
Biochromatography and Molecular Affinity Dijon, May 22-25 ...
Upload
independentCategory
view
1download
0
www.elsevier.com/locate/lifescie
Life Sciences 75 (2004) 1085–1103
Lectinochemical studies on the affinity of Anguilla anguilla
agglutinin for mammalian glycotopes
Albert M. Wua,*, June H. Wub, Tanuja Singha, Jia-Hau Liua, Anthony Herpa
aGlyco-Immunochemistry Research Laboratory, Institute of Molecular and Cellular Biology,
Chang-Gung University, Kwei-san, Tao-yuan, 333, TaiwanbDepartment of Microbiology and Immunology, Chang-Gung University,
Kwei-san, Tao-yuan, 333, Taiwan
Received 13 November 2003; accepted 17 February 2004
Abstract
Anguilla anguilla agglutinin (AAA) is a fucose-specific lectin found in the serum of the fresh water eel.
It is suggested to be associated with innate immunity by recognizing disease-associated cell surface glycans,
and has been widely used as a reagent in hematology and glycobiology. In order to gain a better
understanding of AAA for further applications, it is necessary to elucidate its binding profile with
mammalian glycotopes. We, therefore, analyzed the detailed carbohydrate specificity of AAA by enzyme-
linked lectinosorbent assay (ELLSA) with our extended glycan/ligand collection and lectin-glycan inhibition
assay. Among the glycans tested, AAA reacted well with nearly all human blood group Ah
(GalNAca1! 3[LFuca1! 2]Gal), Bh (Gala1! 3[LFuca1! 2]Gal), H LFuca1! 2Gal) and Leb (Fuca1!2Galh1! 3[Fuca1! 4]GlcNAc) active glycoproteins (gps), but not with blood group Lea
(Galh1! 3[Fuca1! 4]GlcNAc) substances, suggesting that residues and optimal density of a1-2 linked
LFuc to Gal at the non-reducing end of glycoprotein ligands are essential for lectin-carbohydrate interactions.
Blood group precursors, Galh1-3GalNAc (T), GalNAca1-Ser/Thr (Tn) containing glycoproteins and N-linked
plasma gps, gave only negligible affinity. Among the mammalian glycotopes tested, Ah, Bh and H
determinants were the best, being about 5 to 6.7 times more active than LFuc, but were weaker than p-
nitrophenylaFuc indicating that hydrophobic environment surrounding the LFuc moiety enhance the
reactivity. The hierarchy of potency of oligo- and monosaccharides can be ranked as follows: p-nitrophenyl-
aFuc > Ah, Bh and H > LFuc > LFuca1! 2Galh1! 4Glc (2V-FL) and Galh1! 4[LFuca1! 3]Glc (3V-FL),while LNDFH I (Leb hexa-), Lea, Lex (Galh1! 4[Fuca1! 3]GlcNAc), and LDFT (gluco-analogue of Ley)
were inactive. From the present observations, it can be concluded that the combining site of AAA should be
0024-3205/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.lfs.2004.02.016
* Corresponding author. Tel.: +886-3-211-8966 (Lab.); fax: +886-3-211-8456 (Lab.), +886-3-211-8700 (Col.).
E-mail address: [email protected] (A.M. Wu).

A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031086
a small cavity-type capable of recognizing mainly H/crypto H and of binding to specific polyvalent ABH
and Leb glycotopes.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Carbohydrate specificity; Glycoproteins; Lectins; Combining site; Anguilla anguilla
Introduction
Lectins are carbohydrate-binding proteins, some of which have great value as specific probes for
investigating the structure and function of carbohydrate chains of glycoconjugates. The complexity of
glycans and the growing evidence of the importance of glycans in various normal and pathological
conditions present substantial assets to portray carbohydrates as sugar codes in glycobiology (Gabius et
al., 2002; Reuter and Gabius, 1999). Matching glycan diversity, the increasing level of refinement of
our knowledge about lectin specificities epitomizes the sphere of action of the carbohydrate structural
units.
During the past few decades, various fucose-specific lectins have been derived from different
sources revealing the specificity of a-linked LFucose for the blood group H determinant and are widely
used as reagents in blood typing (Judd, 1980; Liener et al., 1986) and histochemistry (Danguy et al.,
1988; Wagner, 1988). Anguilla anguilla agglutinin (AAA), a fucose-specific lectin present in the serum
of eel has been reported to participate in the recognition of bacterial lipopolysaccharides by the animal
innate immunity system suggesting a role for fucose-specific lectins in host defense (Bianchet et al.,
2002).
In previous studies, knowledge of the carbohydrate specificity of AAA was limited to a few
oligosaccharides tested by the inhibition assay (Baldus et al., 1996; Matsumoto and Osawa, 1971;
Springer and Desai, 1971; Watkins and Morgan, 1952) and by binding with crystal lectin (Bianchet et
al., 2002). In order to broaden our knowledge of AAA required for its application and interpretation, it is
necessary to further characterize this lectin. For instance, jacalin was deemed to be an anti-T specific
lectin for decades; however, recent studies revealed that it can also react well with the blood group
precursor type II sequence (Galh1-4GlcNAc) prompting us to reevaluate its binding profile (Wu et al.,
2003b). Thus, systematic analysis of the carbohydrate binding spectrum of a lectin is important
particularly when used as a diagnostic tool. In this study, we have examined: (i) the combining site
of AAA by our biotin/avidin-mediated microtiter plate lectin-enzyme binding assay, which is more
sensitive and rapid (Duk et al., 1994; Lisowska et al., 1996) than the quantitative precipitin assay that we
have used previously (Wu et al., 1992, 1997); (ii) the interactions of AAA with a panel of glycans to
establish its binding profile; (iii) the effect of glycan polyvalency on lectin binding by inhibition of this
lectin-glycan binding with glycans; and (iv) its relative affinity for mammalian glycotopes from our
collections. The results indicate that AAA reacts most strongly with human blood group A (cryptic H), B
(cryptic H), H and Leb active gps, but weakly or not at all with their precursors and other glycoproteins
containing Galh1-3GalNAc (T), GalNAca1-Ser/Thr (Tn) or Galh1-3/4GlcNAc (I/II) gps; it recognizesmainly LFuca1! 2Gal (H) and the crypto H active sequence, but not other Gal/GalNAc glycotopes
frequently found in mammalians. The hierarchy of potency of oligo- and monosaccharides can be ranked
as follows: p-nitrophenyl-aFuc > Ah, Bh and H > LFuc > LFuca1! 2Galh1! 4Glc (2V-FL) and
Galh1! 4[LFuca1! 3]Glc (3V-FL), while LNDFH I (Leb hexa-), Lea, Lex (Galh1!

A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1087
4[Fuca1! 3]GlcNAc), and LDFT (gluco-analogue of Ley) were inactive. Comparison of the inhibition
profile of AAA obtained from this study with those of earlier studies (Baldus et al., 1996; Bianchet et al.,
2002; Matsumoto and Osawa, 1971; Springer and Desai, 1971; Watkins and Morgan, 1952) is also
included.
Material and methods
Lectin and biotinylation of the lectin
Anguilla anguilla agglutinin from fresh water eel was purchased from Sigma Chemical Company (St.
Louis, MO, USA). For AAA biotinylation by biotinamidocaproate-N-hydroxy-succinimide ester (biotin
ester; Sigma), the lectin [200 Ag/250 Al PBS (phosphate buffered saline; 0.14 M NaCl, 0.027 M KCl,
0.081 M Na2HPO4, 0.0014 M KH2PO4, pH 7.3)] was mixed with 400 Al of the biotin ester solution (100Ag biotin ester per 200 Ag lectin) for 30 min at room temperature. The biotinylated lectin was dialyzed
for 2–3 h against ddH2O and overnight against TBS (Tris buffered saline; 0.05 M Tris HCl, 0.15 M
NaCl, pH 7.35). After dialysis, the sample volume was adjusted to 1 ml with TBS and 20 Al of 5%sodium azide (equivalent to 200 Ag/ml lectin in 0.1% NaN3) (Duk et al., 1994; Lisowska et al., 1996).
Glycoproteins and polysaccharide
The blood group A, B, H, Lea, Leb and Ii active substances were purified from human ovarian cyst
fluid (HOC) by digestion with pepsin and precipitation with increasing concentrations of ethanol (Kabat,
1956; Lloyd and Kabat, 1968; Maisonrouge-McAuliffe and Kabat, 1976); the dried ethanol precipitates
were extracted with 90% phenol, the insoluble fraction being named after the blood group substance
(e.g., Cyst Tighe phenol insoluble). The ‘‘Tighe’’ denotes the code of the HOC sample. A similar
procedure was applied to the other HOC collections (e.g., Beach, MSS, JS, Mcdon, N-1 and HOC 350).
The supernatant was fractionally precipitated by addition of 50% ethanol in 90% phenol to the indicated
concentrations (Kabat, 1956). The designation ‘‘10 (or 20)% (ppt)’’ denotes a fraction precipitated from
phenol at an ethanol concentration of 10 or 20%; ‘‘2� ’’ signifies that a second round of phenol
extraction and ethanol precipitation was carried out (e.g., Cyst MSS 10% 2� and Cyst N-1 Lea 20%
2� ). The carbohydrate chains of HOC consist of multiple saccharide branches attached by O-glycosidic
linkages at their reducing ends to serine or threonine residues of the polypeptide backbone (Lloyd and
Kabat, 1968; Wu, 1988). HOC 350, a sialic-acid-rich and Lea-active gp isolated from human ovarian
cyst fluid (Pusztai and Morgan, 1961), was kindly provided by Dr. W.M. Watkins (Department of
Haematology, Imperial College School of Medicine, Hammersmith Hospital, Du Cane Road, London,
W12 ONN, U.K.).
In general, the ‘‘P-1’’ fractions (e.g., Cyst Beach P-1 and Cyst Tighe P-1) represent the
nondialyzable portion of the blood group substances after mild hydrolysis at pH 1.5–2.0 for 2
h which removes most of the LFuc end groups, as well as some blood group A and B active
oligosaccharide side chains (Allen and Kabat, 1959; Kabat et al., 1948; Leskowitz and Kabat, 1954).
The ‘‘Smith degraded’’ products of blood group active substances (e.g., Cyst MSS 1st Smith
degraded, Cyst JS 1st Smith degraded), in which almost all of the sugar groups at the nonreducing
ends are removed, were prepared as described earlier (Wu et al., 1982, 1984). Both P-1 fractions and

A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031088
Smith degraded products from HOC gps exposing the internal structures equivalent to those on the
blood group precursors are defined as ‘‘precursor equivalent gps’’.
Hog gastric mucin #4, a blood group A + H substance (Scheme 1), was derived from crude hog
stomach mucin as described previously (van Halbeek et al., 1982). Treatment of mucin #4 with HCl (pH
2.0, 90 min, 100jC) yields hog gastric mucin #9, while acid hydrolysis (pH 1.5, 100jC, 2 and 5 hrs)
gives hog gastric mucins #14 and #21, respectively. Extensive hydrolysis leads to destruction of blood
group activities (Wu et al., 1984).
Ovine (OSM) and bovine (BSM) submandibular/salivary glycoproteins were purified according to the
method of Tettamanti and Pigman (1968) with some modifications (Herp et al., 1988). About 75% of the
carbohydrate side chains of asialo OSM were GalNAca1-Ser/Thr (Tn) (Herp et al., 1988).
Glycophorin A was prepared from the membranes of outdated human blood group O erythrocytes by
phenol/saline extraction and purified by gel filtration in the presence of SDS (Lisowska et al., 1987).
Desialylation of glycophorin was performed as described below (Tettamanti and Pigman, 1968; Wu and
Pigman, 1977). The Tn-type glycophorin (Tn-glycophorin) was obtained by removing galactose
residues from asialo-glycophorin by periodate oxidation and mild acid hydrolysis (Smith degradation)
(Duk et al., 1994).
Human a1-acid glycoprotein was purchased from Sigma. It contains complex-type N-glycans and the
carbohydrate units of this asialoglycoprotein can be grouped into bi-antennary and tri-antennary Galh1-4GlcNAc (II) structures with and without a fucosyl residue (Fournet et al., 1978). Fetuin (Gibco, Grand
Island, New York), which is the major glycoprotein in fetal calf serum and has six oligosaccharide side
chains per molecule, three of them (of two types) are O-glycosyl-linked to Ser or Thr residues of the
protein core, and the others are triantennary II (Galh1-4GlcNAc) N-glycans and a small amount of the
Galh1,3-linked isomer at the nonreducing terminal ends (Nilsson et al., 1979).
Mucus gp (or native bird-nest gp), the so-called nest-cementing substance (Wieruszeski et al., 1987)
from the salivary gland of Chinese swiftlets (genus Collocalia), was extracted with distilled water at
60jC for 20 min from commercial bird-nest substance (Kim Hing Co., Singapore).
The rat sublingual glycoprotein (RSL) was prepared by the method of Moschera and Pigman (1975).
The established carbohydrate side chains were found to be composed of 9, 10, 12, 13, and 15-sugar
Scheme 1. Proposed structure of carbohydrate side chains of Hog A + H gastric glycoproteins (van Halbeek et al., 1982).

A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1089
residues, respectively, and contain sialic acid, N-acetylglucosamine, galactose, and N-acetylgalactosami-
nitol (Slomiany and Slomiany, 1978). RSLmay also contain the Tn reactive determinants (Wu et al., 1995).
Desialylation of sialo-glycoproteins was performed by mild acid hydrolysis in 0.01 N HCl at 80jC for
90 min, and dialysed against distilled water for 2 days to remove small fragments (Tettamanti and
Pigman, 1968; Wu and Pigman, 1977).
Human blood group P1-active glycoprotein, isolated from sheep hydatid cyst (Morgan and Watkins,
1964; Cory et al., 1974), was kindly provided by Dr. W.M. Watkins.
The Pneumococcus type 14 polysaccharide, isolated from Streptococcus pneumoniae capsule, was a
generous gift from the late Dr. E.A. Kabat (Department of Microbiology, Columbia University, New
York, NY, U.S.A.) (Lindberg et al., 1977) and poly-2,8-N-acetylneuraminic acid capsular polysaccharide
from Escherichia coli (colominic acid) was purchased from Sigma.
Monosaccharides and oligosaccharides used for the inhibition assay
DGal, DFuc, LFuc, DMan, DGlc, DGalNAc, DGlcNAc, DAra, LAra, methyl-aFuc, methyl-hFuc, p-NO2phenyl-aFuc, p-NO2phenyl-hFuc, Galh1! 4Glc, Gala1! 4Gal, Galh1! 3GalNAc,
Galh1! 3GlcNAc, and Galh1! 4GlcNAc were purchased from Sigma. 2V-Fucosyllactose, 3V-Fucosyllactose, Lacto-N-fucopentaose I (LNFP I), Lacto-N-fucopentaose II (LNFP II), Lacto-N-
fucopentaose III (LNFP III), Lacto-N-difucohexaose I (LNDFH I), Lactodifucotetraose (LDFT), sialyl
Lewisa, sialyl Lewisx, Lewisx, Lewisa, blood group H disaccharide and blood group A and B
trisaccharides were purchased from Dextra (Berkshire, UK).
Binding assay
Enzyme-linked lectinosorbent binding assay (ELLSA) was performed according to the procedures
described by Duk et al. (1994). The volume of each reagent applied to the plate was 50 Al/well,and all incubations, except for coating, were performed at room temperature (20jC). The reagents,
if not indicated otherwise, were diluted with TBS containing 0.05% Tween 20 (TBS-T). The TBS
buffer or 0.15 M NaCl containing 0.05% Tween 20 was used for washing the plate between
incubations.
The 96-well microtiter plates (Nunc, MaxiSorp, Vienna, Austria) were coated with glycoproteins
at 50 to less than 10 ng (Fig. 1a), and 5000 to less than 1000 ng per well (Fig. 1d) in 0.05 M
carbonate buffer, pH 9.6, overnight at 4jC. After washing the plate, biotinylated lectins (12.5 ng)
were added and incubated for 30 min. The plates were washed to remove unabsorbed lectin and the
ExtrAvidin/alkaline phosphatase solution (Sigma, diluted 1:10,000) was added. After 1 h, the plates
were washed at least four times and incubated with p-nitrophenyl phosphate (Sigma 104 phosphatase
substrate 5 mg tablets) in 0.05 M carbonate buffer, pH 9.6, containing 1 mM MgCl2 (1 tablet/5 ml).
The absorbance was read at 405 nm in a microtiter plate reader, after 2 h incubation with the
substrate.
For inhibition studies, serially diluted inhibitor samples were mixed with an equal volume of lectin
solution containing a fixed amount of lectin. The control lectin sample was diluted two-fold with TBS-T.
After 30 min at 20jC, samples were tested in the binding assay, as described above. The inhibitory
activity was estimated from the inhibition curve and is expressed as the amount of inhibitor (nmol/well)
giving 50% inhibition of binding of the control lectin.
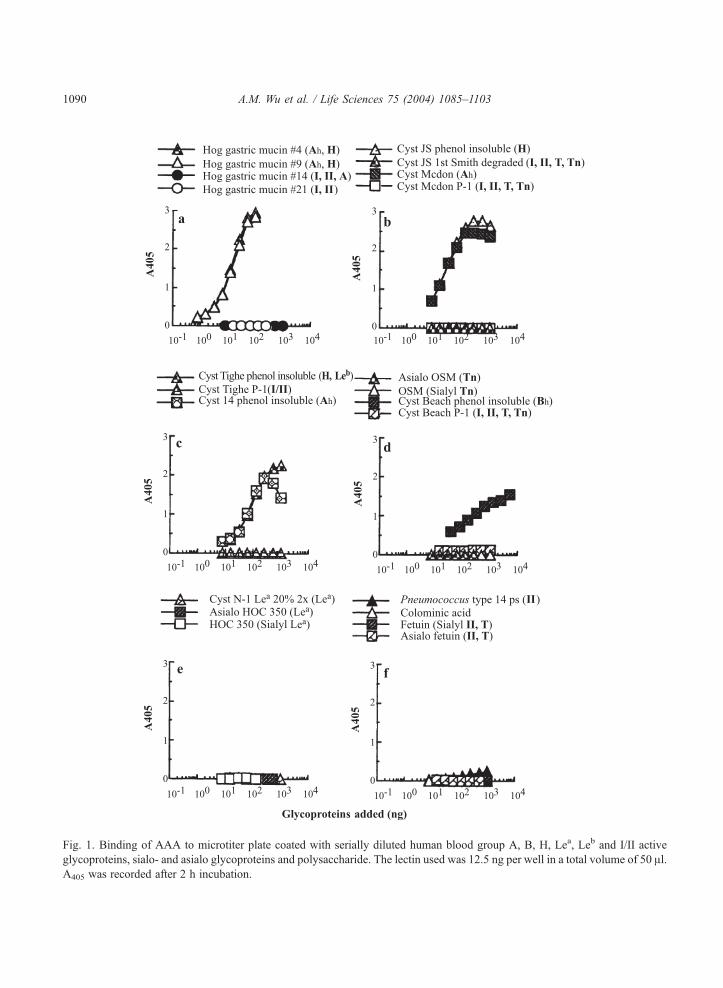
Fig. 1. Binding of AAA to microtiter plate coated with serially diluted human blood group A, B, H, Lea, Leb and I/II active
glycoproteins, sialo- and asialo glycoproteins and polysaccharide. The lectin used was 12.5 ng per well in a total volume of 50 Al.A405 was recorded after 2 h incubation.
A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031090
A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1091
All experiments were done in duplicates or triplicates, and data are presented as the mean value of the
results. The standard deviation did not exceed 12% and in most experiments was less than 5% of the
mean value. The blank wells, where coating or addition of biotinylated lectin was omitted, gave low
absorbance values (below 0.1). It showed that blocking the wells before lectin addition was not
necessary when Tween 20 was present in the TBS.
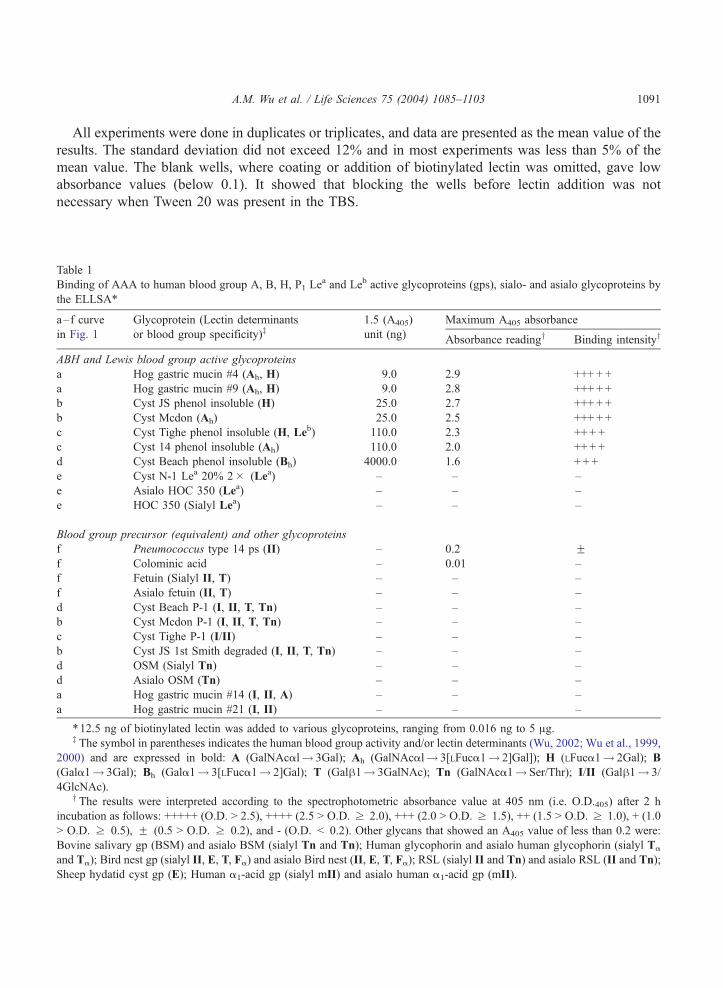
Table 1
Binding of AAA to human blood group A, B, H, P1 Lea and Leb active glycoproteins (gps), sialo- and asialo glycoproteins by
the ELLSA*
a–f curve Glycoprotein (Lectin determinants 1.5 (A405) Maximum A405 absorbance
in Fig. 1 or blood group specificity)z unit (ng) Absorbance readingy Binding intensityy
ABH and Lewis blood group active glycoproteins
a Hog gastric mucin #4 (Ah, H) 9.0 2.9 +++ + +
a Hog gastric mucin #9 (Ah, H) 9.0 2.8 +++ + +
b Cyst JS phenol insoluble (H) 25.0 2.7 +++ + +
b Cyst Mcdon (Ah) 25.0 2.5 +++ + +
c Cyst Tighe phenol insoluble (H, Leb) 110.0 2.3 ++ + +
c Cyst 14 phenol insoluble (Ah) 110.0 2.0 ++ + +
d Cyst Beach phenol insoluble (Bh) 4000.0 1.6 + + +
e Cyst N-1 Lea 20% 2� (Lea) – – –
e Asialo HOC 350 (Lea) – – –
e HOC 350 (Sialyl Lea) – – –
Blood group precursor (equivalent) and other glycoproteins
f Pneumococcus type 14 ps (II) – 0.2 Ff Colominic acid – 0.01 –
f Fetuin (Sialyl II, T) – – –
f Asialo fetuin (II, T) – – –
d Cyst Beach P-1 (I, II, T, Tn) – – –
b Cyst Mcdon P-1 (I, II, T, Tn) – – –
c Cyst Tighe P-1 (I/II) – – –
b Cyst JS 1st Smith degraded (I, II, T, Tn) – – –
d OSM (Sialyl Tn) – – –
d Asialo OSM (Tn) – – –
a Hog gastric mucin #14 (I, II, A) – – –
a Hog gastric mucin #21 (I, II) – – –
*12.5 ng of biotinylated lectin was added to various glycoproteins, ranging from 0.016 ng to 5 Ag.z The symbol in parentheses indicates the human blood group activity and/or lectin determinants (Wu, 2002; Wu et al., 1999,
2000) and are expressed in bold: A (GalNAcal! 3Gal); Ah (GalNAcal! 3[LFuca1! 2]Gal]); H (LFuca1! 2Gal); B
(Gala1! 3Gal); Bh (Gala1! 3[LFuca1! 2]Gal); T (Galh1! 3GalNAc); Tn (GalNAca1! Ser/Thr); I/II (Galh1! 3/
4GlcNAc).y The results were interpreted according to the spectrophotometric absorbance value at 405 nm (i.e. O.D.405) after 2 h
incubation as follows: +++++ (O.D. > 2.5), ++++ (2.5 > O.D. z 2.0), +++ (2.0 > O.D. z 1.5), ++ (1.5 > O.D. z 1.0), + (1.0
> O.D. z 0.5), F (0.5 > O.D. z 0.2), and - (O.D. < 0.2). Other glycans that showed an A405 value of less than 0.2 were:
Bovine salivary gp (BSM) and asialo BSM (sialyl Tn and Tn); Human glycophorin and asialo human glycophorin (sialyl Ta
and Ta); Bird nest gp (sialyl II, E, T, Fa) and asialo Bird nest (II, E, T, Fa); RSL (sialyl II and Tn) and asialo RSL (II and Tn);
Sheep hydatid cyst gp (E); Human a1-acid gp (sialyl mII) and asialo human a1-acid gp (mII).
A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031092
Results
Lectin-glycan interaction
The avidity of AAA for various glycoproteins as studied by the microtiter plate enzyme-linked
lectinosorbent assay (ELLSA) is summarized in Table 1 according to the interaction profiles shown in
Fig. 1. Among the glycoproteins (gps) tested, AAA reacted best with blood group A + H active
glycoproteins from hog gastric mucin (#4 and #9; Fig. 1a; Table 1; Scheme 1), in which less than 9.0 ng
of glycoprotein coating per well was required to interact with 12.5 ng of AAA lectin to yield an
absorbance (A405) value of 1.5 within 2 h. It also reacted strongly with many ABH and Leb substances
from human ovarian cyst fluid (cyst JS phenol insoluble and cyst Mcdon in Fig. 1b; cyst Tighe phenol
insoluble and cyst 14 phenol insoluble in Fig. 1c; cyst Beach phenol insoluble in Fig. 1d). In this assay,
all blood group ABH inactive mucins or polysaccharides, T/Tn containing gps and N-linked plasma
glycoproteins were either poorly active or inactive (Fig. 1, Table 1). These included mild acid-treated
(pH 1.5, 100jC for 2 hr) or Smith degraded human blood group active I/II containing gps (cyst Beach P-
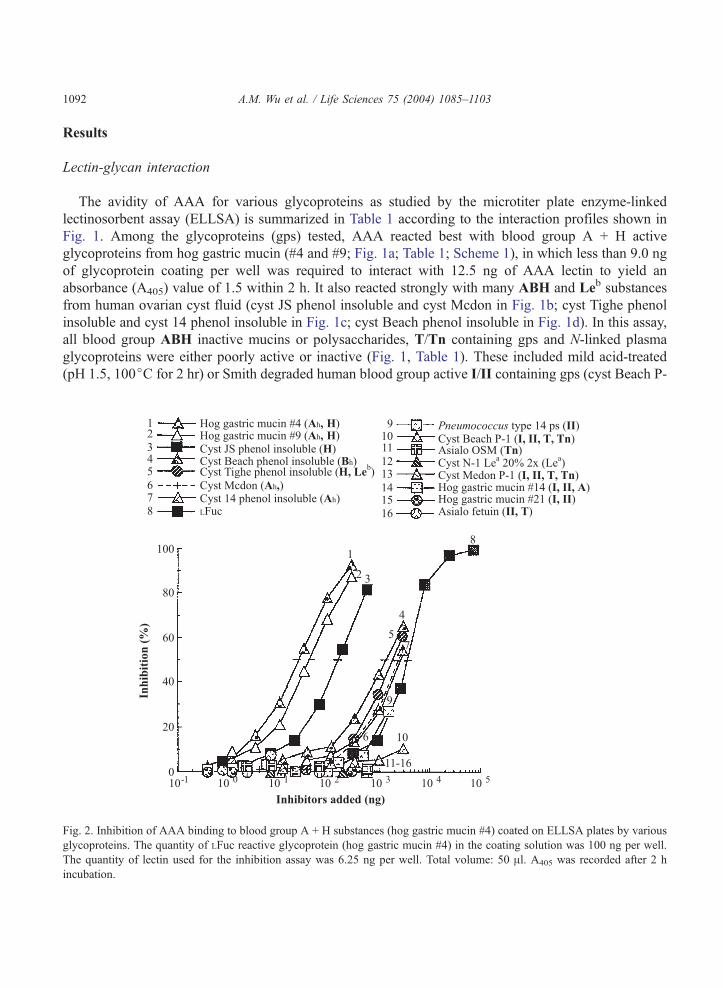
Fig. 2. Inhibition of AAA binding to blood group A + H substances (hog gastric mucin #4) coated on ELLSA plates by various
glycoproteins. The quantity of LFuc reactive glycoprotein (hog gastric mucin #4) in the coating solution was 100 ng per well.
The quantity of lectin used for the inhibition assay was 6.25 ng per well. Total volume: 50 Al. A405 was recorded after 2 h
incubation.

A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1093
1 in Fig. 1d; cyst Mcdon P-1, Tighe P-1 and JS 1st Smith degraded gp in Fig. 1b and 1c), blood group
Lea gp from human ovarian cyst fluid (native and asialo HOC 350 and cyst N-1 Lea 20% 2� in Fig. 1e),
mild-acid hydrolyzed blood group A + H substance (pH 1.5, 100jC for 2–5 hrs; hog gastric mucin #14
and #21 in Fig. 1a), II containing N-glycans (fetuin and its asialo product in Fig. 1f; native and asialo
human a1-acid gp in Table 1; native and asialo RSL; Pneumococcus type 14 polysaccharide in Fig. 1f),
T/Tn containing gps (native and asialo ovine salivary gp in Fig. 1d; bovine salivary gp, human
glycophorin and its asialo products) and colominic acid (Fig. 1f). Since the percentage of glycans
adsorbed on the microtiter plate had not been established in this test, the amount of glycans required to
reach maximum interaction could not be evaluated. Therefore, their binding reactivities were further
assessed by the inhibition of AAA-glycan interaction with various glycans as described below.
Inhibition of AAA - glycan interaction by various glycoproteins
The ability of various glycoproteins to inhibit the binding of AAA by hog gastric mucin #4 (hog A +
H substance) is shown in Fig. 2 and the amount (in nanograms) of glycans required for 50% inhibition of
the lectin-glycan interaction is illustrated in Table 2. Among the glycans tested for inhibition of this
Table 2
Amount of various glycoproteins giving 50% inhibition of AAA binding (6.25 ng/50 Al) by Hog gatric mucin A + H substance
(Hog gastric mucin #4, 100 ng/50 Al)*
Order of
activity
Curve no. Inhibitor Quantity giving 50%
inhibition (ng)yRelative potencyz
1 1 Hog gastric mucin #4 (Ah, H) 20.0 150.0
2 2 Hog gastric mucin #49 (Ah, H) 30.0 100.0
3 3 Cyst JS phenol insoluble (H) 120.0 25.0
4 4 Cyst Beach phenol insoluble (Bh) 1200.0 2.5
5 5 Cyst Tighe phenol insoluble (H, Leb) 1500.0 2.0
6 6 Cyst Mcdon (Ah) 2000.0 1.5
7 7 Cyst 14 phenol insoluble (Ah) 2000.0 1.5
8 8 LFuc 3000.0 1.0
9 9 Pneumococcus type 14 ps (II) >1388.9 (27.0% inhibition) –
10 10 Cyst Beach P-1 (I, II, T, Tn) >2777.8 (10.3% inhibition) –
11 11 Asialo OSM (Tn) >555.6 (1.0% inhibition) –
12 OSM (Sialyl Tn) >277.8 (0% inhibition) –
13 12 Cyst N-1 Lea 20% 2� (Lea) >555.6 (0% inhibition) –
14 13 Cyst Mcdon P–1 (I, II, T, Tn) >555.6 (0% inhibition) –
15 Cyst Tighe P-1 (I/II) >555.6 (0% inhibition) –
16 14 Hog gastric mucin #14 (I, II, A) >555.6 (0% inhibition) –
17 15 Hog gastric mucin #21 (I, II) >555.6 (0% inhibition) –
18 16 Asialo fetuin (II, T) >555.6 (0% inhibition) –
19 Colominic acid >277.8 (0% inhibition) –
*The inhibitory activity was estimated from the inhibition curve in Fig. 2 and is expressed as the amount of inhibitor
(nanogram) giving 50% inhibition. Total volume: 50 Al.y Other gps in which 277.8 ng were used for inhibition of AAA-glycoprotein binding but did not reach 50% inhibition: BN,
asialo BN; RSL, asialo RSL; Human a1-acid gp, asialo Human a1-acid gp; Human glycophorin and asialo human glycophorin.z Reciprocal of relative potency of sugars compared to LFuc which is taken as 1.0.
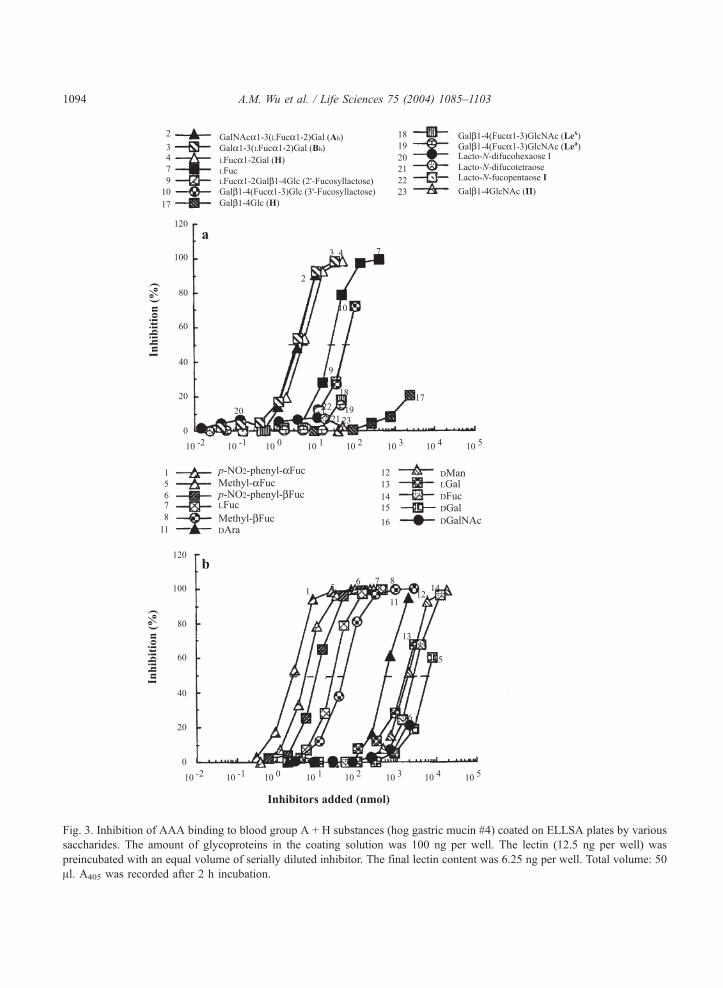
Fig. 3. Inhibition of AAA binding to blood group A + H substances (hog gastric mucin #4) coated on ELLSA plates by various
saccharides. The amount of glycoproteins in the coating solution was 100 ng per well. The lectin (12.5 ng per well) was
preincubated with an equal volume of serially diluted inhibitor. The final lectin content was 6.25 ng per well. Total volume: 50
Al. A405 was recorded after 2 h incubation.
A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031094
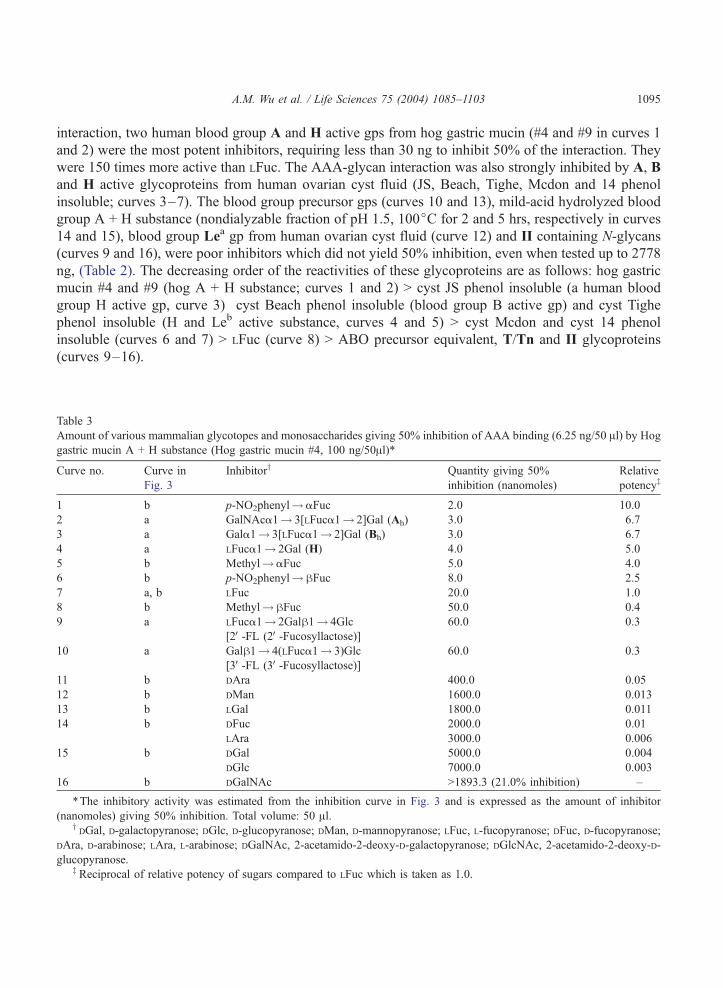
A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1095
interaction, two human blood group A and H active gps from hog gastric mucin (#4 and #9 in curves 1
and 2) were the most potent inhibitors, requiring less than 30 ng to inhibit 50% of the interaction. They
were 150 times more active than LFuc. The AAA-glycan interaction was also strongly inhibited by A, B
and H active glycoproteins from human ovarian cyst fluid (JS, Beach, Tighe, Mcdon and 14 phenol
insoluble; curves 3–7). The blood group precursor gps (curves 10 and 13), mild-acid hydrolyzed blood
group A + H substance (nondialyzable fraction of pH 1.5, 100jC for 2 and 5 hrs, respectively in curves
14 and 15), blood group Lea gp from human ovarian cyst fluid (curve 12) and II containing N-glycans
(curves 9 and 16), were poor inhibitors which did not yield 50% inhibition, even when tested up to 2778
ng, (Table 2). The decreasing order of the reactivities of these glycoproteins are as follows: hog gastric
mucin #4 and #9 (hog A + H substance; curves 1 and 2) > cyst JS phenol insoluble (a human blood
group H active gp, curve 3) cyst Beach phenol insoluble (blood group B active gp) and cyst Tighe
phenol insoluble (H and Leb active substance, curves 4 and 5) > cyst Mcdon and cyst 14 phenol
insoluble (curves 6 and 7) > LFuc (curve 8) > ABO precursor equivalent, T/Tn and II glycoproteins
(curves 9–16).
Table 3
Amount of various mammalian glycotopes and monosaccharides giving 50% inhibition of AAA binding (6.25 ng/50 Al) by Hoggastric mucin A + H substance (Hog gastric mucin #4, 100 ng/50Al)*
Curve no. Curve in
Fig. 3
Inhibitory Quantity giving 50%
inhibition (nanomoles)
Relative
potencyz
1 b p-NO2phenyl!aFuc 2.0 10.0
2 a GalNAca1! 3[LFuca1! 2]Gal (Ah) 3.0 6.7
3 a Gala1! 3[LFuca1! 2]Gal (Bh) 3.0 6.7
4 a LFuca1! 2Gal (H) 4.0 5.0
5 b Methyl!aFuc 5.0 4.0
6 b p-NO2phenyl!hFuc 8.0 2.5
7 a, b LFuc 20.0 1.0
8 b Methyl!hFuc 50.0 0.4
9 a LFuca1! 2Galh1! 4Glc
[2V-FL (2V-Fucosyllactose)]60.0 0.3
10 a Galh1! 4(LFuca1! 3)Glc
[3V-FL (3V-Fucosyllactose)]60.0 0.3
11 b DAra 400.0 0.05
12 b DMan 1600.0 0.013
13 b LGal 1800.0 0.011
14 b DFuc 2000.0 0.01
LAra 3000.0 0.006
15 b DGal 5000.0 0.004
DGlc 7000.0 0.003
16 b DGalNAc >1893.3 (21.0% inhibition) –
*The inhibitory activity was estimated from the inhibition curve in Fig. 3 and is expressed as the amount of inhibitor
(nanomoles) giving 50% inhibition. Total volume: 50 Al.yDGal, D-galactopyranose; DGlc, D-glucopyranose; DMan, D-mannopyranose; LFuc, L-fucopyranose; DFuc, D-fucopyranose;
DAra, D-arabinose; LAra, L-arabinose; DGalNAc, 2-acetamido-2-deoxy-D-galactopyranose; DGlcNAc, 2-acetamido-2-deoxy-D-
glucopyranose.z Reciprocal of relative potency of sugars compared to LFuc which is taken as 1.0.
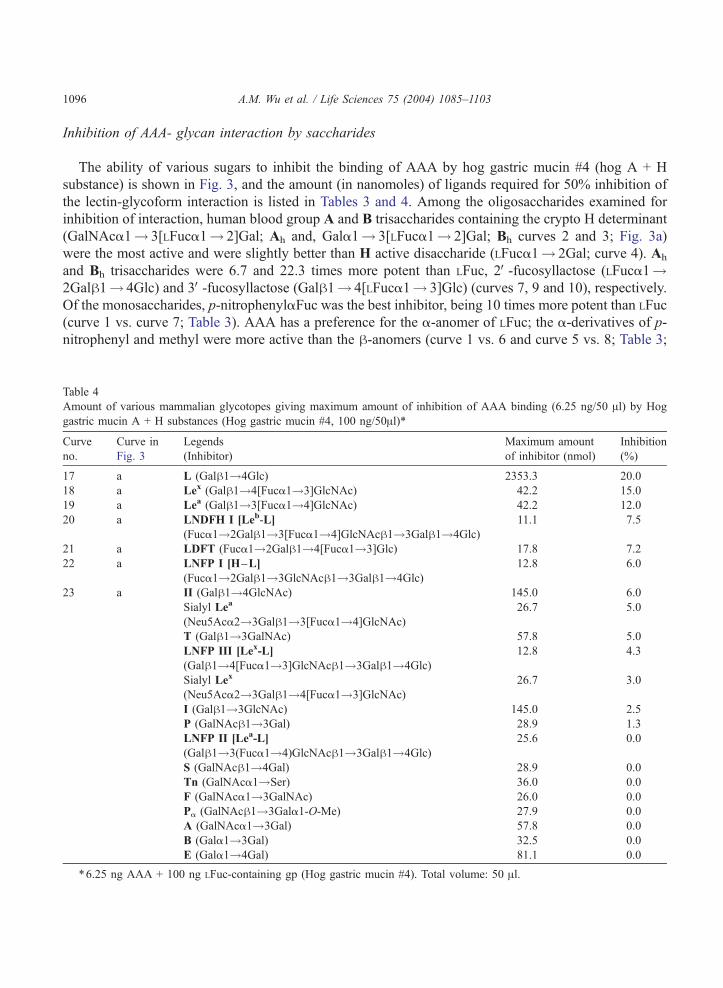
Inhibition of AAA- glycan interaction by saccharides
The ability of various sugars to inhibit the binding of AAA by hog gastric mucin #4 (hog A + H
substance) is shown in Fig. 3, and the amount (in nanomoles) of ligands required for 50% inhibition of
the lectin-glycoform interaction is listed in Tables 3 and 4. Among the oligosaccharides examined for
inhibition of interaction, human blood group A and B trisaccharides containing the crypto H determinant
(GalNAca1! 3[LFuca1! 2]Gal; Ah and, Gala1! 3[LFuca1! 2]Gal; Bh curves 2 and 3; Fig. 3a)
were the most active and were slightly better than H active disaccharide (LFuca1! 2Gal; curve 4). Ah
and Bh trisaccharides were 6.7 and 22.3 times more potent than LFuc, 2V-fucosyllactose (LFuca1!2Galh1! 4Glc) and 3V-fucosyllactose (Galh1! 4[LFuca1! 3]Glc) (curves 7, 9 and 10), respectively.
Of the monosaccharides, p-nitrophenylaFuc was the best inhibitor, being 10 times more potent than LFuc
(curve 1 vs. curve 7; Table 3). AAA has a preference for the a-anomer of LFuc; the a-derivatives of p-
nitrophenyl and methyl were more active than the h-anomers (curve 1 vs. 6 and curve 5 vs. 8; Table 3;
A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031096
Table 4
Amount of various mammalian glycotopes giving maximum amount of inhibition of AAA binding (6.25 ng/50 Al) by Hog
gastric mucin A + H substances (Hog gastric mucin #4, 100 ng/50Al)*
Curve
no.
Curve in
Fig. 3
Legends
(Inhibitor)
Maximum amount
of inhibitor (nmol)
Inhibition
(%)
17 a L (Galh1!4Glc) 2353.3 20.0
18 a Lex (Galh1!4[Fuca1!3]GlcNAc) 42.2 15.0
19 a Lea (Galh1!3[Fuca1!4]GlcNAc) 42.2 12.0
20 a LNDFH I [Leb-L]
(Fuca1!2Galh1!3[Fuca1!4]GlcNAch1!3Galh1!4Glc)
11.1 7.5
21 a LDFT (Fuca1!2Galh1!4[Fuca1!3]Glc) 17.8 7.2
22 a LNFP I [H–L]
(Fuca1!2Galh1!3GlcNAch1!3Galh1!4Glc)
12.8 6.0
23 a II (Galh1!4GlcNAc) 145.0 6.0
Sialyl Lea
(Neu5Aca2!3Galh1!3[Fuca1!4]GlcNAc)
26.7 5.0
T (Galh1!3GalNAc) 57.8 5.0
LNFP III [Lex-L]
(Galh1!4[Fuca1!3]GlcNAch1!3Galh1!4Glc)
12.8 4.3
Sialyl Lex
(Neu5Aca2!3Galh1!4[Fuca1!3]GlcNAc)
26.7 3.0
I (Galh1!3GlcNAc) 145.0 2.5
P (GalNAch1!3Gal) 28.9 1.3
LNFP II [Lea-L]
(Galh1!3(Fuca1!4)GlcNAch1!3Galh1!4Glc)
25.6 0.0
S (GalNAch1!4Gal) 28.9 0.0
Tn (GalNAca1!Ser) 36.0 0.0
F (GalNAca1!3GalNAc) 26.0 0.0
Pa (GalNAch1!3Gala1-O-Me) 27.9 0.0
A (GalNAca1!3Gal) 57.8 0.0
B (Gala1!3Gal) 32.5 0.0
E (Gala1!4Gal) 81.1 0.0
*6.25 ng AAA + 100 ng LFuc-containing gp (Hog gastric mucin #4). Total volume: 50 Al.

A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1097
Fig. 3b), indicating that the a-anomer is important for binding. p-nitrophenyl a- and h-Fuc were more
active than their corresponding methyl a- and h-Fuc derivative, suggesting that hydrophobicity
surrounding fucose is important for binding. LFuc was 20 and 100 times more active than DAra, LGal
and DFuc, respectively, and 250 times more active than DGal, while DGalNAc showed weak inhibition.
DAra (curve 11), which has the same configuration as LFuc (curve 7), but with an absence of carbon-6,
was 20 times less potent than LFuc, indicating that the CH3 at carbon-6 in LFuc is an important functional
group required for binding. LGal (curve 13; Table 3), which has the same structure as LFuc (curve 7)
except that it contains an oxygen (�OH) in carbon-6, was 100 times less active than LFuc, suggesting
that the extra oxygen (�OH) in carbon-6 interferes with the interaction and that the combining site of this
lectin toward LFuc related ligands forms a small cavity.
Among the mammalian glycotopes, lactose (Galh1! 4Glc; Table 4) was tested at a concentration
exceeding that required for 50% inhibition by 2V-fucosyllactose (2V-FL) and 3V-fucosyllactose (3V-FL) by39-fold but yielded only aweak inhibition (20%), suggesting that the fucosyl residue in 2V-FL and 3V-FL is
required for the binding activity (Fig. 3a; curve 17 vs. curves 9 and 10). Furthermore, LFuca1! or
LFuca1! 2Gal (H) was 16 times more active than 2V-FL and 3V-FL, indicating that the major combining
size of AAA is as large as LFuca1! (curve 4 vs. curves 9 and 10; Table 3). Other human milk
oligosaccharides such as, LFuca1! 2 Galh1! 3[Fuca1! 4]GlcNAch1! 3Galh1! 4Glc (LNDFH I,
Leb hexa-saccharide; curve 20), LFuca1! 2Galh1! 4[Fuca 1! 3]Glc (LDFT; gluco-analogue of the
Ley hapten; curve 21), LFuca1! 2Galh1! 3GlcNAch1! 3Galh1! 4Glc (LNFP I, blood group O(H)
type I determinant; curve 22), Galh1! 4[Fuca1! 3]GlcNAch1! 3Galh1! 4Glc (LNFP III; penta-
saccharide) and Galh1! 3[Fuca1! 4]GlcNAch1! 3Galh1! 4Glc (LNFP II) failed to yield 50%
inhibition, indicating that the length of the carbohydrate sequence in an LFuc-containing ligand can reduce
reactivity. The combining site of lectin is a small cavity-type. All the mammalian disaccharide structural
units tested were inactive [Galh1-4GlcNAc (II; curve 23), Galh1-3GlcNAc (I), Gala1-3Gal (B), Gala1-4Gal (E), GalNAca1-3GalNAc (F), GalNAch1-4Gal (S), GalNAca1-3Gal (A), GalNAca1-Ser/Thr (Tn),Galh1-3GalNAc (T), GalNAch1-3Gal (P); Table 4].
Discussion
Anguilla anguilla agglutinin together with fucose-binding lectins such as asparagus pea (Lotus
tetragonolobus) and gorse seed (Ulex europaeus) have been used as powerful tools for studying
glycobiology and hematology (Bianchet et al., 2002; Danguy et al., 1988; Judd, 1980; Liener et al.,
1986). In order to broaden the essential knowledge required for application and interpretation, their
recognition should be thoroughly characterized.
In previous studies, knowledge of the carbohydrate specificity of AAA was limited to a few
oligosaccharides tested by the inhibition assay (Table 5; Baldus et al., 1996; Goldstein and Poretz,
1986; Matsumoto and Osawa, 1971; Morgan and Watkins, 2000; Springer and Desai, 1971; Watkins and
Morgan, 1952) and by binding with crystal lectin (Bianchet et al., 2002). In the present study, we
focused on the interactions of AAAwith mammalian glycotopes by our established ELLSA method (Fig.
1 and Table 1) (Duk et al., 1994; Song et al., 1999; Wu et al., 1999, 2000). The great advantage of this
method is that it is rapid, sensitive and reagent-saving (Wu et al., 1992, 1999). In order to get reliable
data, the interactions of AAA with various glycoforms were examined through three parameters: (1)
amount of the gps added to wells that gave 1.5 units at A405 (Table 1); (2) the maximum A405 value for
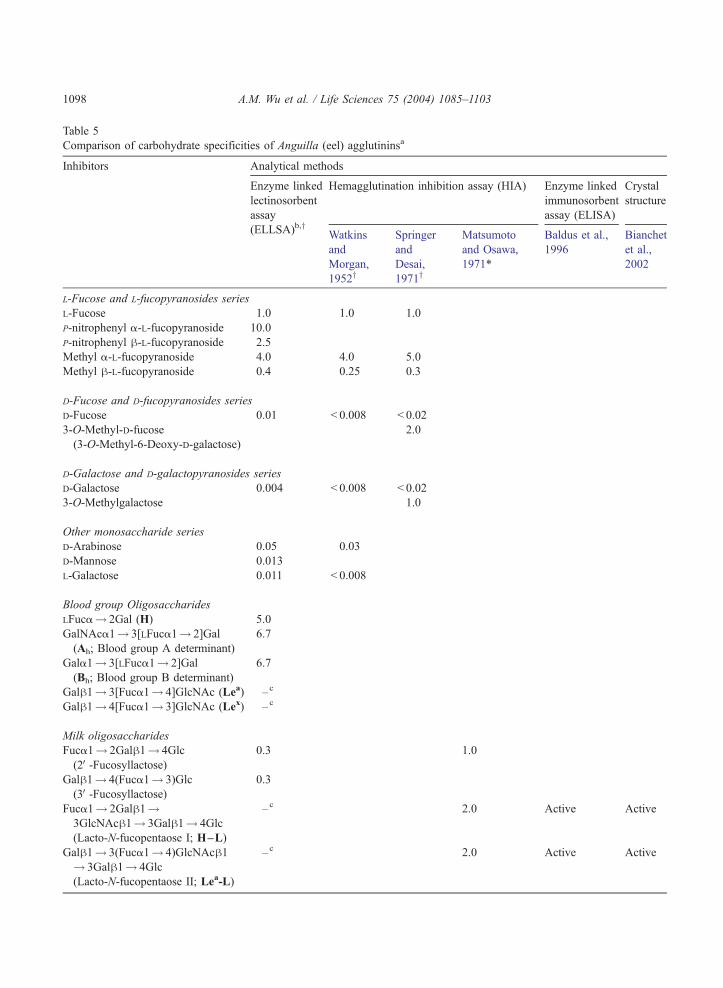
Table 5
Comparison of carbohydrate specificities of Anguilla (eel) agglutininsa
Inhibitors Analytical methods
Enzyme linked
lectinosorbent
assay
Hemagglutination inhibition assay (HIA) Enzyme linked
immunosorbent
assay (ELISA)
Crystal
structure
(ELLSA)b,y Watkins
and
Morgan,
1952y
Springer
and
Desai,
1971y
Matsumoto
and Osawa,
1971*
Baldus et al.,
1996
Bianchet
et al.,
2002
L-Fucose and L-fucopyranosides series
L-Fucose 1.0 1.0 1.0
P-nitrophenyl a-L-fucopyranoside 10.0
P-nitrophenyl h-L-fucopyranoside 2.5
Methyl a-L-fucopyranoside 4.0 4.0 5.0
Methyl h-L-fucopyranoside 0.4 0.25 0.3
D-Fucose and D-fucopyranosides series
D-Fucose 0.01 < 0.008 < 0.02
3-O-Methyl-D-fucose
(3-O-Methyl-6-Deoxy-D-galactose)
2.0
D-Galactose and D-galactopyranosides series
D-Galactose 0.004 < 0.008 < 0.02
3-O-Methylgalactose 1.0
Other monosaccharide series
D-Arabinose 0.05 0.03
D-Mannose 0.013
L-Galactose 0.011 < 0.008
Blood group Oligosaccharides
LFuca! 2Gal (H) 5.0
GalNAca1! 3[LFuca1! 2]Gal
(Ah; Blood group A determinant)
6.7
Gala1! 3[LFuca1! 2]Gal
(Bh; Blood group B determinant)
6.7
Galh1! 3[Fuca1! 4]GlcNAc (Lea) –c
Galh1! 4[Fuca1! 3]GlcNAc (Lex) –c
Milk oligosaccharides
Fuca1! 2Galh1! 4Glc
(2V-Fucosyllactose)0.3 1.0
Galh1! 4(Fuca1! 3)Glc
(3V-Fucosyllactose)0.3
Fuca1! 2Galh1!3GlcNAch1! 3Galh1! 4Glc
(Lacto-N-fucopentaose I; H–L)
–c 2.0 Active Active
Galh1! 3(Fuca1! 4)GlcNAch1! 3Galh1! 4Glc
(Lacto-N-fucopentaose II; Lea-L)
–c 2.0 Active Active
A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031098

Table 5 (continued)
Inhibitors Analytical methods
Enzyme linked
lectinosorbent
assay
Hemagglutination inhibition assay (HIA) Enzyme linked
immunosorbent
assay (ELISA)
Crystal
structure
(ELLSA)b,y Watkins
and
Morgan,
1952y
Springer
and
Desai,
1971y
Matsumoto
and Osawa,
1971*
Baldus et al.,
1996
Bianchet
et al.,
2002
Galh1! 4(Fuca1!3)GlcNAch1! 3Galh1! 4Glc
(Lacto-N-fucopentaose III; Lex-L)
– c Inactive Inactive
Fuca1! 2Galh1! 4[Fuca1!3]Glc (Lacto-difucotetraose)
–c Active
Fuca1! 2Galh1! 3[Fuca1!4]GlcNAch1! 3Galh1! 4Glc
(Lacto-N-difucohexaose I)
– c Inactive
Blanks indicate where testing was not done.
The relative potency of different mammalian glycotopes and monosaccharides were calculated from Tables 3 and 4 for AAA
(Anguilla anguilla agglutinin, fresh water eel). Their relative potencies obtained by hemagglutination inhibition were calculated
from the minimum concentration (mg/ml) giving complete inhibition of hemagglutinating doses.a Data from Bianchet et al., 2002 (Anguilla anguilla, European eel; Sigma); Baldus et al., 1996 (Anguilla anguilla
agglutinin; E–Y Laboratories, San Mateo, California, USA); Morgan and Watkins, 2000; Watkins and Morgan, 1952 (Anguilla
anguilla); Matsumoto and Osawa, 1971 (eel ofunspecified source); Goldstein and Poretz, 1986; Springer and Desai, 1971
(Anguilla rostrata, fresh or brackish water eel).b See Tables 3 and 4.y Reciprocal of inhibitory potency of LFuc was taken as 1.0.
*Reciprocal of inhibitory potency of Fuca1! 2Galh1! 4Glc was taken as 1.0.c 2 to 10 times more amount was tested than that of H epitope (LFuca1! 2Gal) require for 50% inhibition.
A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1099
each gp after 2 h incubation (Fig. 1 and Table 1), and (3) the amount of gps required to give 50%
inhibition of AAA- binding (Fig. 2 and Table 2).
Compared to previous AAA binding property results (Table 5), we have performed a more
systematic analysis using a series of mammalian glycotopes and polyvalent structural units in
glycoproteins and have established the decreasing order of ligand reactivities as follows: p-nitro-
phenyl-a-LFuc z Ah and Bh trisaccharides z H disaccharide > methyl-a-LFuc, which were 5 to 10
times more active than LFuc, while Lea, I, II, T, Tn, P, S, A, B, E and F oligosaccharides were inactive
(Tables 3–5).
As shown in Table 2 (curves 3–7) and Table 3 (curves 2–4), AAA demonstrated parallel reactivity
with the ABH blood group system, demonstrating that this lectin recognizes polyvalent Ah, Bh and H
glycotopes. Although AAA has a greater affinity for polyvalent ligands in glycoproteins than
monovalent haptens (Table 2 vs. Table 3) (e.g. enhancement of the poor oligosaccharides Leb reactivity
due to polyvalency effects, Table 4, curve 20 vs. Table 2, curve 5), the scale of enhancement is small
compared to that found in other lectins, such as Agaricus bisporus agglutinin (ABA; Wu et al., 2003a)
and Artocarpus integrifolia (Jacalin; Wu et al., 2003b). This indicates that the power of polyvalency
effects on carbohydrate-protein interactions have to be evaluated case by case.

A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031100
The location of LFuc linkage is an important binding factor. AAA recognized only the human blood
group A, B and Leb active glycoproteins (a form of H and crypto H substances that are composed of
LFuca1-2Gal) from human ovarian cyst fluid (cyst JS, Mcdon, Tighe and Beach phenol insoluble, Fig.
1b and 1c) and hog gastric mucin #4 and #9 (a blood group A + H substance, Fig. 1a), but not with Lea
substances, in which the LFuc is linked through the penultimate LFuca1-4Gal rather than the terminal
LFuca1-2Gal (cyst N-1 Lea 20% 2� , native and asialo HOC 350, Fig. 1e), suggesting that a1-2 linked
LFuc to Gal (LFuca1-2Gal) at the non-reducing end of glycoproteins is essential for lectin-carbohydrate
interaction. As all the non-LFuc containing gps (II and T/Tn mammalian structures and N-glycans) are
inactive with AAA, it implies that the binding of AAA to blood group gps should be mainly attributed to
its interactions with Ah, Bh and H glycotopes instead of the adjacent structures.
It has been reported that the Lea ligand is an active AAA ligand (Baldus et al., 1996; Bianchet et al.,
2002; Matsumoto and Osawa, 1971; Table 5). However, our study indicates that it is actually a weak
inhibitor. This may be due to the different eel species sources from which the lectin was isolated and/or
different batches of preparation (Bianchet et al., 2002; Matsumoto and Osawa, 1971) or due to the
amount of carbohydrate ligand used (Baldus et al., 1996). Although our study with Lewis oligosac-
charides and their derivatives showed differences in reactivity from Baldus’ report (Table 4), the binding
differences of these monovalent glycotopes are negligible when compared with the activity of polyvalent
forms (Table 4, curve 20 vs. Table 2, curve 5). Furthermore, the binding of glycoproteins to AAA in this
investigation were similar to natural features and should be closer to the in vivo interactions than with
lectin-oligosaccharide interaction. Many reports revealed that lectins with the same mono- or oligosac-
charide specificity may demonstrate completely different specificities for macromolecules, and have
even shown a shift of binding specificity from one type of carbohydrate ligand to another when the
surface density of the carbohydrate increases (Horan et al., 1999; Lee, 1992; Lee and Lee, 2000; Wu,
1988, 2001; Wu et al., 2003a). Therefore, to provide a more satisfactory and realistic depiction of the
carbohydrate specificity of a lectin, the following five criteria are suggested to be considered (Wu,
2001): (i) monosaccharide specificity (Fuc, Gal, GalNAc, GlcNAc, and/or Man); (ii) expression of
reactivities towards mammalian structural units in decreasing order (disaccharide and Tn specificity);
(iii) the most active ligand; (iv) simple multi-antennary or cluster effect, using either the natural
glycopeptides or synthetic neoglycoconjugates with branching sugar side chains; and (v) complex
multivalent or cluster effects, displayed by macromolecules with characterized reactive glycotopes. In
our current study, we have provided almost all the information required for this purpose.
ABH and Lewis histo-blood group antigens are among the most abundant epitopes in the human body
(Morgan and Watkins, 2000). Aberrant expression of these antigens are associated with several
pathological conditions and malignancies (Greenwell, 1997). Therefore, the high selectivity of AAA
for ABH/Leb blood groups without crossreactivity to their precursors and other glycoproteins may make
this lectin an excellent reagent for the characterization and detection of alterations to cell surface
glycoconjugates during malignant transformation.
The overall findings of this study are as follows: (a) illustration of the high specificity of AAA for H
active and crypto H active gps, such as blood group A, B, H and Leb determinants (Figs. 1 and 2), a
property which is rare among animal lectins; (b) the establishment of the binding relationship between
AAA and mammalian O-glycan structural units (Fig. 3); (c) the fact that AAA interacts well with blood
group ABH and Leb determinants but not with Lea substances infers that the optimal density of a1-2
linked LFuc to Gal at the non-reducing end of glycoproteins is essential for lectin-carbohydrate
interactions; (d) the demonstration that AAA has a preference for the a-anomer of LFuc rather than

A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1101
for the h-anomer; hydrophobicity surrounding fucose enhances the binding (Fig. 3); (e) the configu-
ration at carbon-6 of LFuc as well as absence of oxygen at carbon-6, are important factors for successful
binding; (f) unlike many Galh1! specific lectins, such as Agaricus bisporus agglutinin (Wu et al.,
2003a) and Artocarpus integrifolia (Wu et al., 2003b), the effect of glycotope polyvalency are relatively
limited in AAA; and (g) the hierarchy of potency of mono- and oligosaccharides can be ranked as: p-
nitrophenyl-aFuc > Ah, Bh and H > LFuc > LFuca1! 2Galh1! 4Glc (2V-FL) and Galh1! 4[L-
Fuca1! 3]Glc (3V-FL) (the human milk oligosaccharides); while in oligosaccharide forms, LNDFH
I (Leb hexa-), Lea, Lex (Galh1! 4[LFuca1! 3]GlcNAc), and LDFT (gluco-analogue of Ley) were
weak or inactive. From the present results, it can be deduced that the combining site of AAA towards
LFuc containing glycans should be of a small cavity-type capable recognizing mainly H/crypto H
binding well to ABH and Leb glycoproteins and that hydrophobic interaction is important for binding of
the a anomeric LFuc. The reaction profile of AAA supports the concept that every lectin with unique
amino acid sequence has its own binding characteristics (Wu, 2001, 2002, 2003; Wu and Sugii, 1988).
Although most mammalian glycotopes have been tested in this study, it would also be informative to
examine glycotopes from bacterial liposaccharides. Unfortunately, this is hampered due to the
unavailability of reagents.
Acknowledgements
This work was supported by Grants from the Chang-Gung Medical Research Project (CMRP No.
1028), Kwei-san, Tao-yuan, Taiwan, and the National Science Council (NSC 92-2311-B-182-005, 92-
2320-B-182-045, and 92-2320-B-182-046), Taipei, Taiwan.
References
Allen, P.Z., Kabat, E.A., 1959. Immunochemical studies on blood groups. 22. Immunochemical studies on the non-
dialyzable residue from partially hydrolyzed blood group A, B, O(H) substances (P1 fraction). Journal of Immunology
82, 340–357.
Baldus, S.E., Thiele, J., Park, Y.-O., Hanisch, F.-G., Bara, J., Fischer, R., 1996. Characterization of the binding specificity of
Anguilla anguilla agglutinin (AAA) in comparison to Ulex europaeus agglutinin I (UEA-I). Glycoconjugate Journal 13,
585–590.
Bianchet, M.A., Odom, E.W., Vasta, G.R., Amzel, L.M., 2002. A novel fucose recognition fold involved in innate immunity.
Nature Structural Biology 9, 628–634.
Cory, H.T., Yates, A.D., Donald, A.S., Watkins, W.M., Morgan, W.T.J., 1974. The nature of the human blood group P1
determinant. Biochemical Biophysical Research Communication 61, 1289–1296.
Danguy, A., Kiss, R., Pasteels, J.L., 1988. Lectins in histochemistry. A survey. Biological Structures and Morphogenesis 1,
93–106.
Duk, M., Lisowska, E., Wu, J.H., Wu, A.M., 1994. The biotin/avidin-mediated microtiter plate lectin assay with the use of
chemically modified glycoprotein ligand. Analytical Biochemistry 221, 266–272.
Fournet, B., Montreuil, J., Strecker, G., Dorland, L., Haverkamp, J., Vliegenthart, J.F.G., Binette, J.P., Schmid, K., 1978.
Determination of the primary structures of 16 asialo-carbohydrate units derived from human plasma alpha 1-acid glyco-
protein by 360-MHz 1H-NMR spectroscopy and permethylation analysis. Biochemistry 17, 5206–5214.
Gabius, H.-J., Andre, S., Kaltner, H., Siebert, H.-C., 2002. The sugar code: functional lectinomics. Biochimica et Biophysica
Acta 1572, 165–177.
Goldstein, I.J., Poretz, R.D., 1986. Isolation, physicochemical characterization, and carbohydrate binding specificity of lectins.

A.M. Wu et al. / Life Sciences 75 (2004) 1085–11031102
In: Liener, I.E., Sharon, N., Goldstein, I.J. (Eds.), The lectins, properties, function and applications in biology and medicine,
33–247.
Greenwell, P., 1997. Blood group antigens: molecules seeking a function? Glycoconjugate Journal 14, 159–173.
Herp, A., Borelli, C., Wu, A.M., 1988. Biochemistry and lectin binding properties of mammalian salivary mucous glycopro-
teins. Advances in Experimental Medicine and Biology 228, 395–435.
Horan, N., Yan, L., Isobe, H., Whitesides, G.M., Kahne, D., 1999. Nonstatistical binding of a protein to clustered carbohy-
drates. Proceedings of the National Academy of Sciences of the United States of America 96, 11782–11786.
Judd, W.J., 1980. The role of lectins in blood group serology. Critical Review in Clinical Lab Science 12, 171–214.
Kabat, E.A., Baer, H., Bezer, A.E., Knaub, V., 1948. Immunochemical studies on blood groups. 7. Chemical changes associated
with destruction of blood group activity and enhancement of the type XIV cross reactivity by partial hydrolysis of hog and
human blood group A, B and O substances. Journal of Experimental Medicine 88, 43–57.
Kabat, E.A., 1956. Blood group substances: their chemistry and immunochemistry. Academic Press, New York, pp. 135–139.
Lee, Y.C., 1992. Biochemistry of carbohydrate-protein interaction. FASEB Journal 6, 3193–3200.
Lee, R.T., Lee, Y.C., 2000. Affinity enhancement by multivalent lectin-carbohydrate interaction. Glycoconjugate Journal 17,
543–551.
Leskowitz, S., Kabat, E.A., 1954. Immunochemical studies on blood groups. XV. The effect of mild acid hydrolysis on the
glucosamine and galactosamine in blood group substances. Journal of the American Chemical Society 76, 5060–5065.
Liener, I.E., Sharon, N., Goldstein, I.J., 1986. The Lectins. Academic Press, Orlando.
Lindberg, B., Lonngren, J., Powell, D.A., 1977. Structural studies on the specific type-14 pneumococcal polysaccharide.
Carbohydrate Research 58, 177–186.
Lisowska, E., Messeter, L., Duk, M., Czerwinski, M., Lundblad, A., 1987. A monoclonal ant-glycophorin A antibody
recognizing the blood group M determinant: studies on the subspecificity. Molecular Immunology 24, 605–613.
Lisowska, E., Duk, M., Wu, A.M., 1996. Preparation of biotinylated lectins and application in microtiter plate assays and
western blotting. The BioMethod-series, vol. 7, pp. 115–129.
Lloyd, K.O., Kabat, E.A., 1968. Immunochemical studies on blood groups. 41. Proposed structures for the carbohydrate portion
of A, B, H, Lewisa and Lewisb substances. Proceedings of the National Academy of Sciences United States of America 61,
1470–1477.
Maisonrouge-McAuliffe, F., Kabat, E.A., 1976. Immunochemical studies on blood groups: Heterogeneity of oligosaccharides
liberated by degradation with alkaline borohydride of two human ovarian cyst fractions differing in B, I, and i activities and
in reactivity toward concanavalin A. Archives of Biochemistry and Biophysics 175, 81–89.
Matsumoto, I., Osawa, T., 1971. On the specificity of various heterologous anti-H hemagglutinins. Vox Sanguins 21, 548–557.
Morgan, W.T.J., Watkins, W.M., 1964. Blood group P1 substance. 1. Chemical properties. Proceeding 9th Congress Interna-
tional Society. Blood Transfusion, Mexico, 225–229.
Morgan, W.T., Watkins, W.M., 2000. Unravelling the biochemical basis of blood group ABO and Lewis antigenic specificity.
Glycoconjugate Journal 17, 501–530.
Moschera, J., Pigman, W., 1975. The isolation and characterization of rat sublingual mucus-glycoprotein. Carbohydrate
Research 40, 53–67.
Nilsson, B., Norden, N.E., Svensson, S., 1979. Structural studies on the carbohydrate portion of fetuin. Journal of Biological
Chemistry 254, 4545–4553.
Pusztai, A., Morgan, W.T.J., 1961. Studies in immunochemistry. 18. The isolation and properties of a sialomucopolysaccharide
possessing blood-group Lea specificity and virus-receptor activity. Biochemical Journal 78, 135–146.
Reuter, G., Gabius, H.-J., 1999. Eukaryotic glycosylation: whim of nature or multipurpose tool? Cellular and Molecular Life
Sciences 55, 368–422.
Slomiany, A., Slomiany, B.L., 1978. Structures of the acidic oligosaccharides isolated from rat sublingual glycoprotein. Journal
of Biological Chemistry 253, 7301–7306.
Song, S.-C., Chen, Y.-Y., Herp, A., Tsai, M.-S., Wu, A.M., 1999. Lectinochemical characterization of a GalNAc and multi-
Galh1-4GlcNAc reactive lectin from Wistaria sinensis seeds. European Journal of Biochemistry 266, 1–16.
Springer, G.F., Desai, P.R., 1971. Monosaccharides as specific precipitinogens of eel anti-human blood-group H(O) antibody.
Biochemistry 10 (20), 3749–3761.
Tettamanti, G., Pigman, W., 1968. Purification and characterization of bovine and ovine submaxillary mucins. Archives of
Biochemistry and Biophysics 124, 41–50.
van Halbeek, H., Dorland, L., Vliegenthart, J.F.G., Kochetkov, N.K., Arbatsky, N.P., Derevitskaya, V.A., 1982. Character-

A.M. Wu et al. / Life Sciences 75 (2004) 1085–1103 1103
ization of the primary structure and the microheterogeneity of the carbohydrate chains of porcine blood-group H substance
by 500-MHz 1H-NMR spectroscopy. European Journal of Biochemistry 127, 21–29.
Wagner, M., 1988. Light and electron microscopic lectin histochemistry using fluorochromes and ferritin as labels. Acta
Histochemica Supplementband 36, 115–123.
Watkins, W.M., Morgan, W.T.J., 1952. Neutralisation of the anti-H agglutinin in eel serum by simple sugars. Nature 169,
825–826.
Wieruszeski, J.M., Michalski, J.C., Montreuil, J.G., Strecker, J., Peter-Katalinic, H., Eagge, H., Van Halbeek, J.H., Mutsaers,
G.M., Vliegenthart, J.F.G., 1987. Structure of the monosialyl oligosaccharides derived from salivary gland mucin glyco-
proteins of the Chinese swiftlet (genus Collocalia). Journal of Biological Chemistry 262, 6650–6657.
Wu, A.M., 1988. Structural concepts of the blood group A, B, H, Lea, Leb, I and i active glycoproteins purified from human
ovarian cyst fluid. Advances in Experimental Medicine and Biology 228, 351–392.
Wu, A.M., 2001. Expression of binding properties of Gal/GalNAc reactive lectins by mammalian glycotopes. Advances in
Experimental Medicine and Biology 491, 55–64.
Wu, A.M., 2002. Carbohydrate structural units in glycosphingolipids as receptors for Gal and GalNAc reactive lectins.
Neurochemical Research 27, 593–600.
Wu, A.M., 2003. Carbohydrate structural units glycoproteins and polysaccharides as important ligands for Gal and GalNAc
reactive lectins. Journal of Biomedical Science 10, 676–688.
Wu, A.M., Pigman, W., 1977. Preparation and characterization of armadillo submandibular glycoproteins. Biochemical Journal
161, 37–47.
Wu, A.M., Sugii, S., 1988. Differential binding properties of GalNAc and/or Gal specific lectins. Advances in Experimental
Medicine and Biology 228, 205–263.
Wu, A.M., Kabat, E.A., Pereira, M.E.A., Gruezo, F.G., Liao, J., 1982. Immunochemical studies on blood groups: the internal
structure and immunological properties of water-soluble human blood group A substance studied by Smith degradation,
liberation and fractionation of oligosaccharides and reaction with lectins. Archives of Biochemistry and Biophysics 215,
390–404.
Wu, A.M., Kabat, E.A., Nilsson, B., Zopf, D.A., Gruezo, F.G., Liao, J., 1984. Immunochemical studies on blood groups.
Purification and characterization of radioactive 3H-reduced di- to hexasaccharides produced by alkaline beta-elimination-
borohydride 3H-reduced of Smith degraded blood group A active glycoproteins. Journal of Biological Chemistry 259,
7178–7186.
Wu, A.M., Lin, S.-R., Chin, L.-K., Chow, L-P., Lin, J.-Y., 1992. Defining the carbohydrate specificities of Abrus precatorius
agglutinin as T (Galh1! 3GalNAc) > I/II (Galh1! 3/4GlcNAc). Journal of Biological Chemistry 267, 19130–19139.
Wu, A.M., Herp, A., Song, S.C., Wu, J.H., Chang, K.S.S., 1995. Interaction of native and asialo rat sublingual glycoproteins
with lectins. Life Sciences 57, 1841–1852.
Wu, A.M., Song, S.-C., Chang, S.-C., Wu, J.H., Chang, K.S.S., Kabat, E.A., 1997. Further characterization of the binding
properties of a GalNAc specific lectin from Codium fragile subspecies Tomentosoides. Glycobiology 7, 1061–1066.
Wu, A.M., Wu, J.H., Chen, Y.-Y., Song, S.-C., Kabat, E.A., 1999. Further characterization of the combining sites of Bandeiraea
(Griffonia) simplicifolia lectin-I, isolectin A4 (GSI-A4). Glycobiology 9, 1161–1170.
Wu, A.M., Song, S.C., Chen, Y.Y., Gilboa-Garber, N., 2000. Defining the carbohydrate specificities of Aplysia gonad lectin
(AGL) exhibiting a peculiar d-galacturonic acid affinity. Journal of Biological Chemistry 275, 14017–14024.
Wu, A.M., Wu, J.H., Herp, A., Liu, J.H., 2003a. Effect of polyvalencies of glycotopes on the binding of a lectin from the edible
mushroom, Agaricus bisporus. Biochemical Journal 371, 311–320.
Wu, A.M., Wu, J.H., Lin, L.H., Lin, S.H., Liu, J.H., 2003b. Binding profile of Artocarpus integrifolia agglutinin (Jacalin). Life
Sciences 72, 2285–2302.
Copyright © 2022 FDOKUMEN















![Anguilla anguilla L. Biochemical and Genotoxic Responses to Benzo[ a]pyrene](https://static.fdokumen.com/doc/165x107/631d4597f26ecf94330a787a/anguilla-anguilla-l-biochemical-and-genotoxic-responses-to-benzo-apyrene.jpg)




