
Feeding and drinking responses to neuropeptide Y injections in the paraventricular hypothalamic...
7
Brain Research, 575 (1992) 265-271 265 t~) 1992 Elsevier Science Publishers B.V. All rights reserved. 0006-8993/92/$05.00 BRES 17550 Feeding and drinking responses to neuropeptide Y injections in the paraventricular hypothalamic nucleus of aged rats Emilio Merlo Pich 1'2, Beatrice Messori 1, Michele Zoli 1, Francesco Ferraguti 1, Paolo Marrama 2, Giuseppe Biagini 1, Kjell Fuxe 3 and Luigi F. Agnati 1 1Institute of Human Physiology, and 2Department of Endocrinology, University of Modena (Italy) and 3Department of Histology and Neurobiology, Karolinska Institutet, Stockholm (Sweden) (Accepted 5 November 1991) Key words: Neuropeptide Y; Feeding; Drinking behavior; Paraventricular hypothalamic nucleus; Aging; Rat Neuropeptide Y (NPY), a peptide of the pancreatic polypeptide family, exerts a potent stimulatory action on eating when injected into the paraventricular hypothalamic nucleus (PVN) in rats. Several NPY-containing systems are altered with advancing age, and aged rodents de- velop anorexia and a modified daily cycle pattern of feeding. These findings suggest that a relationship may exist between the aging-related anorexia and the reduced function of NPY-containing systems projecting to the PVN. In the present study eating and drinking behavior in satiated or fasted young (3 months) and aged (24 months) rats have been investigated over 22 h after NPY injection into the PVN. The levels of NPY immunoreactivity (IR) in PVN were also evaluated by means of semiquantitative immunocytochemistry. NPY injections into PVN increased food and water consumption in both young and aged satiated rats 30, 90 and 240 min after injection. However, the feeding and drinking responses elicited by 0.05, 0.10 and 1.0 nmol of NPY were significantly attenuated in the aged rats when compared to young rats. In aged rats, 24 h of food and water deprivation produced significant increase of food consumption measured at 30, 90 min and 22 h, which was equivalent to that induced by 1.0 nmol NPY injection. Administration of 1.0 nmol NPY in PVN did not further increase the 24 h dep- rivation effect on feeding in both groups of rats, but enhanced drinking in deprived young rats. This effect was not present in aged rats. In addition, aged rats showed a stronger response to 24 h deprivation than to 1.0 nmol NPY administration. These results and the severe re- duction of NPY IR levels in PVN nerve terminals of aged rats suggest that NPY deficiency may be a factor responsible for anorexia in the aged. INTRODUCTION Converging experiments suggest that neuropeptide Y (NPY), a 36-amino acid member of the pancreatic poly- peptide family 34, is involved as a chemical signal in brain control mechanisms of eating behavior. Injections into the third ventricle or in different brain nuclei elicit a marked increase of food and water consumption in sati- ated rodents 1°'17'3°'31. Among these nuclei, the most re- sponsive is the paraventricular hypothalamic nucleus (PVN) 19'30, an area characterized by a high density of NPY-containing nerve terminals 9'12'2s and known to be involved in eating and drinking control 16, and in modu- lating energy substrate utilization 18'31. Moreover, NPY concentrations in PVN have been demonstrated to in- crease in response to severe food deprivation in adult rats 723. High concentrations of NPY were also found in the PVN of the hyperphagic obese Zucker falfa rat strain 6. In contrast, low NPY concentrations 24 and re- duced NPY immunoreactivity (IR) 35 were demonstrated in PVN of aged rats. Aged rodents develop anorexia and a modified light/dark daily cycle pattern of feeding 2°'2~. Interestingly, a decline in behavioral responses mediated by a series of brain peptides, in particular opioid pep- tides, occurs in rodents with advancing age 2'21. For ex- ample, aged rats seem to be less sensitive to the enhanc- ing effects of morphine on food consumption 14 and analgesia 11A5. In some cases, i.e. for opioid peptides, these functional impairments have been related to a re- duced efficiency of synaptic transmission of the peptide- containing systems, involving both pre- and postsynaptic levels 3A6. Recent evidence indicates that also some NPY- containing systems are altered with advancing age. Re- duced NPY IR was found in the C1-C2 adrenergic groups 4, the locus coeruleus 1 and the nucleus arcuatus (Agnati, unpublished results) of aged rats, systems which project to the PVN 5'8'23. These findings suggest that a relationship may exist between the aging-related anorexia and the reduced function of NPY-containing systems projecting to the PVN. Accordingly, a lower rate of NPY-induced food intake was described in aged mice when compared with young mice 21. The aim of the present work was to study NPY-elic- ited feeding and drinking responses in the PVN of young Correspondence: L.F. Agnati, Institute of Human Physiology, University of Modena, via Campi 287, 41100 Modena, Italy.
Transcript of Feeding and drinking responses to neuropeptide Y injections in the paraventricular hypothalamic...

Brain Research, 575 (1992) 265-271 265 t~) 1992 Elsevier Science Publishers B.V. All rights reserved. 0006-8993/92/$05.00
BRES 17550
Feeding and drinking responses to neuropeptide Y injections in the paraventricular hypothalamic nucleus of aged rats
Emilio Merlo Pich 1'2, Beatrice Messori 1, Michele Zoli 1, Francesco Ferraguti 1, Paolo Marrama 2, Giuseppe Biagini 1, Kjell Fuxe 3 and Luigi F. Agnati 1
1Institute of Human Physiology, and 2Department of Endocrinology, University of Modena (Italy) and 3Department of Histology and Neurobiology, Karolinska Institutet, Stockholm (Sweden)
(Accepted 5 November 1991)
Key words: Neuropeptide Y; Feeding; Drinking behavior; Paraventricular hypothalamic nucleus; Aging; Rat
Neuropeptide Y (NPY), a peptide of the pancreatic polypeptide family, exerts a potent stimulatory action on eating when injected into the paraventricular hypothalamic nucleus (PVN) in rats. Several NPY-containing systems are altered with advancing age, and aged rodents de- velop anorexia and a modified daily cycle pattern of feeding. These findings suggest that a relationship may exist between the aging-related anorexia and the reduced function of NPY-containing systems projecting to the PVN. In the present study eating and drinking behavior in satiated or fasted young (3 months) and aged (24 months) rats have been investigated over 22 h after NPY injection into the PVN. The levels of NPY immunoreactivity (IR) in PVN were also evaluated by means of semiquantitative immunocytochemistry. NPY injections into PVN increased food and water consumption in both young and aged satiated rats 30, 90 and 240 min after injection. However, the feeding and drinking responses elicited by 0.05, 0.10 and 1.0 nmol of NPY were significantly attenuated in the aged rats when compared to young rats. In aged rats, 24 h of food and water deprivation produced significant increase of food consumption measured at 30, 90 min and 22 h, which was equivalent to that induced by 1.0 nmol NPY injection. Administration of 1.0 nmol NPY in PVN did not further increase the 24 h dep- rivation effect on feeding in both groups of rats, but enhanced drinking in deprived young rats. This effect was not present in aged rats. In addition, aged rats showed a stronger response to 24 h deprivation than to 1.0 nmol NPY administration. These results and the severe re- duction of NPY IR levels in PVN nerve terminals of aged rats suggest that NPY deficiency may be a factor responsible for anorexia in the aged.
INTRODUCTION
Converging exper iments suggest that neuropept ide Y
(NPY), a 36-amino acid m e m b e r of the pancreat ic poly-
pept ide family 34, is involved as a chemical signal in bra in
control mechanisms of eat ing behavior . Injections into
the third ventricle or in different bra in nuclei elicit a
marked increase of food and water consumpt ion in sati- a ted rodents 1°'17'3°'31. A m o n g these nuclei, the most re-
sponsive is the paraventr icular hypothalamic nucleus
(PVN) 19'30, an area character ized by a high density of
NPY-containing nerve terminals 9'12'2s and known to be
involved in eat ing and dr inking control 16, and in modu-
lating energy substrate uti l ization 18'31. Moreover , NPY
concentrat ions in PVN have been demons t ra ted to in-
crease in response to severe food depr ivat ion in adult
rats 723. High concentrat ions of NPY were also found in
the PVN of the hyperphagic obese Zucker falfa rat strain 6. In contrast , low NPY concentrat ions 24 and re-
duced NPY immunoreact iv i ty ( IR) 35 were demons t ra ted
in PVN of aged rats. A g e d rodents develop anorexia and a modified l ight/dark daily cycle pa t te rn of feeding 2°'2~.
Interest ingly, a decline in behavioral responses media ted
by a series of bra in pept ides , in par t icular opioid pep-
t ides, occurs in rodents with advancing age 2'21. For ex-
ample , aged rats seem to be less sensitive to the enhanc-
ing effects of morphine on food consumption 14 and
analgesia 11A5. In some cases, i .e. for opioid pept ides ,
these functional impairments have been re la ted to a re-
duced efficiency of synaptic transmission of the pept ide-
containing systems, involving both pre- and postsynaptic
levels 3A6. Recent evidence indicates that also some NPY-
containing systems are al tered with advancing age. Re-
duced NPY I R was found in the C1-C2 adrenergic
groups 4, the locus coeruleus 1 and the nucleus arcuatus
(Agnat i , unpubl ished results) of aged rats, systems which project to the PVN 5'8'23. These findings suggest that a
relat ionship may exist be tween the aging-related anorexia
and the reduced function of NPY-containing systems
project ing to the PVN. Accordingly, a lower rate of
NPY-induced food intake was descr ibed in aged mice when compared with young mice 21.
The aim of the present work was to study NPY-elic-
i ted feeding and drinking responses in the PVN of young
Correspondence: L.F. Agnati, Institute of Human Physiology, University of Modena, via Campi 287, 41100 Modena, Italy.

266
a n d aged rats . T h e levels of N P Y I R were also m e a s u r e d
in P V N of a g r o u p of y o u n g a n d aged ra t s by m e a n s of
s e m i q u a n t i t a t i v e i m m u n o c y t o c h e m i s t r y 4'37.
MATERIALS AND METHODS
Animals Male rats (CD, Sprague-Dawley) were purchased from Charles
River Breeding Labs (Calco, Italy). They were maintained at stan- dard temperature (22°C) and lighting conditions (light on 08.00- 20.00 h) with free access to water and food (MIL, S. Polo d'Enza, Italy). Two groups of rats were used; young rats (3-month-old) and aged rats (24-month-old). Mean body weights (b.wt. + S.E.M.) at the time of the experiment were 364.4 + 5.3 g and 593.4 + 22.6 g, for young and aged rats, respectively. The rats were stereotaxically implanted under ketamine anesthesia (100 mg/kg i.p.) with a uni- lateral chronic 22-gauge stainless-steel guide cannula 1.5 mm dor- sal to the PVN according to the following coordinates: 7.2 mm an- terior and 2.0 mm above the interaural line, and 0.4 mm lateral to the midline 22. This guide-cannula was secured to the skull using stainless-steel screws and dental acrylic cement. The rats were han- dled during a one week recovery period in order to accustom them to the experimental setting.
Experiment 1 After surgery the two groups of rats (n = 12 per group),
3-month-old and 24-month-old, were housed four per cage and maintained on a standard diet. Two days before testing the rats were placed in a clear plexiglass cage with a grid floor in which a weighed amount of dry food (MIL, water content < 13.0%) was placed in a Petri dish. A valved bottle (Tecniplast, Gazzada, Italy) was used to prevent accidental water leakage and to mea- sure water intake. Daily food and water consumption was recorded twice before testing. To ensure that the rats were satiated, they were given a freshly prepared diet consisting of standard pellets, previously dipped into milk, one hour before testing. The rats were then removed from the cages and injected with NPY at various doses (0.01, 0.05, 0.10 and 1.00 nmol dissolved in 0.5/A saline) or an equivalent volume of saline in a counterbalanced order. Injec- tions into PVN were performed using a 26-gauge stainless-steel in- jection cannula connected via a PE-10 tubing to a Hamilton syringe mounted on a high precision infusion pump (Carnegie Medicin, Solna, Sweden). The injection cannula extended 1.5 mm beyond the guide tip. The infusion time was 2 min. Injection cannula was left in situ 3 min after the end of infusion. Then the rats were re- turned to their cage with weighed food and water.
Food consumption was measured by subtracting residual pellets and spillage from initial weight values, and water consumption was assessed by weighing the bottle. Both measurements were per- formed at 30, 90, 240 min and 22 h (1320 min) postinjection. Re- suits were expressed as cumulative values (+ 0.1 g). Testing began at 14.00-14.30 h. Each rat was injected for a maximum of 4-5 times, with an interval between two subsequent injections of at least 48 h. Each rat received at least one saline injection.
Experiment 2 Two groups of rats (n = 8 per group), 3-month-old and 24-
month-old, were used. The rats were injected with 1.0 nmol NPY or saline when satiated, as described in the previous experiment. The same rats were then deprived of food and water for 24 h and tested for food and water consumption after injection of 1.0 nmol NPY or saline, as described above. Food and water consumption was measured 30, 90 min and 22 h (1320 min) postinjection (+ 0.1 g). Injections and deprivation were carried out in a counterbal- anced order.
Histology and immunocytochemistry Within one week after the end of testing, the rats were sacrificed
by transcardiac perfusion with 100 ml of warm saline followed by
100 ml of ice-cold 4% paraformaldehyde +0.3% picric acid in 0.1 M phosphate buffer. The brains were rapidly dissected out, post- fixed overnight in the same fixative and frozen for cryotome cut- ting. Coronal sections (30 ~m) were stained with Cresyl violet and the location of the cannula tips was determined in the microscope with the aid of a stereotaxic atlas.
Young and aged naive rats (n = 4 to 5 rats per group) were sac- rificed (as described above) for NPY immunocytochemistry. Fixed brains were cut using a vibratome (50/~m coronal sections). The sections were then processed for immunocytochemistry using the indirect immunoperoxidase procedure as described elsewhere 4. The sections were incubated free-floating with a rabbit NPY antiserum (Amersham Laboratories, Buckinghamshire, U.K.) diluted 1:500. For control experiments NPY antiserum preabsorbed with NPY (50 #g/ml) was used. In order to improve the linearity of the antigen content/staining intensity relationship, diaminobenzidine and H20 2 were used in optimal concentrations and the antibody dilution was far from saturation 8'33.
Image analysis The semiquantitative evaluation of the intensity of immunostain-
ing was performed using an automatic image analyzer (IBAS I-II, Zeiss Kontron, Munich, E R.G.) as described elsewhere 4"36. Briefly, the sampled fields were selected and the specifically stained pro- files were interactively discriminated from the non-specific staining. The sampled field was the entire parvocellular component of .the PVN. Various rostrocaudal levels were measured from A 5800 to A 5050 (K6nig and Klippel, 1963). The field area, i.e. the area covered by specific profiles, and the mean grey tone value of the specicific profiles (X) were measured. The total immunoreactivity (TIR) was estimated by the product FA x X.
Statistics The data of the first experiment were analyzed by a 3-way anal-
ysis of variance for repeated measures, with 'age' as between fac- tor, 'dose of NPY' as first within factor and 'time after injection' as second within factor. In the second experiment the between factor was 'age', the first within factor was 'dose of NPY' and the second within factor was 'deprivation'. This analysis was performed enter- ing the data obtained at each time interval after the injection. Post- hoc pair-wise comparisons were carried out using Newman-Keuls' or Tukey's tests. Non-parametric tests and Bonferroni's correction for multiple comparison were used to compare image analysis mea- surements obtained in PVN of young and aged rats.
RESULTS
T h e h i s to log ica l a s s e s s m e n t of b r a i n s in b o t h exper i -
m e n t s i nd i ca t e t h a t t he m a j o r i t y of the c a n n u l a t rac t s
t e r m i n a t e d in the vic ini ty of t he P V N . W h e n t he t rac t s
we re f o u n d a b o u t 0.3 m m a n t e r i o r , p o s t e r i o r or dor sa l
to P V N , the a n i m a l s we re no t i n c l u d e d in t he s ta t is t ica l
analys is of f ood a n d w a t e r c o n s u m p t i o n da ta . Fig. 1 il-
l u s t r a t e s the l oca t i on of un i l a t e r a l i n j e c t i o n si tes in y o u n g
a n d aged ra ts s e l ec t ed for f o o d a n d w a t e r m e a s u r e m e n t
analys is of t he first e x p e r i m e n t .
Experiment 1
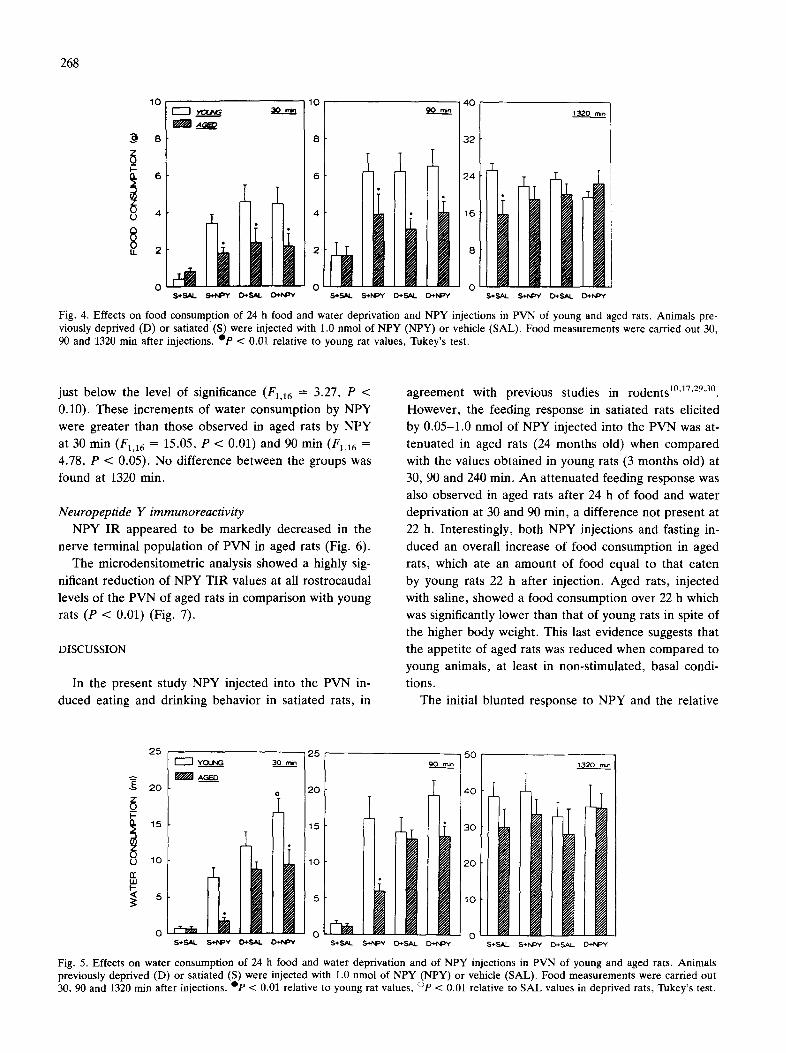
T h e resu l t s of the first e x p e r i m e n t a re s h o w n in Figs.
2 a n d 3. I n j e c t i o n s of N P Y in to t he P V N p r o d u c e d an
inc rease in f o o d c o n s u m p t i o n in b o t h y o u n g a n d aged
ra t s (F4,48 = 15.61, P < 0.01) , t he lowes t e f fec t ive dose
b e i n g 0.05 nmol . H o w e v e r , aged ra t s c o n s u m e d less food
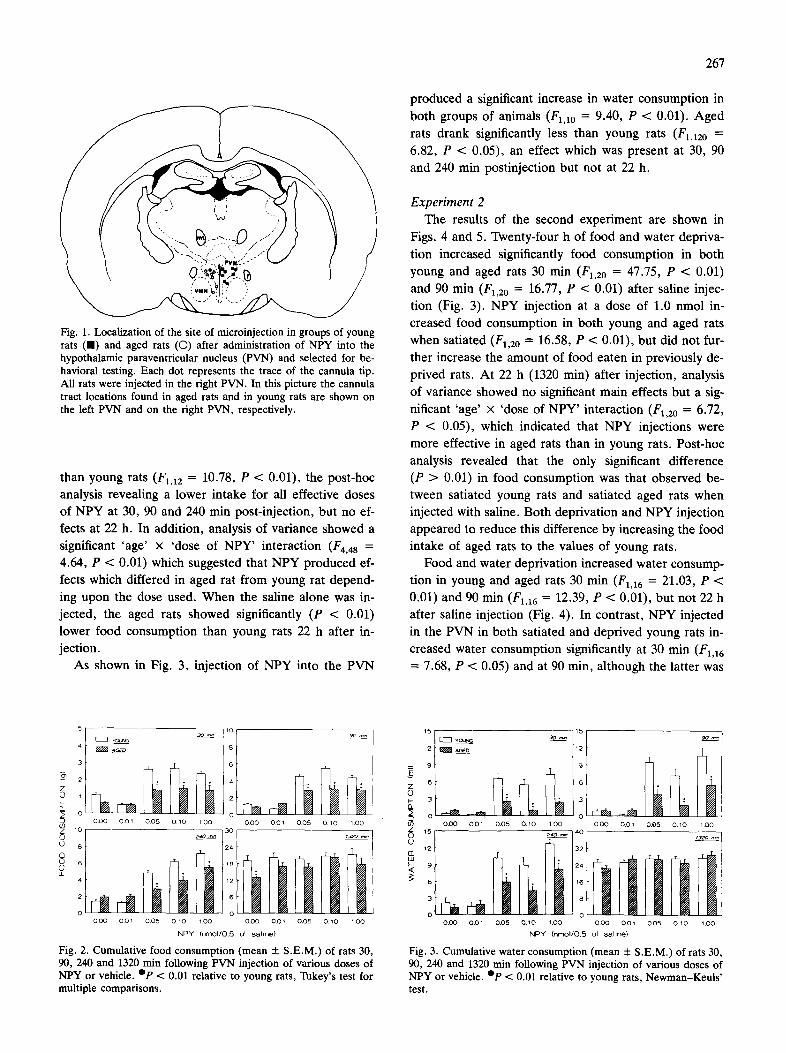
Fig. 1. Localization of the site of microinjection in groups of young rats (l l) and aged rats (O) after administration of NPY into the hypothalamic paraventricular nucleus (PVN) and selected for be- havioral testing. Each dot represents the trace of the cannula tip. All rats were injected in the fight PVN. In this picture the cannula tract locations found in aged rats and in young rats are shown on the left PVN and on the right PVN, respectively.
than young rats (F1,12 = 10.78, P < 0.01), the post-hoe analysis revealing a lower intake for all effective doses of NPY at 30, 90 and 240 min post-injection, but no ef- fects at 22 h. In addition, analysis of variance showed a significant 'age' x 'dose of NPY' interaction (/74,48 = 4.64, P < 0.01) which suggested that NPY produced ef- fects which differed in aged rat from young rat depend- ing upon the dose used. When the saline alone was in- jected, the aged rats showed significantly (P < 0.01) lower food consumption than young rats 22 h after in- jection.
As shown in Fig. 3, injection of NPY into the PVN
267
produced a significant increase in water consumption in both groups of animals (Fl,10 = 9.40, P < 0.01). Aged rats drank significantly less than young rats (F1,120 = 6.82, P < 0.05), an effect which was present at 30, 90 and 240 min postinjection but not at 22 h.
Experiment 2 The results of the second experiment are shown in
Figs. 4 and 5. Twenty-four h of food and water depriva- tion increased significantly food consumption in both young and aged rats 30 min (F1,20 = 47.75, P < 0.01) and 90 min (F1,20 = 16.77, P < 0.01) after saline injec- tion (Fig. 3). NPY injection at a dose of 1.0 nmol in- creased food consumption in both young and aged rats when satiated (F1,20 = 16.58, P < 0.01), but did not fur- ther increase the amount of food eaten in previously de- prived rats. At 22 h (1320 min) after injection, analysis of variance showed no significant main effects but a sig- nificant 'age' x 'dose of NPY' interaction (/71,20 = 6.72, P < 0.05), which indicated that NPY injections were more effective in aged rats than in young rats. Post-hoe analysis revealed that the only significant difference (P > 0.01) in food consumption was that observed be- tween satiated young rats and satiated aged rats when injected with saline. Both deprivation and NPY injection appeared to reduce this difference by increasing the food intake of aged rats to the values of young rats.
Food and water deprivation increased water consump- tion in young and aged rats 30 min (FL16 = 21.03, P < 0.01) and 90 min (F1,16 = 12.39, P < 0.01), but not 22 h after saline injection (Fig. 4). In contrast, NPY injected in the PVN in both satiated and deprived young rats in- creased water consumption significantly at 30 min (F~,16 = 7.68, P < 0.05) and at 90 min, although the latter was
0.00 001 0.05 0 1 0 1.00 0 0 0 001 0.05 0.10 1 .O0
0 _240 ~ iJ2o
{J 8 24
0 ~fl
0(30 0 0 1 0 0 5 0 1 0 1 0 0 0.00 0.01 0 .05 0 .10 1.00
N P Y (n rno l /0 .5 ul sahne }
Fig. 2. Cumulative food consumption (mean + S.E.M.) of rats 30, 90, 240 and 1320 min following PVN injection of various doses of NPY or vehicle, e p < 0.01 relative to young rats, Tukey's test for multiple comparisons.
Z _o
S r r
<
0.05 0.10 1.00 0 0 0 0.01 0.~)5 0.10 100 0.00 0.01
0.00 0.01 0.05 0.10 1.00 0,00 0.01 0.05 O. 10 1.00
N P Y (nmo l /O .5 ul sa l ine )
Fig. 3. Cumulative water consumption (mean + S.E.M.) of rats 30, 90, 240 and 1320 rain following PVN injection of various doses of NPY or vehicle. Op < 0.01 relative to young rats, Newman-Keuls' test.
I-
o
DISCUSSION
lO
8
6
4
2
268
i
i
lO
o i 9 0 rain
i'l .J_
~4
8
o
1:320 rn!n
ii Fig. 4. Effects on food consumption of 24 h food and water deprivation and NPY injections in PVN of young and aged rats. Animals pre- viously deprived (D) or satiated (S) were injected with 1.0 nmol of NPY (NPY) or vehicle (SAL). Food measurements were carried out 30, 90 and • 1320 min after injections. P < 0.01 relative to young rat values, Tukey's test.
just below the level of significance (/71,16 -~- 3.27, P < 0.10). These increments of water consumption by NPY were greater than those observed in aged rats by NPY
at 30 min (El ,16 = 1 5 . 0 5 , P < 0.01) and 90 min (FI j 6 = 4.78, P < 0.05). No difference between the groups was found at 1320 min.
Neuropeptide Y immunoreactivity NPY IR appeared to be markedly decreased in the
nerve terminal population of PVN in aged rats (Fig. 6). The microdensitometric analysis showed a highly sig-
nificant reduction of NPY TIR values at all rostrocaudal levels of the PVN of aged rats in comparison with young rats (P < 0.01) (Fig. 7).
25
In the present study NPY injected into the PVN in- duced eating and drinking behavior in satiated rats, in
agreement with previous studies in r o d e n t s 1°'17'29'3°.
However, the feeding response in satiated rats elicited by 0.05-1.0 nmol of NPY injected into the PVN was at- tenuated in aged rats (24 months old) when compared with the values obtained in young rats (3 months old) at 30, 90 and 240 min. An attenuated feeding response was also observed in aged rats after 24 h of food and water deprivation at 30 and 90 min, a difference not present at 22 h. Interestingly, both NPY injections and fasting in- duced an overall increase of food consumption in aged rats, which ate an amount of food equal to that eaten by young rats 22 h after injection. Aged rats, injected with saline, showed a food consumption over 22 h which was significantly lower than that of young rats in spite of the higher body weight. This last evidence suggests that the appetite of aged rats was reduced when compared to young animals, at least in non-stimulated, basal condi- tions.
The initial blunted response to NPY and the relative
S+S~-
I--
o n ~ w I - < 3:
I - ' - 7 Y<:~NG
AGEO 20
15
i o ~ S+,SAL S+NPY D+SAL
3 0 rain
2
$+NPY D+S~4-
9 0 rmn
1
S+NPY C;~,SAL I~-NPy
50
4 0
30
20
10
0 S÷SAL
1320 mm
)÷NPY
Fig. 5. Effects on water consumption of 24 h food and water deprivation and of NPY injections in PVN of young and aged rats. Animals previously deprived (D) or satiated (S) were injected with 1.0 nmol of NPY (NPY) or vehicle (SAL). Food measurements were carried out 30, 90 and 1320 min after injections. Op < 0.01 relative to young rat values, ©P < 0.01 relative to SAL values in deprived rats, "Ihkey's test.
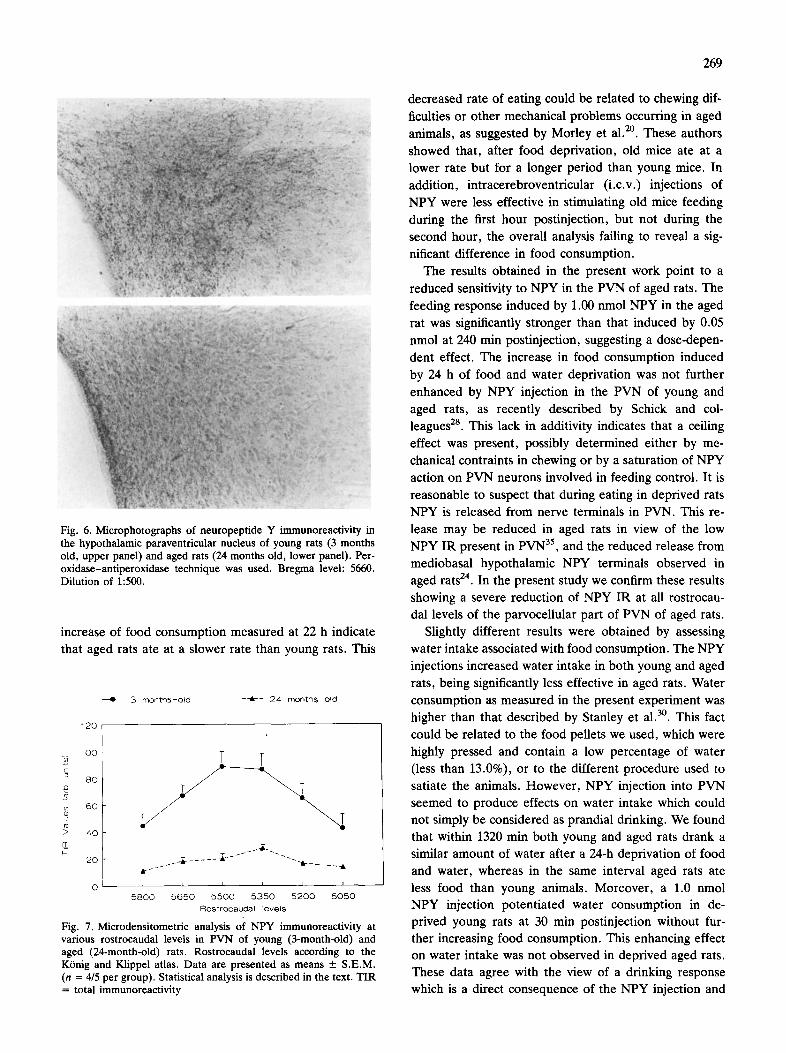
Fig. 6. Microphotographs of neuropeptide Y immunoreactivity in the hypothalamic paraventricular nucleus of young rats (3 months old, upper panel) and aged rats (24 months old, lower panel). Per- oxidase-antiperoxidase technique was used. Bregma level: 5660. Dilution of 1:500.
increase of food consumption measured at 22 h indicate that aged rats ate at a slower rate than young rats. This
• 3 monthS-O+Cl
12O
--A-- 2 4 m o n t h s - o l d
1 O0
8 0 ~5
~ 60
~ 4o _co F-
2 0
0 h I ~ t I f
5 8 0 0 5 6 5 0 5 5 0 0 5 3 5 0 5 2 0 0 5 0 5 0
FqostrocauOal levels
Fig. 7, Microdensitometric analysis of NPY immunoreactivity at various rostrocaudal levels in PVN of young (3-month-old) and aged (24-month-old) rats. Rostrocaudal levels according to the K6nig and Klippel atlas. Data are presented as means + S.E.M. (n = 4/5 per group). Statistical analysis is described in the text. TIR = total immunoreactivity
269
decreased rate of eating could be related to chewing dif-
ficulties or other mechanical problems occurring in aged animals, as suggested by Morley et al. 2°. These authors
showed that, after food deprivation, old mice ate at a lower rate but for a longer period than young mice. In addition, intracerebroventricular (i.c.v.) injections of NPY were less effective in stimulating old mice feeding during the first hour postinjection, but not during the second hour, the overall analysis failing to reveal a sig- nificant difference in food consumption.
The results obtained in the present work point to a
reduced sensitivity to NPY in the PVN of aged rats. The feeding response induced by 1.00 nmol NPY in the aged rat was significantly stronger than that induced by 0.05 nmol at 240 min postinjection, suggesting a dose-depen- dent effect. The increase in food consumption induced by 24 h of food and water deprivation was not further enhanced by NPY injection in the PVN of young and aged rats, as recently described by Schick and col- leagues 28. This lack in additivity indicates that a ceiling
effect was present, possibly determined either by me- chanical contraints in chewing or by a saturation of NPY action on PVN neurons involved in feeding control. It is reasonable to suspect that during eating in deprived rats NPY is released from nerve terminals in PVN. This re- lease may be reduced in aged rats in view of the low NPY IR present in PVN 35, and the reduced release from
mediobasal hypothalamic NPY terminals observed in aged rats 24. In the present study we confirm these results
showing a severe reduction of NPY IR at all rostrocau- dal levels of the parvoceUular part of PVN of aged rats.
Slightly different results were obtained by assessing water intake associated with food consumption. The NPY injections increased water intake in both young and aged rats, being significantly less effective in aged rats. Water consumption as measured in the present experiment was higher than that described by Stanley et al. 3°. This fact
could be related to the food pellets we used, which were highly pressed and contain a low percentage of water (less than 13.0%), or to the different procedure used to satiate the animals. However, NPY injection into PVN seemed to produce effects on water intake which could not simply be considered as prandial drinking. We found that within 1320 min both young and aged rats drank a similar amount of water after a 24-h deprivation of food and water, whereas in the same interval aged rats ate less food than young animals. Moreover, a 1.0 nmol NPY injection potentiated water consumption in de- prived young rats at 30 min postinjection without fur- ther increasing food consumption. This enhancing effect on water intake was not observed in deprived aged rats. These data agree with the view of a drinking response which is a direct consequence of the NPY injection and

270
not completely feeding-dependent w'28-3°. A similar dis-
sociation has also been described after i.c.v, administra-
tion of noradrenaline (NA) and bombesin, substances
which affect feeding but not drinking 2°. In addition, the
volume of water drunk by deprived aged rats 90 min af-
ter injection was significantly greater than the volume of
water drunk by aged rats injected with NPY. This result
suggests that the low water intake observed in aged rats
was not likely to be related to oral mechanical impair-
ments, but to a reduced effect of NPY on PVN neurons
which control drinking behavior. Consequently, we are
tempted to suggest that a diminished secretory function of NPY-containing neurons projecting to the PVN of
aged rats, which may involve reduced synthesis, process-
ing and storage of the peptide, may contribute to the impaired water balance reported in senescence.
All these data are in line with the view that the ca- pacity of NPY neurons to sustain high levels of secre- tory functions gradually decrease with advancing age 3'4'
2t,24. Interestingly, a reduction of NPY IR was observed
in the C1 group of aged rats, whereas the co-stored phe-
nylethanolamine-N-methyltransferase (PNMT) IR, the enzyme involved in the conversion of N A into adrena-
line (A), did not change when compared to the values
obtained from young rats, suggesting that C1 neurons have a reduced ability to synthesize and/or process NPY but not PNMT 4. Reduced NPY IR was also observed in
arcuate nucleus of aged rats (Agnati, unpublished data), a nucleus which projects to PVN and whose NPY levels are affected by changes of the feeding status 7. In addi-
tion, a decreased density and impaired regulation of
adrenergic a2-receptors have been described in various brain regions of the aged rats 13'27'37. These findings are
of interest since the eating response induced by NA in- jection in PVN in satiated rats is elicited through activa-
tion of a2-receptor16,and NPY-enhancing effect on feed-
ing is partially blocked by pretreatment with a2-receptor antagonists in young adult rats 29. Thus, the reduced ac-
tion of NPY in PVN with regard to feeding may also be
related to a reduced interaction with the a2-adrenergic receptor 3.
Alternatively, a decreased support of peripheral hor- mones in aging may be of relevance in determining the
decrease in the secretory and biosynthetic activity of NPY-containing neurons. Adrenal steroids have been found to interact with the hypothalamic NPY systems 13.
In rats NPY-elicited eating responses are corticosterone- dependent 32. In aged rats no reduction of nuclear gluco-
corticoid receptors are detected in PVN 35, suggesting
that corticosterone is still capable of activating glucocor- ticoid receptors in neurons which mediate NPY feeding
responses. However, in aged rats NPY-containing neu-
ronal systems projecting to hypothalamic nuclei did not respond to testosterone as in young rats 26, and the same
may be true for corticosterone.
In conclusion, NPY injections into PVN increase food
and water consumption in both young and aged rats and improve the low basal food consumption of aged rats
over a period of 22 h. However, in aged rats NPY in-
jections were less effective in increasing food and water
intake during the first period (30-240 min) after injec- tion. Fasting produced effects similar to those of NPY injections into PVN on food intake in both young and
aged rats, whereas in aged rats fasting effects on water intake were more intense than NPY effects. Finally, a
severe reduction of NPY IR levels in PVN nerve termi- nals was observed in aged rats using semiquantitative
immunocytochemistry. The impaired response to exoge- nous administration of NPY in PVN and the severe re- duction of NPY immunoreactivity in the same nucleus
observed in aged rats suggest that NPY deficiency in PVN may be one of the factors responsible for anorexia reported in the aged.
Acknowledgements. This work has been supported by C.N.R. (Progetto Finalizzato Invecchiamento), publication 913043 and by a grant from the Leo Osterman Foundation.
REFERENCES
1 Agnati, L.E, Fuxe, K., Calza, L., Giardino, L., Zini, I., Toffano, G., Goldstein, M., Marrama, P., Gustafsson, J.A., Yu, Z.Y., Cuello, A.C., Terenius, L., Lang, R. and Ganten, D., Morphometrical and microdensitometrical studies on monoaminergic and peptidergic neurons in the aging brain. In L.E Agnati, K. Fuxe (Eds.), Quantitative Neuroanatomy in Transmitter Research, McMillan Press, London, 1985, pp. 91- 112.
2 Agnati, L.E, Fuxe, K., Zoli, M., Zini, I., Toffano, G. and Ferraguti, F., A correlation analysis of the regional distribution of central enkephalin and/%endorphin immunoreactive termi- nals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring
transmission, Acta Physiol. Scand., 128 (1986) 201-207. 3 Agnati, L.F., Fuxe, K., Zoli, M., Merlo Pich, E., Benfenati,
F., Zini, I. and Goldstein, M., Aspects on the information han- dling by the central nervous system: focus on cotransmission in the aged rat brain. In T. H6kfelt, K. Fuxe and B. Pernow (Eds.), Coexistence of Neuronal Messengers, Progress in Brain Research, Vol. 68, Elsevier, Amsterdam, 1986, pp. 291-301.
4 Agnati, L.F., Fuxe, K., Zoli, M., Zini, I., H~rfstrand, A., Toffano, G. and Goldstein, M., Morphometrical and microden- sitometrical studies on phenylethanolamine-N-methyltransferase and neuropeptide Y immunoreactive neurons in the rostral me- dulla oblongata of the adult and old rats, Neuroscience, 26 (1988) 461-478.
5 Bai, F.L., Yamano, M., Shiotani, Y., Emson, P.C., Smith, A.D., Powell, J.E and Tahyama, M., An arcuato-paraventric- ular and dorsomedial neuropeptide Y-containing system which

lacks noradrenalin in the rat, Brain Research, 331 (1985) 172-175. 6 Beck, B., Burlet, A., Nicolas, J.-P. and Burlet, C., Hypotha-
lamic neuropeptide Y (NPY) in obese Zucker rats: implication in feeding and sexual behaviors, Physiol. Behav., 47 (1990) 449- 453.
7 Beck, B., Jhanwar-Uniyal, M., Burlet, A., Chapleur-Chateau, M., Leibowitz, S. and Burlet, C., Rapid and localized alteration of neuropeptide Y in discrete hypothalamic nuclei with feeding status, Brain Research, 528 (1990) 245-249.
8 Benno, R.H., Tucker, L.W., Joh, T.H. and Reis, D.J., Quan- titative immunocytochemistry of tyrosine hydroxylase in rat brain. I. Development of a computer-assisted method using the peroxidase antiperoxidase technique, Brain Research, 246 (1982) 225-236.
9 Chronwall, B.M., DiMaggio, D.A., Massari, V.J., Pickel, V.M., Ruggiero, D.A. and O'Donohue, T.L., The anatomy of neu- ropeptide Y-containing neurons in the rat brain, Neuroscience, 15 (1985) 1159-1181.
10 Clark, J.T., Kalra, P.S., Crowley, W.R. and Kalra, S.P., Neu- ropeptide Y and pancreatic polypeptide stimulate feeding be- havior in rats, Endocrinology, 115 (1984) 427-429.
11 Dupont, A., Savard, P., Merand, Y., Labrie, F. and Boissier, J.R., Age-related changes in central nervous system enkepha- lins and substance P, Life Sci., 29 (1981) 2317-2322.
12 Everitt, B.J., H0kfelt, T., Terenius, L., Tatemoto, K., Mutt, V. and Goldstein, M., Differential co-existence of neuropeptide Y (NPY)-like immunoreactivity with catecholamines in the central nervous system of the rat, Neuroscience, 11 (1984) 443-462.
13 Fuxe, K., Agnati, L.E, H~iffstrand, A., Eneroth, P., Cintra, A., Tinner, B., Merlo Pich, E., Aronsson, M., Bunnemann, B., Lang, R. and Ganten, D., Studies on the neurochemical mech- anisms underlying the neuroendocrine actions of neuropeptide Y. In V. Mutt, K. Fuxe, T. H0kfelt and J.M. Lundberg (Eds.), Neuropeptide Y, Raven, New York, 1989, pp. 115-136.
14 Gosnell, B.A., Levine, A.S. and Morley, J.E., The effects of aging on opioid modulation of feeding in rats, Life Sci., 32 (1983) 2793-2799.
15 Hess, G.D., Joseph, J.A. and Roth, G.S., Effects of age on sensitivity to pain and brain opiate receptors, NeurobioL Ag- ing, 2 (1981) 49-55.
16 Leibowitz, S.F., Paraventricular nucleus: a primary site mediat- ing adrenergic stimulation of feeding and drinking, Pharmacol. Biochem. Behav., 8 (1978) 163-178.
17 Levine, A.S. and Morley, J.E., Neuropeptide Y: a potent in- ducer of consummatory behavior in rats, Peptides, 5 (1984) 1025-1030.
18 Menendez, J.A., McGregor, I.S., Healey, P.A., Atrens, D.M. and Leibowitz, S.E, Metabolic effects of neuropeptide Y injec- tions into paraventricular nucleus of the hypothalamus, Brain Research, 516 (1990) 8-14.
19 Morley, J.E., Levine, A.S., Gosnell, B.A., Kneip, J. and Grace, M., Effects of neuropeptide Y on ingestive behaviours in the rat, Am. J. Physiol., 252 (1987) R599-R609.
20 Morley, J.E., Hernandez, E.N. and Flood, J.E, Neuropeptide Y increases food intake in mice, Am. J. Physiol., 253 (1987) R516-R522.
21 Morley, J.E. and Silver, A.J., Anorexia in the elderly, Neuro- biol. Aging, 9 (1988) 9-16.
22 Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic Co- ordinates, Academic Press, New York, 1982.
271
23 Sahu, A., Kalra, S.P. and Kalra, ES., Food deprivation and in- gestion induce reciprocal changes in neuropeptide Y concentra- tions in the paraventricular nucleus, Peptides, 9 (1988) 83-86.
24 Sahu, A., Kalra, S.P., Crowiey, W.R. and Kalra, ES., Evidence that hypothalamic neuropeptide Y secretion decreases in aged male rats: implication for reproductive aging, Endocrinology, 122 (1988) 376-378.
25 Sahu, A., Kalra, S.E, Crowiey, W.R. and Kalra, ES., Evidence that NPY-containing neurons in the brainstem project into se- lected hypothalamic nuclei: implications in feeding behavior, Brain Research, 457 (1988) 376-378.
26 Sahu, A., Kalra, S.P., Crowley, W.R. and Kalra, P.S., Aging in male rats modifies castration and testosterone-induced neu- ropeptide Y response in various microdissected brain nuclei, Brain Research, 515 (1990) 287-291.
27 Scarpace, EJ. and Abrass, I.B., Alpha- and beta-adrenergic re- ceptor function in the brain during senescence, Neurobiol. Ag- ing, 9 (1988) 53-58.
28 Schick, R.R., Schusdziarra, V., Nussbaumer, C. and Classen, M., Neuropeptide Y and food intake in fasted rats: effect of naloxone and site of action, Brain Research, 552 (1991) 232- 239.
29 Stanley, B.G. and Leibowitz, S.E, Neuropeptide Y: stimulation of feeding and drinking by injections into the paraventricular nucleus, Life Sci., 33 (1984) 2635-2642.
30 Stanley, B.G. and Leibowitz, S.E, Neuropeptide Y injected in the paraventricular hypothalamus: a powerful stimulant of feed- ing behavior, Proc. Nag. Acad. Sci. U.S.A., 82 (1985) 3940- 3943.
31 Stanley, B.G., Kirkouli, S.E., Lampert, S. and Leibowitz, S.E, Neuropeptide Y chronically injected into the hypothalamus: a powerful neurochemical inducer of hyperphagia and obesity, Peptides, 7 (1986) 1189-1192.
32 Stanley, G.B., Lanthier, D., Chin, A.S. and Leibowitz, S.E, Suppression of neuropeptide Y-elicited eating by adrenalectomy or hypophysectomy: reversal with corticosterone, Brain Re- search, 501 (1989) 32-36.
33 Sternberger, L.A. and Sternberger, N.H., The unlabeled anti- body method: comparison of peroxidase-antiperoxidase with avidin-biotin complex by a new method of quantification, J. Histochem. Cytochem., 34 (1986) 509-605.
34 Tatemoto, K., Neuropeptide Y: complete amino acid sequence of the brain peptide, Proc. Natl. Acad. Sci. U.S.A., 79 (1982) 5485-5489.
35 Zoli, M., Agnati, L.E, Fuxe, K., Zini, I., Merlo Pich, E., Grimaldi, R., Harfstrand, A., Goldstein, M., Wikstrom, A.C. and Gustafsson, J.-A., Morphometrical and microdensitometri- cal studies on phenylethanolamine-N-methyltransferase and neuropeptide Y immunoreactive nerve terminals and on gluco- corticoid receptor immunoreactive nerve cell nuclei in the paraventricular hypothalamic nucleus in adult and old male rats, Neuroscience, 26 (1988) 479-492.
36 Zoli, M., Zini, I., Ferraguti, E, Guidolin, D., Fuxe, K. and Agnati, L.E, Aspects of neural plasticity in the central nervous system. I. Computer-assisted image analysis methods, Neuro- chem. Int., 16 (1990) 383-418.
37 Zoli, M., Benfenati, E, Merlo Pich, E., Toffano, G., Fuxe, K. and Agnati, L.E, Aspects of neural plasticity in the central nervous system. IV. Chemical anatomy studies on the aging brain, Neurochem. Int., 16 (1990) 437-449.


![Medial septal [beta]-amyloid 1-40 injections alter septo-hippocampal anatomy and function](https://static.fdokumen.com/doc/165x107/6329e172e9556f820801538c/medial-septal-beta-amyloid-1-40-injections-alter-septo-hippocampal-anatomy-and.jpg)
















