
Neuropeptide Y Promotes Neurogenesis in Murine Subventricular Zone
Upload
independentCategory
view
3download
0
Developmental Brain Research, 51 (1990) 45-61 45 Elsevier
BRESD 50994
Morphometrical and microdensitometrical studies on peptide- and tyrosine hydroxylase-like immunoreactivities in the forebrain of rats
prenatally exposed to methylazoxymethanol acetate
Michele Zoli 1, Emilio Merlo Pich 1, Mauro Cimino 2, Gabriella Lombardelli 2, Guido Peruzzi 2, Kjell Fuxe 3, Luigi F. Agnati 1 and Flaminio Cattabeni 2
~ Institute of Human Physiology, University of Modena, Modena (Italy), 21nstitute of Pharmacology and Pharmacognosy, University of Urbino, Urbino (Italy) and 3Department of Histology and Neurobiology, Karolinska Institutet, Stockholm (Sweden)
(Accepted 13 June 1989)
Key words: Methylazoxymethanol acetate; Neuropeptide; Immunocytochemistry; Computer-assisted image analysis; Microencephaly; Rat
Methylazoxymethanol acetate (MAM Ac) injected into pregnant rats at a dose of 25 mg/kg at gestational day 15 causes microcephaly due to an atrophy of various telencephalic areas, mainly neocortex, hippocampus and basal ganglia. Previous studies demonstrated alterations in various neurochemical markers of classical transmitter systems in these regions. The present paper deals with changes in peptide and tyrosine hydroxylase (TH)-containing neurons in MAM Ac-induced microcephaly using immunocytochemistry coupled with computer-assisted morphometry and microdensitometry. No change in the number of vasoactive intestinal polypeptide (VIP)-immunoreactive neurons in the neocortex and neuropeptide Y (NPY)-immunoreactive neurons in the nucleus caudatus-putamen was found whereas cholecystokinin (CCK)- and NPY-immunoreactive neurons in the neocortex and CCK- and VIP-immunoreactive neurons in the hippocampus were decreased. The reduction of the latter peptide containing neuronal populations led to a maintained density of cells in MAM Ae-exposed rats, due to the parallel reduction of the overall mass of these regions. TH immunoreactivity was found to be unchanged in the basal ganglia, and increased in the cerebral cortex in agreement with previous reports on noradrenaline cortical system after MAM Ac exposure. The present results show a heterogeneous vulnerability of different peptide immunoreactive neuronal populations to MAM Ac exposure. The sparing of VIP- and NPY-immunoreactive neurons may be due to their late development in the neocortex and striatum, respectively. The hypothesis is introduced that cortical VIP interneurons can develop independent of marked alterations in the intrinsic circuitry of the cortical region.
INTRODUCTION
Methy lazoxymethano l acetate ( M A M Ac) , injected
into pregnant rats during a par t icular per iod of gestat ion,
can cause a dose -dependen t microcephaly together with
a relat ively main ta ined body weight 24'69. The cytotoxic
effect of M A M Ac exposure is due to the antimitot ic
action of the molecule based on the methyla t ion of D N A
and R N A bases 58. Af te r exposure to M A M Ac on
gestat ional day 15, the most affected regions of the
central nervous system (CNS) are the cerebral cortex and
the h ippocampal format ion, while o ther te lencephal ic
regions such as s t r ia tum show a less p ronounced alter-
ation. Histological examinat ions of the cerebral cortex
and h ippocampus from exposed rats grown to adul thood
revealed hypoplas ia and cytoarchi tectonic abnor-
malit ies 18"4°. The rest of the CNS is only slightly al tered at doses up to 25 mg/kg 32.
Previous neurochemical studies carr ied out both at the
cortical and striatal levels showed that prenata l exposure
to M A M Ac leads to increased concentrat ion of nor-
adrenal ine , acetylcholine, dopamine and serotonin con-
tents in the cerebra l cor tex and of monoamines in the
str iatum, whereas the total amount per region was unchanged or slightly reduced 11'33~4~'43"4s. In contrast , no
changes in the concentra t ion of presynapt ic markers of
y-aminobutyr ic acid ( G A B A ) and glutamic acid neurons
in the cerebral cortex 4° and of acetylcholine and G A B A
neurons in the s t r ia tum were found in M A M Ac-exposed
animals, whereas total amounts per region were great ly
reduced 11. Al though there is a large number of reports
describing the effects of M A M Ac exposure on classical
neurot ransmit ters , few data are available on the fate of
neuropept ides 15.
In spite of the fact that M A M Ac-exposed rats show no
gross behavioural deficit, impai rments of various behav-
ioural pa ramete r s such as general hyperact ivi ty 14 and
various cognitive deficits 8"61'66 have been described.
However , these behavioura l abnormal i t ies have not been
satisfactorily corre la ted to any specific neuroanatomica l
or neurochemical al terat ions.
In the present paper the effects of M A M Ac exposure
Correspondence: L.F. Agnati, Institute of Human Physiology, University of Modena, Via Campi 287, 41100 Modena, Italy.
0165-3806/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)
MATERIALS AND METHODS
o n t y r o s i n e h y d r o x y l a s e ( T H ) - , g lucoco r t i co id r e c e p t o r
( G R ) - , v a s o a c t i v e i n t e s t i n a l p o l y p e p t i d e (V1P)- , n e u r o -
p e p t i d e Y ( N P Y ) - a n d c h o l e c y s t o k i n i n ( C C K ) - l i k e im-
m u n o r e a c t i v i t i e s h a v e b e e n i n v e s t i g a t e d in the r a t fore-
b r a i n by m e a n s of i m m u n o c y t o c h e m i s t r y c o u p l e d wi th
c o m p u t e r - a s s i s t e d i m a g e analys is 3"s3.
KK 8920
Subjects and treatment MAM Ac (Sigma, St.Louis) was administered i.p. in a dose of 25
mg/kg to female rats (Sprague-Dawley, Charles River) at gesta- tional day 15. Controls were injected i.p. with an equal volume of vehicle (saline) under the same experimental conditions. Concep- tion was determined by vaginal smear and the day of discovery of a sperm-plug was designated gestational day 1. The litters born on gestational day 22 were culled to 8 pups and weaned for 3 weeks. The male rats were selected and kept under standardized temper- ature, humidity and lighting conditions (lights on at 8.00 h, lights off at 20.00 h). They had free access to food and water. The rats were sacrificed at the age of 3 months.
Immunocytochemical procedure The rats (n = 4/group) were sacrificed in the morning (between
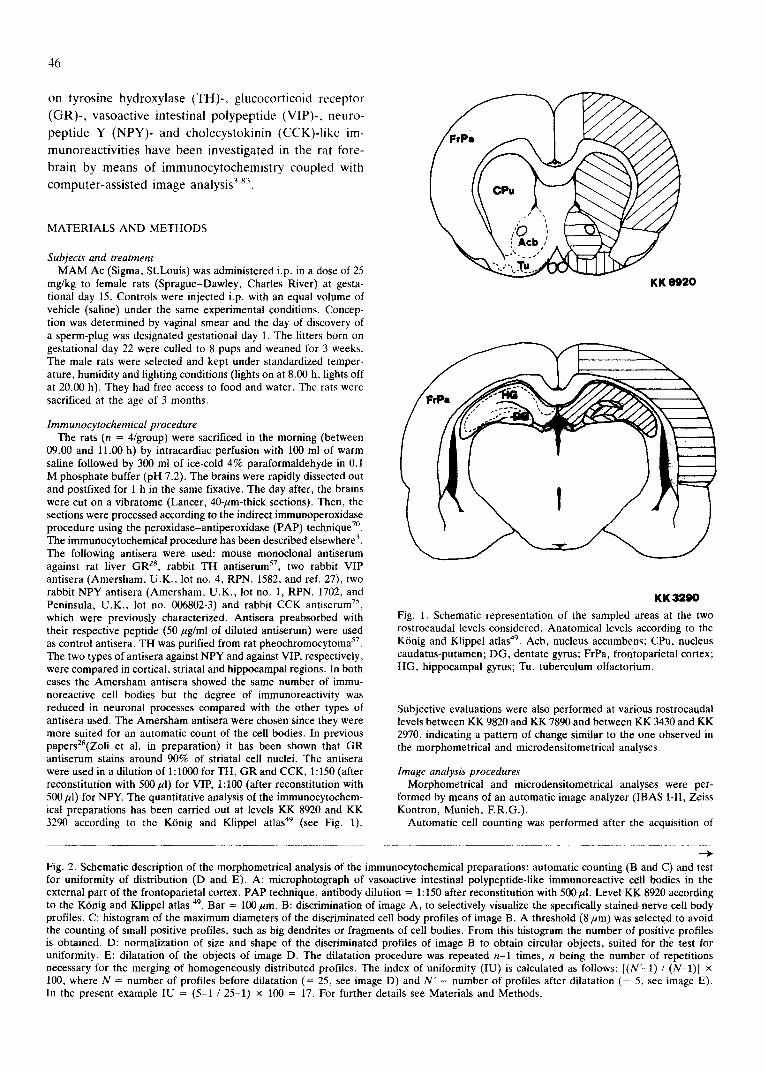
09.00 and 11.00 h) by intracardiac perfusion with 100 ml of warm saline followed by 300 ml of ice-cold 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.2). The brains were rapidly dissected out and postfixed for 1 h in the same fixative. The day after, the brains were cut on a vibratome (Lancer, 40-/~m-thick sections). Then, the sections were processed according to the indirect immunoperoxidase procedure using the peroxidase-antiperoxidase (PAP) technique TM. The immunocytochemical procedure has been described elsewhere 3. The following antisera were used: mouse monoclonal antiserum against rat liver GR 28, rabbit TH antiserum 57, two rabbit VIP antisera (Amersham, U.K., lot no. 4, RPN. 1582, and ref. 27), two rabbit NPY antisera (Amersham, U.K., lot no. 1, RPN. 1702, and Peninsula, U.K., lot no. 006802-3) and rabbit CCK antiserum 75, which were previously characterized. Antisera preabsorbed with their respective peptide (50/~g/ml of diluted antiserum) were used as control antisera. TH was purified from rat pheochromocytoma 5r. The two types of antisera against NPY and against VIP, respectively, were compared in cortical, striatal and hippocampal regions. In both cases the Amersham antisera showed the same number of immu- noreactive cell bodies but the degree of immunoreactivity was reduced in neuronal processes compared with the other types of antisera used. The Amersham antisera were chosen since they were more suited for an automatic count of the cell bodies. In previous papers2S(Zoli et al. in preparation) it has been shown that GR antiserum stains around 90% of striatal cell nuclei. The antisera were used in a dilution of 1:1000 for TH, GR and CCK, 1:150 (after reconstitution with 500/~1) for VIP, 1:100 (after reconstitution with 500/A) for NPY. The quantitative analysis of the immunocytochem- ical preparations has been carried out at levels KK 8920 and KK 3290 according to the K6nig and Klippel atlas 49 (see Fig. 1).
46
K K 3290
Fig. 1. Schematic representation of the sampled areas at the two rostrocaudal levels considered. Anatomical levels according to the KOnig and Klippel atlas 49. Acb, nucleus accumbens; CPu, nucleus caudatus-putamen; DG, dentate gyrus; FrPa, frontoparietal cortex; HG, hippocampal gyrus; Tu, tuberculum olfactorium.
Subjectwe evaluations were also performed at various rostrocaudal levels between KK 9820 and KK 7890 and between KK 3430 and KK 2970, indicating a pattern of change similar to the one observed in the morphometrical and microdensitometrical analyses.
Image analysis procedures Morphometrical and microdensitometrical analyses were per-
formed by means of an automatic image analyzer (IBAS I-II, Zeiss Kontron, Munich, F.R.G.).
Automatic cell counting was performed after the acquisition of
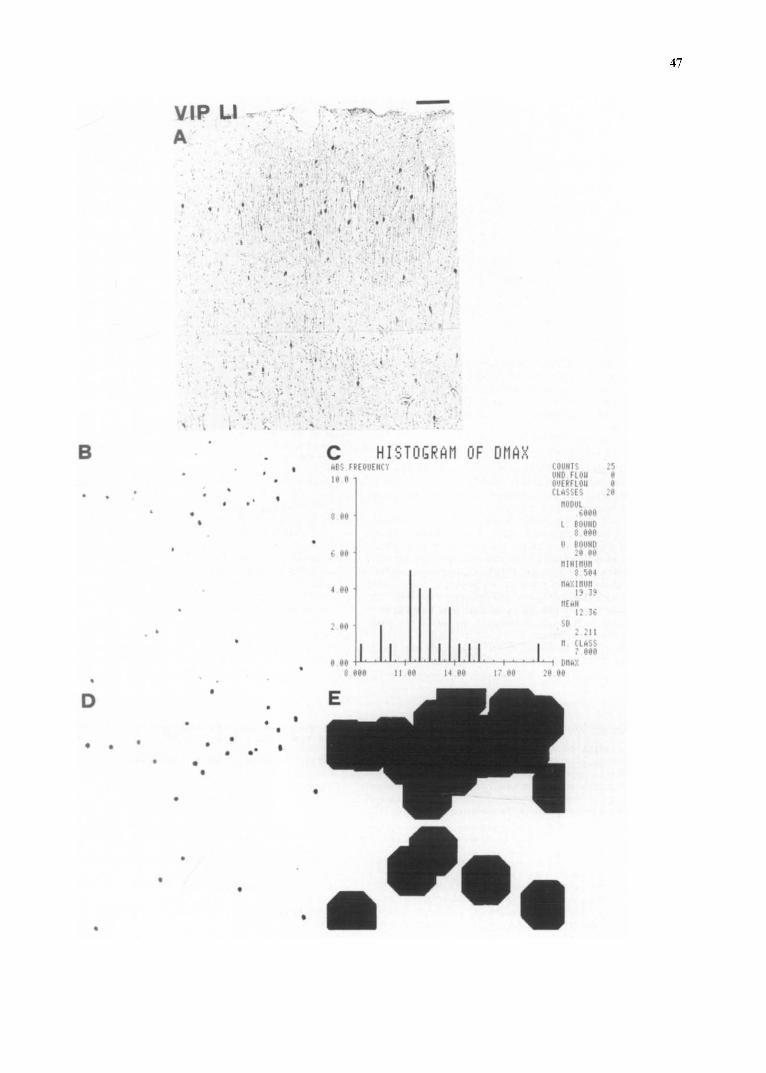
Fig. 2. Schematic description of the morphometrical analysis of the immunocytochemical preparations: automatic counting (B and C) and test for uniformity of distribution (D and E). A: microphotograph of vasoactive intestinal polypeptide-like immunoreactive cell bodies in the external part of the frontoparietal cortex. PAP technique, antibody dilution = 1:150 after reconstitution with 500/A. Level KK 8920 according to the K6nig and Klippel atlas 49. Bar = 100 #m. B: discrimination of image A, to selectively visualize the specifically stained nerve cell body profiles. C: histogram of the maximum diameters of the discriminated cell body profiles of image B. A threshold (8/~m) was selected tO avoid the counting of small positive profiles, such as big dendrites or fragments of cell bodies. From this histogram the number of positive profiles is obtained. D: normalization of size and shape of the discriminated profiles of image B to obtain circular objects, suited for the test for uniformity. E: dilatation of the objects of image D. The dilatation procedure was repeated n-1 times, n being the number of repetitions necessary for the merging of homogeneously distributed profiles. The index of uniformity (IU) is calculated as follows: [ (N ' - I ) / (N- l ) ] × 100, where N = number of profiles before dilatation (= 25, see image D) and N" = number of profiles after dilatation (= 5, see image E). In the present example IU = (5-1 / 25-1) × 100 = 17. For further details see Materials and Methods.
47
B
D
$ •
VI -P L I ~.~ .
A
• +
: . I+ t . .
" . . . " t ~+..+ "+ . " : . 7 ; . +" + , . " ' ~
" " ' + ' ! 41+~ + | • : + : , . + ! •
• i '-: f + . i !
+ | . +
+. p
$ L
$
$
I
l I I
I
I
I $ a |
, " . I $
. " . . . ~
I
v
.+
: - t
t
C H 1:7 T 0 G R A M 0 F D PIA,~.:: AE',S F PEOI.IEN£','
150
:7 ~15
2 (If1 (
::: 585 ]l 05 14 05
CSUNTS Ull[i FL5W 5HEIi'FLSH CLHSSES
1150UL
L BOUND
il E:5UHD LV rm
II]II IMVII
IIH', 1111111 i? ~q
IIEHN
s[b 2 ::'lt
H r L H L--: L-, I 7 5rm
' ' I DMS',: 17 UV .~A gU
I v
Lg IJ
20

48
Fig. 3. Schematic representation of the sampling procedure used to analyze the pattern of neuropeptide Y-positive profile distribution in the nucleus caudatus-putamen. Level KK 8920 according to the Ktinig and Klippel atlas 49. DL, dorsolateral; DM, dorsomedial; FS, fundus striati; VL, ventrolateral; VM, ventromedial. For further details see text.
the sampled areas by the TV camera from the microscope. The fields analyzed at each time were usually 597 × 597 /~m. A discriminative procedure was performed in order to separate the positive cell bodies from the non-specific background3(see Fig. 2b). Counting of small positive profiles, such as fragments of cell bodies or large dendrites, was avoided by setting a lower threshold for the acceptance of profiles to be counted. Some morphometrical param- eters of the resulting profiles were obtained: number, area and maximal diameter of profiles (see Fig. 3). In the present analysis, due to the scattered distribution of cell bodies, a discriminated profile rarely corresponded to more than one positive cell body. The results obtained with the automatic procedure were found to be in good agreement with those obtained by means of the direct inspection of the specimen.
An analysis of the homogeneity of VIP-positive cell body profile distribution in the cerebral cortex was also performed (see Fig. 2).
To this end, the discriminated images (Fig. 2b) were processed t¢~ erase small positive profiles (see above) and to normalize the size and shape of the remaining profiles (see Fig. 2d). On the resulting image the 'dilate' function was used (see Fig. 2e). The 'dilate' function increases the size of discriminated objects by 2 pixels at the boundaries. The number of profiles was evaluated on the discrim- inated image (N) and after dilatation (N'). Thus, close by profiles merge and the number of profiles present m the resulting image will decrease. Dilatation can be repeated several times. A criterion to select the number of times to repeat the dilatation step was assessed as follows:
By assuming the presence of circular profiles of equal size, it is possible to obtain the side (S) of the theoretical unitary square which contains one profile from the square root of the ratio S = ~/SATN, where SA = sampled area, N = number of profiles. Obviously, S also corresponds to the mean distance between the centers of two profiles. Then, the mean free distance (A) between the borders of two close by profiles is given by the formula A - S-D, where D = mean diameter of the profile. Finally, it can be stated that the theoretical number of pixels (P) by which the profiles should be dilated to merge together is given by P = (A/K) × 4, where K = conversion factor from microns to pixels. Thus, according to this procedure we can state that the number of repetitions (R) of the dilatation function just enough not to reach the merging of evenly distributed profiles is equal to R = 17--l.
On this basis an index of uniformity (IU) can be developed: the IU is the percent ratio between ( N ' - I ) and (N--l). This index can take any value between 100%, when an even distribution is present (so that no reduction in the number of profiles will occur upon dilatation), and 0%, when a maximum concentration of profiles is present (so that the number of profiles after dilatation will be equal to 1).
With our equipment the IU procedure can be used when the size of the field to be analyzed is not too large (less than 1000/~m by side). Since VIP-positive neurons are evenly distributed in the frontoparietal cortex 63, 3 representative fields in the lateral part were selected. In the analysis of MAM Ac-exposed rats, the SA was corrected according to the reduction of the cortical area.
In the case of nucleus caudatus-putamen a semiautomatic ap- proach was employed. In fact, the nucleus caudatus-putamen had to be considered in its entireness in view of the possible heterogeneity of NPY-positive neuron distribution. The homogeneity of NPY- positive cell bodies in the nucleus caudatus-putamen was analyzed by means of the Gini index 2. To this aim the sampled areas were subdivided in unitary squares whose side was equal to the
TABLE I
M orphometrical analysis of some telencephalic regions
The values are expressed as area of the selected region per section. The location of the areas measured at level KK 8920 (a) and/or at level KK 3290 (b) is shown in Fig. 1. Means + S.E.M. are shown, n = 4/group. Acb, nucleus accumbens; CPu, nucleus caudatus-putamen; DG, dentate gyrus; FrPa, frontoparietal cortex; HF, hippocampat formation; HG, hippocampal gyrus; Tu, tuberculum olfaetorium.
Brain Control MAM Ac-exposed (MAM/CTRL) region (ram 2) (ram 2) ×t00
FrPa (a) 13.8+0.2 8 .6+0.1"* 62.3 FrPa (b) 12.1 +0.1 6 .0+0 .2 ** 49.6 HF(b) 5 .7+0 .1 3 .0+0.1"* 52.6 H G ( b ) 4 .7+0 .1 2.2_+0.1 ** 46.8 D G ( b ) 1.0 + 0.1 0.8_+0.1"* 80.0 CPu (a) 8.1 + 0.2 6.1 -+ 0.2** 75.0 Acb (a) 3.1 +0.1 2.1 _+ 0.1"* 68.4 Tu(a) 1 .7+0.1 1.1_+0.1 ** 64.7
**P < 0.01, Mann-Whitney U-test.
TABLE II
Morphometrical analysis of peptide-like immunoreactivities in the frontoparietal cortex (level KK 8920)
Means _+ S.E.M. are shown, n = 4 rats/group.
Treatment Number of Number of Maximum cells in the cells per diameter region mm 2 (t~m)
(a) Vasoactive intestinal polypeptide-like immunoreactivity Control 194.5 _+ 16.2 14.1 + 1.2 12.3 + 1.3 MAM Ac-exposed 188.5 _+ 5.2 21.9 + 0.6** 12.1 + 1.1
(b) Cholecystokinin-like immunoreactivity Control 166.9+7.3 12.1 +0.5 11.6+0.5 MAM Ac-exposed 101.5+4.9"* 11.8_+0.8 I1 .6+0.1
(c) Neuropeptide Y-like immunoreactivity Control 85.2_+4.1 6.2_+0.3 13.3___0.6 MAM Ac-exposed 55.7 + 2.3** 6.5 _+ 0.3 13.3 _+ 0.4
**P < 0.01, Mann-Whitney U-test.

49
i . ~ ~1 i./{• • • . ; e ~ • .~' ~ ,
" . ' " - " . J ' : t
I I ° ' I~
" " ~ II
.!
i ,
I
i /
. ~t~
P l
L
j." . .
/
L
, j
, " ** . 0
I '
D . . .
|
lip
¢
!
' I
w .
f , . ~
°
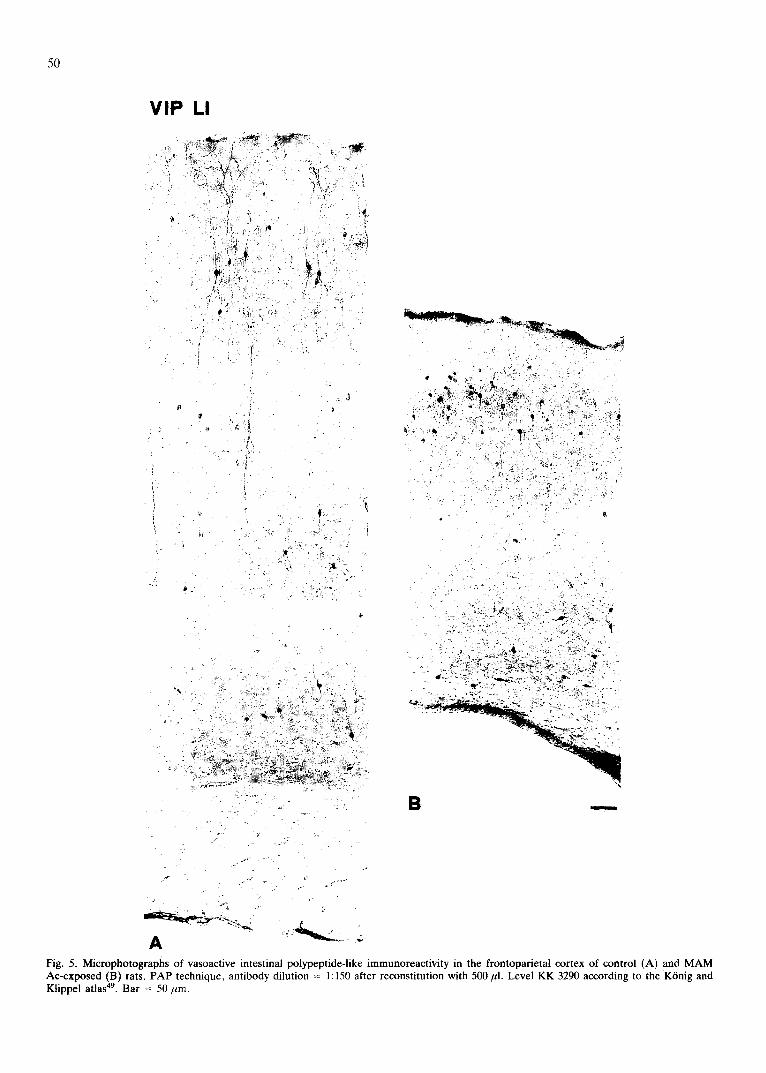
Fig. 4. Microphotographs of vasoactive intestinal polypeptide-like immunoreactivity in the frontoparietal cortex of control (A) and MAM Ac-exposed (B) rats. PAP technique, antibody dilution = 1:150 after reconstitution with 500 ~1. Level KK 8920 according to the KOnig and Klippel atlas 49. Bar = 100 ~m.
50
VIP LI
• ~ ; ~ , ~ _
".ei ;,
' ~4
d
• 0
. . . . . . . .
):~ . . . . ,?:
-d
B _ _ .
Fig. 5. Microphotographs of vasoactive intestinal polypeptide-like immunoreactivity in the frontoparietal cortex of control (A) and MAM Ac-exposed (B) rats. PAP technique, antibody dilution = 1:150 after reconstitution with 500 pl. Level KK 3290 according to the K6nig and Klippel atlas 49. Bar = 50 pro.
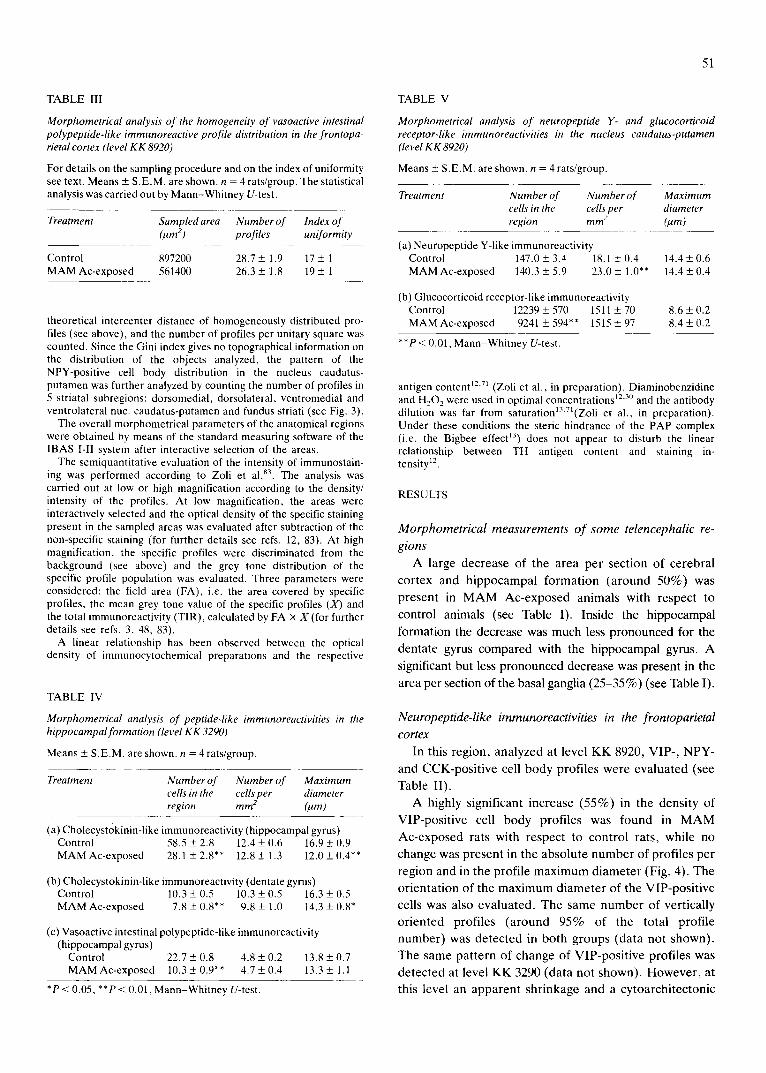
TABLE III
Morphometrical analysis of the homogeneity of vasoactive intestinal polypeptide-like immunoreactive profile distribution in the frontopa- rietal cortex (level KK 8920)
For details on the sampling procedure and on the index of uniformity see text. Means _+ S.E.M. are shown, n = 4 rats/group. The statistical analysis was carried out by Mann-Whitney U-test.
Treatment Sampled area Number of Index of (~din 2) profiles uniformity
Control 897200 28.7 + 1.9 17 + 1 MAM Ac-exposed 561400 26.3 +- 1.8 19 +- 1
theoretical intercenter distance of homogeneously distributed pro- files (see above), and the number of profiles per unitary square was counted. Since the Gini index gives no topographical information on the distribution of the objects analyzed, the pattern of the NPY-positive cell body distribution in the nucleus caudatus- putamen was further analyzed by counting the number of profiles in 5 striatal subregions: dorsomedial, dorsolateral, ventromedial and ventrolateral nuc. caudatus-putamen and fundus striati (see Fig. 3).
The overall morphometrical parameters of the anatomical regions were obtained by means of the standard measuring software of the IBAS I-II system after interactive selection of the areas.
The semiquantitative evaluation of the intensity of immunostain- ing was performed according to Zoli et al. ~3. The analysis was carried out at low or high magnification according to the density/ intensity of the profiles. At low magnification, the areas were interactively selected and the optical density of the specific staining present in the sampled areas was evaluated after subtraction of the non-specific staining (for further details see refs. 12, 83). At high magnification, the specific profiles were discriminated from the background (see above) and the grey tone distribution of the specific profile population was evaluated. Three parameters were considered: the field area (FA), i.e. the area covered by specific profiles, the mean grey tone value of the specific profiles (X) and the total immunoreactivity (TIR), calculated by FA × X (for further details see refs. 3, 48, 83).
A linear relationship has been observed between the optical density of immunocytochemical preparations and the respective
TABLE IV
Morphometrical analysis of peptide-like immunoreactivities in the hippocampal formation (level KK 3290)
Means + S.E.M. are shown, n = 4 rats/group.
Treatment Number of Number of Maximum cells in the cells per diameter reg ion m m 2 (txm)
(a) Cholecystokinin-like immunoreactivity (hippocampal gyrus) Control 58.5 +_ 2.8 12.4 _+ 0.6 16.9 _+ 0.9 MAM Ac-exposed 28.1 -+ 2.8** 12.8 _+ 1.3 12.0 _+ 0.4**
(b) Cholecystokinin-like immunoreactivity (dentate gyrus) Control 10.3 + 0.5 10.3 + 0.5 16.3 _+ 0.5 MAM Ac-exposed 7.8 _+ 0.8** 9.8 + 1.0 14.3 + 0.8*
(c) Vasoactive intestinal polypeptide-like immunoreactivity (hippocampal gyrus)
Control 22.7 + 0.8 4.8 _+ 0.2 13.8 _+ 0.7 MAM Ac-exposed 10.3+0.9"* 4.7_+0.4 13.3_+1.1
*P < 0.05, **P < 0.01, Mann-Whitney U-test.
51
TABLE V
Morphometrical analysis of neuropeptide Y- and glucocorticoid receptor-like immunoreactivities in the nucleus caudatus-putamen (level KK 8920)
Means + S.E.M. are shown, n = 4 rats/group.
Treatment Number of Number of Maximum cells in the cells per diameter region mm 2 (/~m)
(a) Neuropeptide Y-like immunoreactivity Control 147.0+3.4 18.1 +_0.4 14.4+-0.6 MAM Ac-exposed 140.3 +_ 5.9 23.0 + 1.0"* 14.4 _+ 0.4
(b) Glucocorticoid receptor-like immunoreactivity Control 12239 _+ 570 1511 + 70 8.6 + 0.2 MAM Ac-exposed 9241 + 594** 1515 +_ 97 8.4 + 0.2
**P < 0.01, Mann-Whitney U-test.
antigen content ~2'7~ (Zoli et al., in preparation). Diaminobenzidine and H 2 O 2 w e r e used in optimal concentrations '2"3° and the antibody dilution was far from saturation13"71(Zoli et al., in preparation). Under these conditions the steric hindrance of the PAP complex (i.e. the Bigbee effect 13) does not appear to disturb the linear relationship between TH antigen content and staining in- tensity 12.
RESULTS
Morphometrical measurements o f some telencephalic re-
gions
A large dec rease of the a rea pe r sec t ion of ce rebra l
cor tex and h i p p o c a m p a l f o r m a t i o n ( a round 5 0 % ) was
p resen t in M A M A c - e x p o s e d animals wi th respect to
control animals (see Table I). Inside the h ippocampal
format ion the decrease was much less p ronounced for the
denta te gyrus compared with the h ippocampal gyrus. A
significant but less p ronounced decrease was present in the
area per section of the basal ganglia ( 2 5 - 3 5 % ) (see Table I).
Neuropeptide-like immunoreactivities in the frontoparietal
cortex
In this reg ion , ana lyzed at level K K 8920, VIP- , N P Y -
and C C K - p o s i t i v e cell b o d y prof i les were e v a l u a t e d (see
Table II) .
A highly s ignif icant increase (55%) in the dens i ty of
VIP-pos i t ive cell body prof i les was found in M A M
A c - e x p o s e d rats with respec t to con t ro l rats, whi le no
change was p re sen t in the abso lu te n u m b e r of prof i les pe r
reg ion and in the prof i le m a x i m u m d i a m e t e r (Fig. 4). T h e
o r i en ta t ion of the m a x i m u m d i a m e t e r of the VIP-pos i t i ve
cells was also eva lua ted . T h e s a m e n u m b e r of ver t ical ly
o r i en t ed prof i les ( a round 95% of the total prof i le
n u m b e r ) was d e t e c t e d in bo th g roups (da ta no t shown) .
The same pa t t e rn of change of VIP-pos i t i ve prof i les was
d e t e c t e d at level K K 3290 (da ta no t shown) . H o w e v e r , at
this level an appa ren t sh r inkage and a cy toa rch i t ec ton ic
52
CCK, rL|
t , - . -
• ,72:"
q
A
• 4 '
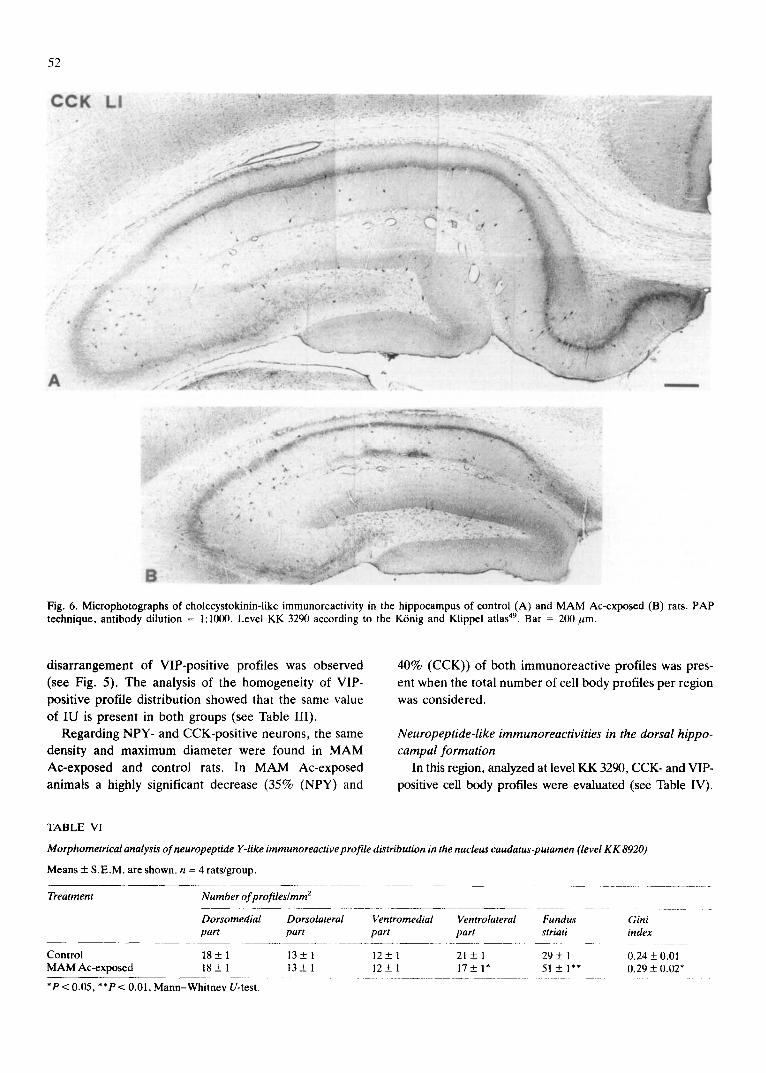
B:ii'i Fig. 6. Microphotographs of cholecystokinin-like immunoreactivity in the hippocampus of control (A) and M A M Ac-exposed (B) rats. PAP technique, antibody dilution = 1:1000. Level KK 3290 according to the K6nig and Klippel atlas 49. Bar = 200 rum.
disarrangement of VIP-positive profiles was observed (see Fig. 5). The analysis of the homogeneity of VIP- positive profile distribution showed that the same value of IU is present in both groups (see Table III).
Regarding NPY- and CCK-positive neurons, the same density and maximum diameter were found in MAM Ac-exposed and control rats. In MAM Ac-exposed animals a highly significant decrease (35% (NPY) and
40% (CCK)) of both immunoreactive profiles was pres- ent when the total number of cell body profiles per region was considered.
Neuropeptide-like immunoreactivities in the dorsal hippo- campal formation
In this region, analyzed at level KK 3290, CCK- and VIP- positive cell body profiles were evaluated (see Table IV).
TABLE VI
Morphometrical analysis of neuropeptide Y-like immunoreactive profile distribution in the nucleus caudatus-putamen (level KK 8920)
Means + S.E.M. are shown, n = 4 rats/group.
Treatment Number of profiles/mm 2
Dorsomedial Dorsolateral part part
Control 18 + 1 13 + 1 M A M Ac-exposed 18 + 1 13 _+ 1
*P < 0.05, **P < 0.01, Mann-Whitney U-test.
Ventromedial Ventrolateral Fundus Gini part part striati index
12_+ l 21 + 1 29_+ 1 0.24 + 0.01 1 2 + 1 17_+ l* 51 _+ 1"* 0 . 2 9 + 0 . 0 2 *
53
NP¥ U "
%
~En
A
?
\
i
P
t •
°
x , o . ~
!
L
I
~qk,
j -
. k ~ . . ~ ~ . , ~ : ~ •
¢
E.* ,~ -. :.r~i~ , - *" " .-2
@ -
B "
zv " ].,° ,~,
, , f " 1
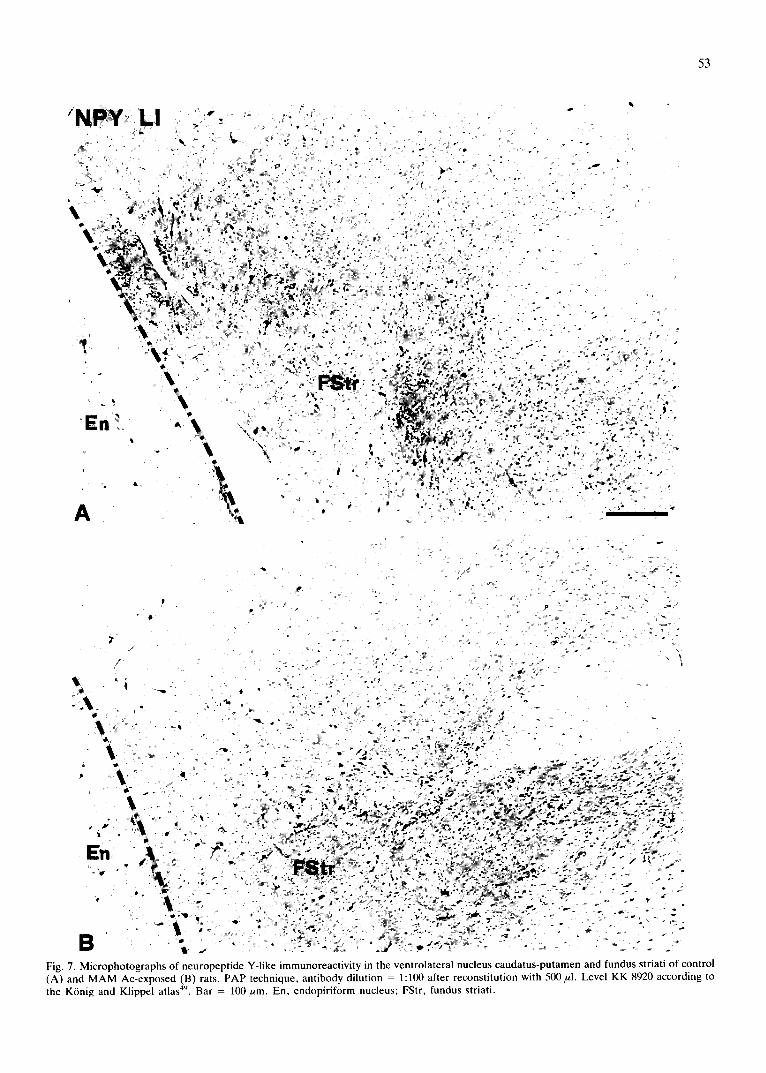
Fig. 7. Microphotographs of neuropeptide Y-like immunoreactivity in the ventrolateral nucleus caudatus-putamen and fundus striati of control (A) and MAM Ac-exposed (B) rats. PAP technique, antibody dilution = 1:100 after reconstitution with 500/~l. Level KK 8920 according to the K6nig and Klippel atlas 49. Bar = 100/~m. En, endopiriform nucleus; FStr, fundus striati.
54
k !
B
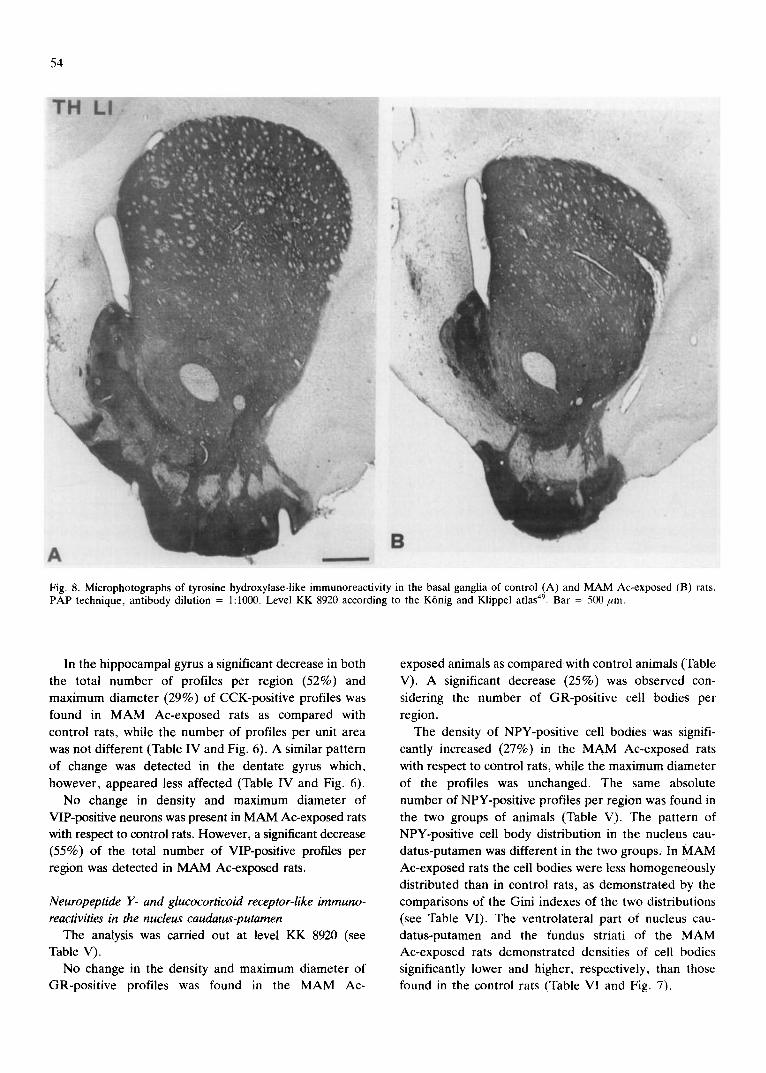
Fig. 8. Microphotographs of tyrosine hydroxylase-like immunoreactivity in the basal ganglia of control (A) and MAM Ac-exposed (B) rats. PAP technique, antibody dilution = 1:1000. Level KK 8920 according to the K6nig and Klippel atlas 49. Bar = 500 pro.
In the hippocampal gyrus a significant decrease in both the total number of profiles per region (52%) and maximum diameter (29%) of CCK-positive profiles was found in MAM Ac-exposed rats as compared with control rats, while the number of profiles per unit area was not different (Table IV and Fig. 6). A similar pattern of change was detected in the dentate gyrus which, however, appeared less affected (Table IV and Fig. 6).
No change in density and maximum diameter of VIP-positive neurons was present in MAM Ac-exposed rats with respect to control rats. However, a significant decrease (55%) of the total number of VIP-positive profiles per region was detected in MAM Ac-exposed rats.
Neuropeptide Y- and glucocorticoid receptor-like immuno- reactivities in the nucleus caudatus-putamen
The analysis was carried out at level KK 8920 (see Table V).
No change in the density and maximum diameter of GR-positive profiles was found in the MAM Ac-
exposed animals as compared with control animals (Table V). A significant decrease (25%) was observed con- sidering the number of GR-positive cell bodies per region.
The density of NPY-positive cell bodies was signifi- cantly increased (27%) in the MAM Ac-exposed rats with respect to control rats, while the maximum diameter of the profiles was unchanged. The same absolute number of NPY-positive profiles per region was found in the two groups of animals (Table V). The pattern of NPY-positive cell body distribution in the nucleus cau- datus-putamen was different in the two groups. In MAM Ac-exposed rats the cell bodies were less homogeneously distributed than in control rats, as demonstrated by the comparisons of the Gini indexes of the two distributions (see Table VI). The ventrolateral part of nucleus cau- datus-putamen and the fundus striati of the MAM Ac-exposed rats demonstrated densities of cell bodies significantly lower and higher, respectively, than those found in the control rats (Table VI and Fig. 7).

[ \ t
- t J _.,'U
[~ 240 pm/side
1 FrPa external layers
2 CPu central part
3 Perirhinal Cortex
Fig. 9. Schematic representation of the sampling procedure used for the high magnification microdensitometrical analysis of tyrosine hydroxylase-like immunoreactivity. Level KK 8920 according to the K6nig and Klippel atlas 4'*.
Tyrosine hydroxylase-like immunoreactivity in various
forebrain areas
Microdensitometrical analysis of TH-like immunore-
activity in the forebrain (level KK 8920) was carried out
at low and high magnification according to the density/ intensity of positive profiles.
The analysis at low magnification was carried out in nucleus caudatus-putamen, nucleus accumbens and tu-
berculum olfactorium, areas where a high density/inten-
TABLE VIII
High magnification densitometrical analysis of tyrosine hydroxylase- like immunoreactivity in the forebrain
The sampled areas (square of 57600/xm ~) are shown in Fig. 9. Means +_ S.E.M. are shown, n = 4 rats/group).
Treatment Mean grey Field area Total value (~m 2 ) immunoreactivity (arbitrary (arbitrary units) units)
(a) Nucleus caudatus-putamen (central part) Control 54 ± 2 48360 ± 1097 2613 + 65 MAM Ac-exposed 53 ± 2 48955 + 1960 2590 ± 108
(b) Frontoparietal cortex (external layers) Control 38 ± 1 290 ± 40 11 + 1 MAM Ac-exposed 38 ± 1 696 _+ 49** 26 + 2"*
(e) Perirhinal cortex Control 38 _+ 1 794 + 73 30 ± 3 MAMAc-exposed 38± 1 1877 ± 278** 71 ± 10"*
**P < 0.01, Mann-Whitney U-test.
55
TABLE Vll
Low magnification densitometrical analysis of tyros±he hydroxylase- like immunoreactivity in the forebrain
The sampled areas are shown in Fig. 1. Means ± S.E.M. are shown. n = 4 rats/group. The statistical analysis was carried out by Mann-Whitney U-test.
Treatment Nucleus Nucleus Tuberculum caudatus- accumbens olfactorium putamen (optical (optical (optical density, density, density, arbitrary arbitrary arbitrary units) units') units')
Control 175 _+ 8 187 + 8 238 ± 7 MAM-Ac-exposed 172 ± 4 192 _+ 4 240 ± 5
sity of TH-positive profiles is present. No change in the
optical density of either region was present in the 2
groups (Table VII and Fig. 8).
The analysis at high magnification was carried out on the external part of the frontoparietal cortex and the
perirhinal cortex, where a low density of TH-positive
profiles is present, and on the central part of the nucleus
caudatus-putamen (Fig. 9). In M A M Ac-exposed rats, a
highly significant increase of the field area and total immunoreactivity of TH-positive profiles was found both
in the frontoparietal and perirhinal cortices, while the
mean grey tone value was unchanged (Table VII and Fig.
10). No change of the field area and mean grey tone value was detected in the nucleus caudatus-putamen.
DISCUSSION
The present morphometrical results are in agreement with previously reported data on the hypoplasia of telencephalic areas in M A M Ac-exposed animals 32'
4°.The hippocampal gyrus (but not the dentate gyrus) and
cerebral cortex appear to be the most affected CNS areas
following M A M Ac exposure at day E15. In addition, a
decrease of similar entity was observed in the area per section of the basal ganglia in M A M Ac-exposed rats, as
could be expected, since these areas develop in the same prenatal period 67.
In agreement with previous neurochemicai data on
classical transmitters 1~'4°, the results obtained in the cerebral cortex, the hippocampal formation and the nucleus caudatus-putamen on peptide-containing neuro- nal populations in M A M Ac-exposed rats demonstrated two patterns of changes: (1) a maintained total number of neurons per region together with an increased density (due to the fact that the regions are smaller in the M A M Ac-exposed rats), or (2) a decreased total number per

56
TH LI i 1 ~ •
" I
| -
4
v
f
o - . ,
: " 2i
J
!
t
[ , " - o
' ¶ . -
1
A Fig. 10. Microphotographs of tyrosine hydroxylase-like immunoreactivity in the external part of the frontoparietal cortcx m control (A) and MAM Ac-exposed (B) rats. PAP technique, antibody dilution = 1:1000. Level KK 8920 according to the KOnig and Klippel atlas 49. Bar = 20/~m.
region together with a maintained density. The first pattern was recognized for VIP-positive neurons in the cerebral cortex and for NPY-positive neurons in the nucleus caudatus-putamen. The second pattern was detected for NPY- and CCK-positive neurons in the cerebral cortex and for VIP- and CCK-positive neurons in the hippocampal formation•
These different patterns of changes do not seem to be related to the cell content of neuropeptides or other known substances. In fact, striatal and cortical NPY neurons costore somatostatin-like immunoreactivity 76'79 and are positive for N A D P H diaphorase enzyme 77"78. N A D P H diaphorase-positive cells, called 'solitary active cells' by Thomas and Pearse72,were shown to be resistant to various toxic agents 9"73,. However, the vulnerability of
cortical NPY-positive neurons to M A M Ac exposure suggests that the resistance shown by striatal NPY- positive neurons is not related to their content of neuropeptides or N A D P H diaphorase.
It can be hypothesized that the differential vulnera- bility of peptide-containing neurons after M A M Ac exposure is related to their developmental characteristics. Peptide-containing neurons appear in the cortical plate in the late prenatal/early postnatal period 21'59~6°'74"81. In the
perinatai days these neurons migrate and reach their adult location in the cortex, The adult morphology is reached within 2 -4 weeks. Cortical VIP neurons seem to develop somewhat later than other peptidergic neurons, such as the CCK-, somatostatin- and NPY-immunoreac- tive neurons 16'59'6°'74'81 . It was demonstrated that starting

from postnatal days 5-7, clusters of weakly VIP-immu- noreactive cell bodies can be detected in the deep layers.
At the beginning of the second postnatal week the cell bodies have a sparse distribution similar to that of the adult animal. The number of varicose terminals increase from postnatal day 7 to day 2821. Based on the evidence of the late expression, migration and maturation of VIP-immunoreactive neurons in the cortex, it has been hypothesized that these neurons have a late period of mitosis 21, even if direct evidence is still lacking.
Neurons of the cortical region actively divide in the period between El3 and E2137, following a reverse order of layer formation so that the cells which will be localized
in the deep layers of the adult animal have an early division time 4"36. The alkylating agent M.AM Ac injected
on day El5 kills rapidly dividing cells which will migrate to layers I I - I V in the adult animal, whereas it spares neurons of layers VI and V which already completed their division phase 4°. In the adult rat, VIP-positive
perikarya are mainly located in layers I I - I I I , which are severely damaged after MAM Ac exposure on day E15. The present evidence of the sparing of VIP-immunore- active cortical neurons after M.AM Ac exposure gives further support to the hypothesis of a relatively delayed
time of final mitosis of this class of neurons with respect to other neurons of the same layers.
Furthermore, the maintenance of the number of VIP-positive neurons in adulthood indicates that the survival of these cells is not influenced by the reduction
in the number of the surrounding neuronal populations. It can be hypothesized that VIP cortical neurons, like brainstem monoamine neurons, which hyperinnervate the telencephalic areas of the microencephalic animal, strictly follow a genetical programme, which is not influenced by the density of other populations of intrinsic neurons. -Alternatively, the selectivity of VIP neuron sparing after M.AM .Ac exposure could be due to the epigenetic influence of specific anatomical and/or func- tional arrangements. In particular, in the adult cerebral cortex, it has been demonstrated 54'55 that VIP-immuno-
reactive interneurons have strict functional interactions with noradrenaline nerve terminals, which are spared by MAM .Ac exposure. Several authors investigated the possible relevance of the monoaminergic innervation for the development of the cerebral cortex 17'26"65. A detailed
analysis of the effects of prenatal treatment with 6- hydroxydopamine on cortical structure did not support the hypothesis of a major regulatory role of monoamines on cortical development 51. However, it was demon- strated that the postnatal lesion of monoaminergic pathways affects plastic changes related to the ocular d o m i n a n c e shift 29"47 and cognitive performance in rats housed in an enriched environment 62. The possible
57
influence of the noradrenergic innervation on the devel-
opment of discrete neuronal populations, such as VIP-
immunoreactive neurons, remains to be investigated. VIP-positive neurons are bipolar cells with vertical
dendritic arborization, which spans from layer I to layer V and only minimally extend in the horizontal plane 27"54.
They are uniformly distributed in the rat cerebral cortex 63
at a mean distance of 60/~m and represent approximately 1% of the overall number of cortical neurons 54. The
anatomical evidence indicates that VIP neurons exert
input-output functions in small, radially oriented cortical domains, so that all these domains of the cerebral cortex
are under the influence of one VIP neuron. Both anatomical, biochemical and functional data suggest a specific involvement of VIP-immunoreactive neurons in the local control of cortical blood f low 20"39"50"80 and
energetic metabolism of cortical ceUs 56. On this basis, it
was hypothesized that VIP neurons can play a central role in the control of the coupling between neuronal activity, blood flow and energy metabolism of the smallest morphofunctional unit of the cerebral cortex, the minicolumn53.54,64.
The analysis of the distribution of VIP-positive neu- rons indicates that the absolute number and spatial relationships among VIP neurons are not altered in M.AM Ac-exposed rats. Based on the present evidence it may be hypothesized that the number of basic morpho- functional elements in the cerebral cortex, i.e. the number of VIP-controlled minicolumns 54, is preserved
after MAM .Ac exposure, whereas their size, i.e. the number of neurons in the minicolumn, is decreased. This hypothesis is indirectly supported by the finding that, in spite of the observed cytoarchitectonic abnormalities 4°,
normal efferent and afferent connections are established by surviving cortical cells 44 and no change is present in
the pattern of vascularization and cytochrome oxidase activity in the cerebral cortex after prenatal exposure to M.AM Ac 6.
In the hippocampal formation the number of CCK- and VIP-positive neurons shows a decrease proportional to the reduction of the overall dimension and, possibly, of the overall number of cells of the region. CCK- positive, but not VIP-positive, cell bodies of M.AM .Ac-exposed rats were significantly smaller than those in the control ra t s . .A previous report 68 demonstrated a severe disarrangement of hippocampal cytoarchitecture, in particular a reduction and a deformation of dendritic branches and spines. The reduction in the maximum diameter of CCK-positive neurons could be due either to a real decrease in the size of the CCK-positive cells or to a decrease in the content of CCK-like immunoreactive material. In both cases, an alteration of this neuronal population in M.AM Ac-exposed rats can be suggested,

58
notwithstanding the maintenance of the cell density. This alteration may tentatively be related to some aspects of cognitive deficits observed in MAM Ac-exposed rats, such as passive avoidance and maze learning s''~34.
Evidence indicates that CCK peptides may play a role in facilitating memory, since intraventricularly injected CCK-related peptides increase, and CCK-8 anti-
serum decreases the latency of passive avoidance in rats23,46
The density of nerve cells in the nucleus caudatus- putamen was evaluated by counting the number of GR-positive profiles per sampled area. In this region the GR antiserum stains the nuclei of the majority (around 90%) of the cell bodies28(Zoli et al., in preparation). By
means of this method no change in GR-positive profile density and maximum diameter was found. This evidence confirms that the observed decrease in striatal volume
and weight in MAM Ac-exposed rats is due to a decrease of the total number of striatal cell bodies 7. However, it
may be suggested that these cells still maintain at least the main features of their reciprocal morphological relations and input-output connections. In fact, there is evidence for intact glutamate 7'1~ and dopamine innervation, as shown by the unchanged density of TH-positive nerve terminals and TH-positive islands (Zoli et al., unpub- lished observations).
The maintenance of the total number of NPY-positive cell bodies (which are not GR-positive 35) indicates that
some striatal cell body subpopulations are differentially affected by MAM Ac exposure. Note that this neuronal population shows an altered distribution in the striatal region. Developmental studies 25's~ demonstrated that
NPY-positive neurons first appear in a region corre- sponding to the ventrolateral striatum at El8 and spread to the medial and dorsal part of nucleus caudatus- putamen as well as to the nucleus accumbens and tuberculum olfactorium between P4 and P10. This evi- dence suggests that the concentration of NPY-positive cells in the most ventral and lateral portion of the nucleus caudatus-putamen (fundus striati) of MAM Ac-exposed rats might be due to a partial blockade of the develop- mental spreading of NPY-positive cells in the basal ganglia. Therefore, the known alterations in striatal neurochemistry H'33 and in striatally controlled behaviour a4 could be due not only to the overall reduction of the striatal mass but also to the changes of the entity and distribution inside the striatum of small transmitter-identified neuronal populations.
The present findings of an increased density of TH-like immunoreactive profiles in the external part of the frontoparietal cortex suggests the presence of a norad- renergic hyperinnervation of the neocortex of MAM Ac-exposed rats, in agreement with previous data 42. A
similar increase of TH-like immunoreactivity was ob- served in the perirhinal cortex, where TH is a marker also of dopamine-containing nerve terminals 223s,52. On the
other hand, no effect of MAM Ac exposure on TH-like immunoreactivity was found in the basal ganglia. The
densitometrical analysis in the nucleus caudatus-puta- men, both at a low and high magnification, did not show any change in the density/intensity of TH-like immuno- reactive nerve terminal profiles, suggesting that a similar density of dopamine terminals is present in MAM Ac-exposed and control rats. Previous data showed both an increase of TH enzyme activity ~ and of dopamine levels and turnover 33. However, these findings may be
related to an increased activity of the striatal dopamine terminal system rather than to an increase of dopamine nerve terminal number. Alternatively, these differences may be related to the different age of the rats used in the present and previous studies. In fact, recent evidence indicates that the forementioned phenomena are age-
dependent. Measurement of noradrenergic and dopamin- ergic markers in the cerebral cortex and striatum, respectively, of MAM Ac-exposed rats killed at 1, 2 and 4 months of age showed an increase of these markers at 1 and 2 months of age with respect to control animals, followed by a partial normalization at 4 months of age ~. This fact may reflect an adaptive reorganization of noradrenergic cortical projections by age, or a delayed
target dependent neuronal death. The existence of an extrapyramidal impairment in MAM Ac-exposed rats is supported by evidence of general hyperactivity ~4'3~ and of increased apomorphine-induced stereotypies sz.
Postnatal target dependent cell degeneration in the dorsal lateral geniculate nucleus due to severe neuron depletion in the occipital cortex induced by prenatal exposure to MAM Ac in rats was recently described by Ashwell 5. These thalamic neurons, which make direct synaptic contacts with cells of laminae III and IV of the occipital cortex, were shown to die in the first week of postnatal life. Neuronal systems characterized, at least in part, by non-synaptic mode of transmission, such as noradrenergic cortical x° and dopaminergic striataP 9 sys- tems, may at first not be affected by target cell degen- eration, probably because of their relative structural and functional independency from target neurons. However, the partial normalization of cortical noradrenaline con- taining terminals in 4-month-old rats exposed prenatally to MAM Ac I may support the existence of a delayed target-dependent neuronal degeneration process.
Acknowledgements. This work has been supported by grants (OYX-715) from the Swedish Medical Research Council, the Swedish Work Environmental Health Fund, Progetti Obiettivo Regione Marche and MPI 60%.

REFERENCES
1 Abbracchio, M.P., Cimino, M., Di Luca, M., Mennuni, L., Zaratin, P. and Cattabeni, F., Target-dependency of catechol- aminergic afferents in an animal model of microencephaly induced by prenatal methylazoxymethanol administration, Soc. Neurosci. Abstr., 14 (1988) 77.7.
2 Agnati, L.E, Fuxe, K., Benfenati, F., Zini, I,, Zoli, M., Fabbri, L. and H~irfstrand, A., Computer assisted morphometry and microdensitometry of transmitter identified neurons with special reference to the mesostriatal dopamine pathway. I. Method- ological aspects. Acta Physiol. Scand., Suppl. 532 (1984) 5-36.
3 Agnati, L.E, Fuxe, K., Zoli, M., Zini, I., H~irfstrand, A., Toffano, G. and Goldstein, M., Morphometrical and microden- sitometrical studies on phenylethanolamine-N-methyltransferase and neuropeptide Y immunoreactive neurons in the rostral medulla oblongata of the adult and old rats, Neuroscience, 26 (1988) 461-478.
4 Angevine, J.B. and Sidman, R.L., Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse, Nature (Lond.), 192 (1961) 766-768.
5 Ashwell, K.W.S., Direct and indirect effects on the lateral geniculate nucleus neurons of prenatal exposure to methyl- azoxymethanol acetate, Dev. Brain Res., 35 (1987) 199-214.
6 Ashwell, K.W.S. and Webster, W.S., Vascularity and cyto- chrome oxidase distribution in the occipital cortex in MAM Ac-induced microencephaly, Dev. Brain Res., 33 (1987) 301- 304.
7 Balduini, W., Abbracchio, M.P., Lombardelli, G. and Catta- beni, F., Loss of intrinsic striatal neurons after methylazoxy- methanol acetate treatment in pregnant rats, Dev. Brain Res., 15 (1984) 133-136.
8 Banff, S., Dorigotti, L., Abbracchio, M.P., Balduini, W., Coen, E., Ragusa, M.C. and Cattabeni, F., Methylazoxymethanol microcephaly in rats: neurochemical characterization and behav- ioural studies with the nootropic oxiracetam, Pharmacol. Res. Commun., 16 (1984) 67-83.
9 Beal, M.F., Kowall, N.W., Ellison, D.W., Mazurek, M.E, Swartz, K.J. and Martin, J.B., Replication of the neurochemical characteristics of Huntington's disease by quinolinic acid, Nature (Lond.). 321 (1986) 168-171.
10 Beaudet, A. and Descarries, L., The monoamine innervation of rat cerebral cortex: synaptic and non-synaptic axon terminals, Neuroscience, 3 (1978) 851-860.
11 Beaulieu, M. and Coyle, J.T., Effects of fetal methylazoxymeth- anol acetate lesion on the synaptic neurochemistry of the adult rat striatum, J. Neurochem., 37 (1981) 878-887.
12 Benno, R.H., Tucker, L.W., Joh, T.H. and Reis, D.J., Quan- titative immunocytochemistry of tyrosine hydroxylase in rat brain. I. Development of a computer assisted method using the peroxidase antiperoxidase technique, Brain Res., 246 (1982) 225-236.
13 Bigbee, J.W., Kosek, J.C. and Eng, L.E, Effects of primary antiserum dilution on staining of antigen-rich tissues with the peroxidase antiperoxidase technique, J. Histochem. Cytochem., 25 (1977) 443-447.
14 Cannon-Spoor, H.E. and Freed, W.J., Hyperactivity induced by prenatal administration of methylazoxymethanol: association with altered performance on conditioning tasks in rats, Phar- macol. Biochem. Behav., 20 (1984) 189-193.
15 Cattabeni, F., Abbracchio, M.P., Cimino, M., Cocchi, D., Di Luca, M., Mennuni, L., Rosi, F. and Zaratin, P., Methylazoxy- methanol-induced microencephaly: persistent increase of corti- cal somatostatin like immunoreactivity, Dev. Brain Res., in press.
16 Cho, H.J., Shiotani, Y., Shiosaka, S., Inagaki, S., Kubota, Y., Kiyama, H., Umegaki, K., Tateishi, K., Hashimura, E., Hamaoka, T. and Tohyama, M., Ontogeny of cholecystokinin- 8-containing neuron system of the rat: an immmunohistoche- mical analysis. I. Forebrain and upper brainstem, J. Comp.
59
Neurol., 218 (1983) 25-41. 17 Coyle, J.T. and Molliver, M.E., Major innervation of newborn
rat cortex by monoaminergic neurons, Science, 196 (1977) 444-447.
18 Dambska, M., Haddad, R., Kozlowski, P.B., Lee, M.H. and Shek, J., Telencephalic architectonics in the brains of rats with graded degree of microencephaly, Acta Neuropathol., 58 (1982) 203-209.
19 Descarries, L., Bosler, O., Berthelet, F. and Des Rosiers, M.H., Dopaminergic nerve endings visualized by high-resolution ra- dioautography in adult rat neostriatum, Nature (Lond.), 284 (1980) 620-622.
20 Eckenstein, F. and Baughmann, R.W., Two types of cholinergic innervation in cortex, one co-localized with vasoactive intestinal polypeptide, Nature (Lond.), 309 (1984) 153-155.
21 Emson, P.C., Gilbert, R.F.T., Loren, I., Fahrenkrug, J., Sundler, B.E and Schaffalitzhay, O.B., Development of vasoac- tive intestinal polypeptide (VIP) containing neurons in the brain, Brain Res., 177 (1979) 437-444.
22 Fallon, J.H., Koziell, D.A. and Moore, R.Y., Catecholamine innervation of the basal forebrain. II. Amygdala, suprarhinal cortex and entorhinal cortex, J. Comp. Neurol., 180 (1978) 509-532.
23 Fekete, M., Lonovics, J. and Telegdy, G., Modulation of passive avoidance behavior of rats by intracerebroventricular adminis- tration of cholecystokinin antiserum, Neuropeptides, 1 (1981) 363-369.
24 Fisher, M.H., Welker, C. and Waisman, H.A., Generalized growth retardation in rats induced by prenatal exposure to methylazoxymethanol acetate, Teratology, 5 (1972) 223-232.
25 Foster, G.A. and Schultzberg, M., Immunohistochemical anal- ysis of the ontogeny of neuropeptide Y immunoreactive neurons in the foetal rat brain, Int. J. Dev. Neurosci., 2 (1984) 387-407.
26 Fuxe, K., Lidbrink, P., Olson, L. and Seiger, A., The relation between paradoxical sleep and central monoamines. Ontogene- tic aspects. In W.P. Koella and P. Levin (Eds.), Sleep: Physiol- ogy, Biochemistry, Psychology, Pharmacology, Clinical Impli- cations, Karger, Basel, 1973, pp. 224-225.
27 Fuxe, K., H6kfelt, T., Said, S.I. and Mutt, V., Vasoactive intestinal polypeptide and the nervous system: immunohisto- chemical evidence for localization in central and peripheral neurons, particularly intracortical neurons of the cerebral cortex, Neurosci. Lett., 5 (1977) 241-246.
28 Fuxe, K., Wikstr6m, A.-C., Okret, S., Agnati, L.F., H~irfstrand, A., Yu, Z.-Y., Granholm, L., Zoli, M., Vale, W. and Gustafs- son, J.-A,, Mapping of glucocorticoid receptor immunoreactive neurons in the rat tel- and diencephalon using a monoclonal antibody against rat liver glucocorticoid receptor, Endocrinol- ogy, 117 (1985) 1803-1812.
29 Gordon, B., Moran, J., Trombley, P. and Soyke, J., Visual behavior of monocularly deprived kittens treated with 6- hydroxydopamine, Dev. Brain Res., 24 (1986) 21-29.
30 Gross, D.S. and Rothfeld, J.M., Quantitative immunocyto- chemistry of hypothalamic and pituitary hormones: validation of an automated, computerized image analysis system, J. Histo- chem. Cytochem., 33 (1985) 11-20.
31 Haddad, R.K. and Rabe, A., Intellectual deficit associated with transplacentally induced microcephaly in the rat, Science, 163 (1969) 88-90.
32 Hallman, H., Neurochernical Studies on Central Monoamine Neurons: Consequences of Selective Toxin Lesions, Ph.D. The- sis, Karolinska Institute, Stockholm, 1984.
33 Hallman, H. and Jonsson, G., Monoamine neurotransmitter metabolism in microencephalic rat brain after prenatal methyl- azoxymethanol treatment, Brain Res. Bull., 13 (1984) 383-389.
34 Hanada, S.T., Nakastuka, I. and Fuju, T., Effects of prenatal treatment with methylazoxymethanol acetate on growth, devel- opment, reproductive performance, learning ability and behav- ior in the rat offspring, J. Toxicol. Sci., 7 (1982) 93-110.
35 H~irfstrand, A., Cintra, A., Fuxe, K., Aronsson, A.M., Wik-

60
strom, A.-C., Okret, S., Gustafsson. J.-A. and Agnati, L.E, Regional differences in glycocorticoid receptor immunoreactiv- ity among neuropeptide Y immunoreactive neurons in the rat brain, Acta Physiol. Scand,, 135 (1989) 3-9.
36 Hicks, S.P. and D'Amato, C.J., Cell migration to isocortex in the rat, Anat. Rec., 160 (1968) 619-634.
37 Hicks, S.P.. D'Amato, C.J. and Lowe, M.J., The development of mammalian nervous system. I. Malformations of the brain esOecially cerebral cortex induced in rats by radiation. II. Some mechanisms of malformation of the cortex, J. Comp. Neurol.. 113 (1959) 435-469.
38 H6kfelt, T., Fuxe, K., Johansson, O. and Ljungdahl, A.. Pharmacohistochemical evidence of the existence of dopamine nerve terminals in the limbic cortex, Eur. J. Pharmacol., 25 (1974) 108-112.
39 Huang, M. and Rorstad, O.P., Effects of vasoactive intestinal polypeptide, monoamines, prostaglandins and 2-chloroadeno- sine on adenylate cyclase in rat cerebral microvessels, J. Neurochem., 40 (1983) 719-726.
40 Johnston, M.V. and Coyle, J.T., Histological and neurochemical effects of fetal treatment with methylazoxymethanol on rat neocortex in adulthood, Brain Res., 170 (1979) 135-155.
41 Johnston, M.V. and Coyle, J.T., Cytotoxic lesions and the development of transmitter system, Trends Neurosci.. 5 (1982) 153-156.
42 Johnston, M.N., Grzanna, R. and Coyle, J.T., Methylazoxy- methanol treatment of fetal rats results in abnormally dense noradrenergic innervation of neoeortex, Science, 203 (1979) 369-371.
43 Johnston, M.V., Carman, A.B. and Coyle, J.T., Effects of fetal treatment with methylazoxymethanol acetate at various gesta- tional dates on the neurochemistry of the adult neocortex of the rat, J. Neurochem., 36 (1981) 124-128.
44 Jones, E.G., Valentino, K.L. and Fleshman, J.W. Jr., Adjust- ment of connectivity in rat neocortex after prenatal destruction of precursor cells of layer II-IV, Dev, Brain Res., 2 (1982) 425-431.
45 Jonsson, G. and Hallman, H., Effects of prenatal methylazoxy- methanol treatment on the development of central monoamine neurons, Dev. Brain Res., 2 (1982) 513-530.
46 Kadar, T., Fekete, M., Penke, B., Kovacs, K. and Telegdy, G., Effect of cholecystokinin octapeptide sulfate ester on passive avoidance behaviour and open-field activity in rats, Neurosci. Lett., Suppl. 5 (1980) 208.
47 Kasamatsu, T. and Pettigrew, J.D., Preservation of binocularity after monocular deprivation in the striate cortex of kittens treated with 6-hydroxydopamine, J. Comp. Neurol., 5 (1979) 139-162.
48 Kitayama, I., Janson, A.M., Fuxe, K., Agnati, L.E, Cintra, A., Ogren, S.-O., H~irfstrand, A., Eneroth, P., Tsutsumi, T., Jonsson, G., Steinbusch, H.W.M. and Visser, T.J., Effects of acute and chronic treatment with imipramine on 5-hydroxytryp- tamine nerve cell groups and on bulbospinal 5-hydroxytrypta- mine/substance P/thyrotropin releasing hormone immunoreac- rive neurons in the rat. A morphometric and microdensitometric analysis, J. Neural Transm., 70 (1987) 251-285.
49 K6nig, J .ER. and Klippel, R.A., The Rat Brain. A Stereotaxic Atlas of the Forebrain and Lower Parts of the Brain Stem, Williams and Wilkins, Baltimore, 1963.
50 Lee, T.J.-E, Saito, A. and Berezin, I., Vasoactive intestinal polypeptide-like substance: the potential transmitter for cerebral vasodilatation, Science, 224 (1984) 898-901.
51 Lidov, H.G.W. and Molliver, M.E,. The structure of cerebral cortex in the rat following prenatal administration of 6-hydroxy- dopamine, Dev. Brain Res., 3 (1982) 81-108.
52 Lindvall, O., Bj6rklund, A. and Divac, I., Organization of catecholamine neurons projecting to the frontal cortex in the rat, Brain Res., 142 (1978) 1-24.
53 Magistretti, P.J. and Morrison, J.H., VIP neurons in the neocortex, Trends Neurosci., 1 (1985) 7-8.
54 Magistretti, P.J. and Morrison, J.H., Noradrenaline and vasoac- tive intestinal peptide containing neuronal systems in neocortex: functional convergence with contrasting morphology, Neurosci- ence, 24 (1988) 367-378.
55 Magistretti, P.J. and Schorderet, M., VIP and noradrenaline act synergistically to increase cyclic AMP in the cerebral cortex, Nature (Lond.), 308 (1984) 280-282.
56 Magistretti, P.J., Morrison, J.H., Shoemaker, W.J., Sapin, V. and Bloom, F.E., Vasoactive intestinal polypeptide induces glycogenolysis in mouse cortical slices: a possible regulatory mechanism for the local control of energy metabolism, Proc. Natl. Acad. Sci. U.S.A., 78 (1981) 6535-6539.
57 Markey, K., Kondo, A., Shenkman, L. and Goldstein, M., Purification and characterization of tyrosine hydroxylase from clonal chromocytoma cell line, Mol. Pharmacol., 17 (1980) 79-85.
58 Matsumoto, H. and Higa, H.M., Studies on methylazoxymeth- anol, the aglycone of cycasin: methylation of nucleic acid in vitro, Biochem. J., 98 (1966) 20c.
59 McDonald, J.K., Parnavelas, J.G., Karamanlidis, A., Brecha, N. and Koenig, J.I., The morphology and distribution of peptide-containing neurons in the adult and developing visual cortex of the rat. I. Somatostatin, J. Neurocytol., 11 (1982) 809-824.
60 McDonald, J.K., Parnavelas, J.G., Karamantidis, A., Brecha, N. and Koenig, J.I., The morphology and distribution of peptide-containing neurons in the adult and developing visual cortex of the rat. II. Vasoactive intestinal polypeptide, J. Neurocytol., 11 (1982) 825-837.
61 Mohammed, A.K., Jonsson, G., Sundstrom, E., Minor, B.G., Soderberg, U. and Archer, T., Selective attention and place navigation in rats treated prenatally with methylazoxymethanol, Dev. Brain Res., 30 (1986) 145-155.
62 Mohammed, A.K., Jonsson, G. and Archer, T , Selective lesioning of forebrain noradrenaline neurons at birth abolishes the improved maze learning performance induced by rearing in complex environment, Brain Res., 398 (1986) 6-10.
63 Morrison, J.H., Magistretti, P.J., Benoit, R. and Bloom, EE., The distribution and morphological characteristics of the intra- cortical VIP-positive cell: an immunohistochemical analysis, Brain Res., 292 (1984) 269-282.
64 Mountcastle, V.B., An organizing principle for cerebral func- tion: the unit module and the distributed system. In G.M. Edelman and V.B. Mountcastle (Eds.), The Mindful Brain, MIT Press, Cambridge, MA, 1982, pp. 7-50.
65 Olson, L. and Seiger, A., Early prenatal ontogeny of central monoamine neurons in the rat: fluorescence histochemieal observations, Z. Anat. Entwickl.-Gesch., 137 (1972) 301-316.
66 Rabe, A. and Haddad, R.K., Methylazoxymethanol induced microencephaly in rats: behavioral studies, Fed. Proc., 31 (1972) 1536-1539.
67 Rodier, P.M., Correlations between prenatally-induced alter- ations in CNS cell populations and postnatal function, Teratol- ogy, 16 (19771 235-246.
68 Singh, S.C., Deformed dendrites and reduced spine number on ectopic neurons in the hippocampus of rats exposed to methyl- azoxymethanol acetate. A Golgi-Cox study, Acta Neuropathol., 49 (1980) 193-198.
69 Spatz, M. and Laquer, G.L., Transplacental induction of microencephaly in two strains of rats, Proc. Soc. Exp. Biol. Med., 129 (1986) 705-710.
70 Sternherger L.A., lmmunocytochemistry, 2nd edn., Wiley, New York, 1979.
71 Sternberger, L.A. and Sternberger, N.H., The unlabeled anti- body method: comparison of peroxidase-antiperoxidase with avidin-biotin complex by a new method of quantification, J. Histochem. Cytochem., 34 (19861 509-605.
72 Thomas, E. and Pearse, A.G.E., The fine localization of dehydrogenases in the nervous system, Histochemistry, 2 (19611 266-282.

73 Thomas, E. and Pearse, A.G.E., The solitary active cells. Histochemical demonstration of damage-resistant nerve cells with a TPN-Diaphorase reaction, Acta Neuropathol., 3 (1964) 238-249.
74 Tohyama, M. and Shiotani, Y., Development of the peptidergic systems in the central and peripheral nervous system. In P. Panula, H. Paivarinta and S. Soinila (Eds.) Neurohistochernistry: Modern Methods and Applications, Liss, New York, 1986, pp. 295-328.
75 Vanderhaeghen. J-J., Lotstra, F., De Mey, J. and Gilles, C., Immunohistochemical localization of cholecystokinin and gas- trin-like peptides in the brain and hypophysis of the rat, Proc. Natl. Acad. Sci. U.S.A., 77 (1980) 1190-1194.
76 Vincent, S.R., Neuropeptide coexistence in mammalian fore- brain. In V.Chan-Palay and S.L. Palay (Eds.) Coexistence of Neuroactive Substances in Neurons, Wiley, New York, 1984, pp. 127-135.
77 Vincent, S.R., NADPH-diaphorase histochemistry and neuro- transmitter coexistence. In P. Panula, H. Paivarinta and S. Soinila (Eds.), Neurohistochemistry: Modern Methods and Ap- plications, Liss, New York, 1986, pp. 375-396.
78 Vincent, S.R. and Johansson, O., Striatal neurons containing both somatostatin- and avian pancreatic polypeptide (APP)-like
61
immunoreactivities and NADPH-diaphorase activity: A light and electron microscopy study, J. Comp. Neurol., 217 (1983) 252-263.
79 Vincent, S.R., Skirboll, L., H6kfelt, T., Johansson, O., Lund- berg, J.M., Elde, R.P., Terenius, L. and Kimmel, J., Coexist- ence of somatostatin- and avian pancreatic polypeptide (APP)- like immunoreactivity in some forebrain neurons, Neuroscience, 7 (1982) 439-446.
80 Wei, E.P., Kontos, H.A. and Said, S.I., Mechanism of action of vasoactive intestinal polypeptide on cerebral arterioles, Am. J. Physiol., 239 (1980) H765-H768.
81 Woodhams, P.L., Allen, Y.S., McGovern, J., Alien, J.M., Bloom, S.R., Balasz, R. and Polak, J.M., Immunohistochemical analysis of the early ontogeny of the neuropeptide Y system in the rat brain, Neuroscience, 15 (1985) 173-302.
82 Yasuda, K., Pharmacological assessment of functional alter- ations in the brain of rats exposed prenatally to methylazoxy- methanol, Nippon Yakurigaku Zasshi, 89 (1987) 203-221.
83 Zoli. M., Agnati, L.F.. Fuxe, K., Cintra, A.. Grimaldi, R., Vanderhaeghen, J.-J., Eneroth, P, and Goldstein, M., Effects of chronic uridine treatment on regional neuropeptide and tyrosine hydroxylase like immunoreactivities in the brain of 12 month-old male rats, Neurochem. Int., 13 (1988) 499-508.
Copyright © 2022 FDOKUMEN




















