
Electrophysiological correlates of default-mode processing in macaque posterior cingulate cortex
Upload
independentCategory
view
3download
0
B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Short-term facilitation in the anterior cingulate cortexfollowing stimulation of the medial thalamus in the rat
Jyh-Jang Sun a, Jeng Chuang Kung a, Chia-Chuan Wang b,Shin-Lang Chen a, Bai-Chuang Shyu a,⁎aInstitute of Biomedical Sciences, Academia Sinica, Taipei 115, Taiwan, Republic of ChinabCollege of Medicine, Fu-Jen Catholic University, Taipei Hsien 242, Taiwan, Republic of China
A R T I C L E I N F O
⁎ Corresponding author. Fax: +886 2 2782 9224E-mail address: [email protected]
0006-8993/$ – see front matter © 2006 Elsevidoi:10.1016/j.brainres.2006.04.065
A B S T R A C T
Article history:Accepted 17 April 2006Available online 24 May 2006
The present study examined the distribution and localization of synaptic activities (fieldpotentials, multiunit activities and sink source currents) evoked in the anterior cingulatecortex (ACC) by electrical paired pulse stimulation of the ipsilateral medial thalamus (MT).Male Sprague–Dawley rats were anesthetized with halothane (1.0–1.5%), and electricalpaired pulses stimuli (100–300 μA, inter-pulse interval, 100 ms) were delivered to the MT.Tungsten microelectrodes and a multichannel Michigan probe were used to record theevoked field potentials and multiunit activities in the ACC. Paired pulse stimulationfacilitated field potentials and multiunit activities elicited from several MT nuclei. Thesecond component of the negative field potential (com2) was augmented to about 2.5 timesthat of the first component (com1), and the integratedmultiunit activities were facilitated byabout 1.6-fold. Paired stimulation produced an expansion of themaximal negative potentialfrom layer II/III into the deeper layers of the cingulate cortex area 1 (Cg1). Furthermore, thepotentiated activity spread into adjacent secondary motor cortex (M2) and prelimbic cortex(PrL). Meanwhile, the area covered by the maximal integrated multiunit activities expandedfrom layer V (com1) to layers II–V (com2) in M2, Cg1 and PrL. The current source density(CSD) analysis revealed that the short latency sinks were located in layer II/III and layer V/VI.The sink currents were potentiated and expanded to more superficial and to deeper layerswhen a second pulse was delivered with a 100-ms time delay. Sink currents and the pairedpulse facilitation (PPF) were reduced by morphine treatment (5 mg/kg, i.v.), and this effectcould be blocked by naloxone. Electrical stimulation at 10 Hz in the MT induced morepronounced c-fos immunolabeling of neurons in the medial prefrontal cortex than did 1-Hzstimulation. The short-term facilitation occurred in the middle layers and expanded to thedeeper layers of the ACC. These changes may mediate the effective signal transference inthe specific frequency associated with painful responses.
© 2006 Elsevier B.V. All rights reserved.
Keywords:Current source densityField potentialMedial thalamusAnterior cingulate cortexPainPlasticityPaired pulse facilitation
.(B.-C. Shyu).
er B.V. All rights reserved.

102 B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
Abbreviations:ACC, anterior cingulate cortexBDA, biotinylated dextran amineCA1, field 1 of Ammon's hornCg1, cingulate cortex area 1CL, centrolateral thalamic nucleusCM, central medial thalamic nucleuscom1, component 1com2, component 2CSD, current source densityD3V, dorsal third ventricleDG, dentate gyrusLD, laterodorsal thalamic nucleusLHb, lateral habenular nucleusMDC, central mediodorsal thalamicnucleusMDL, lateral mediodorsal thalamicnucleusMDM, medial mediodorsal thalamicnucleusMT, medial thalamusM1, primary motor cortexM2, secondary motor cortexPo, posterior thalamic nuclear groupPPF, paired pulse facilitationPrL, prelimbic cortexPVP, posterior paraventricularthalamic nucleussm, stria medullaris of thalamusVL, ventrolateral thalamic nucleusVPM, ventral posteromedialthalamic nucleus
1. Introduction
The importance of the anterior cingulate cortex (ACC) inmediating the experience of pain has consistently beenemphasized in clinical reports (Foltz and White, 1968;Wilkinson et al., 1999; Wong et al., 1997; Santo et al.,1990; Sherma, 1973; Talbot et al., 1995) as well as in recentfunctional neuroimaging studies (Casey et al., 1994; Coghillet al., 1994; Jones et al., 1991; Talbot et al., 1991; Rainville etal., 1997). These findings have prompted investigation ofthe functional role of the ACC in the affective aspect ofpain in animal models. Studies employing conditioningparadigms that measure learned behaviors following phasicor tonic noxious stimuli have indicated that the ACC playsa crucial role in associating the affective and anticipatoryaspects of pain responses (Johansen et al., 2001; Kung et al.,2003; Koyama et al., 2001; Johansen and Fields, 2004;Donahue et al., 2001).
Electrophysiological studies of the nociceptive neurons inthe ACC provide the basis for elucidating the neuronalmechanisms underlying pain-related learned behaviors. Theelectrophysiological profile characteristics of rodent ACCnociceptive neurons include a bilateral receptive field andwide dynamic range responses (Yamamura et al., 1996);comparable nociceptive responses were found in rabbit,monkey and human (Sikes and Vogt, 1992; Koyama et al.,
2001; Rios et al., 1999). Thus, these ACC nociceptive neuronsexhibit properties suitable for nondiscriminative pain re-sponse processing. Recent studies have revealed that synaptictransmission within the ACC is mediated by glutamatesynapses (Pirot et al., 1994; Gigg et al., 1992; Sah and Nicoll,1991; Tanaka and North, 1994; Wei et al., 1999) and regulatedby opioid receptor activation (Tanaka and North, 1994; Wei etal., 1999). Moreover, short-term and long-term plastic changeshave been reported in the ACC in in vivo and in vitro studies(Wei and Zhuo, 2001; Kung and Shyu, 2002; Gemmell andO'Mara, 2002).
The neuronal plasticity revealed by paired pulse facili-tation (PPF) is a short-term enhancement of synaptictransmission thought to be important for regulatinginformation filtering and integration (Zucker and Regehr,2002). Studies examining augmenting responses and short-term synaptic plasticity in the sensory thalamocorticalpathways have furthered our understanding of the dynamiccortical processes activated by specific inputs (Castro-Alamancos and Connors, 1996; Steriade, 1999). Thosefindings indicate that different types of activation patternsmay account for functional specifications in differentpathways (Castro-Alamancos et al., 1995). The medialthalamus (MT) serves as a major relay in the medial painsystem and in the conveyance of nociceptive informationto the ACC (Albe-Fessard et al., 1985; Bushnell and Duncan,

103B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
1989; Vogt, 1993). Studies using electrical stimulation of theMT have revealed the temporal events and the pharmaco-logical sensitivities of MT-ACC activation (Sikes andDeFrance, 1985; Gigg et al., 1992; Pirot et al., 1994). Asimilar approach was undertaken in a study of ACCplasticity evoked by stimulation of the anterior thalamus(Gemmell and O'Mara, 2002). Our recent study of short-term plastic changes induced by paired pulse stimulationin the MT-ACC pathway therefore provide a useful modelfor functional analyses of the thalamo-cingulate pathwayin terms of its modulation of affective responses to noxiousstimuli (Kung and Shyu, 2002).
The aim of the present study was to examine thedistribution and localization of short-term neuronal plasticityin the ACC. Specifically, plasticity in the form of PPF evoked byelectrical stimulation of the MT was examined. Evoked-fieldpotential andmultiunit activities were recorded, and the sink/source current distributions across cortical layers of the ACCwere determined by analysis of current source density (CSD).Functional activation patterns of the MT-ACC pathwaystimulated by different frequencies were examined by c-fosprotein immunohistochemistry.
Fig. 1 – PPF in the ACC evoked by MT nuclei. (A) PPF of the MT-eactivities (middle sweep). The inter-pulse interval was 100ms. Tyand a positive potential. The PPF was evident in the increased ampulse. The com1 and com2 indicated the first and second responsare shown in the lower panel. (B) Anatomical location of the stimuan arrow) is shown in the upper panel. the stimulation sites (ACCin the MT nuclei from different rats are shown in the lower panelresponses shown in panel A. Abbreviations: Cg1, cingulate cortexthalamic nucleus; LD, laterodorsal thalamic nucleus; M1, primarmediodorsal thalamic nucleus; MDL, lateral mediodorsal thalamposterior thalamic nuclear group; PrL, prelimbic cortex; sm, striaVPM, ventral posteromedial thalamic nucleus. Scale bar = 1 mm
2. Results
2.1. Paired pulse facilitation of MT-evoked ACC fieldpotentials
Constant paired pulse currents were delivered as test stimulithrough the stimulating electrode, and the optimal place-ments of the stimulating and recording electrodes weredetermined to obtain the maximum amplitude of the evokedpotentials. Typical MT-elicited PPF of field potentials andmultiunit activities along the border of the cingulate cortexarea 1 (Cg1) and the ACC was observed as shown in Fig. 1 (A,typical sweeps in upper andmiddle panels; B, recording site inupper panel and stimulating site in lower panel indicated byan arrowhead).
Evoked potentials consisted of an initial negative deflectionfollowed by a positive potential. The peak initial negativepotential coincided in latency with the maximal multiunitactivity excitation. The com1 and com2 were defined as thenegative field potential, multiunit activity, and sink/sourcecurrents evoked by the first and second stimuli, respectively.
voked ACC field potentials (upper sweep) and multiunitpical field potentials consisted of an initial negative potentialplitude of the negative field potential evoked by the secondes. The superimposed sweeps evoked by different MT nucleilation and recording site. The PPF recording site (indicated byresponding, filled circles; ACC-nonresponding, open circles)
. The arrowhead indicates the stimulation site of the examplearea 1; CL, centrolateral thalamic nucleus; CM, central medialy motor cortex; M2, secondary motor cortex; MDC, centralic nucleus; MDM, medial mediodorsal thalamic nucleus; Po,medullaris of thalamus; VL, ventrolateral thalamic nucleus;
.
104 B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
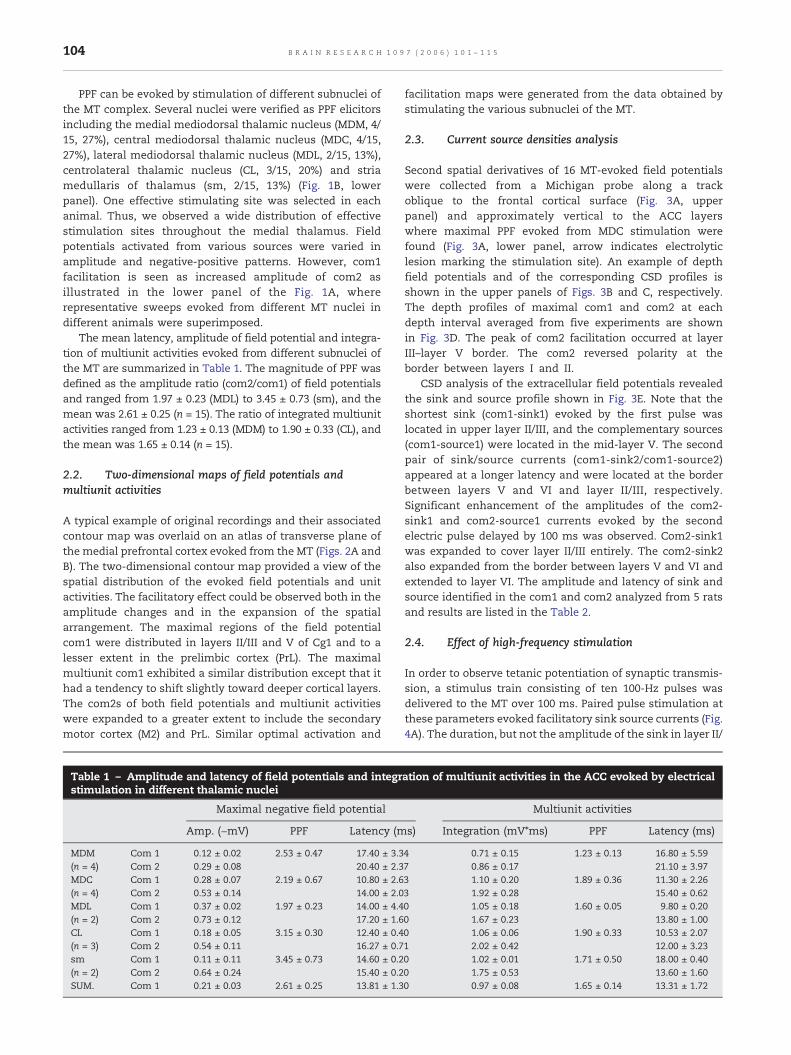
PPF can be evoked by stimulation of different subnuclei ofthe MT complex. Several nuclei were verified as PPF elicitorsincluding the medial mediodorsal thalamic nucleus (MDM, 4/15, 27%), central mediodorsal thalamic nucleus (MDC, 4/15,27%), lateral mediodorsal thalamic nucleus (MDL, 2/15, 13%),centrolateral thalamic nucleus (CL, 3/15, 20%) and striamedullaris of thalamus (sm, 2/15, 13%) (Fig. 1B, lowerpanel). One effective stimulating site was selected in eachanimal. Thus, we observed a wide distribution of effectivestimulation sites throughout the medial thalamus. Fieldpotentials activated from various sources were varied inamplitude and negative-positive patterns. However, com1facilitation is seen as increased amplitude of com2 asillustrated in the lower panel of the Fig. 1A, whererepresentative sweeps evoked from different MT nuclei indifferent animals were superimposed.
The mean latency, amplitude of field potential and integra-tion of multiunit activities evoked from different subnuclei ofthe MT are summarized in Table 1. The magnitude of PPF wasdefined as the amplitude ratio (com2/com1) of field potentialsand ranged from 1.97 ± 0.23 (MDL) to 3.45 ± 0.73 (sm), and themean was 2.61 ± 0.25 (n = 15). The ratio of integrated multiunitactivities ranged from 1.23 ± 0.13 (MDM) to 1.90 ± 0.33 (CL), andthe mean was 1.65 ± 0.14 (n = 15).
2.2. Two-dimensional maps of field potentials andmultiunit activities
A typical example of original recordings and their associatedcontour map was overlaid on an atlas of transverse plane ofthe medial prefrontal cortex evoked from the MT (Figs. 2A andB). The two-dimensional contour map provided a view of thespatial distribution of the evoked field potentials and unitactivities. The facilitatory effect could be observed both in theamplitude changes and in the expansion of the spatialarrangement. The maximal regions of the field potentialcom1 were distributed in layers II/III and V of Cg1 and to alesser extent in the prelimbic cortex (PrL). The maximalmultiunit com1 exhibited a similar distribution except that ithad a tendency to shift slightly toward deeper cortical layers.The com2s of both field potentials and multiunit activitieswere expanded to a greater extent to include the secondarymotor cortex (M2) and PrL. Similar optimal activation and
Table 1 – Amplitude and latency of field potentials and integrstimulation in different thalamic nuclei
Maximal negative field potential
Amp. (−mV) PPF Latency (m
MDM Com 1 0.12 ± 0.02 2.53 ± 0.47 17.40 ± 3.3(n = 4) Com 2 0.29 ± 0.08 20.40 ± 2.3MDC Com 1 0.28 ± 0.07 2.19 ± 0.67 10.80 ± 2.6(n = 4) Com 2 0.53 ± 0.14 14.00 ± 2.0MDL Com 1 0.37 ± 0.02 1.97 ± 0.23 14.00 ± 4.4(n = 2) Com 2 0.73 ± 0.12 17.20 ± 1.6CL Com 1 0.18 ± 0.05 3.15 ± 0.30 12.40 ± 0.4(n = 3) Com 2 0.54 ± 0.11 16.27 ± 0.7sm Com 1 0.11 ± 0.11 3.45 ± 0.73 14.60 ± 0.2(n = 2) Com 2 0.64 ± 0.24 15.40 ± 0.2SUM. Com 1 0.21 ± 0.03 2.61 ± 0.25 13.81 ± 1.3
facilitation maps were generated from the data obtained bystimulating the various subnuclei of the MT.
2.3. Current source densities analysis
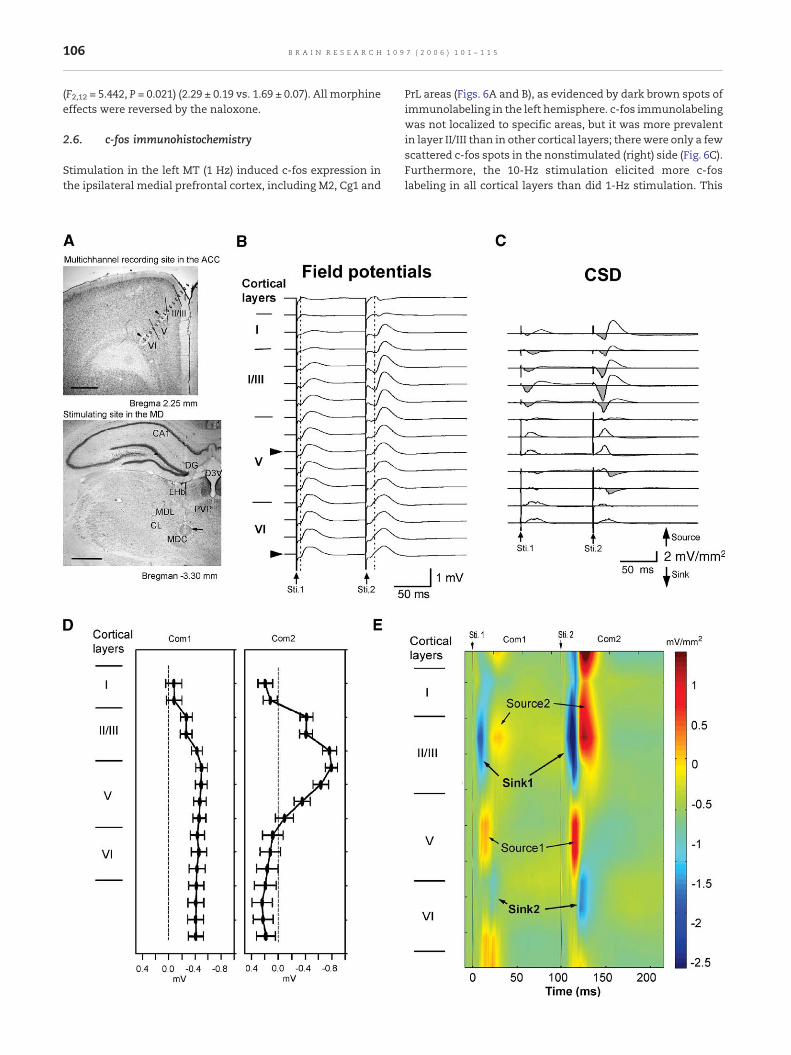
Second spatial derivatives of 16 MT-evoked field potentialswere collected from a Michigan probe along a trackoblique to the frontal cortical surface (Fig. 3A, upperpanel) and approximately vertical to the ACC layerswhere maximal PPF evoked from MDC stimulation werefound (Fig. 3A, lower panel, arrow indicates electrolyticlesion marking the stimulation site). An example of depthfield potentials and of the corresponding CSD profiles isshown in the upper panels of Figs. 3B and C, respectively.The depth profiles of maximal com1 and com2 at eachdepth interval averaged from five experiments are shownin Fig. 3D. The peak of com2 facilitation occurred at layerIII–layer V border. The com2 reversed polarity at theborder between layers I and II.
CSD analysis of the extracellular field potentials revealedthe sink and source profile shown in Fig. 3E. Note that theshortest sink (com1-sink1) evoked by the first pulse waslocated in upper layer II/III, and the complementary sources(com1-source1) were located in the mid-layer V. The secondpair of sink/source currents (com1-sink2/com1-source2)appeared at a longer latency and were located at the borderbetween layers V and VI and layer II/III, respectively.Significant enhancement of the amplitudes of the com2-sink1 and com2-source1 currents evoked by the secondelectric pulse delayed by 100 ms was observed. Com2-sink1was expanded to cover layer II/III entirely. The com2-sink2also expanded from the border between layers V and VI andextended to layer VI. The amplitude and latency of sink andsource identified in the com1 and com2 analyzed from 5 ratsand results are listed in the Table 2.
2.4. Effect of high-frequency stimulation
In order to observe tetanic potentiation of synaptic transmis-sion, a stimulus train consisting of ten 100-Hz pulses wasdelivered to the MT over 100 ms. Paired pulse stimulation atthese parameters evoked facilitatory sink source currents (Fig.4A). The duration, but not the amplitude of the sink in layer II/
ation of multiunit activities in the ACC evoked by electrical
Multiunit activities
s) Integration (mV*ms) PPF Latency (ms)
4 0.71 ± 0.15 1.23 ± 0.13 16.80 ± 5.597 0.86 ± 0.17 21.10 ± 3.973 1.10 ± 0.20 1.89 ± 0.36 11.30 ± 2.263 1.92 ± 0.28 15.40 ± 0.620 1.05 ± 0.18 1.60 ± 0.05 9.80 ± 0.200 1.67 ± 0.23 13.80 ± 1.000 1.06 ± 0.06 1.90 ± 0.33 10.53 ± 2.071 2.02 ± 0.42 12.00 ± 3.230 1.02 ± 0.01 1.71 ± 0.50 18.00 ± 0.400 1.75 ± 0.53 13.60 ± 1.600 0.97 ± 0.08 1.65 ± 0.14 13.31 ± 1.72
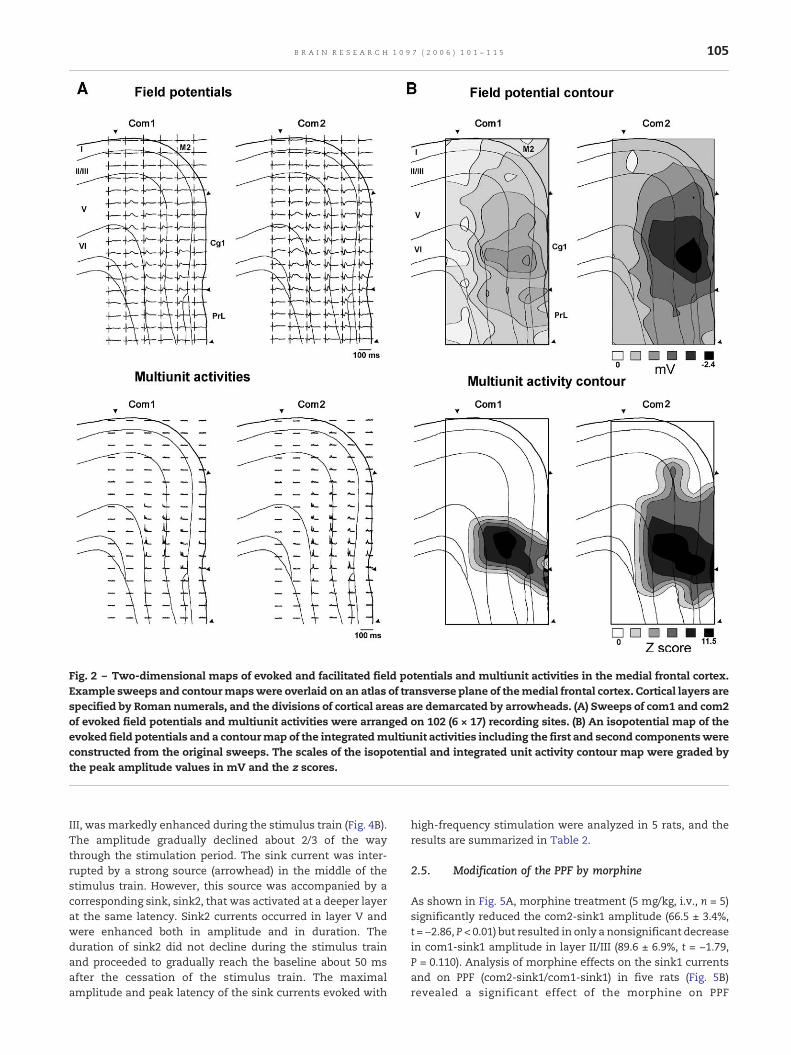
Fig. 2 – Two-dimensional maps of evoked and facilitated field potentials and multiunit activities in the medial frontal cortex.Example sweeps and contourmapswere overlaid on an atlas of transverse plane of themedial frontal cortex. Cortical layers arespecified by Roman numerals, and the divisions of cortical areas are demarcated by arrowheads. (A) Sweeps of com1 and com2of evoked field potentials and multiunit activities were arranged on 102 (6 × 17) recording sites. (B) An isopotential map of theevoked field potentials and a contourmap of the integratedmultiunit activities including the first and second componentswereconstructed from the original sweeps. The scales of the isopotential and integrated unit activity contour map were graded bythe peak amplitude values in mV and the z scores.
105B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
III, was markedly enhanced during the stimulus train (Fig. 4B).The amplitude gradually declined about 2/3 of the waythrough the stimulation period. The sink current was inter-rupted by a strong source (arrowhead) in the middle of thestimulus train. However, this source was accompanied by acorresponding sink, sink2, that was activated at a deeper layerat the same latency. Sink2 currents occurred in layer V andwere enhanced both in amplitude and in duration. Theduration of sink2 did not decline during the stimulus trainand proceeded to gradually reach the baseline about 50 msafter the cessation of the stimulus train. The maximalamplitude and peak latency of the sink currents evoked with
high-frequency stimulation were analyzed in 5 rats, and theresults are summarized in Table 2.
2.5. Modification of the PPF by morphine
As shown in Fig. 5A, morphine treatment (5 mg/kg, i.v., n = 5)significantly reduced the com2-sink1 amplitude (66.5 ± 3.4%,t = −2.86, P < 0.01) but resulted in only a nonsignificant decreasein com1-sink1 amplitude in layer II/III (89.6 ± 6.9%, t = −1.79,P = 0.110). Analysis of morphine effects on the sink1 currentsand on PPF (com2-sink1/com1-sink1) in five rats (Fig. 5B)revealed a significant effect of the morphine on PPF
106 B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
(F2,12 = 5.442, P = 0.021) (2.29 ± 0.19 vs. 1.69 ± 0.07). All morphineeffects were reversed by the naloxone.
2.6. c-fos immunohistochemistry
Stimulation in the left MT (1 Hz) induced c-fos expression inthe ipsilateral medial prefrontal cortex, including M2, Cg1 and
PrL areas (Figs. 6A and B), as evidenced by dark brown spots ofimmunolabeling in the left hemisphere. c-fos immunolabelingwas not localized to specific areas, but it was more prevalentin layer II/III than in other cortical layers; therewere only a fewscattered c-fos spots in the nonstimulated (right) side (Fig. 6C).Furthermore, the 10-Hz stimulation elicited more c-foslabeling in all cortical layers than did 1-Hz stimulation. This

Table 2 – The amplitude and latency of sink currentsevoked by first and second pulses and high frequencystimulation
Sink/Source Current source densities
Com1 Com2 Highfrequency
Sink 1 Amplitude −3.91 ± 1.81 −7.34 ± 2.07 −5.73 ± 1.42Peak latency 9.7 ± 1.39 11.9 ± 0.85 14.8 ± 2.34
Source 1 Amplitude 0.52 ± 0.93 2.32 ± 0.83 3.78 ± 1.13Peak latency 10.40 ± 1.58 11.12 ± 1.12 15.2 ± 1.24
Sink 2 Amplitude −1.45 ± 0.42 −2.78 ± 0.68 −4.53 ± 1.62Peak latency 17.6 ± 1.75 18.3 ± 1.05 21.21 ± 2.41
Source 2 Amplitude 0.72 ± 0.87 2.58 ± 1.53 4.72 ± 1.37Peak latency 18.3 ± 2.47 19.7 ± 2.34 23.21 ± 2.71
Unit for amplitude: (mV/mm2); unit for peak latency: ms. Dataexpressed as means ± SEM (n = 5)
107B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
frequency-dependent effect was especially noticeable in layerII/III (Fig. 6D). A two-way ANOVA showed that the c-fos labeledneurons in the ACC were affected significantly by theapplication of stimulation in the left MT (F2,24 = 43.037,P < 0.001, Fig. 6E). There was significantly more c-fosexpression in the left ACC than that in the right ACC at 10-Hz stimulation (237.0 ± 16.5 vs. 66.6 ± 11.1, P < 0.01) and at 1-Hzstimulation (64.8 ± 8.3 vs. 16.0 ± 4.5, P < 0.01). The amount of c-fos observed in the left ACC after 10-Hz stimulation wassignificantly more than that seen in the left ACC after 1 Hz(P < 0.01) and sham (18.4 ± 3.8, P < 0.01) stimulation.
3. Discussion
The present experiments demonstrated the spatial patternsand temporal sequence of primary and facilitatory neuronalactivities distributed in themedial prefrontal cortex evoked bypaired pulse stimulation in the MT. The cortical activationsources were discrete medial thalamic nuclei, e.g., MD, CL etc.Electrophysiological studies have demonstrated that neuronsin the MD and CL respond to peripheral nociceptive inputs(Berkley et al., 1995; Dostrovsky and Guilbaud, 1990; Shyu etal., 1992; Guilbaud et al., 1986). Likewise, we also previously
Fig. 3 – Localization of MT-evoked sink/source currents in the Amultichannel recording site in the ACC. Arrows indicate two recoindividual recording sites. Short lines mark the cortical layer divAbbreviations: CA1, field 1 of Ammon's horn; CL, centrolateral thaLHb, lateral habenular nucleus; MDL, lateral mediodorsal thalamposterior paraventricular thalamic nucleus. Scale bar = 1mm. (B) Smultichannel electrode along the track with an oblique angle to thaveraged. The inter-stimulus interval was 100 ms. Arrowheads isource density (CSD) profile calculated from the multichannel fiedirected. (D) The peak negative potentials evoked from first anddashed lines in panel B. The depth profile across the cortical layerexperiments. There was a reversal of the com2 negative field ponegative field potentials reached their maximal at the border betcoded. The amplitude of sink and source currents are representesource1 and sink2/source2 pairs labeled in the two-dimensionalcurrents by the second stimulus delivered 100 ms after the first
reported stimulation sites in the CL and other MT nuclei thatwere identified as nociceptive units tested by peripheralnoxious stimuli (Hsu and Shyu, 1997; Kung and Shyu, 2002).The specificity of the electrical stimulation that produced thedistinct cortical responses, the stimulating electrode andapplication of stimulation currents were the same as thatreported in our previous study in which the spread of thecurrent flow was calculated to be localized in a narrow rangewithin a radius of less than 350 μm around the stimulatingelectrode tip (Kung and Shyu, 2002). Thus, nonspecificexcitation of different parts of the medial thalamus resultingfrom current spreading could be excluded.
Topographic maps of field potentials and multiunit activ-ities showed that the primary responses induced in the ACC bydifferentMT stimuli are limited and overlapped in the Cg1 andPrL areas. In a recent study examining the thalamic inputs andanatomical origins of these excitations, we used anterogradetracing of MT projection fibers and found biotinylated dextranamine (BDA) labeled terminals mainly in layers II and III of theACC, in the areas that covered M2, Cg1 and PrL (Wang andShyu, 2004). In the present study c-fos labeled neurons,presumably postsynaptic neurons, following MT stimulationwere distributed in a large region of the medial prefrontalcortex especially in layer II/III. These results are consistentwith previous findings indicating that terminals of differentthalamic nuclei may have extensive overlap in the medialprefrontal cortex (Gioanni et al., 1999; Berendse and Groene-wegen, 1991; Krettek and Price, 1977).
The topographic distribution area covered by maximalactivities of the field potentials and multiunit activities differslightly in that the maximal unit activity extends to a deepercortical layer. The discrepancy may reflect different aspects ofneuronal population responses revealed by the two samplingmethods. It is thought that extracellular local field potentialsrepresent the subthreshold activity from a larger population ofsynchronously excited neurons that reflect the synaptic inputin a cortical region and its intracortical activation (Logothetisand Oeltermann, 2001). Multiunit activity reflects the supra-threshold spiking activity from a small group of neurons, andthe sampling of unit activities may be biased to large cells(Mitzdorf, 1987; Gray et al., 1995; Juergens and Eckhorn, 1999;Logothetis and Oeltermann, 2001). The extension of multiunit
CC using the CSD analysis method. (A) Upper panel,rding site lesions separated by 1000 μM. Dots mark theisions. Lower panel, stimulating site (arrow) in the MDC.lamic nucleus; D3V, dorsal third ventricle; DG, dentate gyrus;ic nucleus; MDC, central mediodorsal thalamic nucleus; PVP,ixteen channel MT-evoked field potentials recorded from thee ACC layers. Twenty sweeps of evoked field potentials werendicate the two recording channel lesion marks. (C) Currentld potentials. Sink currents were shaded and downwardsecond stimuli were selected and indicated by the verticals of the peak negative field potentials were averaged from fivetential at the border of layer I and layer II/III. And the peakween layer II/III and layer V. (E) The CSD profile was colord by blue and red color gradients, respectively. The sink1/map. Note the strong facilitation of the sink and sourcestimulus.
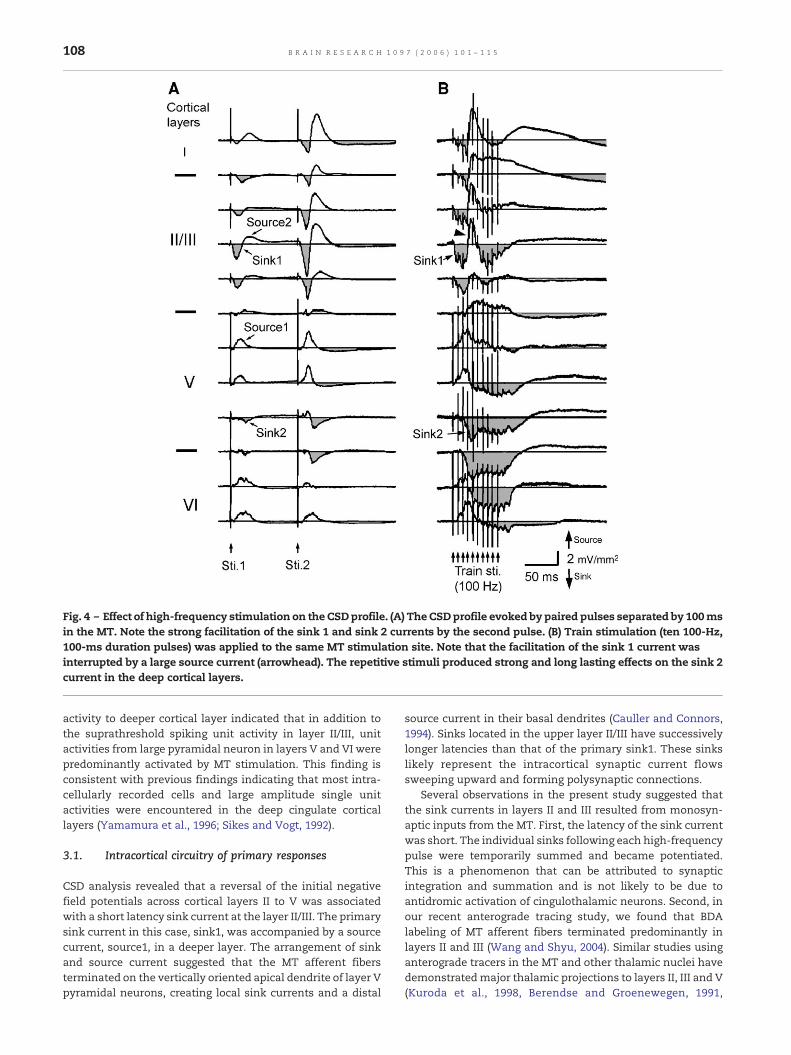
Fig. 4 – Effect of high-frequency stimulation on the CSDprofile. (A) The CSDprofile evoked by paired pulses separated by 100msin the MT. Note the strong facilitation of the sink 1 and sink 2 currents by the second pulse. (B) Train stimulation (ten 100-Hz,100-ms duration pulses) was applied to the same MT stimulation site. Note that the facilitation of the sink 1 current wasinterrupted by a large source current (arrowhead). The repetitive stimuli produced strong and long lasting effects on the sink 2current in the deep cortical layers.
108 B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
activity to deeper cortical layer indicated that in addition tothe suprathreshold spiking unit activity in layer II/III, unitactivities from large pyramidal neuron in layers V and VI werepredominantly activated by MT stimulation. This finding isconsistent with previous findings indicating that most intra-cellularly recorded cells and large amplitude single unitactivities were encountered in the deep cingulate corticallayers (Yamamura et al., 1996; Sikes and Vogt, 1992).
3.1. Intracortical circuitry of primary responses
CSD analysis revealed that a reversal of the initial negativefield potentials across cortical layers II to V was associatedwith a short latency sink current at the layer II/III. The primarysink current in this case, sink1, was accompanied by a sourcecurrent, source1, in a deeper layer. The arrangement of sinkand source current suggested that the MT afferent fibersterminated on the vertically oriented apical dendrite of layer Vpyramidal neurons, creating local sink currents and a distal
source current in their basal dendrites (Cauller and Connors,1994). Sinks located in the upper layer II/III have successivelylonger latencies than that of the primary sink1. These sinkslikely represent the intracortical synaptic current flowssweeping upward and forming polysynaptic connections.
Several observations in the present study suggested thatthe sink currents in layers II and III resulted from monosyn-aptic inputs from the MT. First, the latency of the sink currentwas short. The individual sinks following each high-frequencypulse were temporarily summed and became potentiated.This is a phenomenon that can be attributed to synapticintegration and summation and is not likely to be due toantidromic activation of cingulothalamic neurons. Second, inour recent anterograde tracing study, we found that BDAlabeling of MT afferent fibers terminated predominantly inlayers II and III (Wang and Shyu, 2004). Similar studies usinganterograde tracers in the MT and other thalamic nuclei havedemonstratedmajor thalamic projections to layers II, III and V(Kuroda et al., 1998, Berendse and Groenewegen, 1991,
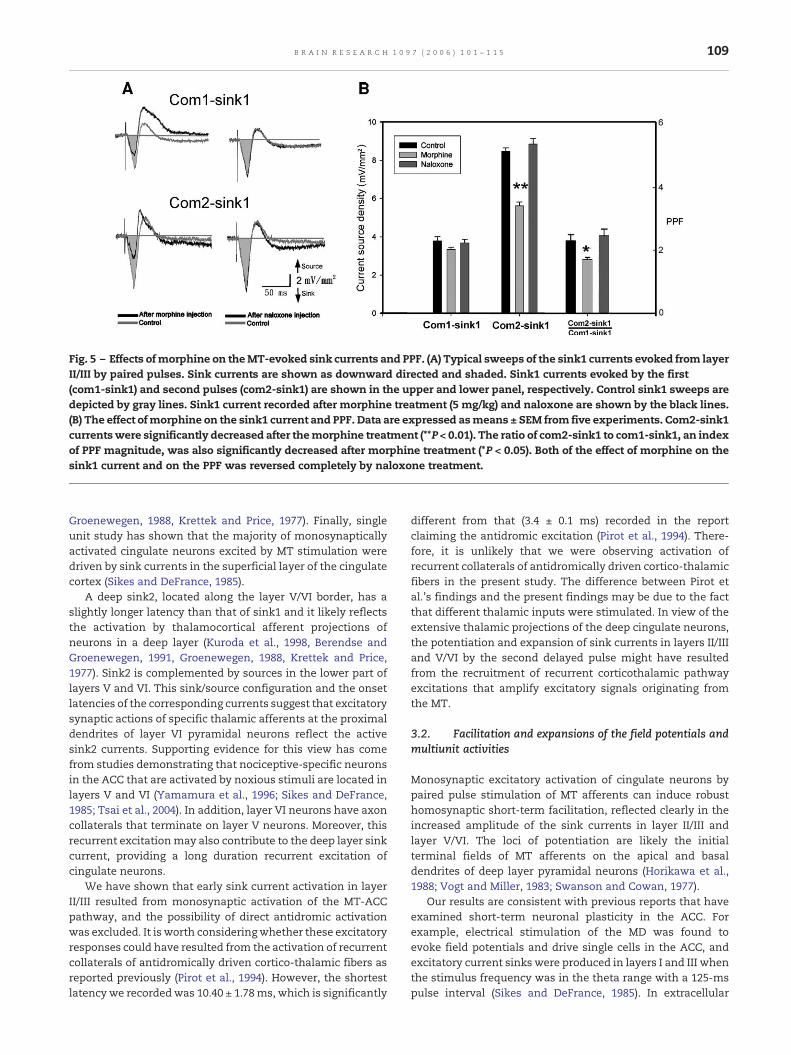
Fig. 5 – Effects ofmorphine on theMT-evoked sink currents and PPF. (A) Typical sweeps of the sink1 currents evoked from layerII/III by paired pulses. Sink currents are shown as downward directed and shaded. Sink1 currents evoked by the first(com1-sink1) and second pulses (com2-sink1) are shown in the upper and lower panel, respectively. Control sink1 sweeps aredepicted by gray lines. Sink1 current recorded after morphine treatment (5 mg/kg) and naloxone are shown by the black lines.(B) The effect ofmorphine on the sink1 current and PPF. Data are expressed asmeans ± SEM from five experiments. Com2-sink1currentswere significantly decreased after themorphine treatment (**P < 0.01). The ratio of com2-sink1 to com1-sink1, an indexof PPF magnitude, was also significantly decreased after morphine treatment (*P < 0.05). Both of the effect of morphine on thesink1 current and on the PPF was reversed completely by naloxone treatment.
109B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
Groenewegen, 1988, Krettek and Price, 1977). Finally, singleunit study has shown that the majority of monosynapticallyactivated cingulate neurons excited by MT stimulation weredriven by sink currents in the superficial layer of the cingulatecortex (Sikes and DeFrance, 1985).
A deep sink2, located along the layer V/VI border, has aslightly longer latency than that of sink1 and it likely reflectsthe activation by thalamocortical afferent projections ofneurons in a deep layer (Kuroda et al., 1998, Berendse andGroenewegen, 1991, Groenewegen, 1988, Krettek and Price,1977). Sink2 is complemented by sources in the lower part oflayers V and VI. This sink/source configuration and the onsetlatencies of the corresponding currents suggest that excitatorysynaptic actions of specific thalamic afferents at the proximaldendrites of layer VI pyramidal neurons reflect the activesink2 currents. Supporting evidence for this view has comefrom studies demonstrating that nociceptive-specific neuronsin the ACC that are activated by noxious stimuli are located inlayers V and VI (Yamamura et al., 1996; Sikes and DeFrance,1985; Tsai et al., 2004). In addition, layer VI neurons have axoncollaterals that terminate on layer V neurons. Moreover, thisrecurrent excitationmay also contribute to the deep layer sinkcurrent, providing a long duration recurrent excitation ofcingulate neurons.
We have shown that early sink current activation in layerII/III resulted from monosynaptic activation of the MT-ACCpathway, and the possibility of direct antidromic activationwas excluded. It is worth consideringwhether these excitatoryresponses could have resulted from the activation of recurrentcollaterals of antidromically driven cortico-thalamic fibers asreported previously (Pirot et al., 1994). However, the shortestlatency we recordedwas 10.40 ± 1.78ms, which is significantly
different from that (3.4 ± 0.1 ms) recorded in the reportclaiming the antidromic excitation (Pirot et al., 1994). There-fore, it is unlikely that we were observing activation ofrecurrent collaterals of antidromically driven cortico-thalamicfibers in the present study. The difference between Pirot etal.'s findings and the present findings may be due to the factthat different thalamic inputs were stimulated. In view of theextensive thalamic projections of the deep cingulate neurons,the potentiation and expansion of sink currents in layers II/IIIand V/VI by the second delayed pulse might have resultedfrom the recruitment of recurrent corticothalamic pathwayexcitations that amplify excitatory signals originating fromthe MT.
3.2. Facilitation and expansions of the field potentials andmultiunit activities
Monosynaptic excitatory activation of cingulate neurons bypaired pulse stimulation of MT afferents can induce robusthomosynaptic short-term facilitation, reflected clearly in theincreased amplitude of the sink currents in layer II/III andlayer V/VI. The loci of potentiation are likely the initialterminal fields of MT afferents on the apical and basaldendrites of deep layer pyramidal neurons (Horikawa et al.,1988; Vogt and Miller, 1983; Swanson and Cowan, 1977).
Our results are consistent with previous reports that haveexamined short-term neuronal plasticity in the ACC. Forexample, electrical stimulation of the MD was found toevoke field potentials and drive single cells in the ACC, andexcitatory current sinks were produced in layers I and III whenthe stimulus frequency was in the theta range with a 125-mspulse interval (Sikes and DeFrance, 1985). In extracellular
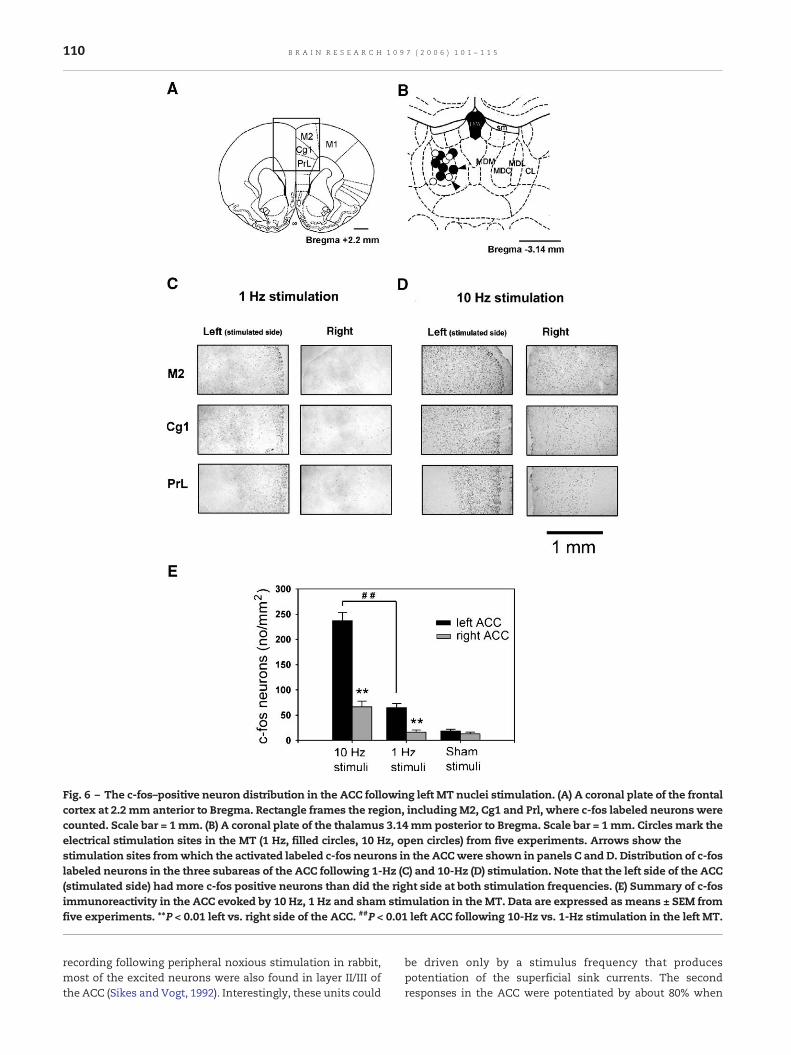
Fig. 6 – The c-fos–positive neuron distribution in the ACC following left MT nuclei stimulation. (A) A coronal plate of the frontalcortex at 2.2 mm anterior to Bregma. Rectangle frames the region, including M2, Cg1 and Prl, where c-fos labeled neurons werecounted. Scale bar = 1mm. (B) A coronal plate of the thalamus 3.14 mm posterior to Bregma. Scale bar = 1 mm. Circles mark theelectrical stimulation sites in the MT (1 Hz, filled circles, 10 Hz, open circles) from five experiments. Arrows show thestimulation sites fromwhich the activated labeled c-fos neurons in the ACCwere shown in panels C and D. Distribution of c-foslabeled neurons in the three subareas of the ACC following 1-Hz (C) and 10-Hz (D) stimulation. Note that the left side of the ACC(stimulated side) had more c-fos positive neurons than did the right side at both stimulation frequencies. (E) Summary of c-fosimmunoreactivity in the ACC evoked by 10 Hz, 1 Hz and sham stimulation in the MT. Data are expressed as means ± SEM fromfive experiments. **P < 0.01 left vs. right side of the ACC. ##P < 0.01 left ACC following 10-Hz vs. 1-Hz stimulation in the left MT.
110 B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
recording following peripheral noxious stimulation in rabbit,most of the excited neurons were also found in layer II/III ofthe ACC (Sikes and Vogt, 1992). Interestingly, these units could
be driven only by a stimulus frequency that producespotentiation of the superficial sink currents. The secondresponses in the ACC were potentiated by about 80% when

111B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
the anterior thalamus was stimulated with a 100-ms pulseinterval (Gemmell and O'Mara, 2002). A similar observationwas reported in layer V neocortical neurons; intracellularrecording showed that action potentials could be initiated bythe second pulse delivered at 100 ms with stimulation ofventrolateral thalamic nucleus (Castro-Alamancos and Con-nors, 1996).
3.3. Excitatory and modulatory neurotransmittersinvolved
Thalamocortical pathways are mediated by glutaminergictransmission and synaptic plastic changes due to the activa-tion of cortical glutaminergic projections have been describedpreviously (Greengard et al., 1991; Hedberg and Stanton, 1995;Kirkwood et al., 1993; Wei et al., 1999). Paired pulse facilitationis usually considered to be a presynaptic phenomenoninvolving changes in intracellular calcium levels broughtabout by previous stimuli (Dittman et al., 2000; Emptage etal., 2001; Zucker and Regehr, 2002). Castro-Alamancos andConnors (1996) reported in vivo and in vitro experimentsconducted in the thalamocortical pathway that demonstratedthat one of the short-term plasticity forms, the augmentingresponse, may be due, at least in part, to a post-synapticinhibitory system. Our recent findings showed that thecellular mechanism of short-term synaptic facilitation in theACC could be mediated by postsynaptic AMPA and GABAreceptors (Sylantyev et al., 2004, 2005); these findings in theACC were consistent with the prior results obtained in lateralamygdala (Zinebi et al., 2001) and in CA1 area of thehippocampus (Berton et al., 1999). Demonstrations of post-synaptic regulation of PPF have also been reported recently,but the possible mechanisms that may be mediating thisregulation are quite diverse and inconclusive relative to that inpresynaptic actions (Jones and Westbrook, 1995; Wang andKelly, 1997; Rozov and Burnashev, 1999; Kirischuk et al., 2002).
The level of excitability in the ACC is under constantmodification and monitoring from several ascending modu-latory inputs, including those from the cholinergic, monoam-inergic (Woolf et al., 1984; Mesulam et al., 1983) andmesolimbic dopaminergic systems (Pirot et al., 1996). Intra-cortical circuitries by which the ACC regulates responses tonoxious stimuli can also be influenced by opioid interneuronsthat terminate directly on deep pyramidal neurons. Moderateto high levels of the μ and σ opioid receptors are present in thedeep layers of cingulate cortex (Mansour et al., 1987). In thepresent study, the action of themorphine on the sink currentsand the reduction of the PPF provided further evidence thatopioid receptor activation influences intra-cortical trans-synaptic interaction.
3.4. Frequency filtering and functional implications
Paired stimuli in the MT produced a striking temporal spatialresponse profile in the medial prefrontal cortex in that thespatial maps of cortical activation attributable to varioussites of MT stimulation overlapped considerably. Thisoverlap may lead to interactions between different medialthalamic afferent systems. We observed in the present studythat when different parts of the MT were excited by
peripheral noxious afferent inputs, distinct patterns oflaminar and topographic profiles in the medial prefrontalcortex were activated. Thus, thalamo-cingulate afferentsoriginating from different nuclei and projecting to differentlayers of the same limbic cortical areas may give rise todiscrete and diverse spatiotemporal responses, which mayunderlie the functional specializations of these pathways. Asimilar functional operation model was described for senso-rimotor cortical projections of the ventrolateral and ventralposterior lateral thalamic nuclei (Castro-Alamancos andConnors, 1996).
The induction of short-term changes in the strength ofcritical synaptic pathways could play a significant role in theprocessing and storage of amplitude and frequency modulat-ed inputs and in the gating of MT output. c-fos labeling in theACC further indicates that this facilitatory effect can beinduced by 10-Hz stimulation in the MT. For effectivetransmission, the thalamic inputs must modulate in the 8 to12 Hz or the theta frequency range. Activities in this frequencyrangewould be amplified and thus able to stimulate the cortexhighly effectively.
It has been proposed that PPF may act as a filter thatenables the cortical circuit to respond only to a specificrange of frequencies. This property can be used to extractsignals from noise and can transform temporal informa-tion into a spatial code (Zucker and Regehr, 2002;Buonomano and Merzenich, 1995). The observation thatthe short-term facilitation occurred first in the middlelayers and then expanded to the deeper layers indicatesthat neural plasticity may involve information transfer toother brain structures such as the hypothalamus, somato-sensory cortex and hippocampus (Izaki et al., 2001). Recentfindings indicating that the ACC may play an importantrole in the plastic changes following the peripheral injury(Wei and Zhuo, 2001) and that the ACC may mediate placeconditioning in the formalin test (Johansen et al., 2001)suggest that the excitability changes and potentiation inthe ACC observed here may have important functionalsignificance.
4. Experimental procedures
4.1. Preparation of animals
Male Sprague–Dawley rats (body weight 250–350 g) were used.They were housed individually in standard wire-mesh cagesin an air-conditioned room (21–23 °C, humidity 50%, 12/12-h light/dark cycle, lights on at 6 am) with free access to foodand water. Fifteen rats were randomly assigned for electro-physiological study and for morphine treatment. Functionalmapping of the distribution of c-fos labeled neurons wasperformed in another group of fifteen rats. All experimentswere carried out in accordance with the guidelines of theAcademia Sinica Institutional Animal Care and UtilizationCommittee.
Rats were initially anesthetized with 4% halothane (in100% O2) in an acrylic box. After tracheal catheterization,anesthetized animals were mounted on a small animalstereotaxic apparatus (David Kopf Instruments, Tujunga,

112 B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
CA, USA) and maintained in a state of anesthesia with 2%halothane (in 30%/70% nitrous oxide/oxygen mixture) duringthe surgical procedures. A venous catheter was inserted fordrug delivery. Body temperature was maintained at ≥36.5 °Cvia a homeothermo blanket system. Electrocardiogram,respiratory rate and CO2 concentration were monitored foreach rat throughout the recording session. To provide accessto the ACC for recording, a small hole was made in the skullfrom 2 to 5 mm anterior to Bregma and 0.5 mm lateral to themidline. Another opening was made from 1 to 3 mmposterior to Bregma and 0.5 to 2 mm lateral to the midlinethrough which the MT was stimulated. The dura wasremoved, and the brain surface was covered with warmparaffin oil during recording. After surgical preparation, theanimals were maintained under anesthesia with 1–1.5%halothane in 70%/30% nitrous oxide/oxygen. The localanesthetic EMLA® Cream (lidocaine 2.5% and prilocaine2.5% cream, AstraZeneca, Sweden) was applied topicallyonto the surgical wounds. To monitor the anestheticdisassociation level, withdrawal responses were tested bysqueezing and pinching of the tail or hind limb. Higherhalothane concentration was given when withdrawalresponses appeared or heart rate increased.
4.2. Electrical stimuli in the MT and recording of evokedfield potentials in the ACC
The electrophysiological procedures conducted in the ACCand MT were similar to those described previously (Kung andShyu, 2002). Mono-polar tungsten electrodes (A-M system Inc.,Carlsborg, WA, USA) with an impedance of 12 MΩ were placedin the MT. Most of the thalamic units that responded toperipheral stimulation were detected by the recording electro-des that were inserted into the cortical surface near 2.5 mmposterior to Bregma, 1 mm lateral of the midline and 5 mmbelow the dura. We used these observations to guide ourplacement of recording electrodes in the present study. Pairedpulse stimuli (0.1 ms, 100–300 μA) were delivered to the MTfrom an isolated pulse stimulator (Model 2100, A-M SystemsInc., USA). To obtain the optimal ratio of facilitation, the inter-pulse interval was set to 80–120 ms.
Tungsten microelectrodes with an impedance of 5 MΩwere used to record evoked field potentials and multiunitactivities in the ACC ipsilateral to the MT site wherestimulation was delivered. To enable a series of evokeddepth field potentials to be recorded, the intra-ACC electro-des were inserted with systematic spacing at a regular depthinterval of 250 μm in the transverse plane of the medialfrontal cortex in each animal. Successive electrodes wereinserted in this manner until a rectangular recording arraycomposed of 6 medial to lateral columns and 17 dorsal toventral rows was formed. This rectangular recording arraywas situated 2.5 mm anterior to and 0–2 mm lateral ofBregma and 0–4 mm below the dura. Sampling of 102recording points was completed within 30 min. The entirepool of recording sites covering an area of approximately1.5 × 4 mm2 were obtained in each experiment and used toconstruct an isopotential map and an integrated multiunitactivity contour map that included the first and secondresponses.
4.3. Multichannel recording
We employed a specially designed multichannel siliconrecording electrode probe provided by the NIH Center forNeural Communication Technology at the University ofMichigan. The probe had 16 recording sites spaced 150 μmapart between the recording sites on a vertical shank. Fieldpotentials were collected along the track of an oblique angle ofabout 45° that was approximately vertical to the ACC layersfrom the surface down to a depth of 6000 μmbelow the corticalsurface. Twenty sweeps of evoked field potentials wereaveraged.
4.4. Recording devices and data acquisition and analysis
The extracellular field potentials produced in the ACCduring PPF were amplified by a high input impedanceamplifier (Axoclamp 2A, Axon Instrument Inc., USA). Multi-unit activities were obtained by passing the field potentialsignals through a high-pass (200–5 kHz) filter in an ACdifferential amplifier (Model 1700, A-M Systems Inc., USA).Both the field potential and multiunit activity analog signalswere sent to a PC-based data acquisition system for on-lineA/D conversion (Sampling rate, 5 kHz, DAS-16F, Metrabyte,USA). Groups of 10–20 sweeps of evoked potentials andfiltered potentials were averaged separately. All signal datawere stored on the hard disk for off-line processing in a PC-based data analysis system. The field potentials wereanalyzed according to their amplitude and latency. Multiunitactivities were rectified, and the area of the rectified poten-tials was integrated.
Signals recorded by the multichannel Michigan probe wereamplified and digitalized by a Medusa BioAmp System(Tucker-Davis Tech, USA). The data were recorded by OpenExsoftware with a custom design circuit with a 6-kHz samplingrate and a 0.1-Hz low-pass filter. The data were analyzed withthe Matlab program. The sink and source currents werederived from the second spatial derivations of the 16simultaneously recorded field potentials (Mitzdorf, 1985;Freeman and Nicholson, 1975). A nine-point formula tosmooth the spatial sampling variability was adopted, andthe final sink and source data values were calculated with thefinite difference formula (Freeman and Nicholson, 1975; Shyuet al., 1989).
4.5. Isopotential map of field potentials
The sweep with the largest negative field potential from the102 sweeps was selected, and the peak latencies of com1 andcom2 in this sweep were used as the time point at which thecom1 and com2 voltage values from all sweeps werecalculated. An isopotential map of the com1 and com2 datafor each rat was constructed based on these values.
4.6. Integration contour map of multiunit activities
Two time periods of integrated multiunit activity data werecalculated. The value calculated from the 30-ms time periodpreceding the stimulation artifact was taken as the basalactivity. The value measured from the 30-ms time period

113B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
after the stimulation artifact encompassed the evokedresponses and was taken to represent the response activity.The z score values were obtained by dividing the responseactivity of each of the 102 sweeps by the mean of the basalactivity values of all sweeps. If the z score was less than2.326 (P < 0.01), the response activity was considered notdifferent than zero. A contour map of the 102 multiunitresponse activities was constructed based on their z scorevalues.
4.7. Verification of electrodes sites
A small lesion was made by passing direct electric current(30 μA for 10 s) at the location of the tip of the recording andstimulating tungsten electrode.When theMichigan probewasused, a small lesion was made by passing an anodal current(30 μA for 5 s) to the 16th (deepest) contact lead at the end ofthe experiment. Another lesion was made at the same leadafter the Michigan probe was withdrawn 1 mm. Rats werefixed by perfusion with physiological saline followed by 10%formalin. Sixty-micrometer-thick coronal sections were col-lected using a cryostat and the sections were stained withcresyl violet (Sigma, USA). Drawings representing each sectionwere made that included clear electrode tracks and lesionmarkers relative to the ACC and MT anatomy; the drawingswere used to reconstruct the recording and stimulating sites.The rat atlas of Paxinos and Waston (1998) served as areference when detailed histological structures wereestimated.
4.8. C-fos immunohistochemistry
Rats were prepared as described above for the electrophys-iological study. PPF was verified in the ACC by placing thestimulating electrode in the position in the left MT thatdemonstrated effective facilitation. Based on our observa-tions of the optimal paradigm for generating the pairedpulse facilitatory effect, we chose to use 10-Hz and 1-Hzfrequency stimulation trains for making comparisons be-tween effective (10 Hz) and noneffective (1 Hz) thalamicstimulation. Animals were killed 1.5 h after 1 Hz, 10-minstimulation (in 5 rats), 1.5 h after 10 Hz, 1-min (in 5 rats)electrical stimulation, or 1.5 h after sham stimulation (in 5rats) in the MT. Rats were perfused transcardially with300 ml of physiological saline followed by 300 ml of 4%paraformaldehyde in 0.1 M, pH 7.4 phosphate-bufferedsaline. The c-fos immunohistochemistry procedureemployed was the same as that described in our previousstudy (Shyu et al., 2004). The number of c-fos labeledneurons was counted in 4 randomly chosen areas in theACC. In each area, labeled neurons within a standard area(0.01 mm2) were counted independently by two experimen-ters, and the mean of the two counts in each of the 4 areaswas averaged.
Acknowledgments
Multichannel silicon probeswere provided by theUniversity ofMichigan Center for Neural Communication Technology,
sponsored by NIH NIBIB grant P41-RR09754. The presentstudy was supported by grants from the National ScienceCouncil and Academia Sinica of the Republic of China.
R E F E R E N C E S
Albe-Fessard, D., Berkley, K.J., Kruger, L., Ralston III, H.J., Willis Jr.,W.D., 1985. Diencephalic mechanisms of pain sensation. BrainRes. 356, 217–296.
Berendse, H.W., Groenewegen, H.J., 1991. Restricted corticaltermination fields of the midline and intralaminar thalamicnuclei in the rat. Neuroscience 42, 73–102.
Berkley, K.J., Benoist, J.M., Gautron, M., Guilbaud, G., 1995.Responses of neurons in the caudal intralaminar thalamiccomplex of the rat to stimulation of the uterus, vagina, cervix,colon and skin. Brain Res. 695, 92–95.
Berton, F., Brancucci, A., Beghe, F., Cammalleri, M., Demuro, A.,Francesconi, W., Gessa, G.L., 1999. Gamma-hydroxybutyrateinhibits excitatory postsynaptic potentials in rat hippocampalslices. Eur. J. Pharmacol. 380, 109–116.
Buonomano, D.V., Merzenich, M.M., 1995. Temporal informationtransformed into a spatial code by a neural network withrealistic properties. Science 267, 1028–1030.
Bushnell, M.C., Duncan, G.H., 1989. Sensory and affective aspectsof pain perception: is medial thalamus restricted to emotionalissues? Exp. Brain Res. 78, 415–418.
Casey, K.L., Minoshima, S., Berger, K.L., Koeppe, R.A., Morrow, T.J.,Frey, K.A., 1994. Positron emission tomographic analysis ofcerebral structures activated specifically by repetitive noxiousheat stimuli. J. Neurophysiol. 71, 802–807.
Castro-Alamancos, M.A., Connors, B.W., 1996. Cellularmechanisms of the augmenting response: short-termplasticity in a thalamocortical pathway. J. Neurosci. 16,7742–7756.
Castro-Alamancos, M.A., Donoghue, J.P., Connors, B.W., 1995.Different forms of synaptic plasticity in somatosensory andmotor areas of the neocortex. J. Neurosci. 15, 5324–5333.
Cauller, L.J., Connors, B.W., 1994. Synaptic physiology of horizontalafferents to layer I in slices of rat SI neocortex. J. Neurosci. 14,751–762.
Coghill, R.C., Talbot, J.D., Evans, A.C., 1994. Distributed processingof pain and vibration by the human brain. J. Neurosci. 14,4095–4108.
Dittman, J.S., Kreitzer, A.C., Regehr, W.G., 2000. Interplay betweenfacilitation, depression, and residual calcium at threepresynaptic terminals. J. Neurosci. 20, 1374–1385.
Donahue, R.R., LaGraize, S.C., Fuchs, P.N., 2001. Electrolytic lesionof the anterior cingulate cortex decreases inflammatory, butnot neuropathic nociceptive behavior in rats. Brain Res. 897,131–138.
Dostrovsky, J.O., Guilbaud, G., 1990. Nociceptive responses inmedial thalamus of the normal and arthritic rat. Pain 40,93–104.
Emptage, N.J., Reid, C.A., Fine, A., 2001. Calcium stores inhippocampal synaptic boutons mediate short-term plasticity,store-operated Ca2+ entry, and spontaneous transmitterrelease. Neuron 29, 197–208.
Foltz, E.L., White, L.E., 1968. The role of rostral cingulumotomy in“pain” relief. Int. J. Neurol. 6, 353–373.
Freeman, J.A., Nicholson, C., 1975. Experimental optimization ofcurrent source-density technique for anuran cerebellum. J.Neurophysiol. 38, 369–382.
Gemmell, C., O'Mara, S.M., 2002. Plasticity in the projection fromthe anterior thalamic nuclei to the anterior cingulate cortex inthe rat in vivo: paired-pulse facilitation, long-term potentiationand short-term depression. Neuroscience 109, 401–406.

114 B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
Gigg, J., Tan, A.M., Finch, D.M., 1992. Glutamatergic excitatoryresponses of anterior cingulate neurons to stimulation of themediodorsal thalamus and their regulation by GABA: an in vivoiontophoretic study. Cereb. Cortex 6, 477–684.
Gioanni, Y., Rougeot, C., Clarke, P.B., Lepouse, C., Thierry, A.M.,Vidal, C., 1999. Nicotinic receptors in the rat prefrontal cortex:increase in glutamate release and facilitation of mediodorsalthalamo-cortical transmission. Eur. J. Neurosci. 11, 18–30.
Gray, C.M., Maldonado, P.E., Wilson, M., McNaughton, B., 1995.Tetrodesmarkedly improve the reliability and yield of multiplesingle-unit isolation from multi-unit recordings in cat striatecortex. J. Neurosci. Methods 63, 43–54.
Greengard, P., Jen, J., Nairn, A.C., Stevens, C.F., 1991. Enhancementof the glutamate response by cAMP-dependent protein kinasein hippocampal neurons. Science 253, 1135–1138.
Groenewegen, H.J., 1988. Organization of the afferent connectionsof the mediodorsal thalamic nucleus in the rat, related to themediodorsal–prefrontal topography. Neuroscience 24, 379–431.
Guilbaud, G., Peschanski, M., Briand, A., Gautron, M., 1986. Theorganization of spinal pathways to ventrobasal thalamus in anexperimental model of pain (the arthritic rat). Anelectrophysiological study. Pain 26, 301–312.
Hedberg, T.G., Stanton, P.K., 1995. Long-term potentiation anddepression of synaptic transmission in rat posterior cingulatecortex. Brain Res. 670, 181–196.
Horikawa, K., Kinjo, N., Stanley, L.C., Powell, E.W., 1988.Topographic organization and collateralization of theprojections of the anterior and laterodorsal thalamic nuclei tocingulate areas 24 and 29 in the rat. Neurosci. Res. 6, 31–44.
Hsu, M.M., Shyu, B.C., 1997. Electrophysiological study of theconnection between medial thalamus and anterior cingulatecortex in the rat. NeuroReport 8, 2701–2707.
Izaki, Y., Takita, M., Nomura, M., 2001. Mousehippocampo-prefrontal paired-pulse facilitation andlong-term potentiation in vivo. NeuroReport 12, 1191–1193.
Johansen, J.P., Fields, H.L., 2004. Glutamatergic activation ofanterior cingulate cortex produces an aversive teaching signal.Nat. Neurosci. 7, 398–403.
Johansen, J.P., Fields, H.L., Manning, B.H., 2001. The affectivecomponent of pain in rodents: direct evidence for acontribution of the anterior cingulate cortex. Proc. Natl. Acad.Sci. 98, 8077–8082.
Jones, M.V., Westbrook, G.L., 1995. Desensitized states prolongGABAA channel responses to brief agonist pulses. Neuron 15,181–191.
Jones, A.K., Brown, W.D., Friston, K.J., Qi, L.Y., Frackowiak, R.S.,1991. Cortical and subcortical localization of response to painin man using positron emission tomography. Proc. R. Soc.Lond., B Biol. Sci. 244, 39–44.
Juergens, E., Eckhorn, R., 1999. Visual stimulation elicits lockedand induced gamma oscillations in monkey intracortical- andEEG-potentials, but not in human EEG. Exp. Brain Res. 129,247–259.
Kirischuk, S., Clements, J.D., Grantyn, R., 2002. Presynaptic andpostsynaptic mechanisms underlie paired pulse depression atsingle GABAergic boutons in rat collicular cultures. J. Physiol.543, 99–116.
Kirkwood, A., Dudek, S.M., Gold, J.T., 1993. Common forms ofsynaptic plasticity in the hippocampus and neocortex in vitro.Science 260, 1518–1521.
Koyama, T., Kato, K., Tanaka, Y.Z., Mikami, A., 2001. Anteriorcingulate activity during pain-avoidance and reward tasks inmonkeys. Neurosci. Res. 39, 421–430.
Krettek, J.E., Price, J.L., 1977. The cortical projections of themediodorsal nucleus and adjacent thalamic nuclei in the rat.J. Comp. Neurol. 171, 157–191.
Kung, J.C., Shyu, B.C., 2002. Potentiation of local field potentials inthe anterior cingulate cortex evoked by the stimulation of themedial thalamic nuclei in rats. Brain Res. 953, 37–44.
Kung, J.C., Su, N.M., Fan, R.J., Chai, S.C., Shyu, B.C., 2003.Contribution of the anterior cingulate cortex to laser-painconditioning in rats. Brain Res 970, 58–72.
Kuroda, M., Yokofujita, J., Murakami, K., 1998. An ultrastructuralstudy of the neural circuit between the prefrontal cortex andthe mediodorsal nucleus of the thalamus. Prog. Neurobiol. 54,417–458.
Logothetis, N., Oeltermann, A., 2001. Neurophysiologicalinvestigation of the basis of the fMRI signal. Nature 412,150–157.
Mansour, A., Khachaturian, H., Lewis, M.E., Akil, H., Watson, S.J.,1987. Autoradiographic differentiation of mu, delta, and kappaopioid receptors in the rat forebrain and midbrain. J. Neurosci.7, 2445–2464.
Mesulam,M.M., Mufson, E.J.,Wainer, B.H., Levey, A.I., 1983. Centralcholinergic pathways in the rat: an overview based on analternative nomenclature (Ch1-Ch6). Neuroscience 10,1185–1201.
Mitzdorf, U., 1985. Current source-density method and applicationin cat cerebral cortex: investigation of evoked potentials andEEG phenomena. Physiol. Rev. 65, 37–100.
Mitzdorf, U., 1987. Properties of the evoked potential generators:current source-density analysis of visually evoked potentials inthe cat cortex. Int. J. Neurosci. 33, 33–59.
Paxinos, G., Watson, C., 1998. The Rat Brain in StereotaxicCoordinates, 4th Edition. Academic Press, San Diego.
Pirot, S., Jay, T.M., Glowinski, J., Thierry, A.M., 1994. Anatomicaland electrophysiological evidence for an excitatory amino acidpathway from the thalamic mediodorsal nucleus to theprefrontal cortex in the rat. Eur. J. Neurosci. 6, 1225–1234.
Pirot, S., Glowinski, J., Thierry, A.M., 1996. Mediodorsal thalamicevoked responses in the rat prefrontal cortex: influence of themesocortical DA system. NeuroReport 7, 1437–1441.
Rainville, P., Duncan, G.H., Price, D.D., Carrier, B., Bushnell, M.C.,1997. Pain affect encoded in human anterior cingulate but notsomatosensory cortex. Science 277, 968–971.
Rios, M., Treede, R., Lee, J., Lenz, F.A., 1999. Direct evidence ofnociceptive input to human anterior cingulate gyrus andparasylvian cortex. Curr. Rev. Pain 3, 256–264.
Rozov, A., Burnashev, N., 1999. Polyamine-dependent facilitationof postsynaptic AMPA receptors counteracts paired-pulsedepression. Nature 401, 594–598.
Sah, P., Nicoll, R.A., 1991. Mechanisms underlying potentiation ofsynaptic transmission in rat anterior cingulate cortex in vitro.J. Physiol. 433, 615–630.
Santo, J.L., Arias, L.M., Barolat, G., Schwartzman, R.J., Grossman, K.,1990. Bilateral cingulotomy in the treatment of reflexsympathetic dystrophy. Pain 41, 55–59.
Sherma, T., 1973. Absence of cognitive deficits from bilateralcingulotomy for intractable pain in humans. Tex. Med. 69,79–82.
Shyu, B.C., Olausson, B., Rydenhag, B., 1989. Field potentialanalysis of the cortical projection of the central lateral nucleusin the cat. Acta Physiol. Scand. 137, 503–512.
Shyu, B.C., Kiritsy-Roy, J.A., Morrow, T.J., Casey, K.L., 1992.Neurophysiological, pharmacological and behavioral evidencefor medial thalamic mediation of cocaine-induceddopaminergic analgesia. Brain Res. 572, 216–223.
Shyu, B.C., Lin, C.Y., Sun, J.J., Chen, S.L., Chang, C., 2004. BOLDresponse of direct thalamic stimulation reveals functionalconnection of the medial thalamus and anterior cingulatecortex in the rat. Magn. Reson. Med. 52, 47–55.
Sikes, R.W., DeFrance, J.F., 1985. Cingulate cortex response toelectrical stimulation of the mediodorsal thalamic nucleus.Exp. Neurol. 89, 428–441.
Sikes, R.W., Vogt, B.A., 1992. Nociceptive neurons in area 24 ofrabbit cingulate cortex. J. Neurophysiol. 68, 1720–1732.
Steriade, M., 1999. Coherent oscillations and short-term plasticityin corticothalamic networks. TINS 22, 337–345.

115B R A I N R E S E A R C H 1 0 9 7 ( 2 0 0 6 ) 1 0 1 – 1 1 5
Swanson, L.W., Cowan, W.M., 1977. An autoradiographic studyof the organization of the efferent connections of thehippocampal formation in the rat. J. Comp. Neurol. 172,49–84.
Sylantyev, S.O., Lee, C.-M., Shyu, B.-C., 2004. Quantitativeassessment of the effect of the CNQX in the paired-pulsefacilitation in the anterior cingulate cortex. J. Neurosci.Methods 137, 207–214.
Sylantyev, S.O., Lee, C.-M., Shyu, B.-C., 2005. A parametricassessment of GABA antagonist effects on paired-pulsefacilitation in the rat anterior cingulate cortex. Neurosci. Res.52, 362–370.
Talbot, J.D., Marrett, S., Evans, A.C., Meyer, E., Bushnell, M.C.,Duncan, G.H., 1991. Multiple representations of pain in humancerebral cortex. Science 251, 1355–1358.
Talbot, J.D., Villemure, J.G., Bushnell, M.C., Duncan, G.H., 1995.Evaluation of pain perception after anterior capsulotomy: acase report, Somatosens. Mot. Res. 12, 115–126.
Tanaka, E., North, R.A., 1994. Opioid actions on rat anteriorcingulate cortex neurons in vitro. J. Neurosci. 14, 1106–1113.
Tsai, M.L., Kuo, C.C., Sun, W.Z., Yen, C.T., 2004. Differentialmorphine effects on short- and long-latency laser-evokedcortical responses in the rat. Pain 110, 665–674.
Vogt, B.A., Sikes, R.W., Vogt, L.J., 1993. Anterior cingulate cortexand the medial pain system. In: Vogt, B.A., Gabriel, M. (Eds.),Neurobiology of Cingulate Cortex and Limbic Thalamus: AComprehensive Handbook. Birkhauser, Boston, pp. 313–344.
Vogt, B.A., Miller, M.W., 1983. Cortical connections between ratcingulate cortex and visual, motor, and postsubicular cortices.J. Comp. Neurol. 216, 192–210.
Wang, J.H., Kelly, P.T., 1997. Attenuation of paired-pulse
facilitation associated with synaptic potentiation mediated bypostsynaptic mechanisms. J. Neurophysiol. 78, 2707–2716.
Wang, C.C., Shyu, B.C, 2004. Differential projections from themediodorsal and centrolateral thalamic nuclei to the frontalcortex in rats. Brain Res. 995, 226–235.
Wei, F., Zhuo, M., 2001. Potentiation of synaptic responses in theanterior cingulate cortex following digital amputation in rat.J. Physiol. (Lond.) 532, 823–833.
Wei, F., Li, P., Zhuo, M., 1999. Loss of synaptic depression inmammalian anterior cingulate cortex after amputation.J. Neurosci. 19, 9346–9354.
Wilkinson, H.A., Davidson, K.M., Davidson, R.I., 1999. Bilateralanterior cingulotomy for chronic noncancer pain.Neurosurgery 45, 1129–1136.
Wong, E.T., Gunes, S., Gaughan, E., Patt, R.B., Ginsberg, L.E.,Hassenbusch, S.J., Payne, R., 1997. Palliation of intractablecancer pain by MRI-guided cingulotomy. Clin. J. Pain 13,260–263.
Woolf, N.J., Eckenstein, F., Butcher, L.L., 1984. Cholinergic systemsin the rat brain: I. projections to the limbic telencephalon. BrainRes. Bull. 6, 751–784.
Yamamura, H., Iwata, K., Tsuboi, Y., Toda, K., Kitajima, K.,Shimizu, N., Nomura, H., Hibiya, J., Fujita, S., Sumino, R., 1996.Morphological and electrophysiological properties of ACCxnociceptive neurons in rats. Brain Res. 735, 83–92.
Zinebi, F., Russell, R.T., McKernan, M., Shinnick-Gallagher, P.,2001. Comparison of paired-pulse facilitation of AMPA andNMDA synaptic currents in the lateral amygdala. Synapse 42,115–127.
Zucker, R.S, Regehr, W.G., 2002. Short-term synaptic plasticity.Annu. Rev. Physiol. 64, 355–405.
Copyright © 2022 FDOKUMEN




















