
Neuropeptide Y immunoreactivity in the cat claustrum: A light- and electron-microscopic...
13
Neuropeptide Y immunoreactivity in the cat claustrum: A light- and electron-microscopic investigation D.V. Hinova-Palova a , B. Landzhov a, *, E. Dzhambazova b , M. Minkov c , L. Edelstein d , L. Malinova a , A. Paloff a , W. Ovtscharoff a a Department of Anatomy, Histology and Embryology, Medical University of Sofia, 1431 Sofia, Bulgaria b Department of Chemistry, Biochemistry, Physiology and Pathophysiology, Sofia University ‘‘St. Kliment Ohridski’’, 1407 Sofia, Bulgaria c Department of Anatomy, Histology and Embryology, Medical University of Varna, 9002 Varna, Bulgaria d Medimark Corporation, P.O. Box 2316, Del Mar, CA 92014, USA 1. Introduction The claustrum is a telencephalic nucleus in the mammalian brain that has long been known as a site of extensive reciprocal heterosensory convergence (Olson and Graybiel, 1980; Ashwell et al., 2004; Hinova-Palova et al., 2007, 2008). Typically, it is subdivided into the dorsal claustrum (or ‘‘insular claustrum’’) and ventral claustrum (or ‘‘endopiriform nucleus’’) (Guirado et al., 2003; Edelstein and Denaro, 2004a; Ashwell et al., 2004). The dorsal claustrum is located deep to the insular cortex and is extensively connected with most if not all neocortex (Druga, 1966a,b; Otellin and Makarov, 1972; Kunzle, 1975, 1978; Norita, 1977; Riche and Lanoir, 1978; Olson and Graybiel, 1980; Carey et al., 1980; Hinova-Palova et al., 1980a,b; Hinova-Palova, 1981; Hinova-Palova and Paloff, 1982, 1984; Carey and Neal, 1985; Edelstein, 1986; Neal et al., 1986; Sloniewski et al., 1986; Tanne- Gariepy et al., 2002; Ashwell et al., 2004). The ventral claustrum is situated beneath the piriform cortex; its interconnections with prepiriform and entorhinal cortex are well documented (Druga, 1966a,b, 1971; Sherk, 1986; Witter et al., 1988; Dinopoulos et al., 1992). From a phylogenetic perspective, the dimensions of the claustrum vary greatly. In lower mammals such as the mouse and rat it is a small and discrete nucleus ventrolateral to the Journal of Chemical Neuroanatomy 61–62 (2014) 107–119 A R T I C L E I N F O Article history: Received 19 May 2014 Received in revised form 15 August 2014 Accepted 16 August 2014 Available online 23 August 2014 Keywords: Cat Claustrum Neuropeptide Y Light microscopy Electron microscopy Ultrastructure A B S T R A C T The claustrum is a telencephalic nucleus located ventrolateral to the basal ganglia in the mammalian brain. It has an extensive reciprocal connectivity with most if not all of the cerebral cortex, in particular, primary sensory areas. However, despite renewed and growing interest amongst investigators, there remains a paucity of data concerning its peptidergic profile. The aim of the present study was to examine the presence, morphology, distribution and ultrastructure of neuropeptide Y-immunoreactive (NPY-ir) neurons and fibers in the claustrum of the cat. Ten adult healthy cats from both sexes were used. All animals received human and ethical treatment in accordance with the Principles of Laboratory Animal Care. Subjects were irreversibly anesthetized and transcardially perfused with fixative solution containing glutaraldehyde and paraformaldehyde. Brains were promptly removed, postfixed and sectioned. Slices were incubated with polyclonal anti-NPY antibodies according to the standard avidin– biotin–peroxidase complex method adopted by our Department of Anatomy, Histology and Embryology. NPY-ir neurons and fibers were found to be diffusely distributed throughout the claustrum, with no obvious topographic or functional patterning other than larger numbers in its central/broadest part (stereotaxic planes A12–A16). Neurons were generally classified by diameter into three sizes: small (under 17 mm), medium (17–25 mm) and large (over 25 mm). Staining density is varied with some neurons appearing darker than others. At the electron-microscopic level NPY immunoproduct was observed within neurons, dendrites and terminal boutons, each differing relative to their ultrastructural attributes. Two types of NPY-ir synaptic boutons were found. Lastly, it is of interest to note that gender- specific differences were not observed. ß 2014 Elsevier B.V. All rights reserved. * Corresponding author at: Medical University of Sofia, Faculty of Medicine, Department of Anatomy, Histology and Embryology, 2 Zdrave Str., 1431 Sofia, Bulgaria. Tel.: +359 29172608; fax: +359 29172601. E-mail address: [email protected] (B. Landzhov). Contents lists available at ScienceDirect Journal of Chemical Neuroanatomy jo ur n al ho mep ag e: www .elsevier .c om /lo cate/jc h emn eu http://dx.doi.org/10.1016/j.jchemneu.2014.08.007 0891-0618/ß 2014 Elsevier B.V. All rights reserved.
Transcript of Neuropeptide Y immunoreactivity in the cat claustrum: A light- and electron-microscopic...

Journal of Chemical Neuroanatomy 61–62 (2014) 107–119
Neuropeptide Y immunoreactivity in the cat claustrum:A light- and electron-microscopic investigation
D.V. Hinova-Palova a, B. Landzhov a,*, E. Dzhambazova b, M. Minkov c,L. Edelstein d, L. Malinova a, A. Paloff a, W. Ovtscharoff a
a Department of Anatomy, Histology and Embryology, Medical University of Sofia, 1431 Sofia, Bulgariab Department of Chemistry, Biochemistry, Physiology and Pathophysiology, Sofia University ‘‘St. Kliment Ohridski’’, 1407 Sofia, Bulgariac Department of Anatomy, Histology and Embryology, Medical University of Varna, 9002 Varna, Bulgariad Medimark Corporation, P.O. Box 2316, Del Mar, CA 92014, USA
A R T I C L E I N F O
Article history:
Received 19 May 2014
Received in revised form 15 August 2014
Accepted 16 August 2014
Available online 23 August 2014
Keywords:
Cat
Claustrum
Neuropeptide Y
Light microscopy
Electron microscopy
Ultrastructure
A B S T R A C T
The claustrum is a telencephalic nucleus located ventrolateral to the basal ganglia in the mammalian
brain. It has an extensive reciprocal connectivity with most if not all of the cerebral cortex, in particular,
primary sensory areas. However, despite renewed and growing interest amongst investigators, there
remains a paucity of data concerning its peptidergic profile. The aim of the present study was to examine
the presence, morphology, distribution and ultrastructure of neuropeptide Y-immunoreactive (NPY-ir)
neurons and fibers in the claustrum of the cat. Ten adult healthy cats from both sexes were used. All
animals received human and ethical treatment in accordance with the Principles of Laboratory Animal
Care. Subjects were irreversibly anesthetized and transcardially perfused with fixative solution
containing glutaraldehyde and paraformaldehyde. Brains were promptly removed, postfixed and
sectioned. Slices were incubated with polyclonal anti-NPY antibodies according to the standard avidin–
biotin–peroxidase complex method adopted by our Department of Anatomy, Histology and Embryology.
NPY-ir neurons and fibers were found to be diffusely distributed throughout the claustrum, with no
obvious topographic or functional patterning other than larger numbers in its central/broadest part
(stereotaxic planes A12–A16). Neurons were generally classified by diameter into three sizes: small
(under 17 mm), medium (17–25 mm) and large (over 25 mm). Staining density is varied with some
neurons appearing darker than others. At the electron-microscopic level NPY immunoproduct was
observed within neurons, dendrites and terminal boutons, each differing relative to their ultrastructural
attributes. Two types of NPY-ir synaptic boutons were found. Lastly, it is of interest to note that gender-
specific differences were not observed.
� 2014 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Journal of Chemical Neuroanatomy
jo ur n al ho mep ag e: www .e lsev ier . c om / lo cate / jc h emn eu
1. Introduction
The claustrum is a telencephalic nucleus in the mammalianbrain that has long been known as a site of extensive reciprocalheterosensory convergence (Olson and Graybiel, 1980; Ashwellet al., 2004; Hinova-Palova et al., 2007, 2008). Typically, it issubdivided into the dorsal claustrum (or ‘‘insular claustrum’’) andventral claustrum (or ‘‘endopiriform nucleus’’) (Guirado et al.,2003; Edelstein and Denaro, 2004a; Ashwell et al., 2004). The
* Corresponding author at: Medical University of Sofia, Faculty of Medicine,
Department of Anatomy, Histology and Embryology, 2 Zdrave Str., 1431 Sofia,
Bulgaria. Tel.: +359 29172608; fax: +359 29172601.
E-mail address: [email protected] (B. Landzhov).
http://dx.doi.org/10.1016/j.jchemneu.2014.08.007
0891-0618/� 2014 Elsevier B.V. All rights reserved.
dorsal claustrum is located deep to the insular cortex and isextensively connected with most if not all neocortex (Druga,1966a,b; Otellin and Makarov, 1972; Kunzle, 1975, 1978; Norita,1977; Riche and Lanoir, 1978; Olson and Graybiel, 1980; Careyet al., 1980; Hinova-Palova et al., 1980a,b; Hinova-Palova, 1981;Hinova-Palova and Paloff, 1982, 1984; Carey and Neal, 1985;Edelstein, 1986; Neal et al., 1986; Sloniewski et al., 1986; Tanne-Gariepy et al., 2002; Ashwell et al., 2004). The ventral claustrum issituated beneath the piriform cortex; its interconnections withprepiriform and entorhinal cortex are well documented (Druga,1966a,b, 1971; Sherk, 1986; Witter et al., 1988; Dinopoulos et al.,1992).
From a phylogenetic perspective, the dimensions of theclaustrum vary greatly. In lower mammals such as the mouseand rat it is a small and discrete nucleus ventrolateral to the

D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119108
caudoputamen. In the cat it is a relatively large structure thatrivals, or even exceeds, the size of the putamen. In primates it is athin gray slab varying in thickness up to several millimeters,bounded laterally by the extreme capsule and medially by theexternal capsule (De Vries, 1910; Landau, 1923; Berlucchi, 1927;Loo, 1931; Brockhaus, 1940; Macchi, 1984; Rae, 1954; Stelmasiak,1955; Berke, 1960; Pilleri, 1961, 1962; Filimonoff, 1966; Druga,1974, 1975; Zilles and Zilles, 1980; Paxinos and Watson, 1989;Kowianski et al., 2004).
We have long been interested in the claustrum’s cytoarchi-tecture, ultrastructure, immunocytochemistry and connectivity(Hinova-Palova et al., 1979, 1980a,b, 1988, 1997, 2001, 2007, 2008,2012, 2013a,b,c; Hinova-Palova, 1981, 1986; Hinova-Palova andUsunoff, 1981; Hinova-Palova and Christova, 1988; Hinova-Palovaand Braak, 1993; Edelstein et al., 2011a,b, 2012a,b). Theclaustrum’s syncytium of GABAergic interneurons and glutama-tergic projection neurons, as well as its heterosensory reciprocalconnectivity, leads one to ponder its functional interplay with therest of the brain.
Neuropeptide Y is a novel 36-amino acid peptide and one of thekey regulators of food intake, circadian rhythm and reproduction inmany mammalian species, including humans. Originally isolatedfrom the porcine hypothalamus (Tatemoto, 1982; Tatemoto et al.,1982), NPY is widely distributed throughout the central andautonomic nervous system of various vertebrates, such asmammals (Adrian et al., 1983; Allen et al., 1983; Whale andAlbus, 1984; Emson and De Quidt, 1984; Smith et al., 1985;Chronwall et al., 1985; Sasek and Elde, 1985; de Quidt and Emson,1986; Dumont et al., 1992; Krivukuca et al., 2010), amphians(Danger et al., 1985; Perroteau et al., 1988), teleosts (Pontet et al.,1989; Danger et al., 1991), elasmobranchs (Vallarino et al., 1988;Chiba and Honma, 1992), cyclostomes (Chiba et al., 1993) and thewhite sturgeon (Chiba and Honma, 1994). With respect tophylogenetic comparisons specific to the neurochemical architec-ture of the claustrum, a major focus of our lab, there are but ahandful of studies on NPY immunoreactivity, namely, in thesquirrel monkey (Smith et al., 1985), the cat (Edelstein et al.,2011b) and humans (Edelstein et al., 2010).
The aim of the present study is to make evident the features andcharacteristics of NPY-immunoreactive neurons and fibers in thecat claustrum at the light- and electron-microscopic level, as wellas to provide a description of their distribution.
2. Material and methods
2.1. Perfusion protocol
Ten adult healthy cats from both sexes with average weight 2.3 kg (from 1.9 to
3.0 kg) were used. Our perfusion protocol was previously described in detail
(Papantchev, 2008). In brief, all animals were deeply and irreversibly anesthetized
with an intraperitoneal injection of Urethan (40 mg/kg) and transcardially perfused
with 500 ml of heparinized saline, followed by 3000 ml of phosphate buffered
saline (PBS; pH 7.4) containing 2.5% glutaraldehyde and 4% paraformaldehyde.
After 2 h the brains were removed and postfixed in the same perfusate for
additional 2 h. The portion of each cerebral hemisphere containing the claustrum
was removed, dissected and cut into a total of fourteen blocks. Each block was then
sectioned in the coronal plane at 40–80 mm on a Vibratome (Technical Products
International, St. Louis, MO, USA). The claustrum was examined in accordance with
the stereotaxic atlas of Reinoso-Suarez (1961).
2.2. Immunohistochemistry
Our protocol for the immunohistochemical visualization of NP immunoreactivity
was previously described in detail (Paloff et al., 2004). In brief, all coronal sections as
described above were treated with sodium borohydride for 45 min followed by
three consecutive 2-min rinses in 0.01 M PBS. This was immediately followed by
30 min in a solution of 1% bovine serum albumin (BSA) and overnight incubation in
a 1:1000 solution of a polyclonal anti-NPY-antibody (Sigma, St. Louis, MO, USA).
Afterwards, sections were given three consecutive 2-min rinses in 0.01 M PBS,
incubated for 20 min in 1% BSA in 0.01 M PBS and followed by a 2-h incubation in
1:500 biotinylated anti-mouse IgG (Vector, Burlingame, CA, USA). This was then
followed by three consecutive 2-min rinses in 0.01 M PBS, with sections incubated
for 1-h in a solution of avidin–biotin–peroxidase complex (Vector, Burlingame, CA,
USA). All incubations were carried out on a shaker table at room temperature. For
controls, fifteen sections were incubated as described but with the omission of the
primary or secondary antibody. All controls were negative.
After this 1-h incubation a new series of rinses were performed according to a
protocol used in our Department (Paloff et al., 2004; Hinova-Palova et al., 2007) –
first in 0.01 M PBS and then in Tris buffer, pH 7.6 – which preceded the visualization
of peroxidase activity using H2O2 and 3,30-diaminobenzidine as substrates. The
protocol continued with three 2-min rinses in Tris buffer followed by another three
2-min rinses in phosphate buffer. Afterwards, some of the sections was processed
for electron microscopy. Sections were postfixed for 1-h with 1% OsO4 in phosphate
buffer, dehydrated in graded series of ethanol and flat-embedded in Durcupan
(Fluka, Buchs, Switzerland) between plastic sheets (Paloff et al., 2004; Hinova-
Palova et al., 2007). Sections were trimmed-out under a dissecting microscope and
cemented to epoxy blanks. Thin sections were cut with an ultramicrotome (LKB,
Stockholm-Bromma, Sweden), counterstained with uranyl acetate and lead citrate
according to a standard protocol used in our Department (Paloff et al., 2004;
Hinova-Palova et al., 2007) and examined with an Hitachi H-500 electron
microscope (Hitachi, Tokyo, Japan). A light microscope (Olympus, Tokyo, Japan)
was used for the low-power examination of NPY-ir neurons.
One-hundred neuronal perikarya were identified and measured via electron
micrographs. The NPY-ir neurons (or neuronal groups) were first identified by light
microscope and then serial sections were performed. All consecutive measure-
ments were performed on sections where a prominent nucleolus was present. The
ratio of the mean nuclear diameter to the mean diameter of the perikaryon was
calculated as the nucleocytoplasmic ratio (Paloff et al., 2004; Hinova-Palova et al.,
2007) using an ultrastructural size calculator (Ted Pella, Tustin, CA, USA). In
addition, a total of 200 NPY-ir terminal boutons were analyzed in order to evaluate
the synaptic morphology and characteristics of the vesicular populations.
2.3. Morphometric analysis
Our morphometric analysis protocol was previously detailed (Hinova-Palova
et al., 2007, 2008). Briefly, stereotaxic planes from A10 to A19 (Reinoso-Suarez,
1961) were examined using an image analyzer (CUE-2, Olympus America, Center
Valley, PA, USA) and a 40� objective. A total of 42 slides (3 randomly selected slides
from each of the 14 tissue blocks examined) per stereotaxic plane were studied. The
number of NPY-ir neurons seen in each section were counted and stored in the
database. The number of NPY-ir neurons was calculated for each plane as an average
of the number of counted neurons from all analyzed sections per plane. The number
of NPY-ir neurons counted was presented as a percentage of all NPY-ir neurons seen
in the entire claustrum. Afterwards, standard planar morphometry and linear
analysis (i.e. line length and width) were performed. The maximum diameter of 500
neurons was measured, and the cells were divided into groups. A mean of the
maximum and minimum diameter of all neurons in each group was then calculated.
For all measurements, only those neurons with clearly visible nuclei were used.
Explanatory marks were added to all images using Adobe Photoshop CS3.
3. Results
3.1. Light microscopy
Neurons and fibers immunopositive for NPY-ir were foundthroughout the claustrum, from rostral to caudal planes (Figs. 1and 2). In general, the NPY-ir neurons were typically stained withimmunoproduct visible in the cell cytoplasm and processes, whilethe nucleus remained free of label (Fig. 3). Labeling intensity,however, varied. Some neurons were lightly stained (Fig. 4), whilein others the staining was so dense that they appeared as if theywere silver impregnated (Fig. 5). Immunopositive neurons werealso found proximal to and within the external and extremecapsules (Fig. 6).
Although NPY-ir neurons were seen throughout the claustrum,their distribution displayed a lack of uniformity. Based on ourmorphometric analysis, approximately 70% of labeled cells werelocated in stereotaxic planes A12–A16. Of these cells, the majorityappeared clustered within the claustrum’s central triangle region(planes A13–A15; Fig. 7), with but a handful found proximal to theexternal and extreme capsules. Most commonly, the neurons weresituated parallel to the fiber tracts, though some were positioned ina perpendicular fashion. Moving caudally, the number of NPY-irneurons gradually diminished (Fig. 8), and at the level ofstereotaxic planes A10–A11 only 15% of all counted NPY-irneurons were identified. Approximately 10% of all NPY-ir neurons

Fig. 1. Coronal section through claustrum in different planes from rostral to caudal:
(A) 19 mm in front of the interaural line (arrows delineate claustrum). Nissl staining
20�; (B) 15 mm in front of the interaural line (arrows delineate claustrum). Nissl
staining 20�; (C) 11 mm in front of the interaural line (arrows delineate claustrum).
Nissl staining 20�.
Fig. 2. Schematic drawings of 5 coronal sections through the cat claustrum showing
the distribution of NPY-positive neurons (black dots) from rostral to caudal planes.
Cn – caudate nucleus, Ic – internal capsule, Put – putamen, Sp – septum pellucidum,
Pal – pallidum (globus pallidus), Amg – amygdala, Th – thalamus, Ac – anterior
commissure, Hy – hypothalamus, Ent – entopeduncular nucleus, NII – second
cranial nerve (optic).
Fig. 3. NPY-immunoproduct is clearly visible in the cell cytoplasm and processes,
with the nucleus stain-free. Scale bar = 50 mm.
D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119 109
were identified in the more rostral stereotaxic planes A17–A18.Finally, the rostral pole of the claustrum contained no more than5% of NPY-ir neurons.
The NPY-ir neurons differed in size and were generallysubdivided into large (over 25 mm), medium (17–25 mm), small(under 17 mm) and dwarf cells (under 8 mm). They were furthercategorized into spiny and aspiny types relative to their dendriticarchitecture. The various shapes of NPY-ir neurons seen was notunlike what has previously been noted in similar studies (perhapsowing to the likelihood that most if not all claustral neurons co-express proteins and peptides of one type or another): polygonal(multipolar), triangular, fusiform (bipolar), globular, elliptical, ovaland round.
3.1.1. Large spiny neurons
Large spiny NPY-ir neurons were classified as being greater than25 mm in diameter, with polygonal (multipolar), triangular,fusiform, or globular perikarya (Figs. 9 and 10). They were seengiving rise to as many as five thick dendritic processes, each withmultiple and multidirectional branches. These branches weretypically quite long and could sometimes be followed up to700 mm from the cell body. The secondary and tertiary dendriteswere covered with spines. Axons with highly visible hillocks andinitial segments were frequently seen emanating from these cells.The intensity of immunostaining varied widely – from highlydense (i.e. impregnation-like) to weakly stained. Typically, thenuclei were non-immunoreactive.
3.1.2. Medium spiny neurons
Medium spiny NPY-ir neurons measured from 18 to 24 mm indiameter with multipolar, fusiform, or oval perikarya. They usuallyhad long bifurcated dendrites, each covered with well-developedspines (Fig. 11).

Fig. 4. Large NPY-positive lightly stained neuron. Scale bar = 50 mm.
Fig. 5. Large spiny NPY-positive darkly stained neuron with long dendrites. Scale
bar = 50 mm.
Fig. 6. Large fusiform NPY-positive neuron close to external capsule. Scale
bar = 50 mm.
Fig. 7. Low magnification of triangular part of claustrum showing the distribution of
NPY-positive neurons. Scale bar = 1 mm.
Fig. 8. Low magnification of the caudal part of claustrum showing a lesser number
of NPY-positive neurons. Scale bar = 1 mm.
Fig. 9. Large spiny NPY-positive neuron with fusiform cell body. Scale bar = 50 mm.
D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119110

Fig. 10. Large spiny pear-shaped NPY-positive neuron. Scale bar = 50 mm.
Fig. 11. Two medium spiny NPY-positive neurons with long bifurcated dendrites
rich with spines. Scale bar = 50 mm.
Fig. 12. Small spiny NPY-positive neurons with long spiny dendrites. Scale
bar = 50 mm.
Fig. 13. Large aspiny NPY-positive neuron with long prominent bulbous dendrites
(arrows show the varicosities) Scale bar = 50 mm.
Fig. 14. Medium aspiny multipolar NPY-positive neuron with varicose dendrites
(arrows). Scale bar = 50 mm.
D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119 111
3.1.3. Small spiny neurons
Small spiny NPY-ir neurons measured from 15 to 17 mm indiameter and were typically oval or fusiform in shape with two tofour long and thin dendrites covered with spines (Fig. 12).
3.1.4. Large aspiny neurons
Large aspiny NPY-ir neurons were similar to the large spinyneurons. These cells also averaged over 25 mm in diameter. Theirperikarya were multipolar, triangular, fusiform, or oval in shape(Fig. 13), each with as many as five dendritic processes which –unlike the large spiny neurons – were rarely seen to branch.
3.1.5. Medium aspiny neurons
Medium aspiny NPY-ir neurons ranged from 18 to 23 mm indiameter. Typically, they had three or four dendrites (Fig. 14), withthe primary dendrites being smooth and normally non-branching,becoming progressively thinner with irregularly spaced varicosi-ties along their lengths.
3.1.6. Small aspiny neurons
Small aspiny NPY-ir neurons were typically 14–17 mm indiameter (Fig. 15) with predominantly oval or slightly ellipticalperikarya and typically two or three thin, varicose, non-branchingdendrites.
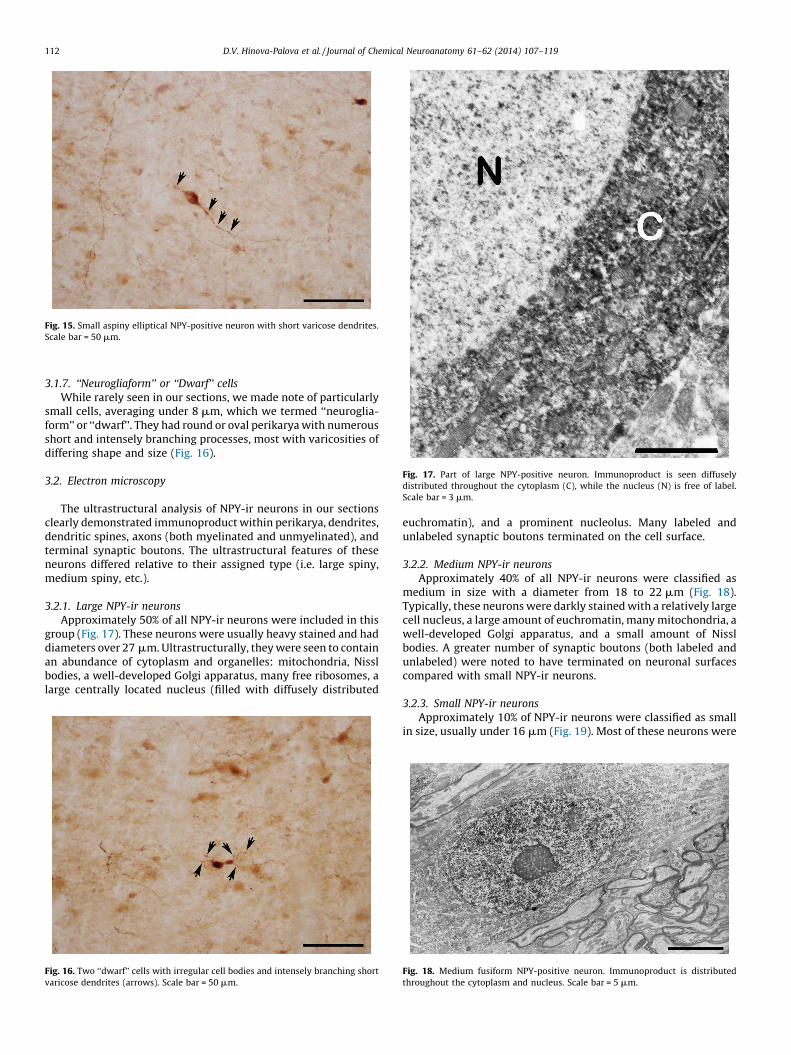
Fig. 15. Small aspiny elliptical NPY-positive neuron with short varicose dendrites.
Scale bar = 50 mm.
Fig. 17. Part of large NPY-positive neuron. Immunoproduct is seen diffusely
distributed throughout the cytoplasm (C), while the nucleus (N) is free of label.
Scale bar = 3 mm.
D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119112
3.1.7. ‘‘Neurogliaform’’ or ‘‘Dwarf’’ cells
While rarely seen in our sections, we made note of particularlysmall cells, averaging under 8 mm, which we termed ‘‘neuroglia-form’’ or ‘‘dwarf’’. They had round or oval perikarya with numerousshort and intensely branching processes, most with varicosities ofdiffering shape and size (Fig. 16).
3.2. Electron microscopy
The ultrastructural analysis of NPY-ir neurons in our sectionsclearly demonstrated immunoproduct within perikarya, dendrites,dendritic spines, axons (both myelinated and unmyelinated), andterminal synaptic boutons. The ultrastructural features of theseneurons differed relative to their assigned type (i.e. large spiny,medium spiny, etc.).
3.2.1. Large NPY-ir neurons
Approximately 50% of all NPY-ir neurons were included in thisgroup (Fig. 17). These neurons were usually heavy stained and haddiameters over 27 mm. Ultrastructurally, they were seen to containan abundance of cytoplasm and organelles: mitochondria, Nisslbodies, a well-developed Golgi apparatus, many free ribosomes, alarge centrally located nucleus (filled with diffusely distributed
Fig. 16. Two ‘‘dwarf’’ cells with irregular cell bodies and intensely branching short
varicose dendrites (arrows). Scale bar = 50 mm.
euchromatin), and a prominent nucleolus. Many labeled andunlabeled synaptic boutons terminated on the cell surface.
3.2.2. Medium NPY-ir neurons
Approximately 40% of all NPY-ir neurons were classified asmedium in size with a diameter from 18 to 22 mm (Fig. 18).Typically, these neurons were darkly stained with a relatively largecell nucleus, a large amount of euchromatin, many mitochondria, awell-developed Golgi apparatus, and a small amount of Nisslbodies. A greater number of synaptic boutons (both labeled andunlabeled) were noted to have terminated on neuronal surfacescompared with small NPY-ir neurons.
3.2.3. Small NPY-ir neurons
Approximately 10% of NPY-ir neurons were classified as smallin size, usually under 16 mm (Fig. 19). Most of these neurons were
Fig. 18. Medium fusiform NPY-positive neuron. Immunoproduct is distributed
throughout the cytoplasm and nucleus. Scale bar = 5 mm.
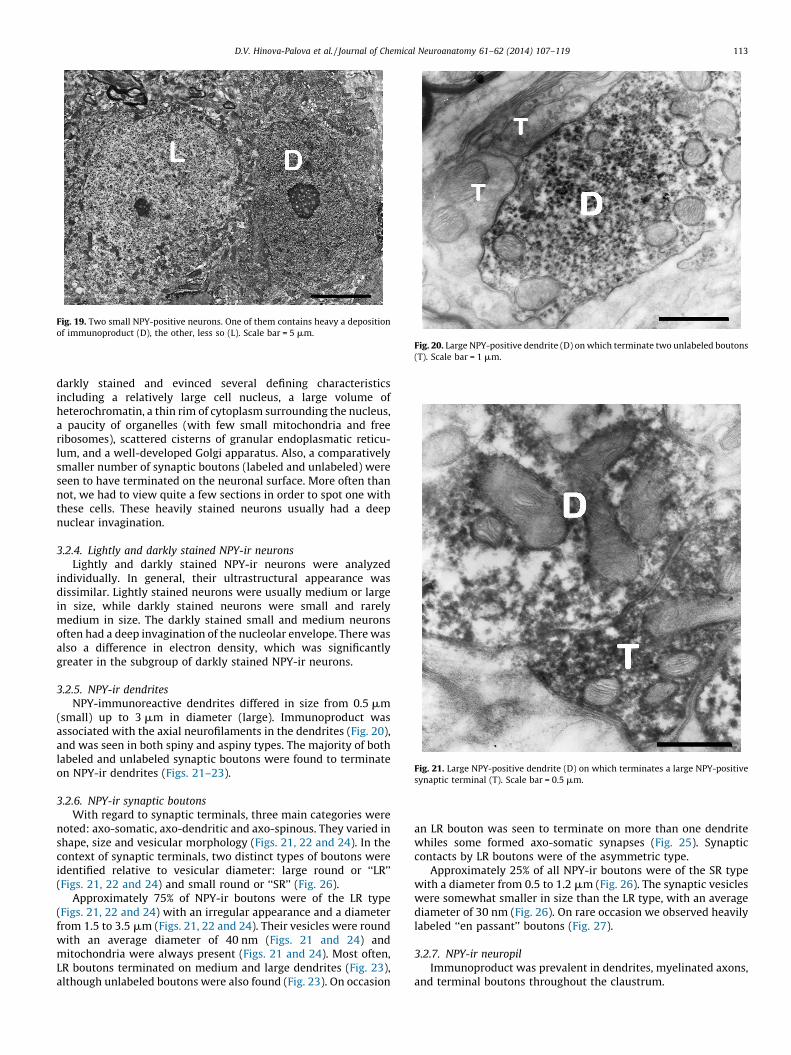
Fig. 19. Two small NPY-positive neurons. One of them contains heavy a deposition
of immunoproduct (D), the other, less so (L). Scale bar = 5 mm.
Fig. 20. Large NPY-positive dendrite (D) on which terminate two unlabeled boutons
(T). Scale bar = 1 mm.
Fig. 21. Large NPY-positive dendrite (D) on which terminates a large NPY-positive
synaptic terminal (T). Scale bar = 0.5 mm.
D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119 113
darkly stained and evinced several defining characteristicsincluding a relatively large cell nucleus, a large volume ofheterochromatin, a thin rim of cytoplasm surrounding the nucleus,a paucity of organelles (with few small mitochondria and freeribosomes), scattered cisterns of granular endoplasmatic reticu-lum, and a well-developed Golgi apparatus. Also, a comparativelysmaller number of synaptic boutons (labeled and unlabeled) wereseen to have terminated on the neuronal surface. More often thannot, we had to view quite a few sections in order to spot one withthese cells. These heavily stained neurons usually had a deepnuclear invagination.
3.2.4. Lightly and darkly stained NPY-ir neurons
Lightly and darkly stained NPY-ir neurons were analyzedindividually. In general, their ultrastructural appearance wasdissimilar. Lightly stained neurons were usually medium or largein size, while darkly stained neurons were small and rarelymedium in size. The darkly stained small and medium neuronsoften had a deep invagination of the nucleolar envelope. There wasalso a difference in electron density, which was significantlygreater in the subgroup of darkly stained NPY-ir neurons.
3.2.5. NPY-ir dendrites
NPY-immunoreactive dendrites differed in size from 0.5 mm(small) up to 3 mm in diameter (large). Immunoproduct wasassociated with the axial neurofilaments in the dendrites (Fig. 20),and was seen in both spiny and aspiny types. The majority of bothlabeled and unlabeled synaptic boutons were found to terminateon NPY-ir dendrites (Figs. 21–23).
3.2.6. NPY-ir synaptic boutons
With regard to synaptic terminals, three main categories werenoted: axo-somatic, axo-dendritic and axo-spinous. They varied inshape, size and vesicular morphology (Figs. 21, 22 and 24). In thecontext of synaptic terminals, two distinct types of boutons wereidentified relative to vesicular diameter: large round or ‘‘LR’’(Figs. 21, 22 and 24) and small round or ‘‘SR’’ (Fig. 26).
Approximately 75% of NPY-ir boutons were of the LR type(Figs. 21, 22 and 24) with an irregular appearance and a diameterfrom 1.5 to 3.5 mm (Figs. 21, 22 and 24). Their vesicles were roundwith an average diameter of 40 nm (Figs. 21 and 24) andmitochondria were always present (Figs. 21 and 24). Most often,LR boutons terminated on medium and large dendrites (Fig. 23),although unlabeled boutons were also found (Fig. 23). On occasion
an LR bouton was seen to terminate on more than one dendritewhiles some formed axo-somatic synapses (Fig. 25). Synapticcontacts by LR boutons were of the asymmetric type.
Approximately 25% of all NPY-ir boutons were of the SR typewith a diameter from 0.5 to 1.2 mm (Fig. 26). The synaptic vesicleswere somewhat smaller in size than the LR type, with an averagediameter of 30 nm (Fig. 26). On rare occasion we observed heavilylabeled ‘‘en passant’’ boutons (Fig. 27).
3.2.7. NPY-ir neuropil
Immunoproduct was prevalent in dendrites, myelinated axons,and terminal boutons throughout the claustrum.
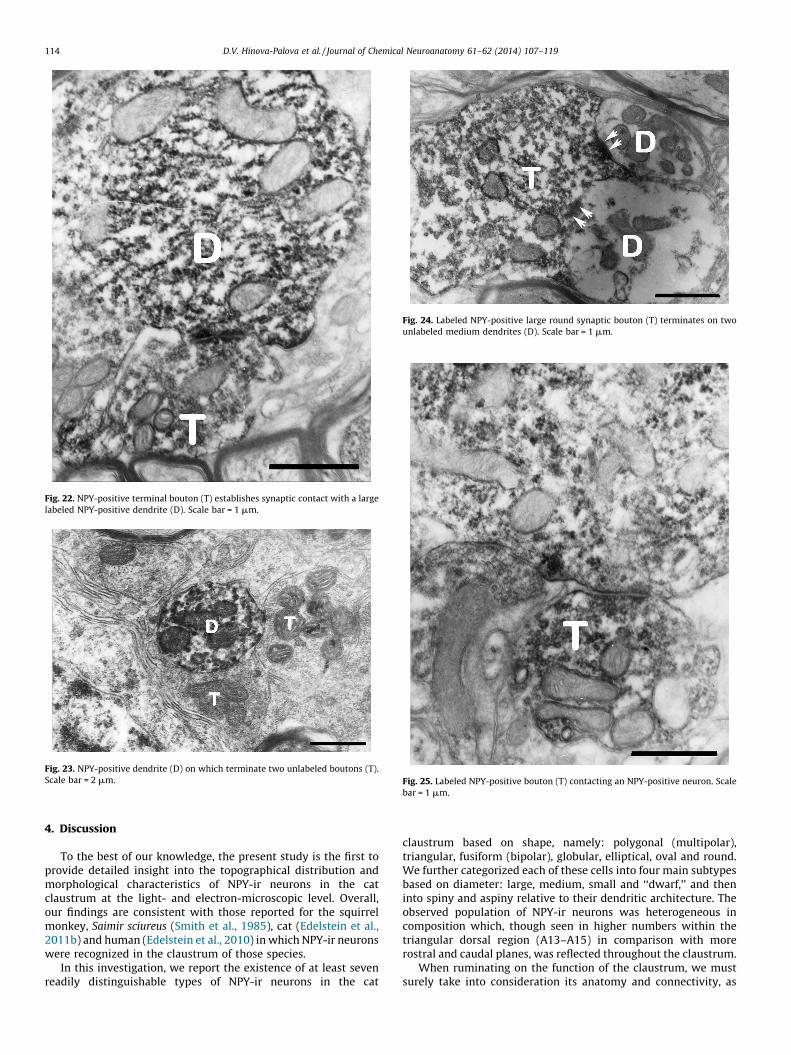
Fig. 22. NPY-positive terminal bouton (T) establishes synaptic contact with a large
labeled NPY-positive dendrite (D). Scale bar = 1 mm.
Fig. 23. NPY-positive dendrite (D) on which terminate two unlabeled boutons (T).
Scale bar = 2 mm.
Fig. 24. Labeled NPY-positive large round synaptic bouton (T) terminates on two
unlabeled medium dendrites (D). Scale bar = 1 mm.
Fig. 25. Labeled NPY-positive bouton (T) contacting an NPY-positive neuron. Scale
bar = 1 mm.
D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119114
4. Discussion
To the best of our knowledge, the present study is the first toprovide detailed insight into the topographical distribution andmorphological characteristics of NPY-ir neurons in the catclaustrum at the light- and electron-microscopic level. Overall,our findings are consistent with those reported for the squirrelmonkey, Saimir sciureus (Smith et al., 1985), cat (Edelstein et al.,2011b) and human (Edelstein et al., 2010) in which NPY-ir neuronswere recognized in the claustrum of those species.
In this investigation, we report the existence of at least sevenreadily distinguishable types of NPY-ir neurons in the cat
claustrum based on shape, namely: polygonal (multipolar),triangular, fusiform (bipolar), globular, elliptical, oval and round.We further categorized each of these cells into four main subtypesbased on diameter: large, medium, small and ‘‘dwarf,’’ and theninto spiny and aspiny relative to their dendritic architecture. Theobserved population of NPY-ir neurons was heterogeneous incomposition which, though seen in higher numbers within thetriangular dorsal region (A13–A15) in comparison with morerostral and caudal planes, was reflected throughout the claustrum.
When ruminating on the function of the claustrum, we mustsurely take into consideration its anatomy and connectivity, as

Fig. 26. NPY-positive dendrites (D) and terminal boutons of differing size and shape
(arrows). Scale bar = 1 mm.
Fig. 27. Heavy labeled ‘‘en passant’’ NPY-positive bouton (arrows) terminates on the
periphery of an NPY-positive neuron. Scale bar = 2 mm.
D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119 115
well as the ultrastructure of its neurons. Our electron-microscopicanalysis of the cat claustrum revealed that large NPY-ir neuronscontained an abundance of cytoplasm rich in a variety oforganelles. In addition, their nuclei displayed a preponderanceof euchromatin, with mitochondria-laden terminal boutons.Characteristics such as these are reflective of cells with significantmetabolic activity. Furthermore, the long and multibranched
dendrites of these large neurons – especially those of the spinyvariety – offer support the contention that these neurons receive asignificant volume of synaptic input. Both the light- and electron-microscopic characteristics of these large cells are indicative ofprojection neurons rather than interneurons. This was confirmedby our previous research (Hinova-Palova, 1986).
Medium NPY-ir neurons typically had a lesser volume ofcytoplasm with a relative paucity of granular endoplasmicreticulum. In point of fact, several subtypes of medium neuronshave been found in the cat claustrum (Hinova-Palova, 1986).However, for the purposes of this study, we were unable to confirmthat the medium NPY-ir neurons we found corresponded to aspecific subtype. Nonetheless, it is not unreasonable to assumethat some of these cells are projection neurons owing to their light-microscopic and ultrastructural morphology.
Small NPY-ir neurons corresponded to the subtypes previouslydescribed by Hinova-Palova (1986). These neurons displayed arelatively large nucleus, a thin rim of cytoplasm, a paucity oforganelles and very few axo-somatic synapses – morphologicalcriteria that are commonly attributed to local circuit neurons, alsoreferred to as interneurons (Paloff, 1985; Paloff et al., 1989).Furthermore, most of the small NPY-ir neurons that we found wereseemingly filled or impregnated with immunoproduct. On thebasis of their light-microscopic and ultrastructural characteristics,we propose that these densely labeled small NPY-ir neurons arepart of a subpopulation of local circuit inhibitory interneurons.
We also observed NPY-ir dwarf cells in the present study.Ramon and Cajal (1911) was the first to recognize these cells in thestriatum by using the Golgi impregnation method. In previousinvestigations (Hinova-Palova, 1986; Hinova-Palova et al., 1987)we demonstrated the existence of dwarf cells in the cat claustrumvia light- and electron-microscopy. Mounting evidence leads us tobelieve that these cells are also interneurons.
To elaborate, the small and medium aspiny NPY-ir neurons seenin the present investigation were predominantly oval-to-fusiformin shape. In contrast, the large and medium spiny NPY-ir neuronswere often multipolar-to-pyramidal in shape. Based on the extantliterarture on this topic, it is not unreasonable to assume that theaspiny neurons represent a subpopulation of local circuitinhibitory interneurons (along with the dwarf cells), while thespiny neurons represent projection neurons. It was previouslyreported that the large spiny neurons in the cat claustrum receivedirect input from cortex and were also shown to project back to thesame areas of cortex, establishing excitatory synapses (Hinova-Palova, 1986; Hinova-Palova et al., 1988; Reynhout and Baizer,1999). Using the Golgi impregnation method in the monkeyclaustrum, Brand (1981) reported that the axons of large neuronswere efferent in nature, projected out of the claustrum. Moreover,in some species, it was reported that the overwhelming majority ofclaustral neurons are glutamatergic projection neurons, with only6–12% of cells engaged in local network activity (Wojcik et al.,2004).
The existence of projection neurons in the cat claustrum wasalso supported by our present study concerning the features oflarge, medium and small spiny NPY-ir neurons. In some sections,the entire initial segment of an NPY-ir myelinated axon waspresent. However, it must be noted that some of these axons werefound not to originate in the claustrum, and thus were afferent innature. Neverthless, the existence of afferent and efferentprojection fibers in claustrum has been suggested by Wojciket al. (2004).
Golgi impregnation studies of the mammalian claustrum havelead to the description of two major functional classes of neurons –projection neurons with polymorphic perikarya and spinydendrites, and inhibitory interneurons characterized by round-to-oval perikarya and aspiny dendrites (Brand, 1981; Braak and

D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119116
Braak, 1982; Spahn and Braak, 1985; Hinova-Palova, 1986; Drugaet al., 1993; Wojcik et al., 2004). Our current findings also revealtwo major classes of NPY-ir neurons in the cat claustrum: spinyand aspiny. In a previous study on the ultrastructure of the NPY-labeled cat claustrum (Edelstein et al., 2011b), we reported on theexistence of immunoproduct within dendritic spines of differingshape and size. Thus, our light- and electron-microscopicinvestigations strongly suggest that NPY-immunoreactivity ispresent in all neuronal types within the claustrum, and thatNPY-ir neurons are not reflective of a specific subpopulation. Thefact that NPY-immunoreactivity was observed in neurons ofvarying shapes and sizes suggests that these cells play differingroles in the context of claustral function (Hinova-Palova et al.,2007, 2008). Present results were comparable to our previousstudy of the synaptic organization of the cat claustrum (Hinova-Palova, 1986).
It has been established that neurons of differing shapes andsizes can have different functions (Papantchev et al., 2006). One ofthe aims of the present study was to identify whether NPYimmunoproduct was present in possibly functionally distinctsubpopulations of claustral neurons. It is generally accepted thatthe number and types of synapses are principal factors withrespect to influencing neuronal function (Paloff, 1985; Paloff et al.,1989). This explains why otherwise morphologically identicalneurons function and respond differently, as they are dependentupon on the characteristics and spike codes of their synapticinputs. Therefore, one could speculate that the morphologicallysimilar NPY-ir neurons we have found dispersed throughout thecat claustrum may not be as functionally similar as one mightthink.
As we have shown in this study, the observed population ofNPY-ir neurons was heterogeneous in nature, consisting of small,medium and large neurons of various shapes. In support of theabove-mentioned conclusions, we must emphasize that Golgiimpregnation studies of the claustrum in various species ofmammal have lead to the description of two major functionalclasses of neurons: projection neurons with polymorphic perikar-ya, spiny dendrites, prominent mitochondria and numerousaxodendritic synapses, and inhibitory interneurons characterizedby round or oval perikarya and aspiny dendrites (Braak and Braak,1982; Hinova-Palova, 1986; Wojcik et al., 2004). Another aim ofthis study was to establish whether NPY-ir neurons in theclaustrum have a well-defined topographical distribution. In pointof fact, such a distribution was not found. On the contrary, despitethe fact that the central part of the claustrum (stereotaxic planesA12–A16) was seen to contain the majority of NPY-ir neurons,these cells were diffusely distributed throughout the claustrum.Indeed, though NPY-ir neuronal clustering or grouping wasapparent within the central stereotaxic planes (which generallyhas the highest concentration of neurons), this was not seen withinthe ascribed functional zones of claustrum.
Immunohistochemical and in situ hybridization studies haveshown that NPY is one of most abundant and widely distributedpeptides in the central and autonomic nervous system ofmammals, particularly in rats and humans. In addition to theclaustrum, NPY is highly expressed in the cerebral cortex,hypothalamus, amygdala, hippocampus, nucleus of the solitarytract, locus coeruleus, nucleus accumbens, periaqueductual gray,nucleus raphe pallidus, striatum and the bed nucleus of the striaterminalis (Allen et al., 1983; Chronwall et al., 1985; Sasek andElde, 1985; Smith et al., 1985; de Quidt and Emson, 1986; Dumontet al., 1992; Todd and Spike, 1993; Krivukuca et al., 2010; Edelsteinet al., 2010, 2011a,b). Neuropeptide Y in involved in the control of abroadly diverse array of physiological functions including feedingbehavior, water consumption, learning and memory, locomotion,body temperature regulation, sexual behavior, emotional behavior,
neuronal excitability, cardiovascular homeostasis, hormone secre-tion and circadian rhythms (Baraban et al., 1997; Colmers andBleakman, 1994; Hokfelt et al., 1998; Inui, 1999; Munglani et al.,1996; Vezzani et al., 1999; Wahlested and Reis, 1993). In addition,it has been suggested that NPY plays a role in psychiatric disordersincluding depression, eating disorders, anxiety and epilepsy(Baraban et al., 1997; Colmers and Bleakman, 1994; Grundemaret al., 1992; Hokfelt et al., 1998; Munglani et al., 1996; Wahlestedand Reis, 1993). What more, hypothalamic injections of NPYincrease food intake (Stanley and Leibowitz, 1985); in thehippocampus, NPY has been suggested as an endogenous anti-epileptic (Colmers et al., 1991); in the cortex NPY is co-expressed inmany inhibitory GABAergic neurons that modulate pyramidal cellfunction in the other regions of the brain (Kubota et al., 2011); NPYcells play a role in cardiovascular function and emotion (Colmersand Wahlested, 1993).
From a clinical perspective, given that the claustrum and NPYhave been implicated in the development, symptoms and/orsequelae of serious maladies such as autism, schizophrenia,epilepsy, Alzheimer’s disease, Parkinson’s disease, and Hunting-ton’s disease (Wegiel et al., 2014; Cascella and Sawa, 2014; Venneriand Shanks, 2014; Kalaitzakis, 2014; Corcoran, 2004; Hallidayet al., 1990; Koide et al., 1995; Croom and Taylor, 2001; Flint andMartin, 1986; Ramos et al., 2006), it behooves both basic andclinical investigators to learn as much as possible about theinterplay between the two.
5. Conclusion
In conclusion, this is the first detailed investigation of the lightand electron-microscopic features of NPY-ir neurons and fibers inthe cat claustrum. It is hoped that our results and wide-rangingdiscussion will provide deeper insight into the functionalneuroanatomy of the claustrum, further elucidating its involve-ment with a host of critical physiological parameters and biologicprocesses, especially those impacted by the aforementionedneurological disorders. Lastly, much work remains to be done interms of studying the claustrum from an ontogenetic andphylogenetic perspective, with respect to homologues in non-mammalian species as well as its contribution in the context ofhigher cortical functions and prevalent hypotheses (Smythieset al., 2012, 2014a,b). In this regard, it must be mentioned that oneof the greatest minds in modern biology, the late Nobel laureateand co-discoverer of the structure of DNA, Francis Crick, spent thelast hours of his remarkable life dictating corrections to a draft ofwhat was to become his final paper, entitled ‘‘What is the functionof the claustrum?’’ (Crick and Koch, 2005; Edelstein and Denaro,2004b). Francis Crick and his co-author Christof Koch have laid thefoundation for future investigative efforts into the claustrum’sputative role in that most complex and elusive of neurophysiologicattributes, consciousness.
Ethical statement
All animals in this study were afforded human care incompliance with the ‘‘Principles of Laboratory Animal Care’’formulated by the National Society for Medical Research and the‘‘Guide for the Care and Use of Laboratory Animals’’ prepared bythe National Institutes of Health (NIH publication No. 86-23,revised 1996).
References
Adrian, T.E., Allen, J.M., Bloom, S.R., Ghatei, M.A., Rossor, M.N., Roberts, G.W., Crow,T.J., Tatemoto, K., Polak, J.M., 1983. Neuropeptide Y distribution in human brain.Nature 306 (5943) 584–586.

D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119 117
Allen, Y.S., Adrian, T.E., Allen, J.M., Tatemoto, K., Crow, T.J., Bloom, S.R., Polak, J.M.,1983. Neuropeptide Y distribution in the rat brain. Science 221 (4613) 877–879.
Ashwell, K.W.S., Hardman, C., Paxinos, G., 2004. The claustrum is not missing fromall monotreme brains. Brain Behav. Evol. 64, 223–241.
Baraban, S.C., Hollopeter, G., Erickson, J.C., Schwartzkroin, P.A., Palmiter, R.D., 1997.Knock-out mice reveal a critical antiepileptic role for neuropeptide Y. J. Neu-rosci. 17 (23) 8927–8936.
Berke, J.J., 1960. The claustrum, the external capsule and the extreme capsule ofMacaca mulatta. Neurology 115, 297–321.
Berlucchi, C., 1927. Microscopic anatomy of the claustrum and insula of the cat. Riv.Sper. Freniatr. 51, 125–157.
Braak, H., Braak, E., 1982. Neuronal types in the claustrum of man. Anat. Embryol.(Berl.) 163 (4) 447–460.
Brand, S., 1981. A serial section Golgi analysis of the primate claustrum. Anat.Embryol. (Berl.) 162, 447–460.
Brockhaus, H., 1940. Cytoarchitectural and myeloarchitectural study of claustralcortex and claustrum in man. J. Psychol. Neurol. 49, 249–348.
Carey, R.G., Bear, M.F., Diamond, I.T., 1980. The laminar organization of the recip-rocal projections between the claustrum and the striate cortex in the treeshrew, Tupaia glis. Brain Res. 184, 193–198.
Carey, R.G., Neal, T.L., 1985. The rat claustrum: afferent and efferent connectionswith visual cortex. Brain Res. 329, 185–193.
Cascella, N.G., Sawa, A., 2014. The Claustrum in Schizophrenia. In: Smythies, J.R.,Edelstein, L.R., Ramachandran, V.S. (Eds.), The Claustrum – Structural, Func-tional and Clinical Neuroscience. Elsevier, Oxford, pp. 237–243.
Chiba, A., Honma, Y., 1992. FMRFamide-immunoreactive structures in the brain ofthe brown hagfish, Paramyxine atami: relationship with neuropeptide Y-im-munoreactive structures. Histochemistry 98 (1) 33–38.
Chiba, A., Honma, Y., Oka, S., 1993. Immunohistochemical localization of neuro-peptide Y-like substance in the brain and hypophysis of the brown hagfish,Paramyxine atami. Cell Tissue Res. 271, 289–295.
Chiba, A., Honma, Y., 1994. Neuropeptide Y-immunoreactive structures in thetelencephalon and diencephalon of the white sturgeon, Acipenser transmonta-nus, with special regard to the hypothalamo-hypophyseal system. Arch. Histol.Cytol. 57 (1) 77–86.
Chronwall, B.M., Di Maggio, D.A., Massari, V.J., Pickel, V.M., Ruggiero, D.A., O’Dono-hue, T.L., 1985. The anatomy of neuropeptide-Y-containing neurons in rat brain.Neuroscience 15 (4) 1159–1181.
Colmers, W.F., Klapstein, G.J., Fournier, A., St-Pierre, S., Treherne, K.A., 1991.Presynaptic inhibition by neuropeptide Y in rat hippocampal slice in vitro ismediated by a Y2 receptor. Br. J. Pharmacol. 102 (1) 41–44.
Colmers, W.F., Wahlested, C., 1993. The Biology of Neuropeptide Y and RelatedPeptides. Humana Press, New Jersey.
Colmers, W.F., Bleakman, D., 1994. Effects of neuropeptide Y on the electricalproperties of neurons. Trends Neurosci. 17 (9) 373–379.
Corcoran, M.E., 2004. Clinical relations: epilepsy. In: Smythies, J.R., Edelstein, L.R.,Ramachandran, V.S. (Eds.), The Claustrum – Structural, Functional and ClinicalNeuroscience. Elsevier, Oxford, pp. 245–261.
Crick, F.C., Koch, C., 2005. What is the function of the claustrum? Philos. Trans. R.Soc. Lond. B: Biol. Sci. 360 (1458) 1271–1279.
Croom, J., Taylor, I.L., 2001. Neuropeptide Y, peptide YY and aluminum in Alzhei-mer’s disease: is there an etiological relationship? J. Inorg. Biochem. 87 (1–2)51–56.
Danger, J.M., Guy, J., Benyamina, M., Jegou, S., Leboulenger, F., Cote, J., Tonon, M.C.,Pelletier, G., Vaudry, H., 1985. Localization and identification of neuropeptide Y(NPY)-like immunoreactivity in the frog brain. Peptides 6 (6) 1225–1236.
Danger, J.M., Breton, B., Vallarino, M., Fournier, A., Pelletier, G., Vaudry, H., 1991.Neuropeptide-Y in the trout brain and pituitary: localization, characterization,and action on gonadotropin release. Endocrinology 128 (5) 2360–2368.
de Quidt, M.E., Emson, P.C., 1986. Distribution of neuropeptide Y-like immunore-activity in the rat central nervous system – II. Immunohistochemical analysis.Neuroscience 18 (3) 545–618.
De Vries, E., 1910. Bemerkungen zur ontogenie und vergleichenden anatomie desckaustrums. Folia Neurobiol. 4, 481–513.
Dinopoulos, A., Papadopoulos, G.C., Michaloudi, H., Parnavelas, J.G., Uylings, H.B.,Karamanlidis, A.N., 1992. Claustrum in the hedgehog (Erinaceus europaeus)brain: cytoarchitecture and connections with cortical and subcortical struc-tures. J. Comp. Neurol. 316 (2) 187–205.
Druga, R., 1966a. The claustrum of the cat (Felis domestica). Folia Morphol. (Praha)14, 7–16.
Druga, R., 1966b. Cortico-claustral connections, I. Fronto-claustral connections.Folia Morphol. (Praha) 14, 391–399.
Druga, R., 1971. Projection of prepyriform cortex into claustrum. Folia Morphol.(Praha) 19, 405–410.
Druga, R., 1974. The claustrum and the transitional neopaleocortical area of thehedgehog (Erinacea europaeus). Anat. Anz. 135, 442–454.
Druga, R., 1975. Claustrum (Struktura, Ontogenese a Spoje). (Doctoral disserta-tion)Charles University, Praha, pp. 193.
Druga, R., Chen, S., Bentivoglio, M., 1993. Parvalbumin and calbindin in the ratclaustrum: an immunocytochemical study combined with retrograde tracingfrom frontoparietal cortex. J. Chem. Neuroanat. 6, 399–406.
Dumont, Y., Martel, J.C., Foumier, A., St-Pierre, S., Quirion, R., 1992. Neuropeptide Yand Neuropeptide Y receptor subtypes in brain and peripheral tissues. Prog.Neurobiol. 38, 125–167.
Edelstein, L.R., 1986. The anatomy of the claustrum: a light- and electron-micro-scopic analysis in rat and monkey incorporating the technique of HRPcytochemistry. (Doctoral thesis)State University of New York at Stony Brook,New York, pp. 279.
Edelstein, L.R., Denaro, F.J., 2004a. The claustrum: a historical review of its anatomy,physiology, cytochemistry and functional significance. Cell. Mol. Biol. 50, 675–702.
Edelstein, L.R., Denaro, F.J., 2004b. The neurobiology of consciousness and SirFrancis Crick. Cell. Mol. Biol. 50, 671–673.
Edelstein, L.R., Cozzi, B., Castagna, M., Quilici, F., Lenzi, C., Piano, I., Pirone, A., 2010.Parvalbumin and neuropeptide Y immunoreactivity in the human claustrum.In: Society for Neuroscience, 40th Annual Meeting, Abstract #900.1.
Edelstein, L., Denaro, F., Stamm, J.S., Landzhov, B., Malinova, L., Hinova-Palova, D.,Paloff, A., Bozhilova-Pastirova, A., Dzhambazova, E., Bocheva, A., Ovtscharoff,W., 2011a. Distribution of CB1 receptors in the claustrum of rats undergoingacute stress: an immunohistochemical study. In: Society for Neuroscience, 41stAnnual Meeting, Abstract #734.12.
Edelstein, L., Hinova-Palova, D., Malinova, L., Papantchev, V., Landzhov, B., Paloff, A.,Ovtscharoff, W., 2011b. Distribution of neuropeptide Y immunoreactivity in thedorsal claustrum of the cat: light- and electron-microscopic identification ofdistinct neuronal populations. In: Society for Neuroscience, 41st Annual Meet-ing, Abstract #817.19.
Edelstein, L., Hinova-Palova, D., Denaro, F.J., Landzhov, B., Malinova, L., Minkov, M.,Paloff, A., Ovtscharoff, W., 2012a. NADPH-diaphorase-positive neurons in thehuman claustrum. In: Society for Neuroscience, 42nd Annual Meeting, Abstract#895.20.
Edelstein, L., Hinova-Palova, D., Landzhov, B., Malinova, L., Minkov, M., Paloff, A.,Ovtscharoff, W., 2012b. Neuronal nitric oxide synthase immunoreactivity in thehuman claustrum: light- and electron microscopic investigation. In: Society forNeuroscience, 42nd Annual Meeting, Abstract #895.21.
Emson, P.C., De Quidt, M.E., 1984. NPY – a new member of the pancreatic polypep-tide family. Trends Neurosci. 7, 31–35.
Filimonoff, I.N., 1966. The claustrum: its origin and development. J. Hirnforsch. 8,503–528.
Flint, B., Martin, J.B., 1986. Neuropeptides in neurological disease. Ann. Neurol. 20(5) 547–565.
Grundemar, L., Jonas, S.E., Morner, N., Hogestatt, E.D., Wahlested, C., Hakanson, R.,1992. Characterization of vascular neuropeptide Y receptors. Br. J. Pharmacol.105 (1) 45–50.
Guirado, S., Real, M.A., Olmos, J.L., Davila, J.C., 2003. Distinct types of nitric oxide-producing neurons in the developing and adult mouse claustrum. J. Comp.Neurol. 465, 431–444.
Halliday, G.M., Li, Y.W., Blumbergs, P.C., Joh, T.H., Cotton, R.G., Howe, P.R., Blessing,W.W., Geffen, L.B., 1990. Neuropathology of immunohistochemicallyidentified brainstem neurons in Parkinson’s disease. Ann. Neurol. 27 (4)373–385.
Hinova-Palova, D.V., Paloff, A.M., Penev, D.I., 1979. Synaptic organization of theclaustrum in the cat. C. R. Acad. Bulg. Sci. 32 (6) 831–834.
Hinova-Palova, D.V., Paloff, A.M., Usunoff, K.G., 1980a. Identification of three typesof degenerated boutons in claustrum dorsale of the cat after lesion of thetemporal cortex. C. R. Acad. Bulg. Sci. 33, 125–128.
Hinova-Palova, D.V., Paloff, A.M., Usunoff, K.G., 1980b. Identification of three typesof degenerated boutons in claustrum dorsale of the cat after lesion of the frontalcortex. C. R. Acad. Bulg. Sci. 33, 129–132.
Hinova-Palova, D., Usunoff, K., 1981. Electron-microscopic evidence for theexistence of a nigroclaustral projection in the cat. C. R. Acad. Bulg. Sci. 34 (5)729–732.
Hinova-Palova, D., 1981. Identification of degenerated boutons in claustrum dorsaleafter lesion of visual cortex. C. R. Acad. Bulg. Sci. 34, 449–452.
Hinova-Palova, D.V., Paloff, A.M., 1982. Corticoclaustral connections. An electron-microscopic study. Verh. Anat. Ges. 76, 503–504.
Hinova-Palova, D., Paloff, A., 1984. Identification of degenerated synaptic boutons inthe claustrum of cat after lesion of the parietal cortex. Contemp. Probl. Neu-romorphol. (Sofia) 13–14, 154–160.
Hinova-Palova, D., 1986. Light-microscopic and ultrastructural organization of theclaustrum in the cat. Afferent and efferent connections (Thesis)Medical Acade-my, Sofia.
Hinova-Palova, D., Dimova, R., Ivanov, D.P., 1987. Identification of small neurons(dwarf cells) in the claustrum of the cat. Light and electron microscopicobservation. Verh. Anat. Ges. Leipzig 66.
Hinova-Palova, D., Christova, T., 1988. Immunohistochemical investigation ofsomatostatin (SRIF), vasoactive polypeptide (VIP) and glial fibrillary acidicprotein (GFAP) in the claustrum of the cat. In: Third symposium and schoolof histochemistry and cytochemistry, Varna, Bulgaria.
Hinova-Palova, D., Paloff, A., Usunoff, K., Dimova, R., Wossifov, T., Ivanov, D., 1988.Reciprocal connections between the claustrum and the auditory cortical field inthe cat. An experimental study using light and electron microscopic antero-grade degeneration methods and the horseradish peroxidase retrograde axonaltransport. J. Hirnforsch. 29, 255–278.
Hinova-Palova, D., Braak, E., 1993. Somatostatin-like immunoreactive neurons inthe human claustrum. In: XIth Congress of Anatomists and Embriologists, Sofia,Bulgaria.
Hinova-Palova, D.V., Paloff, A.M., Christova, T., Ovtcsharoff, W., 1997. Topographicaldistribution of NADPH-diaphorase-positive neurons in the cat’s claustrum. Eur.J. Morphol. 35, 105–116.

D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119118
Hinova-Palova, D.V., Christova, T., Yotovski, P.V., Logofetov, A.P., Paloff, A.M., 2001.Somatostatin-like neurons and fibres in the cat claustrum. C. R. Acad. Bulg. Sci.54, 81–84.
Hinova-Palova, D., Edelstein, L., Paloff, A., Hristov, S., Papantchev, V., Ovtscharoff, W.,2007. Parvalbumin in the cat claustrum: ultrastructure, distribution and func-tional implications. Acta Histochem. 109, 61–77.
Hinova-Palova, D., Edelstein, L., Paloff, A., Hristov, S., Papantchev, V., Ovtscharoff, W.,2008. Neuronal nitric oxide synthase immunopositive neurons in cat claustrum– a light and electron microscopic study. J. Mol. Histol. 39, 447–457.
Hinova-Palova, D., Edelstein, L., Papantchev, V., Landzhov, B., Malinova, L., Todor-ova-Papantcheva, D., Minkov, M., Paloff, A., Ovtscharoff, W., 2012. Light- andelectron-microscopic study of leucine enkephalin immunoreactivity in the catclaustrum. J. Mol. Histol. 43 (6) 641–649.
Hinova-Palova, D., Edelstein, Landzhov, B., Braak, E., Malinova, L., Minkov, M., Paloff,A., Ovtscharoff, W., 2013a. Parvalbumin-immunoreactive neurons in thehuman claustrum. Brain Struct. Funct., http://dx.doi.org/10.1007/s00429-013-0603-x.
Hinova-Palova, D., Edelstein, Landzhov, B., Minkov, M., Malinova, L., Alexandrov, A.,Hristov, S., Paloff, A., Ovtscharoff, W., 2013b. Light microscopic immunocyto-chemical identification of leucine enkephalin in human claustrum. Scr. Sci. Med.45 (l) 23–28.
Hinova-Palova, D., Edelstein, Landzhov, B., Minkov, M., Malinova, L., Hristov, S.,Denaro, F.J., Alexandrov, A., Kiriakova, T., Brainova, I., Paloff, A., Ovtscharoff, W.,2013c. Distribution and morphological characteristics of NADPH-diaphorase-positive neurons and fibers in the human claustrum. Front. Syst. Neurosci.,http://dx.doi.org/10.3389/fnsys.2014.00096.
Hokfelt, T., Broberger, C., Zhang, X., Diez, M., Koop, J., Xu, Z.Q., Landry, M., Bao, L.,Schalling, M., Koistinaho, J., Dearmond, S.J., Prusiner, S., Gong, J., Walsh, J.H.,1998. Neuropeptide Y: some viewpoints on a multifaceted peptide in thenormal and diseased nervous system. Brain Res. Rev. 26, 154–166.
Inui, A., 1999. Neuropeptide Y feeding receptors: are multiple subtypes involved?Trends Pharmacol. Sci. 20 (2) 43–46.
Kalaitzakis, M.E., 2014. Parkinson’s disease and the claustrum. In: Smythies J.R.,Edelstein, L.R., Ramachandran, V.S. (Eds.), The Claustrum – Structural, Func-tional and Clinical Neuroscience. Elsevier, Oxford, pp. 277–297.
Koide, S., Onishi, H., Hashimoto, H., Kai, T., Yamagami, S., 1995. Plasma neuropeptideY is reduced in patients with Alzheimer’s disease. Neurosci. Lett. 198 (2) 149–151.
Kowianski, P., Morys, J.M., Wojcik, S., Dziewiatkowski, J., Luczynska, A., Spodnik, E.,Timmermans, J.P., Morys, J., 2004. Neuropeptide-containing neurons in theendopiriform region of the rat: morphology and colocalization with calcium-binding proteins and nitric oxide synthase. Brain Res. 996, 97–110.
Krivukuca, D., Puskas, L., Puskas, N., Eric, M., 2010. Morphometric characteristics ofneuropeptide Y immunoreactive neurons in cortex of human inferior parietallobule. Coll. Antropol. 34 (Suppl. 1) 99–104.
Kubota, Y., Shigematsu, N., Karube, F., Sekigawa, A., Kato, S., Yamaguchi, N., Hirai, Y.,Morishima, M., Kawaguchi, Y., 2011. Selective coexpression of multiple chemi-cal markers defines discrete populations of neocortical GABAergic neurons.Cereb. Cortex 21, 1803–1817, http://dx.doi.org/10.1093/cercor/bhq252.
Kunzle, H., 1975. Bilateral projection from precentral motor cortex to the putamenand other parts of the basal ganglia. An autoradiographic study in Macacafascicularis. Brain Res. 88, 195–209.
Kunzle, H., 1978. An autoradiographic analysis of the efferent connections from thepremotor and adjacent prefrontal regions (area 6 and 9) in Macaca fascicularis.Brain Behav. Evol. 15, 185–234.
Landau, E., 1923. Zur kenntnis der Beziehungen des claustrums zum nucleusamygdalae und zur area piriformis im speziellen zum tractus olfactorius.Schweiz. Arch. Neurol. Psychiatr. 13, 391–400.
Loo, T.T., 1931. The forebrain of the opossum, Didelphis virginiana. J. Comp. Neurol.52 (1) 1–148.
Macchi, G., 1984. Morphology and structure of human claustrum. Cervello 24, 1–26.Munglani, R., Hudspith, M.J., Hunt, S.P., 1996. The therapeutic potential of neuro-
peptide Y. Analgesic, anxiolytic and antihypertensive. Drugs 52 (3) 371–389.Neal, J.W., Pearson, R.C.A., Powell, T.P.S., 1986. The relationship between the
auditory cortex and the claustrum in the cat. Brain Res. 366, 145–151.Norita, M., 1977. Demonstration of bilateral claustro-cortical connections in the cat
with the method of retrograde axonal transport of horseradish peroxidase.Arch. Histol. Jpn. 40, 1–10.
Olson, C.R., Graybiel, A.M., 1980. Sensory maps in the claustrum of the cat. Nature288, 479–481.
Otellin, V.A., Makarov, F.N., 1972. Descending connections of the auditory cortex ofthe cat with contralateral neostriatal complex and claustrum. Dokl. Akad. NaukSSSR (Ser. Biol.) 202, 723–725.
Paloff, A.M., 1985. Somatodendritic synapses in the central nucleus of colliculusinferior (CI) in the cat. J. Hirnforsch. 26, 353–358.
Paloff, A.M., Usunoff, K.G., Hinova-Palova, D., Ivanov, D.P., 1989. The fine structure ofthe inferior colliculus in the cat. I. Neuronal perikarya in the central nucleus. J.Hirnforsch. 30, 69–90.
Paloff, A.M., Usunoff, K.G., Yotovski, P., Hinova-Palova, D.V., Ovtscharoff, W.A., 2004.Parvalbumin-like immunostaining in the cat inferior colliculus. Light andelectron microscopic investigation. Acta Histochem. 106 (3) 219–234.
Papantchev, V., Paloff, A., Hinova-Palova, D., Hristov, S., Todorova, D., Ovtshcaroff,W., 2006. Neuronal nitric oxide synthase immunopositive neurons in the catvestibular nuclear complex: a light and electron microscopic study. J. Mol.Histol. 37, 343–352.
Papantchev, V., 2008. Cytoarchitectonics, ultrastructure and immunocytochemistryof vestibular complex of the cat. (Ph.D. thesis)Medical University, Sofia,Bulgaria.
Paxinos, G., Watson, C., 1989. The Rat Brain in Stereotaxic Coordinates, second ed.Academic Press, New York.
Perroteau, I., Danger, J.M., Biffo, S., Pelletier, G., Vaudry, H., Fasolo, A., 1988.Distribution and characterization of neuropeptide Y-like immunoreactivityin the brain of the crested newt. J. Comp. Neurol. 275 (3) 309–325.
Pilleri, G., 1961. The claustrum of Didelphis marsupialis Lin (Marsupialis, Didelphoi-dea). Acta Anat. Basel 45, 310–314.
Pilleri, G., 1962. The claustrum of the Canadian beaver (Castor canadensis Kuhl):structure and comparative anatomy. J. Hirnforsch. 5, 59–81.
Pontet, A., Danger, J.M., Dubourg, P., Pelletier, G., Vaudry, H., Calas, A., Kah, O.,1989. Distribution and characterization of neuropeptide Y-like immuno-reactivity in the brain and pituitary of the goldfish. Cell Tissue Res. 255 (3)529–538.
Rae, A.S.L., 1954. The connections of the claustrum. Confin. Neurol. Basel. 14, 211–219.
Ramon, Cajal, S., 1911. Histologie du Systeme Nerveux de l’Homme et des Verteb-retes, Paris.
Ramos, B., Baglietto-Vargas, D., del Rio, J.C., Moreno-Gonzalez, I., Santa-Maria, C.,Jimenez, S., Caballero, C., Lopez-Tellez, J.F., Khan, Z.U., Ruano, D., Gutierrez, A.,Vitorica, J., 2006. Early neuropathology of somatostatin/NPY GABAergic cells inthe hippocampus of a PS1xAPP transgenic model of Alzheimer’s disease.Neurobiol. Aging 27 (11) 1658–1672.
Reinoso-Suarez, F., 1961. Topographischer hirnatlas der katze fur experimental-physiologische untersuchungen. E Merck AG.
Reynhout, K., Baizer, J.S., 1999. Immunoreactivity for calcium-binding proteins inthe claustrum of the monkey. Anat. Embryol. (Berl.) 199 (1) 75–83.
Riche, D., Lanoir, J., 1978. Some claustrocortical connections in the cat and baboonas studied by retrograde HRP transport. J. Comp. Neurol. 177, 435–444.
Sasek, C.A., Elde, R.P., 1985. Distribution of neuropeptide Y-like immunoreactivityand its relationship to FMRF-amide-like immunoreactivity in the sixthlumbar and first sacral spinal cord segments of the rat. J. Neurosci. 5 (7)1729–1739.
Sherk, H., 1986. The claustrum and the cerebral cortex. In: Jones, E.G., Peters, A.(Eds.), Cerebral Cortex 5. Plenum Press, New York, pp. 467–499.
Sloniewski, P., Usunoff, K.G., Pilgrim, C., 1986. Diencephalic and mesencephalicafferents of the rat claustrum. Anat. Embryol. (Berl.) 173, 401–411.
Smith, Y., Parent, A., Kerkerian, L., Pelletier, G., 1985. Distribution of neuropeptide Yimmunoreactivity in the basal forebrain and upper brainstem of the squirrelmonkey (Saimiri sciureus). J. Comp. Neurol. 236 (1) 71–89.
Smythies, J., Edelstein, L., Ramachandran, V.S., 2012. Hypotheses relating to thefunction of the claustrum. Front. Integr. Neurosci. 6, 53, http://dx.doi.org/10.3389/fnint.2012.00053.
Smythies, J., Edelstein, L., Ramachandran, V., 2014a. Hypotheses relating tothe function of the claustrum II: instructional oscillations and dendriticintegration. Front. Integr. Neurosci. 8, 7, http://dx.doi.org/10.3389/fnint.2014.00007.
Smythies, J., Edelstein, L., Ramachandran, V.S., 2014b. Hypotheses relating tothe function of the claustrum. In: Smythies, J.R., Edelstein, L.R., Ramachandran,V.S. (Eds.), The Claustrum – Structural, Functional and Clinical Neuroscience.Elsevier, Oxford, pp. 299–352.
Spahn, B., Braak, H., 1985. Percentage of projection neurons and various types ofinterneurons in the human claustrum. Acta Anat. Basel 122, 245–248.
Stanley, B.G., Leibowitz, S.F., 1985. Neuropeptide Y injected in the paraventricularhypothalamus: a powerful stimulant of feeding behavior. Proc. Natl. Acad. Sci.U. S. A. 82 (11) 3940–3943.
Stelmasiak, M., 1955. Volume of the claustrum in man. Folia Morphol. Warszawa 6,137–144.
Tanne-Gariepy, J., Boussaoud, D., Rouiller, E.M., 2002. Projections of the claustrumto the primary motor, premotor, and prefrontal cortices in the macaquemonkey. J. Comp. Neurol. 454, 140–157.
Tatemoto, K., 1982. Neuropeptide Y: complete amino acid sequence of the brainpeptide. Proc. Natl. Acad. Sci. U. S. A. 79 (18) 5485–5489.
Tatemoto, K., Carlquist, M., Mutt, V., 1982. Neuropeptide Y – a novel brain peptidewith structural similarities to peptide YY and pancreatic polypeptide. Nature296 (5858) 659–660.
Todd, A.J., Spike, R.C., 1993. The localization of classical transmitters and neuropep-tides within neurons in laminae I–III of the mammalian spinal dorsal horn. Prog.Neurobiol. 41 (5) 609–645.
Vallarino, M., Danger, J.M., Fasolo, A., Pelletier, G., Saint-Pierre, S., Vaudry, H., 1988.Distribution and characterization of neuropeptide Y in the brain of an elasmo-branch fish. Brain Res. 448 (1) 67–76.
Venneri, A., Shanks, M., 2014. The claustrum and Alzheimer’s disease. In: Smythies,J.R., Edelstein, L.R., Ramachandran, V.S. (Eds.), The Claustrum – Structural,Functional and Clinical Neuroscience. Elsevier, Oxford, pp. 263–275.
Vezzani, A., Sperk, G., Colmers, W.F., 1999. Neuropeptide Y: emerging evidence for afunctional role in seizure modulation. Trends Neurosci. 22 (1) 25–30.
Wahlested, C., Reis, D.J., 1993. Neuropeptide Y-related peptides and their receptors– are the receptors potential therapeutic drug targets? Annu. Rev. Pharmacol.Toxicol. 33, 309–352.
Wegiel, J., Morys, J., Kowianski, P., Ma, S.Y., Kuchna, I., Nowicki, K., Imaki, H., Wegiel,J., Flory, M., Brown, W.T., Wisniewski, T., 2014. Delayed development of theclaustrum in autism. In: Smythies, J.R., Edelstein, L.R., Ramachandran, V.S.

D.V. Hinova-Palova et al. / Journal of Chemical Neuroanatomy 61–62 (2014) 107–119 119
(Eds.), The Claustrum – Structural, Functional and Clinical Neuroscience.Elsevier, Oxford, pp. 225–235.
Whale, P., Albus, K., 1984. Localization of an NPY-like immunoreactivity in the cat’scentral nervous system. Soc. Neurosci. 10, 433 (Abstract).
Witter, M.P., Room, P., Groenewegen, H.J., Lohman, A.H.M., 1988. Reciprocalconnections of the insular and piriform claustrum with limbic cortex: ananatomical study in the cat. Neuroscience 24, 519–539.
Wojcik, S., Dziewiatkowski, J., Spodnik, E., Ludkiewicz, B., Domaradzka-Pytel, B.,Kowianski, P., Morys, J., 2004. Analysis of calcium-binding protein immuno-reactivity in the claustrum and the endopiriform nucleus of the rabbit. ActaNeurobiol. Exp. (Warsz.) 64, 449–460.
Zilles, K., Zilles, B., 1980. A quantitative approach to cytoarchitectonics. VI. Theareal pattern of the cortex of the albino rat. Anat. Embryol. (Berl.) 159 (3) 335–360.




















