
Antagonistic Mechanism of Iturin A and Plipastatin A ... - PLOS

ojp ELSEVIER European Journal of Pharmacology 271 (1994) 201-212
Antagonistic regulation of a2-adrenoceptors by neuropeptide Y receptor subtypes in the nucleus tractus solitarii
Shao-Nian Yang a, D6bora R. Fior a, Peter B. Hedlund a, Luigi F. Agnati b, Kjell Fuxe a,, a Division of Cellular and Molecular Neurochemistry, Department of Neuroscience, Karolinska Instituter, S-171 77 Stockholm, Sweden
6 Institute of Human Physiology, University of Modena, Modena, Italy
Received 14 April 1994; revised MS received 9 August 1994; accepted 13 September 1994
Abstract
The modulation of a2-adrenoceptors by neuropeptide Y Y1 and neuropeptide Y Ye receptor subtypes has been studied in the nucleus tractus solitarii of the male rat. The autoradiographical experiments showed that neuropeptide Y-(1-36), neuropeptide Y-(13-36), a selective neuropeptide Y Y2 receptor agonist, and [Leu31,Pro34]neuropeptide Y, a selective neuropeptide Y Y1 receptor agonist, in the nanomolar range increased the K d value of the [3H]p-aminoclonidine binding sites in the above rank order of potency without changing the Bma x values. In contrast, in the competition experiments, the neuropeptide Y Y1 and the neuropeptide Y Y2 receptor agonists decreased and increased, respectively, with the same potency the IC50 value of/-adrenaline and especially of clonidine for the a2-adrenoceptor agonist binding sites associated with an increase and a decrease of the B 0 value, respectively. Cardiovascular experiments showed that microinjections of clonidine into the nucleus tractus solitarii induced dose-dependent vasodepressor and bradycardiac responses. Threshold doses for vasodepressor effects of neuropeptide Y-(1-36) and of the neuropeptide Y Ya receptor agonist and for vasopressor effects of the neuropeptide Y Y2 receptor agonist significantly counteracted the vasodepressor action elicited by an EDso dose of clonidine in the nucleus tractus solitarii, the bradycardiac action of clonidine also being counteracted by the neuropeptide Y Yz but not the neuropeptide Y Yt receptor agonist. The present results give indications for the existence of an antagonistic modulation of high affinity a2-adrenoceptors by the neuropeptide Y Y1 and neuropeptide Y Y2 receptor subtype in the nucleus tractus solitarii which may contribute to a reduction of a2-adrenoceptor-mediated cardiovascular depression.
Keywords: Nucleus tractus solitarii; Neuropeptide Y receptor; a2-Adrenoceptor; Receptor autoradiography; Cardiovascular
1. Introduction
The nucleus tractus solitarii contains a high density of high affinity a2-adrenoceptors (Fuxe et al., 1987a; Rosin et al., 1993; Unnerstall et al,, 1984; Young and Kuhar, 1980) and of the neuropeptide Y Y1 and neu- ropeptide Y Y2 receptors (Aicher et al., 1991; Fuxe et al., 1987a; Gehlert et al., 1992; H~irfstrand et al., 1986; Leslie et al., 1988). By using membrane preparations of the dorsomedial medulla oblongata as well as quantita- tive receptor autoradiography it has been shown that neuropeptide Y-(1-36) can reduce the affinity of the a2-adrenoceptor agonist binding sites within the dorso- medial medulla oblongata including the nucleus tractus
* Corresponding author. Tel. + 46 (8) 728 7080, fax + 46 (8) 337 941.
0014-2999/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0 0 1 4 - 2 9 9 9 ( 9 4 ) 0 0 5 6 2 - 1
solitarii itself (Agnati et al., 1983; Fuxe et al., 1987a; H~irfstrand et al., 1989). Furthermore, studies using pertussis toxin treatment have shown that this interac- tion between neuropeptide Y receptors and aa-adren- oceptors in the dorsomedial medulla oblongata is de- pendent upon G i proteins (Von Euler et al., 1989). This Gi protein-dependent receptor-receptor interac- tion is also true for the central cardiovascular actions of neuropeptide Y-(1-36) and the a2-adrenoceptor agonist clonidine (Fuxe et al., 1989). In view of the existence of a high density of neuropeptide Y and adrenaline costoring nerve terminal networks within the nucleus tractus solitarii it seems likely that the ligands which interact with neuropeptide Y receptors and a2-adrenoceptors originate mainly from the adrenal ine /neuropept ide Y costoring terminals within the nucleus tractus solitarii itself (Everitt et al., 1984; Fuxe et al., 1986). Previous work has demonstrated

202 S.-N. Yang et al. / European Journal of Pharmacology 271 (1994) 201-212
that intraventricular injections of an a2-adrenoceptor agonist such as clonidine (Kobinger, 1978) and of neu- ropeptide Y-(1-36) (Fuxe et al., 1990) lead to marked vasodepressor and bradycardiac responses in the anaesthetized rat and the awake unrestrained rat. When given together they were found to antagonize the cardiovascular actions of one another (Fuxe et al., 1987b; H~irfstrand and Fuxe, 1987). The mechanism underlying this antagonistic interaction has been sug- gested to be a receptor-receptor interaction between neuropeptide Y receptors and a2-adrenoceptors mainly located within the nucleus tractus solitarii (Fuxe et al., 1987a; H~irfstrand et al., 1989).
It has been demonstrated that microinjections of neuropeptide Y-(1-36) and of neuropeptide Y Y1 re- ceptor agonists as well as of clonidine and adrenaline into the nucleus tractus solitarii lead to vasodepressor responses (Barraco et al., 1990; Rockhold and Cald- well, 1980; Yang et al., 1993, 1994; Zandberg et al., 1979). In contrast, microinjections of the neuropeptide Y Y2 receptor agonist, neuropeptide Y-(13-36), into the nucleus tractus solitarii lead to vasopressor re- sponses in the femtomolar dose range (Narvfiez et al., 1993). It is well known that C-terminal neuropeptide Y fragments have a much higher potency at neuropeptide Y Y2 versus neuropeptide Y Y1 receptors (Wahlestedt et al., 1990). In the present study, we have evaluated if in fact the demonstrated neuropeptide Y receptor/ a2-adrenoceptor interaction takes place within the nu- cleus tractus solitarii as determined by coinjections of neuropeptide Y Y1 and/or neuropeptide Y Y2 recep- tor agonists together with clonidine in the nucleus tractus solitarii and by quantitative receptor auto- radiography of the binding characteristics of Otz-adren- oceptors in the nucleus tractus solitarii in the absence or presence of neuropeptide Y peptides. Furthermore, in view of the differential cardiovascular actions of neuropeptide Y Y1 and neuropeptide Y Y2 receptor agonists the role of neuropeptide Y Y1 and neuropep- tide Y Y2 receptor agonists in the control of az-adren- oceptor agonist binding sites in the nucleus tractus solitarii needs to be clarified.
2. Materials and methods
2.1. Animals
Male specific pathogen free Sprague-Dawley rats (body weight: 150 g for in vitro studies and 200-265 g for in vivo studies) obtained from B&K Universal, Stockholm, Sweden, were maintained on a regular light-dark cycle (light on at 06:00 h and off at 20:00 h) in temperature- and humidity-controlled rooms. The animals had free access to food pellets and tap water.
2.2. Quantitative receptor autoradiography
The procedure to perform receptor autoradiography has been described elsewhere (Hedlund et al., 1991). The rats were killed by decapitation and their brains were rapidly removed from the skull and frozen in CO2 gas. Coronal sections (20 /zm thick) were made in a Leitz cryostat at bregma levels -13.68 mm to -14.08 mm according to the atlas of Paxinos and Watson (1986) and thaw-mounted on gelatin-coated slides.
The in vitro effects of neuropeptide Y-(1-36), neu- ropeptide Y-(13-36) and [Leu31,Pro34]neuropeptide Y on az-adrenoceptors were studied by both competition and saturation experiments. A group of six animals were used in each competition experiment in which a total of 40 sections of each rat, divided into four groups of ten sections, were analysed. Adjacent sections were used for control and the treated groups. Competition experiments were performed with ten concentrations (10/zM to 0.1 nM) of/-adrenaline or clonidine versus 10 nM of the a2-adrenoceptor agonist [3H]p-amino- clonidine in the presence or absence of neuropeptide Y-(1-36), neuropeptide Y-(13-36) and [Leu 31, Pro34]neuropeptide Y in concentrations of 1, 10 and 30 nM. Sections were incubated in a 50 mM Tris-HCl buffer (pH 7.7) containing 5 mM MgC12, 5 mM CaCI 2, 1 ~M bacitracin, 0.25% bovine serum albumin, 0.01% ascorbic acid and 10 nM [3H]p-aminoclonidine. Satu- ration experiments were also performed using [3H]p- aminoclonidine in concentrations ranging from 1 to 40 nM. A group of six animals was used. In each satura- tion experiment, four groups of ten sections were di- vided in order to perform the total binding, the non- specific binding, the total binding and the non-specific binding in the presence of neuropeptide Y-(1-36), neuropeptide Y-(13-36) or [Leu31,Pro34]neuropeptide Y in concentrations ranging from 0.1 to 30 nM. In this way, each concentration tested had its own paired control. Non-specific binding was defined as the bind- ing in the presence of 10/zM of l-noradrenaline. The binding was performed under equilibrium conditions (30 min at room temperature, 22°C). Following incuba- tion, the sections were washed in 50 mM Tris-HCl buffer (pH 7.7) two times 5 min, rinsed twice in dis- tilled water (0-4°C) and dried in a stream of cold air. The sections were then exposed to a tritium-sensitive film for 6 weeks.
2.3. Computer-assisted image analysis
The autoradiograms were analysed as described pre- viously (Hedlund et al., 1991) using a computer-as- sisted image analysis system. The computer software was developed by Imaging Reseach (Brock University, Canada). The mean grey values were determined by bilateral measurements in the nucleus tractus solitarii
S.-N. Yang et al. / European Journal of Pharmacology 271 (1994) 201-212 203
using a square as a sampling field (0.06 1"Ilil12). Pre- fabricated 3H-labeled polymer strips (Amersham mi- croscale, UK) were used to convert the grey values into fmol/mg protein values (Benfenati et al., 1986; Cum- mins et al., 1987).
2.4. In vivo cardiovascular experiments
Rats were anaesthetized with a mixture of a-chlo- ralose (35 mg/kg i.p.) and urethane (1 g/kg i.p.). The trachea was cannulated to avoid obstruction of the airway, and body temperature was maintained at 37.5°C by means of a thermostatic blanket. A heparinized (Heparin, 50 IU/ml in 0.9% saline) catheter was in- serted in the femoral artery and connected to a Statham PC23 DC transducer (Statham Co., Puerto Rico), con- nected to a Grass polygraph (model 7, Grass Instru- ments, USA) to monitor the arterial blood pressure and heart rate. The transducer and polygraph recorder were calibrated with a mercury manometer prior to the blood pressure recordings. The animals were then placed in a stereotaxic frame (Kopf, USA) and the head was adjusted to a 45 ° angle from the horizontal plane. The electrocautery of the neck muscles was conducted to expose the posterior atlanto-occipital membrane and then fine dissection was employed to reveal the caudal medulla in the region of the calamus scriptorius. The stereotaxic coordinates for microinjec- tions were 0.5 mm rostral, 0.5 mm lateral to the caudal tip of the area postrema (Zandberg and De Jong, 1977), and 0.5 mm below the dorsal surface of the brainstem. Each animal received only one unilateral microinjection into the dorsomedial part of the nucleus tractus solitarii made by a glass micropipette (tip diam- eter 40-50/zm) connected to a Hamilton micro-syringe (0.5 /xl). In a first set of experiments, five different doses of clonidine (0.05-5 nmol) dissolved in artificial cerebrospinal fluid (aCSF) (120 mM NaC1, 20 mM NaHCO 3, 2 mM KCI, 0.5 mM K H 2 P O 4 , 1.2 mM CaC12, 1.8 mM MgCI2, 0.5 mM Na2SO 4, 5.8 mM D-glucose; pH 7.35), and aCSF (50 nl) alone as a control were injected in the nucleus tractus solitarii to evaluate dose-dependent effects of clonidine. The dose-dependent effects of neuropeptide Y-(1-36), neu- ropeptide Y-(13-36) and [Leu31,Pro34]neuropeptide Y were determined in previous papers (Narv~ez et al., 1993; Yang et al., 1993). In a second set of experiments coinjections of a close to EDs0 dose of clonidine (200 pmol) with a subthreshold (neuropeptide Y-(1-36): 100 fmol; [Leu3~,Pro3a]neuropeptide Y: 100 fmol; neu- ropeptide Y-(13-36): 10 fmol) and a threshold dose (neuropeptide Y-(1-36): 1 pmol; [Leu3;,Proaa]neuro - peptide Y: 2.5 pmol; neuropeptide Y-(13-36): 50 fmol) of neuropeptide Y peptides were given to evaluate any modulatory effect of these peptides on the a2-adreno- ceptor-mediated vasodepressor and bradycardiac re-
sponses. Drug and control solutions were injected in 50 nl during a period of 10 s. Recordings were performed as described earlier (H~irfstrand et al., 1987). Basal values were registered every 5 min during a period of 15 min before the microinjection and the measure- ments of the mean arterial blood pressure and heart rate were made during the following 1 h time interval. The placement of the injections and the diffusion area were evaluated by injecting Evans blue dye (50 nl) at the end of the experiment.
2.5. Data analys& and statistics
All data from the competition and saturation experi- ments were analysed by iterative non-linear regression
A
150 -
: lOO-
B
A
o
~ 1000-
O
m
T _ T
F i • t J s
/ 1 ¢ • / / x ~ ,
¢ 1 ~ ¢ / /
T
10 3O
! " - -
• / , ~ / x J , ~
• t • , ¢ ~ ¢ 4
r e
,h-•"" i:!:t
. T
7 , / ,
/ / J
r . - i
• / i
r / l
F / ,
F / •
10 3O c o n c a n t r a t l o n ( n M )
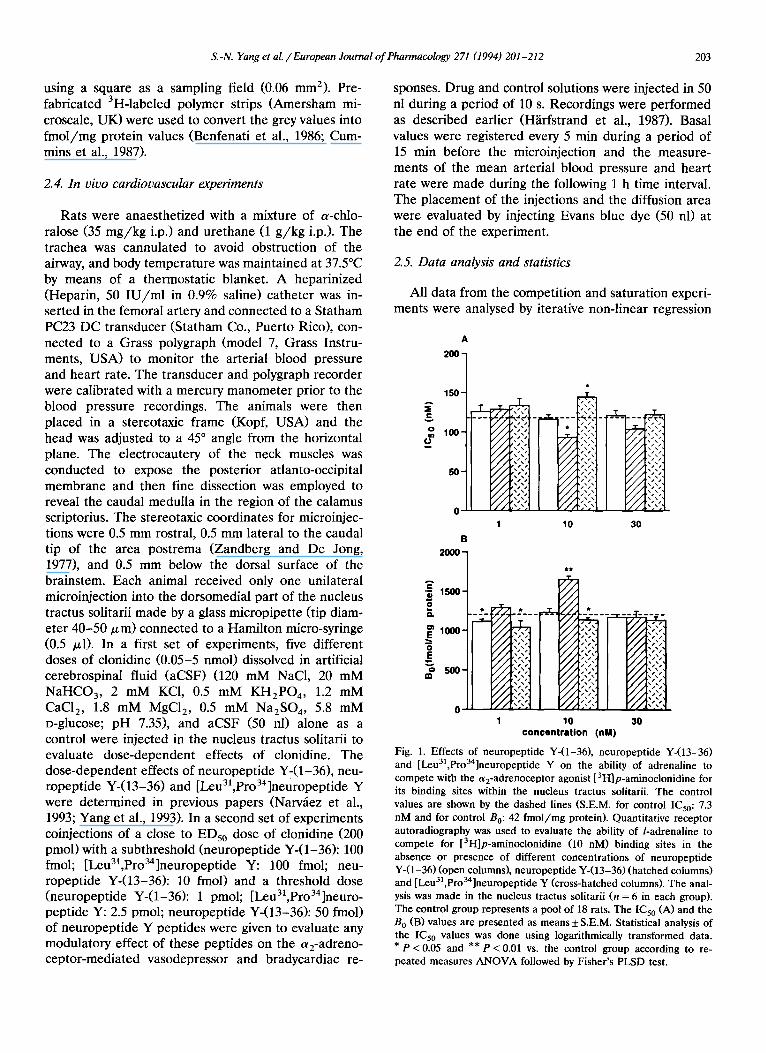
Fig. 1. Effects of neuropeptide Y-(1-36), neuropeptide Y-(13-36) and [Leu31,Pro3a]neuropeptide Y on the ability of adrenaline to compete with the a2-adrenoceptor agonist [3H]p-aminoclonidine for its binding sites within the nucleus tractus solitarii. The control values are shown by the dashed lines (S.E.M. for control IC50:7.3 nM and for control B0:42 fmol/mg protein). Quantitative receptor autoradiography was used to evaluate the ability of /-adrenaline to compete for [3H]p-aminoclonidine (10 nM) binding sites in the absence or presence of different concentrations of neuropeptide Y-(1-36) (open columns), neuropeptide Y-(13-36) (hatched columns) and [Leu31,Proaa]neuropeptide Y (cross-hatched columns). The anal- ysis was made in the nucleus tractus solitarii (n = 6 in each group). The control group represents a pool of 18 rats. The IC50 (A) and the B 0 (]3) values are presented as means + S.E.M. Statistical analysis of the IC50 values was done using logarithmically transformed data. * P < 0.05 and * * P < 0.01 vs. the control group according to re- peated measures ANOVA followed by Fisher's PLSD test.

204 S.-N. Yang et al. / European Journal of Pharmacology 271 (1994) 201-212
fitting procedures allowing the use of non-transformed data (Motulsky and Ransnas, 1987). A best fit was formed for a model of one binding site, used to deter- mine the ICs0 (drug concentration inhibiting 50% of radioligand binding) and B 0 (amount bound in the absence of unlabeled aE-adrenoceptor agonist) values in the competition experiments and the K d (the equi- librium dissociation constant) and the Bm~ x (the maxi- mum number of binding sites) values in the saturation experiments (Rodbard, 1984). Hill coefficients were calculated for both the saturation and competition curves (Hrdina, 1986). To achieve homogeneity of vari- ance and allow parametric statistical analysis, the IC50 and K d values were logarithmically transformed
A
200-
150 -
:S v o 10o
(J
50
B
2000 -
1500- o Q.
O1
__.E 1000- O
o~ 500-
T
: : : : i
I / / / , ' , "
I / / / . ' ~ "
1
] ' " 4
I t t
10
.T
t
.,~ r" - - . . . . 4 x x
T
J t
, , " " ••. " r/Ai:i: • i V / . /
,t# I f / / , . .
Y / / q . ' ~ . J V / / l . • . - J
30
T
- 7r- 1 "J"~ - ~ " / / / T / / / ' . / . / J
. , a / / / , l ~ j / / / . ' ~ ' 4
l e / / . - - j
/ / / . ' . ' 4 / / / . . . .
/ / / / / / / / /
z / /
Y/ Y,4,
/ / / / / /
10 30 Concentration (nM)
Fig. 2. Effects of neuropeptide Y-(1-36), neuropeptide Y-(13-36) and [Leu31,Pro34]neuropeptide Y on the ability of clonidine to compete with the a2-adrenoceptor agonist [3H]p-aminoclonidine for its binding sites within the nucleus tractus solitarii. The control values are shown by the dashed lines (S.E.M. for control ICs0:6.1 nM and for control B0:27 fmol/mg protein). Quantitative receptor autoradiography was used to evaluate the ability of clonidine to compete for [3H]p-aminoclonidine (10 nM) binding sites in the absence or presence of different concentrations of neuropeptide Y-(1-36) (open columns), neuropeptide Y-(13-36) (hatched columns) and [Leu31,Proaa]neuropeptide Y (cross-hatched columns). The anal- ysis was made in the nucleus tractus solitarii (n = 6 in each group). The control group represents a pool of 18 rats. The IC50 (A) and the B 0 (B) values are presented as means + S.E.M. Statistical analysis of the IC50 values was done using logarithmically transformed data. * P < 0.05 and * * P < 0.01 vs. the control group according to re- peated measures ANOVA followed by Fisher's PLSD test.
125 : NPY(13-36) [Leo 31 Pro 34]NpY
0 " I I I I I "1"
- 1 0 - 9 - 8 - 7 - 6 - 5
Clorddlne (log M)
Fig. 3. Representative competition curves showing the shift to the left of the clonidine displacement curve in the nucleus tractus solitarii in the presence of neuropeptide Y-(13-36) (10 nM) ( • ) ; the shift to the right of the clonidine displacement curve in the presence of [Leu31,Pro34]neuropeptide Y (10 nM) (o); and a control curve (0) . The concentration of [3H]p-aminoclonidine was 10 nM.
(Snedecor and Cochran, 1980). However, in the figures the untransformed values are shown. In the competi- tion experiments, repeated measures ANOVA was used which when showing heterogeneity was followed by Fisher's protected least square difference (PLSD) test (Ludbrook, 1991) to compare four different treatments in the same rats (one control group of sections and three groups of sections each treated with different concentrations of the respective neuropeptide Y pep- tides). In the saturation experiments, Student's paired t-test was used to compare the control group with the respective experimental group in the same rats.
In the in vivo cardiovascular experiments the area created by the curve for each parameter (the mean arterial blood pressure and heart rate) and for each animal was calculated using computer programs devel- oped by Guna Consult, Stockholm, Sweden. The area values (overall effects) were expressed as absolute val- ues in arbitrary units, mainly reflecting the duration of the effect under 30 min, and the peak effects (maximal responses) were shown as percent changes from the respective mean basal values. The possible dose-re- sponse relationship was evaluated with the Jonck- heere-Terpstra test (Hollander and Wolf, 1973) and the non-parametric Kruskal-Wallis test modified for treatments versus control (Siegel and Castellan, 1988) was used to compare peak and overall effects between experimental groups and the control group.
2.6. Materials
Neuropeptide Y-(1-36), neuropeptide Y-(13-36) and [Leu3t,Pro34]neuropeptide Y were purchased from Peninsula Laboratories, USA. [3H]p-Aminoclonidine, specific activity 50 Ci/mmol, was purchased from

S.-N. Yang et al. / European Journal of Pharmacology 271 (1994) 201-212 205
Amersham, UK. l-Noradrenaline bitartrate salt, l- adrenaline bitartrate salt and clonidine hydrochloride were purchased from Sigma Chemical Co., USA. The tritium-sensitive film was purchased from 3H-Hyper- film, Amersham, UK.
3. Results
3.1. Receptor autoradiographical analysis
3.1.1. Competition experiments We have studied the modulation by neuropeptide
Y-(1-36), by the C-terminal fragment, neuropeptide
A
30-
20-
C
2 lO-
0 o.1 0.3
B
3500-
A 3000-
i 2500 - o. m 2000 E
o 1500
x 10oo E
m 5OO
I ,
T /
:::::::
_I_ .
10 30
/ A
. . i ~. /
:::::::: :::::: ::::: iiii! :!:
/ "
..1.,
. . . ' /
...... : .... / /
0.3 Concentration
• " r~ - . _
.e/ " 9
• y .., : / ~:. ::.:: ,::., . .......... / '.. " . ..:/ ,.'t , / ,
I 10 (nM)
0.1 30
Fig. 4. Effects of neuropeptide Y-(1-36), neuropeptide Y-(13-36) and [Leu31,Pro34]neuropeptide Y on the binding characteristics of the a2-adrenoceptor agonist [3H]p-aminoclonidine binding sites in the nucleus tractus solitarii. The control values are shown by the dashed lines (S.E.M. for control Kd: 1.4 nM and for control Brna×: 170 fmol/mg protein). Saturation experiments with ten concentra- tions of [3H]p-aminoclonidine (1-40 nM) were performed in the absence or presence of different concentrations of neuropeptide Y-(1-36) (open columns), neuropeptide Y-(13-36) (hatched columns) and [Leu31,Pro34]neuropeptide Y (cross-hatched columns). Non- specific binding was defined using 10 /.~M of l-noradrenaline. The Kd(A) and the Bmax(B) values are shown as means ± S.E.M. Statisti- cal analysis of the K d values was done using logarithmically trans- formed data. * P < 0.05 and ** P < 0.01 vs. the control group accord- ing to Student's paired t-test (two-tailed). n = 6 rats in each group. The control group represents a pool of 72 animals.
2000 =
I 1500-
1000-
500-
~ r , • NPY(1-36) (1 nM)
t~'~AC {~)
Fig. 5. Representative non-linear saturation curves showing the ef- fect of 1 nM of neuropeptide Y-(1-36) on the binding characteristics of the [3H]p-aminoclonidine binding sites within the nucleus tractus solitarii as analysed by quantitative receptor autoradiography. 10/zM of /-noradrenaline was used for the determination of non-specific binding. K d values as calculated by computer-assisted non-linear regression procedures were 11 nM for the control group (©) and 25 nM for neuropeptide Y-(1-36) (e).
Y-(13-36), and by [Leu31,Pro34]neuropeptide Y of the ability of /-adrenaline and clonidine to compete for [3H]p-aminoclonidine binding sites in the nucleus trac- tus solitarii. The Hill coefficients in all these competi- tion experiments with and without neuropeptide Y peptides were found to be close to unity (Hill coeffi- cients: 0.95 + 0.048, 1.06 + 0.067 and 0.99 + 0.074).
The effects of these peptides are shown on the IC50 and B 0 values of /-adrenaline when competing for [3H]p-aminoclonidine binding sites in the nucleus trac- tus solitarii (Fig. 1). Neuropeptide Y-(1-36) did not produce any change in the IC50 value of/-adrenaline in the concentrations tested. However, in a concentra- tion of 1 nM, neuropeptide Y-(1-36) induced a small decrease in the B 0 value (8%) of /-adrenaline. The C-terminal fragment, neuropeptide Y-(13-36) (10 nM) produced a significant decrease in the IC50 value of /-adrenaline by about 27%. A significant increase in the B 0 value (35%) was also observed with this concen- tration of the peptide. On the other hand, the neu- ropeptide Y Y1 receptor agonist [Leu3],Pro34]neuro - peptide Y (10 nM) produced a significant increase in the IC50 value by 18%. A significant decrease in the B 0 value (15%) was observed with 1 nM of [Leu 31, Pro34]neuropeptide Y (Fig. 1).
Competition experiments were also carried out us- ing clonidine to compete for the [3H]p-aminoclonidine binding sites in the nucleus tractus solitarii. Neuropep- tide Y-(1-36) (1-30 nM) did not exert any significant effect on the IC50 value of clonidine, but, in a concen- tration of 30 nM, neuropeptide Y-(1-36) decreased the B 0 value (14%) of clonidine (Fig. 2). The other neu- ropeptide Y peptides, neuropeptide Y-(13-36) and [Leu31,Pro34]neuropeptide Y were more potent in
206 S.-N. Yang et al. / European Journal of Pharmacology 271 (1994) 201-212
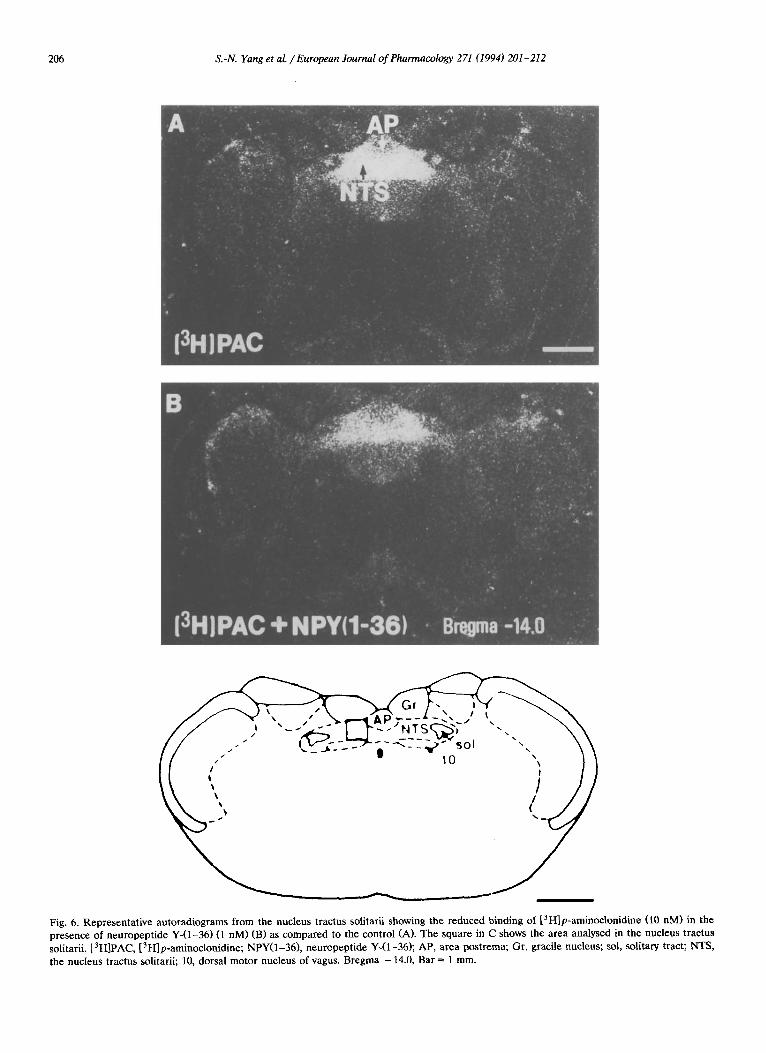
Fig. 6. Representative autoradiograms from the nucleus tractus solitarii showing the reduced binding of [~H]p-aminoclonidine (10 nM) in the presence of neuropeptide Y-(1-36) (1 nM) (B) as compared to the control (A). The square in C shows the area analysed in the nucleus tractus solitarii. [3H]PAC, [3H]p-aminoclonidine; NPY(1-36), neuropeptide Y-(1-36); AP, area postrema; Gr, gracile nucleus; sol, solitary tract; NTS, the nucleus tractus solitarii; 10, dorsal motor nucleus of vagus. Bregma - 14.0, Bar = 1 mm.
S.-N. Yang et al. / European Journal of Pharmacology 271 (1994) 201-212 207
modulating the ability of clonidine to compete for [3H]p-aminoclonidine binding sites when compared with the results obtained with /-adrenaline as a com- petitor. Neuropeptide Y-(13-36) produced a signifi- cant decrease by 27% in the IC50 value of clonidine in a concentration of 1 nM and the maximal effect of this peptide was observed at 10 nM, which produced a decrease by 50% in the IC50 value for clonidine. The B 0 value was also altered and a significant increase by 34% was observed with the concentration of 10 nM of neuropeptide Y-(13-36) (Fig. 2). [Leu31,Pro34]neuro - peptide Y produced a significant increase in the ICs0 value by 30% at a concentration of 1 nM, and the maximal effect (a 56% increase) was observed with 10 nM of the peptide. The B 0 value was also significantly affected by [Leual,Proa4]neuropeptide Y as seen from a 14% reduction at a concentration of 1 and 10 nM of the peptide. Representative competition curves by clonidine for [3H]p-aminoclonidine binding sites are shown in the presence or absence of neuropeptide Y-(13-36) (10 nM) or [Leual,Proa4]neuropeptide Y (10 nM) using non-linear regression analysis (Fig. 3).
3.1.2. Saturation experiments The effect of neuropeptide Y-(1-36) and selective
neuropeptide Y Y1 and neuropeptide Y Y2 receptor agonists on the modulation of the binding character- istics of [3H]p-aminoclonidine in the nucleus tractus solitarii was also examined using saturation analysis (Figs. 4 and 5). In this case the Hill coefficients were found to be significantly below one (0.75 _+ 0.037, P <
0.05; 0.69 ___ 0.055, P < 0.01 and 0.72 _+ 0.024, P < 0.05), possibly indicating the presence of high and low affin- ity binding sites. Neuropeptide Y-(1-36) potently and substantially increased the K d value of high affinity [3H]p-aminoclonidine binding sites in the nucleus trac- tus solitarii by about 118% at a concentration of 1 nM. A significant increase was also observed with a concen- tration of 0.3 nM (26%) and 10 nM (58%) of this peptide. No significant change was seen in the Bma x values (Fig. 4). Neuropeptide Y-(13-36) (1 nM) and [Leu31,Pro34]neuropeptide Y (10 nM) were also able to significantly increase the Kd value in this case by about 63% and 40%, respectively. The Bma x value was also not significantly changed in these experiments (Fig. 4). In Fig. 5 representative saturation curves show the effect of 1 nM of neuropeptide Y-(1-36) on the binding characteristics of [3H]p-aminoclonidine in the nucleus tractus solitarii. As seen in Fig. 6, neuropep- tide Y-(1-36) (1 nM) substantially reduced the binding of [3H)p-aminoclonidine in the nucleus tractus soli- tarii.
3.2. Cardiovascular analysis
Microinjections of clonidine into the nucleus tractus solitarii alone or together with neuropeptide Y-(1-36), neuropept ide Y-(13-36) and [Leu31,Pro34]neuro - peptide Y were carried out in order to evaluate the existence of in vivo receptor-receptor interactions. As seen in Fig. 7, the microinjection site was located in the
Fig. 7. Coronal section at the level of the area postrema showing the injection site (arrow) in the caudal dorsomedial part of the nucleus tractus solitarii. AP, area postrema; Gr, gracile nucleus; sol, solitary tract; NTS, the nucleus tractus solitarii; 10, dorsal motor nucleus of vagus. Bar = 200 /.~m.
208 S.-N. Yang et al. /European Journal of Pharmacology 271 (1994) 201-212
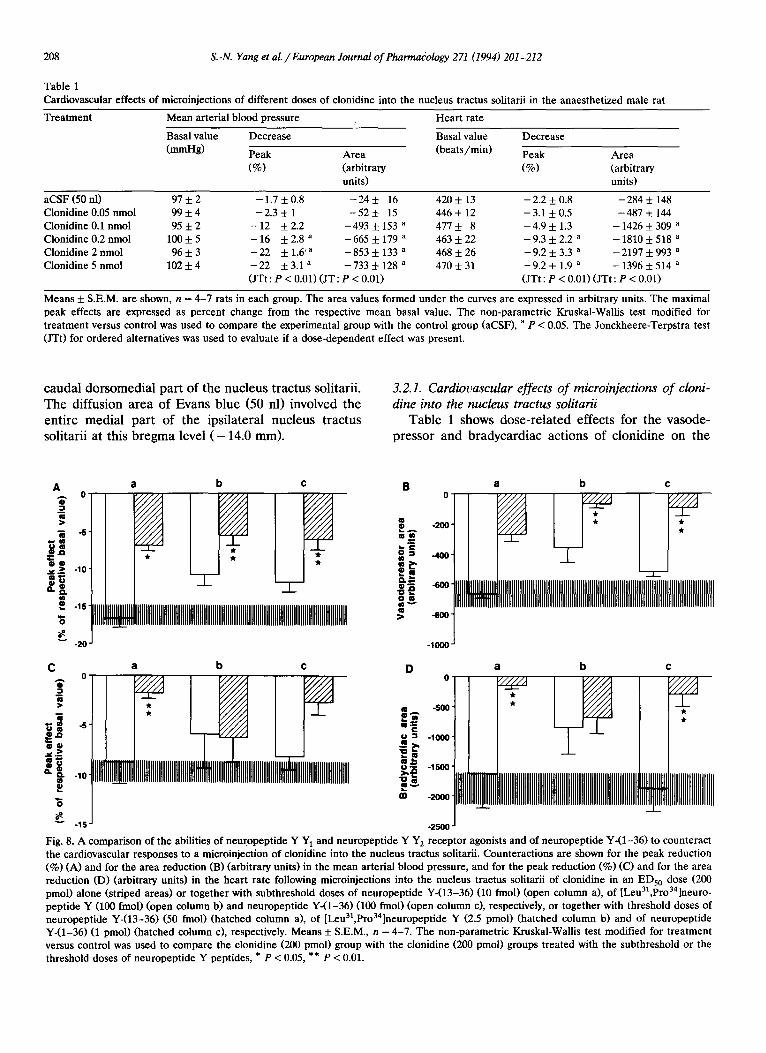
Table 1 Cardiovascular effects of microinjections of different doses of clonidine into the nucleus tractus solitarii in the anaesthetized male rat
Trea tment Mean arterial blood pressure Hear t rate
Basal value Decrease Basal value Decrease
(mmHg) Peak Area (bea t s /min ) Peak Area
(%) (arbitrary (%) (arbitrary units) units)
aCSF (50 nl) 97 + 2 Clonidine 0.05 nmol 99 + 4 Clonidine 0.1 nmol 95 + 2 Clonidine 0.2 nmol 100 + 5 Clonidine 2 nmoi 96 +- 3 Clonidine 5 nmol 102 -t- 4
- 1 . 7 + 0 . 8 -24+_ 16 420+- 13 - 2 . 2 + 0 . 8 - 2 8 4 + 148 - 2 . 3 + 1 - 5 2 +- 15 446 +- 12 - 3 . 1 + 0.5 - 4 8 7 + 144
- 1 2 +2 .2 - 4 9 3 5 : 1 5 3 a 4 7 7 + 8 - 4 . 9 + 1 . 3 - 1 4 2 6 + 3 0 9 a - 1 6 +2 .8 a - 6 6 5 + 179 a 463+-22 - 9 . 3 + 2.2 a - 1 8 1 0 + 518 a - 2 2 +1 .6 'a - 8 5 3 + 133 a 4 6 8 + 2 6 - 9 . 2 + 3.3 a - 2 1 9 7 + 993 a - 2 2 +3 .1 a - 7 3 3 + 128 a 4 7 0 + 3 1 - 9 . 2 + 1 . 9 a - 1396 + 514 a (JTt : P < 0.01) (JT : P < 0.01) (JTt : P < 0.01) (JTt : P < 0.01)
Means + S.E.M. are shown, n = 4 - 7 rats in each group. The area values formed under the curves are expressed in arbitrary units. The maximal peak effects are expressed as percent change from the respective mean basal value. The non-parametr ic Kruskal-Wallis test modified for t reatment versus control was used to compare the experimental group with the control group (aCSF), a p < 0.05. The Jonckheere-Terpstra test (JTt) for ordered alternatives was used to evaluate if a dose-dependent effect was present.
caudal dorsomedial part of the nucleus tractus solitarii. The diffusion area of Evans blue (50 nl) involved the entire medial part of the ipsilateral nucleus tractus solitarii at this bregma level ( - 14.0 mm).
3.2.1. Cardiovascular effects of microinjections of cloni- dine into the nucleus tractus solitarii
Table 1 shows dose-related effects for the vasode- pressor and bradycardiac actions of clonidine on the
A A 0 ID
ca >
~ -5"
-15" "8
g -20
c 0
Ill >
, ~ W - S ' ,D
O O
m
v . 1 5 ¸
III
a b c
!N!,,H II II II I111 II U l l illi IIII i l [I HII I
a
B
i .
O m ~ v Q
>
a
° t -200 °]
-1111111
b c
;-'///.-~ I ~"///A V///A
i llllllilWIIWIIIlilillWilillillililllllllillililllllillltllllllillililll0ilili
b c D a b c o
~ - S ~
lil I lllli IIIII
- 2 5 0 0
Fig. 8. A comparison of the abilities of neuropept ide Y Y1 and neuropept ide Y Y2 receptor agonists and of neuropept ide Y-(1-36) to counteract the cardiovascular responses to a microinjection of clonidine into the nucleus tractus solitarii. Counteract ions are shown for the peak reduction (%) (A) and for the area reduction (B) (arbitrary units) in the mean arterial blood pressure, and for the peak reduction (%) (C) and for the area reduction (D) (arbitrary units) in the heart rate following microinjections into the nucleus tractus solitarii of clonidine in an EDs0 dose (200 pmol) alone (striped areas) or together with subthreshold doses of neuropept ide Y-(13-36) (10 frnol) (open column a), of [Leu31,Pro34]neuro - peptide Y (100 fmol) (open column b) and neuropept ide Y-(1-36) (100 fmol) (open column c), respectively, or together with threshold doses of neuropept ide Y-(13-36) (50 fmol) (hatched column a), of [Leu31,Pro34]neuropeptide Y (2.5 pmol) (hatched column b) and of neuropept ide Y-(1-36) (1 pmol) (hatched column c), respectively. Means + S.E.M., n = 4-7. The non-parametr ic Kruskal-Wallis test modified for t reatment versus control was used to compare the clonidine (200 pmol) group with the clonidine (200 pmol) groups treated with the subthreshold or the threshold doses of neuropept ide Y peptides, * P < 0.05, ** P < 0.01.
S.-N. Yang et aL / European Journal of Pharmacology 271 (1994) 201-212 209
mean arterial blood pressure and heart rate (maximal peak and overall effects). The EDs0 value for the vasodepressor action was 200 pmol / ra t , and this con- centration was used to evaluate the functional interac- tion between clonidine and neuropeptide Y-(1-36) or the selective neuropeptide Y receptor agonists.
3.2.2. Interactions between clonidine (ED5o dose) and neuropeptide Y-(1-36) or selective neuropeptide Y recep- tor agonists (threshold doses) after coadministration into the nucleus tractus solitarii
The basal values of the mean arterial blood pressure and heart rate for all groups studied were within nor- mal limits and not significantly different from each other. A threshold dose of neuropeptide Y-(1-36) coadministered with clonidine (ED50), counteracted the vasodepressor effect of the ot2-adrenoceptor agonist as can be seen from the peak values (counteraction by 62%) and area values (counteraction by 86%) (Fig.
A Q
t~
CU
. a
O
..J
"6
15 7
-15
-25 O 1 'O 2'0 30 40 50 6'0
B 15"
o
ell
al
5" . Q
o .2
o o. -5 w
2 O
-15 0 ,'0 2'0 3'0 ,'0 5', 5', Tlme(mln)
Fig. 9. T i m e courses of the m e a n ar ter ia l b lood p res su re (A) and
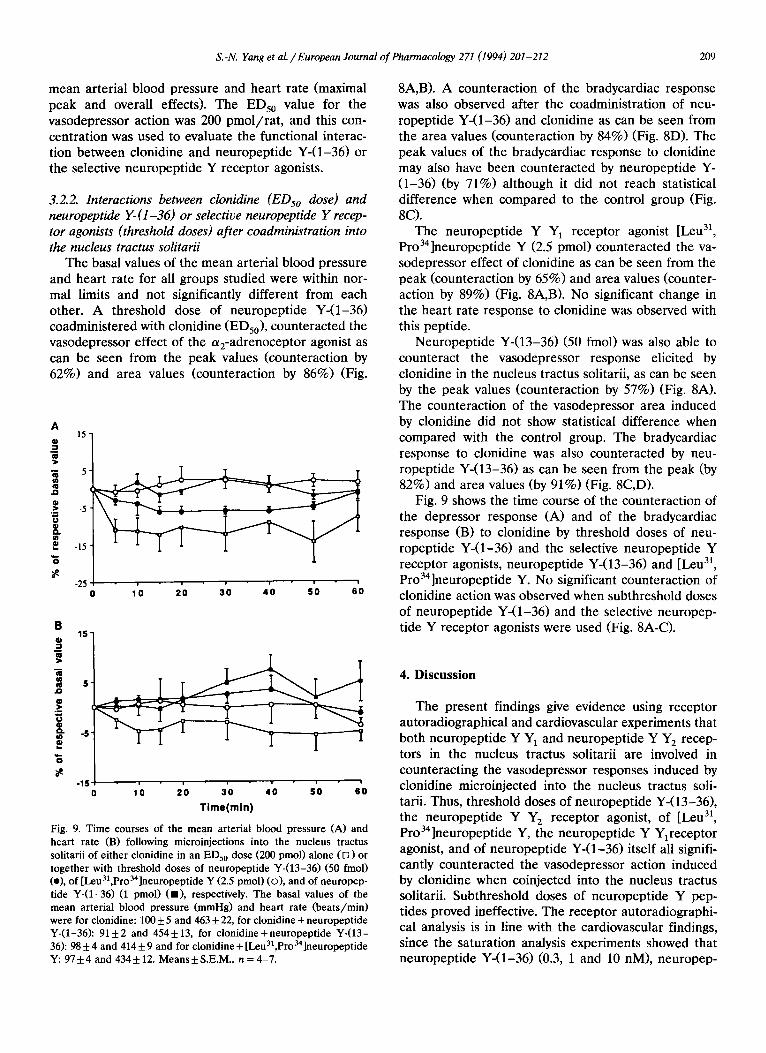
heart rate (B) following microinjections into the nucleus tractus solitarii of either clonidine in an EDs0 dose (200 pmol) alone (1~) or together with threshold doses of neuropeptide Y-(13-36) (50 fmol) (e), of [Leu3],Pro34]neuropeptide Y (2.5 pmol) (O), and of neuropep- tide Y-(1-36) (1 pmol) (11), respectively. The basal values of the mean arterial blood pressure (mmHg) and heart rate (beats/min) were for clonidine: 100 + 5 and 463 + 22, for clonidine + neuropeptide Y-(1-36): 91 + 2 and 454 + 13, for clonidine + neuropeptide Y-(13- 36): 98 + 4 and 414 + 9 and for clonidine + [Leual,Pro34]neuropeptide Y: 97+4 and 4344-12. Means+S.E.M., n = 4-7.
8A, B). A counteraction of the bradycardiac response was also observed after the coadministration of neu- ropeptide Y-(1-36) and clonidine as can be seen from the area values (counteraction by 84%) (Fig. 8D). The peak values of the bradycardiac response to clonidine may also have been counteracted by neuropeptide Y- (1-36) (by 71%) although it did not reach statistical difference when compared to the control group (Fig. 8C).
The neuropeptide Y Y1 receptor agonist [Leu 31, Pro34]neuropeptide Y (2.5 pmol) counteracted the va- sodepressor effect of clonidine as can be seen from the peak (counteraction by 65%) and area values (counter- action by 89%) (Fig. 8A, B). No significant change in the heart rate response to clonidine was observed with this peptide.
Neuropeptide Y-(13-36) (50 fmol) was also able to counteract the vasodepressor response elicited by clonidine in the nucleus tractus solitarii, as can be seen by the peak values (counteraction by 57%) (Fig. 8A). The counteraction of the vasodepressor area induced by clonidine did not show statistical difference when compared with the control group. The bradycardiac response to clonidine was also counteracted by neu- ropeptide Y-(13-36) as can be seen from the peak (by 82%) and area values (by 91%) (Fig. 8C,D).
Fig. 9 shows the time course of the counteraction of the depressor response (A) and of the bradycardiac response (B) to clonidine by threshold doses of neu- ropeptide Y-(1-36) and the selective neuropeptide Y receptor agonists, neuropeptide Y-(13-36) and [Leu 3], Pro34]neuropeptide Y. No significant counteraction of clonidine action was observed when subthreshold doses of neuropeptide Y-(1-36) and the selective neuropep- tide Y receptor agonists were used (Fig. 8A-C).
4. Di scuss ion
The present findings give evidence using receptor autoradiographical and cardiovascular experiments that both neuropeptide Y Y] and neuropeptide Y Y2 recep- tors in the nucleus tractus solitarii are involved in counteracting the vasodepressor responses induced by clonidine microinjected into the nucleus tractus soli- tarii. Thus, threshold doses of neuropeptide Y-(13-36), the neuropeptide Y Y2 receptor agonist, of [Leu 31, Pro34]neuropeptide Y, the neuropeptide Y Ylreceptor agonist, and of neuropeptide Y-(1-36) itself all signifi- cantly counteracted the vasodepressor action induced by clonidine when coinjected into the nucleus tractus solitarii. Subthreshold doses of neuropeptide Y pep- tides proved ineffective. The receptor autoradiographi- cal analysis is in line with the cardiovascular findings, since the saturation analysis experiments showed that neuropeptide Y-(1-36) (0.3, 1 and 10 nM), neuropep-

210 S.-N. Yang et al. / European Journal of Pharmacology 271 (1994) 201-212
tide Y-(13-36) (1 nM), and [Leu31,Pro34]neuropeptide Y (10 nM) all could significantly reduce the affinity of the a2-adrenoceptor agonist binding sites within the nucleus tractus solitarii without altering the Bma x val- ues, which, therefore, may be part of the mechanism involved. As shown, the Hill coefficient was signifi- cantly below unity in these experiments, indicating the possible presence of both high and low affinity binding sites of the a2-adrenoceptors, where the high affinity binding sites may dominate (Kdvalues around 10 nM). These results are in agreement with previous findings, where it was shown that neuropeptide Y-(1-36) can reduce the affinity of the high affinity a2-adrenoceptor agonist binding sites in membrane preparation from the medulla oblongata (Agnati et al., 1983; Fuxe et al., 1984, 1987a; H~irfstrand et al., 1989), an action which has been shown to be G protein-dependent (Von Euler et al., 1989). The inverse U-shaped concentration re- sponse curve obtained is a previously well documented phenomenon and true for all receptor modulations by neuropeptides so far described (Zoli et al., 1993).
The present findings for the first time show that both neuropeptide Y Y1 and neuropeptide Y 5(2 recep- tors may be involved in the antagonistic regulation of the aE-adrenoceptors and that the neuropeptide Y Y2 receptors are more potent than the neuropeptide Y Y1 receptors in this regard, since the neuropeptide Y-(1- 36) and the neuropeptide Y Y2 receptor agonist were effective already at 0.3 and 1 nM, respectively in con- trast to the neuropeptide Y YI receptor agonist (10 nM). Thus, in line with the cardiovascular findings the neuropeptide Y Y2 receptor agonist is more potent than the neuropeptide Y Y1 receptor agonist, but the combined activation of neuropeptide Y Y1 and neu- ropeptide Y Y2 receptors gives the most potent and strongest reduction in the affinity of the aE-adrenocep- tors in the nucleus tractus solitarii. The higher potency of the neuropeptide Y Y2 versus the neuropeptide Y Y~ receptors to modulate the a2-adrenoceptors in terms of affinity may be related to the existence of neuropep- tide Y Y2 receptors together with aE-adrenergic au- toreceptors also on the noradrenaline and adrenaline nerve terminals within the nucleus tractus solitarii (Martire et al., 1993). Thus, the neuropeptide Y Y2 receptor agonist may modulate the affinity of the a 2- adrenoceptors both pre- and postsynapticaUy, which may contribute to the higher potency of the neuropep- tide Y Y2 receptor agonist in the in vitro experiments compared with the neuropeptide Y Y1 receptor ago- nist, since the neuropeptide Y Y~ receptors may mainly be located postsynaptically (Potter and McCloskey, 1992; Wahlestedt et al., 1986, 1990). Also in the cardio- vascular experiments considering the vasodepressor re- sponses, the neuropeptide Y Y2 receptor agonist showed a higher potency in counteracting the clonidine induced vasodepressor response compared with the
neuropeptide Y Y1 receptor agonist and neuropeptide Y-(1-36) itself (50 fmol versus 2.5 pmol and 1 pmol respectively). On the other hand, it has been demon- strated that the vasodepressor responses induced by microinjecting clonidine into the nucleus tractus soli- tarii is a postsynaptic action (Feldman and Moises, 1988; Howe, 1985; Kobinger, 1978). Therefore, the counteraction by threshold doses of neuropeptide Y Y2 and neuropeptide Y Y1 receptor agonists of the cloni- dine-induced vasodepressor responses, probably re- fleets the existence of both postsynaptic antagonistic neuropeptide Y Y2 receptor/a2"adrenoceptor and neuropeptide Y Y1 receptor/az-adrenoceptor interac- tions in the nucleus tractus solitarii.
The demonstrated receptor-receptor interactions in the saturation experiments discussed above may possi- bly mainly involve a2-adrenoceptors of the high affinity type, although the low affinity receptors may also con- tribute to the [3H]p-aminoclonidine binding (Hill coef- ficient significantly below unity). In contrast, in the competition experiments with clonidine versus [3H]p- aminoclonidine and adrenaline versus [3H]p-amino- clonidine only the low affinity state of the a2-adreno- ceptors may contribute to the IC50 value obtained, since the Hill coefficient is not significantly different from unity and the ICs0 value of clonidine is above 100 nM. The best fit for the competition curves obtained was for one binding site. The opposing results obtained with the neuropeptide Y Y2 receptor agonist versus the neuropeptide Y Y1 receptor agonist in the regulation of the ICs0 value for clonidine and adrenaline in the competition experiments may therefore be related to the fact that the neuropeptide Y Y2 receptors upon activation increase the affinity of the low affinity bind- ing sites, while the neuropeptide Y Y1 receptors upon activation reduce the affinity of the low affinity state of the az-adrenoceptor. These observations are consistent with the fact that similar results are obtained using either clonidine or /-adrenaline to compete with [3H]p-aminoclonidine for its binding sites in the nu- cleus tractus solitarii, although the effects are stronger when using clonidine as a displacer. The results on the ICs0 values are also supported by the fact that in the competition experiments with /-adrenaline and cloni- dine, the B 0 values were increased upon neuropeptide Y Y2 receptor activation and reduced upon neuropep- tide Y Y1 receptor activation. The present explanation also accounts for the failure of the neuropeptide Y-(1- 36), activating both neuropeptide Y Y~ and neuropep- tide Y Yz receptors, to significantly modulate the IC50 values in both the clonidine and adrenaline competi- tion experiments versus [3H]p-aminoclonidine. These results are also of substantial interest since they open up the possibility that it is the modulation of the high and not the low affinity state of the a2-adrenoceptors, which is of relevance for the ability of the neuropep-

S.-N. }Tang et al. / European Journal of Pharmacology 271 (1994) 201-212 211
tide Y peptides to counteract the vasodepressor re- sponses elicited at ot2-adrenoceptors within the nucleus tractus solitarii, since all the neuropeptide Y peptides counteracted the vasodepressor action of clonidine. Furthermore, since differential effects can be exerted on the low versus the high affinity state of the a 2- adrenoceptors by neuropeptide Y Y2 receptors, the findings indicate that the G protein coupling of the a2-adrenoceptor, which determines the proportion of receptors in the high affinity state (Jakobs, 1985; Dophin and Scott, 1991), influences the type of modu- lation obtained in terms of heteroregulation by the neuropeptide Y Y1 and neuropeptide Y Y2 receptors.
The fact that the neuropeptide Y Y2 but not the neuropeptide Y Y1 receptor agonist could counteract the clonidine-induced bradycardiac action may be ex- plained by the higher density of neuropeptide Y Y2 than of neuropeptide Y Y1 receptors in the nucleus tractus solitarii (Aicher et al., 1991), so that it could interact with a larger number of a2-adrenoceptors, including those involved in the control of the heart rate.
In conclusion, evidence has been obtained for the existence of antagonistic neuropeptide Y YI receptor/ aE-adrenoceptor and neuropeptide Y Y2 receptor/t~2- adrenoceptor interactions in the cardiovascular regions of the nucleus tractus solitarii involved in the regula- tion of vasodepressor responses evoked via the a 2- adrenoceptors in this region. The antagonistic neu- ropeptide Y Y2receptor/aE-adrenoceptor interaction also appears to be involved in the counteraction of the bradycardiac response to clonidine microinjected into the nucleus tractus solitarii. In this interaction the affinity reduction by neuropeptide Y Y1 and neuropep- tide Y Y2 receptors of the high but not the low affinity state of the aE-adrenoceptor agonist binding site ap- pears to be involved.
Acknowledgements
This work has been supported by a grant (04X-715) from the Swedish Medical Research Councial and by a grant from the Knut and Alice Wallenberg Founda- tion. S.-N. Yang is supported by the Wenner-Gren Center Foundation, Sweden and D.R. Fior by CNPq (20.1409/92.5), Brasilia, Brazil.
References
Agnati, L.F., K. Fuxe, F. Benfenati, N. Battistini, A. H~irfstrand, K. Tatemoto, T. H6kfelt and V. Mutt, 1983, Neuropeptide Y in vitro selectively increases the number of a2-adrenergic binding sites in membranes of the medulla oblongata of the rat, Acta Physiol. Scand. 118, 293.
Aicher, S.A., M. Springston, S.B. Berger, D.J. Reis and C. Wahle- stedt, 1991, Receptor-selective analogs demonstrate NPY/PYY receptor heterogeneity in rat brain, Neurosci. Lett. 130, 32.
Barraco, R.A., E. Ergene, J.C. Dunbar and M.R. EI-Ridi, 1990, Cardiorespiratory response patterns elicited by microinjections of neuropeptide Y in the nucleus tractus solitarius, Brain Res. Bull. 24, 465.
Benfenati, F., M. Cimino, L.F. Agnati and K. Fuxe, 1986, Quantita- tive autoradiography of central neurotransmitter receptors: methodological and statistical aspects with special reference to computer-assisted image analysis, Acta Physiol. Scand. 128, 129.
Cummins, J.T., G. Von Euler, K. Fuxe, S.-O. ()gren and L.F. Agnati, 1987, Chronic imipramine treatment reduces (+)2-[125I]iodoly - sergic acid, diethylamide but not t25I-neuropeptide Y binding in layer IV of rat cerebral cortex, Neurosci. Lett. 75, 152.
Dolphin, A.C. and R.H. Scott, 1991, Identification of G-protein- mediated processes, in: Molecular Neurobiology. A Practical Approach, eds. J. Chad and H. Wheal (Oxford University Press, Oxford) p. 95.
Everitt, B.J., T. H6kfelt, L. Terenius, K. Tatemoto, V. Mutt and M. Goldstein, 1984, Differential co-existence of NPY (NPY)-like immunoreactivity with catecholamines in the central nervous sys- tem of the rat, Neuroscience 11, 443.
Feldman, P.D. and H.C. Moises, 1988, Electrophysiological evidence for a 1- and a2-adrenoceptors in solitary tract nucleus, Am. J. Physiol. 254, H756.
Fuxe, K., L.F. Agnati, A. H/irfstrand, M. Martire, M. Goldstein, R. Grimaldi, P. Bernardi, I. Zini, K. Tatemoto and V. Mutt, 1984, Evidence for a modulation by neuropeptide Y of t~-2 adrenergic transmission line in central adrenaline synapses. New possibilities for treatment of hypertensive disorders, Clin. Exp. Hypertens. Theory Pract. A6, 1951.
Fuxe, K., L.F. Agnati, A. H~irfstrand, A.M. Janson, A. Neumeyer, K. Andersson, M. Ruggeri, M. Zoli and M. Goldstein, 1986, Mor- phofunctional studies on the neuropeptide Y/adrenaline costor- ing terminal systems in the dorsal cardiovascular region of the medulla oblongata. Focus on receptor-receptor interactions in cotransmission, Prog. Brain Res. 68, 303.
Fuxe, K., L.F. Agnati, A. H~irfstrand, B.B. Fredholm, M. Kalia and M. Goldstein, 1987a, On the role of receptor-receptor interac- tions in synaptic transmission: biochemical and autoradiographi- cal studies on the interactions between aE-adrenergic and neu- ropeptide Y receptors in the nucleus tractus solitarius, in: Recep- tor-Receptor Interactions. A Now Intramembrane Integrative Mechanism, eds. K. Fuxe and L.F. Agnati (MacMillan Press, London) p. 222.
Fuxe, IC, A. H/irfstrand, L.F. Agnati, M. Kalia, B. Fredholm, T. Svensson, J.A. Gustafsson, R. Lung and D. Ganten, 1987b, Cen- tral catecholamine-neuropeptide Y interactions at the pre- and postsynaptic level in cardiovascular centers, J. Cardiovasc. Phar- macol. 10, $1.
Fuxe, K., G. Von Euler, I. Van der Ploeg, B.B. Fredholm and L.F. Agnati, 1989, Pertussis toxin treatment counteracts the cardio- vascular effects of neuropeptide Y and clonidine in the awake unrestrained rat, Neurosci. Lett. 101,337.
Fuxe, K., J.A. Aguirre, L.F. Agnati, G. Von Euler, P. Hedlund, R. Covenas, M. Zoli, B. Bjelke and P. Eneroth, 1990, Neuropeptide Y and central cardiovascular regulation. Focus on its role as a cotransmitter in cardiovascular adrenergic neurons, Ann. N.Y. Acad. Sci. 611, 111.
Gehlert, D.R., S.L. Gackenheimer and D.A. Schober, 1992, [Leu31,Pro34]neuropeptide Y identifies a subtype of 125I-labeled peptide YY binding sites in the rat brain, Neurochem. Int. 21, 45.
Hedlund, P., G. Von Euler and K. Fuxe, 1991, Activation of 5-hy- droxytryptamineiA receptors increases the affinity of galanin re- ceptors in di- and telencephalic areas of the rat, Brain Res. 560, 251.

212 S.-N. Yang et aL / European Journal of Pharmacology 271 (1994) 201-212
Hollander, M. and D.A. Wolf, 1973, Nonparametric Statistical Meth- ods (Wiley, New York) p. 120.
Howe, P.R.C., 1985, Blood pressure control by neurotransmitters in the medulla oblongata and spinal cord, J. Auton. Nerv. Syst. 12, 95.
Hiirfstrand, A., 1987, Brain neuropeptide Y mechanism. Basic as- pects and involvement in cardiovascular and neuroendocrine reg- ulation, Acta Physiol. Scand. $565, 1.
H~irfstrand, A. and K. Fuxe, 1987, Simultaneous central administra- tion of adrenaline and neuropeptide Y leads to antagonistic interactions in vasodepressor responses in awake male rats, mcta Physiol. Scand. 130, 529.
Hiirfstrand, A., K. Fuxe, L.F. Agnati, F. Benfenati and M. Goldstein, 1986, Receptor autoradiographical evidence for high densities of 125I-neuropeptide Y binding sites in the nucleus tractus solitarius of the normal male rat, Acta Physiol. Scand. 128, 195.
H~irfstrand, A., K. Fuxe, L.F. Agnati and B.B. Fredholm, 1989, Reciprocal interactions between a2-adrenoceptor agonist and neuropeptide Y binding sites in the nucleus tractus solitarius of the rat. A biochemic and autoradiographic analysis, J. Neural Transm. 75, 83.
Hrdina, P.D., 1986, General principles of receptor binding, in: Neu- romethods 4 Receptor Binding, eds. A.A. Boulton, G.B. Baker and P.D. Hrdina (Humana Press, Clifton, N J) p. 1.
Jakobs, K.H., 1985, Coupling mechanisms of a2-adrenoceptors, J. Cardiovasc. Pharmacol. 7, S109.
Kobinger, W., 1978, Central a-adrenergic systems as targets for hypotensive drugs, Rev. Physiol. Biochem. Pharmacol. 81, 39.
Leslie, R.A., T.J. McDonald and H.A. Robertson, 1988, Autoradio- graphic localization of peptide YY and neuropeptide Y binding sites in the medulla oblongata, Peptides 9, 1071.
Ludbrook, J., 1991, On making multiple comparisons in clinical and experimental pharmacology and physiology, Clin. Exp. Pharma- col. Physiol. 18, 379.
Martire, M., G. Pistritto, N. Mores, L.F. Agnati and K. Fuxe, 1993, Region-specific inhibition of potassium-evoked [3H]noradrena- line release from rat brain synaptosomes by neuropeptide Y-(13- 36). Involvement of NPY receptors of the Y2 type, Eur. J. Pharmacoi. 230, 231.
Motulsky, H.J. and L.A. Ransnas, 1987, Fitting curves to data using nonlinear regression: a practical and nonmathematical review, FASEB J. 1, 365.
Narv~tez, J.A., J.A. Aguirre and K. Fuxe, 1993, Subpicomolar amounts of NPY(13-36) injected into the nucleus tractus solitarius of the rat counteract the cardiovascular responses to L-glutamate, Neu- rosci. Lett. 151, 182.
Paxinos, G. and C. Watson, 1986, The Rat Brain in Stereotaxic Coordinates (Academic Press, Sidney).
Potter, E.K. and M.J.D. McCloskey, 1992, [Leu31,Pro34]NpY, a selective functional postjunctional agonist at neuropeptide- Y receptors in anaesthetised rats, Neurosci. Lett. 134, 183.
Rockhold, R.W. and R. W. Caldwell, 1980, Cardiovascular effects
following clonidine microinjection into the nucleus tractus soli- tarii of the rat, Neuropharmacoiogy 19, 919.
Rodbard, D., 1984, Lessons from the computerization of radioum- munoassays: an introduction to the basic principles of modeling, in: Computers in Endocrinology, eds. D. Rodbard and G. Forti (Raven, New York) p. 75.
Rosin, D.L., D. Zeng, R.L. Stornetta, F.R. Norton, T. Riley, M.D. Okusa, P.G. Guyenet and K.R. Lynch, 1993, Immunohistochemi- cal localization of a2A-adrenergic receptors in catecholaminergic and other brainstem neurons in the rat, Neuroscience 56, 139.
Siegel, S. and N.J. Castellan, Jr., 1988, Nonparametric Statistics for the Behavioral Sciences (McGraw-Hill, New York) p. 214.
Snedecor, G.W. and W.C. Cochran, 1980, Statistical Methods (Iowa State University, Ames) p. 291.
Unnerstall, J.R., T.A. Kopajtic and M.J. Kuhar, 1984, Distribution of a 2 agonist binding sites in the rat and human central nervous system: analysis of some functional, anatomic correlates of the pharmacologic effects of clonidine and related adrenergic agents, Brain Res. Rev. 7, 69.
Von Euler, G., K. Fuxe, I. Van der Ploeg, B.B. Fredholm and L.F. Agnati, 1989, Pertussis toxin treatment counteracts intramem- brahe interactions between neuropeptide Y receptors and a 2- adrenoceptors, Eur. J. Pharmacol. 172, 435.
Wahlestedt, C., N. Yanaihara and R. Hfikanson, 1986, Evidence for different pre- and postjunctional receptors for neuropeptide Y and related peptides, Reg. Pept. 13, 307.
Wahlestedt, C., L. Grundemar, R. H~kanson, M. Heilig, G.H. Shen, Z. Zukowska-Grojec and D.J. Reis, 1990, Neuropeptide Y recep- tor subtypes, YI and Y2, Ann. N.Y. Acad. Sci. 611, 7.
Yang, S.-N., J.A. Narvfiez, B. Bjelke, L.F. Agnati and K. Fuxe, 1993, Microinjections of subpicomolar amounts of NPY(13-36) into the nucleus tractus solitarius of the rat counteract the vasodepressor responses of NPY(1-36) and of a NPY Y1 receptor agonist, Brain Res. 621, 126.
Yang, S.-N., D.R. Fior, P.B. Hedlund, J.A. Narvfiez, L.F. Agnati and K. Fuxe, 1994, Coinjections of NPY(1-36) or [Leual,Pro34]NPY with adrenaline in the nucleus tractus solitarius of the rat coun- teract the vasodepressor responses to adrenaline, Neurosci. Lett. 171, 27.
Young, W.S., III. and M.J. Kuhar, 1980, Noradrenergic ct I and t~ z receptors: light microscopic autoradiographic localization, Proc. Natl. Acad. Sci. USA 77, 1696.
Zandberg, P. and W. De Jong, 1977, a-Methylnoradrenaline-induced hypotension in the nucleus tractus solitarii of the rat: a localiza- tion study, Neuropharmacology 16, 219.
Zandberg, P., W. De Jong and D. De Wied, 1979, Effects of catecholamine-receptor stimulating agents on blood pressure af- ter local application in the nucleus tractus solitarii of the medulla oblongata, Eur. J. Pharmacol. 55, 43.
Zoli, M., L.F. Agnati, P.B. Hedlund, X.M. Li, S. Ferr6 and K. Fuxe, 1993, Receptor-receptor interactions as an integrative mechanism in nerve cells, Mol. Neurobiol. 7, 293.
Copyright © 2022 FDOKUMEN




















