cancer Res. 44, 5086–5091.
Monod, G., Vindimian, E., 1991. Effect of storage conditions and
subcellular fraction of fish and cytochrome P450 dependent
enzymatic activities used for the monitoring of water pollution.
Water Res. 25, 173–177.
Otto, D.M., Moon, T.W., 1995. 3, 30, 4, 40-Tetrachlorobiphenyl effectson antioxidant enzymes and glutathione status in different tissues
of rainbow trout. Pharmacol. Toxicol. 77, 281–287.
Pacheco, M., Santos, M.A., 1998. Anguilla anguilla L. liver EROD and
erythrocytic nuclear abnormalities induction by cyclophosphamide
and PAHs. Ecotoxicol. Environ. Saf. 40, 71–76.
Pacheco, M., Santos, M.A., 2001. Biotransformation, endocrine and
genetic responses of Anguilla anguilla L. to petroleum distillate
products environmental contaminated waters. Ecotoxicol. Environ.
Saf. 49, 64–75.
Radi, A.A.A.R., Matkovics, B., 1988. Effects of metal ions on the
antioxidant enzyme activities, protein content and lipid peroxida-
tion of carp tissues. Comp. Biochem. Physiol. C90, 69–72.
Radi, A.A.A.R., Hai, D.Q., Matkovics, B., Gabrielak, T., 1985.
Comparative antioxidant enzyme study in freshwater fish with
different types of feeding behaviour. Comp. Biochem. Physiol. C81,
395–399.
Raisuddin, S., Parmer, D., Zaidi, S.I.A., Singh, K.P., Verma, A.S.,
Seth, P.K., Ray, P.K., 1994. Aflatoxin induces depletion of
activities of phase I biotransformation enzymes in growing rats.
Eur. J. Drug Metab. Pharmacokinet. 19, 163–168.
Regoli, F., Principato, G., 1995. Glutathione, glutathione-dependent
and antioxidant enzymes in mussel, Mytilus galloprovincialis,
exposed to metal under field and laboratory condition: implication
for the use of biomarkers. Aquat. Toxicol. 31, 143–164.
Roesjadi, G., 1993. Metallothionein induction as a measure of
response to metal exposure in aquatic animals. Environ. Health
Perspect. Suppl. 102, 91–96.
Stadtman, E.R., 1991. Ascorbic acid and oxidative inactivation of
proteins. Am. J. Clin. Nutr. 54, 1125–1128.
Thomas, R.E., Rice, S.D., 1981. Excretion of aromatic hydrocarbons
and their metabolites by freshwater and seawater Dolly Varden
char. In: Vernberg, F.J., Calabrese, A., Thurberg, F.P., Venberg,
W.B. (Eds.), Biological Monitoring of Marine Pollutants.
Academic Press, New York, pp. 425–448.
Tort, L., Kargacin, B., Torres, P., Giralt, M., Hidalgo, J., 1996.
The effect of cadmium exposure and stress plasma cortisol,
metallothionein levels and oxidative status in rainbow trout
(Oncorhynchus mykiss) liver. Comp. Biochem. Physiol. C114,
29–34.
Utley, H.C., Bernheim, F., Hachslein, P., 1967. Effect of sulfhydryl
reagent on peroxidation in microsome. Arch. Biochem. Physiol.
C114, 29–34.
Van Veld, P.A., 1990. Absorption and metabolism of dietary
xenobiotics by the intestine of fish. Rev. Aquat. Sci. 2, 185–203.
Varanasi, U. (Ed.), (1989). Metabolism of Polycyclic Aromatic
Hydrocarbons in the Aquatic Environment. CRC Press, Boca
Raton, FL.
Walker, C.H., 1980. Species variations in some hepatic microsomal
enzymes that metabolise xenobiotics. Prog. Drug Metab. 5,
113–164.
Walker, C.H., Livingstone, D.R., 1992. Persistent Pollutants in Marine
Ecosystems, SETAC Special Publication. Pergamon Press, Oxford.
Walker, C.H., Hopkin, S.P., Sibley, R.M., Peakall, D.B., 1996.
Principles of Ecotoxicology. Taylor and Francis, London.
Walsh, A.R., O’Halloran, J., 1988. Accumulation of chromium by
a population of mussels (Mytilus edulis L.) exposed to leather
tannery effluent. Environ. Toxicol. Chem. 17, 1429–1438.
Winston, G.W., Di Giulio, R.T., 1991. Prooxidant and anti-
oxidant mechanisms in aquatic organisms. Aquat. Toxicol. 19,
137–161.











![Anguilla anguilla L. Biochemical and Genotoxic Responses to Benzo[ a]pyrene](https://static.fdokumen.com/doc/165x107/631d4597f26ecf94330a787a/anguilla-anguilla-l-biochemical-and-genotoxic-responses-to-benzo-apyrene.jpg)








