
Anguilla anguilla L. oxidative stress biomarkers: An in situ study of freshwater wetland ecosystem...
11
Anguilla anguilla L. oxidative stress biomarkers: An in situ study of freshwater wetland ecosystem (Pateira de Fermentelos, Portugal) Iqbal Ahmad, Ma ´rio Pacheco, Maria Ana Santos * Animal Physiology/Ecotoxicology Sector, Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal Received 24 November 2005; received in revised form 17 February 2006; accepted 18 March 2006 Available online 24 May 2006 Abstract Pateira de Fermentelos (PF) is a natural freshwater wetland in the central region of Portugal. In the last decade, the introduction of agricultural chemicals, heavy metals, domestic wastes, as well as eutrophication and incorrect utility of resources resulted in an increased water pollution. The present research work was carried out to check the various oxidative stress biomarker responses in European eel (Anguilla anguilla L.) gill, kidney and liver due to this complex water pollution. Eels were caged and plunged at five different PF sites (A– E) for 48 h. A reference site (R) was also selected at the river spring where no industrial contamination should be detected. Lipid per- oxidation (LPO), catalase (CAT), glutathione peroxidase (GPX), glutathione S-transferase (GST) and reduced glutathione (GSH) were the oxidative stress biomarkers studied. In gill, site A exposure induced a significant GST activity increase and site B exposure induced CAT activity increase when compared to R. Site C exposure showed a significant CAT and GPX activity increase. Data concerning site D exposure were not determined due to cage disappearance. Site E exposure displayed a significant CAT and GST activity increase. In kidney, site A exposure induced a significant CAT and GPX decrease as well as a GST increase. Site B exposure showed a significant decrease in GPX activity and GSH content. However, site C exposure demonstrated a significant increase in CAT and a decrease in GPX. Site E exposure showed a significant decrease in GPX and increase in GST. In liver, site A exposure showed a significant GST activity decrease as well as GSH content increase. Site B exposure showed a significant CAT, GST and LPO decrease. Site C exposure showed only GST activity decrease, while site E exposure induced a significant increase in GPX. These investigation findings provide a rational use of oxidative stress biomarkers in freshwater ecosystem pollution biomonitoring using caged fish, and the first attempt reported in Portugal as a study of this particular watercourse under the previous conditions. The presence of pollutants in the PF water was denunciated even without a clear relation to the main pollution source distance. The organ specificity was evident for each parameter but without a clear pattern. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Freshwater ecosystem; Pateira de Fermentelos; Anguilla anguilla L.; Oxidative stress; Antioxidants; Organ specificity 1. Introduction Lakes and reservoirs are more prone to receive and accumulate contaminants discharging from sewages, domestic wastes and agriculture runoff due to their specific water dynamic and configuration differing from other aquatic ecosystems. Pateira de Fermentelos (PF), a natural freshwater wetland, is an important fishing and recreation place in the central region of Portugal, receiving water mainly from the Ce ´rtima River and discharging into the A ´ gueda River. In the last decade, eutrophication caused by sewage and agricultural fields’ nutrient runoff, pesticides leachates, electroplating industry discharges as well as incorrect utility of resources with the introduction of new organisms have aggravated the pollution problems, leading to an increased organisms’ health threat where fish are included (Calado et al., 1991; Calado and Craveiro, 1995). PF heavy metal analysis demonstrated the presence of nickel, zinc, aluminium and manganese, as a clear dem- onstration of water quality deterioration (Almeida, 1998). 0045-6535/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.chemosphere.2006.03.042 * Corresponding author. Tel.: +351 234370965; fax: +351 234426408. E-mail addresses: [email protected] (I. Ahmad), mpacheco@bio. ua.pt (M. Pacheco), [email protected] (M.A. Santos). www.elsevier.com/locate/chemosphere Chemosphere 65 (2006) 952–962
Transcript of Anguilla anguilla L. oxidative stress biomarkers: An in situ study of freshwater wetland ecosystem...

www.elsevier.com/locate/chemosphere
Chemosphere 65 (2006) 952–962
Anguilla anguilla L. oxidative stress biomarkers: An in situ studyof freshwater wetland ecosystem (Pateira de Fermentelos, Portugal)
Iqbal Ahmad, Mario Pacheco, Maria Ana Santos *
Animal Physiology/Ecotoxicology Sector, Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal
Received 24 November 2005; received in revised form 17 February 2006; accepted 18 March 2006Available online 24 May 2006
Abstract
Pateira de Fermentelos (PF) is a natural freshwater wetland in the central region of Portugal. In the last decade, the introduction ofagricultural chemicals, heavy metals, domestic wastes, as well as eutrophication and incorrect utility of resources resulted in an increasedwater pollution. The present research work was carried out to check the various oxidative stress biomarker responses in European eel(Anguilla anguilla L.) gill, kidney and liver due to this complex water pollution. Eels were caged and plunged at five different PF sites (A–E) for 48 h. A reference site (R) was also selected at the river spring where no industrial contamination should be detected. Lipid per-oxidation (LPO), catalase (CAT), glutathione peroxidase (GPX), glutathione S-transferase (GST) and reduced glutathione (GSH) werethe oxidative stress biomarkers studied. In gill, site A exposure induced a significant GST activity increase and site B exposure inducedCAT activity increase when compared to R. Site C exposure showed a significant CAT and GPX activity increase. Data concerning siteD exposure were not determined due to cage disappearance. Site E exposure displayed a significant CAT and GST activity increase. Inkidney, site A exposure induced a significant CAT and GPX decrease as well as a GST increase. Site B exposure showed a significantdecrease in GPX activity and GSH content. However, site C exposure demonstrated a significant increase in CAT and a decrease inGPX. Site E exposure showed a significant decrease in GPX and increase in GST. In liver, site A exposure showed a significant GSTactivity decrease as well as GSH content increase. Site B exposure showed a significant CAT, GST and LPO decrease. Site C exposureshowed only GST activity decrease, while site E exposure induced a significant increase in GPX. These investigation findings provide arational use of oxidative stress biomarkers in freshwater ecosystem pollution biomonitoring using caged fish, and the first attemptreported in Portugal as a study of this particular watercourse under the previous conditions. The presence of pollutants in the PF waterwas denunciated even without a clear relation to the main pollution source distance. The organ specificity was evident for each parameterbut without a clear pattern.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Freshwater ecosystem; Pateira de Fermentelos; Anguilla anguilla L.; Oxidative stress; Antioxidants; Organ specificity
1. Introduction
Lakes and reservoirs are more prone to receive andaccumulate contaminants discharging from sewages,domestic wastes and agriculture runoff due to their specificwater dynamic and configuration differing from otheraquatic ecosystems. Pateira de Fermentelos (PF), a naturalfreshwater wetland, is an important fishing and recreation
0045-6535/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.chemosphere.2006.03.042
* Corresponding author. Tel.: +351 234370965; fax: +351 234426408.E-mail addresses: [email protected] (I. Ahmad), mpacheco@bio.
ua.pt (M. Pacheco), [email protected] (M.A. Santos).
place in the central region of Portugal, receiving watermainly from the Certima River and discharging into theAgueda River. In the last decade, eutrophication causedby sewage and agricultural fields’ nutrient runoff, pesticidesleachates, electroplating industry discharges as well asincorrect utility of resources with the introduction of neworganisms have aggravated the pollution problems, leadingto an increased organisms’ health threat where fish areincluded (Calado et al., 1991; Calado and Craveiro,1995). PF heavy metal analysis demonstrated the presenceof nickel, zinc, aluminium and manganese, as a clear dem-onstration of water quality deterioration (Almeida, 1998).

I. Ahmad et al. / Chemosphere 65 (2006) 952–962 953
In these environments, organisms are exposed to mix-tures of pollutants, whose synergistic/antagonistic effectsare hardly interpreted and predicted exclusively from thechemical analyses; some contaminants strongly accumulatein tissues without inducing toxic effects, while others arecharacterised by elevated toxicity at low levels of exposure.Moreover, the impracticality to analyze all the individualchemicals pooled in a mixture of contaminants alsoincreases the problem of aquatic pollutant characterization.Thus, during the past two decades, the use of biologicalresponses (biomarkers) on particular test species has becomerelevant in toxicological assessments since it allows the earlydetection of overall effects of contaminants, providing infor-mation, even at the sub-lethal level, which reflects eventualchemical interactions (Passino, 1984; Goksøyr and Forlin,1992; Livingstone, 1993; Peakall and Shugart, 1993).
In this direction, the study of oxidative stress seems tobe particularly promising (Lackner, 1998; Livingstone,2001). Many environmental pollutants are capable ofinducing oxidative stress in aquatic animals including fish.An important role in toxicity of several pollutants isassumed by the enhancement of intracellular reactive oxy-gen species (ROS) and perturbation of antioxidant effi-ciency which often prelude the onset of alterations likeDNA damage, lipid peroxidation (LPO) and enzyme inhi-bition (Winston and Di Giulio, 1991). The overall enhance-ment or reduction of pollutant-induced toxicity greatlydepends on the imbalance between pro-oxidant and antiox-idant status as a result of xenobiotic interferences. Hence,enhanced pollutant oxygenation rates may increase toxicityvia oxidative stress, rendering fish antioxidants less effec-tive. In this perspective, a number of studies confirmedthe successful employment of antioxidant enzymes andnon-enzymatic antioxidant modulation in identifying envi-ronmental stress (Regoli et al., 1998; Ahmad et al., 2000,2004, 2005; Livingstone, 2001; Santos et al., 2004). In addi-tion, LPO estimation in particular has also been found tohave a high predictive importance as revealed from a cred-ible number of research papers describing its suitability as abiomarker of effect (Lackner, 1998; Ahmad et al., 2000,2004, 2005; Santos et al., 2004).
Previous field studies using European eel (Anguilla angu-
illa L.) as a model, on harbour water and paper mill effluentcontaminated river exposure under natural environmentalcondition as well as laboratory studies on sub-lethal naph-thalene, b-naphthoflavone/copper or chromium exposuresshowed that antioxidants are useful biomarkers of pollu-tion. Moreover, a correlation between exposure and bio-marker response, especially the induction of antioxidantswas also observed (Santos et al., 2004, 2006; Ahmadet al., 2005).
As per the best of our knowledge, none of the previouslypublished papers includes PF water-body assessment in thiscontext. Keeping in view the applicability and suitability ofA. anguilla L. oxidative stress responses for different classesof compounds, the same approach was applied in the pres-ent study as a rational use for PF monitoring which is
likely to present a complex mixture of pollutants. Theduration of exposure (48 h) is based on our previous fieldinvestigation (Ahmad et al., 2004; Santos et al., 2004,2006) showing measurable oxidative stress responses at48 h exposure. Thus, A. anguilla were caged and plungedin situ at five study sites located at increasing distancesfrom the entrance point of the Certima River, as a mainsource of contamination, following the same exposurestrategy as adapted by Fenet et al. (1996, 1998), Doyotteet al. (2001), Livingstone et al. (2000) and Pacheco andSantos (1998) for different aquatic ecosystems biomonitor-ing using A. anguilla and other fish species.
The study was focussed on the role of antioxidantenzymes such as catalase (CAT), glutathione peroxidase(GPX), glutathione S-transferase (GST) and non-enzymaticantioxidants such as reduced glutathione (GSH) againstperoxidative damage (measured as LPO increase). It isintended to evaluate the relationships between different PFsite exposure and the A. anguilla oxidative stress responsemodulation in the gill, kidney and the liver with a key ques-tion of various antioxidant status against peroxidative dam-age. Moreover, the assessment A. anguilla organ specificitywas also intended. In addition, the suitability and sensitivityof A. anguilla oxidative stress biomarkers in early detectionfor the freshwater ecosystem health was evaluated.
2. Materials and methods
2.1. Chemicals
1-Chloro-2,4-dinitrobenzene (CDNB), 2,5-dithiobis-tet-ranitrobenzoic acid (DTNB), 2-thiobarbituric acid (TBA),trichloroacetic acid (TCA), NADPH were purchased fromSigma (Spain) and other routine chemicals and reagents(analytical grade) were purchased from local sources.
2.2. Test animals
European eels (A. anguilla L.) with an average weight49.50 ± 0.6 g were captured from a non-polluted Aveirolagoon area – Murtosa, Portugal. Eels were acclimatedfor 7 days in 160-l aquaria under standard laboratoryconditions as described by Santos and Pacheco (1996).Briefly, fish were kept under a natural photoperiod at roomtemperature in aerated, filtered, dechlorinated and recircu-lating tap water with the following physico-chemicalconditions: salinity 0&, pH 7.4 ± 0.2, dissolved oxygen8.67 ± 0.5 mg/l. Fish were fed neither during laboratoryadaptation nor during field exposure.
2.3. Experimental design
In order to evaluate the A. anguilla oxidative stressresponses to PF water contamination, five groups of eelwere caged and plunged into the five selected sites of PFwater-body (sites A–E), differing in their distances to themain known pollution source (Certima River), for 48 h

954 I. Ahmad et al. / Chemosphere 65 (2006) 952–962
(Fig. 1). The study sites (A–E) are affected primarily by thepollutants input from the Certima River, since point-spe-cific sources around the study sites are not known, exclud-ing agricultural leachates and domestic discharges. Onemore site as a reference (site R) was selected at the CertimaRiver spring, without any industrial and domestic sewagecontamination. The geographical location of study siteshas been described in Table 1. The experiment was carriedout in December 2004.
Fish were placed into an 80-l net cage, transported andplunged into PF pre-selected sites according to Santoset al.’s (2006) previous technique, adopted for this fish spe-cies. Fish cages were maintained 15 cm from the bottom toavoid a direct contact with the sediment. After 48 h expo-sure, A. anguilla liver, kidney and gill were sampled fromeach fish and individually assayed for LPO, CAT, GPX,GST and GSH.
2.4. Water physico-chemical parameters
Water samples collected from the bottom of each PFstudy site were analyzed (Table 2) for some physico-chemi-
Fig. 1. Representation of A. anguilla L. caging sites (A–E) on Patei
Table 1Geographical positions of different Pateira de Fermentelos study sites
Geographical positions Exposure sites
Site R Site A Site B
Longitude 40�2005000N 40�3303400N 40�3402Lattitude 008�2400800W 008�3003700W 008�300
cal characteristics such as temperature, pH, conductivity,dissolved oxygen (DO), biological oxygen demand (BOD),total solid (TS), total dissolved solid (TDS), total suspendedsolids (TSS) and depth using procedures described inAPHA (1998). Other parameters such as ammonium,nitrate, nitrite, phosphate, carbonate hardness (acid-binding capacity). Total hardness tests were performedusing Compact laboratory for water testing (Merck,Germany).
2.5. Estimation of lipid peroxidation (LPO) in fish tissues
LPO was estimated in liver, kidney and gill of allexposed fish. The gills were excised out of the sacrificedfish, gill rackers removed and gill lamellae were used forhomogenization. All the tissues were homogenized inchilled phosphate buffer (0.1 M, pH 7.4) containing KCl(1.17%), using a Potter–Elvehjem homogenizer. The LPOwas determined in the homogenate as per the method ofUtley et al. (1967), modified and adapted by Fatimaet al. (2000). The absorbance of each aliquot was measuredat 535 nm. The rate of lipid peroxidation was expressed as
ra de Fermentelos water-body in the central region of Portugal.
Site C Site D Site E
400N 40�3403400N 40�34044.6000N 40�35019.5500N4800W 008�3100600W 008�31038.0600W 008�31044.1900W

Table 2Physico-chemical analysis of water at different Pateira de Fermentelosstudy sites
Physico-chemicalparameters
Exposure sites
Site R Site A Site B Site C Site E
Temperature (�C) 10.9 9.3 9.3 9.1 9.1pH 7.937 7.999 8.175 9.640 8.203Conductivity (ls/cm) 289 492 435 151 449DO (mg/1) 10.34 8.86 6.62 8.45 8.84BOD (mg/1) 2.33 1.38 0.56 1.03 3.73Ammonium (mg/1) 0.5 0.5 0.8 0.3 0.3Nitrate (mg/1) 0 0 0 0 0Nitrite (mg/1) 0.075 <0.025 >0.5 <0.025 0.5Phosphate (mg/1) 0 0.25 0.125 0.25 0.125Carbonate hardness
(acid-binding capacity)(mmol/l)
0.9 2.5 2.3 1.0 2.1
Total hardness (mmol/l) 0.95 2.85 2.4 1.4 2.4Total solids (mg/l) 1388.8 294 440 136 188Total dissolved solids
(mg/l)170 256 122 124 14
Total suspended solids(mg/l)
1218 38 318 12 174
Depth (m) 1.0 1.5 2.4 2.25 1.50
I. Ahmad et al. / Chemosphere 65 (2006) 952–962 955
nmol of thiobarbituric acid reactive substance (TBARS)formed per hour per milligram of protein using a molarextinction coefficient of 1.56 · 105 M�1 cm�1.
2.6. Antioxidant evaluation
Antioxidant parameters were measured on the postmitochondrial supernatant (PMS) fraction after tissuehomogenization as described above.
2.6.1. Preparation of PMS
PMS was obtained according to the method of Ahmadet al. (2000). Briefly, the homogenate was filtered througha muslin cloth and centrifuged at 800g for 5 min at 4 �Cto separate the nuclear debris. The supernatant was centri-fuged in a refrigerated ultracentrifuge (Optima TL, Beck-man) at 10000 rpm for 30 min at 4 �C to obtain the PMSwhich was used for biochemical analysis.
2.6.2. Glutathione peroxidase (GPX) activity
GPX activity was assayed according to the methoddescribed by Mohandas et al. (1984) as modified by Atharand Iqbal (1998). The assay mixture consisted of 1.44 mlphosphate buffer (0.05 M, pH 7.0), 0.1 ml EDTA(1 mM), 0.1 ml sodium azide (1 mM), 0.05 ml glutathionereductase (1 IU/ml), 0.1 ml GSH (1 mM), 0.1 ml NADPH(0.2 mM), 0.01 ml H2O2 (0.025 mM) and 0.1 ml ofPMS (10%) in a total volume of 2 ml. Oxidation ofNADPH was recorded spectrophotometrically (JascoUV/VIS, V-530) at 340 nm at room temperature. Theenzyme activity was calculated as nmol NADPH oxi-dized/min/mg of protein, using a molar extinction coeffi-cient of 6.22 · 103 M�1 cm�1.
2.6.3. Catalase (CAT) activity
CAT activity was assayed by the method of Claiborne(1985) as described by Giri et al. (1996). Briefly, the assaymixture consisted of 1.95 ml phosphate buffer (0.05 M,pH 7.0), 1 ml hydrogen peroxide (0.019 M) and 0.05 ml10% PMS in a final volume of 3 ml. The change in absor-bance was recorded at 240 nm. CAT activity was calculatedin terms of nmol H2O2 consumed/min/mg protein.
2.6.4. Glutathione S-transferase (GST) activity
GST activity was determined by the method of Habiget al. (1974) with some modifications (Raisuddin et al.,1994). The reaction mixture consisted of 1.65 ml sodiumphosphate buffer (0.1 M, pH 6.5), 0.1 ml reduced glutathi-one (1 mM), 0.05 ml CDNB (1 mM) and 0.2 ml PMS (10%)in a total volume of 2 ml. The change in absorbance wasrecorded at 340 nm and enzyme activity calculated as nmolCDNB conjugate formed/min/mg protein using a molarextinction coefficient of 9.6 · 103 M�1 cm�1.
2.6.5. Estimation of reduced glutathione (GSH)
GSH content was determined by the method of Jollowet al. (1974) as described by Iqbal et al. (1999). PMS(1 ml) was precipitated with 1 ml of sulfosalicylic acid(4.0%). The samples were kept at 4 �C for 1 h and then sub-jected to centrifugation at 3000 rpm for 15 min at 4 �C. Theassay mixture contained 0.5 ml of filtered aliquot, 2.3 ml ofsodium phosphate buffer (0.1 M, pH 7.4) and 0.2 mlDTNB (stock = 100 mM in 0.1 M sodium phosphate buf-fer, pH 7.4) in a total volume of 3 ml. The reaction productoptical density (OD) was read immediately at 412 nm on aspectrophotometer.
2.7. Protein measurement
Total protein contents were determined according to theBiuret method (Gornall et al., 1949), using bovine serumalbumin (E. Merck-Darmstadt, Germany) as a standard.
2.8. Statistical analysis
A statistical data analysis was done using the Statisticasoftware (StatSoft, Inc., Tulsa, OK) on five eels each(n = 5). The final results were based on the mean of the fiveindividual eels by each group. The assumptions of normal-ity and homogeneity of data were verified (p > 0.05) usingthe Shapiro–Wilk and Levene test respectively. Analysisof variance was followed by the LSD test to compareresults between fish groups (Zar, 1996) and the significanceof the results was ascertained at p < 0.05.
3. Results
3.1. Physico-chemical parameter of water (Table 2)
The water temperature showed no important variationsneither to reference site nor PF study sites. All the sites

956 I. Ahmad et al. / Chemosphere 65 (2006) 952–962
were alkaline in nature with an incremental pH trend fromsite A (minimum) to site C (maximum), followed by adecrease at site E. Conductivity measurement showed nomajor differences among all the study sites. Concerningthe DO level, the observed variation ranged from 6.62 (siteE) to 10.34 mg/l (site R). BOD also showed a slight varia-tion along all the sites, being highest at site E. Among theammonium, nitrates, nitrites, phosphate, carbonates hard-ness and total hardness parameters, only nitrite concentra-tion showed its considerable presence at sites B and E. Interms of TS and TSS, major differences were measuredsince a higher level was detected at the reference site. Therecorded depth was minimum at the reference site, whereasPF study sites showed an increase from sites A to C, fol-lowed by a decrease at site E.
Site R Site A Site B Site C Site E 0
5
10
15
20
nmol
H2O
2co
nsum
ed/m
in/ m
g pr
otei
n
*
* *
(CAT)
0
100
200
300
nmol
CD
NB
form
ed/m
in/m
gpr
otei
n * (GST) *
0
0.5
1
1.5
2 (LPO)
nmol
TBAR
S fo
rmed
/hr/m
gpr
otei
n
Site R Site A Site B Site C Site E
Site R Site A
Fig. 2. Represents CAT, as expressed in nanomol H2O2 consumed/min/mg prGST, as expressed in nanomol CDNB formed/min/mg protein; GSH, as expreleased during 1 h incubation in gill PMS of A. anguilla L. exposed to Patei*p < 0.05 when compared with the values of reference (site R).
3.2. Oxidative stress profile
Biochemical findings have been described in an organ-specific manner, keeping in view only significant changesat PF exposure sites in comparison to reference site (siteR). Site D exposed fish could not be processed for variousbiochemical analyses because of the cage disappearanceafter 48 h exposure.
3.2.1. Gill responses (Fig. 2)
In general, no clear relation could be establishedbetween gill oxidative stress responses and the distance tothe main source of pollution (river Certima). However,an overall induction of antioxidant enzymes is perceptible.Specifying, site A exposure induced a significant GST
0
30
60
90
120
150nm
olN
ADPH
oxi
dize
d/m
in/m
g pr
otei
n *
(GPX)
0
0.005
0.01
0.015
nmol
/g ti
ssue
(GSH)
Site R Site A Site B Site C Site E
Site R Site A Site B Site C Site E
Site B Site C Site E
otein; GPX, as expressed in nanomol NADPH oxidized/min/mg protein;ressed in nanomole/g tissue and LPO, as expressed in nanomole TBARSra de Fermentelos water-body for 48 h. The significance level observed is
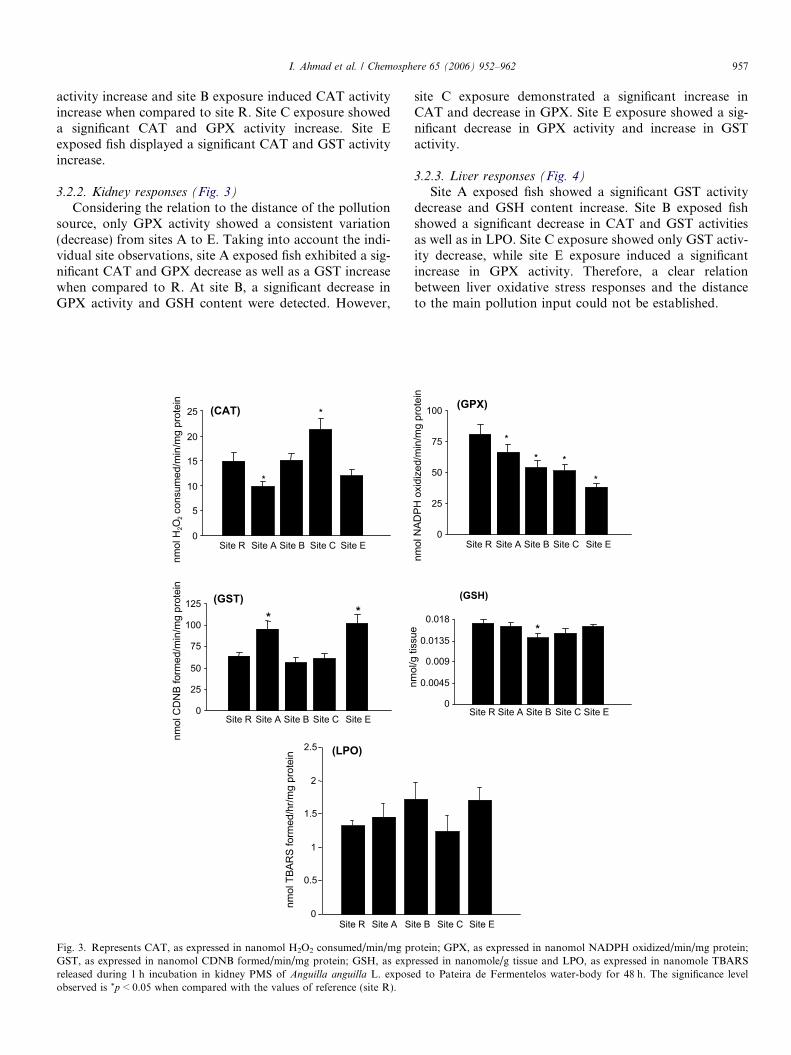
I. Ahmad et al. / Chemosphere 65 (2006) 952–962 957
activity increase and site B exposure induced CAT activityincrease when compared to site R. Site C exposure showeda significant CAT and GPX activity increase. Site Eexposed fish displayed a significant CAT and GST activityincrease.
3.2.2. Kidney responses (Fig. 3)
Considering the relation to the distance of the pollutionsource, only GPX activity showed a consistent variation(decrease) from sites A to E. Taking into account the indi-vidual site observations, site A exposed fish exhibited a sig-nificant CAT and GPX decrease as well as a GST increasewhen compared to R. At site B, a significant decrease inGPX activity and GSH content were detected. However,
Site R Site A Site B Site C Site E 0
5
10
15
20
25
nmol
H2O
2co
nsum
ed/m
in/m
g pr
otei
n
(CAT)
*
*
0
25
50
75
100
125
nmol
CD
NB
form
ed/m
in/m
g pr
otei
n
* * (GST)
0
0.5
1
1.5
2
2.5
nmol
TBAR
S fo
rmed
/hr/m
gpr
otei
n (LPO)
Site R Site A Site B Site C Site E
Site R Site A S
Fig. 3. Represents CAT, as expressed in nanomol H2O2 consumed/min/mg prGST, as expressed in nanomol CDNB formed/min/mg protein; GSH, as expreleased during 1 h incubation in kidney PMS of Anguilla anguilla L. exposeobserved is *p < 0.05 when compared with the values of reference (site R).
site C exposure demonstrated a significant increase inCAT and decrease in GPX. Site E exposure showed a sig-nificant decrease in GPX activity and increase in GSTactivity.
3.2.3. Liver responses (Fig. 4)Site A exposed fish showed a significant GST activity
decrease and GSH content increase. Site B exposed fishshowed a significant decrease in CAT and GST activitiesas well as in LPO. Site C exposure showed only GST activ-ity decrease, while site E exposure induced a significantincrease in GPX activity. Therefore, a clear relationbetween liver oxidative stress responses and the distanceto the main pollution input could not be established.
0
25
50
75
100nm
olN
ADPH
oxi
dize
d/m
in/m
g pr
otei
n
* *
(GPX)
*
*
*
0
0.0045
0.009
0.0135
0.018
nmol
/g ti
ssue *
(GSH)
Site R Site A Site B Site C Site E
ite B Site C Site E
Site R Site A Site B Site C Site E
otein; GPX, as expressed in nanomol NADPH oxidized/min/mg protein;ressed in nanomole/g tissue and LPO, as expressed in nanomole TBARSd to Pateira de Fermentelos water-body for 48 h. The significance level

Site R Site A Site B Site C Site E0
25
50
75
100
nmol
H 2O
2co
nsum
ed/m
in/m
g pr
otei
n*
(CAT)
0
10
20
30
40
nmol
NAD
PHO
xidi
zed/
min
/mg
prot
ein *
(GPX)
0
100
200
300
nmol
CD
NB
form
ed/m
in/m
g pr
otei
n
* * *
(GST)
0
0.006
0.012
0.018
0.024
0.03
nmol
/g ti
ssue
*(GSH)
0
0.25
0.5
0.75
1
nmol
TBAR
S fo
rmed
/hr/m
gpr
otei
n
*
(LPO)
Site R Site A Site B Site C Site E
Site R Site A Site B Site C Site E Site R Site A Site B Site C Site E
Site R Site A Site B Site C Site E
Fig. 4. Represents CAT, as expressed in nanomol H2O2 consumed/min/mg protein; GPX, as expressed in nanomol NADPH oxidized/min/mg protein;GST, as expressed in nanomol CDNB formed/min/mg protein; GSH, as expressed in nanomole/g tissue and LPO, as expressed in nanomole TBARSreleased during 1 h incubation in liver PMS of Anguilla anguilla L. exposed to Pateira de Fermentelos water-body for 48 h. The significance level observedis *p < 0.05 when compared with the values of reference (site R).
958 I. Ahmad et al. / Chemosphere 65 (2006) 952–962
4. Discussion
The physico-chemical analysis has long been employedto assess the water quality. In the current study, waterquality parameters are, in general, on acceptable levels con-sidering criteria given in APHA (1998) and in Aquamerck�
(Germany) guide lines, as well as the A. anguilla require-ments in particular. However, some exceptions as observedfor pH, BOD and nitrite levels must be considered. WaterpH showed a considerably high level at site C (9.640) whichcan be assumed as an indication of strongly polluted toextremely polluted condition. All the other sites presentedpH levels in a range belonging either to an unpolluted orto slightly polluted state. In the direction of pollutionmarking, BOD levels revealed site E as the most oxygendemanding, suggesting the existence of a moderate pollu-tion state. Nitrite levels evidenced a moderately polluted
state at sites B and E. In terms of depth, slight differenceswere found along PF study sites; however, those differenceswere inversely related to DO levels, constituting a possibleexplanation for DO variations.
Considering the previous water quality characterization,the observed fish oxidative stress responses are mainly dis-cussed according to the different distances to the main pol-lution source. Nevertheless, some site-specific responses arealso discussed keeping in view the respective water qualityvariables.
Fish were not fed during holding period mainly to avoidthe water quality depletion. We cannot exclude the hypoth-esis that starvation may pre-dispose fish to oxidative stress.However, results have been compared with control groupskept in the same conditions, reducing the relevance of theprevious factor. In addition, fish starvation and fish withoutfood often occur in the aquatic environment, mainly in winter.

I. Ahmad et al. / Chemosphere 65 (2006) 952–962 959
4.1. Gill responses
An overall antioxidant enzyme inducing trend wasobserved in the present gill responses. However, this trendwas not clearly related to the distance of the main pollutionsource. Thus, CAT induction provides an indication of thehigher H2O2 amount production at sites B, C and E sug-gesting the presence of redox active components in thePF water-body; however, GPX activity only corroboratesthis explanation for site C. Hence, fish exposed to sites Band E seem to cope efficiently with H2O2 production onlythrough CAT activity as no peroxidative damage wasobserved, though, GPX involvement cannot be ignoredsince a slight induction was observed at those sites, despitebeing statistically non-significant.
The absence of CAT induction at site A (closest to themain pollution source) appears to be unexpected since cur-rent gill GST as well as some kidney and liver responses, asdiscussed in the respective sections, showed the presence ofpollutants at site A. In the author’s opinion it can demon-strate a fish incapability to induce gill CAT activity due toa high concentration of pollutants specific to CAT in-hibition.
Detoxification enzymes, such as GST, helps in eliminat-ing reactive compounds by forming conjugates with gluta-thione and subsequently eliminating them as mercapturicacid, thereby protecting cells against ROS induced damage(Rodriguez-Ariza et al., 1991). A high level of GST wasobserved at sites A and E suggesting an activation of gilldetoxification processes probably due to the presence oforganic contaminants. This statement is partially in agree-ment with BOD data, since the highest oxygen demand wasmeasured at site E. The conjugation of GSH with a xeno-biotic can occur either spontaneously, or can be catalysedby GST (Elia et al., 2003). Thus, current results suggest aGST catalysed conjugation for sites A and E exposure.Moreover, the unchanged GSH levels, as observed in thepresent study, indicates that this potent free radical scaven-ger was not a limiting factor for GST or GPX activities.
As a contributory part for water pollutants inducedstress, the occurrence of high nitrite levels can be an impor-tant source of pro-oxidants for fish (Das et al., 2004), lead-ing to nitric radicals production (nitrosative stress) asdemonstrated in mammals by Lijima et al. (2003). In thisdirection, antioxidant responses observed at site B (CATinduction) and at site E (GST induction) could be corre-lated with the nitrite levels detected at these particular sites.However, Tomosso and Grosell (2005) demonstrated thatfreshwater adapted A. anguilla exhibit a high degree ofresistance to NO�2 due to an absent or minimal gill Cl�
uptake activity. Thus, the observed eel’s antioxidantresponses should be attributable mainly to other classesof compounds subsiding the nitrite effects.
Since the typical reaction during oxidative stress is per-oxidative damage to unsaturated fatty acids, the oxidativestress response could be conveniently used as biomarkersof oxidative stress inducing chemical pollutants (Ahmad
et al., 2004; Santos et al., 2006). Thus, no gill LPO induc-tion may be explained by an effective antioxidant actionagainst the peroxidative PF pollutant’s potential.
4.2. Kidney response
An overall trend on kidney stress responses is notdiscernible, since each studied parameter displayed a par-ticular pattern. Therefore, the CAT activity displayed adecrease at site A and an increase at site C, reflecting anincremental trend in relation to the distance from PFentrance (from sites A to C). The significant CAT inhibi-tion (site A) could be attributed to a high production ofsuperoxide anion radical, which has been reported to inhi-bit CAT activity in case of excess production (Kono andFridovich, 1982). In addition, the heavy metal presencepreviously reported by Almeida (1998) and their role inCAT activity decrease should also be considered, sinceRadi and Matkovics (1988) and Ahmad et al. (2000)reported the CAT decrease due to a high concentrationof copper.
GPX activity is reported to be induced by environmentalpollutants (Radi et al., 1985). However, its protectionagainst ROS requires more attention (Aksnes and Njaa,1981; Hasspielar et al., 1994). Accordingly, the presentresults showed a regular kidney GPX activity decreasefor all exposure sites, displaying a GPX decreasing patternin concomitance to the increasing distance from the mainpollution source. Bainy et al. (1996) has also reported aGPX decrease in Nile tilapia from a polluted lake, but anappropriated explanation was not provided. On the otherhand, Janssens et al. (2000) found that general metabolicactivity depletion associated with the depth increase maycause a GPX activity decrease. This explanation can beapplicable to current observations since all PF sites werehigher in depth as compared to the adopted reference site.This condition, in conjugation with a low water transpar-ency, provides a less stimulating environment renderingeels more unperturbed and consequently reducing theirmetabolic activity. According to Janssens et al. (2000) theCAT activity behaves in a different manner being affectedneither by the metabolic activity nor by the depth, corrob-orating our current CAT findings.
Kidney GST showed a significant increase at sites A andE as observed in gills; thus, the same explanation can besuggested as described above for gills. The decrease inGSH content observed for site B exposed fish may be dueto an increased use of GSH in spontaneous conjugationprocesses and an inefficient GSH regeneration. This GSHreduction coincides with the highest LPO level, though sta-tistically insignificant, revealing a potential risk increasedue to this non-enzymatic antioxidant depletion. Surpris-ingly, kidney LPO measurement revealed that fish wereable to cope with the pollution stress, despite the depletionon some antioxidants. This is in agreement with Cossuet al. (2000) who stated that LPO increase cannot be pre-dicted only on the basis of antioxidant depletion.

960 I. Ahmad et al. / Chemosphere 65 (2006) 952–962
4.3. Liver response
Antioxidant responses in liver from exposure sites A to Eshowed no defined pattern suggesting that all the study sitesare not equally polluted. Thus, CAT activity showed adecrease at site B indicating the presence of CAT inhibitorsas described above for kidney at site A. The significant GPXincrease at site E and no observable effect at other sites’exposure may be due to the enzyme inducing effect ofH2O2 on that particular site. The presence of mixtures ofchemicals on PF water seems to affect the crucial roleof GPX protection and therefore, current GPX responsesreflect changes on the GPX inhibitors/inducers balancealong the PF area, favouring the later on site E.
GST activity decrease displayed at sites A–C suggests asignificant reduction of fish capacity to detoxify chemicals.Although GST induction has been widely demonstrated fol-lowing exposure to some organic contaminants (Eggenset al., 1995; Goksøyr, 1995; Beyer et al., 1996; Peterset al., 1997; Grinwis et al., 2000; Schlezinger and Stegeman,2000; Stephensen et al., 2000), its inhibition has also beenreported as a non-specific response to chemical challenge(Regoli et al., 2003).
GSH exhibited a significant increase at site A and unal-tered levels at the other sites. Hence, a GST decrease wasnot related to the liver GSH content suggesting a spontane-ous GSH conjugation to xenobiotics at site A. In additionto its role as a cofactor for GPX and GST, GSH is by itselfan effective protectant capable of oxyradical quenching(Ross, 1988). Therefore, the high liver GSH contentobserved in A. anguilla at site A indicates a protective rolefor GSH against chemical contaminants induced oxidativestress. A similar observation of high GSH levels in fishexposed to polluted sediment and river water have beenreported by Di Giulio et al. (1993) and Pandey et al. (2003).
The current liver LPO decrease at site B may be theresult of a decreased DO level at this site (though in anacceptable level) when compared to the reference site. Thisfinding agrees with Janssens et al.’s (2000) observations ondeep-sea fish which were less prone to oxidative dangerbecause of their poorly oxygenated environment andrevealing that, under certain conditions, hypoxic environ-ments can be beneficial as it relieves fish from a major oxi-dative stress risk.
4.4. Organ specificity
Current results demonstrated an organ-specific antioxi-dant modulation, varying at different exposure sites.Organ-specific toxic responses are related to their anatomiclocation determining the exposure route and distribution ofpollutants, as well as their defensive capacity. This is partic-ularly evident on CAT activity as an overall increase wasobserved in gill, whereas liver and kidney showed no definedpattern. Both GPX and GST data demonstrated the pres-ence of chemical inducers in the PF water; however, clearorgan specificity is evident on their responses. In this direc-
tion, GPX activity showed a decrease in kidney, whereas thegill and the liver displayed their ability to induce thisenzyme. On the other hand, GST activity showed a decreasein liver, while gill and kidney exhibited their ability toinduce this conjugase. Therefore, the kidney seems to bethe most vulnerable organ to GPX inhibition whereas theliver is most sensitive to GST inhibition, probably due toits specific propensity to bioaccumulate either chemicalinhibitors or substrates up to inhibitory levels. Moreover,neither kidney GPX nor liver GST depletion can be justifiedby GSH overuse and insufficient regeneration.
The overall analysis of GPX and GST data suggests twodifferent explanations occurring either simultaneously oralternatively: first, the chemicals responsible for GPXand GST inhibition are different in nature; second, anorgan-specific detoxification skill determining the overallresponses in each organ. The latter explanation is corrobo-rated by the difference in baseline levels (recorded for thereference site) among organs found for GPX or GST.Thus, kidney showed a higher basal GPX level than didthe liver (around 80 vs. 24 nmol NADPH oxidized/min/mg protein), whereas the opposite was observed in GST(around 60 vs. 250 nmol CDNB formed/min/mg protein).
GSH showed its protective capability (though point spe-cific) in kidney and liver, signed either by its increase (liverat site A) or decrease (kidney at site B); being the latter caseprobably related with an insufficient glutathione regen-eration.
Fish gill is in close contact with environmental watercontaminants, being the main absorption organ for thoseaquatic toxicants (Sayeed et al., 2000). In our earlier fieldinvestigation on the recovery of bleached kraft paper milleffluent affected areas, we observed gill highest susceptibilitytowards peroxidative damage compared to kidney and liver,rendering gill antioxidants less effective (Santos et al., 2004,2006). Moreover, studies have shown that gill and kidneyantioxidant enzymes and non-enzymatic antioxidant mole-cules are less efficient than liver, thus increasing their vulner-ability towards reactive oxidative radicals (Di Giulio et al.,1989; Winston and Di Giulio, 1991; Ahmad et al., 2000;Santos et al., 2004). Despite the previous statements, thepresent study showed significant gill antioxidant inductioncoupled with no peroxidative damage, demonstrating thegill efficiency to cope with the rapid contact of pro-oxidantspresent in the PF water body. In addition, a long-termexposure study may be suggested to validate the role of anti-oxidants to this particular ecosystem exposure. The non-accepting trend of antioxidant induction in kidney and livercould be explained through differences in exposure routeand pollutant uptake which is corroborated by Pitchardand Bend (1984) who stated that kidney is the target organfor chemicals after being taken up through the gill.
5. Conclusions
The present results demonstrate that, despite the com-plexity of A. anguilla antioxidant mechanisms aggravated

I. Ahmad et al. / Chemosphere 65 (2006) 952–962 961
by the presence of mixtures of contaminants, the assessedbiomarkers were able to express site-specific responses,demonstrating their utility on freshwater health assess-ment. The presence of pollutants in the PF water wasdenunciated even without a clear relation to the main pol-lution source distance. In this direction, the occurrence ofnon-pointed pollution sources and their contribution forantioxidant modulation should also be considered.
Fish antioxidant responses in each organ proved to bedependent either on the anatomic position determiningthe uptake and distribution of xenobiotics or organ-specificphysiology. The organ specificity was evident for eachparameter but without a clear pattern. However, all theorgans revealed a similar resistance to peroxidative dam-age, suggesting that the antioxidants are more responsivebiomarkers than LPO for short-term exposure.
Besides the activation of antioxidant enzymes (asobserved in gill), their inhibition (as observed in kidneyand liver) should also be considered as a clear indication ofpollution presence and environmental health degradation.The overall fish responses should be explained by the typeand degree of contamination as well as their interference toeach exposure site. Moreover, the presence of different clas-ses of contaminants both at the water column and sedimentscan undergo diverse dilution and degradation along thewatercourse resulting in site specific chemical interactionsand consequent antagonistic or synergistic effects.
Keeping in view all the assessed parameters and theorgan’s responsiveness, the authors suggest, for future bio-monitoring studies, that a multiple organ strategy shouldbe adopted in order to avoid misinterpretations.
A. anguilla in situ trial proves its high ability for fresh-water monitoring, especially on the basis of oxidative stressassessment.
Acknowledgements
The financial supports from FCT (Government of Por-tugal) provided through contract S.F.R.H/B.P.D/3603/2000 and by the Aveiro University Research Institute/CESAM are gratefully acknowledged.
References
Ahmad, I., Hamid, T., Fatima, M., Chand, H.S., Jain, S.K., Athar, M.,Raisuddin, S., 2000. Induction of hepatic antioxidants in freshwaterfish (Channa punctatus Bloch) is a biomarker of paper mill effluentexposure. Biochim. Biophys. Acta 1523, 37–48.
Ahmad, I., Pacheco, M., Santos, M.A., 2004. Enzymatic and non-enzymatic antioxidants as an adaptation to phagocyte-induced damagein Anguilla anguilla L. following in situ harbor water exposure.Ecotoxicol. Environ. Saf. 57, 290–302.
Ahmad, I., Oliveira, M., Pacheco, M., Santos, M.A., 2005. Anguilla
anguilla L. oxidative stress biomarkers responses to copper exposurewith or without b-naphthoflavone pre-exposure. Chemosphere 61,267–275.
Aksnes, A., Njaa, L.R., 1981. Catalase, glutathione peroxidase andsuperoxide dismutase in different fish species. Comp. Biochem. Physiol.69, 893–896.
Almeida, S., 1998. Utilizacao das diatomacias na avaliacao da qualidadedas aguas doces, Ph.D. thesis, Department of Biology, University ofAveiro, Portugal (in Portuguese).
APHA, 1998. In: Clesceri, L.S., Greenberg, A.E., Eaton, A.D. (Eds.),Standard Methods for the Examination of Water and Waste-water,20th ed. American Public Health Association, Washington.
Athar, M., Iqbal, M., 1998. Ferric nitrilotriacetate promotes N-diethylni-tros-amine-induced renal tumorigenesis in the rat: implications for theinvolvement of oxidative stress. Carcinogenesis 19, 1133–1139.
Bainy, A.C.D., Saito, E., Carvello, P.S.M., Junqueria, V.B.C., 1996.Oxidative stress in gill and kidney of Nile tilapia (Oreochromis
niloticus) from a polluted site. Aquat. Toxicol. 34, 151–162.Beyer, J., Sandvik, M., Hylland, K., Fjeld, E., Egaas, E., Aas, E., Skare,
J.U., Goksøyr, A., 1996. Contaminant accumulation and biomarkerresponses in flounder (Platichthys flesus L.) and Atlantic cod (Gadus
morhua L.) exposed by caging to polluted sediments in Sørfjorden,Norway. Aquat. Toxicol. 36, 75–98.
Calado, A.J., Craveiro, S.C., 1995. Notes on the ecology of Synurophy-cean algae found in Portugal. Nord. J. Bot. 15, 641–654.
Calado, A.J., Freitas, A.M., Veloso, V.M., 1991. Algas Pateira deFermentelos numa situacao de Inverno. Rev. Biol. U. Aveiro 4, 55–71.
Claiborne, A., 1985. Catalase activity. In: Greenwald, R.A. (Ed.), CRCHandbook of Methods in Oxygen Radical Research. CRC Press, BocaRaton, FL, pp. 283–284.
Cossu, C., Doyotte, A., Babut, M., Exinger, A., Vasseur, P., 2000.Antioxidant biomarkers in freshwater bivalves, unio tumidus, inresponse to different contamination profiles of aquatic sediments.Ecotoxicol. Environ. Saf. 45, 106–121.
Das, P.C., Ayyappan, S., Das, B.K., Jena, J.K., 2004. Nitrite toxicity inIndian major carps: sublethal effect on selected enzymes in fingerlingsof Catla catla, Labeo rohita and Cirrhinus mrigala. Comp. Biochem.Physiol. C. Toxicol. Pharmicol. 138, 3–10.
Di Giulio, R.T., Washburn, P.C., Wenning, R.J., Winston, G.W., Jewell,C.S., 1989. Biochemical responses in aquatic animals: a review ofdeterminants of oxidative stress. Environ. Toxicol. Chem. 8, 1103–1123.
Di Giulio, R.T., Habig, C., Gallagher, E.P., 1993. Effects of black riverharbour sediments on indices of biotransformation, oxidative stress,and DNA integrity in channel catfish. Aquat. Toxicol. 26, 1–22.
Doyotte, A., Mitchelmore, C.L., Ronisz, D., McEvoy, J., Livingstone,D.R., Peters, L.D., 2001. Hepatic 7-ethoxyresorufin O-deethylaseactivity in eel (Anguilla anguilla) from the Thames Estuary andcomparisons with other United Kingdom estuaries. Mar. Pollut. Bull.42, 1313–1322.
Eggens, M., Bergman, A., Vethaak, D., van der Weiden, M., Celander,M., Boon, J.P., 1995. Cytochrome P4501A indices as biomarkers ofcontaminant exposure: results of a field study with plaice (Pleuronectes
platessa) and flounder (Platichthys flesus) from the southern NorthSea. Aquat. Toxicol. 32, 211–225.
Elia, A.C., Galarini, R., Taticchi, M.I., Dorr, A.J.M., Mantilacci, L.,2003. Antioxidant responses and bioaccumulation in Ictalurus melas
under mercury exposure. Ecotoxicol. Environ. Saf. 55, 162–167.Fatima, M., Ahmad, I., Sayeed, I., Athar, M., Raisuddin, S., 2000.
Pollutant-induced overactivation of phagocytes is concomitantlyassociated with peroxidative damage in fish tissues. Aquat. Toxicol.49, 243–250.
Fenet, H., Casellas, C., Bontoux, J., 1996. Hepatic enzymatic activities ofthe European eel Anguilla anguilla as a tool for biomonitoringfreshwater streams: laboratory and field caging studies. Water Sci.Technol. 33, 321–329.
Fenet, H., Casellas, C., Bontoux, J., 1998. Laboratory and field-cagingstudies on hepatic enzymatic activities in European eel and rainbowtrout. Ecotoxicol. Environ. Saf. 40, 137–143.
Giri, U., Iqbal, M., Athar, M., 1996. Porphyrine-mediated photosensiti-zation has a weak tumor promoting effect in mouse skin: possible roleof in situ generated reactive oxygen species. Carcinogenesis 17, 2023–2028.

962 I. Ahmad et al. / Chemosphere 65 (2006) 952–962
Goksøyr, A., Forlin, L., 1992. The cytochrome P-450 system in fish,aquatic toxicology and environmental monitoring. Aquat. Toxicol. 22,287–312.
Goksøyr, S.G., 1995. Use of cytochrome P4501A (CYP1A) in fish as abiomarker of aquatic pollution. Arch. Toxicol. 17, 80–95.
Gornall, A.C., Bardawill, C.J., David, M.M., 1949. Determination ofserum proteins by means of the biuret reaction. J. Biol. Chem. 177,751–766.
Grinwis, G.C.M., Besselink, H.T., van der Brandof, E.J., Bulder, A.S.,Engelsma, M.Y., Kuiper, R.V., Wester, P.W., Vaal, M.A., Vethaal,A.D., Vos, J.C., 2000. Toxicity of TCDD in European flounder(Platichthys flesus) with emphasis on histopathology and cytochromeP450 1A induction in several organ systems. Aquat. Toxicol. 50, 387–401.
Habig, W.H., Pabst, M.J., Jokoby, W.B., 1974. Glutathione S-transferase.The first enzymatic step in mercapturic acid formation. J. Biol. Chem.249, 7130–7139.
Hasspielar, B.M., Behar, J.V., Di Giulio, R.T., 1994. Glutathine depen-dant defense in channel catfish (Ictalurus punctatus) and brownbullhead (Ameiurus nebulosus). Ecotoxicol. Environ. Saf. 28, 82–90.
Iqbal, M., Giri, U., Giri, D.K., Alam, M.S., Athar, M., 1999. Age-dependent renal accumulation of 4-hydroxy-2 nonenal (HNE)-modifiedproteins following parenteral administration of ferric nitrilotriacetatecommensurates with its differential toxicity: implications for theinvolvement of HNE-protein adducts in oxidative stress and carcino-genesis. Arch. Biochem. Biophys. 365, 101–112.
Janssens, B.J., Childress, J.J., Baguet, F., Rees, J., 2000. Reducedenzymatic antioxidative defense in deep-sea fish. J. Exp. Biol 203,3717–3725.
Jollow, D.W., Mitchell, J.R., Zampagilone, N., Gilete, J.R., 1974.Bromobenzene-induced liver necrosis: protective role of glutathioneand evidence for 3,4-bromobenzeneoxide as the hepatotoxic interme-diate. Pharmacology 11, 151–169.
Kono, Y., Fridovich, I., 1982. Superoxide radicals inhibit catalase. J. Biol.Chem. 257, 5751–5754.
Lackner, R., 1998. Oxidative stress in fish by environmental pollutants. In:Braunbeck, T., Hinton, D.E., Streit, B. (Eds.), Fish Ecotoxicology.Birkhause Verlag, Basel, pp. 203–224.
Lijima, K., Grant, J., McElroy, K., Fyfe, V., Preston, T., McColl, K.E.,2003. Novel mechanism of nitrosative stress from dietary nitrate withrelevance to gastro-oesophageal junction cancers. Carcinogenesis 24,1951–1960.
Livingstone, D.R., 1993. Biotechnology and pollution monitoring: use ofmolecular biomarkers in the aquatic environment. J. Chem. Technol.Biotechnol. 57, 195–211.
Livingstone, D.R., 2001. Contaminant-stimulated reactive oxygen speciesproduction and oxidative damage in aquatic organisms. Mar. Pollut.Bull., 656–666.
Livingstone, D.R., Mitchelmore, C.L., Peters, L.D., O’Hara, S.C.M.,Shaw, J.P., Chesman, B.S., et al., 2000. Development of hepaticCYP1A and blood vitellogenin in eel (Anguilla anguilla) for use asbiomarkers in the Thames Estuary, UK. Mar. Environ. Res. 50, 367–371.
Mohandas, J., Marshall, J.J., Duggins, G.G., Horvath, J.S., Tiller, D.,1984. Differential distribution of glutathione and glutathione relatedenzymes in rabbit kidney. Possible implications in analgesic neurop-athy. Cancer Res. 44, 5086–5091.
Pacheco, M., Santos, M.A., 1998. Induction of liver EROD anderythrocytic nuclear abnormalities by cyclophosphamide and PAHsin Anguilla anguilla L. Ecotoxicol. Environ. Saf. 40, 71–76.
Pandey, S., Parvez, S., Sayeed, I., Haque, R., Bin-Hafeez, B., Raisuddin,S., 2003. Biomarkers of oxidative stress: a comparative study of riverYamuna fish Wallago attu (Bl. & Schn.). Sci. Total Environ. 20 (309),105–115.
Passino, D.R.M., 1984. Biochemical indicators of stress of fishes: anoverview. In: Cairns, V.W., Hodson, P.V., Nriagu, J.O. (Eds.),
Contaminant Effects on Fisheries, vol. 16. Wiley, New York, NY,pp. 37–50.
Peakall, D., Shugart, L., 1993. Biomarker: Research and Application inthe Assessment of Environmental Health. Springer, Berlin.
Peters, L.D., Morse, H.R., Waters, R., Livingstone, D.R., 1997.Responses of hepatic cytochrome P-450 1A and formation of DNA-adducts in juveniles of turbot (Scophthalmus maximus L.) exposed towater-borne benzo[a]pyrene. Aquat. Toxicol. 38, 67–82.
Pitchard, J.B., Bend, J.R., 1984. Mechanisms controlling the renalexcretion of xenobiotics in fish: effect of chemical structure. DrugMetabol. Rev. 15, 655–671.
Radi, A.A.R., Matkovics, B., 1988. Effects of metal ions on theantioxidant enzyme activities, protein content and lipid peroxidationof carp tissues. Comp. Biochem. Physiol. 90C, 69–72.
Radi, A.A.R., Hai, D.Q., Matkovics, B., Gabrielak, T., 1985. Compar-ative antioxidant enzyme study in fresh water fish with different typesof feeding behaviour. Comp. Biochem. Physiol. 81C, 395–399.
Raisuddin, S., Parmer, D., Zaidi, S.I.A., Singh, K.P., Verma, A.S., Seth,P.K., Ray, P.K., 1994. Aflatoxin induces depletion of activities ofphase I biotransformation enzymes in growing rats. Eur. J. DrugMetabol. Pharmacokinet. 19, 163–168.
Regoli, F., Nigro, M., Orlando, E., 1998. Lysosomal and antioxidantresponses to metals in the Antarctic scallop Adamussium colbecki.Aquat. Toxicol. 40, 375–392.
Regoli, F., Winston, G.W., Gorbi, S., Frenzilli, G., Nigro, M., Corsi, I.,Focardi, S., 2003. Integrating enzymatic responses to organic chemicalexposure with total oxyradical absorbing capacity and DNA damagein the European eel Anguilla anguilla. Environ. Toxicol. Chem. 22, 56–65.
Rodriguez-Ariza, A., Dorado, G., Peinado, J., Pueyo, C., Lopez-Barea, J.,1991. Biochemical effects of environmental pollution in fishes fromSpanish south-Atlantic littoral. Biochem. Soc. Trans. 19, 301S.
Ross, D., 1988. Glutathione, free radicals and chemotherapeutic agents.Mechanisms of free radical induced toxicity and glutathione-depen-dent protection. Pharmacol. Therapeut. 37, 231–249.
Santos, M.A., Pacheco, M.G., 1996. Anguilla anguilla L. stress biomarkersrecovery in clean water and secondary-treated pulp mill effluent.Ecotoxicol. Environ. Saf. 35, 96–100.
Santos, M.A., Pacheco, M., Ahmad, I., 2004. Anguilla anguilla L.antioxidants responses to in situ bleached kraft pulp mill effluentoutlet exposure. Environ. Int. 30, 301–308.
Santos, M.A., Pacheco, M., Ahmad, I., 2006. Responses of European eel(Anguilla anguilla L.) circulating phagocytes to an in situ closed pulpmill effluent exposure and its association with organ-specific peroxi-dative damage. Chemosphere 63, 794–801.
Sayeed, I., Ahmad, I., Fatima, M., Hamid, T., Islam, F., Raisuddin, S.,2000. Inhibition of brain Na+, K+-ATPase in freshwater catfish(Channa puncatatus Bloch) exposed to paper mill effluent. Bull.Environ. Contam. Toxicol. 65, 161–167.
Schlezinger, J., Stegeman, J.J., 2000. Induction of cytochrome P450 1A inthe American eel by model halogenated and non-halogenated arylhydrocarbon receptor agonists. Aquat. Toxicol. 50, 375–386.
Stephensen, E., Svavarsson, J., Sturve, J., Ericson, G., Adolfsson-Erici,M., Forlin, L., 2000. Biochemical indicators of pollution exposure inshorthorn sculpin (Myoxocephalus scorpius) caught in four harbourson the southwest coast of Iceland. Aquat. Toxicol. 48, 431–442.
Tomosso, J.R., Grosell, M., 2005. Physiological basis for large differencesin resistance to nitrite among freshwater and freshwater acclimatedeurihaline fishes. Environ. Sci. Technol. 39, 98–102.
Utley, H.C., Bernheim, F., Hachslien, P., 1967. Effects of sulfhydrylreagent on peroxidation in microsome. Arch. Biochem. Biophys. 260,521–531.
Winston, G.W., Di Giulio, R.T., 1991. Prooxidant and antioxidantmechanisms in aquatic organisms. Aquat. Toxicol. 19, 137–161.
Zar, J.H., 1996. Biostatistical Analysis, third ed. Prentice Hall Interna-tional, Inc., USA.




















