
Innervation of the paraventricular nucleus of the thalamus from cocaine- and amphetamine-regulated...
11
Innervation of the Paraventricular Nucleus of the Thalamus from Cocaine- and Amphetamine-Regulated Transcript (CART) Containing Neurons of the Hypothalamus GILBERT J. KIROUAC*, MATTHEW P. PARSONS, AND SA LI Division of Basic Medical Sciences, Faculty of Medicine, Memorial University of Newfoundland, St. John’s, Newfoundland and Labrador, A1B 3V6, Canada ABSTRACT The paraventricular nucleus of the thalamus (PVT) is a midline thalamic nucleus with heavy projections to the nucleus accumbens and other limbic regions. Previous studies have shown that the PVT contains fibers immunoreactive for cocaine- and amphetamine-related transcript (CART). The purpose of the present study was to determine the location of CART neurons innervating the PVT of the rat by using retrograde tracing with cholera toxin B (CTb) combined with immunofluorescence for CTb and CART (amino acid sequence 55–102). Immunohistochemical analysis of CART in the dorsal thalamus showed that the PVT is densely innervated by CART fibers whereas adjacent midline and intralaminar thalamic nuclei are unlabeled. Injections of CTb in the dorsal midline thalamus retrogradely labeled neurons in several areas of the hypothalamus and brainstem which also contained CART neurons. The largest number of double-labeled neurons (CTb/CART) was found in the arcuate nucleus of the hypothalamus. CTb/CART neurons were also found in the lateral hypothala- mus, zona incerta, and periventricular hypothalamus. These results indicate that the arcuate nucleus is a major source of CART fibers in the PVT. CART neurons in the arcuate nucleus monitor circulating hormonal signals and may regulate food intake and hypothalamic- pituitary-adrenal (HPA) activity. Consequently, CART neurons in the arcuate nucleus may transmit signals to the PVT which in turn may influence limbic regions involved in regulating food intake and the HPA. J. Comp. Neurol. 497:155–165, 2006. © 2006 Wiley-Liss, Inc. Indexing terms: arcuate nucleus; stress food intake; peptide Cocaine- and amphetamine-regulated transcript (CART) was initially described in the rat brain as an activated transcript in response to injection of the psycho- stimulants cocaine and amphetamine (Douglass et al., 1995). Subsequent analysis indicated that the transcript coded for a novel neuropeptide called CART with several potentially active fragments (Kuhar and Yoho, 1999; Thim et al., 1999). CART peptides are putative transmitters that have been implicated in reinforcement, satiety, stress, and endocrine regulation (Dominguez et al., 2004; Hunter and Kuhar, 2003; Hunter et al., 2004; Kuhar et al., 2002). For example, central injections of CART fragments in rats inhibit food intake (Kong et al., 2003; Kristensen et al., 1998; Larsen et al., 2000; Rohner-Jeanrenaud et al., 2002) and increase hypothalamic-pituitary-adrenal (HPA) activity (Smith et al., 2004; Stanley et al., 2001; Vrang et al., 2000). Antisera against CART have been used to map the distribution of neuronal cell bodies and fibers in the CNS (Koylu et al., 1998; Koylu et al., 1997). Dense immu- nostaining for CART in cell bodies was reported for neu- rons in the arcuate, paraventricular, and supraoptic nu- clei as well as the zona incerta of the hypothalamus. Grant sponsor: Canadian Institutes for Health Research. *Correspondence to: G.J. Kirouac, Division of Basic Medical Sciences, Faculty of Medicine, Memorial University of Newfoundland and Labrador, A1B 3V6, Canada. E-mail: [email protected] Received 4 November 2005; Revised 14 December 2005; Accepted 14 January 2006 DOI 10.1002/cne.20971 Published online in Wiley InterScience (www.interscience.wiley.com). THE JOURNAL OF COMPARATIVE NEUROLOGY 497:155–165 (2006) © 2006 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Innervation of the paraventricular nucleus of the thalamus from cocaine- and amphetamine-regulated...

Innervation of the ParaventricularNucleus of the Thalamus from Cocaine-and Amphetamine-Regulated Transcript
(CART) Containing Neurons of theHypothalamus
GILBERT J. KIROUAC*, MATTHEW P. PARSONS, AND SA LI
Division of Basic Medical Sciences, Faculty of Medicine, Memorial University ofNewfoundland, St. John’s, Newfoundland and Labrador, A1B 3V6, Canada
ABSTRACTThe paraventricular nucleus of the thalamus (PVT) is a midline thalamic nucleus with
heavy projections to the nucleus accumbens and other limbic regions. Previous studies haveshown that the PVT contains fibers immunoreactive for cocaine- and amphetamine-relatedtranscript (CART). The purpose of the present study was to determine the location of CARTneurons innervating the PVT of the rat by using retrograde tracing with cholera toxin B(CTb) combined with immunofluorescence for CTb and CART (amino acid sequence 55–102).Immunohistochemical analysis of CART in the dorsal thalamus showed that the PVT isdensely innervated by CART fibers whereas adjacent midline and intralaminar thalamicnuclei are unlabeled. Injections of CTb in the dorsal midline thalamus retrogradely labeledneurons in several areas of the hypothalamus and brainstem which also contained CARTneurons. The largest number of double-labeled neurons (CTb/CART) was found in the arcuatenucleus of the hypothalamus. CTb/CART neurons were also found in the lateral hypothala-mus, zona incerta, and periventricular hypothalamus. These results indicate that the arcuatenucleus is a major source of CART fibers in the PVT. CART neurons in the arcuate nucleusmonitor circulating hormonal signals and may regulate food intake and hypothalamic-pituitary-adrenal (HPA) activity. Consequently, CART neurons in the arcuate nucleus maytransmit signals to the PVT which in turn may influence limbic regions involved in regulatingfood intake and the HPA. J. Comp. Neurol. 497:155–165, 2006. © 2006 Wiley-Liss, Inc.
Indexing terms: arcuate nucleus; stress food intake; peptide
Cocaine- and amphetamine-regulated transcript(CART) was initially described in the rat brain as anactivated transcript in response to injection of the psycho-stimulants cocaine and amphetamine (Douglass et al.,1995). Subsequent analysis indicated that the transcriptcoded for a novel neuropeptide called CART with severalpotentially active fragments (Kuhar and Yoho, 1999; Thimet al., 1999). CART peptides are putative transmittersthat have been implicated in reinforcement, satiety,stress, and endocrine regulation (Dominguez et al., 2004;Hunter and Kuhar, 2003; Hunter et al., 2004; Kuhar et al.,2002). For example, central injections of CART fragmentsin rats inhibit food intake (Kong et al., 2003; Kristensen etal., 1998; Larsen et al., 2000; Rohner-Jeanrenaud et al.,2002) and increase hypothalamic-pituitary-adrenal (HPA)activity (Smith et al., 2004; Stanley et al., 2001; Vrang et
al., 2000). Antisera against CART have been used to mapthe distribution of neuronal cell bodies and fibers in theCNS (Koylu et al., 1998; Koylu et al., 1997). Dense immu-nostaining for CART in cell bodies was reported for neu-rons in the arcuate, paraventricular, and supraoptic nu-clei as well as the zona incerta of the hypothalamus.
Grant sponsor: Canadian Institutes for Health Research.*Correspondence to: G.J. Kirouac, Division of Basic Medical Sciences,
Faculty of Medicine, Memorial University of Newfoundland and Labrador,A1B 3V6, Canada. E-mail: [email protected]
Received 4 November 2005; Revised 14 December 2005; Accepted 14January 2006
DOI 10.1002/cne.20971Published online in Wiley InterScience (www.interscience.wiley.com).
THE JOURNAL OF COMPARATIVE NEUROLOGY 497:155–165 (2006)
© 2006 WILEY-LISS, INC.

Several brainstem regions including the Edinger-Westphal nucleus, nucleus incertus, locus caeruleus, andother noradrenergic cell groups were reported to containCART-labeled neurons. CART immunoreactive fiberswere found to have a widespread distribution in the brainwith dense innervation found in the nucleus accumbens,ventral pallidum, amygdala, many regions of the hypo-thalamus, periaqueductal gray, parabrachial nucleus, nu-cleus of the solitary tract, and area postrema. Further-more, CART is co-stored with the orexigenic melanin-concentrating hormone (MCH) in neurons of the lateralhypothalamus and with the anorexigenic precursor pro-opiomelanocortin (POMC) in neurons of the arcuate nu-cleus (Vrang et al., 1999). Therefore, the distribution ofCART neurons as well as their extensive fiber projectionsis consistent with the diversity of roles proposed for thisnovel peptide (Dominguez et al., 2004; Hunter and Kuhar,2003; Hunter et al., 2004; Kuhar et al., 2002).
The dorsal midline region of the thalamus including theparaventricular nucleus of the thalamus (PVT) was alsoreported to contain a dense plexus of CART fibers (Koyluet al., 1998). The PVT is part of a group of midline andintralaminar thalamic nuclei that are believed to have arole in arousal (Groenewegen and Berendse, 1994; Vander Werf et al., 2002). Consistent with this hypothesis, themidline and intralaminar nuclei receive convergent inputfrom a large number of brainstem cell groups associatedwith arousal, motor, and somatosensory systems (Chenand Su, 1990; Cornwall and Phillipson, 1988; Krout et al.,2002; Krout and Loewy, 2000a; Krout and Loewy, 2000b;Otake and Ruggiero, 1995; Otake et al., 1995). Each nu-cleus of the midline and intralaminar group innervatesspecific cortical areas as well as restricted regions of thebasal ganglia and amygdala (Groenewegen and Berendse,1994; Van der Werf et al., 2002). The PVT is notable fromthe rest of the midline and intralaminar nuclei because itreceives a large and distinct input from several areas ofthe hypothalamus including projections from the supra-chiasmatic, arcuate and dorsomedial, ventromedial, andanterior hypothalamic nuclei (Chen and Su, 1990; Corn-wall and Phillipson, 1988; Krout et al., 2002; Krout andLoewy, 2000a; Krout and Loewy, 2000b; Otake and Rug-giero, 1995; Otake et al., 1995). In turn, PVT neuronsproject to the nucleus accumbens, amygdala, bed nucleusof the stria terminalis, and prefrontal cortex (Bubser andDeutch, 1998; Moga et al., 1995; Otake and Nakamura,1998; Van der Werf et al., 2002), areas of the forebrainassociated with viscerolimbic functions (Cardinal et al.,2002; Pennartz et al., 1994; Walker et al., 2003). Althoughthe function of the PVT remains poorly understood, exper-iments looking at the expression of Fos as a measure ofneuronal activation have consistently shown that neuronsin the PVT are activated during periods of arousal or bystress protocols (Bhatnagar and Dallman, 1998; Bubserand Deutch, 1999; Novak and Nunez, 1998; Novak et al.,2000; Otake et al., 2002; Peng et al., 1995). Furthermore,the PVT has been linked to food intake and the regulationof HPA activity (Bhatnagar and Dallman, 1998; Jaferi etal., 2003; Nakahara et al., 2004), two functions associatedwith CART.
Although the PVT seems to contain a relatively denseplexus of CART fibers, the source of these fibers is notknown. Previous tracing studies showed that the PVT isinnervated by several hypothalamic and brainstem re-gions associated with CART neuron immunoreactivity
(Koylu et al., 1997; Kristensen et al., 1998). The purpose ofthe present study was to determine the source of CARTfibers in the PVT by combining retrograde tracing withimmunofluorescence for CART. Knowledge of the source ofCART fibers in the PVT will be helpful for understandingthe functional relationship between this peptide and amidline thalamic nucleus connected to the limbic system.
MATERIALS AND METHODS
The results presented in this study represent experi-ments done in male Sprague-Dawley rats (200–250 g) thatwere obtained from the Memorial University of New-foundland vivarium and housed on a 12/12 light/dark cyclewith food and water available ad libitum. All efforts weremade to minimize animal suffering and the number ofanimals used. All experiments were done in accordancewith the Canadian Council on Animal Care and wereapproved by Memorial University of Newfoundland.
CART immunoreactivity in the PVT
Four rats were deeply anesthetized with equithesin (0.4ml/100 g) and perfused transcardially with 150 ml of hep-arinized saline followed by 400–500 ml of ice-cold 4%paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Thebrains were removed immediately, post-fixed in the samefixative for 2–4 hours, and cryoprotected in graded su-crose concentrations (10 and 20% w/v) over 2 days at 4°C.Sections of the PVT were cut on a cryostat at 50 �m andplaced in wells containing 0.1M phosphate-buffered saline(PBS). Briefly, every fourth section of the PVT was prein-cubated in blocking solution containing 5% normal donkeyserum, 0.3% Triton-X 100, and 0.1% sodium azide, fol-lowed by overnight incubation in a polyclonal rabbit anti-CART (55–102) primary antibody (1:5,000; Phoenix Phar-maceuticals, Belmont, CA, catalog #H-003-62, lot #00324)prepared in the same blocking solution. The antiserumstains a single band of 5-kD molecular weight on Westernblot (manufacturer’s technical information) and preab-sorption of this antiserum with the CART (55–102) pep-tide fragments eliminated immunostaining of neuronsand fibers in the brain of rats (Dun et al., 2000a; Dun etal., 2000b; Dun et al., 2002). In addition, staining through-out the hypothalamus produced a pattern of immunoreac-tivity that was identical to that previously published byanother group using a different antibody against CART(Koylu et al., 1998; Koylu et al., 1997). Sections were thenrinsed in PBS and incubated for 2–3 hours at room tem-perature in biotinylated donkey anti-rabbit immunoglob-ulin G (1:500; Jackson Immunoresearch). Sections wererinsed again and then exposed to an avidin–biotin com-plex for 60 minutes at room temperature (Elite ABC Kit;Vector Laboratories, Burlingame, CA). After a few morerinses, the tissue was reacted for 5–10 minutes with dia-minobenzidine (DAB) with nickel intensification (VectorDAB Kit) to produce black CART fiber labeling. The DABreaction was terminated by thorough rinsing in PBS be-fore sections were mounted onto gelatin-coated slides andcoverslipped. Representative sections were examined andphotographed using an Olympus BX51 microscopeequipped with real-time digital camera (SPOT RT Slider;Diagnostic Instruments Inc., Sterling Heights, MI). Cali-bration bars were inserted using SPOT software (Version3.2; Diagnostic Instruments) and images were transferred
The Journal of Comparative Neurology. DOI 10.1002/cne
156 G.J. KIROUAC ET AL.

to Adobe Photoshop 5.5 to optimize light and contrastlevels.
Co-localization of CART and synaptophysin
Every fourth section of the PVT in three rats perfusedand prepared as described above was incubated in a pri-mary antibody cocktail of rabbit anti-CART (1:5,000) andmouse anti-synaptophysin (1:500; Chemicon, catalog#MAB329, lot #24110798) for 2–3 days at 4°C. This anti-serum stains a single band of 38-kD molecular weight onWestern blot (Honer et al., 1993). Examination of sectionsthrough the hippocampus showed that the antiserum usedin the present study produced heavy staining in the mossyfiber layer which was similar to that previously publishedusing a different antiserum against synaptophysin (Xu etal., 2004). Sections were rinsed and then transferred to acocktail of Cy2 conjugated donkey anti-rabbit and Cy5conjugated donkey anti-mouse secondary antibodies (1:500; Jackson Immunoresearch Laboratories, West Grove,PA) for 3 hours at room temperature. Sections were thenexamined using the Olympus FV300 confocal microscopeequipped with blue argon (488 nm) and red helium neon(633 nm) lasers. Co-localization was visualized using a100� oil immersion lens combined with a 3� digital zoomwith images collected at 4-�m intervals. Because synap-tophysin antibodies penetrate fixed tissue with difficulty(Calhoun et al., 1996; Hiscock et al., 2000), only the sur-face of the tissue sections was examined for co-localizationof CART and synaptophysin. Optical sections were exam-ined individually to ensure that appearance of doublelabeling was confined to a 0.4-�m thickness of tissue. Inan effort to increase contrast, color settings were adjustedusing the Fluoview 3.0 software (Olympus) so that Cy2labeling was assigned to the red channel whereas Cy5labeling was assigned to the green channel. Image fileswere imported into Adobe Photoshop 5.5 to further opti-mize light and contrast levels.
Retrograde tracing experiments
Six rats were anesthetized with equithesin (0.3 ml/100g, intraperitoneally) and given supplementary doses (0.15ml/100 g) if necessary. Subjects were placed in a Kopfstereotaxic frame and a hand drill was used to expose thebrain surface above the PVT. Iontophoretic injections of0.5% cholera toxin B (CTb; List Biological Laboratory,Campbell, CA) were performed by applying a 3- to 5-�Apositive current (200-ms pulses at 2 Hz for 15 minutes)through a chlorinated silver wire placed in a glass pipette(7–10 �m tip diameter). The coordinates used for injectingCTb into the PVT region were 1.5–2.5 mm posterior and0.8 mm lateral to bregma (angled at 10°), and 5.1 mmventral to the brain surface. The scalp incision was su-tured and rats were returned to their home cages forrecovery. After a 7- to 10-day postoperative survival, ratswere deeply anesthetized and perfused as describedabove. The brains were removed, post-fixed, and cryopro-tected as before. Alternate sections of the brain weretaken at 50 �m and used for the retrograde double-labeling experiments.
A double-labeling immunofluorescent protocol was usedto assess the distribution of CART neurons projecting tothe PVT. Brain sections were preincubated in blockingsolution for 1 hour at room temperature. Sections werethen incubated (2–3 days at room temperature) in a pri-mary antibody cocktail containing goat anti-CTb
(1:40,000; List Biological, catalog #703, lot #7032H) andrabbit anti-CART (1:5,000). After rinsing in PBS, sectionswere transferred to a secondary antibody cocktail of Cy2conjugated donkey anti-rabbit and Cy3 conjugated donkeyanti-goat (1:500; Jackson). Sections were rinsed again,mounted, and a coverslip was placed on the slide.
Double-labeled neurons were analyzed with an Olym-pus fluorescent microscope (BX51) equipped with appro-priate filter cubes for Cy2 (U-MNB2, Olympus), Cy3 (U-MNG2, Olympus), or combination narrow green and bluefilter (U-M51006ZZ, Olympus) combined with a blue/greenexcitation balancer (U-EXBABG, Olympus) to optimizethe contrast under the single filter cube. We counted thenumber of CART, CTb, and double-labeled CTb/CARTneurons from every fourth section of the hypothalamus innuclei containing double-labeled neurons. Brain sectionswere examined and photographed as before. Coronal sec-tions through the PVT taken from a stereotaxic atlas(Paxinos and Watson, 1997) were scanned into Photoshopand tracer injection sites for each subject were drawn intothe corresponding area using the CART immunoreactivityto define the anatomical boundaries of the PVT. Represen-tative sections at different levels of the hypothalamuswere photographed, traced, and schematic drawings weremade in Photoshop to show the spatial location of CTb,CART, and double-labeled neurons.
Control experiments were done for all experiments byremoving either the primary (CTb, CART, synaptophysin)or secondary antibodies. This resulted in the absence ofthe labeling observed when the immunochemical reac-tions were done with the antibodies present.
RESULTS
A dense CART-like immunoreactivity was observed inall levels of the PVT with the heaviest innervation foundin the middle to posterior PVT (Fig. 1). Despite changes inthe shape of the PVT along its anteroposterior axis (Groe-newegen and Berendse, 1994), CART staining was limitedwithin the boundaries of the PVT. Therefore, CART stain-ing can be used to distinguish PVT from other thalamicnuclei adjacent to the PVT (Fig. 1A–D). CART immunore-activity was weak in the paratenial, centromedial, andintermediodorsal nuclei (Fig. 1A–D). CART staining wasslightly above background levels in the paracentral andmediodorsal nuclei (Fig. 1B–D). All regions of the anteriorthalamic nuclei contained a moderate-to-dense plexus ofimmunoreactive CART fibers, as previously described(Koylu et al., 1998).
Examination of CART fibers in the PVT at high magni-fication revealed unevenly spaced fiber swellings of vari-ous sizes which are characteristic of en passant enlarge-ments (Fig. 2B). Experiments were done to determine ifsynaptophysin, a synaptic vesicle protein associated withpresynaptic terminals (Matthew et al., 1981; Wiedenmannand Franke, 1985), is co-localized with CART staining onfiber enlargements in the PVT. Examination of 0.4-�moptical sections of the PVT revealed considerable overlapbetween synaptophysin and CART immunofluorescence(Fig. 2). This overlap was consistently found on the en-largements and not on fiber segments between enlarge-ments.
Results from the tract tracing experiments were ob-tained from six animals with CTb injections largely re-stricted within the PVT (Fig. 3). Most of the injections had
The Journal of Comparative Neurology. DOI 10.1002/cne
157SOURCE OF CART INNERVATION OF THE PVT

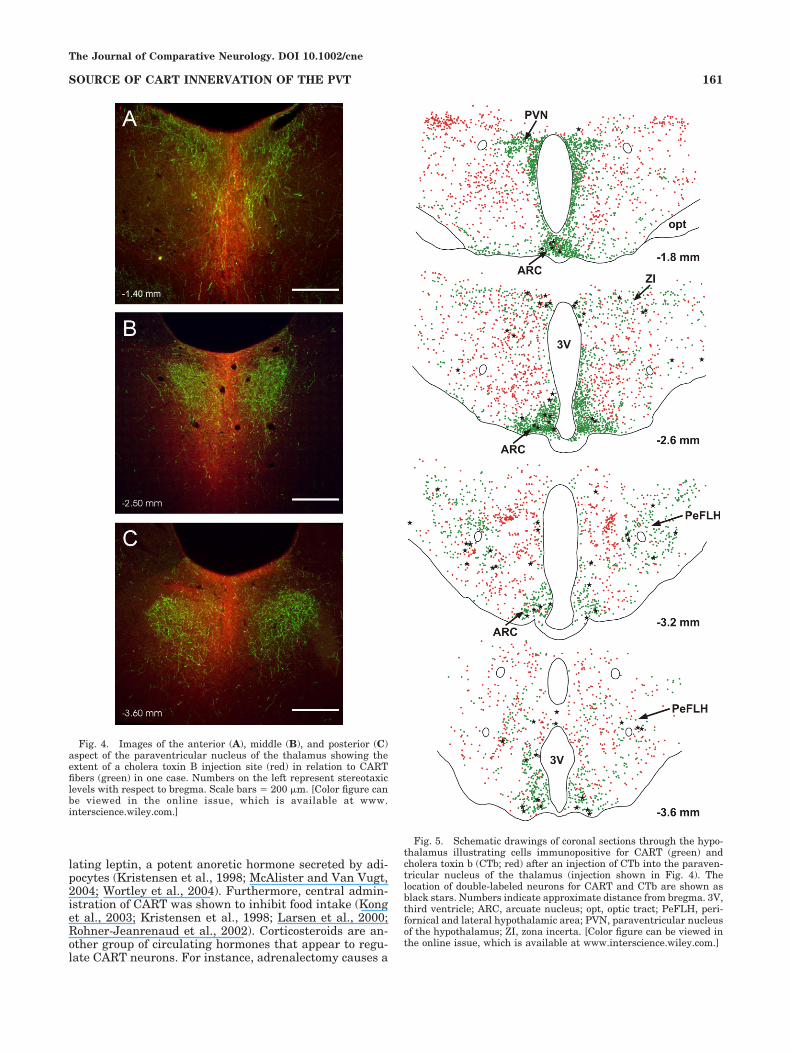
dense cores that extended the anterior to posterior extentof the PVT. Some CTb staining could be seen in nucleiadjacent to the PVT where CART fibers were absent.Figure 4 shows a representative CTb injection in the PVTin relation to CART fibers in the dorsal thalamus. TheCTb injections did not diffuse in the anterior thalamicnucleus or the habenula and could not account for CTb/CART neurons labeled in our experiments. All regionsknown to contain CART neurons were examined for evi-dence of double-labeled CTb/CART neurons, with a focuson areas of the brain with projections to the PVT includingthe hypothalamus, parabrachial nucleus, noradrenergicbrainstem groups, and nucleus of the solitary tract. Figure5 schematically depicts the distribution of CTb, CART,and double-labeled cells in the hypothalamus of a repre-sentative case (injection shown in Fig. 4). A large numberof CTb-labeled cells were found in many regions of thehypothalamus as previously described (Otake et al., 1995)including the areas containing CART immunoreactiveneurons. Close examination of all the cases showed thatCTb/CART double-labeled neurons were located in thearcuate nucleus, lateral hypothalamus, zona incerta, and
along the periventricular regions. The numbers of CART,CTb, and double-labeled neurons in three hypothalamicregions are shown in Table 1. The arcuate nucleus con-tained the largest number of double-labeled neurons de-spite the relatively low number of CTb-labeled neurons inthis nucleus compared with other regions of the hypothal-amus (Figs. 5 and 6). Although the number of CARTneurons in the arcuate nucleus that were retrogradelylabeled by CTb injections in the PVT was low (4.1%), weestimated that 42.8% of CTb-labeled neurons in the arcu-ate nucleus contained CART. The remainder of thedouble-labeled cells were scattered throughout the hypo-thalamus (Fig. 5). The lateral hypothalamus area andzona incerta contained double-labeled neurons but wereless frequent in the arcuate nucleus. The average numberof neurons counted in the six cases is as follows: arcuatenucleus � 86.0 � 27.3; lateral hypothalamus � 19.6 � 4.9;and zona incerta � 17.4 � 3.7 (representing the total in50-�m sections counted bilaterally and taken at every 200�m through the hypothalamus). The periventricular re-gion around the third ventricle also contained somedouble-labeled neurons intermixed with the CART and
Fig. 1. Photomicrographs showing the distribution of CART im-munoreactivity in the dorsal midline and intralaminar nuclei. Theshape of the paraventricular nucleus of the thalamus (PVT) variesthrough its anterior (A,B), middle (C), and posterior (D) extent but iswell defined at each level by heavy CART immunoreactivity. In gen-eral, the PVT is easily distinguished from adjacent thalamic nuclei bydense CART innervation. Numbers indicate approximate distancefrom bregma. AN, anterior nucleus of the thalamus; CM, centrome-
dial nucleus of the thalamus; Hb, habenula; IMD, intermediodorsalnucleus of the thalamus; MD, mediodorsal nucleus of the thalamus;PC, paracentral nucleus of the thalamus; PT, paratenial nucleus ofthe thalamus; PVA, anterior paraventricular nucleus of the thalamus;PVP, posterior paraventricular nucleus of the thalamus; PVT, para-ventricular nucleus of the thalamus; sm, stria medullaris. Scalebars � 450 �m.
The Journal of Comparative Neurology. DOI 10.1002/cne
158 G.J. KIROUAC ET AL.

CTb-labeled cells. The percentage of CART neurons retro-gradely labeled with CTb was low (2.6% with a range of1.5–4.1% for individual hypothalamic area; Table 1). Theparabrachial nucleus, locus caeruleus, and the nucleus ofthe solitary tract are regions of the brainstem that provideafferents to the PVT. However, we found no evidence forprojections to the PVT from brainstem CART neurons(Fig. 7) except for occasional immunoreactive CART neu-rons located in the C1 region of the medulla (Fig. 7D–F).The lack of double labeling in the parabrachial nucleuswas noteworthy because of the relatively large number ofCART and CTb-labeled neurons in lateral parabrachialregion. (Fig. 7A,B).
DISCUSSION
The present study shows that CART is found in fiberslocated in all regions of the PVT whereas the adjacentdorsal midline and intralaminar thalamic nuclei do notcontain CART fibers. We also show that synaptophysin is
co-localized with CART immunoreactivity on fiber en-largements in the PVT. The source of CART fibers in thePVT are from neurons in the arcuate nucleus, lateralhypothalamus, zona incerta, and periventricular region ofthe hypothalamus. These observations provide the firstdescription of CART innervation of the PVT as well as thesource of these fibers. The innervation of the PVT byCART fibers is of interest because the PVT is part of themidline and intralaminar group of nuclei which may func-tion as an ascending arousal system for the entire basalganglia-thalamocortical system (Groenewegen and Be-rendse, 1994). The fact that CART exclusively innervatesthe PVT suggests that this member of the midline andintralaminar group may have unique functions associatedwith peptidergic signals originating from the hypothala-mus.
Although the effect of CART peptide application on PVTneurons is unknown, the present study demonstrates thatCART fiber enlargements express synaptophysin, amarker for potential sites of peptide release (Matthew et
Fig. 2. Confocal imaging showing immunostaining for synaptophysin (A) and CART (B) in theparaventricular nucleus of the thalamus. Merged image (C) showing the colocalization of CART andsynaptophysin on fiber enlargements (examples shown by arrows). Scale bars � 10 �m. [Color figure canbe viewed in the online issue, which is available at www.interscience.wiley.com.]
The Journal of Comparative Neurology. DOI 10.1002/cne
159SOURCE OF CART INNERVATION OF THE PVT

al., 1981; Wiedenmann and Franke, 1985). Electron mi-croscopy has confirmed that CART is packaged withinvesicles in presynaptic terminals making synaptic contactwith postsynaptic neurons in a variety of brain regions(Dallvechia-Adams et al., 2001; Dun et al., 2002; Smith etal., 1999). Although specific receptors for CART have notbeen identified, extracellular and patch-clamp electro-physiological approaches have shown that CART can haveeither inhibitory or excitatory effects depending on theregion of the brain studied (Davidowa et al., 2005; Yermo-laieva et al., 2001). Furthermore, we recently reportedthat CART fiber terminals make putative contacts on pro-jection neurons in the PVT (Parsons et al., 2005). Thepresent study used an antibody against the CART 55–102sequence, which is the most frequently used fragmentinjected when examining the physiological effects of CART(Hunter et al., 2004; Kuhar et al., 2002). Therefore, mul-tiple lines of evidence suggest that CART may be released
in the PVT where it may exert effects on postsynapticneurons.
CART peptides have been implicated in the regulationof food intake and the neuroendocrine system (Hunter andKuhar, 2003; Hunter et al., 2004; Kuhar et al., 2002). Thetracing experiments in this study demonstrate that CARTfibers in the PVT originate from neurons in the hypothal-amus with a major source being the arcuate nucleus. Infact, approximately 43% of arcuate neurons labeled withCTb were found to be CART positive. The arcuate nucleushas been associated with energy homeostasis and neu-roendocrine regulation (Chronwall, 1985; Schwartz et al.,2000), two functions also linked to CART. For example,fasting attenuates CART mRNA (Kristensen et al., 1998;Li et al., 2002; McAlister and Van Vugt, 2004) whereasintake of a high-fat diet leads to an increase in CARTmRNA (Wortley et al., 2004). Evidence also shows thatCART neurons are under stimulatory control from circu-
Fig. 3. Schematic drawings of coronal sections through the dorsalthalamus illustrating the location of cholera toxin b (CTb) injectionsin the paraventricular nucleus of the thalamus (PVT). Six cases (A–F)are each drawn through three different stereotaxic levels represent-ing the anterior (�1.4 mm from bregma), middle (�2.5 mm), andposterior (�3.6 mm) levels. The dense core and diffuse CTb spread are
represented by black and gray shading, respectively. AM, anterome-dial nucleus of the thalamus; CM, centromedial nucleus of the thal-amus; Hb, habenula; IMD, intermediodorsal nucleus of the thalamus;MD, mediodorsal nucleus of the thalamus; PT, paratenial nucleus ofthe thalamus; PVT, paraventricular nucleus of the thalamus. Scalebar � 1 mm.
The Journal of Comparative Neurology. DOI 10.1002/cne
160 G.J. KIROUAC ET AL.
lating leptin, a potent anoretic hormone secreted by adi-pocytes (Kristensen et al., 1998; McAlister and Van Vugt,2004; Wortley et al., 2004). Furthermore, central admin-istration of CART was shown to inhibit food intake (Konget al., 2003; Kristensen et al., 1998; Larsen et al., 2000;Rohner-Jeanrenaud et al., 2002). Corticosteroids are an-other group of circulating hormones that appear to regu-late CART neurons. For instance, adrenalectomy causes a
Fig. 4. Images of the anterior (A), middle (B), and posterior (C)aspect of the paraventricular nucleus of the thalamus showing theextent of a cholera toxin B injection site (red) in relation to CARTfibers (green) in one case. Numbers on the left represent stereotaxiclevels with respect to bregma. Scale bars � 200 �m. [Color figure canbe viewed in the online issue, which is available at www.interscience.wiley.com.]
Fig. 5. Schematic drawings of coronal sections through the hypo-thalamus illustrating cells immunopositive for CART (green) andcholera toxin b (CTb; red) after an injection of CTb into the paraven-tricular nucleus of the thalamus (injection shown in Fig. 4). Thelocation of double-labeled neurons for CART and CTb are shown asblack stars. Numbers indicate approximate distance from bregma. 3V,third ventricle; ARC, arcuate nucleus; opt, optic tract; PeFLH, peri-fornical and lateral hypothalamic area; PVN, paraventricular nucleusof the hypothalamus; ZI, zona incerta. [Color figure can be viewed inthe online issue, which is available at www.interscience.wiley.com.]
The Journal of Comparative Neurology. DOI 10.1002/cne
161SOURCE OF CART INNERVATION OF THE PVT

decrease in CART mRNA and CART peptide levels in thearcuate nucleus which can be reversed by corticosteronereplacement (Balkan et al., 2003; Balkan et al., 2001;Savontaus et al., 2002; Vrang et al., 2003). It is of specialinterest that the PVT is innervated by CART neurons ofthe arcuate nucleus because the PVT may also be involvedin the regulation of the food intake and HPA activity(Bhatnagar and Dallman, 1998; Jaferi et al., 2003; Naka-hara et al., 2004).
A number of CTb/CART neurons were found in thelateral hypothalamus after CTb injections in the PVT. Thelateral hypothalamus contains neurons producing orexinsand MCH, both of which have been implicated in arousaland food intake (Kawano et al., 2002; Kukkonen et al.,2002; Siegel, 2004). Similar to the description of CARTfiber staining in the dorsal midline thalamus presented inthis article, we recently reported that orexin peptides pref-erentially innervate the PVT when compared with theother dorsal midline and intralaminar nuclei (Kirouac etal., 2005). Although the function of CART neurons in thelateral hypothalamus is poorly understood, studies showthat mRNA for CART in the lateral hypothalamus isslightly attenuated in genetic rodent models of obesitycharacterized by a dysfunctional leptin system (Kris-tensen et al., 1998). Another area that contained a smallnumber of CART neurons retrogradely labeled with CTbwas the zona incerta. The zona incerta receives and inte-
grates somatosensory information and relays this infor-mation to dorsal thalamic regions involved in the regula-tion of arousal states (Shaw and Mitrofanis, 2002). ManyCTb-labeled cells that were not labeled for CART werefound in the dorsomedial nucleus, ventromedial nucleus,and periventricular hypothalamic region. Although theneurochemical identity of these CTb neurons was not in-vestigated in the present study, a recent investigationreported that half of the neurons in the dorsomedial nu-cleus retrogradely labeled from a PVT injection of CTbwere cholecystokinin positive (Otake, 2005). Because fi-bers in the PVT are immunoreactive to a large number ofneuropeptides (Freedman and Cassell, 1994; Kirouac etal., 2005; Otake, 2005; Otake and Nakamura, 1995), it islikely that other types of peptidergic neurons in the hypo-thalamus project to the PVT. The fact that CART neuronsof the lateral hypothalamus and zona incerta innervatethe PVT is consistent with the arousal functions associ-ated with the PVT.
As shown in the present study, the PVT receives astrong and distinct innervation from CART neurons in thearcuate nucleus of the hypothalamus. Neurons in the ar-cuate nucleus express CART and sense circulating levelsof hormones such as leptin and corticosterone (Balkan etal., 2003; Balkan et al., 2001; Kristensen et al., 1998;McAlister and Van Vugt, 2004; Savontaus et al., 2002;Vrang et al., 2003; Wortley et al., 2004). CART neurons in
Fig. 6. Photomicrographs showing the distribution of CART andcholera toxin B (CTb) neurons in the arcuate nucleus. CART neuronsare shown in green (A,D) and CTb neurons are shown in red (B,E).Images of Cy2-labeled CART and Cy3-labeled CTb taken under sep-
arate filters were merged to show evidence of double labeling (arrows,C,F). Scale bars � 200 �m in A, B, C; 400 �m in D–F. [Color figure canbe viewed in the online issue, which is available at www.interscience.wiley.com.]
TABLE 1. Number of CART and Retrograde CTb Neurons in Areas of the Hypothalamus Innervating the PVT1
CART CTb Double Double/CART2 Double/CTb3
Zona incerta 1089.6 � 123.9 219.2 � 69.7 17.4 � 3.7 1.5% 12.5%Lateral hypothalamus 856.8 � 152.1 524.8 � 178.4 19.6 � 4.9 2.3% 3.7%Arcuate nucleus 2112.0 � 312.1 200.6 � 57.37 86.0 � 27.3 4.1% 42.8%
1Counts done on 50-�m sections every 200 �m, n � 6.2Pecentage of CART neurons that were double labeled.3Percentage of CTb neurons that were double labeled.
The Journal of Comparative Neurology. DOI 10.1002/cne
162 G.J. KIROUAC ET AL.

the lateral hypothalamus and the zona incerta may signalsimilar types of information (Hunter and Kuhar, 2003;Hunter et al., 2004; Kuhar et al., 2002). CART neuronsmay in turn provide feedback to brain regions involved inthe regulation of food intake and HPA activity. The PVT,a major recipient of CART fibers, has also been implicatedin the regulation of the HPA and food intake. For instance,lesions of the PVT in rats increase food intake and bodyweight (Bhatnagar and Dallman, 1998). Lesions of theposterior part of the PVT block the habituation of HPAactivity in chronic restraint stress (Bhatnagar et al., 2002)whereas lesions of the anterior PVT raise the basal corti-costerone levels in nonstressed rats (Fernandes et al.,2002). The present study did not examine the possibilitythat the anterior and posterior regions of the PVT areinnervated by different CART populations. Our CTb injec-tions were concentrated in the middle portion of the PVTwith significant involvement of both the anterior and pos-terior PVT regions. Therefore, we cannot rule out thepossibility that the different segments of the PVT arepreferentially innervated by discrete populations of CARTneurons.
Neurons of the PVT project to several brain regions alsoassociated with food intake and regulation of the HPA. Forexample, the PVT projects to the nucleus accumbenswhich is connected with behavioral motor control systemsof the lateral hypothalamus and has been implicated inthe control of feeding (Kelley, 2004; Pennartz et al., 1994;Reynolds and Berridge, 2003). In fact, a recent reviewplaces the midline and intralaminar nuclei including thePVT in a critical position in a brain model of integration ofenergy balance, arousal, and food reward (Kelley et al.,2005). The PVT also projects to the medial prefrontalcortex, ventral subiculum of the hippocampus, amygdala,
and bed nucleus of the stria terminalis (Bubser and Deu-tch, 1998; Moga et al., 1995; Otake and Nakamura, 1998;Van der Werf et al., 2002), areas that have been impli-cated in the stress response and regulation of HPA activ-ity (Herman et al., 2003). It is possible that CART isreleased in the PVT along with other hypothalamic neu-ropeptides (Freedman and Cassell, 1994; Kirouac et al.,2005; Otake, 2005; Otake and Nakamura, 1995) where itmay act to influence projection targets of the PVT involvedin regulating food intake or HPA activity.
ACKNOWLEDGMENTS
G.J.K. is a recipient of a New Investigator Award fromthe CIHR/Regional Partnership Program and M.P.P. is arecipient of a Natural Sciences and Engineering ResearchCouncil (NSERC) graduate scholarship.
LITERATURE CITED
Balkan B, Koylu E, Pogun S, Kuhar MJ. 2003. Effects of adrenalectomy onCART expression in the rat arcuate nucleus. Synapse 50(1):14–19.
Balkan B, Koylu EO, Kuhar MJ, Pogun S. 2001. The effect of adrenalec-tomy on cocaine and amphetamine-regulated transcript (CART) ex-pression in the hypothalamic nuclei of the rat. Brain Res 917(1):15–20.
Bhatnagar S, Dallman M. 1998. Neuroanatomical basis for facilitation ofhypothalamic-pituitary-adrenal responses to a novel stressor afterchronic stress. Neuroscience 84(4):1025–1039.
Bhatnagar S, Huber R, Nowak N, Trotter P. 2002. Lesions of the posteriorparaventricular thalamus block habituation of hypothalamic-pituitary-adrenal responses to repeated restraint. J Neuroendocrinol 14(5):403–410.
Bubser M, Deutch AY. 1998. Thalamic paraventricular nucleus neuronscollateralize to innervate the prefrontal cortex and nucleus accumbens.Brain Res 787(2):304–310.
Fig. 7. Photomicrographs showing the distribution of CART andcholera toxin B (CTb) neurons in the parabrachial nucleus (A,B), locuscaeruleus (C), and the C1 area (D–F). CART neurons are shown ingreen and CTb neurons are shown in red. Images of Cy2-labeled
CART (D) and Cy3-labeled CTb (E) were merged to show evidence ofdouble labeling (arrows, F) in the C1 area. Scale bars � 200 �m in A;175 �m in B; 75 �m in C–F. [Color figure can be viewed in the onlineissue, which is available at www.interscience.wiley.com.]
The Journal of Comparative Neurology. DOI 10.1002/cne
163SOURCE OF CART INNERVATION OF THE PVT

Bubser M, Deutch AY. 1999. Stress induces Fos expression in neurons ofthe thalamic paraventricular nucleus that innervate limbic forebrainsites. Synapse 32(1):13–22.
Calhoun ME, Jucker M, Martin LJ, Thinakaran G, Price DL, Mouton PR.1996. Comparative evaluation of synaptophysin-based methods forquantification of synapses. J Neurocytol 25(12):821–828.
Cardinal RN, Parkinson JA, Hall J, Everitt BJ. 2002. Emotion and moti-vation: the role of the amygdala, ventral striatum, and prefrontalcortex. Neurosci Biobehav Rev 26(3):321–352.
Chen S, Su HS. 1990. Afferent connections of the thalamic paraventricularand parataenial nuclei in the rat: a retrograde tracing study withiontophoretic application of Fluoro-Gold. Brain Res 522(1):1–6.
Chronwall BM. 1985. Anatomy and physiology of the neuroendocrine ar-cuate nucleus. Peptides 6(Suppl 2):1–11.
Cornwall J, Phillipson OT. 1988. Afferent projections to the dorsal thala-mus of the rat as shown by retrograde lectin transport. I. The me-diodorsal nucleus. Neuroscience 24(3):1035–1049.
Dallvechia-Adams S, Smith Y, Kuhar MJ. 2001. CART peptide-immunoreactive projection from the nucleus accumbens targets sub-stantia nigra pars reticulata neurons in the rat. J Comp Neurol 434(1):29–39.
Davidowa H, Li Y, Plagemann A. 2005. The main effect of cocaine- andamphetamine-regulated transcript (CART) peptide on hypothalamicneuronal activity depends on the nutritional state of rats. Neuro En-docrinol Lett 26(1):29–34.
Dominguez G, Vicentic A, Del Giudice EM, Jaworski J, Hunter RG, KuharMJ. 2004. CART peptides: modulators of mesolimbic dopamine, feed-ing, and stress. Ann NY Acad Sci 1025:363–369.
Douglass J, McKinzie AA, Couceyro P. 1995. PCR differential displayidentifies a rat brain mRNA that is transcriptionally regulated bycocaine and amphetamine. J Neurosci 15(3 Pt 2):2471–2481.
Dun NJ, Dun SL, Wong PY, Yang J, Chang J. 2000a. Cocaine- andamphetamine-regulated transcript peptide in the rat epididymis: animmunohistochemical and electrophysiological study. Biol Reprod63(5):1518–1524.
Dun SL, Chianca DA Jr, Dun NJ, Yang J, Chang JK. 2000b. Differentialexpression of cocaine- and amphetamine-regulated transcript-immunoreactivity in the rat spinal preganglionic nuclei. Neurosci Lett294(3):143–146.
Dun SL, Ng YK, Brailoiu GC, Ling EA, Dun NJ. 2002. Cocaine- andamphetamine-regulated transcript peptide-immunoreactivity in ad-renergic C1 neurons projecting to the intermediolateral cell column ofthe rat. J Chem Neuroanat 23(2):123–132.
Fernandes GA, Perks P, Cox NK, Lightman SL, Ingram CD, Shanks N.2002. Habituation and cross-sensitization of stress-inducedhypothalamic-pituitary-adrenal activity: effect of lesions in the para-ventricular nucleus of the thalamus or bed nuclei of the stria termina-lis. J Neuroendocrinol 14(7):593–602.
Freedman LJ, Cassell MD. 1994. Relationship of thalamic basal forebrainprojection neurons to the peptidergic innervation of the midline thal-amus. J Comp Neurol 348(3):321–342.
Groenewegen HJ, Berendse HW. 1994. The specificity of the ‘nonspecific’midline and intralaminar thalamic nuclei. Trends Neurosci 17(2):52–57.
Herman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, Ostrander MM, ChoiDC, Cullinan WE. 2003. Central mechanisms of stress integration:hierarchical circuitry controlling hypothalamo-pituitary-adrenocorticalresponsiveness. Front Neuroendocrinol 24(3):151–180.
Hiscock JJ, Murphy S, Willoughby JO. 2000. Confocal microscopic estima-tion of GABAergic nerve terminals in the central nervous system.J Neurosci Methods 95(1):1–11.
Honer WG, Hu L, Davies P. 1993. Human synaptic proteins with a heter-ogeneous distribution in cerebellum and visual cortex. Brain Res609(1–2):9–20.
Hunter RG, Kuhar MJ. 2003. CART peptides as targets for CNS drugdevelopment. Curr Drug Targets CNS Neurol Disord 2(3):201–205.
Hunter RG, Philpot K, Vicentic A, Dominguez G, Hubert GW, Kuhar MJ.2004. CART in feeding and obesity. Trends Endocrinol Metab 15(9):454–459.
Jaferi A, Nowak N, Bhatnagar S. 2003. Negative feedback functions inchronically stressed rats: role of the posterior paraventricular thala-mus. Physiol Behav 78(3):365–373.
Kawano H, Honma S, Honma A, Horie M, Kawano Y, Hayashi S. 2002.Melanin-concentrating hormone neuron system: the Wide Web thatcontrols the feeding. Anat Sci Int 77(3):149–160.
Kelley AE. 2004. Ventral striatal control of appetitive motivation: role iningestive behavior and reward-related learning. Neurosci BiobehavRev 27(8):765–776.
Kelley AE, Baldo BA, Pratt WE. 2005. A proposed hypothalamic-thalamic-striatal axis for the integration of energy balance, arousal, and foodreward. J Comp Neurol 493(1):72–85.
Kirouac GJ, Parsons MP, Li S. 2005. Orexin (hypocretin) innervation of theparaventricular nucleus of the thalamus. Brain Res 1059(2):179–188.
Kong WM, Stanley S, Gardiner J, Abbott C, Murphy K, Seth A, ConnoleyI, Ghatei M, Stephens D, Bloom S. 2003. A role for arcuate cocaine andamphetamine-regulated transcript in hyperphagia, thermogenesis,and cold adaptation. FASEB J 17(12):1688–1690.
Koylu EO, Couceyro PR, Lambert PD, Kuhar MJ. 1998. Cocaine- andamphetamine-regulated transcript peptide immunohistochemical lo-calization in the rat brain. J Comp Neurol 391(1):115–132.
Koylu EO, Couceyro PR, Lambert PD, Ling NC, DeSouza EB, Kuhar MJ.1997. Immunohistochemical localization of novel CART peptides in rathypothalamus, pituitary and adrenal gland. J Neuroendocrinol 9(11):823–833.
Kristensen P, Judge ME, Thim L, Ribel U, Christjansen KN, Wulff BS,Clausen JT, Jensen PB, Madsen OD, Vrang N, Larsen PJ, Hastrup S.1998. Hypothalamic CART is a new anorectic peptide regulated byleptin. Nature 393(6680):72–76.
Krout KE, Belzer RE, Loewy AD. 2002. Brainstem projections to midlineand intralaminar thalamic nuclei of the rat. J Comp Neurol 448(1):53–101.
Krout KE, Loewy AD. 2000a. Parabrachial nucleus projections to midlineand intralaminar thalamic nuclei of the rat. J Comp Neurol 428(3):475–494.
Krout KE, Loewy AD. 2000b. Periaqueductal gray matter projections tomidline and intralaminar thalamic nuclei of the rat. J Comp Neurol424(1):111–141.
Kuhar MJ, Adams S, Dominguez G, Jaworski J, Balkan B. 2002. CARTpeptides. Neuropeptides 36(1):1–8.
Kuhar MJ, Yoho LL. 1999. CART peptide analysis by Western blotting.Synapse 33(3):163–171.
Kukkonen JP, Holmqvist T, Ammoun S, Akerman KE. 2002. Functions ofthe orexinergic/hypocretinergic system. Am J Physiol Cell Physiol283(6):C1567–1591.
Larsen PJ, Vrang N, Petersen PC, Kristensen P. 2000. Chronic intracere-broventricular administration of recombinant CART(42–89) peptideinhibits and causes weight loss in lean and obese Zucker (fa/fa) rats.Obes Res 8(8):590–596.
Li HY, Hwang HW, Hu YH. 2002. Functional characterizations of cocaine-and amphetamine-regulated transcript mRNA expression in rat hypo-thalamus. Neurosci Lett 323(3):203–206.
Matthew WD, Tsavaler L, Reichardt LF. 1981. Identification of a synapticvesicle-specific membrane protein with a wide distribution in neuronaland neurosecretory tissue. J Cell Biol 91(1):257–269.
McAlister ED, Van Vugt DA. 2004. Effect of leptin administration versusre-feeding on hypothalamic neuropeptide gene expression in fastedmale rats. Can J Physiol Pharmacol 82(12):1128–1134.
Moga MM, Weis RP, Moore RY. 1995. Efferent projections of the paraven-tricular thalamic nucleus in the rat. J Comp Neurol 359(2):221–238.
Nakahara K, Fukui K, Murakami N. 2004. Involvement of thalamic para-ventricular nucleus in the anticipatory reaction under food restrictionin the rat. J Vet Med Sci 66(10):1297–1300.
Novak CM, Nunez AA. 1998. Daily rhythms in Fos activity in the ratventrolateral preoptic area and midline thalamic nuclei. Am J Physiol275(5 Pt 2):R1620–1626.
Novak CM, Smale L, Nunez AA. 2000. Rhythms in Fos expression in brainareas related to the sleep-wake cycle in the diurnal Arvicanthis niloti-cus. Am J Physiol Regul Integr Comp Physiol 278(5):R1267–1274.
Otake K. 2005. Cholecystokinin and substance P immunoreactive projec-tions to the paraventricular thalamic nucleus in the rat. Neurosci Res51(4):383–394.
Otake K, Kin K, Nakamura Y. 2002. Fos expression in afferents to the ratmidline thalamus following immobilization stress. Neurosci Res 43(3):269–282.
Otake K, Nakamura Y. 1995. Sites of origin of corticotropin-releasingfactor-like immunoreactive projection fibers to the paraventricular tha-lamic nucleus in the rat. Neurosci Lett 201(1):84–86.
Otake K, Nakamura Y. 1998. Single midline thalamic neurons projectingto both the ventral striatum and the prefrontal cortex in the rat.Neuroscience 86(2):635–649.
The Journal of Comparative Neurology. DOI 10.1002/cne
164 G.J. KIROUAC ET AL.

Otake K, Ruggiero DA. 1995. Monoamines and nitric oxide are employed byafferents engaged in midline thalamic regulation. J Neurosci 15(3 Pt1):1891–1911.
Otake K, Ruggiero DA, Nakamura Y. 1995. Adrenergic innervation offorebrain neurons that project to the paraventricular thalamic nucleusin the rat. Brain Res 697(1–2):17–26.
Parsons MP, Li S, Kirouac GJ. 2006. The paraventricular nucleus of thethalamus as an interface between the orexin and CART peptides andthe shell of the nucleus accumbens. Synapse 59(8);480–490.
Paxinos G, Watson C. 1997. The rat brain in stereotaxic coordinates. 3rded. San Diego. Academic Press.
Peng ZC, Grassi-Zucconi G, Bentivoglio M. 1995. Fos-related protein ex-pression in the midline paraventricular nucleus of the rat thalamus:basal oscillation and relationship with limbic efferents. Exp Brain Res104(1):21–29.
Pennartz CM, Groenewegen HJ, Lopes da Silva FH. 1994. The nucleusaccumbens as a complex of functionally distinct neuronal ensembles:an integration of behavioural, electrophysiological and anatomicaldata. Prog Neurobiol 42(6):719–761.
Reynolds SM, Berridge KC. 2003. Glutamate motivational ensembles innucleus accumbens: rostrocaudal shell gradients of fear and feeding.Eur J Neurosci 17(10):2187–2200.
Rohner-Jeanrenaud F, Craft LS, Bridwell J, Suter TM, Tinsley FC, SmileyDL, Burkhart DR, Statnick MA, Heiman ML, Ravussin E, Caro JF.2002. Chronic central infusion of cocaine- and amphetamine-regulatedtranscript (CART 55-102): effects on body weight homeostasis in leanand high-fat-fed obese rats. Int J Obes Relat Metab Disord 26(2):143–149.
Savontaus E, Conwell IM, Wardlaw SL. 2002. Effects of adrenalectomy onAGRP, POMC, NPY and CART gene expression in the basal hypothal-amus of fed and fasted rats. Brain Res 958(1):130–138.
Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, Baskin DG. 2000. Centralnervous system control of food intake. Nature 404(6778):661–671.
Shaw V, Mitrofanis J. 2002. Anatomical evidence for somatotopic maps inthe zona incerta of rats. Anat Embryol (Berl) 206(1–2):119–130.
Siegel JM. 2004. Hypocretin (orexin): role in normal behavior and neuro-pathology. Annu Rev Psychol 55:125–148.
Smith SM, Vaughan JM, Donaldson CJ, Rivier J, Li C, Chen A, Vale WW.2004. Cocaine- and amphetamine-regulated transcript activates thehypothalamic-pituitary-adrenal axis through a corticotropin-releasingfactor receptor-dependent mechanism. Endocrinology 145(11):5202–5209.
Smith Y, Kieval J, Couceyro PR, Kuhar MJ. 1999. CART peptide-immunoreactive neurones in the nucleus accumbens in monkeys: ul-
trastructural analysis, colocalization studies, and synaptic interactionswith dopaminergic afferents. J Comp Neurol 407(4):491–511.
Stanley SA, Small CJ, Murphy KG, Rayes E, Abbott CR, Seal LJ, MorganDG, Sunter D, Dakin CL, Kim MS, Hunter R, Kuhar M, Ghatei MA,Bloom SR. 2001. Actions of cocaine- and amphetamine-regulated tran-script (CART) peptide on regulation of appetite and hypothalamo-pituitary axes in vitro and in vivo in male rats. Brain Res 893(1–2):186–194.
Thim L, Kristensen P, Nielsen PF, Wulff BS, Clausen JT. 1999. Tissue-specific processing of cocaine- and amphetamine-regulated transcriptpeptides in the rat. Proc Natl Acad Sci USA 96(6):2722–2727.
Van der Werf YD, Witter MP, Groenewegen HJ. 2002. The intralaminarand midline nuclei of the thalamus. Anatomical and functional evi-dence for participation in processes of arousal and awareness. BrainRes Brain Res Rev 39(2–3):107–140.
Vrang N, Larsen PJ, Clausen JT, Kristensen P. 1999. Neurochemicalcharacterization of hypothalamic cocaine-amphetamine-regulatedtranscript neurons. J Neurosci 19(10):RC5.
Vrang N, Larsen PJ, Kristensen P, Tang-Christensen M. 2000. Centraladministration of cocaine-amphetamine-regulated transcript activateshypothalamic neuroendocrine neurons in the rat. Endocrinology141(2):794–801.
Vrang N, Larsen PJ, Tang-Christensen M, Larsen LK, Kristensen P. 2003.Hypothalamic cocaine-amphetamine regulated transcript (CART) isregulated by glucocorticoids. Brain Res 965(1–2):45–50.
Walker DL, Toufexis DJ, Davis M. 2003. Role of the bed nucleus of the striaterminalis versus the amygdala in fear, stress, and anxiety. EurJ Pharmacol 463(1–3):199–216.
Wiedenmann B, Franke WW. 1985. Identification and localization of syn-aptophysin, an integral membrane glycoprotein of Mr 38,000 charac-teristic of presynaptic vesicles. Cell 41(3):1017–1028.
Wortley KE, Chang GQ, Davydova Z, Fried SK, Leibowitz SF. 2004.Cocaine- and amphetamine-regulated transcript in the arcuate nucleusstimulates lipid metabolism to control body fat accrual on a high-fatdiet. Regul Pept 117(2):89–99.
Xu H, He J, Richardson JS, Li XM. 2004. The response of synaptophysinand microtubule-associated protein 1 to restraint stress in rat hip-pocampus and its modulation by venlafaxine. J Neurochem 91(6):1380–1388.
Yermolaieva O, Chen J, Couceyro PR, Hoshi T. 2001. Cocaine- andamphetamine-regulated transcript peptide modulation of voltage-gated Ca2� signaling in hippocampal neurons. J Neurosci 21(19):7474–7480.
The Journal of Comparative Neurology. DOI 10.1002/cne
165SOURCE OF CART INNERVATION OF THE PVT




















