
FORMULATION AND EVALUATION OF CONTROLLED RELEASE TABLETS OF PIOGLITAZONE
Upload
centralunipunjabCategory
view
5download
0
6) 2209–2216www.elsevier.com/locate/lifescie
Life Sciences 79 (200
Chronic administration of pioglitazone attenuates intracerebroventricularstreptozotocin induced-memory impairment in rats
Asif R. Pathan, Bhoomi Viswanad, Swapnil K. Sonkusare, Poduri Ramarao ⁎
Department of Pharmacology and Toxicology, National Institute of Pharmaceutical Education and Research (NIPER), S.A.S. Nagar-160062, Punjab, India
Received 9 December 2005; accepted 17 July 2006
Abstract
Memory impairment induced by intracerebroventricular (ICV) injection of streptozotocin (STZ) in rats is associated with impaired brainglucose and energy metabolism, oxidative stress and impaired cholinergic neurotransmission. Treatment with antioxidants and cholinergicagonists has been reported to produce beneficial effect in this model. However, no reports are available on drugs that improve glucose utilizationand metabolism. In the present study, we evaluated the effects of pioglitazone on cognitive performance, oxidative stress and glucose utilization inICV STZ injected rats (3 mg/kg, on day 1 and 3). Pioglitazone (10 and 30 mg/kg) was administered per oral (p.o.) for 14 days, starting 5 days priorto STZ injection. Cognitive performance was assessed using step-through passive avoidance and Morris water maze task. Malondialdehyde(MDA) and glutathione levels in brain were estimated as parameters of oxidative stress. Glucose utilization by brain was assessed as the amount ofglucose consumed from the media by the brain. ICV STZ injected rats showed a severe deficit in learning and memory associated with increasedMDA levels (+67.5%), decreased glutathione levels (−29.2%) and impaired cerebral glucose utilization (−44.4%). In contrast pioglitazonetreatment improved cognitive performance, lowered oxidative stress and improved cerebral glucose utilization in ICV STZ rats. The present studydemonstrates the beneficial effects of pioglitazone in the ICV STZ induced cognitive deficits, which can be exploited for the dementia associatedwith diabetes and age-related neurodegenerative disorder, where oxidative stress and impaired glucose and energy metabolism are involved.© 2006 Published by Elsevier Inc.
Keywords: Intracerebroventricular streptozotocin; Learning and memory; Pioglitazone; Oxidative stress and glucose utilization
Introduction
Impaired cerebral glucose utilization and energy metabolismrepresent very early abnormalities in initial stages of cognitiveimpairment (Cao et al., 2003). Sporadic dementia of Alzheimer'stype (SDAT) in particular, is characterized by a progressive dete-rioration of both cognitive function and energy metabolism (Hoyeret al., 1991). The reduction in cerebral glucose utilization, rangingfrom10% inmild tomore than 40% in severe cases of dementia hasbeen reported (Kumar et al., 1991). Besides diminished glucoseutilization, insulin receptor signal transduction cascade is severelyimpaired in hippocampus and hypothalamus of AD (Alzheimer'sdisease) brain (Steen et al., 2005). Another major feature of SDATpathophysiology is oxidative damage. Oxidative stress can affectall classes of macromolecules (sugar, lipids, proteins, and DNA),leading inevitably to neuronal dysfunction (Polidori and Mecocci,
⁎ Corresponding author. Tel.: +91 172 2214697; fax: +91 172 2214692.E-mail address: [email protected] (P. Ramarao).
0024-3205/$ - see front matter © 2006 Published by Elsevier Inc.doi:10.1016/j.lfs.2006.07.018
2002). Moreover, impaired insulin signaling has already beenlinked to increased oxidative stress and mitochondrial dysfunctionin neuronal cells (Hoyer and Lannert, 1999). These pathologicalhallmarks of SDAT are very similar to type 2 diabetes (NIDDM),therefore SDAT is considered as NIDDM of the brain (Hoyer,2004). Thus, treatment with ligands that improve glucoseutilization and insulin signaling and decrease oxidative stressmay help to improve viability and function of neuronal cells at therisk of AD type neurodegeneration.
The Intracerebroventricular streptozotocin (ICV STZ) injectedrat has been described as an appropriate animal model for SDATcharacterized by progressive deterioration of memory and cere-bral glucose and energy metabolism, along with oxidative stress(Nitsch and Hoyer, 1991; Lannert and Hoyer, 1998; Sharma andGupta, 2001; Sonkusare et al., 2005). Activities of glycolyticenzyme are reduced (Plaschke and Hoyer, 1993), leading toreduced formation of acetyl CoA and thereof acetylcholine(Blokland and Jolles, 1994; Weinstock et al., 2001). Preliminaryreports suggest the desensitization of neuronal insulin receptor as

2210 A.R. Pathan et al. / Life Sciences 79 (2006) 2209–2216
the cause of impaired cerebral glucose and energy metabolism inthis animal model (Hoyer and Lannert, 1999).
Treatment with antioxidants and drugs that facilitate choliner-gic neurotransmission has been reported to have beneficial effectsin this model (Sharma and Gupta, 2002; Sonkusare et al., 2005;Weinstock et al., 2001). However, no reports are available on thedrugs that improve glucose utilization and metabolism leading toimprovement in cognitive performance in rats. The present study,therefore, was designed to evaluate the effect of pioglitazone (aninsulin sensitizer) on learning and memory, oxidative stress andcerebral glucose utilization in ICV STZ injected rats.
Materials and methods
Animals
Adult male Wistar rats weighing 320–350 g were procuredfrom Central Animal Facility (CAF), National Institute ofPharmaceutical Education and Research (NIPER). Animalswere housed three per cage prior to STZ administration and oneper cage after STZ administration. The animals were fed withnormal pellet diet (PranawAgro Industries, NewDelhi) andwaterad libitum and maintained in standard environmental conditions(temperature 22±2 °C, humidity 55±5% and 12 h light/darkcycle). All animal protocols of the experiment were approved byInstitutional Animal Ethics Committee (IAEC), NIPER andperformed in accordance with the guidelines of Committee forControl and Supervision of Experimentation on Animals(CPCSEA), Government of India on animal experimentation.
Intracerebroventricular administration of streptozotocin
ICV injection of STZ was performed as described previously(Sonkusare et al., 2005). Briefly, adult male Wistar rats weighing320–350 gwere anesthetisedwith thiopentone (NeonLaboratories,India, 45 mg/kg, i.p.). The head was placed in position in thestereotaxic apparatus and a midline sagittal incision was made inthe scalp. Following coordinates were used for ICV injection:0.8 mm posterior to bregma, 1.5 mm lateral to saggital suture and3.6 mm ventral from the surface of the brain. STZ (Sigma, USA)was dissolved in citrate buffer (pH 4.4) just prior to injection. TheSTZ group was injected bilaterally with ICV STZ (3mg/kg) in twodivided doses, on days 1 and 3. The concentration of STZ in citratebuffer was adjusted so as to deliver 10 μl of the solution. Rats insham operated group were given ICV injection of the same volumeof citrate buffer on the first and third day as in STZ injected rat.After ICV injection, povidone–iodine solution was applied and thecut skin was sutured after second injection followed by dailyapplication of antiseptic powder (Neosporin®). Post operatively,the rats were fedwithmilk by oral gavage and normal pellet diet for4 days, followed by normal pellet diet alone. The body weightswere monitored throughout the study period.
Animal groups and drug administration
The animals were randomly divided into four experimentalgroups with 12 animals in each viz. ICV citrate buffer injected
(control), vehicle treated ICVSTZ rats, pioglitazone (10mg/kg) andpioglitazone (30mg/kg) treated ICVSTZ rats.After behavioral tasks6 animals from each groupwere sacrificed and used for estimationof oxidative stress parameters. Remaining 6 animals in each groupwere used for brain glucose utilization study. Pioglitazone HCl(gifted by Ranbaxy Laboratories Ltd, India, 10 and 30mg/kg/day,p.o.) treatment was started 5 days prior to the first ICV STZinjection and continued for 14 days. The suspension of drug wasprepared in 1% carboxymethyl cellulose (CMC) and administeredorally by gavage. The control group received 1% CMC orally.
Behavioral tests
Step-through passive avoidance taskStep-through passive avoidance task was used on days 14 and
15 to evaluatememory retention deficit. The apparatus (UgoBasile,Type-7552, Italy) consisted of a chamber illuminated with a 40 Wbulb and a dark chamber, separated by a guillotine door. The floorwas made of a metal grid with a shock scrambler. During acqui-sition trial, rat was placed in the illuminated chamber. After initialhabituation period of 60 s, the guillotine door was opened and thetime taken by the rat to enter the dark chamber was noted. As soonas the rat entered the dark chamber, it was given amild footshock of0.5mA for 2 s through the grid floor. The rat was allowed to remainin the dark compartment for 5 s and thenwas taken out. The latencyto step into the dark compartmentwas recorded as initial trial or pre-shock latency (ITL). Rats showing ITL of more than 60 s wereexcluded from further experiments. 24 h later, retention trial wasperformed and retention trial or post-shock latency (RTL) to stepinto the dark compartment was noted. Cut-off time was fixed at300 s (Sharma and Gupta, 2001; Sonkusare et al., 2005).
Morris water maze testMorris water maze (MWM) procedure, originally described by
Morris, ismost commonly used to test spatialmemory.Watermazeconsists of a large circular black pool (150 cm in diameter, 40 cm inheight, filled to a depth of 30 cmwith water at 28±1 °C), placed ina darkened room illuminated by a sparse light. A round, submergedplatform, painted black, is placed inside this pool, 2 cm below thesurface of water. Rat climbs this platform to escape the necessity ofswimming. Region within 15 cm from border was called as borderzone, whereas inner regionwas called central zone that was furtherdivided into four quadrants. Acquisition trials were performedkeeping the platform in different quadrants.On 5 consecutive days,rats were given 2 acquisition trials per day with an inter-trial in-terval of 15 min. Rats were allowed a maximum of 120 s to locatethe platform and permitted to stay on it for amaximumof 30 s. Ratsthat failed to locate the platformwere placed on the platform by theexperimenter for a maximum of 30 s. Learning was assisted withextra maze cues and two parameters were noted in each expe-riment: escape latency (time taken to find the hidden platform) andtime spent in the border zone. All the trials were completedbetween 10.00 to 16.00 h (Manschot et al., 2003).
Closed field activityClosed field activity was measured to rule out the interference
of change in locomotor activity in the parameters of learning and
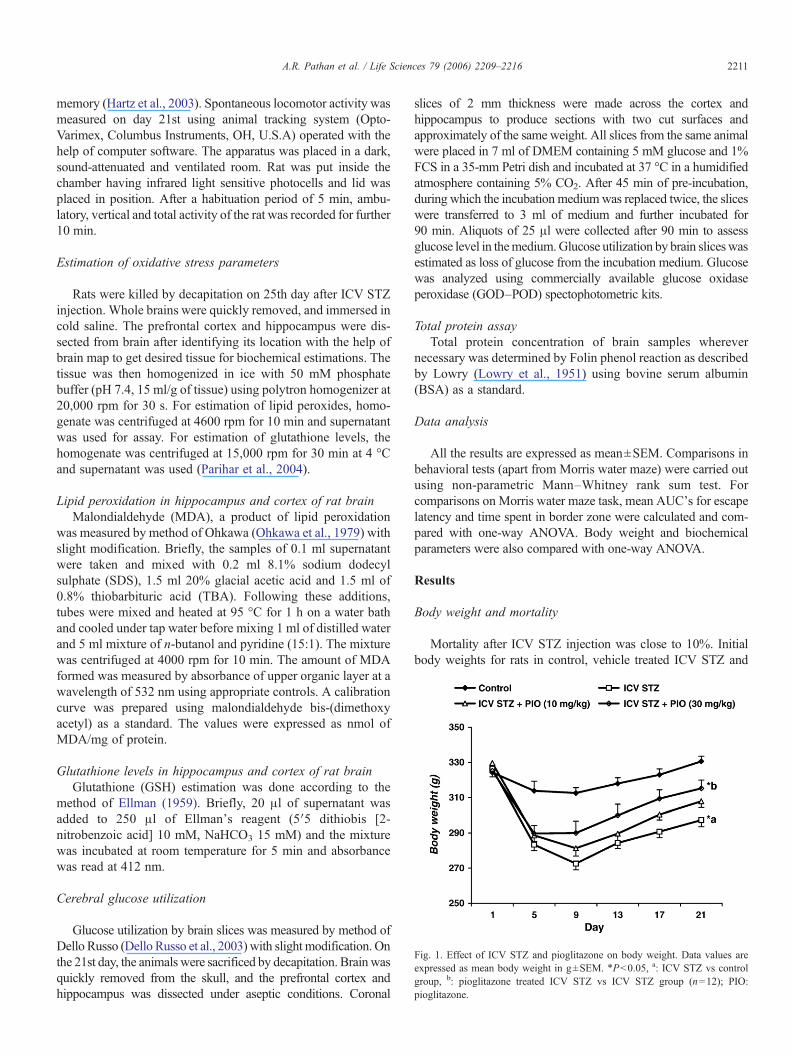
Fig. 1. Effect of ICV STZ and pioglitazone on body weight. Data values areexpressed as mean body weight in g±SEM. ⁎Pb0.05, a: ICV STZ vs controlgroup, b: pioglitazone treated ICV STZ vs ICV STZ group (n=12); PIO:pioglitazone.
2211A.R. Pathan et al. / Life Sciences 79 (2006) 2209–2216
memory (Hartz et al., 2003). Spontaneous locomotor activity wasmeasured on day 21st using animal tracking system (Opto-Varimex, Columbus Instruments, OH, U.S.A) operated with thehelp of computer software. The apparatus was placed in a dark,sound-attenuated and ventilated room. Rat was put inside thechamber having infrared light sensitive photocells and lid wasplaced in position. After a habituation period of 5 min, ambu-latory, vertical and total activity of the rat was recorded for further10 min.
Estimation of oxidative stress parameters
Rats were killed by decapitation on 25th day after ICV STZinjection. Whole brains were quickly removed, and immersed incold saline. The prefrontal cortex and hippocampus were dis-sected from brain after identifying its location with the help ofbrain map to get desired tissue for biochemical estimations. Thetissue was then homogenized in ice with 50 mM phosphatebuffer (pH 7.4, 15 ml/g of tissue) using polytron homogenizer at20,000 rpm for 30 s. For estimation of lipid peroxides, homo-genate was centrifuged at 4600 rpm for 10 min and supernatantwas used for assay. For estimation of glutathione levels, thehomogenate was centrifuged at 15,000 rpm for 30 min at 4 °Cand supernatant was used (Parihar et al., 2004).
Lipid peroxidation in hippocampus and cortex of rat brainMalondialdehyde (MDA), a product of lipid peroxidation
was measured by method of Ohkawa (Ohkawa et al., 1979) withslight modification. Briefly, the samples of 0.1 ml supernatantwere taken and mixed with 0.2 ml 8.1% sodium dodecylsulphate (SDS), 1.5 ml 20% glacial acetic acid and 1.5 ml of0.8% thiobarbituric acid (TBA). Following these additions,tubes were mixed and heated at 95 °C for 1 h on a water bathand cooled under tap water before mixing 1 ml of distilled waterand 5 ml mixture of n-butanol and pyridine (15:1). The mixturewas centrifuged at 4000 rpm for 10 min. The amount of MDAformed was measured by absorbance of upper organic layer at awavelength of 532 nm using appropriate controls. A calibrationcurve was prepared using malondialdehyde bis-(dimethoxyacetyl) as a standard. The values were expressed as nmol ofMDA/mg of protein.
Glutathione levels in hippocampus and cortex of rat brainGlutathione (GSH) estimation was done according to the
method of Ellman (1959). Briefly, 20 μl of supernatant wasadded to 250 μl of Ellman's reagent (5′5 dithiobis [2-nitrobenzoic acid] 10 mM, NaHCO3 15 mM) and the mixturewas incubated at room temperature for 5 min and absorbancewas read at 412 nm.
Cerebral glucose utilization
Glucose utilization by brain slices was measured by method ofDello Russo (Dello Russo et al., 2003) with slight modification. Onthe 21st day, the animals were sacrificed by decapitation. Brain wasquickly removed from the skull, and the prefrontal cortex andhippocampus was dissected under aseptic conditions. Coronal
slices of 2 mm thickness were made across the cortex andhippocampus to produce sections with two cut surfaces andapproximately of the same weight. All slices from the same animalwere placed in 7 ml of DMEM containing 5 mM glucose and 1%FCS in a 35-mm Petri dish and incubated at 37 °C in a humidifiedatmosphere containing 5% CO2. After 45 min of pre-incubation,during which the incubation mediumwas replaced twice, the sliceswere transferred to 3 ml of medium and further incubated for90 min. Aliquots of 25 μl were collected after 90 min to assessglucose level in themedium.Glucose utilization by brain sliceswasestimated as loss of glucose from the incubation medium. Glucosewas analyzed using commercially available glucose oxidaseperoxidase (GOD–POD) spectophotometric kits.
Total protein assayTotal protein concentration of brain samples wherever
necessary was determined by Folin phenol reaction as describedby Lowry (Lowry et al., 1951) using bovine serum albumin(BSA) as a standard.
Data analysis
All the results are expressed as mean±SEM. Comparisons inbehavioral tests (apart from Morris water maze) were carried outusing non-parametric Mann–Whitney rank sum test. Forcomparisons on Morris water maze task, mean AUC's for escapelatency and time spent in border zone were calculated and com-pared with one-way ANOVA. Body weight and biochemicalparameters were also compared with one-way ANOVA.
Results
Body weight and mortality
Mortality after ICV STZ injection was close to 10%. Initialbody weights for rats in control, vehicle treated ICV STZ and
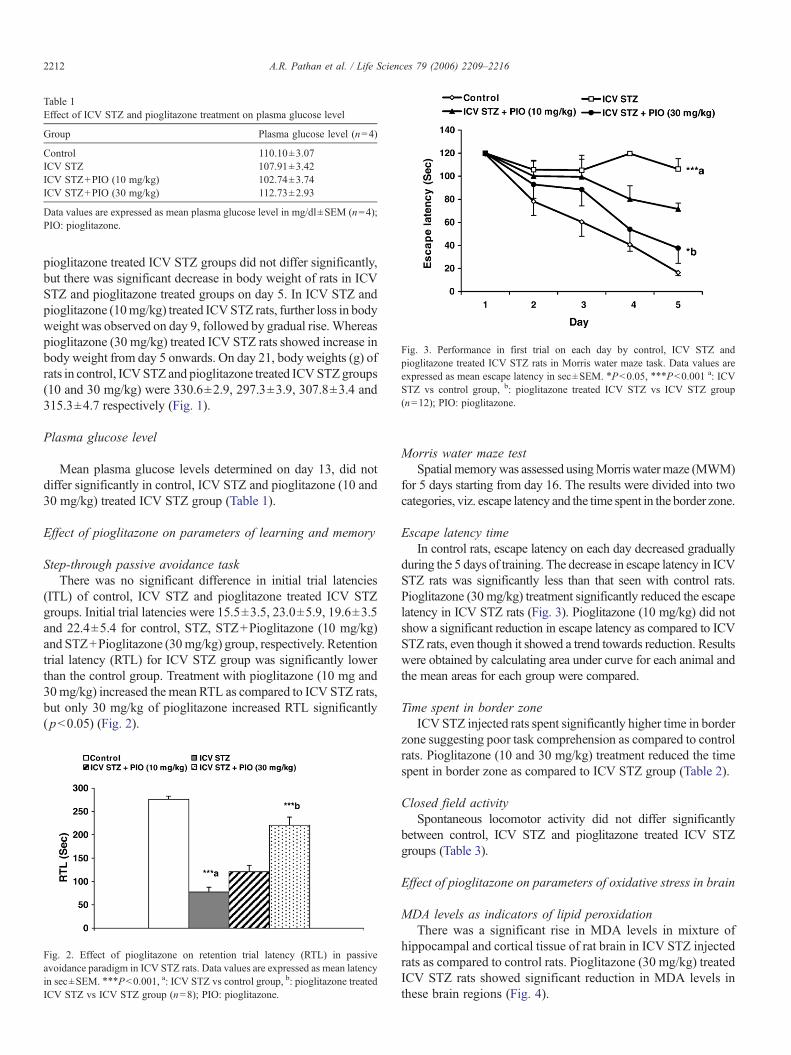
Table 1Effect of ICV STZ and pioglitazone treatment on plasma glucose level
Group Plasma glucose level (n=4)
Control 110.10±3.07ICV STZ 107.91±3.42ICV STZ+PIO (10 mg/kg) 102.74±3.74ICV STZ+PIO (30 mg/kg) 112.73±2.93
Data values are expressed as mean plasma glucose level in mg/dl±SEM (n=4);PIO: pioglitazone.
Fig. 3. Performance in first trial on each day by control, ICV STZ andpioglitazone treated ICV STZ rats in Morris water maze task. Data values areexpressed as mean escape latency in sec±SEM. ⁎Pb0.05, ⁎⁎⁎Pb0.001 a: ICVSTZ vs control group, b: pioglitazone treated ICV STZ vs ICV STZ group(n=12); PIO: pioglitazone.
2212 A.R. Pathan et al. / Life Sciences 79 (2006) 2209–2216
pioglitazone treated ICV STZ groups did not differ significantly,but there was significant decrease in body weight of rats in ICVSTZ and pioglitazone treated groups on day 5. In ICV STZ andpioglitazone (10mg/kg) treated ICVSTZ rats, further loss in bodyweight was observed on day 9, followed by gradual rise. Whereaspioglitazone (30 mg/kg) treated ICV STZ rats showed increase inbody weight from day 5 onwards. On day 21, body weights (g) ofrats in control, ICVSTZ and pioglitazone treated ICVSTZgroups(10 and 30 mg/kg) were 330.6±2.9, 297.3±3.9, 307.8±3.4 and315.3±4.7 respectively (Fig. 1).
Plasma glucose level
Mean plasma glucose levels determined on day 13, did notdiffer significantly in control, ICV STZ and pioglitazone (10 and30 mg/kg) treated ICV STZ group (Table 1).
Effect of pioglitazone on parameters of learning and memory
Step-through passive avoidance taskThere was no significant difference in initial trial latencies
(ITL) of control, ICV STZ and pioglitazone treated ICV STZgroups. Initial trial latencies were 15.5±3.5, 23.0±5.9, 19.6±3.5and 22.4±5.4 for control, STZ, STZ+Pioglitazone (10 mg/kg)and STZ+Pioglitazone (30mg/kg) group, respectively. Retentiontrial latency (RTL) for ICV STZ group was significantly lowerthan the control group. Treatment with pioglitazone (10 mg and30mg/kg) increased the mean RTL as compared to ICV STZ rats,but only 30 mg/kg of pioglitazone increased RTL significantly(pb0.05) (Fig. 2).
Fig. 2. Effect of pioglitazone on retention trial latency (RTL) in passiveavoidance paradigm in ICV STZ rats. Data values are expressed as mean latencyin sec±SEM. ⁎⁎⁎Pb0.001, a: ICV STZ vs control group, b: pioglitazone treatedICV STZ vs ICV STZ group (n=8); PIO: pioglitazone.
Morris water maze testSpatial memorywas assessed usingMorris watermaze (MWM)
for 5 days starting from day 16. The results were divided into twocategories, viz. escape latency and the time spent in the border zone.
Escape latency timeIn control rats, escape latency on each day decreased gradually
during the 5 days of training. The decrease in escape latency in ICVSTZ rats was significantly less than that seen with control rats.Pioglitazone (30 mg/kg) treatment significantly reduced the escapelatency in ICV STZ rats (Fig. 3). Pioglitazone (10 mg/kg) did notshow a significant reduction in escape latency as compared to ICVSTZ rats, even though it showed a trend towards reduction. Resultswere obtained by calculating area under curve for each animal andthe mean areas for each group were compared.
Time spent in border zoneICV STZ injected rats spent significantly higher time in border
zone suggesting poor task comprehension as compared to controlrats. Pioglitazone (10 and 30 mg/kg) treatment reduced the timespent in border zone as compared to ICV STZ group (Table 2).
Closed field activitySpontaneous locomotor activity did not differ significantly
between control, ICV STZ and pioglitazone treated ICV STZgroups (Table 3).
Effect of pioglitazone on parameters of oxidative stress in brain
MDA levels as indicators of lipid peroxidationThere was a significant rise in MDA levels in mixture of
hippocampal and cortical tissue of rat brain in ICV STZ injectedrats as compared to control rats. Pioglitazone (30 mg/kg) treatedICV STZ rats showed significant reduction in MDA levels inthese brain regions (Fig. 4).
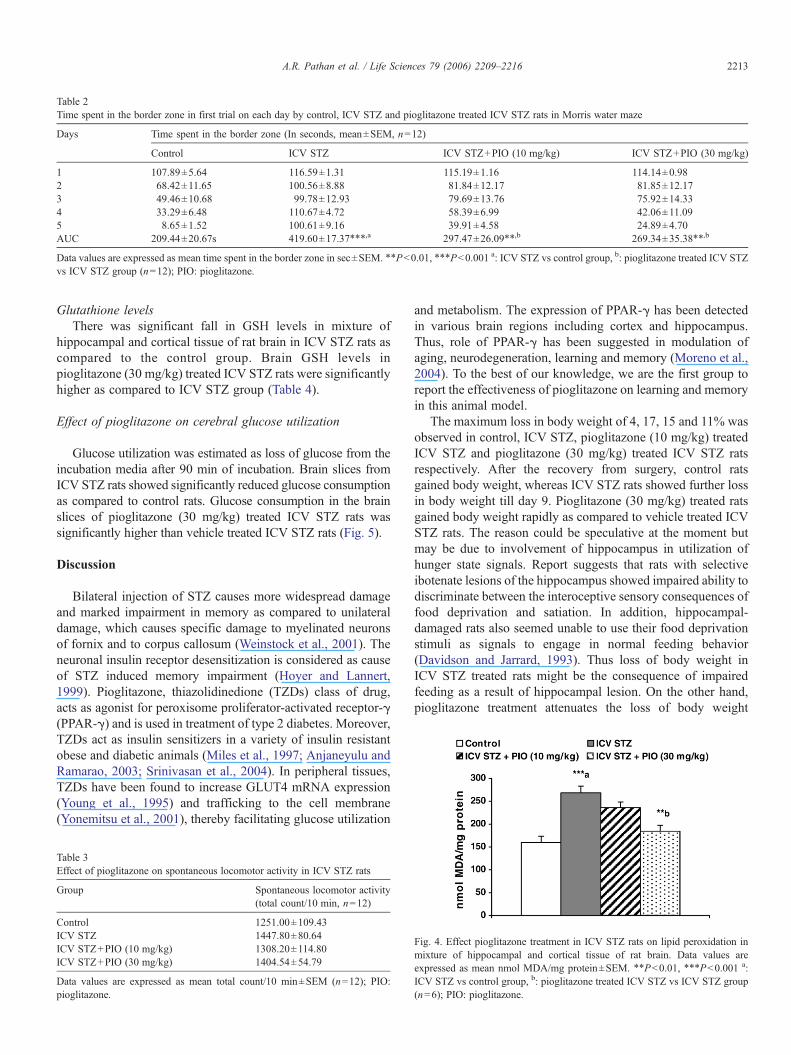
Table 2Time spent in the border zone in first trial on each day by control, ICV STZ and pioglitazone treated ICV STZ rats in Morris water maze
Days Time spent in the border zone (In seconds, mean±SEM, n=12)
Control ICV STZ ICV STZ+PIO (10 mg/kg) ICV STZ+PIO (30 mg/kg)
1 107.89±5.64 116.59±1.31 115.19±1.16 114.14±0.982 68.42±11.65 100.56±8.88 81.84±12.17 81.85±12.173 49.46±10.68 99.78±12.93 79.69±13.76 75.92±14.334 33.29±6.48 110.67±4.72 58.39±6.99 42.06±11.095 8.65±1.52 100.61±9.16 39.91±4.58 24.89±4.70AUC 209.44±20.67s 419.60±17.37⁎⁎⁎,a 297.47±26.09⁎⁎,b 269.34±35.38⁎⁎,b
Data values are expressed as mean time spent in the border zone in sec±SEM. ⁎⁎Pb0.01, ⁎⁎⁎Pb0.001 a: ICV STZ vs control group, b: pioglitazone treated ICV STZvs ICV STZ group (n=12); PIO: pioglitazone.
2213A.R. Pathan et al. / Life Sciences 79 (2006) 2209–2216
Glutathione levelsThere was significant fall in GSH levels in mixture of
hippocampal and cortical tissue of rat brain in ICV STZ rats ascompared to the control group. Brain GSH levels inpioglitazone (30 mg/kg) treated ICV STZ rats were significantlyhigher as compared to ICV STZ group (Table 4).
Effect of pioglitazone on cerebral glucose utilization
Glucose utilization was estimated as loss of glucose from theincubation media after 90 min of incubation. Brain slices fromICV STZ rats showed significantly reduced glucose consumptionas compared to control rats. Glucose consumption in the brainslices of pioglitazone (30 mg/kg) treated ICV STZ rats wassignificantly higher than vehicle treated ICV STZ rats (Fig. 5).
Discussion
Bilateral injection of STZ causes more widespread damageand marked impairment in memory as compared to unilateraldamage, which causes specific damage to myelinated neuronsof fornix and to corpus callosum (Weinstock et al., 2001). Theneuronal insulin receptor desensitization is considered as causeof STZ induced memory impairment (Hoyer and Lannert,1999). Pioglitazone, thiazolidinedione (TZDs) class of drug,acts as agonist for peroxisome proliferator-activated receptor-γ(PPAR-γ) and is used in treatment of type 2 diabetes. Moreover,TZDs act as insulin sensitizers in a variety of insulin resistantobese and diabetic animals (Miles et al., 1997; Anjaneyulu andRamarao, 2003; Srinivasan et al., 2004). In peripheral tissues,TZDs have been found to increase GLUT4 mRNA expression(Young et al., 1995) and trafficking to the cell membrane(Yonemitsu et al., 2001), thereby facilitating glucose utilization
Table 3Effect of pioglitazone on spontaneous locomotor activity in ICV STZ rats
Group Spontaneous locomotor activity(total count/10 min, n=12)
Control 1251.00±109.43ICV STZ 1447.80±80.64ICV STZ+PIO (10 mg/kg) 1308.20±114.80ICV STZ+PIO (30 mg/kg) 1404.54±54.79
Data values are expressed as mean total count/10 min±SEM (n=12); PIO:pioglitazone.
and metabolism. The expression of PPAR-γ has been detectedin various brain regions including cortex and hippocampus.Thus, role of PPAR-γ has been suggested in modulation ofaging, neurodegeneration, learning and memory (Moreno et al.,2004). To the best of our knowledge, we are the first group toreport the effectiveness of pioglitazone on learning and memoryin this animal model.
The maximum loss in body weight of 4, 17, 15 and 11% wasobserved in control, ICV STZ, pioglitazone (10 mg/kg) treatedICV STZ and pioglitazone (30 mg/kg) treated ICV STZ ratsrespectively. After the recovery from surgery, control ratsgained body weight, whereas ICV STZ rats showed further lossin body weight till day 9. Pioglitazone (30 mg/kg) treated ratsgained body weight rapidly as compared to vehicle treated ICVSTZ rats. The reason could be speculative at the moment butmay be due to involvement of hippocampus in utilization ofhunger state signals. Report suggests that rats with selectiveibotenate lesions of the hippocampus showed impaired ability todiscriminate between the interoceptive sensory consequences offood deprivation and satiation. In addition, hippocampal-damaged rats also seemed unable to use their food deprivationstimuli as signals to engage in normal feeding behavior(Davidson and Jarrard, 1993). Thus loss of body weight inICV STZ treated rats might be the consequence of impairedfeeding as a result of hippocampal lesion. On the other hand,pioglitazone treatment attenuates the loss of body weight
Fig. 4. Effect pioglitazone treatment in ICV STZ rats on lipid peroxidation inmixture of hippocampal and cortical tissue of rat brain. Data values areexpressed as mean nmol MDA/mg protein±SEM. ⁎⁎Pb0.01, ⁎⁎⁎Pb0.001 a:ICV STZ vs control group, b: pioglitazone treated ICV STZ vs ICV STZ group(n=6); PIO: pioglitazone.

Table 4Effect of pioglitazone treatment in ICV STZ rats on glutathione levels in mixtureof hippocampal and cortical tissue of rat brain
Group Glutathione levels (nmol GSH/mg protein, n=6)
Control 46.07±1.52ICV STZ 32.60±1.41⁎⁎⁎,a
ICV STZ+PIO (10 mg/kg) 34.79±1.22ICV STZ+PIO (30 mg/kg) 41.38±1.63⁎⁎,b
Data values are expressed as mean nmol of GSH/mg protein±SEM. ⁎⁎Pb0.01,⁎⁎⁎Pb0.001 a: ICV STZ vs control group, b: pioglitazone treated ICV STZ vsICV STZ group (n=6); PIO: pioglitazone.
Fig. 5. Effect of ICV STZ injection and pioglitazone treatment in ICV STZ ratson glucose utilization by slices of hippocampal and cortical tissue of rat brainafter 90 min of incubation in media. Data values are expressed as mean % ofglucose utilization±SEM. ⁎⁎Pb0.01, ⁎⁎⁎Pb0.001, a: ICV STZ vs controlgroup, b: pioglitazone treated ICV STZ vs ICV STZ group (n=6); PIO:pioglitazone.
2214 A.R. Pathan et al. / Life Sciences 79 (2006) 2209–2216
suggesting its neuroprotective action. There was no significantdifference in plasma glucose level and spontaneous locomotoractivity among the four groups (Tables 1 and 3), which rules outthe interference by changes in blood glucose and locomotoractivity by pioglitazone and/or STZ in performance in the testsof learning and memory. In step-through passive avoidance,higher dose of pioglitazone (30 mg/kg) increased the retentiontrial latency in ICV STZ rats, signifying improvementacquisition and retention of memory (Fig. 2). In Morris watermaze task, ICV STZ rats treated with 30 mg/kg pioglitazoneshowed shorter escape latencies to reach the platform. Also,time spent in the border zone was significantly less than ICVSTZ rats (Fig. 3 and Table 2). The MWM procedure is based ona reward principle and provides measure of spatial memory. Asanimal dislike swimming, their tendency is to escape from thewater being accomplished by finding an escape platform. Timespent in border zone of the pool provides a measure of taskcomprehension. Trained rats try to escape by more centrallyoriented platform and consequently spent less time in borderzone whereas untrained rats try to escape by the border of pool.Shorter escape latencies and time spent in border zone by30 mg/kg pioglitazone treated rats indicates improvement ofspatial memory by pioglitazone treatment in ICV STZ rats. Itshould be noted that pioglitazone at the lower dose used in thisstudy (10 mg/kg), showed no significant effects on the per-formance in the tasks of learning and memory, although it didshow a trend towards improvement.
Because of their involvement in behavioral and cognitivetasks, prefrontal cortex and hippocampus were used for the bio-chemical estimations. We observed a reduction in MDA levelswith pioglitazone treatment (30 mg/kg) as compared to vehicletreated ICV STZ rats (Fig. 4). In addition, glutathione levels inpioglitazone treated rats were significantly higher than vehicletreated ICV STZ rats (Table 4). MDA, an end product of lipidperoxidation is a measure of oxidative damage by free radicals(Polidori and Mecocci, 2002) whereas glutathione is a tripeptide,an endogenous antioxidant found in all animal cells in variableamounts (Schuessel et al., 2004). An earlier report has shownincrease in MDA levels and decrease in levels of glutathione inICV STZ rats (Sharma and Gupta, 2001). However, it is stillunclear whether STZ induced oxidative stress is a result of directneuronal damage by STZ or a consequence of impaired glucoseand energy metabolism. STZ is known to produce intracellularfree radicals, NO and H2O2, both of which may constitute directcomponent of STZ-induced neuronal damage (Prickaerts et al.,
2000). On the other hand, STZ inhibits tyrosine kinase activity ofinsulin receptors by releasing NO intracellularly, which leads toS-nitrosylation of receptor tyrosine kinase (Simon et al., 1996).Plaschke and Hoyer (1993) reported increased extracellular con-centration of glucose in brain of ICV STZ injected rats. This maylead to increased nonenzymatic glycosylation of proteins andglucose auto-oxidation, resulting into formation of advancedglycation end-products (AGE), which in turn causes activation ofreceptors for advanced glycation end-products (RAGE). Thissequence of events results into oxidative stress and cellulardamage (Ott et al., 1999). Therefore at this juncture, it is difficultto establish whether the protective effect of pioglitazone againstoxidative stress is primarily as an antioxidant or secondary to itsinsulin sensitizing action on glucose metabolism. Several otherstudies have reported antioxidant effect of pioglitazone that ac-companies its beneficial effect in diabetes (Dobrian et al., 2004;Gumieniczek, 2003, 2005; Ishida et al., 2004).
Glucose utilization by brain slices from ICV STZ rats wassignificantly less than the control group, whereas pioglitazonetreatment (30 mg/kg) increased glucose utilization by brain slicesin ICV STZ rats (Fig. 5). Some studies have reported reducedglycolytic enzyme activities such as hexokinase and phospho-fructokinase in cerebral cortex and hippocampus after ICV STZadministration (Plaschke and Hoyer, 1993). In contrast, ICVinsulin increases the activities of these enzymes (Hoyer et al.,1993). Phillips and Coxon (1976) reported increased glucoseutilization by brain slices from young rats after insulin admi-nistration. Transfection of Chinese hamster ovary cells with theexpression vector containing rabbit brain HepG2-type glucosetransporter cDNA respond to insulin application with more than40% increase in deoxyglucose uptake (Asano et al., 1989). Insulinreportedly stimulated glucose uptake in neonatal glial cells fromrat brain (Clarke et al., 1984). Thus, insulin dysfunction has beensuggested to be responsible for impaired glucose uptake byneuronal cells in ICV STZ rats. Our findings are consistent withprevious reports suggesting impaired glucose uptake and uti-lization in brain after ICV STZ injection (Duelli et al., 1994).Increase in glucose utilization with pioglitazone treatment couldbe an insulin sensitizing and a favorable action of pioglitazone on

2215A.R. Pathan et al. / Life Sciences 79 (2006) 2209–2216
glucose metabolism in the brain. However, we believe that mole-cular mechanism needs to be studied to explore this possibility.
Conclusions
Our results suggest impaired glucose utilization and oxidativestress as the causes of memory impairment in ICV STZ rats.Pioglitazone exerts its beneficial effect on memory processing,which may be attributed to its favorable effect on glucose utili-zation and metabolism and antioxidant action. The study demon-strates the beneficial effect of pioglitazone in ICV STZ inducedcognitive deficits however further studies are required to demon-strate the potential use of pioglitazone in dementia-associateddiabetes and age-related neurodegenerative disorders, whereimpaired glucose utilization and metabolism and oxidative stressare involved.
Acknowledgement
The authors acknowledge the gift samples provided byRanbaxy Research Laboratories Ltd, India.
References
Anjaneyulu, M., Ramarao, P., 2003. Protective effect of pioglitazone againstmultiple low-dose streptozotocin-induced diabetes in rats. Methods andFindings in Experimental and Clinical Pharmacology 25 (3), 205–208.
Asano, T., Shibasaki, Y., Ohno, S., Taira, H., Lin, J.L., Kasuga, M., Kanazawa,Y., Akanuma, Y., Takaku, F., Oka, Y., 1989. Rabbit brain glucose transporterresponds to insulin when expressed in insulin-sensitive Chinese hamsterovary cells. Journal of Biological Chemistry 264 (6), 3416–3420.
Blokland, A., Jolles, J., 1994. Behavioral and biochemical effects of an ICVinjection of streptozotocin in old Lewis rats. Pharmacology, Biochemistryand Behavior 47 (4), 833–837.
Cao, Q., Jiang, K., Zhang, M., Liu, Y., Xiao, S., Zuo, C., Huang, H., 2003. Brainglucose metabolism and neuropsychological test in patients with mildcognitive impairment. Chinese Medical Journal (Engl) 116 (8), 1235–1238.
Clarke, D.W., Boyd Jr., F.T., Kappy, M.S., Raizada, M.K., 1984. Insulin binds tospecific receptors and stimulates 2-deoxy-D-glucose uptake in cultured glialcells from rat brain. Journal of Biological Chemistry 259 (19), 11672–11675.
Davidson, T.L., Jarrard, L.E., 1993. A role for hippocampus in the utilization ofhunger signals. Behavioral and Neural Biology 59 (2), 167–171.
Dello Russo, C., Gavrilyuk, V., Weinberg, G., Almeida, A., Bolanos, J.P., Palmer, J.,Pelligrino, D., Galea, E., Feinstein, D.L., 2003. Peroxisome proliferator-activatedreceptor gamma thiazolidinedione agonists increase glucose metabolism inastrocytes. Journal of Biological Chemistry 278 (8), 5828–5836.
Dobrian, A.D., Schriver, S.D., Khraibi, A.A., Prewitt, R.L., 2004. Pioglitazoneprevents hypertension and reduces oxidative stress in diet-induced obesity.Hypertension 43 (1), 48–56.
Duelli, R., Schrock, H., Kuschinsky, W., Hoyer, S., 1994. Intracerebroven-tricular injection of streptozotocin induces discrete local changes in cerebralglucose utilization in rats. International Journal of DevelopmentalNeuroscience 12 (8), 737–743.
Ellman, G.L., 1959. Tissue sulfhydryl groups. Archives of Biochemistry andBiophysics 82 (1), 70–77.
Gumieniczek, A., 2003. Effect of the new thiazolidinedione–pioglitazone on thedevelopment of oxidative stress in liver and kidney of diabetic rabbits. LifeSciences 74 (5), 553–562.
Gumieniczek, A., 2005. Effects of pioglitazone on hyperglycemia-inducedalterations in antioxidative system in tissues of alloxan-treated diabeticanimals. Experimental and Toxicologic Pathology 56 (4–5), 321–326.
Hartz, B.P., Sohoel, A., Berezin, V., Bock, E., Scheel-Kruger, J., 2003. Asynthetic peptide ligand of NCAM affects exploratory behavior and
memory in rodents. Pharmacology, Biochemistry and Behavior 75 (4),861–867.
Hoyer, S., 2004. Glucose metabolism and insulin receptor signal transduction inAlzheimer disease. European Journal of Pharmacology 490 (1–3), 115–125.
Hoyer, S., Lannert, H., 1999. Inhibition of the neuronal insulin receptor causesAlzheimer-like disturbances in oxidative/energy brain metabolism and inbehavior in adult rats. Annals of the New York Academy of Sciences 893,301–303.
Hoyer, S., Nitsch, R., Oesterreich, K., 1991. Predominant abnormality incerebral glucose utilization in late-onset dementia of the Alzheimer type: across-sectional comparison against advanced late-onset and incipient early-onset cases. Journal of Neural Transmission 3 (1), 1–14.
Hoyer, S., Prem, L., Sorbi, S., Amaducci, L., 1993. Stimulation of glycolytic keyenzymes in cerebral cortex by insulin. NeuroReport 4 (7), 991–993.
Ishida, H., Takizawa, M., Ozawa, S., Nakamichi, Y., Yamaguchi, S., Katsuta,H., Tanaka, T., Maruyama, M., Katahira, H., Yoshimoto, K., Itagaki, E.,Nagamatsu, S., 2004. Pioglitazone improves insulin secretory capacity andprevents the loss of beta-cell mass in obese diabetic db/db mice: possibleprotection of beta cells from oxidative stress. Metabolism 53 (4),488–494.
Kumar, A., Schapiro, M.B., Grady, C., Haxby, J.V., Wagner, E., Salerno, J.A.,Friedland, R.P., Rapoport, S.I., 1991. High-resolution PET studies inAlzheimer's disease. Neuropsychopharmacology 4 (1), 35–46.
Lannert, H., Hoyer, S., 1998. Intracerebroventricular administration of streptozo-tocin causes long-term diminutions in learning and memory abilities and incerebral energy metabolism in adult rats. Behavioral Neuroscience 112 (5),1199–1208.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein mea-surement with the Folin phenol reagent. Journal of Biological Chemistry 193(1), 265–275.
Manschot, S.M., Biessels, G.J., Cameron, N.E., Cotter, M.A., Kamal, A.,Kappelle, L.J., Gispen, W.H., 2003. Angiotensin converting enzymeinhibition partially prevents deficits in water maze performance, hippocam-pal synaptic plasticity and cerebral blood flow in streptozotocin-diabeticrats. Brain Research 966 (2), 274–282.
Miles, P.D., Romeo, O.M., Higo, K., Cohen, A., Rafaat, K., Olefsky, J.M., 1997.TNF-alpha-induced insulin resistance in vivo and its prevention bytroglitazone. Diabetes 46 (11), 1678–1683.
Moreno, S., Farioli-Vecchioli, S., Ceru, M.P., 2004. Immunolocalization ofperoxisome proliferator-activated receptors and retinoid X receptors in theadult rat CNS. Neuroscience 123 (1), 131–145.
Nitsch, R., Hoyer, S., 1991. Local action of the diabetogenic drug, streptozotocin,on glucose and energymetabolism in rat brain cortex.Neuroscience Letters 128(2), 199–202.
Ohkawa, H., Ohishi, N., Yagi, K., 1979. Assay for lipid peroxides in animaltissues by thiobarbituric acid reaction. Analytical Biochemistry 95 (2),351–358.
Ott, A., Stolk, R.P., van Harskamp, F., Pols, H.A., Hofman, A., Breteler, M.M.,1999. Diabetes mellitus and the risk of dementia: the Rotterdam study.Neurology 53 (9), 1937–1942.
Parihar, M.S., Chaudhary, M., Shetty, R., Hemnani, T., 2004. Susceptibility ofhippocampus and cerebral cortex to oxidative damage in streptozotocintreated mice: prevention by extracts of Withania somnifera and Aloe vera.Journal of Clinical Neuroscience 11 (4), 397–402.
Phillips, M.E., Coxon, R.V., 1976. Effect of insulin and phenobarbital on uptakeof 2-deoxyglucose by brain slices and hemidiaphragms. Journal ofNeurochemistry 27 (2), 643–645.
Plaschke, K., Hoyer, S., 1993. Action of the diabetogenic drug streptozotocin onglycolytic and glycogenolytic metabolism in adult rat brain cortex andhippocampus. International Journal of Developmental Neuroscience 11 (4),477–483.
Polidori, M.C., Mecocci, P., 2002. Plasma susceptibility to free radical-inducedantioxidant consumption and lipid peroxidation is increased in very old subjectswith Alzheimer disease. Journal of Alzheimer's Disease 4 (6), 517–522.
Prickaerts, J., De Vente, J., Honig, W., Steinbusch, H., Ittersum, M.M.V.,Blokland, A., Steinbusch, H.W., 2000. Nitric oxide synthase does notmediate neurotoxicity after an i.c.v. injection of streptozotocin in the rat.Journal of Neural Transmission 107 (7), 745–766.

2216 A.R. Pathan et al. / Life Sciences 79 (2006) 2209–2216
Schuessel, K., Leutner, S., Cairns, N.J., Muller, W.E., Eckert, A., 2004. Impact ofgender on upregulation of antioxidant defence mechanisms in Alzheimer'sdisease brain. Journal of Neural Transmission 111 (9), 1167–1182.
Sharma, M., Gupta, Y.K., 2001. Intracerebroventricular injection of streptozo-tocin in rats produces both oxidative stress in the brain and cognitiveimpairment. Life Sciences 68 (9), 1021–1029.
Sharma, M., Gupta, Y.K., 2002. Chronic treatment with trans resveratrolprevents intracerebroventricular streptozotocin induced cognitive impair-ment and oxidative stress in rats. Life Sciences 71 (21), 2489–2498.
Simon, D.I., Mullins, M.E., Jia, L., Gaston, B., Singel, D.J., Stamler, J.S., 1996.Polynitrosylated proteins: characterization, bioactivity, and functionalconsequences. Proceedings of the National Academy of Sciences of theUnited States of America, vol. 93 (10), pp. 4736–4741.
Sonkusare, S., Srinivasan, K., Kaul, C., Ramarao, P., 2005. Effect of donepeziland lercanidipine on memory impairment induced by intracerebroventricularstreptozotocin in rats. Life Sciences 77 (1), 1–14.
Srinivasan, K., Patole, P.S., Kaul, C.L., Ramarao, P., 2004. Reversal of glucoseintolerance by pioglitazone in high fat diet-fed rats. Methods and Findings inExperimental and Clinical Pharmacology 26 (5), 327–333.
Steen, E., Terry, B.M., Rivera, E.J., Cannon, J.L., Neely, T.R., Tavares, R., Xu,X.J., Wands, J.R., de la Monte, S.M., 2005. Impaired insulin and insulin-likegrowth factor expression and signaling mechanisms in Alzheimer's disease—is this type 3 diabetes? Journal of Alzheimer's Disease 7 (1), 63–80.
Weinstock, M., Kirschbaum-Slager, N., Lazarovici, P., Bejar, C., Youdim, M.B.,Shoham, S., 2001. Neuroprotective effects of novel cholinesterase inhibitorsderived from rasagiline as potential anti-Alzheimer drugs. Annals of theNew York Academy of Sciences 939, 148–161.
Yonemitsu, S., Nishimura, H., Shintani, M., Inoue, R., Yamamoto, Y., Masuzaki,H., Ogawa, Y., Hosoda, K., Inoue, G., Hayashi, T., Nakao, K., 2001. Tro-glitazone induces GLUT4 translocation in L6 myotubes. Diabetes 50 (5),1093–1101.
Young, P.W., Cawthorne, M.A., Coyle, P.J., Holder, J.C., Holman, G.D., Kozka,I.J., Kirkham, D.M., Lister, C.A., Smith, S.A., 1995. Repeat treatment ofobese mice with BRL 49653, a new potent insulin sensitizer, enhancesinsulin action in white adipocytes. Association with increased insulinbinding and cell-surface GLUT4 as measured by photoaffinity labeling.Diabetes 44 (9), 1087–1092.
Copyright © 2022 FDOKUMEN




















