
Kidney Function in Rats Treated With a High-Fructose Diet and Streptozotocin
Upload
independentCategory
view
0download
0
Chemosphere, Vol. 38, No. 13, pp. 3003-3013, 1999 Pergamon © 1999 Elsevier Science Ltd. All rights reserved
0045-6535/99/$ - see front matter
PII: S0045-6535(98)00502-5
M ODULATION OF HEPATIC AND INTESTINAL GLUTATHIONE S-TRANSFERASES
AND OTHER ANTIOXIDANT ENZYMES BY DIETARY LIPIDS IN STREPTOZOTOCIN
DIABETIC RATS 1
M. Dolores Glr6n , Rafael Salto, Yolanda Gonz~lez, Jose A. Gir6n, Natalia Nieto, Jos6 L. Periago, M.
Dolores Su~ez and Paloma Hortelano
Department of Biochemistry and Molecular Biology, School of Pharmacy,
University of Granada, Spain
(Received in Germany 5 August 1998; accepted 2 October 1998)
ABSTRACT Antioxidant enzymes in liver and small intestine were investigated using control and
streptozotocin diabetic rats fed diets with 5% olive, sunflower or fish oil for five weeks. In liver, Glutathione
Peroxidase and Superoxide Dismutase decreased and in intestine Glutathione-S-transferase (GST) increased
by diabetes. In isolated jejunum and ileum, this increase in GST activity was due to an increase in GST-a and
-~t isoenzymes in jejunum and GST-~, la and -re in ileum. Since GST plays an important role in protecting
tissues from oxidative damage, our results highlight the role of the intestine against free radicals in
physiological or pathological situations. © 1999 Elsevier Science Ltd. All rights reserved
Key Words: Antioxidant defenses, Dietary fat, Streptozotocin diabetic rats
INTRODUCTION
Growing evidence indicates that oxidative stress is increased in the diabetic condition due to
overproduction of reactive oxygen radicals and decreased efficiency of antioxidant defenses. Oxidative stress,
as well as non-enzymatic glycosylation, is now considered a major factor contributing to the extent of chronic
diabetes complications [1,2]. Free radicals meet many of the criteria required for a role in the pathogenesis of
diabetic late syndrome: they are present in tissues affected by the diabetic process, they have a direct toxic
effect on tissues, and under certain conditions, high glucose levels can induce free radical production [3-6]•
The glutathione antioxidant system plays a fundamental role in cellular defense against reactive free
radicals and other oxidant species. It consists of reduced glutathione and an array of functionally-related
enzymes of which Glutathione Reductase (GR) is responsible for the regeneration of glutathione, whereas Se-
Glutathione Peroxidase (Se-GPx) and Glutathione S-transferases (GST) work together with glutathione in the
decomposition of hydrogen peroxide or other organic hydroperoxides [7]. Other enzymes involved in the
l,This study was supported by the CICYT through Project no. PB92- 1 1 17.
To whom correspondence should be addressed: Deparanent of Biochemistry and Molecular Biology, School of Pharmacy, Campus de Cartuja s/n, 18071 Granada, Spain. Phone 34-958-246362. FAX 34-958-248960. E-mail [email protected]
3003

3004
antioxidant defense against free radicals are: Superoxide Dismutase (SOD) and Catalase (CAT) [3,5]. SODs
catalyse the dismutation of superoxide radical to produce hydrogen peroxide and molecular oxygen [9]. CAT
has been reported to be responsible for the detoxification of hydrogen peroxide [9,10].
Glutathione S-Transferases are a large, multigene family ot" proteins that may play an important role in
protecting tissues from oxidative damage because they function in the transport of cellular components or
metabolites (or conjugation of metabolites with glutathione) formed during diabetes l 11-1_3]. This family of
enzymes is divided into four main classes of isoenzymes, ~, [3, n and 0, according to their structural,
immunological and catalytic properties 114,151. GSTs are homodimmers or heterodimmers of 45 kd
comprising at least seven subunits 113].
An adequate diet plays an important role in the treatment of diabetes. Nowadays there is a raising
interest on the supplementation of diabetic diets with lipids. As the quantity and type of dietary fatty acids may
have influence on the increased oxidative stress in diabetes, we considered interesting to investigate the
relationship between the type of dietary lipids and the status of antioxidant defenses in diabetes.
We have focused on the study of antioxidant defenses in response to different types of fat
supplemented diets in control and diabetic rats. Fat supplements have been monounsaturated, n-6 and n-3
polyunsaturated fatty acids, respectively. We have studied the enzymatic activities involved in antioxidam
defense in two locations: liver, as the most important free radical scavenger organ, and small intestine as the
first tissue responding to dietary intake. Our results show a remarkable response in certain of the antioxidant
defense enzymes. In small intestine, antioxidant enzymes and specifically GST isoenzymes changed in
response to the different fatty acid supplemented diets and increased as a result of the elevated oxidative stress
in diabetes. In contrast, liver enzymes showed less pronounced changes in response to the diet.
MATERIALS AND METHODS
Animals and diets. Sixty female Wistar rats (Granada University breeding colony) weighing 12l/- 130 g were
housed with a 12-hour light/dark cycle. Rats were divided into three groups. Each group was fed during one
week the same purified diet with a different 5% (w/w) fat supplement: olive oil (OO), sunflower oil (SO) or
fish oil (FO), respectively. Fatty acid composition of the diets is shown in Table 1. The composition of the
diet was (g/kg): 650 corn starch, 2{X) vittunin-free casein, 50 cellulose, 35 mineral mix, l(I vitamin mix, 3 DL-
methionine and 2 choline bitartrate.
Experimental diabetes was induced by a single stmptozotocin (STZ) in iection (61) mg/kg i.p. in 50 mM
sodium citrate buffer, pH 4.5). Control animals received citrate buffer alone. Only led animals with blood
glucose levels greater than 20mmol/l were considered diabetics. Diabetic rats and the cmTesponding controls
were maintained for five additional weeks on the corresponding dietary regime. Animals were studied in
compliance with our institution's guidelines for animal research.
Tissue prqparation. Rats were killed by decapitation and liver and small intestine removed. The small intestine
from the pylorus to the ileocecal valve was trimmed of fat and mesentery. The segment between the pylorus
and the Treitz angle was considered duodenum and was discarded. The rest of small intestine was divided into
two equal segments. The proximal segment was defined as jejunum and the distal segment as ileum. Each
segment was opened longitudinally and the mucosa was scraped using a glass slide. A combined small
intestine mucosa sample was obtained by pooling aliquots of.jejunum and ileum mucosae. Liver and intestine
mucosae were homogenized in 10 volumes of 0,25 M saccharose, lmM dithiothreitol, l(hnM sodium
3005
phosphate buffer, pH 7.4. Homogenates were centrifuged at 9000 xg for 20 rain and the resultant
supernatants were centrifuged at 150000 xg for lh to produce a cytosolic fraction. All procedures were
performed on ice and samples were stored at -80°C.
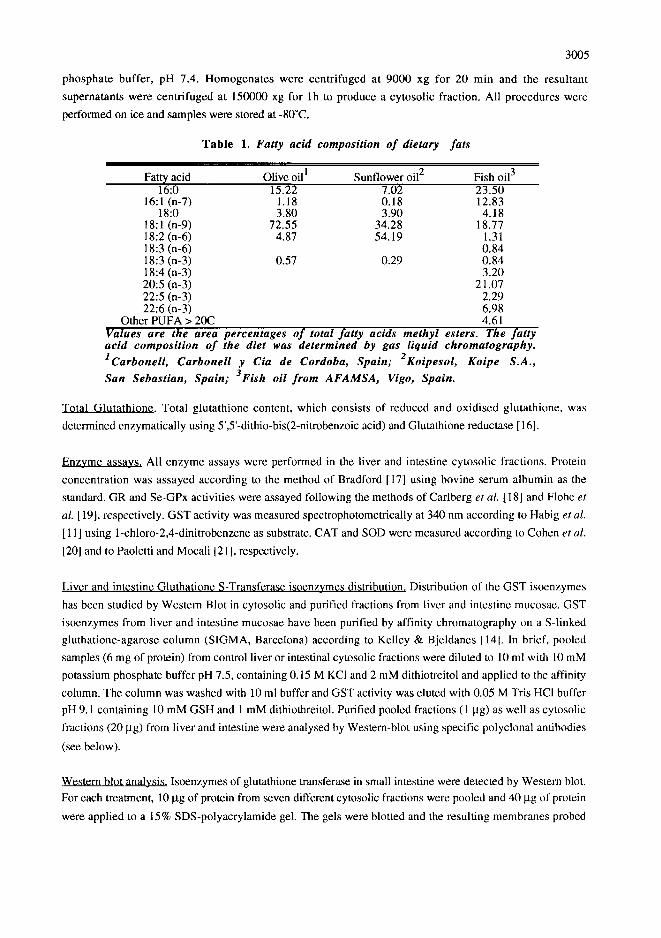
Tab le 1. Fatty acid composition of dietary fats
Fatty acid Olive oil I Sunflower oil 2 Fish oil 3 16:0 15.22 7.02 23.50
16:1 (n-7) 1.18 0.18 12.83 18:0 3.80 3.90 4.18
18:1 (n-9) 72.55 34.28 18.77 18:2 (n-6) 4.87 54.19 1.31 18:3 (n-6) 0.84 18:3 (n-3) 0.57 0.29 0.84 18:4 (n-3) 3.20 20:5 (n-3) 21.07 22:5 (n-3) 2.29 22:6 (n-3) 6.98
Other PUFA > 20C 4.61 Values' are the area percentages o f total fatty acids methyl esters. The fatty acid composition o f the diet was determined by gas liquid chromatography. 1Carbonell, Carbonell y Cia de Cordoba, Spain; 2Koipesol, Koipe S.A.,
San Sebastian, Spain; 3Fish oil from AFAMSA, Vigo, Spain.
Total Glutathione. Total glutathione content, which consists of reduced and oxidised glutathione, was
determined enzymatically using 5',5'-dithio-bis(2-nitrobenzoic acid) and Glutathione reductase [ 161.
Enzvme assavs. All enzyme assays were performed in the liver and intestine cytosolic fractions. Protein
concentration was assayed according to the method of Bradford [17] using bovine serum albumin as the
standard. GR and Se-GPx activities were assayed following the methods of Carlberg et al. [ 18] and Flohe et
al. [ 19], respectively. GST activity was measured spectrophotometrically at 340 nm according to Habig et al.
[ 11 ] using 1-chloro-2,4-dinitrobenzene as substrate. CAT and SOD were measured according to Cohen et al.
[201 and to Paoletti and Mocali [21], respectively.
Liver and intestine Gluthatione S-Transferase isoenzvmes distribution. Distribution of the GST isoenzymes
has been studied by Western Blot in cytosolic and purified fractions from liver and intestine mucosae. GST
isoenzymes from liver and intestine mucosae have been purified by affinity chromatography on a S-linked
gluthatione-agarose column (SIGMA, Barcelona) according to Kelley & Bjeldanes [14]. In brief, pooled
samples (6 mg of protein) from control liver or intestinal cytosolic fractions were diluted to 10 ml with 10 mM
potassium phosphate buffer pH 7.5, containing 0.15 M KC1 and 2 mM dithiotreitol and applied to the affinity
column. The column was washed with 10 ml buffer and GST activity was eluted with 0.05 M Tris HC1 buffer
pH 9.1 containing 10 mM GSH and 1 mM dithiothreitol. Purified pooled fractions (1 lag) as well as cytosolic
fractions (20 lag) from liver and intestine were analysed by Western-blot using specific polyclonal antibodies
(see below).
Western blot analysis. Isoenzymes of glutathione transferase in small intestine were detected by Western blot.
For each treatment, 10 ~tg of protein from seven different cytosolic fractions were pooled and 40 lag of protein
were applied to a 15% SDS-polyacrylamide gel. The gels were blotted and the resulting membranes probed

3006
with polyclonal antibodies against rat GST Ya (GST-ot), GST Ybl (GST-B) and GST Yp (GST-rc) (Biotrin
International, Dublin, Ireland). After incubation of the membranes with anti rabbit IgG antibodies linked to
peroxidase, specific b inding of the ant i -GST antiserum was detected using d iaminobenc id ine and
chemiluminescence (Boeringher Mannheim).
Statistics. The results shown are means __+ S.E.M. The effects of dietary treatments and diabetes were
examined by two-way analysis of variance followed by a Tukey test. Analyses were done with the BMDP
Computer Software program (BMDP Statistical Software, Cork, Ireland).
R E S U L T S
After induction of diabetes by a single dose STZ injection, animals were ted the supplemented diets for
five weeks. Plasma glucose levels were significantly increased in the diabetic animals (Table 2). In all diabetic
groups body weight was moderately decreased while liver weight was increased. As compared to control
animals, the intestine of diabetic animals grew in length and weight.
Table 2 Effects of Diet and Diabetes on Plasma Glucose Levels, Body,
Liver and Intestine Mucosae Weights and Intestine Length.
Olive oil Sunflower oil Fish oil
Glucose (mmol/1) C 8.2 + 0.7 9.2 + 1.0 6.4 + 0.2 cb
D 42.3 + 3.5 a 39.5 + 2.3 a 34.8 + 0.8 a
Body Weight (g) C 201.3 + 5.2 195.3 + 10.3 203.2 + 7.5
D 169.7 + 9.8 a 174.3 _+ 6.1 171.6 + 4.4 a
Liver Weight (g) C 4.7 + 0.1 5.2 + 0.1 5.0 -+ 0.2
D 8.2 + 0.4 a 8.6 + 0.3 a 8.9 + 0.3 a
Intestine Weight (g) C 4.2 + I).3 4.6 + 0.4 5.7 +_ 0.3
D 11.7 + 0.5 a 12.2 + 0.5 a 12.3 + 0.6 a
Intestine Length (cm) C 108.4 + 2.1 107.4 + 2.5 105.6 + 1.2
D 149.8 + 4.5 a 145.0 + 3.6 a 154.8 + 4.4 a Plasma was obtained by centrifuging the heparinized blood at 3000 rpm fo r 10 min at 4°C. Plasma glucose levels were assayed by a glucose oxidase-peroxidase method. Values are means _~ SEM for 6-8 animals per group. C: Control rats; D: Diabetic rats. a: control versus diabetic rats. b: olive oil versus sunflower or f ish oil fed rats. c: sunflower versus fish oil fed rats. P < 0.05.
Variations in dietary lipids cause a significant effect on liver glutathione concentration (Table 3).
Control OO fed rats showed levels which were three-fold higher those obtained for SO or FO-fed animals.
Diabetes produced changes in liver glutathione levels that were also modulated by dietary treatment. While in
the OO-fed rats there was a small but significant decrease in glutathione levels, in SO and FO-fed diabetic rats
glutathione was significantly increased. In small intestine, dietary lipids had no effects on total glutathione

3007
levels in control animals (Table 4). OO and FO groups had similar glutathione content and showed no
modification by diabetes whereas SO fed rats glutathione levels decreased 50% with diabetes.
The effect of diet and diabetes on liver antioxidant defenses is shown in Table 3. Liver GR, GST and
CAT activities showed small or no significant differences in response to diet supplementation or diabetes. On
the contrary, diabetes produced a significant decrease in SOD activity in all groups without any significant
influence of dietary treatments. A third type of response to diet and diabetes is shown by Se-GPx. Diabetes
produced a significant decrease in this enzyme activity. However, the decrease of activity was less
pronounced in the animals fed the OO diet compared to SO and FO fed rats.
Table 3 Effects o f diet and diabetes on liver glutathione concentration and
antioxidant enzymatic defenses
Olive oil Sunflower oil Fish oil
Glutathione C 40.0 + 2.4 14.1 + 1.2 b2 16.0 + 0.8 b
(mmol/mg of protein) D 32.0 + 1.4 a 25.2 + 1.5 ab 40.1 + 0.9 abe
GR C 30.7 + 1.7 31.9 + 3.6 35.8 + 2.6
(nmol/min/mg protein) D 29.9 + 1.9 37.8 + 2.0 b 36.8 + 1.6 b
GST C 128.2 + 12.0 157.8 + 13.2 147.1 + 9.0
(nmol/min/mg protein) D 147.9 + 8.1 138.7 + 6.9 134.0 + 13.6
CAT C 8.1 + 0.5 8.1 + 1.0 7.1 + 1.0
(I.tmol/min/mg protein) D 10.0 + 1.0 10.4 + 1.4 9.5 + 0.7
SOD C 84.5 + 8.7 108.2 + 11.1 86.8 + 7.9
(IU/mg protein) D 55.6 + 4.0 a 59.5 + 6.7 a 52.9 + 6.8 a
Se-GPx C 441.0 + 32.8 372.2 + 39.2 477.1 + 36.2
(nmol/min/m~ protein) D 300.6 + 23.9 a 212.3 + 14.5 ab 231.9 + 15.5 a Values are means _4- SEM for 6-8 animals per group. C: Control rats; D: Diabetic rats. a: control vs diabetic rats. b: olive oil vs sunflower or f ish oil fed rats. c: sunflower vs f ish oil fed rats. P< 0.05.
Antioxidant enzyme activities in small intestine are summarized in Table 4. GR, CAT and SOD
activities were not modified by the type of dietary fat consumed or by experimental diabetes. Se-GPx activity
was similar in control and diabetic groups, but was modulated by dietary supplementation since SO fed rats
had the lowest Se-GPx levels. GST was increased up to a 175% of the control values in diabetic rats
independently of dietary fat consumed.
Due to the role of Glutathione S-transferases in the decomposition of organic hydroperoxides and in
the metabolism of xenobiotics in the intestine [7,24] and to the significant increases found in diabetes, we
have carded out a detailed study of its activity and isoenzyme pattern.
Tissue specific distribution of GST isoenzymes in liver and intestine has been studied by Western blot
in the cytosolic fraction of both tissues and the results have been confirmed by using affinity purified samples.
To detect GST isoenzymes ct, I.t and rc known to occur in small intestine mucosa, GST Ya, GST Ybl and
GST Yp antisera have been used as first antibodies. Figure 1 shows that the most abundant GST isoenzymes
3008
in intestine correspond to the ~ and n classes while GST-B is expressed at the lowest percentage and has
only been detected by chemiluminiscence. GST Ybl antiserum reacts with two GST subunits both in
cytosolic and purified samples. These two bands correspond to subunits 3 and 4 of GST-B since the antisera
is able to recognise both subunits [25]. In liver GST isoenzyme distribution is different since there are no
detectable levels of the GST-r~ isoenzyme and there are higher levels of GST-B. In liver, only one band
corresponding to type 3 subunit of the GST-B was detected.
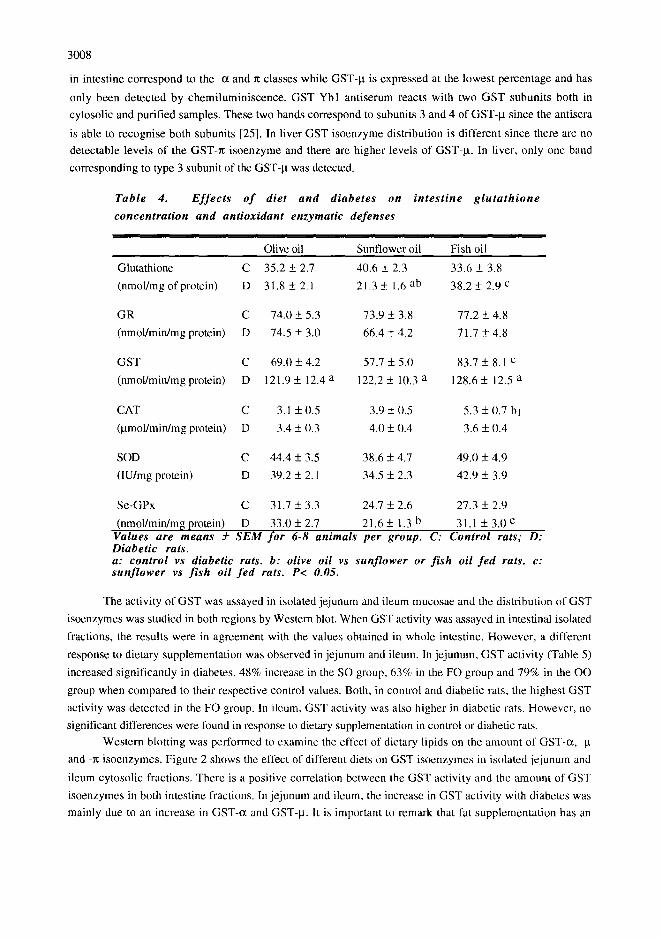
Table 4. E f f e c t s o f diet and diabetes on in tes t ine g l u t a t h i o n e
concentrat ion and antioxidant enzymatic defenses
Olive oil Sunflower oil Fish oil
Glutathione C 35.2 + 2.7 40.6 + 2.3 33.6 + 3.8
(nmol/mg of protein) D 31.8 + 2.1 21.3 +_ 1.6 ab 38.2 + 2.9 c
GR C 74.0 + 5.3 73.9 + 3.8 77.2 + 4.8
(nmol/min/mg protein) D 74.5 + 3.0 66.4 + 4.2 71.7 + 4.8
GST C 69.0 + 4.2 57.7 + 5.0 83.7 + 8.1 c
(nmol/min/mg protein) D 121.9 + 12.4 a 122.2 + 10.3 a 128.6 + 12.5 a
CAT C 3.1 + 0.5 3.9 + 0.5 5.3 + 0.7 bl
(Bmol/min/mg protein) D 3.4 _+ 0.3 4.0 + 0.4 3.6 + 0.4
SOD C 44.4 + 3.5 38.6 + 4.7 49.0 + 4.9
(IU/mg protein) D 39.2 _+ 2.1 34.5 +_ 2.3 42.9 _+ 3.9
Se-GPx C 31.7 _+ 3.3 24.7 +_ 2.6 27.3 + 2.9
(nmol/min/m~ protein) D 33,0 +_ 2.7 21.6 + 1.3 b 31.1 +_ 3.0 c Values are means _-2- S E M for 6-8 animals per group. C: Control rats; D: Diabetic rats. a: control vs diabetic rats. b: olive oil vs sunf lower or f i sh oil f e d rats. c: sun f lower vs f i sh oil f e d rats. P< 0.05.
The activity of GST was assayed in isolated jejunum and ileum mucosae and the distribution of GST
isoenzymes was studied in both regions by Western blot. When GST activity was assayed in intestinal isolated
fractions, the results were in agreement with the values obtained in whole intestine. However, a different
response to dietary supplementation was observed in jejunum and ileum. In jeiunum, GST activity (Table 5)
increased significantly in diabetes, 48% increase in the SO group, 63% in the FO group and 79% in the OO
group when compared to their respective control values. Both, in control and diabetic rats, the highest GST
activity was detected in the FO group. In ileum, GST activity was also higher in diabetic rats. However, no
significant differences were found in response to dietary supplementation in control or diabetic rats.
Western blotting was performed to examine the effect of dietary lipids on the amount of GST-ot, -B
and -n isoenzymes. Figure 2 shows the effect of different diets on GST isoenzymes in isolated jejunum and
ileum cytosolic fractions. There is a positive correlation between the GST activity and the amount of GST
isoenzymcs in both intestine fractions. In jejunum and ileum, the increase in GST activity with diabetes was
mainly due to an increase in GST-c~ and GST-ta. It is important to remark that fat supplementation has an

3009
Table 5 Effects o f diet and diabetes on j e junum and ileum Glutathione S-
Transferase activity (nmol/min/mg protein)
Olive oil Sunflower oil Fish oil
Jejunum C 102.6 + 8.4 124.4 + 11.7 140.5 + 12.1 b
D 184.1 + 16.9 a 184.5 + 21.4 a 229.8 + 9.1 ab
Ileum C 72.6 + 6.5 82.8 + 5.4 65.4 + 6.0
D 120.6 + 12.8 a 119.4 + 5.9 a 119.2 + 7.7 a Values are means +__ SEM for 6.8 animals per group. C: Control rats; D: Diabetic rats. a: control vs diabetic rats. b: olive oil vs sunflower or f i sh oil f ed rats. c: sunflower vs f i sh oil f ed rats. P < 0.05.
Figure I GST isoenzyme distribution in rat liver and small intestine mucosae.
M W ( k D ) 1 2 3 4 1 2 3 4 1 2 3 4
8 4 . 0 m
5 0 . 1
3 5 . 7
2 9 . 3 m . . , , , , , ~ . . _ . - - . - ' ~
2 1 . 2 m
7 . S n
GST--(~ GST-~ GST-~
Aliquots o f cytosols f rom liver (1) and small intestine mucosae (2) as well as purified GST fractions f rom liver (3) and intestine mucosae (4) have been electrophoresed in a 15% SDS-polyacrylamide gel and transfered to nitrocellulose membranes. GST isoenzymes have been detected by using polyclonal antibodies against GST Ya (GST-a), GST Yp (GST-Tr)
and GST Ybl (GST-IO. Bands were detected using diaminobencidine (GST-ot and GST-Jz)
or chemiluminiscence (GST-II).
influence not only on the total amount of class I,t isoenzymes but it also produces modifications in the relative
amounts of type 3 and 4 subunits in control and diabetic animals. GST-n isoenzyme showed a different
behaviour in je junum and ileum mucosae. While in jejunum, the amount of GST-rc was not modified, in
ileum, diabetes produced an increase in the amount of GST-Tt in all dietary groups.

3010
Figure 2 Modulation o f GST isoenzyme amount by dietary fa t in control and diabetic rat
j e junum and ileum mucosae.
, k q ~ m Muco~l J ~ ;~COm
(kD) OO SO FO OO SO FO O 0 SO FO O O SO FO 3S.7
~ . 3 - - . . . . - - - - . , , , ~ , m G S T - ~ J ~ " * ~ ~
21.2
3 S . 7 ~
21.2
Aliquots o f seven different cytosolic fractions per condi t ion were p o o l e d a n d applied (40 pg) to a 15% SDS-polyacrylamide gel. Gels were blotted and GST isoenzymes were
detected using polyclonal antibodies against GST Ya (GST-a), GST Yp (GST-Tr) and GST
Ybl (GST-I~I). B a n d s were d e t e c t e d us ing diaminobencidine (GST-a and GST-~) or
c h e m i l u m i n i s c e n c e (GST-I~).
D I S C U S S I O N
Diabetes mellitus is characterized by long-term complications affecting many organs. Non-enzymatic
glycosylation and autoxidative glycosylation have been considered the main attributable causes, Moreover,
diabetes leads to an increase in active oxygen species production and to changes in the status of antioxidant
defenses. Increases in the levels of oxygen free radicals can be due to increased production and/or decreased
destruction [1,3,4,12]. Changes in antioxidant mechanisms in liver and small intestine mucosa will be
discussed to ascertain the role of type of dietary fat on the oxidative stress in rats with STZ-induced diabetes.
Hepatic glutathione levels decreased or increased and small intestinal mucosal glutathione content did
not change in rats with STZ-induced diabetes. Experimental findings regarding the changes in tissue
glutathione levels in diabetic state have been inconsistent. Hepatic glutathione level was found to be decreased,
increased or unchanged in diabetic rats. Changes in tissue glutathione may reflect a complex interaction
between the diabetes-induced oxidative stress and the glutathione antioxidant system in various organs [3,7].
GR activity was higher in intestine than in liver. No differences in activity occasioned by diabetes or
by dietary fatty acid composition were observed in liver or intestine. It has been reported [7,27] that GR levels
remain unchanged in the liver of diabetic rats. Chen et al. ]10] have described that a diet enriched of the diet
with linoleic or linolenic acid increased GR activity in control rats. These data agreed to our results in liver,
although lower fat percentage in our diets (5% vs 20% in Chert et al. experiments) diminishes the differences

3011
between oleic acid-rich and linoleic or linolenic acids-rich groups. Most likely, the constancy in GR activity is
due to its role in the maintenance of reduced glutathione levels [4,9,26].
The different GST isoenzyme classes have been detected by using specific antibodies. In intestine,
assaying cytosolic and affinity purified fractions, we were able to detect by Western blot GST-tx, -It and -re
isoenzymes, the higher amounts corresponding to GST-ct and -n. Class Ix formed by type 3 and 4 subunits
according to Pickett and Lu [15] classification, was expressed at the lowest levels. Since the antibody used to
detect the GST class It reacts with both subunits, we were able to compare the relative amounts of both
subunits 3 and 4 in intestine. Our results on GST isoenzymes distribution are in agreement to those previously
described by Hatono et al. [17]. Compared to intestine, in liver GST-x was not expressed or expressed under
our detection limits. When the GST Ybl antisera was used, only GST-It type 3 subunit was detected in liver.
Our results show that in small intestine, GST activity increased in diabetic rats independently of dietary
treatment. When GST activity was assayed in isolated jejunum and ileum fractions, we detected an increase in
both in response to diabetes but, in jejunum only GST activity was modulated by dietary treatment with the
higher values corresponding to rats fed the FO diet. The raise in the GST activity in response to diabetes in
intestine can be attributed to two causes. It can either increase due to an elevation of free radicals and oxidative
molecules being therefore a reflection of a higher oxidative stress or it can be also explained as a positive
response to the oxidative damage, increasing the defenses of the organism to this damage. This latter
hypothesis has been used in the study of the importance of GST activity in the defense against xenobiotic and
carcinogenic molecules in intestine [16,24]. Both explanations are an appropriate hypothesis in our study.
Diabetes leads to an increase in free radicals and other oxidative molecules and in response to this situation,
GST levels are raised. This protective role must be more important in intestine than in liver where the GST
activity is similar in control and diabetic rats. It also appear that fat supplements can modulate GST activity
since the fatty acid composition of the diet leads to different levels in free radical production. From this point
of view, both rats fed the FO diet which are more susceptible to lipid peroxidation, have in jejunum the
highest GST activity both in control and diabetic animals. This hypothesis is in agreement to previously
published results where the effect of a FO diet on GST activity has been compared to a fat-free diet [24].
Since there is an increase in GST activity with diabetes, we studied which is the possible GST
isoenzyme responsible for this increase in activity. We have then studied the relative amount of GST
isoenzymes by Western blot. Our results show that in jejunum and ileum, the increase in activity is mainly due
to an increase in GST-tx. While in jejunum, the amount of GST-~ is not modified, in ileum, diabetes produces
an increase in GST-r~ in all dietary groups. Fat supplementation and diabetes also have an effect on the levels
of GST It. This GST class is an heterodimeric enzyme formed by two types of subunits, 3 and 4. Our results
show a modification in the relative amounts of the 3-3, 3-4 and 4-4 dimmers in response to diet and diabetes.
Our results show a specific induction of GST-~ and -kt isoenzymes by diabetes and diet. This selective
induction of different GST classes is similar to the one described in liver and small intestine in response to
different xenobiotic and anticarcinogenic compounds [16,17]. It is possible that the same mechanism of
induction arising in response to these compounds, which likely is a response to oxidative stress, is also
responsible for the increase in GST isoenzymes in response to diabetes and dietary fat.
No set patterns or trends are apparent with regard to changes in tissue Catalase and Peroxidase
activities in either alloxan- or STZ- induced diabetes [3,5]. As an example, CAT activity of rats with alloxan-
or STZ- induced diabetes was increased in liver, kidney, testis and erythrocyte hemolysate but decreased in
spleen [1,3,26]. Nevertheless, other authors have shown decreases in CAT activity in liver, kidney and heart
of rats with induced diabetes [5,8,26]. No references have been found with respect to CAT activity in intestine

3012
from diabetic rats. Our results show no modifications in CAT levels by experiment al diabetes or dietary
supplementation either in liver or in intestine.
Results obtained for GPx activity are not consistent. Different studies describe increases or decreases
in activity in erythrocytes, pancreas, liver and kidney in type II diabetes [1,3,5,26,27]. GPx is the major
defense against the damage produced by hydrogen peroxide [2]. Our results show that liver GPx activity
decreases in the STZ-induced diabetic rats. The decrease in activity occasioned by diabetes is less marked in
OO fed rats. Diabetic OO fed show significant differences with respect to SO and FO led animals. In intestinal
mucosa, GPx activity is much lower than in liver and similar in control and diabetic rats. Interestingly, dietary
effect shows the same pattern than in liver, being the SO fed rats those with lowest activity.
Superoxide Dismutases play an essential role as a defense system against oxidative stress. SOD
activity has been measured in tissues of animals with chemically induced diabetes. Other authors have found
various discrepancies in SOD activity. The loss of SOD activity may be a function of the length [8,27].and
severity of diabetes [3,5]. Other authors have shown that SOD levels did not significantly change with age,
degree of insulitis or level of diabetes [8] or even increased alter 10 weeks of diabetes [ 11. D'Aquino et al.
[28] compared the influence of n-3 fatty acids (highly susceptible to oxidative deterioration) vs saturated fatty
acids and found no significant differences in SOD activity. We show a decrease in SOD levels five weeks after
the onset of diabetes, but fat supplementation has no effect on this activity when comparing groups.
In conclusion, the increased oxidative stress during diabetes causes in liver a decline in the activities of
GPx and SOD but actually challenged the activities of GR, GT and CAT. Moreover, olive oil (supplied 5% in
the diet) partially avoids the decrease in GPX activity accompanying diabetes. This is positive as GPx is one
of the most important antioxidant defenses against the damage produced by peroxides 19].
In intestine, the most important results are related to the increase in GST activity in diabetic rats. In
jejunum, those rats fed the FO diet have the highest GST levels. Glutathione S-transferases play an important
role in the detoxification of organic hydroperoxides. Diabetes and n-3 polyunsaturated fatty acids in the diet
increase the levels of peroxides. In intestine, GPx and SOD, have similar levels in control and diabetic rats.
These results highlight the role of the intestine alleviating the pernicious effects of free radicals.
R E F E R E N C E S
1. R. Kakkar, J. Kalra, S.V. Mantha and K. Prasad, Lipid Peroxidation and Activity of Antioxidant Enzymes
in Diabetic Rats, Mol. Cell.Biochem. 151, 113-119 (1995).
2. J. Remacle, M. Raes, O. Toussaint, P. Renard and G. Rao, Low Levels of Reactive Oxygen Species as
Modulators of Cell Function, Mutation Res. 316, 103-112 (1995).
3. L.W. Oberley, Free Radicals and Diabetes, Free Rad. Biol. Med. 5, 113-124 (1988).
4. J.V. Hunt and S.P. Wolff, Oxidative Glycation and Free-Radical Production - A Causal Mechanism of
Diabetic Complications, Free Rad. Res. Comm. 12-13 (Pt 1), 115-123 (1991).
5. B.P. Yu, Cellular Defenses Against Damage From Reactive Oxygen Species, Physiol. Rev. 74, 139-162
(1994).
6. T.K. Makar, K. Rimpel-Lamhaouar, D.G. Abraham, V.S. Gokhale and A.J.L. Cooper, Antioxidant
Defense Systems in the Brain of Type II Diabetic Mice, J. Neurochem. 65, 287-291 (1995).
7. D.H. Mak, S.P. lp, P.C. Li, M.K. Poon and K.M. Ko, Alterations in Tissue Glutathione Antioxidant
System in Streptozotocin-Induced Diabetic Rats, Mol. Cell. Biochem. 162, 153-158 (1996).

3013
8. K. Asayama, S. Yokota and K. Kato, Peroxisomal Oxidases in Various Tissues of Diabetic Rats, Diabetes
Res.Clin. Pract. 11, 89-94 (1991).
9. M. Martfnez-Cayuela, Oxygen Free Radicals and Human Disease, Biochimie 77, 147-161 (1995).
10. L.C. Chen, G. Boissonneault, M.G. Hayek and C.K. Chow, Dietary Fat Effects on Hepatic Lipid Peroxidation and Enzymes ofH202 Metabolism and NADPH Generation, Lipids 28, 657-662 (1993).
11. W.H. Habig, M.J. Pabst and W.B. Jakoby, Glutathione S-Transferases. The First Enzymatic Step in
Mercapturic Acid Formation, J. BioLChem. 249, 7130-7139 (1984).
12. M.K. Kelley and L.F. Bjeldanes, Modulation of Glutathione S-Transferase Activity and Isozyme Pattern in Liver and Small Intestine of Rats Fed Goitrin- and T3-Supplemented Diets, Food Chem.Toxicol. 33,
129-137 (1995).
13. C.B. Pickett and A.Y.H. Lu, Glutathione S-Transferases: Gene Structure, Regulation and Biological
Function, Annu. Rev. Biochem. 58, 743-764 (1989).
14. W.A. Nijhoff, M.A. Bosboom, M.H. Smidt and W.H.M. Peters, Enhancement of Rat Hepatic and
Gastrointestinal Glutathione and Glutathione S-Transferases by ct-Angelicalactone and Flavone,
Carcinogenesis 16, 607-612 (1995).
15. S. Hatono, A. Jimenez and M.J. Wargovich, Chemopreventive Effect of S-Allylcysteine and its
Relationship to the Detoxification Enzyme Glutathione S-Transferase, Carcinogenesis 17, 1041-1044
(1996).
16. O.W. Griffith, Determination of Glutathione and Glutathione Disulfide Using Glutathione Reductase and
2-Vinylpyridine, Anal, Biochem. 106, 207-212 (1980).
17. M.M. Bradford, A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein
Utilizing the Principle of Protein-Dye Binding, Anal Biochem. 72, 248-254 (1979).
18. E. Carlberg and A. Mannervik, Glutathione Reductase, Methods Enzymol. 113, 484-495 (1985).
19. G. Cohen, D. Dembiec and J. Marcus, Measurement of Catalase Activity in Tissue Extracts, Anal.
Biochem. 34, 30-38 (1970).
20. L. Flohe and W.A. GiJnzler, Assays of Glutathione Peroxidase, Methods Enzymol. 105, 114-121
(1984).
21. F Paoletti and A. Mocali, Determination of Superoxide Dismutase Activity by Purely Chemical System
Based on NAD(P)H Oxidation, Methods Enzymol. 186, 209-221 (1986).
22. A.J. Dannerberg, E.J. Yang and D. Aharon, Dietary Lipids Induce Phase 2 Enzymes in Rat Small
Intestine, Biochim.Biophys. Acta 1210, 8-12 (1993).
23. W.A. Nijhoff, M.J. Grubben, F.M. Nagengast, J.B. Jansen, H. Verhagen, G. van Poppel and W.H.M.
Peters, Effects of Consumption of Brussels Sprouts on Intestinal and Lymphocytic Glutathione S-
Transferases in Humans, Carcinogenesis 16, 2125-2128 (1995).
24. P. Pietarinen, K. Raivio, R.R. Devlin, J.D. Crapo, L.-Y. Chang and V.L. Kinnula, Catalase and
Glutathione Reductase Protection of Human Alveolar Macrophages During Oxidant Exposure in vitro,
Am. J. Respir. Cell Mol. Biol. 13, 434-441 (1995).
25. A.K. Saxena, P. Srivastana, R.K. Kale and N.Z. Baquer, Impaired Antioxidant Status in Diabetic Rat
Liver. Effect of Vanadate, Biochem. Pharmacol. 45, 539-542 (1993).
26. M. D'Aquino, P.C. Benedetti, M. DiFelice, V. Gentili, G. Tomassi, M. Maiorino and F. Ursini, Effects
of Fish Oil and Coconut Oil on Antioxidant Defense System and Lipid Peroxidation in Rat Liver, Free
Rad. Res. Comm. 12-13, 147-152 (1991).
Copyright © 2022 FDOKUMEN







![[Cool] Gas Chromatography and Lipids](https://static.fdokumen.com/doc/165x107/6325a4b1852a7313b70e98e9/cool-gas-chromatography-and-lipids.jpg)












