
Microalgal lipids biochemistry and biotechnological perspectives

0163-7827/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.doi:10.1016/j.plipres.2007.07.001
Abbreviations: AADC, aromatic acid decarboxylase; ABA, abscisic acid; a-Allenic-DOPA allenic analog of amino acid L-3,4-dihydoxyphenylalanine (L-DOPA); GABA, c-aminobutyric acid; HepG2, human hepatocellular liver carcinoma cell line; HUVEC, umbilicvein endothelial cells; LM, leucomycin; LOX, lipoxygenase; LPS, lipopolysaccharide; MGC, mammalian gene collection; MTT assay islaboratory test and a standard colorimetric assay for measuring cellular proliferation; NF-kappa B, factor-kappa B; OAT, optically activtriglycerides; Raji cells are a line of EBV-transformed lymphocytes with surface Fc receptors; TNF-alpha, tumor necrosis factor; VDRvitamin D nuclear receptor.
q See Ref. [1] for a previous article giving additional information.* Corresponding author. Tel./fax: +972 2 590 2947.
E-mail address: [email protected] (V.M. Dembitsky).
Progress in Lipid Research 46 (2007) 328–375
www.elsevier.com/locate/plipres
Progress inLipid Research
Review
Allenic and cumulenic lipids q
Valery M. Dembitsky a,*, Takashi Maoka b
a Department of Medicinal Chemistry and Natural Products, School of Pharmacy,
P.O. Box 12065, Hebrew University, Jerusalem 91120, Israelb Research Institute for Production Development, 15 Shimogamo-Morimoto-Cho, Sakyo-Ku, Kyoto 606-0805,
Kyoto Pharmaceutical University, Japan
Received 14 May 2007; received in revised form 13 June 2007; accepted 2 July 2007
Abstract
Nowadays, about 200 natural allenic metabolites, more than 2700 synthetic allenic compounds, and about 1300 cumu-lenic structures are known. The present review describes research on natural as well as some biological active allenic andcumulenic lipids and related compounds isolated from different sources. Intensive searches for new classes of pharmaco-logically potent agents produced by living organisms have resulted in the discovery of dozens of such compounds possess-ing high anticancer, cytotoxic, antibacterial, antiviral, and other activities. Known allenic and cumulenic compounds canbe subdivided on several structural classes: fatty acids, hydrocarbons, terpenes, steroids, carotenoids, marine bromoal-lenes, peptides, aromatic, cumulenic, and miscellaneous compounds. This review emphasizes the role of natural and syn-thetic allenic and cumulenic lipids and other related compounds as an important source of leads for drug discovery.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Fatty acids; Lipids; Triacylglycerides; Hydrocarbons; Allenic; Cumulenic; Anticancer; Antiviral; Antibacterial; Terpenoids;Carotenoids; Steroids; Peptides; Aromatic; Synthetic
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3292. Allenic fatty acids and derivatives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 330
r-alae,

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 329
2.1. Biosynthesis and synthesis of allenic lipophilic compounds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3322.2. Triacylglycerides of plant species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 335
3. Allenic prostaglandins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3394. Allenic hydrocarbons . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3405. Allenic steroids and derivatives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3416. Allenic terpenoids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3437. Marine bromoallenes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3458. Allenic norisoprenoids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3489. Allenic carotenoids. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 350
9.1. Distribution in nature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 350
99.1.1. Marine and freshwater algal species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 350
.1.2. Higher plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 354
.1.3. Invertebrates and animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 355
99.2. Functions and biological activities. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35810. Aromatic and peptide metabolites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35811. Allenic amino acids and derivatives. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36012. Allenic alkaloids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36013. Cumulenic lipids and related compounds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36114. Addition to distributions, biosynthetic preparation and synthesis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36315. Concluding remarks. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 364
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 364References. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 365
1. Introduction
Fatty acids constitute an abundant class of natural compounds and are major constituents of many com-plex lipids. Fatty acids differ in the number of olefinic bonds, extent of branching, the length of the hydrocar-bon chain, and the number of functional groups. Allenic fatty acids which are one of the most interestinggroups among the naturally occurring lipids, are often neglected in the literature [2,3]. Allenic fatty (carbox-ylic) acids and other liphophilic metabolites contain the [–HC@C@CH–] group, and cumulenic fatty (carbox-ylic) acids and related compounds contain the [–HC@C@C@CH–] group. Both these lipid groups are rare innature [2–4]. Graphic chemical structures both groups are shown in Fig. 1.
Natural occurrence, structures, and biosynthetic pathways have partly been reported in several publications[4–8]. Among naturally occurring allenic fatty acid derivatives are a large group compounds which show cyto-toxic, antibacterial, antiviral and other activities, [2,8,9]. Synthesis of allenes, cumulenes, and acetylenes hasalso been reviewed [8–14].
The chemistry of allenes began from Jacobus Henricus van’t Hoff (awarded the first Nobel Prize in Chem-istry, 1901), who predicted the structures of allenes and cumulenes paper, in 1875 [15]. The first allenic dicar-boxylic acid, named glutinic acid (2,3-pentadienedioic acid) was isolated from leaf resin of Alnus glutinosa(Betulaceae, also called ‘European alder’), in 1908 by Hans [16]. The first allenic carotenoid, named fucoxan-thin was found in extracts of three species of marine brown algae, Fucus, Dictyota and Laminaria, in 1914 byWillstatter and Page [17]. More recently, fucoxanthin was isolated from other algae, invertebrates, plants, andother species [18–24].
The first allenic sesquiterpene named panacene was isolated from mixture of H2O–Et2O–MeOH extract of aperennial shrub Korean ginseng (also called Panax ginseng) in 1915 by Kondo and Tanaka [25]. In 1931, Minre-discovered the sesquiterpene previously named panacene from Panax ginseng and Panax quinquefolius [26].In 1964, Lee and Lee confirmed the presence of sesquiterpene panacene in Panax ginseng [27]. More recently,brominated analog of panacene was isolated from the sea hare Aplysia brasiliana, and it acts as a fish antife-edent [28]. The second allenic fatty acid, named mycomycin, which was produced by a mold-like Actinomyceteand was active against the Bacilli sp. of human tuberculosis was discovered in 1947 by Johnson and Burdon[29]. The structure of mycomycin and its physical, chemical, and tuberculostatic properties were reported soonafter [30–34].

R CH C CH R1 R CH C C CH R1
R1RR1R
Allenes Cumulenes
R R1 R R1
Fig. 1. Graphical display of chemical structures of allenes and cumulenes. R, R1 = H, alkyl, heteroatoms, and etc.
330 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
The first cumulenic acids, such as two c-lactones of 4-hydroxy-2,4,6,7,8-decapentaenoic and 4-hydroxy-2,4,5,6,8-decapentaenoic acids were isolated from several Astereae (Asteraceae) [35].
In the past few decades, natural metabolites containing an allene and cumulene groups have been isolatedfrom plants, marine algae, invertebrates, and other organisms. Extensive pharmacological screening per-formed on both terrestrial and aquatic species resulted in the discovery of novel antitumor, antibacterial,and antiviral agents. The purpose of this review is to summarize more than 300 different natural, syntheticallenic and cumulenic products with pharmacological active properties. These allenic compounds are subdi-vided into several structural classes: fatty acids, hydrocarbons, terpenes, steroids, carotenoids, marine bro-moallenes, peptides, aromatic, as well as cumulenic compounds.
2. Allenic fatty acids and derivatives
The structure of glutinic acid (1) was confirmed in 1958 [36], and its absolute configuration was establishedin 1962 [37]. Glutinic acid has been used as a starting substrate for synthesizing natural allenes [38,39], hetero-cyclic prostanoids [40], alkaloids [41], nucleosides [42] and other heterocyclic compounds [43–45].
Mycomycin (2), antibiotic 07F275 (3), and its methyl ester (4), produced by submerged fermentations offungal culture LL-07F275, were isolated and characterized despite their inherent instability. Since 07F275is a C13-containing allenic diyne, it is closely related to mycomycin, but without double bonds [46]. AntibioticLA-1 (5, 9-oxo-2,4,5,7-decatetraenoic acid) was isolated from the culture broth of LM-producing Streptomy-ces kitasatoensis NU-23-1. During fermentation, before LM is produced, LA-1 is transiently accumulated [47].In 1949, nemotinic acid (6, 4-hydroxy-5,6-undecadiene-8,10-diynoic acid), and nemotin (8) were isolated fromcultures of three Basidiomycete species: Poria tenuis, P. corticola, and an unidentified fungus (B-841). Bothcompounds exhibited antibiotic activity against Staphilococcus aureus [48,49]. Nemotinic acid and 4-b-D-xylo-pyranosyloxy-5,6-undecadiene-8,10-diynoic acid (7) were isolated from the cultures of Basidiomycete B-841[50]. Cortinellin (also known as nemotinic acid) has a broad antibiotic spectrum on Gram-positive andGram-negative bacteria, mycobacteria, and fungi (limited concentration at 0.3 cg/mL) [51]. Nemotin and dro-sophilin D (9) are also produced by some Basidiomycete species [52]. Several natural antibiotics, includingnemotinic acid and neomycin, were tested against 60 strains of bacterial viruses by a paper-disk method. Thesecompounds showed good activity against Enterococcus phage (phylum Firmicutes) [53].
1 Glutinic acid
HOOC COOH
CO2H
HH
H
2 Mycomycin
Z E
H
HO
COORH
H
3 Antibiotic 07F275, R = H4 R = Me
5 Antibiotic LA 1
COOHO
RO
COOH
H
H H
6 Nemotinic acid, R = H7 R = Xyl
O
H
O
HH
8 Nemotin
COOH
9 Drosophilin D

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 331
Odyssic acid (10, 4-hydroxy-5,6-dodecadiene-8,10-diynoic acid), and its methyl ester (11), odyssin (12),nemotinic acid (6), and its c-lacton (8) have been isolated from the higher fungi belonging to Basidiomycetes[54]. Two years later, odyssic acid, its methyl ester, and odyssin were also found in other unidentified higherfungi [55]. Ten years later, nemotinic acid and nemotin were isolated from the cultured media of Poria suba-cida, P. colorea, and P. mutans, P. subacida also yields Me nemotinate (11). Interestingly, Me nemotinate wasalso derived from the fungus Coprinus flocculosus [56,57]. 3,4-Nonadiene-6,8-diynoic acid (13), its methyl ester(14), and nemotinic acid have been isolated from the fungi Aleurodiscus roseus [58].
The natural C18 allene fatty acid has been isolated from seed oil of Leonotis nepetaefolia, which contains upto 16% laballenic (15, (�)-5,6-octadecadienoic) acid [59]. Lamium purpureum seed oil contains 16% of an acidcharacterized as (�)-octadeca-5,6-trans-16-trienoic (lamenallenic) acid (16). This acid was isolated as its Meester from the transesterification products of L. purpureum oil by countercurrent distribution by using a com-bination of recycling and single withdrawal techniques. Me lamenallenate (17) is strongly levorotatory [60].
Phlomic acid (18, 7,8-eicosadienoic acid or 20:2D7,8 allene) was observed in several genera of the Lamia-ceae, subfamily Lamioideae: hemp nettle (Galeopsis tetrahit), horehound (Marrubium vulgare), Lamium
galeobdolon, L. maculatum, Leonurus cardiaca, L. sibiricus, Moluccella laevis, Panzerina canescens, Phlomis
fruticosa, P. samia, P. tuberose, Physostegia virginiana, Stachys byzantina, and S. palustris. The occurrenceof this fatty acid was correlated with the presence of 20:1 acid, 20:1D9 cis- or 20:1n � 11. Phlomic acid wasapparently produced by chain elongation of the major seed oil fatty acid, laballenic acid, or 18:2D5,6 allene(16) [61]. The 2-(1,2-tetra-decadienyl)-cyclopropane-carboxylic acid (19) and laballenic acid were isolated fromLeonotis nepetaefolia seed oil [62]. A triene allenic fatty acid (20) and methyl ester (21) were shown to occur asa sex pheromone in the male dried bean beetle Acanthoscelides obtectus (Coleoptera, Bruchidae) [63]. The neu-tral lipids of both Phlomis species had a significant number of the same basic components. In fact, the seeds ofboth species contain monohydroxyacyldiacyl glycerides, but those of P. oreophilla had monoepoxyacyl- andmonooxoacyldiacyl glycerides, which were absent in seeds of P. regellii. Lipids of P. oreophilla seeds had ahigh linoleic acid content and those of P. regellii seeds had a high oleic acid content [64].
Two rare bromoallenic aliphatic fatty acids (12E,15S,18S)-15-hydroxy-18-bromo-12,16,17-octadecatrienoicacid (22) and (13Z,15R,18R)-15-hydroxy-18-bromo-13,16,17-octadecatrienoic acid (23) have been isolatedfrom several lichen species: Acarospora gobiensis, Cladonia furcata, Lecanora fructulosa, Leptogium saturni-num, Peltigera canina, Rhizoplaca peltata, Xanthoparmelia camtschadalis, Xanthoparmelia tinctina, and Xanth-
oria elegans [65,66]. The distribution of these isolated allenic fatty acids is shown in Table 1.
12 Odyssin
O
O
H H
HO
COORH H
10 Odyssic acid, R = H11 R = Me
H
COOR
13 R = H14 R = Me
20 R = H, R1 = n-Bu21 R = R1 = Me
COOR
R1
18 Phlomic acid
H
H
COOH
16 Lamenallenic acid, R = H17 R = Me
COORH
H
COOHH
H
15 Laballenic acid
COOH19

332 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
Marasin allenic diacetylene (24, 3,4-nonadiene-6,8-diyn-1-ol), an unstable antibiotic, was isolated from theculture medium of Marasmius ramealis as a highly unsaturated, optically active, straight-chain alcohol [67].Marasin belongs to the category of compounds that contain acetylene-allene bonds. Common to these sub-stances, besides their antibiotic action and instability, is their alkali-catalyzed isomerization. This is accompa-nied by changes in ultraviolet and infrared spectra. Marasin was active in vitro against numerous bacteriaincluding Mycobacterium tuberculosis. Marasin, allenic alcohols (25–29), as well as two m-dioxane derivatives(30, 31), were isolated from five basidiomycete fungi (Cortinellus berkeleyanus, Odontia bicolor, Flammula sapi-
nea, Daedalea juniperina, and the unidentified fungus B285) [68]. The sea coral Plexaura homomalla containsprostaglandins of the A and E series. It has been proposed that a key step in the pathway of biosynthesisinvolves the 8(R)-lipoxygenase metabolites of arachidonic acid. The 8-hydroxy-5,9,11,14–20:4, 8-hydroper-oxy-5,9,11,14–20:4, (5Z,8R,11Z,14Z)-8,9-epoxy-5,9,11,14-eicosatetra-enoic acid (32), and prostaglandins wereisolated, but an allenic intermediate (33) has not been discovered [69–71].
31
H
OOH
O
O
30
H
H H
OH26
H
H H
OHR
27 R = H28 R = OH
H
H H
OH29
OH
R
24 Marasin, R = H25 R = Me
HOOCO
32
33
3
3
HOOC
H
H
Br
HOOC
OH
H
Br
H
HOOC
OH22 n = 8
23 n = 7
n
n
2.1. Biosynthesis and synthesis of allenic lipophilic compounds
Biosynthesis of the allene (�)-marasin in Marasmius ramealis was recently reported [72]. [1-14C]-E-Dehy-dromatricaria methyl ester and dimethyl [1-14C]-deca-4,6,8-triyne-1,10-dioate were incorporated into theallene (�)-marasin in M. ramealis without scrambling of the 14C label. These levels of incorporations (0.8%and 4.9%, respectively) strongly suggest that the above esters, or their close relatives, can be converted directly

Table 1Occurrence of bromoallenic acids in lichen species from Tian Shan mountains [65]a
Lichen species 22 23
Acarospora gobiensis 2.8 0.7Cladonia furcata ND 1.9Lecanora fructulosa 3.2 NDLeptogium saturninum 2.5 NDPeltigera canina ND 1.8Rhizoplaca peltata 5.7 NDXanthoparmelia camtschadalis ND 3.6Xanthoparmelia tinctina ND 0.7Xanthoria elegans 1.5 ND
a mg/100 g of dry wet; ND, not detected.
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 333
into (�)-marasin in M. ramealis, and that the diyne-allene moiety in the latter compound arises by rearrange-ment, under enzymic control, of an alkyltriyne moiety. Biosynthesis of marasin takes place via four steps asshown in Scheme 1.
Novel allenic analogs (E)- and (Z)-(34–41) of ABA were synthesized and tested for ABA activity [73].The allene functional group was introduced by base-catalyzed isomerization of an enyne to an enallene con-jugated to a ketone, or by reduction of a 2-butyne-1,4-diol to an allenic alcohol. A 3:1 mixture of (E)-(34)and (Z)-(36) strongly inhibited growth of axenic duckweed (Lemna gibba), consequently causing a 50%reduction in frond number at 60 lg/L over a period of 7 days in continuous light. Racemic (24–37, and38–41) ABA compounds caused the same inhibition at 130 lg/L. This mixture caused the production of tur-ion-like structures, an effect previously known to be induced only by short photoperiods or by certain con-centrations of ABA. a-Allenic acids (42–49) were obtained by oxidation of the corresponding alcohols byPseudomonas aeruginosa (ATCC 17504) [74,75]. P. aeruginosa is a Gram-negative, aerobic rod belongingto the bacterial family Pseudomonadaceae. High yields (approx. 60–87%) were obtained, with partial reso-lution of racemic mixtures via preferential oxidation of one isomer. An enantiomeric excess was alsoobtained with a-secondary and b-allenic alcohols. Substituents close to the allenic function decrease theponderal yield. Allenic acids (32, 34, 36 and 37) were also obtained via oxidation of a-allenic alcohols byyeast alcohol dehydrogenase [76] (Scheme 2).
MeCOOMe
Dehydromatricaria ester
(ii) + (iii) + ester hydrolysis
10 9
1
1
910HOOC
COOH
(i) + (iv) + rearrangement
HCH2OH9
1
E
Marasin
Scheme 1. Apparent pathway from dehydromatricaria ester to marasin via dicarboxylic acid: (i) reduction of the C1 ester group to analcohol; (ii) saturation of the C2@C3 double bond; (iii) oxidation of the terminal C10 methyl group, which yields a hydroxymethyl group,then a carboxyl group; (iv) decarboxylation of the acid formed in (iii) produces a C9 compound (marasin).
334 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
O
OH
ROOC
E/Z
24 E, R = H25 E, R = Me
H
R
OE/Z
28 R = COOMe, E29 R = COOMe, Z
26 Z, R = H27 Z, R = Me
30 R = AcO, E31 R = AcO, Z
R
R COOH
H
COOH
H32 R = Me33 R = H
34
COOH
MeH
H
35
COOH
H
36
COOH
R
38 R = H39 R = Me
H H
COOH
37
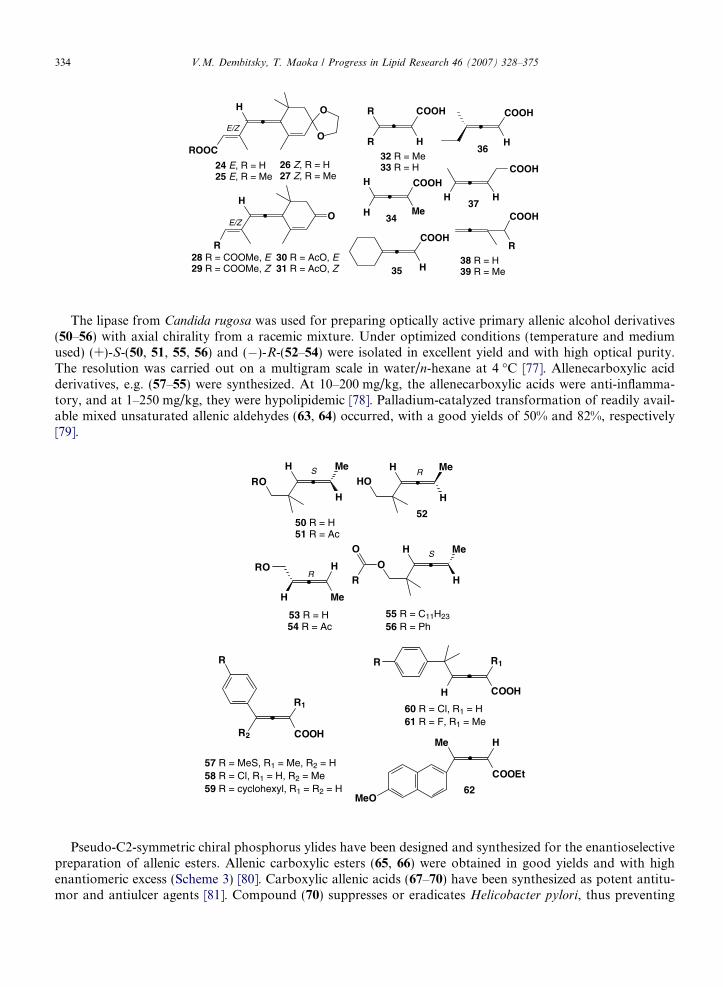
The lipase from Candida rugosa was used for preparing optically active primary allenic alcohol derivatives(50–56) with axial chirality from a racemic mixture. Under optimized conditions (temperature and mediumused) (+)-S-(50, 51, 55, 56) and (�)-R-(52–54) were isolated in excellent yield and with high optical purity.The resolution was carried out on a multigram scale in water/n-hexane at 4 �C [77]. Allenecarboxylic acidderivatives, e.g. (57–55) were synthesized. At 10–200 mg/kg, the allenecarboxylic acids were anti-inflamma-tory, and at 1–250 mg/kg, they were hypolipidemic [78]. Palladium-catalyzed transformation of readily avail-able mixed unsaturated allenic aldehydes (63, 64) occurred, with a good yields of 50% and 82%, respectively[79].
H Me
H
RO
H
H
Me
RO
S
R
H
H
HOR
50 R = H51 R = Ac
53 R = H54 R = Ac
52
SH Me
Me
H
O
O
R
55 R = C11H23
56 R = Ph
H
COOEt
Me
MeO
R1
COOHH
R
R2
R1
COOH
R
57 R = MeS, R1 = Me, R2 = H 58 R = Cl, R1 = H, R2 = Me 59 R = cyclohexyl, R1 = R2 = H
60 R = Cl, R1 = H61 R = F, R1 = Me
62
Pseudo-C2-symmetric chiral phosphorus ylides have been designed and synthesized for the enantioselectivepreparation of allenic esters. Allenic carboxylic esters (65, 66) were obtained in good yields and with highenantiomeric excess (Scheme 3) [80]. Carboxylic allenic acids (67–70) have been synthesized as potent antitu-mor and antiulcer agents [81]. Compound (70) suppresses or eradicates Helicobacter pylori, thus preventing

H
CHO
HHOa
O O
O
Hb
63
Ph
CHO
PhHOa
O O
O
Phb
64
Scheme 2. Synthesis of unsaturated allenic aldehydes: (a) BuLi, tetrahydrofuran (THF)–hexane �20 �C, then COCl2, toluene, �20 �C,then silyl enol ether, MeLi, tetramethylethylenediamine, THF, �40 �C; (b) Pd(PPh3)4, THF, 60 �C.
PhROOC
65 R = Et, 81% ee66 R = t-Bu, 92% ee
P
Ph
Ph
Ph
O
O
R
+ Br- 1. NaHMDS, 25
oC, THF
2.
OPh -78
oC
Scheme 3. Synthesis allenic carboxylic acid esters (NaHMDS, sodium bis(trimethylsilyl)amide).
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 335
recurrence of peptic ulcer, and also (70) dose-dependently inhibited water immersion stress-induced ulcerformation in rats. Synthesis of other allenic compounds were reviewed in some books and other publications[6–8,10–14].
Me
Ph OEt
O
Me COOEt
67
Me
Ph
R
OEt
O
68 R = Me69 R = Ph
70
2.2. Triacylglycerides of plant species
Laballenic acid was present in seeds of both species, mostly as a free acid. The epoxy acids of P. oreophillaconsisted of 9,10-epoxystearic, vernolic, coronaric, and 15,16-epoxyoctadec-9,12-dienoic acids [64]. In the tria-cylglycerols of seeds of three species of the family Labiatae (Phlomis regelii, P. oreophilla, and Lavandula
spica), the saturated acids were distributed either in the sn-1 or in the sn-1 and sn-3 positions. The distributionof C18:1 was characteristic for each oil; C18:2 was esterified primarily in the sn-2 position, with some in sn-1 ofthe two terminals; C18:3 in two of the species preferentially occupied sn-3, and sn-1 occupied the other. In P.
regelii, laballenic acid esterified the 3 position of the triacylglycerols, with preference shown for the two ter-minals. Phosphorylation of sn-1,2- and sn-2,3-diacylglycerols was not accompanied by the formation of appre-ciable amounts of secondary products [82].

336 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
The advantages and disadvantages of HPLC investigations of triglycerides of seed oil from plants of theLamiaceae family: Ballota nigra, Dracocephalum thymiflorum, Leonurus cardiaca, Mentha piperita, Ocimum
basilicum, Origanum majorana, Phlomis tuberose, Salvia aethiopis, S. tesquicola, Stachys palustris, and S. recta
were reported [83]. Special attention was paid to triglycerides formed from laballenic acid. Four main types oftriacylglycerides (71–75) with laballenic acid have been isolated from seed oil of the Lamiaceae family. Theseed oil of Sebastiana commersoniana (Euphorbiaceae) was rich in triacylglycerides. Linolenic acid was themain fatty acid [82]. A unique triacylglyceride (74) was noted, where one a-carbon of the glyceride backbonewas esterified with 8-hydroxy-5,6-octadienoic acid, in the second position with trans-2-cis-4-decadienoic acid(also known as stillingic acid), and in the third position with a-linolenic acid [84].
The optically active lipid isolated from the seed oil of the Chinese tallow tree, Sapium sebiferum, previ-ously considered to be a triglyceride containing 2,4-decadienoic acid esterified with one of the primaryhydroxyls of glycerol, was shown to be tetraester triglyceride. The other primary hydroxyl of glycerol is ester-ified to 8-hydroxyoctanoic acid, and its x-hydroxyl group, which in turn, is esterified to decanoic acid. Intriacylglycerol the predominant common fatty acids are linoleic and linolenic acids. The 8-C hydroxy acidwas 8-hydroxy-5,6-octadienoic acid (see structure 75). The optical activity observed in compounds with 8-hydroxy-5,6-octadienoic acid probably resulted from the allene function rather than an asymmetric b-C ofglycerol [85]. The highly OAT of the seed oil of Sebastiana ligustrina were isolated and characterized [86].As with the similar, known triglycerides from Sapium sebiferum, the optically active 8-hydroxy-5,6-octadie-noic acid was esterified to glycerol and to 2,4-decadienoic acid. The other two acyl moieties in the triglycer-ides of both these species are common fatty acids, principally 16:0, 18:1, 18:2, and 18:3, which occur in pairsranging in degree of unsaturation from 16:0, 18:1 to 18:3, 18:3. Analysis of lipids of parts of the related spe-cies Stillingia sylvatica and S. texana revealed no allene or carbonyl conjugation, and thus by implication, noOAT. A substance having IR absorptions characteristic of allenes and carbonyl, conjugated dienes, but dif-ferent from OAT, was found in the stems of S. texana. The seeds of S. sebiferum are known to contain OAT,but it was not found in fresh leaves [86].
O
O
HO
O O
O
OH
H
7
ZE
4
75
5
5O
O
H
H(CH2)8Me
O
O O
(CH2)6Me
(CH2)4Me
O
5O
O
H
H(CH2)8Me
O
O O
(CH2)6Me
(CH2)4Me
O
129
9
5
O
O
H
H(CH2)8Me
O
O O
(CH2)4Me
(CH2)3Me
O
O
O
H
H(CH2)8Me
O
O O
(CH2)6Me
(CH2)3Me
O
71
72
73
74

α-ketol
RR
O OH
Z
13
76 9,11,12,14-Octadecataetraenoic acid
77 12,13(S)-oxido-9Z,11,15Z-octadecatrienoic acid
Linolenic acid
7-iso-Jasmonic acid
Reductaseβ-oxidation
O
COOH
O
(CH2)7COOHα- and γ-ketolsand racemic OPDA
Spontaneous hydrolysis
Allene oxide cyclase
Allene oxide synthase
+ H2O2
+ H+
HO2C5
5
HO2C
OOH
Z Z
EZ
HO2C
H
5
H
13(S)-Hydroperoxylinolenic acid
5
HO2C
O
Lipoxygenase
13-Hydroxy-9,11-octadecatrienoic acid
Z EHO2C
OH
5
- H2O
HO2CO+
HOOH
+
5 78
Fig. 2. The proposed mechanism involves the formation of reactive intermediate cations and allenic acid when used in oxidation oflinolenic acid. The lipoxygenase-catalyzed formation of the linolenic acid hydroperoxide includes the enzymatic conversion of the alleneoxide (12,13S)-epoxyoctadecatrienoic acid to enantiomeric 12-oxo-(10,15Z)-phytodienoic acid (OPDA) and later to jasmonic acid as wellas its chemical decomposition to a- and c-ketol and racemic OPDA.
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 337
The composition of glycerides was detected for seed oils of Anethum graveolens, Carum carvi, Corian-
drum sativum, Cuminum cyminum, Foeniculum vulgare, Pertroselinum sativum, Piminella anisum, Asclepias
syriaca, A. incarnata, Vincetoxicum nigrum, and Leucas cephalotes. Of these, the rarely occurring fattyacids are petroselinic, vaccenic, and laballenic acids [87,88]. Seed oil from Leonotis nepetaefolia contains16% of laballenic acid [59]; the oil of Leucas urticaefolia seeds contains 24% of 5,6-octadecadienoic(laballenic acid) [89], and Leucas cephalotes was found to be a new seed oil rich in labellenic acid up

338 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
to 28 wt.% [90]. The kernel oil of Chinese tallow (Sapium sebiferum) seed contains tetraester triglyceridescomposed of trans-2,cis-4-decadienoic acid joined in an estolide linkage to 8-hydroxy-5,6-octadienoic acid[91].
Earlier investigations showed that a particular isoform of LOX localized on lipid bodies plays a role ininitiating the mobilization of triacylglycerols during seed germination [92]. Physiological functions of LOXswithin whole cotyledons of cucumber (Cucumis sativus) were analyzed by measuring the endogenousamounts of LOX-derived products. As the dominant metabolite of the LOX pathway in this tissue,lipid-body LOX-derived esterified (13S)-hydroperoxy linoleic acid accumulated to about 14 lM/g freshweight, which represented about 6% of the total amount of linoleic acid in cotyledons. This LOX productwas not only reduced to its hydroxy derivative, leading to degradation by b-oxidation, but alternatively, itwas metabolized by fatty acid hydroperoxide lyase, leading to the formation of hexanal as well. Further-more, the activities of the LOX form metabolizing linolenic acid, were detected by measuring the accumu-lation of volatile aldehydes, ketones, and carboxylic acids [93]. Allene oxides are unstable epoxides that havebeen implicated as intermediates in the biotransformation of hydroperoxy-ecosatetraenoic acids and relatedhydroperoxides to ketols and cyclopentenones. However, direct proof of the structure of the putative alleneoxide intermediates has been hampered by their extreme instability under the conditions of their biosynthe-sis (Fig. 2).
The isolation and structural elucidation of allene oxides prepared from the (13S)-hydroperoxides of lin-oleic and linolenic acids have been described [92]. 12,13(S)-Oxido-9Z,11-octa-decadienoic acid was derivedfrom linoleic acid, and 12,13(S)-oxido-9Z,11,15Z-octadecatrienoic acid (77) was derived from linolenic acid(Fig. 2). Analysis of the breakdown products formed upon exposure to H2O led to identification of thehydrolysis and cyclization products previously characterized as enzymic derivatives of the (13S)-hydroper-oxides in flaxseed. 12,13(S)-Oxido-9Z,11-octadecadienoic and 12,13(S)-oxido-9Z,11,15Z-octadecatrienoic(77) acid were formed via reactive intermediate cations (78) and allenic fatty acid (76). An intermediate cat-ion (78) is unstable and produced allenic acid (76). This allenic fatty acid, with internal hydroperoxide, pro-duced allene oxide (77). Allene oxide (77) is also a reactive intermediate with an unusual structure,characterized by an epoxide ring with an exocyclic carbon–carbon double bond. Allene oxide cyclaseand/or spontaneous hydrolysis acts to form well-known jasmonic acid, a- and c-ketols, and racemic 12-oxo-(10,15Z)-phytodienoic acids, respectively [94–96]. Moreover, allene oxide cyclase catalyzes the stereo-
3
OH
O
O
O
3
COOMe
OH
O
COOHMe
Me
O
3
Me
MeCOOH
O
O
3
COOMe
O
OH OH
3
COOMe
Me
Me
O
OCOOMe
Me
Me COOMe
Me
Me
O
Me
MeCOOH
79
80
81
82
83
84
85
86
87
Fig. 3. Oxidation of allenic acids underwent via allene oxides and formed a-hydroxyketones and lactones.

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 339
specific cyclization of an unstable allene oxide to (9S,13S)-12-oxo-(10,15Z)-phytodienoic cacid, the ultimateprecursor of jasmonic acid [93].
Allenic carboxylic acids (79, 80) have been converted to functionalized lactones and a-ketols by oxidation–cyclization promoted by dimethyldioxirane. These transformations can be rationalized by the involvement ofallene oxide (81, 82) and spirodioxide intermediates (83, 84). When products were formed directly from alleneoxides, keto lactones are formed. The corresponding spirodioxide intermediates produced lactones (87) withappropriately situated a-hydroxyketone moieties (85, 86) (Fig. 3). Use of prepared solutions of the oxidantgenerally proceed via spirodioxides, whereas in situ reactions normally yielded products derived from theallene oxides [97]. Oxidation and epoxidation of allenic acids and other allenes have recently been reviewed[98].
3. Allenic prostaglandins
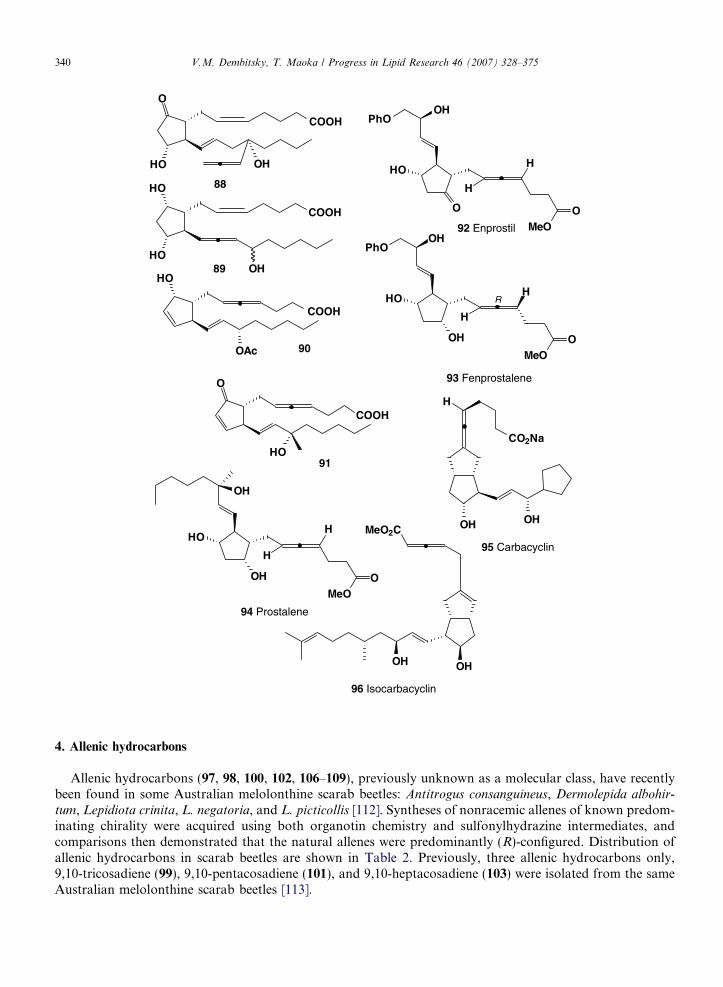
The term prostaglandin derives from the prostate gland. Prostaglandins were first discovered and iso-lated from human semen in 1935 by the Swedish physiologist Ulf Von Euler [99]. Prostaglandins playactive roles in stimulating human muscle contraction and in regulating the cardiovascular and nervous sys-tems. Structurally, they are unsaturated carboxylic acids, consisting of a 20-carbon skeleton that also con-tains a five-member ring; they are biochemically synthesized from arachidonic acid [100,101]. Allenicprostaglandins have not been isolated from natural sources, but some bioactive analogs of natural pros-taglandins have been synthesized and used in pharmaceutical science and clinics. Bronchodilator (88), aPGF2 analog, was prepared [102]; this prostaglandin (89) had 0.05–0.2 times the activity of PGF2a inan antifertility assay in hamsters [103]. Allenic prostaglandins (90) and (91) have physiological activitiessimilar to natural prostaglandins, e.g., as ovarian cycle regulators, broncho dilators, antihypertensives,and sedatives [104].
Prostaglandin-E analogues inhibit gastric acid secretion [105,106]. Enprostil (92), a synthetic analogue ofprostaglandin E2, is effective in treating patients with duodenal or gastric ulcers. Four allenic isomers, whichare prostanoids, are structurally related to PGE2. Enprostil also exhibited a potent agonist effect at FP and TPreceptors in the rat colon and guinea pig aorta (�logEC50 values = 7.34 and 6.54) [107]. The unnatural allenicR- and S-isomers of (92) were much less potent than (92), with the latter being virtually inactive. The (92) andthe natural R- and S-isomers, therefore, were EP3, FP, and TP agonists, being most potent at the EP3 recep-tor. The preferred configurations for these receptors appear to be the R form, and to a lesser extent, the S formof the natural allenic isomer [99]. In a MTT assay, concentrations of enprostil (>10 lM) had cytotoxitic effectson HT-29 cells [108]. Furthermore, enprostil (1 lM) suppressed IL-8 production in HT-29 cells, SW620 andCaCo2 were stimulated with interleukin-1-beta (IL-1-b) or LPS, but did not suppress this response when cellswere stimulated with TNF-a. The obtained results suggest that enprostil affects a point in the pathwaybetween the IL-1 receptor or the LPS receptor and nuclear NF-kappa B, without affecting the pathwaybetween the TNF receptor and NF-kappa B, with the latter factor being required for IL-8 gene transcription[108].
Enprostil and the somatostatin analogue SMS 201 995 affected the growth of a clonal variant of thehuman gastric adenocarcinoma cell line (MKN45) [109]. For example, Enprostil reduced postprandialserum gastrin levels when administered from day 7 to day 14 and prevented gastrin release by MKN45in vitro. It also inhibited growth of colonic cancer cells PROb and REGb [110]. Moreover, Enprostilhas been shown to exert both antisecretory and mucoprotective effects [111]. Another prostaglandin, fen-prostalene (93, also known as Bovilene), could induce abortion during mid-pregnancy in bitches, and thesubsequent estrus may occur early with a low conception rate [104]. It has a known inhibitory effect onprostaglandin synthesis in newborn pigs [105,106]. Prostalene (94) solution may be used under the FederalFood, Drug, and Cosmetic Act for controlling estrus in mares [107,108]. Prostalene (at 1 mg) produced anincreased pregnancy rate after both foal heat and second postpartum estrus breedings in the mare [109].Other pharmacological and toxicological properties of prostalene were reviewed [110]. The prostacyclinanalogs, carbacyclin (95) and isocarbacyclin (96), were found to be potent antiplatelet agents in a ratthrombocytopenia model [111].
340 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
COOH
O
HO OH
88
COOH
HO
HOOH89
COOH
HO
OAc 90
COOH
O
HO91
R
93 Fenprostalene
OH
HO
OHPhO
H
H
MeOO
92 Enprostil
O
HO
OHPhO
H
H
MeOO
96 Isocarbacyclin
95 Carbacyclin
OHOH
MeO2C
H
CO2Na
OH OH
94 Prostalene
OH
HOH
H
MeOO
OH
4. Allenic hydrocarbons
Allenic hydrocarbons (97, 98, 100, 102, 106–109), previously unknown as a molecular class, have recentlybeen found in some Australian melolonthine scarab beetles: Antitrogus consanguineus, Dermolepida albohir-
tum, Lepidiota crinita, L. negatoria, and L. picticollis [112]. Syntheses of nonracemic allenes of known predom-inating chirality were acquired using both organotin chemistry and sulfonylhydrazine intermediates, andcomparisons then demonstrated that the natural allenes were predominantly (R)-configured. Distribution ofallenic hydrocarbons in scarab beetles are shown in Table 2. Previously, three allenic hydrocarbons only,9,10-tricosadiene (99), 9,10-pentacosadiene (101), and 9,10-heptacosadiene (103) were isolated from the sameAustralian melolonthine scarab beetles [113].

Table 2Distribution of D9,10-alkadienes in Canebeetle species [112]
Scarab species D9,10-Alkadienes
A. consanguineus 102, 103,a 104, 107
L. negatoria 103a
L. crinita 108, 109a
L. picticollis 101a
D. albohirtum 101a
a Major allene in each species.
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 341
Examination of the chemistry of a number of Australian insect species provided examples of unusual struc-tures and encouraged researchers to determine their absolute stereochemistry by stereocontrolled synthesesand chromatographic comparisons. Inter alia, studies with the fruit-spotting bug (Amblypelta nitida), certainparasitic wasps (Biosteres sp.), the aposematic shield bug (Cantao parentum), and various species of scarabgrubs were summarized. Determination of enantiomeric excesses (ee’s) for component allenes, epoxides, lac-tones, and spiroacetals was also reported [114].
n
106 n = 6107 n = 8108 n =11109 n =13
n 99 n = 1 100 n = 2101 n = 3102 n = 4
H
H
97
98
10
103 n = 5104 n = 7105 n = 9
5. Allenic steroids and derivatives
It has been established that the Hawaiian sponge Callyspongia diffusa contains 24-keto-cholesterol and anew allenic sterol, 24-ethyl-D5,24(28),28-cholestatrien-3b-ol (110) [115]. Isolated compounds specificallyinhibited the conversion of stigmast-5-en-3b-ol to stigmasta-5,24(28)-dien-3b-ol and/or to 24,28-epoxy-stig-mast-5-en-3b-ol in Bombyx mori larvae [116]. The (110) held the larvae in the second instar for >20 days whenadministered alone or with the dietary sterols. Synthetic cholesta-5,23,24-trien-3b-ol (111) inhibited insectgrowth and development, but did not act via inhibition of sterol dealkylation [117]. A series of allenicderivatives of cholesterol (112–115) was synthesized from pregnenolone [118]. These compounds carry anallenic function at C20 or C22 and were devised with the aim of inhibiting C22 hydroxylation of ecdysonebiosynthesis by a suicide-substrate mechanism. The four compounds synthesized could efficiently inhibitthe synthesis of ecdysone in the prothoracic glands of Locusta. It also was shown that all allenic steroidsinduced a decrease in the synthesis of ecdysone [119]. Three steroidal compounds (116–118) with allenic sidechains substituted for phosphonyl-linked groups were synthesized and tested for their effects on Leishmania
donovani and L. mexicana mexicana culture growth [120]. The compounds inhibited organism proliferationat concentrations in 5–10 lg/mL. Interestingly, the same two di-Et phosphonosteroid compounds (117 and118) that inhibited Leishmania proliferation were also the most active against P. carinii, as detected by thepotent effect they had in reducing cellular ATP content. The most potent inhibitors of this group of com-pounds were characterized by two ethyl groups at the phosphate function. Leishmania organisms treated with(117) exhibited reduced growth after transfer into inhibitor-free medium. Because there are currently no axenicmethods available for the continuous subcultivation of Pneumocystis carinii, the effects of these drugs on thisorganism were evaluated by two alternative screening methods.

342 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
111
HO
H
HH
H
110
HO
H
HH
H
115 R =
114 R =
113 R =
112
OH
H
OH
OH
H
OH
OH
HO
R
HO
The effects of conjugated allenic 3-oxo-5,10-secosteroids, (4R)-5,10-secoestra-4,5-diene-3,10,17-trione (119)and of (4R)-5,10-seco-19-norpregna-4,5-diene-3,10,20-trione (120) on epididymal D4-5a-reductase and 3a-hydroxy steroid dehydrogenase were reported [121]. Assessment by IC50 values showed that D4-5a-reductasewas inhibited approx. 50- and 250-fold more effectively than 3a-hydroxy steroid dehydrogenase by (119) and(120), respectively. It also shown that compound (120) inhibited androgen biosynthesis in the mature rat (0.8–8 mg/kg/day for 7 days, i.p.) [122].
118
H
H H
O
PH
OEtO
OEt
H
116 R = H117 R = Et
H
H H
MeO
PH
ORO
OR
C
O
O
H H
HAc
C
O
O
H H
HO
119
120
123 C22 = S124 C22 = R
22
E
Z
OH
HOH
H
CH
H
OH
E
Z
121 C22 = S122 C22 = R
22
OH
HOH
H C
OH
OH
Dihydroxyvitamin D3
OH
HOH
H
OH

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 343
The steroid hormone 1a,25-dihydroxyvitamin D3 [(3b,5Z,7E)-9,10-secocholesta-5,7,10(19)-trienetriol], inwhich allenic functionality has been incorporated into the side chain (121–124), has been synthesized [123].The hormonal form of vitamin D, 1a,25-dihydroxyvitamin D3, generates many biological actions by interac-tions with its VDR. The presence of a carbon-25 hydroxyl group is necessary for optimizing binding to theVDR. To examine the effect of spatial orientation of the 25-hydroxyl, researchers recently observed two pairsof 22,23-allene side chain analogs [124]. The 22R orientation in analogs HR (52%) and LA (154%) resulted inhigher affinity binding than the 22S orientation of analogs (121 and 123) HQ (21%) and LB (3.5%;1,25D = 100%).
Limited trypsin proteolysis revealed that 22R analogs (122 and 124) induced VDR conformationalchanges better suited to protect VDR from digestion than 22S analogs. 22R analogs were also able toinduce gene transcription at 10–100-fold lower concentrations than 1,25D; however, 22S analogs were lesseffective. Analog LA was at least 10-fold more potent than 1,25D at inducing differentiation, whereas theother analogs were less potent. None of the analogs were as potent as 1,25D in promoting in vivo intes-tinal calcium absorption or bone calcium mobilization. LA was the most potent of the analogs butrequired 20–30-fold higher doses than 1,25D. The 25-hydroxyl orientation, combined with the 16,17-enefunctionality of analog LA, enhances its ability to interact with VDR and induce biological activity. Allen-ic sulfone (125), an analog of dehydroepiandrosterone sulfatide, a potent, natural inhibitor of glucose-6-phosphate dehydrogenase, was obtained from propargylic sulfone (126) by tautomerizing the electrophilicallene (125) [125].
126
H
O
HH
O2S
C13H27
H
O
HH
O2S
H
125
C13H27
6. Allenic terpenoids
Vernonallenolide (127), 4b-hydroxy-4,5-dihydro-vernonallenolide (128), and an epoxide of vernonalleno-lide (129) have been isolated from seven Vernonia species, mainly from northern Brazil [126]. The aerial partsof Vernonia cotoneaster from India afforded vernonallenolide (127), 4b-hydroxy-4,5-dihydrovernonallenolide(128) [127]. Three Vernonia species, namely, V. compactiflora, V. cotoneaster, and V. chalybaea, also containvernonallenolide [128,129]. Acalycixeniolides C (130), D (131), E (132), and four new diterpenoids of the xeni-cane class have been isolated from the gorgonian Acalycigorgia inermis. Isolated compounds exhibited cyto-toxicity against a human leukemia cell line [130]. Novel compounds, acalycixeniolide C, D, and E, isolatedfrom Acalycigorgia inermis, inhibit the growth of leukemia, and were thus used for the treatment of cancer[131]. Acalycixenolide E (132), from Acalycigorgia inermis, exhibits potent anti-angiogenic activity bothin vitro and in vivo. Compound (132) inhibits the bFGF-induced proliferation of HUVEC in a dose-dependentmanner, along with the basic fibroblast growth factor-induced migration, invasion, and tube formation ofHUVECs. Moreover, (132) potently inhibits the in vivo neovascularization of the chorioallantoic membranesof growing chick embryos [132].

344 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
A bioactive metabolite, ginamallene (133), was isolated from four species of Acalycigorgia and was shownto be a xenicin-type norditerpene having terminal allene functionality [133]. Acalycixeniolide B 0 (134), C 0
(135), and the previously reported ginamallene have been isolated from a gorgonian of the genus Acalycigorgia
as inhibitors of cell division of fertilized sea urchin eggs [134,135]. Twelve fungal strains from marine habi-tates, predominantly from the North Sea, were studied in a chemical screening program with respect to theirsecondary metabolite production. The strain WDMH46 (Paraspheaoshaeria sp.) produced several secondarymetabolites with an allene moiety, named lupallenes (136–142), which were isolated, and their structures elu-cidated by Diana Wolff (University of Gottingen, Germany) [136].
129
C
O
OAc
O
OAc
O
128
C
O
OAc
O
OAc
Me OH
127 Vernonallenolide
C
O
OAc
H
O
OAc
132 Acalycixeniolide E
O
H
HO
AcO
131 Acalycixeniolide D
O
O
H
HO
130 Acalycixeniolide C
O
O
H
HO
H
O
OAc
AcO
H
H
133 Ginamallene
O
R
O
134 Acalycixeniolide B', R = OAc 135 Acalycixeniolide C', R = H
Table 3Distribution of major lupallens in the strain WDMH46 (Paraspheaoshaeria sp., Ref. [136])
Metabolite Occurrence, % of total
136 17.9137 18.9138 17.6139 9.3140 29.3

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 345
Lupallene A (138), C (137), and E (139) have shown antimicrobial activity against Candida albicans,
Stapphylococcus aureus, Bacillus subtilis, and Escherichia coli. Distribution of the major allenic metabolitesin the strain WDMH46 (Paraspheaoshaeria sp.) are shown in Table 3. Metabolites not indicated in Table3 are considered as minor compounds. A new unusual diterpene, irciniketene (143, 7-cis-14,15-didehyd-roretinal), was isolated from the marine sponge Ircinia selaginea collected from the sea around Baihai(Guangxi Province, China) [137]. In this structure, a ketene function is connected with a long conjugatedsystem via an allene unit. Irciniketene showed a moderate inhibition toward MGC and HepG2 tumor celllines.OH
OHO
OH136 Lupallene A C-7 S137 Lupallene B C-7 R
7
OH
RO
OH
OH
HO
138 Lupallene C, R = Ac139 Lupallene E, R = H
OH
OAcO
OH
140 Lupallene D
OH
HOO
OH
141 Lupallene F1, R142 Lupallene F2, S
143 Irciniketene
C
C
C
O
MeZ
7. Marine bromoallenes
Although the debrominated sesquiterpene panacene was isolated from the plants Panax ginseng and P.
quinquefolius [22–27], more recently, brominated analog of panacene (144) was isolated from the marine gas-tropod mollusc Aplysia brasiliana [28]. Simple bromoallene (145) was isolated from the red alga Laurencia

346 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
obtusa [138]. Two halogenated C15 acetogenins, itomanallenes A (146) and B (147), with a terminal bromoal-lene moiety, have been isolated from the red alga Laurencia intricata collected in Okinawan waters. The alco-hol corresponding to itomanallene B seems to be a plausible precursor of itomanallene A, which has anunusual 2,10-dioxabicyclo[7.3.0]dodecene skeleton [139]. Laurallene (148) and isolaullene (149), both bro-moallenes, were isolated from Laurencia nipponica [140,141]. Kumausallene (150) is a natural product isolatedfrom the red algae Laurencia nipponica. It contains a 2,6-dioxabicyclo[3.3.0]octane ring system in its core[142,143]. Kumausallene and 1-epi-kumausallene (151), obtained from red algae of the genus Laurencia, weresynthesized [144]. Kumausallene and pannosallene (152) produced major metabolites by the Japanese marinered alga Laurencia nipponica Yamada (Rhodomelaceae, Ceramiales) [145].
Okamurallene (153), a halogenated C15 metabolite, was isolated from some red algal species belongingto the genus Laurencia [146]. The red alga Laurencia okamurai yielded four nonterpenoid C-15 bromoal-lenes (154–156) as minor components, together with laurinterol and debromolaurinterol as major compo-nents [147]. The structures of okamurallene and three metabolites (157–159) were isolated from the redalga Laurencia intricata [148]. The 12-membered O-bridged cyclic ether obtusallene derivatives (160–164)were present in extract of the red seaweed Laurencia obtusa from Kas in the Turkish Mediterranean[149], 10-bromoobtusallene (134) was detected from the same alga [150]. New polyhalogenated (165–169), 12-membered, O-bridged cyclic C15-ethers, having in common oxygenation at C(6) and a bromoal-lene side chain at C(4) [where C(1) is the bromoallene-chain terminus], were isolated from the red seaweedLaurencia obtusa from the Turkish Mediterranean Sea [151]. Two new natural brominated allenes, termednipponallene 1 (170) and neonipponallene 2 (172), were isolated from the red alga Laurencia nipponica,collected near the Russian shore of the Sea of Japan (Troitsa Bay) [152]. Laurallene (171), brominatedcyclic ether extracted from Laurencia nipponica [140,153], was also found in other algal species of thegenus Laurencia.
R
147 Itomanallene B
146 Itomanallene A
Br
H H
O
AcO
O
O
Br
H
Br
145 Bromoallene
H
HBr
H
144 Panacene
O
OH
BrH
H
H
H
S
S
148 Laurallene
O
O
H
H
H
H
Br
Br
149 Isolaurallene
O
O
Br H
H
H
H
Br
152 Pannosallene
S
O
O
H
H
Br
HH
H H
Br
R/S
150 Kumausallene, R151 1-epi-Kumausallene, S
O
OH
H
H
H
Br
Br
H

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 347
A bromoallene metabolite, termed aplysiallene (173), was isolated from the Japanese sea hare, Aplysia
kurodai, (Gastropoda), as an Na,K-ATPase inhibitor. The known metabolites, laurinterol and debromo-laurinterol, isolated from this sponge, were also evaluated for their Na, K-ATPase inhibitory activity[154]. A C-15 nonterpenoid (174) was detected in the red alga Laurencia okamurai [155]. From a singlecollection of the red alga Laurencia implicata, three new unusual secondary metabolites (175–177) wereisolated [156]. Other findings include (a) two C-15 bromoallenes (178, 179); (b) a 12-membered O-bridgedcyclic ether obtained from extract of Laurencia obtusa from Kas in the Turkish Mediterranean, and (c)kasallene (180), a highly oxygenated bromoallene, which has been isolated from the red alga L. obtusa
[157].
O
OH
HH
Br
H
Br
OS
155 Isookamurallene
154 Deoxyokamurallene
ZSO
OH
HH
Br
H
Br
156
O
O
H
Br
BrO
153 Okamurallene
O
OH
HH
Br
H
Br
OH
HS S
159O
O
Br
H
HO
Br
H
H
H
H
Br
H
O
O
Br
Br
157
164 Obtusallene IV
162 Obtusallene III, R = H163 Peracetylobtusallene III, R = Ac
OCl
H
H
O
Br
H
Br
H
R
ORO
H
H
O
OROR
H
Br
H
160 Obtusallene I, R = H, R1 = Cl161 10-Bromoobtusallene, R,R1 = Br
RO
R1
Cl
H
H
Br
H
O
R
O
OH
HH
Br
H
Br
H
Cl
HO
S
158

Br H
S
168 Obtusallene VIII, R = H169 Obtusallene IX, R = Ac
O
H
H
O
OR
Br
H
Br
H
Br
S
H
Br
O
H
R
O
Cl
O
167 Obtusallene VII
O
H
H
OBr
H
Br
HCl
BrOH
H
165 Obtusallene V, R = Br166 Obtusallene VI, R = H
S
172 Neonipponallene 2
O
Br
H
OAc OAc
H
Br
H
H
170 Nipponallene 1, R171 Laurallene, S
R
O
O
Br
HH
H
H
H
Br
HR/S
RO
O H
H
Br
HH
H
H
R
173 Aplysiallene, R = Me174 R = Br
180 Kasallene
O
OH
OHO
OH
Br179
OO
H
H
Cl
Br
Br
178
177
O
OH
HBr
BrAcO
Br
176
175
O
OH
HBr
Br
H
OBr
OO
H
H Br
O
O
H
H
HO
HO OH
Br
8. Allenic norisoprenoids
Natural norisoprenoids (181–189), structurally derived from carotenoids, are widely distributed inplants, algae, and insects, and because of their often low flavor thresholds, many of them are highly
348 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 349
potent aroma compounds and, thus, of great interest for the flavor and fragrance industries [158–160].Norisoprenoids as degradation carotenoid products [160], represent preferable cleavage of 6,7,7,8,8,9,and 9,10 bonds lead to flavor components with 9,10,11, and 13 carbon atoms, respectively. In theintact plant, carotenoid degradation is likely to be affected by oxygenase systems and by photo-oxy-genations and other nonenzymatic oxidations [161]. The sensory-active C9-, C10-, C11-, and C13-nori-soprenoids naturally emerge from the oxidative cleavage of tetra-terpenoid precursor carotenoids[159,162].
Nonselective fissions of the polyene chain induced by abiotic factors such as heat or light, result in abroad spectrum of oxidized breakdown products [163,164]. A smaller range of products, which often dis-play biological activities, was released by enzymatic cleavage reactions. In the last few years, enzymescatalyzing the centrical or excentrical fission of specific carotenoids in vivo have been isolated and char-acterized from plants [165]. Recently, chemical, photochemical, and oxidase-coupled degradation ofcarotenoids to norisoprenoids have been reported [158,166]. The brownish secretion emitted upon distur-bances by the grasshopper, Romalea microptera, produced four allenic norisoprenoids (181–184) [167],which are produced by degradation of related allenic carotenoids [158]. An allenic ketodiol (185) withthe natural stereochemistry observed in samples isolated from the defensive secretions of a large flightlessgrasshopper was synthesized from b-ionone by a sequence of reactions involving photosensitized oxygen-ation and photo-stereomutation [168]. Biosynthesis of fucoxanthin and neoxanthin may involve the samereactions [158,169].
Twenty-four norisoprenoids, including grasshopper ketone, which are either free volatile components ofjuices of Vitis vinifera cv. Chardonnay, Semillon, and Sauvignon Blanc, or were liberated by glycosidaseenzyme, or acid hydrolysis of extracts of these juices, have been identified [170]. The hypothetical 7-oxomeg-astigmane precursors, grasshopper ketone and megastigm-5-en-7-yne-3,9-diol, as well as the related allene, 9-hydroxymegastigma-4,6,7-trien-3-one (183), have been observed for the first time, co-occurring with damasce-none, 3-hydroxy-b-damascone 3-oxo-b-damascone, and 3-oxo-a-damascone. Hydrolytic studies have shownthat megastigm-5-en-7-yne-3,9-diol is a precursor of damascenone and 3-hydroxy-b-damascone during wineconservation.
Allenic norisoprenoid glycoside, icariside B1 (186) was observed in some plant species: Epimedium grandi-florum var. thunbergianum [171], Epimedium diphyllum [172], Vitis vinifera cv. Riesling leaves [173], Nepeta cad-
mea [174], in leaves of Glochidion obovatum [175], Ebenus cretica [176], in the fruit of anise (Pimpinella anisum)[177], in the aerial parts of three species of Erigeron: E. annuus, E. philadelphicus, and E. sumatrensis [178], inthe leaves of Cryptostegia grandiflora [179], Peucedanum japonicum [180], Taraxacum obovatum [181], in theaerial parts of Crepidiastrum lanceolatum [182], Evodia austrosinensis [183], and in leaves of Laurus nobilis
[184].Two glycosides citroside A (187) and B (188), were isolated from the methanol extract of leaves of Citrus
unshiu [185]. On the basis of spectral and chemical evidence, the structures of the new glycosides were detectedas (5-dehydroxygrasshopper ketone-5-yl) 5-b-D-glucopyranoside (187) and a (5-dehydroxy-allenic ketodiol-5-yl) 5-b-D-glucopyranoside (188), from the methanol extracts of coriander (fruit of Coriandrum sativum) [186],and celery seed (fruit of Apium graveolens) [187]. Citroside A was also isolated from the barks of Prunus ssiori
and P. padus [188], the leaves of Hydrangea macrophylla var. thunbergii [189], lulo leaves (Solanum quitoense)[190], the aerial parts of Phlomis grandiflora var. grandiflora [191], a leaf extract of Eriobotrya japonica [192],the aerial parts of Phlomis spinidens [193], the leaves of Melaleuca quinquenervia [194], the leaves of Lasianthus
wallichii [195], the aerial portion of Erythroxylum cambodianum [196], and from the aerial parts of Tribulus
parvispinus [197]. Citroside B was also isolated from the leaves of Solanum quitoense [198]. A potent antiulcer-ogenic compound, cassioside: (3R)-4-[(2 0R,4 0S)-2 0-hydroxy-4 0-(b-D-apiofuranosyl-(1! 6)-b-D-glucopyrano-syl)-2 0,6 0,6 0-trimethylcyclo-hexylidene]-3-buten-2-one (189), was isolated from a hot water extract ofCinnamomi Cortex (the dried stem bark of Cinnamomun cassia) [199], and from the juices of grapes and otherfruits [200]. The allenic epoxycyclohexanes (190, 191) were isolated from the culture medium of Eutypa lata
[201,202].

350 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
OH
OH
O
H
190
OH
OH
O
H
191
O
HO
HO
OH
HO
O OH
H
O
HO O
H
O
O
OH
OH
OH
OH186 Icariside B1
187 Citroside A, R188 Citroside B, S
R/S
O
OHHO
HO
OO
OH
OH
OH
HO OH
H
O
189 Cinnamoside185
HO OH
H
O
HO R
H
O
HO
HOH
OH
181
182 R = Me183 R = OH
AcO
HOH
O
184
9. Allenic carotenoids
9.1. Distribution in nature
Carotenoids, which exhibit purple, red, orange, and yellow color, are tetraterpene pigments distributed inbacteria, fungi, algae, as well as in higher plants and animals. To date, more than 700 carotenoids have beenfound in nature [203]. Of approximately 700 naturally occurring carotenoids, about 43 carotenoids contain theallene group. The principal allenic carotenoids are fucoxanthin (192) in brown algae and diatoms, and perid-inin (193) in dinoflagellates and neoxanthin (194) in higher plants and algae [204–206]. They serve as light-har-vesting pigments in photosynthesis and occur as carotenoid–chlorophyll–protein complexes in chloroplasts[204]. Some marine animals, such as sea anemones, shellfishes, sea urchins, and tunicates also contain alleniccarotenoids [22,24].
9.1.1. Marine and freshwater algal species
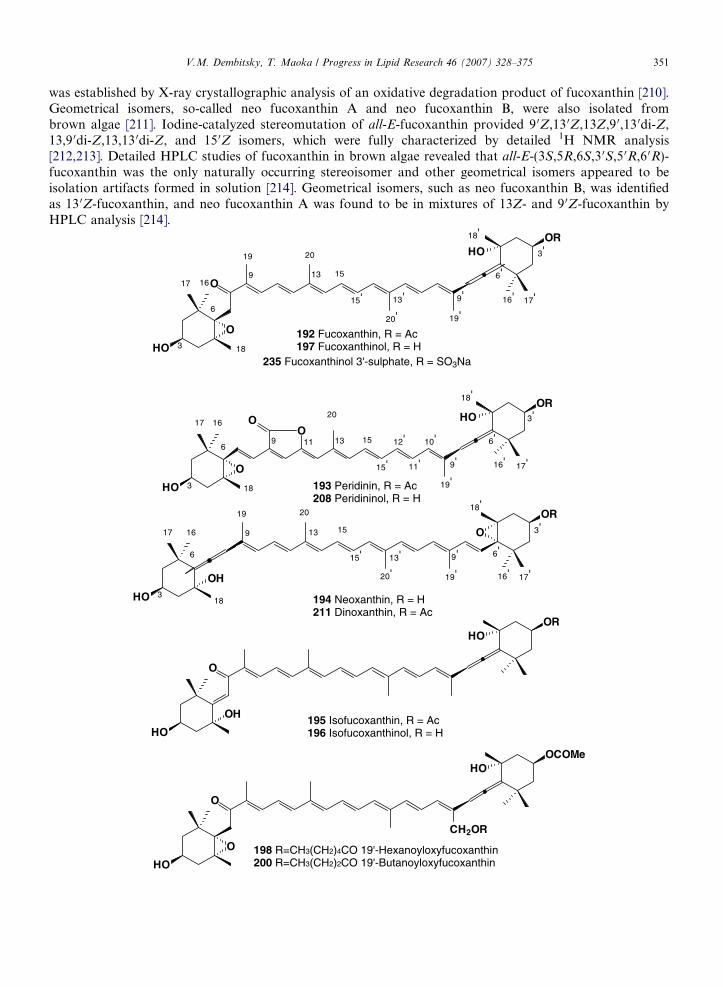
Fucoxanthin (192) is a one of the most abundant marine carotenoids, which is widely distributed in brownalgae and diatoms [204–206]. Fucoxanthin contributes more than 10% of the estimated total production ofcarotenoids in nature [206]. Fucoxanthin was first isolated by Willstatter and Page in 1914 [17]. About 50 yearslater, its structure was found to be 5,6-epoxy-3 0-ethanoyloxy-3,5 0-dihydroxy-6 0,7 0-didehydro-5,6,7,8,5 0,6 0-hexahydro-b,b-caroten-8-one by Bonnett et al. [207–209]. Subsequently, (3S,5R,6S,3 0S,5 0R,6 0R)-chirality
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 351
was established by X-ray crystallographic analysis of an oxidative degradation product of fucoxanthin [210].Geometrical isomers, so-called neo fucoxanthin A and neo fucoxanthin B, were also isolated frombrown algae [211]. Iodine-catalyzed stereomutation of all-E-fucoxanthin provided 9 0Z,13 0Z,13Z,9 0,13 0di-Z,13,9 0di-Z,13,13 0di-Z, and 15 0Z isomers, which were fully characterized by detailed 1H NMR analysis[212,213]. Detailed HPLC studies of fucoxanthin in brown algae revealed that all-E-(3S,5R,6S,3 0S,5 0R,6 0R)-fucoxanthin was the only naturally occurring stereoisomer and other geometrical isomers appeared to beisolation artifacts formed in solution [214]. Geometrical isomers, such as neo fucoxanthin B, was identifiedas 13 0Z-fucoxanthin, and neo fucoxanthin A was found to be in mixtures of 13Z- and 9 0Z-fucoxanthin byHPLC analysis [214].
15'
15
3
6
18
1617
19
9 13
20
13'
20' 19'
9' 6'
18'
3'
16' 17'
OCOMeHO
HO
O
O
CH2OR
235 Fucoxanthinol 3'-sulphate, R = SO3Na
17'16'
18'
19'
20
18
1617
17'16'
18'
19'20'
2019
18
17 16
13119 10'
11'
12'
15'
156
3
9'
6'
3'
3'
15'
15
6
3
139
13' 9'
6'
HO
OH
OR
O
198 R=CH3(CH2)4CO 19'-Hexanoyloxyfucoxanthin200 R=CH3(CH2)2CO 19'-Butanoyloxyfucoxanthin
HO
O
HO
OH
OR
195 Isofucoxanthin, R = Ac196 Isofucoxanthinol, R = H
ORHO
HO
O
OO
193 Peridinin, R = Ac208 Peridininol, R = H
ORHO
HO
O
O
194 Neoxanthin, R = H211 Dinoxanthin, R = Ac
192 Fucoxanthin, R = Ac197 Fucoxanthinol, R = H

352 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
Allenic (6 0S)-isomers of fucoxanthin were obtained from natural (6 0R)-fucoxanthin by iodine-catalyzed1
stereomutation in benzene under strong light and were characterized by H NMR [215]. However,(6 0S)-fucoxanthin has not been found in nature [214]. Treatment of fucoxanthin with base produces yellowhemiketals and isofucoxanthin (195) as well as isofucoxanthinol (196) [216] and acid, which leads to theformation of blue oxonium ions [217]. Fucoxanthin is distributed in marine algae such as Chrysophyceae,Prymnesiophyceae, Bacillariophyceae and Phaeophyceae as a major carotenoid [204–206,218]. Total synthe-sis of all-E-(3S,5R,6S,3 0S,5 0R,6 0R)-fucoxanthin was achieved by Yamano et al. [219]. Fucoxanthiol (197), adeacetyl product of fucoxanthin, was distributed in some marine algae as a minor carotenoid along withfucoxanthin [220–223]. Prymnesiophyceae (Haptophyceae) are characterized by 19 0-Acyloxyfucoxanthinderivatives. 19 0-Hexanoyloxyfucoxanthin (198) was isolated from Emiliania (Coccolithus) huxleyi [224]and Chrysochromulina polylepis [225,226] as a major carotenoid. 4-Keto-19 0-hexanoyloxyfucoxanthin(199), which was first misidentified as 19 0-hexanoyloxyparacentrone 3-acetate [224], was also isolated asa minor component from E. huxleyi, and its structure was fully characterized by detailed spectral analysis[227]. Furthermore, 19 0-butanoyloxyfucoxanthin (200) was found in Pelagococcus subviridis (Chrysophy-ceae) [228].
A series of apo-fucoxanthin, apo-10 0-fucoxanthinal (201), apo-12 0-fucoxanthinal (202), apo-12-fucoxanth-inal (203), and apo-13 0-fucoxanthinone (204) were isolated from the marine diatom Phaeodactylum tricornu-
tum [229]. Apo-9 0-fucoxanthinone (205) was isolated from the cultured marine dinoflagellate Amphidinium sp.[230]. Apo-13-fucoxanthinone (206) was isolated along with 9 0-apo-fucoxanthinone, and apo-13 0-fucoxanthi-none was isolated from brown alga Scytosiphon lomentaria [231]. These apo-fucoxanthins were assumed to beoxidative cleavage products of fucoxanthin in algae. Vaucheriaxanthin (207) was first isolated from Vaucheria
virescens, belonging to Raphidophyceae (Xanthophyceae) and its structure was found to be 19 0-hydroxyneo-xanthin after saponification [232]. Vaucheriaxanthin is characteristic for Eustigmatophyceae but also occurs inXanthophyceae [205,233] and is presented as a free and esterified form in Nannochloropsis salina (Eustigmat-ophyceae) [233].
HO
O
OO
202 Apo-12'-fucoxanthinal
HO
O
OO
201 Apo-10'-fucoxanthinal
HO
O
CH2OCO(CH2)4CH3
O
OCOMeHO
O
199 4-Keto-19'-hexanoyloxyfucoxanthin

OCOMe
206 Apo-13-fucoxanthinone
HO
O
OO
205 Apo-9'-fucoxanthinone
O
OCOMeHO
204 Apo-13'-fucoxanthinone
203 Apo-12-fucoxanthinal
O
HO
O
OCOMeHO
Peridinin (193), which is a unique C37-skeltal carotenoid characteristic of dinoflagellates, was first iso-lated in 1980 by Sch}utt [234]. Its structure was found to be 5,6-epoxy-3 0-ethanoyloxy-3,5 0-dihydroxy-6 0,7 0-didehydro-5,6,5 0,6 0-tetrahydro-12 0,13 0,20 0-trinor-b,b-caroten-19,11-olide by Strain et al. [235]. The(3S,5R,6S,3 0S,5 0R,6 0R) chirality for peridinin was determined by ozonolytic degradation of its p-bro-mobenzoate to derivatives of known chirality obtained from fucoxanthin and violaxanthin [236]. Totalsynthesis of all-E-(3S,5R,6S,3 0S,5 0R,6 0R)-peridinin was achieved by Yamano et al. [237] and Furuichet al. [238]. The (6 0S)-allenic isomer of peridinin was prepared from (6 0R) natural peridinin by iodine cat-alyzed photoisomerization and its stereochemistry was confirmed by chemical synthesis [239,240]. How-ever, (6 0S)-peridinin has not been found in nature. Furthermore, iodine-catalyzed stereomutation of all-
E-(3S,5R,6S,3 0S,5 0R,6 0R)-peridinin provided 9 0Z, 11Z and 13Z isomers, which were fully characterizedby detailed 1H NMR analysis [241]. Incorporation experiment of 14C, 3H-labeled mevalonate in intact cells[242] and 14C labeled zeaxanthin in a cell-free system [243] of the alga Amphidinum carterae (Dinophceae)revealed that C37-skeltal peridinin was formed from neoxanthin by deletion of the three carbon atoms atC-13 0, C-14 0, and C-20 0. Peridinin derivatives were isolated in dinoflagellate. Peridininol (208), a deacetylproduct of peridinin, was isolated from dinoflagellate, along with peridinin [244]. Two C8 epimeric furano-ide derivatives of peridinin (209 and 210) were isolated from the cultured dinoflagellate of the genus Sym-biodinium, a symbiont of the Okinawan soft coral Clavularia virdis [245]. Dinoxanthin(neoxanthin 3-acetate) (211) was also found in several dinoflagellate algae, along with peridinin [244,246]. Two C8 0 epi-meric furanoxide carotenoids, dinochrome A (212) and dinochrome B (213), were isolated from the freshwater red tide Peridinium bipes [247]. Gyroxanthin (214), the first allenic acetylenic carotenoid, was iso-lated from dinoflagellate Gymnodinium galatheanum [248]. A carotenoid termed P457 (215) is a minorcarotenoid disaccharide that is encountered in several dinoflagellates [244,249]. Its structure was foundto be (3S,5R,6R,3 0S,5 0R,6 0S)-13 0Z-7 0,8 0-dihydroneoxanthin-20 0-al 3 0-b-D-lactoside. This carotenoid repre-sents one of the most complex carotenoid structures known, containing structural elements such as a lac-toside, cross-conjugated C40-carotenal, allene, 5,6-epoxide and saturated C-7,8 bond [250,251]. Neoxanthin(194) is widely distributed in algae [204–206].
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 353

354 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
215 P457
.
OH
OH
O
O
O
HOOH
OH
O OH
OHHO
HO
O
O
8'
8
214 Gyroxanthin
OCOMeHO
CH2OH
HO
O
OH
MeCOO
OH
OH
212 (8'R) Dinochrome A 213 (8'S) Dinochrome B
209 (8R) Peridinin furanoxide 210 (8S) Peridinin furanoxide
O
OCOMe
O HO
HOO H
207 Vaucheriaxanthin HO
CH2OH
OH
O
OH
9.1.2. Higher plants
Neoxanthin (194) is a common carotenoid, widely distributed in higher plants and algae, first isolated fromthe green leaves of barley by Strain in 1938 [252] . Its structure was found to be 5 0,6 0-epoxy-6,7-didehydro-5,6,5 0,6 0-tetradehydro-b,b-carotene-3,5,3 0-triol, and 9Z geometry was assigned to a major neoxanthinobtained from maple leaves by Cholnoky et al. in 1969 [253]. all-E-Neoxanthin was isolated from the petalsof flowers and characterized by 1H NMR [254]. The structures of all-E-neoxanthin [255] and 9 0Z-neoxanthin[256] were fully confirmed by chemical synthesis. Takaich and Mimuro reported that only the 9 0Z form wasfound in chloroplasts. On the other hand, the all-E form was found in nonphotosynthetic organs such as fruit,petal, and root [257]. Iodine-catalyzed stereomutation of all-E-neoxanthin provided 9 0Z,9 0Z,13Z,13 0Z,15Z,and three minor di-Z-isomers, which were characterized by 1H NMR, UV, and HPLC [258]. Neoxanthinwas converted from violaxanthin by neoxanthin synthase (NSY) in plants [259,260]. A gene of neoxanthin

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 355
synthase was isolated from tomato [259] and potato [260]. Neoxanthin is converted to two stereoisomers ofneochromes 216 and 217, by acid catalyzed epoxy-furanoxide rearrangement, and these structures were char-acterized [261]. Two C6 epimeric pairs of pentahydroxy allenic carotenoids, neoflor (218) and 6-epineoflor(219), were isolated from the petals of Trollius europaerus [254]. They were assumed to be hydrolytic cleavageproducts of epoxy end groups in neoxanthin [254,261]. A symmetrical di allenic carotenoid, mimulaxanthin(220), was isolated from the petals of the monkey flower Mimulus guttatus [262], and yellow flowers of Lami-
num montanum [263] and its structure was found to be (3S,5R,6R,3 0S,5 0R,6 0R)-6,7,6 0,7 0-tetrahydro-5,6,5 0,6 0-tetrhydro-b,b-carotene-3,5,3 0,5 0-tetrol by chemical and spectroscopic data. Its structure was also confirmedby chemical synthesis [264]. Deepoxyneoxanthin (221) was also found in Mimulus guttatus [263]. Allenicseco-carotenoids termed tobiraxanthin C (222) [265] and 5-hydroxy-tobiraxanthin C (223) [266] were isolatedfrom the seeds of Pittosporum tobira. They were assumed to be oxidative cleavage products of epoxy endgroups in neoxanthin. Furthermore, a novel carotenoid tocopherol complex, termed pittosporumxanthinC1 (224) and C2 (225), were isolated from the seeds of P. tobira. These structures were found to be 11 0,12 0-additional products of 9 0Z-neoxanthin with a-tocopherol [267]. Capsoneoxanthin (226) was isolated fromthe fruits of Asparagus falcatus along with capsanthin, capsorubin, capsocrome, and capsanthin 5,6-epoxide[268]. This compound was assumed to be the pinacol rearrangement product of neoxanthin catalyzed by cap-santhin-capsorbin synthase.
216 (8'R)-Neochrome217 (8'S)-Neochrome
OH
HO
OH
OH
218 Neoflor
8'
OHHO
HO
OH
OH OHHO
HO OH
OH
219 6-Epineoflor
OHHO
220 Mimulaxanthin
HO
OH
221 Deepoxyneoxanthin
OH
HO
OH
9.1.3. Invertebrates and animalsAnimals generally do not synthesize carotenoids de novo and those found in animals represent either a
direct accumulation of carotenoids from the food or are partly modified through metabolic reactions[22,24,269]. Several marine invertebrates, such as sea anemones, shell fishes, sea urchins, and tunicates are

356 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
filter feeders and accumulate carotenoids from dietal micro algae. Peridinin was found in sea anemones[270,271], gorgonians [272], shellfishes [22], and tunicates [273]. Fucoxanthin and fucoxanthinol were alsofound in shellfishes [22,274,275], sea urchins [276–278], and tunicates [279]. They represent free and esterifiedforms in shellfishes [274,275]. 3-Acylated fucoxanthinols were found in the clam Mactra chinensis [275]. 19 0-Hexanoyl-oxyfucoxanthin was isolated from the mussel Mytilus edulis [280,281].
Several metabolites of fucoxanthin and peridinin in marine animals have been isolated. A C31 allenic apoc-arotenoid named paracentrone (227), which is an oxidative cleavage product of fucoxanthin, was isolated fromthe sea urchin Paracentrotus lividis, along with fuxoxanthin and fucoxanthinol [282]. Hydratoperidinin (228), ahydrolytic cleavage product of epoxy end group in peridinin, was isolated from the corbicula clam Corbicula
japonica [283,284]. A C37-skeltal apocarotenoid (229), found in the oyster Crassostrea gigas [285], was assumedto be a dehydroxy and oxidative metabolite of (228). An enol carotenoid (230), which was assumed to be a met-abolic intermediate of fucoxanthin to mytiloxanthin, was isolated from C. gigas [285]. Two allenic carotenoids,named corbiculaxanthin (231) and 7 0,8 0-didehydro-deepoxyneoxanthin (232), were isolated from the clam C.
japonica [283,284]. The 7 0,8 0-didehydro-deepoxyneoxanthin (232) has an interesting structure with both alleneand acetylene bonds, and it was assumed to be one of the possible precursors of the di-acetylenic carotenoid,alloxanthin in shellfish [284]. Amarouciaxanthin A (233), a metabolite of fucoxanthin, was first isolated fromtunicate Amaroucium pliciferum [279,286]. It was also found in bivalve Paphia euglypta as an esterifiedform [287]. 19-Ethanoyloxy derivatives of amarouciaxanthin A, termed muricellaxanthin (234), were isolatedfrom a dark red gorgonian Muricella sp. [288].
R2R1
HHHH
223 5-Hydroxytobiraxanthin C
OHHO
O
OH O
C O
(CH2)12
CH3
CH3
)12OH
HO
O
O O
C O
(CH2
226 Capsoneoxanthin
224 Pittosporumxanthin C1225 Pittosporumxanthin C2
222 Tobiraxanthin C
OHO
O
O
R1
R2
HO
OH
OH
O
HO
OH

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 357
It was reported that allenic carotenoids were converted to corresponding acetylenic carotenoids in mar-ine invertebrate [22,269]. Feeding experiments of M. edulis with the micro algae, Skeletonema costatum
(diatom), Emiliania huxleyi (haptophyte), and Heterocapsa triquetra (dinoflagellate) revealed that fucoxan-thin and peridinin were metabolized to halocynthiaxanthin and pyrrhoxanthinol, respectively [281]. Somefreshwater fishs take-up fucoxanthin from their dietary algae or aquatic insects [289]. However, fish cannotaccumulate fucoxanthin and its derivatives in their bodies [289]. Fucoxanthinol 3 0-sulfate (235) and para-centron were found in egg yolk after feeding a diet supplemented with the seaweed Fucus serratus, themajor carotenoid of which is fucoxanthin [290]. Fucoxanthinol and amarouciaxanthin A were detectedin plasma and liver after oral administration of fucoxanthin in mouse and human hepatoma cell HepG2incubated with fucoxanthin-containing medium [291]. The conversion of fucoxanthinol into amarouciaxan-thin was predominantly shown in liver microsomes of mouse. This conversion required NAD(P)+ as acofactor [291].
232 7', 8'-Didehydrodeepoxyneoxanthin
OH
HO
OH
OOH
OCOMeHO
OO
OH
OHHO
228 Hydratoperidinin
HO
OH
OH
OHHO
HO
OH231 Corbiculaxanthin
230
227 ParacentroneHO
OH
O
OCOMeHO
OO
O
OH229
233 Amarouciaxanthin A, R = H234 Muricellaxanthin, R = OAc
OO
OHR
HO
OH

358 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
9.2. Functions and biological activities
The principal function of fucoxanthin, peridinin, and neoxanthin in plants is light-harvesting, which is theprimary process in photosyntheses. These pigments occurred as carotenoid–chlorophyll–protein complexes inthe thylakoid membrane of chloroplasts [204]. The fucoxanthin–chlorophyll–protein complex was isolatedfrom brown algae and diatoms and characterized [292,293]. The crystal structure of peridinin–chlorophyll–protein in dinoflagellate Amphidinum carterae was determined by X-ray crystallography [294]. Neoxanthinis also a known component of light-harvesting chlorophyll-protein in higher plants. The crystal structure ofmajor light-harvesting protein in spinach was determined by X-ray crystallography [295,296]. Another impor-tant role of carotenoids in plants is quenching of singlet oxygen. Peridinin [297], fucoxanthin, and neoxanthin[298,299] exhibited quenching activity for singlet oxygen. However, these activities are weaker than those of b-carotene [298,299].
The important roles of carotenoids in marine animals are in antioxidation, immune enhancement, repro-duction, growth, maturation and photoprotection [300]. Fucoxanthin also possesses excellent radical scaveng-ing activity [301,302]. Furthermore, fucoxanthin enhances immunological activity and ovulation in sea urchin[303]. The best known biological function of carotenoids in humans is with pro-vitamin A. However, all alleniccarotenoids cannot be converted to vitamin A. Carotenoids also have been known to reduce the risk of cancerand several life style diseases [304]. Some carotenoids exhibit anti-tumor and anti-carcinogenic activities [304].Recently, fucoxanthin has received increased attention as an anticancer carotenoid. Fucoxanthin inhibitsgrowth of human neuroblastoma GOTO cells [305,306], human leukemia cells [307], prostate cancer cells[308], and activation of Epstein-Barr virus early antigen in Raji cells [309]. Fucoxanthin also inhibits duode-num and skin carcinogenesis in mouse [310,311]. Fucoxanthin induces apoptosis and enhances the antiprolif-erative effect of the cancer cells [312,313]. Anti-mutagenic effects of fucoxanthin and peridinin have also beenreported [314]. Recent studies found that fucoxanthin has antiobesity [315] and anti-inflammatory effects [316].
Neoxanthin was reported to inhibit lipid peroxidation [317], carcinogensis [318] and the generation ofsuperoxides from stimulated leukocytes [319]. Peridinin exhibits antitumor prompting activity in a two-stagecarcinogenesis in the skin of mice, and its potency was shown to be higher than that of fucoxanthin [314,320].Other allenic carotenoids, namely, deepoxyneoxanthin, fucoxanthinol, and isofucoxanthin, also inhibit activa-tion of Epstein-Barr virus early antigen in Raji cells [309]. Peridinin furanoxides [245] and dinochromes [247]exhibit significant growth-inhibitory activity toward cancer cells. In the present paper, the IUPAC-IUB carot-enoid nomenclature was based on the new rules described in the ‘‘Carotenoid Handbook’’ [203].
10. Aromatic and peptide metabolites
A few aromatic and peptide metabolites have been isolated from natural sources. A new lignoid (235) with anovel allene structure was isolated from the bark of the Brazilian medicinal plant Brosimum acutifolium (Mor-aceae), whose common name is ‘‘murure’’. This plant, which has been used in folk medicine in the Amazonianregion, is an anti-inflammatory and antirheumatic agent [321]. Two derivatives of 2-propenoic acid, eucalyp-tene A (236) and B (237) are fungal metabolites of Clitocybe eucalyptorum [322]. Improved production of an
Table 4Inhibitory activities of allenic aromatic ethers against tongue cancer KB and KBv200 cells in vitro (IC50, lM; Ref. [325])
No. KB KBv200
239 22.52 20.86240 27.64 18.96241 17.36 18.27242 29.36 23.38243 40.58 49.56244 71.50 42.52245 75.40 52.92246 48.76 55.47247 34.55 30.27

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 359
allenic ether, eucalyptene A (236), from a mangrove fungus Xylaria sp. 2508, was also reported [323]. The fun-gus Hypoxylon chestersii culture solutions yielded a new allenic aromatic ether, chesterseine, 4-(4-methoxy-phenoxy)buta-1,2-diene (238) [324].
Allenic aromatic ethers (239–247), some of which are natural products isolated from the mangrove fungusXylaria sp. 2508, were synthesized [325]. Their antitumor activities against KB and KBv200 cells were deter-mined (see Table 4). All these compounds exhibited cytotoxic potential, ranging from weak to strong activity.Analysis of the structure–activity relationships revealed that the introduction of allenic moiety could generateor enhance cytotoxicity of these phenol compounds.
OHCO
OH
OMe
HO
MeO
OH
235
O
COOMe
O OMe
236 Eucalyptene A, E237 Eucalyptene B, Z
E/Z
238 Chestersiene
O 2
R1
O
O 1
R2
239 R1 = H, R2 = COOMe240 R1 = H, R2 = CH=CHCOOMe241 R1 = OMe, R2 = CH=CHCOOMe242 R1 = OH, R2 = CH=CHCOOMe243 R1 = H, R2 = COOH244 R1 = H, CH=CHCOOH
245 R1 = H, R2 = COOMe246 R1 = H, R2 = CH=CHCOOMe247 R1 = OMe, R2 = CH=CHCOOMe
R
R
A novel cyclic peptide containing an allenic ether of aa N-(p-hydroxycinnamoyl)amide [xyloallenolide A(248)] and two aromatic allenic ethers, the but-2,3-dienyl ether of p-hydroxy-cinnamic acid (249) and the cor-responding Me ester eucalyptene A (250) were isolated from the endophytic fungus Xylaria sp. from the SouthChina Sea [326].
249 R = H250 R = Me
248 Xyloallenolide A
ORO
O
N
N N
NHO
O
O
OH
O

360 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
11. Allenic amino acids and derivatives
L-2-Aminohexa-4,5-dienoic acid (249) was isolated from the toxic mushroom Amanita solitaria (=Amanita
echinocephala, Basidiomycetes, Amanitaceae, also known as European Solitary Lepidella) [327]. This mush-room group often smells like chlorine household bleach (sodium hypochlorite). Amanita solitaria also containsin excess of 1000 ppm 2(S)-amino-4,5-hexadienoic acid, 300 ppm of toxic trans-2-amino-5-chloro-4-hexenoicacid, and a chloride ion concentration (2000 ppm) significantly greater than that found in other basidiomyce-tes [328]. The synthetic c-allenic-GABA (250) and related compounds (251, 252) are potent and specific mech-anism-based inhibitors of mammalian GABA-transaminase [329]. Synthetic allenic amino alcohols (253–257)are neoplasm inhibitors, which inhibited the growth of certain transformed cancer cells in rat tissue culture[330].
The a-allenic a-amino acids are potent irreversible inhibitors of their target amino acid decarboxylases[331]. a-Allenic-DOPA (258) rapidly inactivates porcine kidney AADC. The diastereomeric pairs of chiralallenic aroma amino acids (259) differ in their abilities to inactivate mammalian and bacterial AADC.Although there is little variation in the abilities of (258) or the separate diastereomeric pairs of (259) to inac-tivate bacterial tyrosine decarboxylase, one diastereomeric pair was at least an order of magnitude more effec-tive than the other against mammalian AADC. Bacterial aromatic amino acid decarboxylase underwentinactivation by 3-hydroxy-a-1,2-propadienyl-tyrosine (258) [332].
COOH
NH2
R
R1
249
COOH
H2N
250 R = R1 = H251 R = Me, R1 = H252 R = H, R1 = Me
HO R
NH2
253 R = H254 R = Me
HO
NH2
255
OHNH2
R
256 R = H257 R = Me
258 R = H, R1 = OH259 R = Me, R1 = H
R
COOHH2N
HO R1
12. Allenic alkaloids
Several taxa of small frogs contained alkaloids (260–265) similar or identical to compounds previouslyknown only from neotropical poison frogs of the family Dendrobatidae [333–335]. Dihydroisohistrionico-toxin (H2-HTX, 260), discovered from frog Dendrobates histrionicus skin [333,336], inhibits acetylcholinereceptor-dependent 22Na+ uptake of cultured chick muscle cells with a Ki of 0.2 lM. H2-HTX increasesthe affinity of the desensitized form of the receptor for agonists but not antagonists. The obtained resultssuggest that H2-HTX inhibits the acetylcholine receptor by causing an increase in the affinity of the desen-sitized form of the receptor for agonists and thereby stabilizes the desensitized state [337]. Some of these
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 361
compounds may have arisen convergently from new biosynthetic pathways in several families of frogs, orthese alkaloids may represent parallel expression of shared-primitive pathways that are unexpressed or lostin related frogs. Distribution and biological activities of frog alkaloids have been reviewed in some recentpapers [338–340]. Pipecolic acid, a lysine metabolite, is thought to be a factor responsible for hepaticencephalopathy [341], and it is a potent inhibitor of some liver cancer cells such as Walker 256 carcino-sarcoma and Morris hepatoma 5123 [342]. Three bioactive derivatives of pipecolic acid (266–268) weresynthesized by Agami et al. [343].
HNHO
H260 Dihydroisohistrionicotoxin
264 R = H265 R = Me263
+NMe2O
O
R
NO
O
262
NH
H
HNHO
261 Isotetrahydrohistrionicotoxin
NO
O
Ph
H
HN
O OMe
NO
OMe
MeO O
266267
268
13. Cumulenic lipids and related compounds
Cumulenes are usually highly unstable compounds, and they do not seem to be widely distributed in nature.Several cumulenic fatty acids and other metabolites have been discovered in nature. Thus, the 2,6,7,8-decatet-raen-4-ynoic acid, 9-(methylthio)-, methyl ester (269) was obtained from the extract of roots of Anthemius aus-
triaca [35]. From the roots of the same plant, two d-lactones of 5-hydroxy-9-(methylthio)-2,4,6,7,8-decapentaenoic acid (270) and its isomer (271) were also isolated [344]. The above-ground portions of Erigeron
canadense (fleabane, Compositae) contained unstable cumulenes (272, 273). These metabolites were biogenet-ically closely related to the lachnophyllum ester cis-Pr(C„C)2CH@CHCO2Me [345]. The 5-(1,2,4-hexatrieny-lidene)-2(5H)-furanone, (274) was isolated from Aster bellidiastrum [346]. Cumulenic fatty acid (275), and twocumulene phenolics (277, 278) were isolated from maruna bezwonna (Matricaria inodora = Tripleurospermum

362 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
perforatum, Compositae) [347]. Seven years later 2,3,4-decatrienoic acid, methyl ester (276) was found in oil ofMatricaria inodora [348].
269
SMe
Me
H
MeOOC
271
SMe
Me
H
OO
270
SMe
Me
H
OO
OO
272
OO H
273
OO
274
275 R = H276 R = Me
H COOR
H
CPh
PhC C C C
277 278
Ph
Ph
Families of cumulene compounds (279–290) containing the natural double bonds have been discoveredfrom astronomical sources, where some terminal hydrogen atoms were replaced by deuterium atoms. Thecumulene carbenes are important components of hydrocarbon chemistry in low-mass star-forming cores. Asimple allene (1,2-propadiene-1,3-diylidene) and 1,2,3,4-pentatetraene-1,5-diylidene (279) were detected inthe dark cloud TMC-1 for the first time in the course of a molecular line survey using the Nobeyama 45-mtelescope (Nobeyama Radio Observatory, Nagano, Japan) [349]. More recently, they were also observed bythe IR space observatory in the young planetary nebula NGC 7027 [350].
Several authors from the Jet Propulsion Laboratory (California Institute of Technology, USA) reported thefirst astronomical detection of the long-chain cumulene carbene (279 and 282) in the interstellar cloud TMC-1[351]. In this year other groups from the USA reported the cumulene carbenes (280, 281, 282, and 283) as wellas the cyanopolyynes HC11N and HC13N; the C chain radicals C7H, C8H, C9H, and C11H were observed fromP1 astronomical sources [352]. Cumulene carbenes (284–290) were observed for the evolved carbon star IRC+10216, and discovered by scientists from the Harvard-Smithsonian Center for Astrophysics (Cambridge,MA, USA) [353].
279 ButatrienylideneH2C C C CH2
280 Pentatetraenylidene, R = R1 = D281 Pentatetraenylidene, R,R1 = H
H2C C C C CRR1

290 Decanonaenylidene, R, R1 = D
H2C C C C C C C C C CRR1
288 Nonaoctaenylidene, R, R1 = D289 Nonaoctaenylidene, R,R1 = H
286 Octaheptaenylidene, R, R1 = D287 Octaheptaenylidene, R,R1 = H
H2C C C C C C C C CRR1
H2C C C C C C C CRR1
284 Heptahexaenylidene, R, R1 = D285 Heptahexaenylidene, R,R1 = H
H2C C C C C C CRR1
282 Hexapentaenylidene, R, R1 = D283 Hexapentaenylidene, R,R1 = H
H2C C C C C CRR1
V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 363
14. Addition to distributions, biosynthetic preparation and synthesis
Two acetylenic antibiotics, cepacins A and B, have been isolated from the fermentation broth of Pseudo-monas cepacia SC 11,783 and assigned structures (291) and (292), respectively. Cepacin A has good activityagainst Staphylococci (MIC) 0.2 lg/mL, but weak activity against Streptococci (MIC 50 lg/mL) and themajority of Gram negative organisms (MIC values 6.3 approx. >50 lg/mL). Cepacin B had excellent activityagainst Staphylococci (MIC <0.05 lg/mL) and some Gram negative organisms (MIC values 0.1 approx.>50 lg/mL) [354,355]. Synthesis of cepacin A has recently been reported [356].
HO
OH
O
291 Cepacin A
HO
O
O
OOH
O
292 Cepacin B
Biosynthetic preparation of 2,3-allenols (293–314) using the lipase B from Candida antarctica, lipase fromPseudomonas fluorescens, lipase from Candida cylindracea, lipase from pig pancreas, lipase from Candida
rugosa, and lipase from Pseudomonas cepacia has been reported [357]. The organic solvents were hexane,cyclohexane, vinyl acetate, dichloromethane, and/or acetone. Both allenes (>95% ee) of 2-methyl-3,4-pentadi-en-1-ol were prepared via 2-stage enzymic kinetic resolution, i.e., a sequence of lipase-catalyzed acetylation(Pseudomonas cepacia) and hydrolysis (Candida antarctica) [358]. Unique an allenic sugar derivative 6,7,8-tri-deoxy-1,2:3,4-bis-O-(1-methylethylidene)-a-D-galacto-octa-6,7-dienopyranose (315) was synthesized fromacetylenic precursors [359].
The first synthesis of (�)-panacene (144) has been accomplished in concise, highly stereoselective fashionfrom compound available 2-methoxy-6-methylbenzoic acid (15 steps, 8.3% overall yield) [360]. Two reviewarticles were devoted to syntheses of allenes have recently been published [361,362], and Liu was preparedthe PhD thesis from Brigham Young University on synthesis of allenes [363].

364 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
R
OAc
293 R = Me294 R = Et
R
OH
295 R = Me296 R = Et
PhOR
305 R = H306 R = Ac
R
OH
n
297 n = 1, R = Me298 n = 1, R = Et299 n = 2, R = Me300 n = 2, R = Et301 n = 3, R = Me302 n = 3, R = Et303 n = 4, R = Me304 n = 4, R = Et
PhOR
307 R = H308 R = Ac
Ph
OR
309 R = H310 R = Ac
OR
311 R = H312 R = Ac
HOR
313 R = H314 R = Ac O
OO
O
OH
H
H H
315
15. Concluding remarks
Plant and marine secondary metabolites are unique sources for pharmaceuticals, food additives, flavors,and other industrial materials. Accumulation of such metabolites often occurs in plants subjected to stressesincluding various elicitors or signal molecules. This article consists of the following sections: allenic naturalproducts (linear allenes, carotinoids, and terpenoids, bromoallenes, and other naturally occurring allenesand cumulenes) and pharmacological active allenes: steroids, prostaglandins and carbacyclins, amino acids,nucleoside analogs, and other bioactive allenes. At the present time, more than 200 allenic lipids and relatedcompounds have been isolated from living organisms. Natural and synthetic allenic and cumulenic materials,their analogues and derivatives have been discovered and/or synthesized and evaluated for their biologicalactivity. Inspired by the intriguing biological activities of many allenic natural products, allenic moietiesare now introduced in compounds that have the pharmacologically activity. The many functionalized allenesthus obtained exhibit an impressive array of activities, such as enzyme inhibitor activities, cytotoxic or anti-viral activities. Without doubt, other important new allenic and cumulenic compounds possessing importantbiological activities will be discovered in the near future.
Acknowledgements
Authors are grateful to Professor John L. Harwood (University of Wales, College of Cardiff), and ProfessorJohn Boukouvalas (Laval University, Quebec City, Canada) for critically reading the manuscript, and the con-structive comments, and to Mrs Jannette A. Dembitsky for assistance with the Figures.

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 365
References
[1] Dembitsky VM, Srebnik M. Natural halogenated fatty acids: their analogues and derivatives. Progr Lipid Res 2002;41:315–67.[2] Smith Jr CR. Occurrence of unusual fatty acids in plants. Prog Chem Fats Other Lipids 1970;11:137–77.[3] Bertrand M. From allenes to natural products. Parfums Cosmetiques Aromes 1981;39:29–38.[4] Jones ERH. Natural polyacetylenes and their precursors. Chem Brit 1966;2:6–13.[5] Landor SR. Aliphatic compounds. I. Unsaturated aliphatic compounds. Ann Rep Prog Chem 1970;66B:519–40.[6] Landor SR. Naturally occurring allenes. In: Landor SR, editor. The chemistry of the allenes. London: Academic Press; 1982. p.
679–707.[7] Claesson A. Biologically active allenes. In: Landor SR, editor. The chemistry of the allenes. London: Academic Press; 1982. p.
709–33.[8] Hoffmann-Roeder A, Krause N. Synthesis and properties of allenic natural products and pharmaceuticals. Angew Chem Int Ed
2004;43:1196–216.[9] Krause N, Hoffmann-Roeder A. Allenic natural products and pharmaceuticals. In: Krause N, Hashmi A, Stephen K, editors.
Modern allene chemistry, vol. 2. Weinheim, Germany: Wiley-VCH Verlag GmbH & Co. KGaA; 2004. p. 997–1039.[10] Mavrov MV, Kucherov VF. Chemistry of allenes. Uspekhi Khimii (USSR) 1967;36:553–86.[11] Brandsma L. Acetylenes, allenes and cumulenes. Amsterdam, Netherlands: Elsevier; 2003. pp. 512.[12] Brummond KM, Chen H. Allenes in natural product synthesis. In: Krause N, Hashmi ASK, editors. Modern allene chemistry, vol.
2. Weinheim, Germany: Wiley-VCH Verlag GmbH & Co. KGaA; 2004. p. 1041–89.[13] Schuster HF, Coppola GM. Allenes in organic synthesis. New York: Wiley; 1984.[14] Taylor DR. Chemistry of allenes. Chem Rev 1967;67:317–59.[15] Van’t Hoff JH. Voorstel tot Uitbreiding der tegenwoordig in de scheikunde gebruikte Structuur-Formules in de ruimte; benevens een
daarmee samenhangende opmerkung omtrent het verband tusschen optisch actief Vermogen en Chemische Constitutie vanOrganische Verbindingen, Greven, Utrecht, 1874; La chimie dans l’espace, Bazendijk, Rotterdam, 1875.
[16] Hans EA. Alcohols and resin acids in the leaf resin of Alnus glutinosa. Berich Deut Chem Ges 1908;40:4760–4.[17] Willstatter R, Page HJ. Chlorophyll. XXIV. The pigments of the brown algae. Ann 1914;404:237–71.[18] Kylin H. The carotinoid pigments of algae. Z Physiol Chem 1927;166:39–77.[19] Karrer P, Wehrli H, Helfenstein A. The coloring materials of plants. XIX. Zeaxanthin and xanthophyll. Helv Chim Acta
1930;13:268–73.[20] Karrer P, Helfenstein A, Wehrli H, Pieper B, Morf R. Plant pigments. XXX. The carotenes, the xanthophylls, fucoxanthin and
capsanthin. Helv Chim Acta 1931;14:614–32.[21] Retovsky R. Fucoxanthin, zeaxanthin, and carotene from the mass cultures of Navicula. Coll Czechoslovak Chem Commun
1948;13:631–40.[22] Matsuno T, Hirao S. Marine carotenoids. In: Ackman RG, editor. Marine biogenic lipids, fats, and oils, vol. 1. Boca Raton, FL:
CRC Press; 1989. p. 251–388.[23] Weedon BCL, Moss GP. Structure and nomenclature. In: Britton G, Liaaen-Jensen S, Pfander H, editors. Carotenoids, vol. 1A.
Basel: Birkhauser; 1995. p. 27–70.[24] Matsuno T. Aquatic animal carotenoids. Fish Sci 2001;67:771–83.[25] Kondo H, Tanaka G. Chemical constituents of Korean ginseng. Yakugaku Zasshi (Japan) 1915;401:779.[26] Min P. The experimental investigation of Panax ginseng. V. The comparative study on the chemical constituents and the general
action of Panax ginseng and Panax quinquefolius. Folia Pharmacol Jpn 1931;11:256–60.[27] Lee CV, Lee TY. Constituents of Panax ginseng. In: Symposium on phytochemistry, Proceedings of a meeting held as part of the
golden jubilee congress of the University of Hong Kong, 1964. p. 171.[28] Kinnel R, Duggan AJ, Eisner T, Meinwald J, Miura I. Panacene: an aromatic bromoallene from a sea hare Aplysia brasiliana.
Tetrahedron Lett 1977;18:3913–6.[29] Johnson EA, Burdon KL. Mycomycin – a new antibiotic produced by a moldlike actonomycete active against the Bacilli of human
tuberculosis. J Bacteriol 1947;54:281.[30] Jenkins DE. Mycomycin: a new antibiotic with tuberculostatic properties. J Labor Clin Med 1950;36:841–2.[31] King DS. Tuberculosis. New Eng J Med 1950;243:530–6. and 565–72.[32] Celmer WD, Solomons IA. The structure of the antibiotic mycomycin. J Am Chem Soc 1952;74:1870–1.[33] Celmer WD, Solomons IA. Mycomycin. I. Isolation, crystallization, and chemical characterization. J Am Chem Soc 1952;74:
2245–8.[34] Celmer WD, Solomons IA. Mycomycin. II. The structure of isomycomycin, and alkali-isomerization product of mycomycin. J Am
Chem Soc 1952;74:3838–42.[35] Bohlmann F, Zdero C. Polyacetylenic compounds. 195. Cumulene from Anthemius austriaca. Chem Ber 1971;104:1329–31.[36] Corsano S, Capito L, Bonamico M. Research on the structure of glutinic acid. I. Heterocyclic derivatives of the allenic acids. Ann
Chim (Rome, Italy) 1958;48:140–55.[37] Agosta WC. Absolute configuration of pentadienedioic acid. J Am Chem Soc 1962;84:110–1.[38] Cymerman-Craig J, Moyle M. A model for the natural synthesis of allenes. J Chem Soc 1963:5356–60.[39] Craig JC, Moyle M. An in vitro model for the synthesis of allenes. Proc Chem Soc 1963:56–7.[40] Williams RC. Routes to heterocyclic prostanoids from allene-1,3-dicarboxylic esters. University of Salford, Salford, UK; 1989. 118
pp. Diss Abstr Int B 1991;52:1451.

366 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
[41] Tanida H, Irie T, Hayashi Y. Neighboring group participation by pyridine rings and N-oxides. Synthesis and solvolysis of 5,8-dihydro-5,8-methanoisoquinoline derivatives. J Org Chem 1984;49:2527–33.
[42] Kozikowski AP, Ames A. Total synthesis of the C-nucleoside dl-showdomycin by a Diels–Alder, retrograde Dieckmannstrategy. J Am Chem Soc 1981;103:3923–4.
[43] Suzuki T, Kagaya S, Tomino A, Unno K, Kametani T. Studies on the syntheses of heterocyclic compounds. Part 855. Cycloadditionreaction of bis(trimethylsilyl)allenedicarboxylate with homocyclic dienes. J Chem Soc, Perkin Trans 1 1980;12:2801–4.
[44] Corsano S, Inverardi R. Some reactions of glutinic acid and 1,6-hexadieneoctacarboxylic acid. Ricerca Sci 1959;29:74–7.[45] Nixon NS, Scheinmann F, Suschitzky JL. 45. Heterocyclic synthesis with allene-1,3-dicarboxylic esters and acids: new chromene,
chromone, quinolone, a-pyrone, and coumarin syntheses. Tetrahedron Lett 1983;24:597–600.[46] Schlingmann G, Milne L, Pearce CJ, Borders DB, Greenstein M, Maiese WM, et al. Isolation, characterization and structure of a
new allenic polyine antibiotic produced by fungus LL-07F275. J Antibiot 1995;48:375–9.[47] Omura S, Imai H, Takeshima H, Nakagawa A. Structure of a new antimicrobial unsaturated fatty acid from Streptomyces
kitasatoensis NU-23-1. Chem Pharm Bull 1976;24:3139–43.[48] Kavanagh F, Hervey A, Robbins W J. Antibiotic substances from Basidiomycetes. V. Poria corticola, Poria tenuis, and an
unidentified Basidiomycete. Proc Natl Acad Sci USA 1949;36:1–7.[49] Anchel M, Polatnick J, Kavanagh F. Isolation of a pair of closely related antibiotic substances produced by three species of
Basidiomycetes. Arch Biochem 1950;25:208–20.[50] Bu’Lock JD, Gregory H. Structure and reactions of a polyacetylenic glycoside. J Chem Soc 1960:2280–5.[51] Herrmann H. Cortinellin, an antibiotically active substance from Cortinellus shiitake. Naturwissenschaften. 1962;49:542.[52] Anchel M. Some naturally occurring antibiotic polyacetylenes. Trans New York Acad Sci 1954;16:337–42.[53] Hall EA, Kavanagh F, Asheshov IN. Action of forty-five antibacterial substances on bacterial viruses. Antibiot Chemother
(Washington, DC) 1951;1:369–78.[54] Bu’Lock JD, Jones ERH, Leeming PR. Constituents of the higher fungi. V. The structures of nemotinic acid and nemotin. J Chem
Soc 1955:4270–6.[55] Bu’Lock JD, Jones ERH, Leeming PR. Chemistry of the higher fungi. VII. Odyssic acid and odyssin. J Chem Soc 1957:1097–101.[56] Bew RE, Cambie RC, Jones ERH, Lowe G. Natural acetylenes. XIX. Metabolites from some Poria species. J Chem Soc Org
1966;2:135–8.[57] Gupta AS, Aggarwal JS. Production of 12- and 18-hydroxystearic acids and some useful compounds derived from them. J Indian
Chem Soc 1956;33:804–6.[58] Cambie RC, Hirschberg A, Jones ERH, Lowe G. Chemistry of the higher fungi. XVI. Polyacetylenic metabolites from Aleurodiscus
roseus. J Chem Soc 1963:4120–30.[59] Bagby MO, Smith Jr CR, Wolff IA. Laballenic acid. A new allenic acid from Leonotis nepetaefolia seed oil. J Org Chem
1965;30:4227–9.[60] Mikolajczak KL, Rogers MF, Smith Jr CR, Wolff IA. Octadecatrienoic acid from Lamium purpureum seed oil containing 5,6-allenic
and trans-16-olefinic unsaturation. Biochem J 1967;105:1245–9.[61] Aitzetmuller K, Tsevegsuren N, Vosmann K. A new allenic fatty acid in Phlomis seed oil. Fett/Lipid 1997;99:74–8.[62] Bagby MO, Smith Jr CE, Wolff IA. A naturally occurring allenic acid from Leonotis nepetaefolia seed oil. Chem Ind (London)
1964;45:1861–2.[63] Horler DF. (�)-Methyl n-tetradeca-trans-2,4,5-trienoate, an allenic ester produced by the male dried bean beetle, Acanthoscelides
obtectus (Say). J Chem Soc 1970:859–62.[64] Asilbekova DT, Gusakova SD, Umarov AU. Neutral lipids of Phlomis oreophilla and P. regellii seeds. Khim Prirod Soed (USSR)
1983;5:561–5.[65] Rezanka T, Dembitsky VM. Bromoallenic lipid compounds from lichens of central Asia. Phytochemistry 2001;56:869–74.[66] Dembitsky VM. Lichens: Nature’s pioneers. INFORM, Am Oil Chem Soc 2003;14:30–4.[67] Bendz G. Marasin, an antibiotic polyacetylene isolated from the culture medium of Marasmius ramealis. Arkiv foer Kemi
1959;14:305–21.[68] Bew RE, Chapman JR, Jones ERH, Lowe BE, Lowe G. Natural acetylenes. XVIII. Some allenic polyacetylenes from
basidiomycetes. J Chem Soc 1966;2:129–35.[69] Brash AR. Formation of an allene oxide from (8R)-8-hydroperoxyeicosatetraenoic acid in the coral Plexaura homomalla. J Am
Chem Soc 1989;111:1891–2.[70] Baertschi SW, Brash AR, Harris TM. Formation of a cyclopropyl eicosanoid via an allene oxide in the coral Plexaura homomalla:
implications for the biosynthesis of 5,6-trans-prostaglandin A2. J Am Chem Soc 1989;111:5003–5.[71] Brash AR, Baertschi SW, Ingram CD, Harris TM. On non-cyclooxygenase prostaglandin synthesis in the sea whip coral, Plexaura
homomalla. An 8(R)-lipoxygenase pathway leads to formation of an a-ketol and a racemic prostanoid. J Biol Chem1987;262:15829–39.
[72] Davies DG, Hodge P. Biosynthesis of the allene (�)-marasin in Marasmius ramealis. Org Biomol Chem 2005;3:1690–3.[73] Abrams SR, Milborrow BV. Synthesis and biological activity of allenic analogs of abscisic acid. Phytochemistry 1991;30:3189–95.[74] Bertrand M, Ferre E, Gil G, Le Petit J, Deveze L. Microbial oxidation of a-allenic alcohols. Tetrahedron Lett 1980;21:1711–4.[75] Ferre E, Gil G, Bertrand M, Le Petit J. Microbial transformations: a stereoselective pathway to oxidize allenic alcohols. Appl
Microbiol Biotech 1985;21:258–66.[76] Ferre E, Gil G, Barre M, Bertrand M, Le Petit J. Oxidation of a-allenic alcohols by yeast alcohol dehydrogenase. Enzyme Microb
Technol 1986;8:297–9.

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 367
[77] Cipiciani A, Bellezza F. Primary allenic alcohols of high optical purity via lipase catalyzed resolution. J Mol Catal B: Enzym2002;17:261–6.
[78] Galantay EE, Kathawala FG. Hypolipidemic allene carboxylic acids. 16 pp. US Patent: 4011339 19770308. Application: US 75-605040 19750815. 1977.
[79] Bienayme H. A new synthesis of polyunsaturated allenic carbonyls. Tetrahedron Lett 1994;35:7387–90.[80] Li C-Y, Sun X-L, Jing Q, Tang Y. Enantioselective synthesis of allenic esters via an ylide route. Chem Commun Cambridge
2006;28:2980–2.[81] Sato T. Antiulcer agents containing allenecarboxylic acids as urease inhibitors. Jpn Kokai Tokkyo Koho 1998;9 pp. Japanese Patent:
JP 10182451 A 19980707 Heisei.[82] Khomova TV, Asilbekova DT, Gusakova SD, Umarov AU. Study of the structure of plant triacylglycerols by stereospecific
analysis. Khim Prirod Soed 1983;3:279–83.[83] Deineka VI, Deineka LA, Ermakov AM, Tret’yakov MYu. HPLC investigation of triglycerides from plants of the Lamiaceae
family. Chem Nat Comp 2004;40:413–6.[84] Spitzer V, Tomberg W, Pohlentz G. Structure analysis of an allene-containing estolide tetraester triglyceride in the seed oil of
Sebastiana commersoniana (Euphorbiaceae). Lipids. 1997;32:549–57.[85] Sprecher HW, Maier R, Barber M, Holman RT. Structure of an optically active allene-containing tetraester triglyceride isolated
from the seed oil of Sapium sebiferum. Biochemistry. 1965;4:1856–63.[86] Heimermann WH, Holman RT. Highly optically active triglycerides of Sebastiana ligustrina and related species. Phytochemistry
1972;11:799–802.[87] Seher A, Fiebig HJ. Stereospecific analysis of triacyl glycerides of rarely occurring fatty acids. Fette, Seifen, Anstrichmittel
1983;85:333–8.[88] Plaeek LL. A review on petroselinic acid and its derivatives. J Am Oil Chem Soc 1963;40:319–29.[89] Nasirullah AF, Osman SM. Leucas urticaefolia seed oil – a rich source of laballenic acid. Fette, Seifen, Anstrichmittel 1982;85:314–5.[90] Sinha S, Ansari AA, Osman SM. Leucas cephalotes: a new seed oil rich in labellenic acid. Chem Ind (London) 1978;2:67.[91] Payne-Wahl K, Kleiman R. Quantitation of estolide triglycerides in Sapium seeds by high-performance liquid chromatography with
infrared detection. J Am Oil Chem Soc 1983;60:1011–2.[92] Weichert H, Kolbe A, Kraus A, Wasternack C, Feussner I. Metabolic profiling of oxylipins in germinating cucumber seedlings-
lipoxygenase-dependent degradation of triacylglycerols and biosynthesis of volatile aldehydes. Planta 2002;215:612–9.[93] Brash AR, Baertschi SW, Ingram CD, Harris TM. Isolation and characterization of natural allene oxides: unstable intermediates in
the metabolism of lipid hydroperoxides. Proc Natl Acad Sci USA 1988;85:3382–6.[94] Ziegler J, Stenzel I, Hause B, Maucher H, Hamberg M, Grimm R, et al. Molecular cloning of allene oxide cyclase. The enzyme
establishing the stereochemistry of octadecanoids and jasmonates. J Biol Chem 2000;275:19132–8.[95] Itoh A, Schilmiller AL, McCaig BC, Howe GA. Identification of a jasmonate-regulated allene oxide synthase that metabolizes 9-
hydroperoxides of linoleic and linolenic acids. J Biol Chem 2002;277:46051–8.[96] Kongrit D, Jisaka M, Kobayasi K, Nishigaichi Y, Nishimura K, Nagaya T, et al. Molecular cloning, functional expression, and
tissue distribution of a potato sprout allene oxide synthase involved in a 9-lipoxygenase pathway. Biosci Biotechnol Biochem2006;70:2160–8.
[97] Crandall JK, Rambo E. Allene epoxidation: synthesis of functionalized lactones by the DMDO oxidation of allenic acids.Tetrahedron 2002;58:7027–36.
[98] Horvath A, Baeckvall J-E. Oxidation of allenes. Mod Allene Chem 2004;2:973–94.[99] Von Euler US. Uber die spezifische blutdrucksenkende Substanz des menschlichen Prostata- und Samenblasensekrets. Klin
Wochenschr 1935;14:1182–3.[100] Das UN. Essential Fatty acids – a review. Curr Pharm Biotechnol 2006;7:467–82.[101] Musiek ES, Yin H, Milne GL, Morrow JD. Recent advances in the biochemistry and clinical relevance of the isoprostane pathway.
Lipids. 2005;40:987–94.[102] Floyd MB, Jr. 15-Deoxy-16-hydroxy-16-allenyl prostane derivatives of the E and F series. 19 pp. US Patent: 4233453 19801111,
Application: US 79-46512 19790607, 1980.[103] Fried JH, Muchowski JM, Carpio H. Synthesis of lower chain allenic prostaglandins. 1. Prostaglandins. 1977;14:807–11.[104] Crabbe P, Fried JH, Guzman A. 4,5,10,13-Prostatetraenoic acid derivatives. 25 pp. US Patent: 3873598 19750325, Application: US
73-377106 19730706, 1975.[105] Muller P, Dammann HG, Simon B, Kommerell B. Drug therapy of peptic ulcer. What is coming up? Leber, Magen, Darm
1987;17:50–4.[106] Goa KL, Monk JP. Enprostil. A preliminary review of its pharmacodynamic and pharmacokinetic properties, and therapeutic
efficacy in the treatment of peptic ulcer disease. Drugs. 1987;34:539–59.[107] Eglen RM, Whiting RL. Characterization of the prostanoid receptor profile of enprostil and isomers in smooth muscle and platelets
in vitro. Br J Pharm 1989;98:1335–43.[108] Toshina K, Hirata I, Maemura K, Sasaki S, Murano M, Nitta M, et al. Enprostil, a prostaglandin-E(2) analogue, inhibits
interleukin-8 production of human colonic epithelial cell lines. Scand J Immunol 2000;52:570–5.[109] Watson SA, Durrant LG, Morris DL. The effect of the E2 prostaglandin enprostil, and the somatostatin analogue SMS 201 995, on
the growth of a human gastric cell line, MKN45G. Int J Cancer 1990;45:90–4.[110] Yactayo S, Patrice T, Foultier MT, Berrada A, Xu XN. Effect of gastrin and enprostil, a PGE2 analog, on colonic cancerous cell
growth. Gastroenterol Clin Biol 1991;15:519–24.

368 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
[111] Lartigue S, Corrion F, Bernades P, Bonfils S, Chevalier T, Laverdant C, et al. Effect of enprostil on the healing and the recurrence ofduodenal ulcer. Comparison with ranitidine. Gastroenterol Clin Biol 1994;18:617–22.
[112] McGrath MJ, Fletcher MT, Koenig WA, Moore CJ, Cribb BW, Allsopp PG, et al. A suite of novel allenes from Australianmelolonthine scarab beetles. Structure, synthesis, and stereochemistry. J Org Chem 2003;68:3739–48.
[113] Fletcher MT, McGrath MJ, Kitching W, Konig WA, Moore CJ, Cribb BW, et al. A novel group of allenic hydrocarbons from fiveAustralian (Melolonthine) beetles. Chem Commun 2001;10:885–6.
[114] Hayes PY, Fletcher MT, Chow S, McGrath MJ, Tu YQ, Zhang H, et al. Insect chemistry and chirality. Chirality. 2002;15:S116–27.[115] Theobald N, Shoolery JN, Djerassi C, Erdman TR, Scheuer PJ. Minor and trace sterols in marine invertebrates. 24-Ethyl-
D5,24(28),28-cholestatrien-3b-ol – a naturally occurring allenic marine sterol. J Am Chem Soc 1978;100:5574–5.[116] Awata N, Morisaki M, Fujimoto Y, Ikekawa N. Inhibitory effects of steroidal allenes on growth, development, and steroid
metabolism of the silkworm, Bombyx mori. J Insect Physiol 1976;22:403–8.[117] Morisaki M, Awata N, Fujimoto Y, Ikekawa N. Steroidal allenes as inhibitors of sterol biosynthesis. J Chem Soc, Chem Commun
1975;9:362–3.[118] Burger A, Roussel JP, Hetru C, Hoffmann JA, Luu B. Allenic cholesteryl derivatives as inhibitors of ecdysone biosynthesis.
Tetrahedron 1989;45:155–64.[119] Roussel JP, Burger A, Hetru C. Inhibitory activity of allene derivatives of cholesterol on the biosynthesis of ecdysone. Arch Int
Physiol Biochim 1990;98:253–9.[120] Beach DH, Chen F, Cushion MT, Macomber RS, Krudy GA, Wyder MA, et al. Effects of steroidal allenic phosphonic acid
derivatives on the parasitic protists Leishmania donovani, Leishmania mexicana mexicana, and Pneumocystis carinii carinii.Antimicrob Agents Chemother 1997;41:162–8.
[121] Robaire B, Covey DF, Robinson CH, Ewing LL. Selective inhibition of rat epididymal steroid D4-5a-reductase by conjugated allenic3-oxo-5,10-secosteroids. J Steroid Biochem 1977;8:307–10.
[122] Batzold FH, Covey DF, Robinson CH. Effects of novel acetylenic and allenic steroids on the rat prostate. Cancer Treatment Rep1977;61:255–7.
[123] Craig AS, Norman AW, Okamura WH. Studies of vitamin D (calciferol) and its analogs. 43. Two novel allenic side chain analogs of1a,25-dihydroxyvitamin D3. J Org Chem 1992;57:4374–80.
[124] Collins ED, Bishop JE, Bula CM, Acevedo A, Okamura WH, Norman AW. Effect of 25-hydroxyl group orientation on biologicalactivity and binding to the 1a,25-dihydroxy vitamin D3 receptor. J Steroid Biochem Mol Biol 2005;94:279–88.
[125] Williams JP, Boehm JC. The synthesis and reactivity of 3b-(2-alkynylsulfonyl)- and 3b-(2-alkynylsulfonylmethyl) androst-5-en-17-ones as inhibitors of glucose-6-phosphate dehydrogenase. Steroids. 1995;60:699–708.
[126] Bohlmann F, Jakupovic J, Gupta RK, King RM, Robinson H. Naturally occurring terpene derivatives. Part 296. Allenicgermacranolides, bourbonene-derived lactones, and other constituents from Vernonia species. Phytochemistry 1981;20:473–80.
[127] Krishna VK, Jain R, Singh P. Allenic germacranolides and ent-kaurene derivatives from Vernonia cotoneaster. J Med Arom PlantSci 1998;20:417–20.
[128] Bohlmann F, Zdero C, King RM, Robinson H. Naturally occurring terpene derivatives. Part 390. Hirsutinolides and othersesquiterpene lactones from Vernonia species. Phytochemistry 1982;21:695–9.
[129] Bohlmann F, Gupta RK, Jakupovic J, King RM, Robinson H. Naturally occurring terpene derivatives. 292. The first sesquiterpenelactone with an allenic group from Vernonia cotoneaster. Liebigs Ann Chem 1980;11:1904–6.
[130] Rho J-R, Lee H-S, Seo Y, Cho KW, Shin J. New xenicane diterpenoids from the gorgonian Acalycigorgia inermis. J Nat Prod2000;63:254–7.
[131] Cho GU, Kwon BM, Kwon HJ, Lee HS, Noh JR, Seo YW et al. Method for preparation of novel compounds, acalycixeniolide C,D, E and F, isolated from Acalycigorgia inermis. Republic Korean Kongkae Taeho Kongbo, Korean Patent: KR 2001087678 A20010921, Application: KR 2000-11560 20000308, 2001.
[132] Kwon HJ, Kim JH, Jung HJ, Kwon Y-G, Kim M-Y, Rho J-R, et al. Anti-angiogenic activity of acalycixenolide E, a novel marinenatural product from Acalycigorgia inermis. J Microbiol Biotechnol 2001;11:656–62.
[133] Hokama S, Tanaka J, Higa T, Fusetani N, Asano M, Matsunaga S, et al. Ginamallene, a new norditerpene with allene functionalityfrom four gorgonians of the genus Acalycigorgia. Chem Lett 1988;5:855–6.
[134] Fusetani N, Asano M, Matsunaga S, Hashimoto K. Bioactive marine metabolites. Part 27. Acalycixeniolides, novel norditerpeneswith allene functionality from two gorgonians of the genus Acalycigorgia. Tetrahedron 1989;45:1647–52.
[135] Fusetani N, Asano M, Matsunaga S, Hashimoto K. Bioactive marine metabolites series. Part 22. Acalycixeniolides, novelnorditerpenes which inhibit cell division of fertilized starfish eggs, from the gorgonian Acalycigorgia inermis. Tetrahedron Lett1987;28:5837–40.
[136] Wolff D. Allene, dihydronaphthalenon-derivate und andere Sekundarmetabolite aus Pilzen mariner Habitate sowieBeitrage zu derenBiosynthese. Dissertation, Germany: Gottingen; 2004. p. 202.
[137] Yan S-J, Zhang G-W, Su J-Y, Zeng L-M. Rare long conjugated diterpene ketene from the marine sponge Ircinia selaginea (Lamark).Gaodeng Xuexiao Huaxue Xuebao 2001;22:949–51.
[138] Oztunc A, Imre S, Wagner H, Norte M, Fernandez JJ, Gonzalez R. A new and highly oxygenated bromoallene from a marinesource. Tetrahedron Lett 1991;32:4377–80.
[139] Suzuki M, Takahashi Y, Mitome Y, Itoh T, Abe T, Masuda M. Brominated metabolites from an Okinawan Laurencia intricata.Phytochemistry 2002;60:861–7.
[140] Fukuzawa A, Kurosawa E, Tobetsu I. Constituents of marine plants. Part XXXVI. Laurallene, new bromoallene from the marinered alga Laurencia nipponica Yamada. Tetrahedron Lett 1979;30:2797–800.

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 369
[141] Kurata K, Furusaki A, Suehiro K, Katayama C, Suzuki T. Constituents of marine plants. 49. Isolaurallene, a new nonterpenoidC15- bromoallene, from the red alga Laurencia nipponica Yamada. Chem Lett 1982;7:1031–4.
[142] Suzuki T, Koizumi K, Suzuki M, Kurosawa E. Kumausallene, a new bromoallene from the marine red alga Laurencia nipponica
Yamada. Chem Lett 1983;10:1639–42.[143] Suzuki M, Segawa M, Suzuki T, Kurosawa E. Constituents of marine plants. 58. Structures of halogenated chamigrene derivatives,
minor constituents from the red alga Laurencia nipponica Yamada. Bull Chem Soc Jpn 1983;56:3824–6.[144] Grese TA, Hutchinson KD, Overman LE. General approach to halogenated tetrahydrofuran natural products from red algae of the
genus Laurencia. Total synthesis of (±)-kumausallene and (±)-1-epi-kumausallene. J Org Chem 1993;58:2468–77.[145] Abe T, Masuda M, Suzuki T, Suzuki M. Chemical races in the red alga Laurencia nipponica (Rhodomelaceae, Ceramiales). Phycol
Res 1999;47:87–95.[146] Suzuki M, Kurosawa E. Constituents of marine plants. Part 46. Okamurallene, a novel halogenated C15 metabolite from the red
alga Laurencia okamurai Yamada. Tetrahedron Lett 1981;22:3853–6.[147] Suzuki M, Kurosawa E. Structures of new bromoallenes from the red alga Laurencia okamurai Yamada. Tennen Yuki Kagobutsu
Toronkai Koen Yoshishu 1981;24:47–54.[148] Suzuki M, Sasage Y, Ikura M, Hikichi K, Kurosawa E. Constituents of marine plants. Part 72. Structure revision of
okamurallene and structure elucidation of further C15 non-terpenoid bromoallenes from Laurencia intricata. Phytochemistry1989;28:2145–8.
[149] Guella G, Chiasera G, Mancini I, Oztunc A, Pietra F. Twelve-membered O-bridged cyclic ethers of red seaweeds in the genusLaurencia exist in solution as slowly interconverting conformers. Chem Eur J 1997;3A:1223–31.
[150] Oztunc A, Imre S, Wagner H, Norte M, Fernandez JJ, Gonzalez R. A new haloether from Laurencia possessing alauroxacyclododecane ring. Structural and conformational studies. Tetrahedron 1991;47:2273–6.
[151] Guella G, Mancini I, Oztunc A, Pietra F. Conformational bias in macrocyclic ethers and observation of high solvolytic reactivity ata masked furfuryl (=2-furylmethyl) C-atom. Helv Chim Acta 2000;83:336–48.
[152] Lyakhova EG, Kalinovsky AI, Dmitrenok AS, Kolesnikova SA, Fedorov SN, Vaskovsky VE, et al. Structures and absolutestereochemistry of nipponallene and neonipponallene, new brominated allenes from the red alga Laurencia nipponica. TetrahedronLett 2006;47:6549–52.
[153] Okino T, Machiguchi Y, Kuramata K. Lipid-soluble fractions from marine algae as inhibitors of diatom adhesion. Jpn KokaiTokkyo Koho, 2007. p. 14. Japanese Patent: JP 2007045811 A 20070222.
[154] Okamoto Y, Nitanda N, Ojika M, Sakagami Y. Aplysiallene, a new bromoallene as an Na, K-ATPase inhibitor from the sea hare,Aplysia kurodai. Biosci Biotechnol Biochem 2001;65:474–6.
[155] Suzuki M, Kurosawa E. A C-15 non-terpenoid from the red alga Laurencia okamurai. Phytochemistry 1985;24:1999–2002.[156] Wright AD, Koenig GM, Sticher O. New sesquiterpenes and C15 acetogenins from the marine red alga Laurencia implicata. J Nat
Prod 1991;54:1025–33.[157] Oztunc A, Imre S, Lotter H, Wagner H. Two C15 bromoallenes from the red alga Laurencia obtusa. Phytochemistry 1991;30:255–7.[158] Dembitsky VM. Astonishing diversity of natural surfactants. 3. Carotenoid glycosides and isoprenoid glycolipids. Lipids.
2005;40:535–57.[159] Winterhalter P, Rouseff R. Carotenoid derived aroma compounds: an introduction. In: Winterhalter P, Rouseff R, editors.
Carotenoid derived aroma compounds. ACS symposium series, no 802. Washington, DC: ACS; 2002. p. 1–17.[160] Enzell C. Biodegradation of carotenoids – an important route to aroma compounds. Pure Appl Chem 1985;57:693–700.[161] Wahlberg I, Enzell CR. Tobacco isoprenoids. Nat Prod Rep 1987;4:237–76.[162] Schreier P. Aromaforschung Aktuell. Naturwissenschaften. 1995;82:21–7.[163] Beatriz M, Gloria A, Grulke EA, Gray JI. Effect of type of oxidation on b-carotene loss and volatile products formation in model
systems. Food Chem 1993;46:401–6.[164] Handelman GJ, van Kuijk GM, Chatterjee A, Krinsky NI. Characterisation of products formed during the autoxidation of b-
carotene. Free Radical Biol Med 1991;10:427–37.[165] Schwartz SH, Qin X, Zeevaart JAD. Characterization of a novel carotenoid cleavage dioxygenase from plants. J Biol Chem
2001;276:25208–11.[166] Baumes R, Wirth J, Bureau S, Gunata Y, Razungles A. Biogeneration of C13-norisoprenoid compounds: experiments supportive for
an apo-carotenoid pathway in grapevines. Anal Chim Acta 2002;458:3–14.[167] Meinwald J, Erickson K, Hartshorn M, Meinwald YC, Eisner T. Defensive mechanisms of arthropods. XXIII. An allenic
sesquiterpenoid from the grasshopper Romalea microptera. Tetrahedron Lett 1968;25:2959–62.[168] Russell SW, Weedon BCL. Allenic ketone from grasshoppers. Chem Commun (London) 1969;3:85–6.[169] Isoe S, Katsumura S, Suong BH, Sakan T. Biogenetic type synthesis of grasshopper ketone and loliolide and a possible biogenesis of
allenic carotenoids. Tetrahedron Lett 1971;16:1089–92.[170] Sefton MA, Skouroumounis GK, Massy-Westropp RA, Williams PJ. Norisoprenoids in Vitis vinifera white wine grapes and the
identification of a precursor of damascenone in these fruits. Aust J Chem 1989;42:2071–84.[171] Miyase T, Ueno A, Takizawa N, Kobayashi H, Karasawa H. Studies on the glycosides of Epimedium grandiflorum Morr. var.
thunbergianum (Miq.) Nakai. I. Chem Pharm Bull 1987;35:1109–17.[172] Miyase T, Ueno A, Takizawa N, Kobayashi H, Oguchi H. Ionone and lignan glycosides from Epimedium diphyllum. Phytochemistry
1989;28:3483–5.[173] Skouroumounis GK, Winterhalter P. Glycosidically bound norisoprenoids from Vitis vinifera cv. Riesling Leaves. J Agric Food
Chem 1994;42:1068–72.

370 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
[174] Takeda Y, Ooiso Y, Masuda T, Honda G, Otsuka H, Sezik E, et al. Iridoid and eugenol glycosides from Nepeta cadmea.Phytochemistry 1998;49:787–91.
[175] Takeda Y, Mima C, Masuda T, Hirata E, Takushi A, Otsuka H. Glochidioboside, a glucoside of (7S,8R)-dihydrodehydrodiconiferylalcohol from leaves of Glochidion obovatum. Phytochemistry 1998;49:2137–9.
[176] Mitrocotsa D, Skaltsounis A, Mitaku S, Harvala C, Tillequin F. Flavonoid and terpene glycosides from European Ebenus species.Biochem Syst Ecol 1999;27:305–7.
[177] Fujimatu E, Ishikawa T, Kitajima J. Aromatic compound glucosides, alkyl glucoside and glucide from the fruit of anise.Phytochemistry 2003;63:609–16.
[178] Iijima T, Yaoita Y, Kikuchi M. Constituents of Erigeron annuus, Erigeron philadelphicus and Erigeron sumatrensis. J Nat Med(Tokyo, Japan) 2003:57–75.
[179] Assaf MH, Kamel MS, Bishay DW. Phenolic and aliphatic glucosides from Cryptostegia grandiflora and cardiotonic activity ofcryptostigmin II. Bull Pharm Sci, Assiut University 2003;26:41–8.
[180] Hisamoto M, Kikuzaki H, Nakatani Ni. Constituents of the leaves of Peucedanum japonicum Thunb. and their biological activity. JAgr Food Chem 2004;52:445–50.
[181] Michalska K, Kisiel W. Taxonomically significant guaianolides from Taraxacum obovatum. Biochem Syst Ecol2004;32:765–8.
[182] Takeda Y, Masuda T, Morikawa H, Ayabe H, Hirata E, Shinzato T, et al. Lanceocrepidiasides A–F, glucosides of guaiane-typesesquiterpene from Crepidiastrum lanceolatum. Phytochemistry 2005;66:727–32.
[183] Wang J, Shen Y, He H, Kang W, Hao X. Norsesquiterpenoid and sesquiterpenoid glycosides from Evodia austrosinensis. Planta Med2005;71:96–8.
[184] De Marino S, Borbone N, Zollo F, Ianaro A, Di Meglio P, Iorizzi M. Megastigmane and phenolic components from Laurus nobilis
L. leaves and their inhibitory effects on nitric oxide production. J Agr Food Chem 2004;52:7525–31.[185] Umehara K, Hattori I, Miyase T, Ueno A, Hara S, Kageyama C. Studies on the constituents of leaves of Citrus unshiu Marcov.
Chem Pharm Bull 1988;36:5004–8.[186] Ishikawa T, Kondo K, Kitajima J. Water-soluble constituents of coriander. Chem Pharm Bull 2003;51:32–9.[187] Kitajima J, Ishikawa T, Satoh M. Polar constituents of celery seed. Phytochemistry 2003;64:1003–11.[188] Yoshinari K, Sashida Y, Shimomura H. Two new lignan xylosides from the barks of Prunus ssiori and Prunus padus. Chem Pharm
Bull 1989;37:3301–3.[189] Yoshikawa M, Ueda T, Matsuda H, Yamahara J, Murakami N. Absolute stereostructures of hydramacrosides A and B, new
bioactive secoiridoid glucoside complexes from the leaves of Hydrangea macrophylla Seringe var. thunbergii Makino. Chem PharmBull 1994;42:1691–3.
[190] Osorio C, Duque C, Fujimoto Y. C13-Norisoprenoid glucoconjugates from lulo (Solanum quitoense L.) leaves. J Agr Food Chem1999;47:1641–5.
[191] Takeda Y, Kinugawa M, Masuda T, Honda G, Otsuka H, Sezik E, et al. Phlomisethanoside, a phenylethanoid glycoside fromPhlomis grandiflora var. grandiflora. Phytochemistry 1999;51:323–5.
[192] Ito H, Kobayashi E, Li S-H, Hatano T, Sugita D, Kubo N, et al. Megastigmane glycosides and an acylated triterpenoid fromEriobotrya japonica. J Nat Prod 2001;64:737–40.
[193] Takeda Y, Isai N, Masuda T, Honda G, Takaishi Y, Ito M, et al. K.Phlomisflavosides A and B, new flavonol bisglycosides fromPhlomis spinidens. Chem Pharm Bull 2001;49:1039–41.
[194] Lee T-H, Wang G-J, Lee C-K, Kuo Y-H, Chou C-H. Inhibitory effects of glycosides from the leaves of melaleuca quinquenervia onvascular contraction of rats. Planta Med 2002;68:492–6.
[195] Takeda Y, Shimidzu H, Mizuno K, Inouchi S, Masuda T, Hirata E, et al. An iridoid glucoside dimer and a non-glycosidic iridoidfrom the leaves of Lasianthus wallichii. Chem Pharm Bull 2002;50:1395–7.
[196] Kanchanapoom T, Noiarsa P, Tiengtham P, Otsuka H, Ruchirawat S. Acetophenone diglycosides from Erythroxylum cambodianum.Chem Pharm Bull 2005;53:579–81.
[197] Perrone A, Plaza A, Bloise E, Nigro P, Hamed AI, Belisario MA, et al. Cytotoxic furostanol saponins and a megastigmaneglucoside from Tribulus parvispinus. J Nat Prod 2005;68:1549–53.
[198] Osorio C, Duque C, Fujimoto Y. Glycosides of C13-norisoprenoids as aroma precursors in lulo (Solanum quitoense L.) leaves.Natural product analysis: chromatography, spectroscopy, biological testing, [symposium], Wuerzburg, Germany, September 1997,meeting. Wiesbaden, Germany: Vieweg; 1998. p. 353–5.
[199] Shiraga Y, Okano K, Akira T, Fukaya C, Yokoyama K, Tanaka S, et al. Structures of potent antiulcerogenic compounds fromCinnamomun cassia. Tetrahedron 1988;44:4703–11.
[200] Skouroumounis GK, Massy-Westropp RA, Sefton MA, Williams PJ. Precursors of damascenone in fruit juices. Tetrahedron Lett1992;33:3533–6.
[201] Renaud JM, Tsoupras G, Stoeckli-Evans H, Tabacchi R. A novel allenic epoxycyclohexane and related compounds from Eutypa
lata (Pers: F.) Tul. Helv Chim Acta 1989;72:1262–7.[202] Sanson DR, Gracz H, Tempesta MS, Fukuda DS, Nakasukasa WM, Sands TH, et al. Tetrahedron 1991;47:3633–44.[203] Britton G, Liaaen-Jensen S, Pfander H, editors. Carotenoids handbook. Basel: Birkhauser Verlag; 2004.[204] Goodwin T W. The biochemistry of carotenoids, plants, vol. 1. London: Chapman and Hall; 1980.[205] Liaeen-Jensen S. Carotenoids in chemosystematics. In: Britton G, Liaaen-Jensen S, Pfander H, editors. Carotenoids biosynthesis and
metabolism, vol. 3. Basel: Birkhauser; 1998. p. 217–47.

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 371
[206] Liaaen-Jensen S. Marine carotenoid. In: Scheuer PJ, editor. Marine natural products. Chemical and biochemical perspective, vol. 2.New York: Academic Press; 1978. p. 1–73.
[207] Bonnett R, Spark AA, Tee JL, Weedon BCL. Fucoxanthin. Proc Chem Soc London 1964:419.[208] Bonnett A, Mallams AK, Tee JL, Weedon BCL, McCormick A. Fucoxanthin and related pigments. Chem Commun
1966:515–6.[209] Bonnett A, Mallams AK, Spark AA, Tee JL, Weedon BCL. Carotenoids and related compounds. Part XX. Structure and reactions
of fucoxanthin. J Chem Soc (C) 1969:429–54.[210] De Ville TE, Hursthouse MB, Russell SW, Weedon BCL. Absolute configuration of carotenoids. Chem Commun 1969:1311–2.[211] Strain HH, Manning WM. The occurrence and innerconversion of various fucoxanthin. J Am Chem Soc 1942;64:1235.[212] Englert G, Bjornland T, Liaaen-Jensen S. 1D and 2D NMR study of some allenic carotenoids of the fucoxanthin series. Magn Reson
Chem 1990;28:519–28.[213] Haugan JA, Englert G, Glinz E, Liaaen-Jensen S. Algal carotenoids. 48. Structural assignments of geometrical isomers of
fucoxanthin. Acta Chem Scand 1992;46:389–95.[214] Haugan JA, Liaaen-Jensen S. Naturally occurring stereoisomers of fucoxanthin. Phytochemistry 1992;31:1359–61.[215] Haugan JA, Liaaen-Jensen S. Isolation and characterization of four allenic (6 0S)-isomers of fucoxanthin. Tetrahedron Lett
1994;35:2245–8.[216] Haugan JA, Englert G, Liaaen-Jensen S. Algal carotenoids 50. Alkali lability of fucoxanthin – reactions and products. Acta Chem
Scand 1992;46:614–24.[217] Haugan JA, Liaaen-Jensen S. Blue carotenoids. Part 1. Novel oxonium ions derived from fucoxanthin. Acta Chem Scand
1994;48:68–75.[218] Liaaen-Jensen S. Marine carotenoids-selected topics. New J Chem 1990;14:747–59.[219] Yamano Y, Tode C, Ito M. Carotenoids and related polyenes. Part 3. First total synthesis of fucoxanthin and halocynthiaxanthin
using oxo-metallic catalyst. J Chem Soc, Perkin Trans 1 1995:1895–904.[220] Palermo JA, Gros EG, Seldes AM. Carotenoids from three red algae of the corallinaceae. Phytochemistry 1991;30:2983–6.[221] Fiksdahl A, Withers N, Gullard RL, Liaaen-Jensen S. Carotenoids of the Raphidophyceae – a chemosystematic contributions.
Comp Biochem Physiol 1984;78B:265–71.[222] Haugan JA, Liaaen-Jensen S. Algal carotenoids 54, carotenoids of brown alge Phaeophyceae. Biochem Syst Ecol 1994;22:31–41.[223] Pennington F, Guillard RL, Liaaen-Jensen S. Carotenoid distribution patterns in Bacillariophyceae (diatom). Biochem Syst Ecol
1988;16:589–92.[224] Arprin N, Svbc WA, Liaaen-Jensen S. New fucoxanthin-related carotenoids from Coccolithus huxleyi. Phytochemistry
1976;15:529–32.[225] Bjornland T, Guillard RRL, Liaaen-Jensen S. Algal carotenoids. Part 39. Phaeocystis sp. clone 677-3 – a topical marine planktonic
prymnesiophyte with fucoxanthin and 190-aceloxyfucoxanthins as chemosystematic carotenoid markers. Biochem Syst Ecol1988;16:445–52.
[226] Bjerkeng B, Vernet M, Nielsen V, Liaeen-Jensen S. Carotenoids of Chrysochromulina polylepis (Prymnesiophceae). Biochem SystEcol 1990;18:303–6.
[227] Egeland ES, Garrido JL, Zapata M, Maestro MA, Liaaen-Jensen S. Algal carotenoids. Part 64. Structure and chemistry of 4-keto-190-hexanoyloxyfucoxanthin with a novel carotenoid end group. J Chem Soc, Perkin Trans 1 2000:1223–30.
[228] Bjornland T, Liaaen-Jensen S, Throndsen J. Carotenoids of the marine Chrysophyte Pelagococcus subviridis. Phytochemistry1989;28:3347–53.
[229] Shaw BA, Andersen RJ, Harrison PJ. Feeding deterrence properties of apo-fucoxanthinoids from marine diatoms. I. Chemicalstructures of apo-fucoxanthinoids produced by Phaeodactylum tricornutum. Mar Biol 1995;124:467–72.
[230] Doi Y, Ishibashi M, Yamaguchi N, Kobayashi J. Isolation of apo-90-fucoxanthinone from the cultured marine dinoflagellateAmphidinium sp.. J Nat Prod 1995;58:1097–9.
[231] Mori K, Ooi T, Hiraoka M, Oka N, Hamada H, Tamura M, et al. Fucoxanthin and its metabolites in edible brown algae cultivatedin deep seawater. Mar Drugs 2004;2:63–72.
[232] Nitsche H. The structure of vaucheriaxanthin. Z Naturforsch 1973;28C:641–5.[233] Fiksdahl A, Withers N, Guillard RL, Liaaen-Jensen S. Carotenoids of the Rapidophyceae – a chemosystematic contributions. Comp
Biochem Physiol 1984;78B:265–71.[234] Sch}utt F. Uber Peridineenfarbstoffe. Ber Deut Bot Ges 1980;8:9–32.[235] Strain HH, Svec WA, Aitzetm}uller K, Grandolfo MC, Katz JJ, Kosen H, et al. The structure of peridinin, the characteristic
dinoflagellate carotenoid. J Am Chem Soc 1971;93:1823–5.[236] Johansen JE, Borch G, Liaaen-Jensen S. Chirality of peridinin and dinoxanthin. Phytochemistry 1980;19:441–4.[237] Yamano Y, Ito M. First total synthesis of (±)-peridinin, (±)-pyrrhoxanthin and optical active peridinin. J Chem Soc, Perkin Trans 1
1993:1599–610.[238] Furuichi N, Hara H, Osaki T, Nakano M, Mori H, Katunuma S. Stereocontrolled total synthesis of a polyfunctional carotenoid,
peridinin. J Org Chem 2004;69:7949–59.[239] Yamao Y, Sumiya S, Suzuki K, Kurimoto Y, Koyama Y, Shimamura T, et al. A novel photoproduct of peridinin, A 6S allenic
isomer. Tetrahedron Lett 1992;33:2991–4.[240] Yamao Y, Sumiya S, Ito M. Carotenoids and related poyenes. Part 2. Photoisomerization of an allenic carotenoid, peridinin, and
allenic model compounds. J Chem Soc, Perkin Trans 1 1995:167–76.

372 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
[241] Haugan JA, Englert G, Aakermann T, Glinz E, Liaaen-Jensen S. Algal carotenoids 58. Isomerization studies on peridinin. ActaChem Scand 1994;48:769–79.
[242] Swift IE, Milborrow BV. Stereochemistry of allene biosynthesis and the formation of the acetylenic carotenoid diadinoxanthin andperidinin (C37) from neoxanthin. Biochem J 1981;199:69–74.
[243] Swift IE, Milborrow BV, Jeffery SW. Formation of neoxanthin, diadinoxanthin and peridinin from [14C] zeaxanthin by a cell-freesystem from Amphidinium carterae. Phytochemistry 1982;21:2859–64.
[244] Johansen JE, Svec WA, Liaaen-Jensen S, Haxo FT. Carotenoids of the dinophyceae. Phytochemistry 1974;13:2261–71.[245] Suzuki M, Watanabe K, Fujiwara S, Kurasawa T, Wakabayashi T, Tsuzuki M, et al. Isolation of peridinin-related norcarotenoids
with cell growth-inhibitory activity from the cultured dinoflagellate of Symbiodinium sp., a symbiont of the Okinawan soft coralClavularia virdis, and analysis of fatty acid of the dinoflagellate. Chem Phrm Bull 2003;51:724–7.
[246] Kylin H. The carotenoid pigments of algae. Kgl Fysiograf Sallskap Lund, Forh 1939;9:213–31.[247] Maoka T, Tsushima M, Nishino H. Isolation and characterization of dinochrome A and B, anti-carcinogenic active carotenoids
from the fresh water red tide, Peridinium bipes. Chem Pharm Bull 2002;50:1630–3.[248] Bjornland T, Fiksdahl A, Skjetne T, Krane J, Liaaen-Jensen S. Gyroxanthin – the first allenic acetylenic carotenoid. Tetrahedron
2000;56:9047–56.[249] Loeblich III AR. Dinoflagellate evolution: speculation and evidence. J Protozool 1976;23:13–28.[250] Aakermann T, Guillard RRL, Liaaen-Jensen S. Algal carotenoids 55. Structural elucidation of (3S,5R,6R,3 0S,50R,60S)-130-cis-70,8 0-
dihydroneoxanthin-200-al 30-b-D-lactoside (P457). Part 1. Reisolation, derivatization and synthesis of model compounds. Acta ChemScand 1993;47:1207–13.
[251] Englert G, Aakermann T, Schiedt K, Liaaen-Jensen S. Structure elucidation of the algal carotenoid (3S,5R,6R,30S,5 0R,60S)-130-cis-7 0,8 0-dihydroneoxanthin-200-al 30-b-D-lactoside (P457). Part 2, NMR studies. J Nat Prod 1995;58:1675–82.
[252] Strain HH. Leaf xanthophylls, Carnegic Institute Publication. No 490, Washington; 1938.[253] Cholnoky L, Gyorgyft K, Ronai A, Szabolcs J, Toth G, Galasko G, et al. Carotenoids and related compounds. XXI. Structure of
neoxanthin (foliaxanthin). J Chem Soc (C) 1969:1256–63.[254] Marki-Fischer E, Eugster CH. Neoflor and 6-epineoflor from flowers of Trollius europaeus; highfield 1H-NMR spectra of (all-E)-
neoxanthin and (90Z)-neoxanthin. Helv Chim Acta 1990;73:1637–43.[255] Baumeler A, Eugster CH. Synthesise of (6R, all-E)-neoxanthin and related allenic carotenoids. Helv Chim Acta 1992;75:773–90.[256] Baumeler A, Zerbe O, Kunz R, Eugster CH. (6R,90Z)-Neoxanthin: synthesis, physical properties, spectra and calculation of its
conformation in solution. Helv Chim Acta 1994;77:909–30.[257] Takaichi S, Mimuro M. Distribution and geometrical isomer of neoxanthin in oxygenic phototrophs: 9 0-cis, a sole molecular form.
Plant Cell Physiol 1998;39:968–77.[258] Strand A, Kvernberg K, Karlsen AM, Liaaen-Jensen S. Geometrical E/Z isomers of (6R)- and (6S)-neoxanthin and biological
implications. Biochem Syst Ecol 2000;28:443–55.[259] Bouvier F, d’Harlingue A, Backhaus RA, Kumagai MH, Camara B. Identification of neoxanthin synthase as a carotenoid cyclase
paralog. Eur J Biochem 2000;267:6346–52.[260] Al-Babili S, Hugueney P, Schledz M, Welsch R, Frohnmeyer H, Laule O, et al. Identification of a novel gene coding for neoxanthin
synthase from Solanum tuberosum. FEBS Lett 2000;485:168–72.[261] Marki-Fischer E, Buchecker R, Eugster CH. Separation and absolute configuration of the C(8) epimeric (all-E)-neochrome
(Trollichrome) and -dinochrome. Helv Chim Acta 1984;65:461–6.[262] Nitsch H. Structurs of mimulaxanthin. Z Naturforsch 1973;28C:481–7.[263] Buchecker R, Eugster CH. Mimulaxanthin, the main carotenoid in Laminum montanum, Determination of its absolute
configuration. Absolute configuration of deepoxyneoxanthin by its correlation with neoxanthin. Helv Chim Acta1980;63:2531–7.
[264] Baumeler A, Eugster CH. Synthesis of enantiometrically pure mimulaxanthin and its (9Z,90Z)- and (15Z)-isomers. Helv Chim Acta1991;74:469–86.
[265] Fujiwara Y, Hashimoto K, Manabe K, Maoka T. Structures of tobiraxanthins A1, A2, A3, B, C and D, new carotenoids from theseeds of Pittosporum tobira. Tetrahedron Lett 2002;43:4385–8.
[266] Maoka T, Fujiwara Y, Hashimoto K, Naoshige A. 5-Hydroxy-seco-carotenoids from Pittosporum tobira. Phytochemistry2006;67:2120–55.
[267] Maoka T, Akimoto N, Hashimoto K, Kuroda Y, Fujiwara Y. Structures of novel carotenoids pittosporumxanthins B1, B2, C1 andC2 from the seeds of Pittosporum tobira by using of MS/MS spectrometry. Tennen Yuki Kagobuts Toronkai Koen Yoshishu (Jpn)1995;37:373–8.
[268] Deli J, Molnar P, Osz E, Toth G. Capsoneoxanthin, a new carotenoid isolated from the fruits of Asparagus falcatus. TetrahedronLett 2000;41:8153–5.
[269] Liaeen-Jensen S. Carotenoids in food chains. In: Britton G, Liaaen-Jensen S, Pfander H, editors. Carotenoids biosynthesis andmetabolism, vol. 3. Basel: Birkhauser; 1998. p. 259–371.
[270] Heilbron IM, Jackson H, Jones RN. The lipochromes of sea anemones. 1. Carotenoid pigments of Actinia equina, Anemonia sulcata,Actionoloba dianthus and Tealia felina. Biochem J 1935;29:1384–8.
[271] Fiksdahl A, Hallenstvet M, Beress L, Liaaen-Jensen S. Animal carotenoids 18. Identity of Sulcatoxanthin and Peridinin. BiochemSyst Ecol 1979;7:173–4.
[272] Marinus DL, de A Epifanio R. A new germacrane sesquiterpen from the Brazilian endemic gorgonian Phyllogorgia dilatata Esper. JBraz Chem Soc 1998;9:586–90.

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 373
[273] Matsuno T, Sakaguchi S. Isolation of Peridinin from The Sea Squirt Botrylloides violaceus, ‘Itaboya’ in Japanese. Bull Jpn Soc SciFish 1984;50:1267–77.
[274] Matsuno T, Maoka T, Shiba K, Ookubo M. Isolation of fucoxanthinol from short-necked clam Tapes philippinarum [Asari inJapanese]. Nippon Suisan Gaku 1986;52:167.
[275] Maoka T, Fujiwara Y, Hashimoto K, Akimoto N. Characterizatoin of fucoxanthin and fucoxanthinol esters in the Chinese surfclam, Mactra chinensis. J Agric Food Chem 2007;55:1563–7.
[276] Hallenstvea M, Ryberg E, Liaaen-Jensen S. Animal carotenoids – XIV. Carotenoids of Psammechinus miliaris (sea-urchin). CompBiochem Physiol 1978;60B:173–5.
[277] Tsushima M, Matsuno T. Comparative biochemical studies of carotenoids in sea urchins-I. Comp Biochem Physiol1990;96B:801–10.
[278] Matuno T, Tsushima M. Carotenoids in sea urchins. In: Lawrence JM, editor. Edible sea urchins: biology and biology. Amsterdam:Elsevier; 2001. p. 115–38.
[279] Ookubo M, Matsuno T. Carotenoids of sea squirts – II. Comparative biochemical studies of carotenoids in sea squirts Comp.Biochem Physiol 1985;81B:137–41.
[280] Hertberg S, Partail V, Liaaen-Jensen S. Animal carotenoids. 32. Carotenoids of Mytilus edulis (Edible Mussel). Acta Chem Scand1988;B42:495–503.
[281] Partali V, Tanger K, Liaaen-Jensen S. Carotenoids in food chain studies – III. Resorption and metabolic transformation ofcarotenoids in Mytilus edulis (Edible mussel). Comp Biochem Physiol 1989;92B:239–46.
[282] Galasko G, Hora J, Toube TP, Weedon BCL, Andre D, Barbier M, et al. Carotenoids and related compounds. Part XXII. Alleniccarotenoids in sea urchins. J Chem Soc (C) 1969:1264–5.
[283] Maoka T, Fujiwara Y, Hashimoto K, Akimoto N. Structure of new carotenoids from corbicula clam Corbicula japonica. J Nat Prod2005;68:1341–4.
[284] Maoka T, Fujiwara Y, Hashimoto K, Akimoto N. Carotenoids in three species of corbicula clams Corbicula japonica, Corbicula
sandai, and Corbicula sp. (Chinese freshwater corbicula clam). J Agric Food Chem 2005;53:8357–64.[285] Maoka T, Hashimoto K, Akimoto N, Fujiwara Y. Structures of five new carotenoids from the oyster Crassostrea gigas. J Nat Prod
2001;64:578–81.[286] Matsuno T, Ookubo M, Komori T. Carotenoids of tunicates, III. The structural elucidation of two new marine carotenoids,
amarouciaxanthin A and B. J Nat Prod 1985;48:606–13.[287] Matuno T, Sakaguchi S, Ookubo M, Maoka T. Isolation of amarouciaxanthin A from the bivalve Paphia euglypta (Sudaregai in
Japanese). Bull Jap Soc Sci Fish 1985;51:1909.[288] Rho J-R, Seo Y, Cho KW, Song J-I, Shin J. Isolation of a new carotenoid pigment from an undescribed gorgonian of the genus
Muricella. Bull Korean Chem Soc 1996;17:529–31.[289] Matsuno T, Ohkubo M, Toriiminami Y, Tsushima M, Sakaguchi S, Minamai T, et al. Carotenoids in food chain between
freshwater fish and aquatic insects. Comp Biochem Physiol 1999;124B:341–5.[290] Strand A, Herstad O, Liaaen-Jensen S. Fucoxanthin metabolites in egg yolks of laying hens. Comp Biochem Physiol
1998;119A:963–74.[291] Asai A, Sugawara T, Ono H, Nagao A. Biotranfsormation of fucoxanthinol into amarouciaxanthin A in mice and HEPG2 cells:
Formation and cytotoxicity of fucoxanthin metabolites. Drug Metabol Dispos 2004;32:205–11.[292] Alberte RS, Friedman AL, Gustafson DL, Rudnick MS, Lyman H. Light-harvesting systems of brown algae and diatoms. Isolation
and characterization of chlorophyll a/c and chlorophyll a/fucoxanthin pigment-protein complexes. Biochim Biophys Acta1981;635:304–16.
[293] Buchel C. Fucoxanthin-chlorophyll proteins in diatoms: 18 and 19 kDa subunits assemble into different oligomeric states.Biochemistry. 2003;42:13027–34.
[294] Hofmann E, Wrench PM, Sharples EP, Hiller RG, Welte W, Diederichs K. Structural basis of light harvesting by carotenoids:peridinin-chlorophyll-protein from Amphidinium carterae. Science. 1996;272:1788–91.
[295] Kuhlbrandt W, Wang DN, Fujiyoshi Y. Atomic model of plant light-harvesting complex by electron crystallography. Nature.1994;367:614–21.
[296] Liu Z, Yan H, Wang K, Kuang T, Zhang J, Gui L, et al. Crystal structure of spinach major light-harvesting complex at 2.72 Aresolution. Nature. 2004;428:287–94.
[297] Pinto E, Catalani LH, Lopes NP, Di Mascio P, Colepicolo P. Peridinin as the major biological carotenoid quencher of singlet oxygenin marine algae Gonyaulax polyedra. Biochem Biophys Res Commun 2000;268:496–500.
[298] Hirayama O, Nakamura K, Hamada S, Kobayashi Y. Singlet oxygen quenching ability of naturally occurring carotenoids. Lipids.1994;29:149–50.
[299] Shimidzu N, Goto M, Miki W. Carotenoid as singlet oxygen quenchers in marine organisms. Fish Sci 1996;62:134–7.[300] Miki W. Biological functions and activities of animal carotenoids. Pure Appl Chem 1991;63:141–6.[301] Yan X, Chuda Y, Suzuki M, Nagata T. Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible sea weed.
Biosci Biotechnol Biochem 1999;63:605–7.[302] Nomura T, Kikuchi M, Kubodera A, Kawasaki Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-
picrylhydrazyl (DPPH). Biochem Mol Bio Int 1997;42:361–70.[303] Kawakami T, Tsushima M, Katabami Y, Mine M, Ishida A, Matsuno T. Effect of b,b-carotene, b-echinenone, astaxanthin,
fucoxanthin, vitamin A and vitamin E on the biological defense of the sea urchin Pseudocentrotus depresssus. J Exp Mar Biol Ecol1998;226:125–74.

374 V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375
[304] Krinsky NI, Mayne ST, Sies H. Carotenoids in health and disease. New York: Marcel Dekker; 2004.[305] Okuzumi J. Antiproliferative and antitumor promoting activities of fucoxanthin, a natural carotenoid. J Kyoto Pref Univ Med
1991;100:551–66.[306] Okuzumi J, Nishino H, Murakoshi M, Iwashima A, Tanaka Y, Yamane T, et al. Inhibitory effects of fucoxanthin, a natural
carotenoid, on N-myc expression and cell cycle progression in human malignant tumor cells. Cancer Lett 1990;55:75–81.[307] Hosokawa M, Wanezaki S, Miyauchi K, Kurihara H, Kohono H, Kawabata J, et al. Apoptosis-inducing effect of fucoxanthin on
human leukemia cell HL-60. Food Sci Technol Res 1999;5:243–6.[308] Kotake-Nara E, Kushiro M, Zhang H, Sugawara T, Miyashita K, Nagao A. Carotenoids affect proliferation of human prostate
cancer cells. J Nutr 2001;131:3303–6.[309] Tsushima M, Maoka T, Katuyama M, Kozuka M, Matsuno T, Tokuda H, et al. Inhibitory effect of natural carotenoids on Epstein-
Barr virus activation activity of a tumor promoter in Raji cells. A screening study for anti-tumor promoters. Biol Pharm Bull1995;18:227–33.
[310] Kim JM, Araki S, Kim DJ, Park CB, Takasuka N, Baba-Toriyama H, et al. Chemopreventive effects of carotenoids and curcuminson mouse colon carcinogenesis after 1,2-dimethylhydrazine initiation. Carcinogenesis. 1998;19:81–5.
[311] Okuzumi J, Takahashi T, Yamane T, Kitao T, Inagaki M, Ohya K, et al. Inhibitory effects of fucoxanthin, a natural carotenoid, onN-ethyl-N 0-nitro-N-nitrosoguanidine-induced mouse duodenal carcinogenesis. Cancer Lett 1993;68:159–68.
[312] Hosokawa M, Kudo M, Maeda H, Kohno H, Tanaka T, Miyashita K. Fucoxanthin induces apotosis and enhances theantiproliferative effect on the PPARc ligand, troglitazone, on colon cancer cells. Biochim Biophys Acta 2004;1675:113–9.
[313] Kotake-Nara E, Sugawara T, Nagao A. Antiproliferative effect on neoxnthin and fucoxanthin on cultured cells. Fish Sci2005;71:459–61.
[314] Nishino H. Cancer prevention by carotenoids. Mutat Res 1998;402:159–63.[315] Maeda H, Hosokawa M, Sashima T, Funayama K, Miyashita K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows
aniobesity effect through UCP1 expression in white adipose tissues. Biochem Biophys Res Commun 2005;332:392–7.[316] Shiratori K, Ohigashi K, Ilieva I, Jin X-H, Koyama Y, Miyashita K, et al. Effects of fucoxanthin on lipopolysaccharide-induced
inflammation in vitro and in vivo. Exp Eye Res 2005;81:271–7.[317] Chang JM, Lin JK. Isolation of neoxanthin from spinach and its prevention of lipid peroxidation. J Chin Med 1993;4:235–45.[318] Chang JM, Chen WC, Hong D, Lin JK. The inhibition of DMBA-induced carcinogensis by neoxanthin in hamster buccal pouch.
Nutr Cancer 1995;24:325–33.[319] Murakami A, Nakashima M, Koshiba T, Maoka T, Nishino H, Yano M, et al. Modifying effects of carotenoids on superoxide and
nitric oxide generation from stimulated leukocites. Cancer Lett 2000;149:115–23.[320] Nishino H, Satomi Y, Tokuda H, Nishino A, Iwashima A, Tanaka Y, et al. Antitumor activity of peridinin and its structurally
related butenolide compound. J Kyoto Pref Univ Med 1991;100:831–5.[321] Takashima J, Asano S, Ohsaki A. Structures and activities of constituents from Brazilian medicinal plant, Brosimum acutifolium.
Tennen Yuki Kagobutsu Toronkai Koen Yoshishu 2000;42:487–92.[322] Arnone A, Cardillo R, Nasini G, Vajna de Pava O. Secondary mold metabolites. Part 40. Two cinnamic allenic ethers from the
fungus Clitocybe eucalyptorum. Phytochemistry 1993;32:1279–81.[323] Zeng X, Wang H, He L, Lin Y, Li Z. Medium optimization of carbon and nitrogen sources for the production of eucalyptene A and
xyloketal A from Xylaria sp. 2508 using response surface methodology. Process Biochem (Amsterdam) 2006;41:293–8.[324] Edwards RL, Anderson JR, Whalley AJS. Metabolites of the higher fungi. Part 20. Chesterseine, 4-(4-methoxyphenoxy)buta-1,2-
diene: an allenic ether from the fungus Hypoxylon chestersii. Phytochemistry 1982;21:1721–3.[325] Wang SY, Mao WW, She ZG, Li CR, Yang DQ, Lin YC, et al. Synthesis and biological evaluation of 12 allenic aromatic ethers.
Bioorg Med Chem Lett 2007;17:2785–8.[326] Lin Y, Wu X, Feng S, Jiang G, Zhou S, Vrijmoed LLP, et al. A novel N-cinnamoylcyclopeptide containing an allenic ether from the
fungus Xylaria sp. (strain #2508) from the South China Sea. Tetrahedron Lett 2001;42:449–51.[327] Chilton WS, Tsou G, Kirk L, Benedict RG. Naturally-occurring allenic amino acid. Tetrahedron Lett 1968;60:6283–4.[328] Chilton WS, Tsou G. A chloro amino acid from Amanita solitaria. Phytochemistry 1972;11:2853–7.[329] Castelhano AL, Krantz A. Allenic amino acids. 1. Synthesis of c-allenic GABA by a novel aza-Cope rearrangement. J Am Chem Soc
1984;106:1877–9.[330] Doutheau A, Saba A, Gore J, Quash G. Preparation of allenic amino alcohols. Tetrahedron Lett 1982;23:2461–4.[331] Castelhano AL, Pliura DH, Taylor GJ, Hsieh KC, Krantz A. Allenic suicide substrates. New inhibitors of vitamin B6 linked
decarboxylases. J Am Chem Soc 1984;106:2734–5.[332] Krantz A, Castelhano AL. a-Allenic a-amino acids. European Patent: EP 134561 A1 19850320. 1985. 51 pp.[333] Tokuyama T, Uenoyama K, Brown G, Daly JW, Witkop B. Allenic and acetylenic spiropiperidine alkaloids from the neotropical
frog, Dendrobates histrionicus. Helv Chim Acta 1974;57:2597–604.[334] Daly JW, Wilham JM, Spande TF, Garraffo HM, Gil RR, Silva GL, et al. Alkaloids in bufonid toads (melanophryniscus): temporal
and geographic determinants for two argentinian species. J Chem Ecol 2007;33:871–87.[335] Daly JW, Spande TF, Garraffo HM. Alkaloids from amphibian skin: a tabulation of over eight-hundred compounds. J Nat Prod
2005;68:1556–75.[336] Karle IL. The structure of dihydroisohistrionicotoxin, a unique unsaturated alkaloid and anticholinergic agent. J Am Chem Soc
1973;95:4036–40.[337] Burgermeister W, Catterall WA, Witkop B. Histrionicotoxin enhances agonist-induced desensitization of acetylcholine receptor.
Proc Natl Acad Sci USA 1977;74:5754–8.

V.M. Dembitsky, T. Maoka / Progress in Lipid Research 46 (2007) 328–375 375
[338] St Claire MB, Kennett MJ, Thomas ML, Daly JW. The husbandry and care of dendrobatid frogs. Contemp Top Lab Anim Sci2005;44:8–14.
[339] Daly JW, Garraffo HM, Spande TF, Decker MW, Sullivan JP, Williams M. Alkaloids from frog skin: the discovery of epibatidineand the potential for developing novel non-opioid analgesics. Nat Prod Rep 2000;17:131–5.
[340] Daly JW. Alkaloids from frog skins: selective probes for ion channels and nicotinic receptors. Braz J Med Biol Res 1995;28:1033–42.[341] Matsumoto S, Yamamoto S, Sai K, Maruo K, Adachi M, Saitoh M, et al. Pipecolic acid induces apoptosis in neuronal cells. Brain
Res 2003;980:179–84.[342] Davis JR, Morris RN. Isolation of intermediary metabolites of L-lysine-U-C14 from tissues of tumor-bearing rats. Cancer Res
1963;23:143–7.[343] Agami C, Bihan D, Hamon L, Kadouri-Puchot C, Lusinchi M. Enantioselective synthesis of an allenyl derivative of pipecolic acid.
Eur J Org Chem 1998;11:2461–5.[344] (a) Bohlmann F, Hopf PD. Polyacetylene compounds. 222. Biogenesis of the cumulene isolated from Anthemis austriaca. Chem Ber
1973;106:3772–4;(b) Bohlmann F, Kleine KM, Arndt C. Polyacetylene compounds. C. Natural occurring cumulene as well as various sulfurcompounds derived from matricaria ester. Justus Liebigs Ann Chem 1966;694:149–56.
[345] Bohlmann F, Zdero C. Polyacetylene compounds. 183. New cumulene from Erigeron canadense. Tetrahedron Lett 1970;28:2465–6.[346] Bohlmann F, Zdero C. Polyacetylenic compounds. CLX. Constituents of Aster bellidiastrum. Chem Ber 1969;102:1034–6.[347] Sorensen NA, Stavholt K. A hexahydro matricaria ester, Composit-Cumulene I,’’ from Matricaria inodora. Acta Chem Scand
1950;4:1080–4.[348] Christensen PK, Sorensen NA, Bell I, Jones ERH, Whiting MC. Constitution of the so-called ‘‘composite-cumulene I’’ from
scentless mayweed (Matricaria inodora). Festschr Arthur Stoll 1957:545–50.[349] Kawaguchi K, Kaifu N, Ohishi M, Ishikawa S, Hirahara Y, Yamamoto S, et al. Observations of cumulene carbenes,
butatrienylidene (H2CCCC) and propadienylidene (H2CCC), in TMC-1. Publicat Astronom Soc Jpn 1991;43:607–19.[350] Goicoechea JR, Cernicharo J, Masso H, Senent ML. A new unidentified far-infrared band in NGC 7027. Astrophys J
2004;609:225–30.[351] Langer WD, Velusamy T, Kuiper TBH, Peng R, Mccarthy MC, Travers MJ, et al. First astronomical detection of the cumulene
carbon chain molecule H2C6 in TMC-1. Astrophys J 1997;480:L63–6.[352] McCarthy MC, Travers MJ, Kovacs A, Gottlieb CA, Thaddeus P. Eight new carbon chain molecules. Astrophys J, Suppl Ser
1997;113:105–20.[353] Apponi AJ, McCarthy MC, Gottlieb CA, Thaddeus P. Laboratory detection of four new cumulene carbenes: H2C7, H2C8, H2C9,
and D2C10. Astrophys J 2000;530:357–61.[354] Parker WL, Rathnum ML, Seiner V, Trejo WH, Principe PA, Sykes RB. Cepacin A and Cepacin B, two new antibiotics produced by
Pseudomonas cepacia. J Antib 1984;37:431–40.[355] Sykes RB, Parker WL. Cepacin A and B by fermentation. 1984;12 pp. Germany Patent: DE 3417994 A1 19841122.[356] Tang C-J, Wu Y. On the synthesis of cepacin A. Tetrahedron 2007;63:4887–906.[357] Ma S, Xu D, Li Z. Preparation of optically active 2,3-allenols by enzymic resolution. Faming Zhuanli Shenqing Gongkai
Shuomingshu. 2002;16 pp. Chinese Patent: CN 1386730 A 20021225.[358] Konegawa T, Ohtsuka Y, Ikeda H, Sugai T, Ohta H. Enzyme-mediated optical resolution of 2-methyl-3,4-pentadien-1-ol, a chiral
building block possessing terminal allenyl group. Synlett 1997;11:1297–9.[359] Hems R, Horton D, Nakadate M. Extension of sugar chains through acetylenic intermediates. X. Synthesis of D-glycero-D-galacto-
heptitol (perseitol), L-glycero-D-galacto-heptitol, and an allenic sugar derivative, from acetylenic precursors. Carbohyd Res1972;25:205–16.
[360] Boukouvalas J, Pouliot M, Robichaud J, MacNeil S, Snieckus V. Asymmetric total synthesis of (�)-panacene and correction of itsrelative configuration. Org Lett 2006;8:3597–9.
[361] Brummond KM, DeForrest JE. Synthesizing allenes today (1982–2006). Synthesis 2007;6:795–818.[362] Escudie J, Ranaivonjatovo H. Group 14 and 15 Heteroallenes E:C:C and E:C:E 0. Organometallics 2007;26:1542–59.[363] Liu R. Asymmetric dihydroxylation and aziridination of allenes and related chemistry. Ph.D. Thesis. Brigham Young University,
August 2007; p. 199, Provo, Utah.
Copyright © 2022 FDOKUMEN




















