
Mammuthus meridionalis (Nesti, 1825) from Campo di Pile (L’Aquila, Abruzzo, Central Italy)
11
Mammuthus meridionalis (Nesti, 1825) from Campo di Pile (L’Aquila, Abruzzo, Central Italy) S. Agostini a, * , M.R. Palombo b, c , M.A. Rossi a , E. Di Canzio a , M. Tallini d, e a Soprintendenza per i Beni Archeologici dell’Abruzzo, Via degli Agostiniani 14, Chieti, Italy b Dipartimento di Scienze della Terra, Università di Roma “La Sapienza”, Roma, Italy c CNR, Istituto di Geologia Ambientale e Geoingegneria, Montelibretti, Roma, Italy d Dipartimento di Ingegneria delle Strutture, delle Acque e del Terreno Università dell’Aquila, Italy e Centro di Ricerca e Formazione in Ingegneria Sismica, Università dell’Aquila, Italy article info Article history: Available online 11 May 2012 abstract In the summer of 2009, four molars and two tusks, of a single individual of Mammuthus meridionalis, (a female, 28e30 year old) were discovered at Campo di Pile (L’Aquila, Abruzzo, Central Italy) in fluvial beach sediments of a depositional unit dated to the late Early Pleistocene. A new fragmentary tusk of a second individual has been recently discovered not far from the former. The morphology and dimensions of the penultimate molariform teeth fall within the range of variation of the Upper Valdarno specimens. The enamel pattern is consistent with that commonly found in M. meridionalis. Moreover, the relative thickness of the layers indicates a less advanced Mammuthus representative. In the L’Aquila Basin, at Madonna della Strada-Scoppito, a complete skeleton of M. meridionalis was found in sediments dating to the pre-Jaramillo Early Pleistocene, while some other isolated remains have been recorded at Rocca Santo Stefano and Colle Mancino. During the early Middle Pleistocene, Mammuthus (? Mammuthus trogontherii) and Palaeoloxodon co-occurred in the faunal assemblage of Pagliare di Sassa. Based on stratigraphic evidence, the Campo di Pile findings seem to be intermediate in age between the Madonna della Strada and the Pagliare di Sassa specimens. Ongoing research on the Campo di Pile site contributes to enhancement of knowledge of the Quaternary fauna and paleoenvironmental evolution of the L’Aquila Basin. Ó 2012 Elsevier Ltd and INQUA. All rights reserved. 1. Introduction The L’Aquila Basin (Abruzzo, Central Italy) has been well known since the last century because of the discovery of a number of Quaternary mammals and especially the finding of an almost complete skeleton of a large Mammuthus meridionalis at Madonna della Strada (Scoppito) (Maccagno, 1958, 1962), later chosen as a holotype of the subspecies Mammuthus meridionalis vestinus by Azzaroli (in Ambrosetti et al., 1972). In the summer of 2009, during the numerous interventions conducted in the area after the devastating 2009 L’Aquila earth- quake, some elephant remains (two tusks, two upper penultimate and one unworn, incomplete last upper molariform tooth along with some isolated plates belonging to a second ultimate tooth) were discovered in a sedimentary succession of L’Aquila Basin, cropping out at Campo di Pile (L’Aquila) (Fig. 1). Successively a new partial tusk and a few fragmentary remains of a stenonoid horse (an incomplete metatarsal), a large deer (two incomplete molars) and a large bovid (an incomplete horn-core) were found in the same area, about 50 m far from the first discovery, in a similar stratigraphic context. This paper gives a short account on the main findings of proboscideans recorded in the L’Aquila Basin and illustrates the remains found at Campo di Pile with the aim of further enhance- ment of knowledge of the elephant populations that inhabited this basin during the Quaternary. 2. The proboscidean fossil record of L’Aquila Basin: an overview The L’Aquila Basin represents, in the Abruzzo region, the area with the highest density of elephant remains (Fig. 1). The first discoveries are attributed to Giambattista Brocchi in 1818 (Mozzetti, 1893) and to T. Bonanni (1872). Both recovered elephant bones from the Calcare or Croce locality, close to the small church of San Pietro (Pagliare di Sassa). This site was revisited several times in the scientific literature as regards its fossil record. For instance, in 1913 the geologist P. Zuffardi, during the summer retreat of the * Corresponding author. E-mail address: [email protected] (S. Agostini). Contents lists available at SciVerse ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint 1040-6182/$ e see front matter Ó 2012 Elsevier Ltd and INQUA. All rights reserved. doi:10.1016/j.quaint.2012.05.013 Quaternary International 276-277 (2012) 42e52
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Mammuthus meridionalis (Nesti, 1825) from Campo di Pile (L’Aquila, Abruzzo, Central Italy)

at SciVerse ScienceDirect
Quaternary International 276-277 (2012) 42e52
Contents lists available
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
Mammuthus meridionalis (Nesti, 1825) from Campo di Pile (L’Aquila, Abruzzo,Central Italy)
S. Agostini a,*, M.R. Palombo b,c, M.A. Rossi a, E. Di Canzio a, M. Tallini d,e
a Soprintendenza per i Beni Archeologici dell’Abruzzo, Via degli Agostiniani 14, Chieti, ItalybDipartimento di Scienze della Terra, Università di Roma “La Sapienza”, Roma, ItalycCNR, Istituto di Geologia Ambientale e Geoingegneria, Montelibretti, Roma, ItalydDipartimento di Ingegneria delle Strutture, delle Acque e del Terreno Università dell’Aquila, ItalyeCentro di Ricerca e Formazione in Ingegneria Sismica, Università dell’Aquila, Italy
a r t i c l e i n f o
Article history:Available online 11 May 2012
* Corresponding author.E-mail address: [email protected] (S
1040-6182/$ e see front matter � 2012 Elsevier Ltd adoi:10.1016/j.quaint.2012.05.013
a b s t r a c t
In the summer of 2009, four molars and two tusks, of a single individual of Mammuthus meridionalis,(a female, 28e30 year old) were discovered at Campo di Pile (L’Aquila, Abruzzo, Central Italy) in fluvialbeach sediments of a depositional unit dated to the late Early Pleistocene. A new fragmentary tusk ofa second individual has been recently discovered not far from the former. Themorphology and dimensionsof the penultimate molariform teeth fall within the range of variation of the Upper Valdarno specimens.The enamel pattern is consistent with that commonly found in M. meridionalis. Moreover, the relativethickness of the layers indicates a less advanced Mammuthus representative. In the L’Aquila Basin, atMadonna della Strada-Scoppito, a complete skeleton of M. meridionalis was found in sediments dating tothe pre-Jaramillo Early Pleistocene, while some other isolated remains have been recorded at Rocca SantoStefano and Colle Mancino. During the early Middle Pleistocene,Mammuthus (?Mammuthus trogontherii)and Palaeoloxodon co-occurred in the faunal assemblage of Pagliare di Sassa. Based on stratigraphicevidence, the Campo di Pilefindings seem to be intermediate in age between theMadonna della Strada andthe Pagliare di Sassa specimens. Ongoing research on the Campo di Pile site contributes to enhancement ofknowledge of the Quaternary fauna and paleoenvironmental evolution of the L’Aquila Basin.
� 2012 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
The L’Aquila Basin (Abruzzo, Central Italy) has been well knownsince the last century because of the discovery of a number ofQuaternary mammals and especially the finding of an almostcomplete skeleton of a large Mammuthus meridionalis at Madonnadella Strada (Scoppito) (Maccagno, 1958, 1962), later chosen asa holotype of the subspecies Mammuthus meridionalis vestinus byAzzaroli (in Ambrosetti et al., 1972).
In the summer of 2009, during the numerous interventionsconducted in the area after the devastating 2009 L’Aquila earth-quake, some elephant remains (two tusks, two upper penultimateand one unworn, incomplete last upper molariform tooth alongwith some isolated plates belonging to a second ultimate tooth)were discovered in a sedimentary succession of L’Aquila Basin,cropping out at Campo di Pile (L’Aquila) (Fig. 1). Successively a newpartial tusk and a few fragmentary remains of a stenonoid horse
. Agostini).
nd INQUA. All rights reserved.
(an incomplete metatarsal), a large deer (two incomplete molars)and a large bovid (an incomplete horn-core) were found in thesame area, about 50 m far from the first discovery, in a similarstratigraphic context.
This paper gives a short account on the main findings ofproboscideans recorded in the L’Aquila Basin and illustrates theremains found at Campo di Pile with the aim of further enhance-ment of knowledge of the elephant populations that inhabited thisbasin during the Quaternary.
2. The proboscidean fossil record of L’Aquila Basin: anoverview
The L’Aquila Basin represents, in the Abruzzo region, the areawith the highest density of elephant remains (Fig. 1). The firstdiscoveries are attributed to Giambattista Brocchi in 1818(Mozzetti, 1893) and to T. Bonanni (1872). Both recovered elephantbones from the Calcare or Croce locality, close to the small church ofSan Pietro (Pagliare di Sassa). This sitewas revisited several times inthe scientific literature as regards its fossil record. For instance, in1913 the geologist P. Zuffardi, during the summer retreat of the
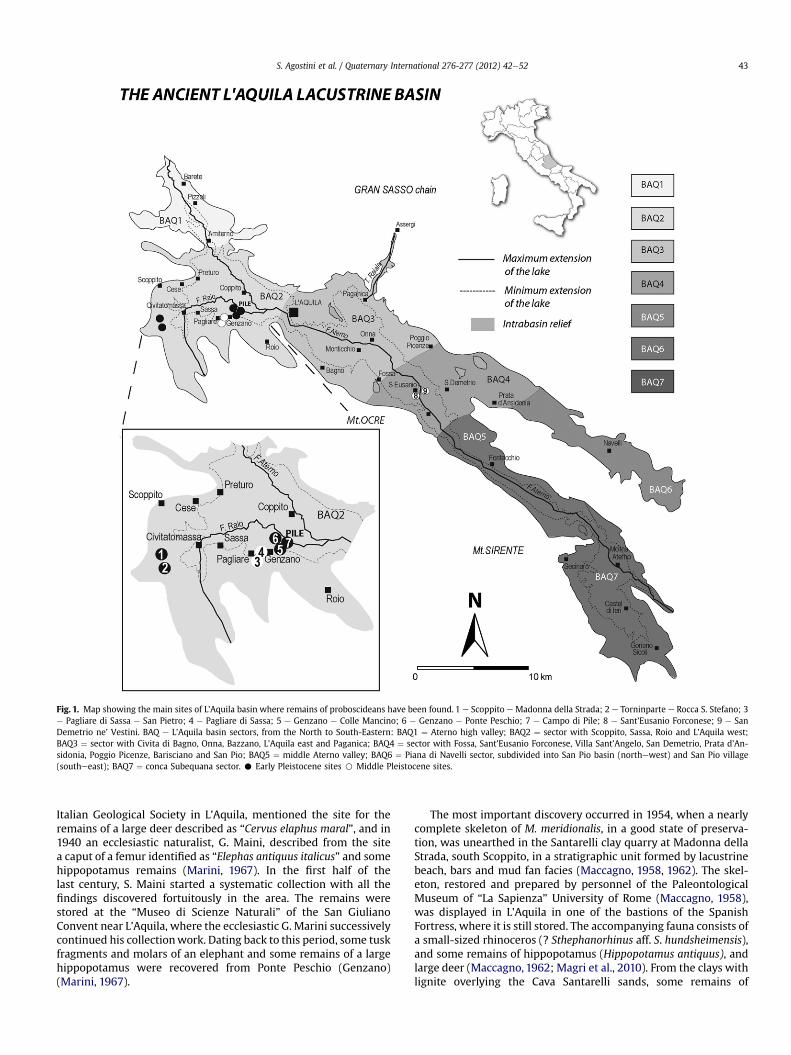
Fig. 1. Map showing the main sites of L’Aquila basin where remains of proboscideans have been found. 1 e Scoppito e Madonna della Strada; 2 e Torninparte e Rocca S. Stefano; 3e Pagliare di Sassa e San Pietro; 4 e Pagliare di Sassa; 5 e Genzano e Colle Mancino; 6 e Genzano e Ponte Peschio; 7 e Campo di Pile; 8 e Sant’Eusanio Forconese; 9 e SanDemetrio ne’ Vestini. BAQ e L’Aquila basin sectors, from the North to South-Eastern: BAQ1 ¼ Aterno high valley; BAQ2 ¼ sector with Scoppito, Sassa, Roio and L’Aquila west;BAQ3 ¼ sector with Civita di Bagno, Onna, Bazzano, L’Aquila east and Paganica; BAQ4 ¼ sector with Fossa, Sant’Eusanio Forconese, Villa Sant’Angelo, San Demetrio, Prata d’An-sidonia, Poggio Picenze, Barisciano and San Pio; BAQ5 ¼ middle Aterno valley; BAQ6 ¼ Piana di Navelli sector, subdivided into San Pio basin (northewest) and San Pio village(southeeast); BAQ7 ¼ conca Subequana sector. C Early Pleistocene sites B Middle Pleistocene sites.
S. Agostini et al. / Quaternary International 276-277 (2012) 42e52 43
Italian Geological Society in L’Aquila, mentioned the site for theremains of a large deer described as “Cervus elaphus maral”, and in1940 an ecclesiastic naturalist, G. Maini, described from the sitea caput of a femur identified as “Elephas antiquus italicus” and somehippopotamus remains (Marini, 1967). In the first half of thelast century, S. Maini started a systematic collection with all thefindings discovered fortuitously in the area. The remains werestored at the “Museo di Scienze Naturali” of the San GiulianoConvent near L’Aquila, where the ecclesiastic G. Marini successivelycontinued his collectionwork. Dating back to this period, some tuskfragments and molars of an elephant and some remains of a largehippopotamus were recovered from Ponte Peschio (Genzano)(Marini, 1967).
The most important discovery occurred in 1954, when a nearlycomplete skeleton of M. meridionalis, in a good state of preserva-tion, was unearthed in the Santarelli clay quarry at Madonna dellaStrada, south Scoppito, in a stratigraphic unit formed by lacustrinebeach, bars and mud fan facies (Maccagno, 1958, 1962). The skel-eton, restored and prepared by personnel of the PaleontologicalMuseum of “La Sapienza” University of Rome (Maccagno, 1958),was displayed in L’Aquila in one of the bastions of the SpanishFortress, where it is still stored. The accompanying fauna consists ofa small-sized rhinoceros (? Sthephanorhinus aff. S. hundsheimensis),and some remains of hippopotamus (Hippopotamus antiquus), andlarge deer (Maccagno, 1962; Magri et al., 2010). From the clays withlignite overlying the Cava Santarelli sands, some remains of

S. Agostini et al. / Quaternary International 276-277 (2012) 42e5244
amphibians (Triturus sp., Discoglossus cf. D. pictus, Bufo cf. B. viridis,Rana dalmatina), reptiles (Emys orbicularis), and scanty remains ofthe arvicolid Mimomys savini, have been reported (Kotsakis, 1988),together with terrestrial and freshwater molluscs (Esu et al., 1992,1993). Pollen analysis carried out in the lowermost portion of thislignitic succession suggests for Madonna della Strada site an EarlyPleistocene age, older than the Jaramillo Subchron (1.07 Ma) andyounger than MIS 40 (about 1.3 Ma), giving a chronologicalconstraint for the southern mammoth skeleton found in theunderlain sandy deposits (Magri et al., 2010 and referencestherein).
A femur ofM. meridionalis has been reported also in Rocca SantoStefano (Torninparte), 1.2 km south of Madonna della Strada, in thesame sands of the stratigraphic unit exposed at Santarelli quarry(Maccagno, 1965). Two last upper molariform teeth, a tusk andsome ribs belonging to the same species were found at ColleMancino, near Genzano (Maccagno, 1965). These specimens arenow stored in the Paleontological Museum of Federico II Universityof Naples.
At the end of the last century, the discovery of the early MiddlePleistocene bearing site of Pagliare di Sassa, 300 m from thehistorical site of San Pietro (see above), was of particular interest(Palombo et al., 2010 and references therein). The fossil-bearingsequence of Pagliare di Sassa is part of a sedimentary successionwhich overlies the lacustrine Madonna della Strada deposits in thesouthernewestern part of the L’Aquila Basin. The exposed lith-ofacies consists of an alluvial fan deposit with grey basal clays,partially pedogenised at the top, and cut by a channel, filled bysandy lenses with fine gravel intercalations, prograding northwardsto the L’Aquila paleo-lake (Palombo et al., 2010). Among thevertebrate remains, which mostly consisted of disarticulated skel-etal elements of large mammals and scanty and isolated fragmen-tary remains of small mammals and other vertebrates,Mammuthus(? M. trogontherii) and Palaeoloxodon antiquus were both present,although with different numbers of remains. Mammuthus (?M. trogontherii) is documented by very fragmentary remainsbelonging to a single, large individual, which was largely damagedby bulldozer excavation at the time of the discovery. Among thespecimens retrieved (two occipital condyles, tusk fragments,vertebrae and vertebral apophyses, ribs, the glenoid joint ofa scapula, a proximal epiphysis of a humerus and an incompleteulna), no specimens with firm diagnostic features were present,although some tusk fragments clearly show Schreger lines whose
Fig. 2. Campo di Pile: tusks of Mammuthus meridionalis discovered during the surveymade by Soprintendenza Archeologica dell’Abruzzo in the summer of 2009.
angles fall in the variability range of Mammuthus. Similarly, thepossible occurrence of a straight-tusked elephant has been inferredon the basis of few tusk fragments, whose Schreger angles aredefinitely wider than those of Mammuthus, falling in the range ofouter angles of Palaeoloxodon antiquus (Palombo and Villa, 2001,2007). The faunal assemblage includes, beside elephants andamong others (S. hundsheimensis, Hippopotamus cf. H. antiquus, Susscrofa, Megaloceros savini, Praemegaceros verticornis, Crocuta cf. C.crocuta, Lepus sp., Crocidura sp.) the voles Mimomys savini andMicrotus ex gr. Microtus hintoni e Microtus gregaloides (Palomboet al., 2010), whose co-occurrence indicates the fauna is notyounger than the early Middle Galerian (see Sala and Masini, 2007for a biochronology of Italian small mammals).
Fig. 3. Sketch of the stratigraphic unit cropping out at Campo di Pile (excavation2009). Star indicate level from which Mammuthus meridionalis remains have beenretrieved. A e agricultural brown soil; B e illuvial clay and silt dark brown level; C e
sands with clay chips, rarely carbonate concretion; D e coarse sands with a carbonateconcretion horizon and a laminar oxidized horizon of silt and sand; E e sands; F e
alternating of silt and fine sands; G e thin laminae of sands; H e gray silt with smallgravel at the bottom; I e pockets of silt and small gravel; L e small gravel partiallycemented, limestone crust at the bottom; M e silty sand; Ne small gravel cementedupon which rested the tusks and the two molars M2; O e yellow and gray sands inwhich were the two molars (M3). Two joints sets with a decimeter of space arepresent: J1 ¼ N60� E � dip S80� , J2 ¼ N60� W e dip S80.

S. Agostini et al. / Quaternary International 276-277 (2012) 42e52 45
Only a few remains have been recorded from the southernmostL’Aquila Basin. A right tusk of P. antiquus has been found at San-t’Eusanio Forconese (Maini, 1956) and is today displayed at the“Museo di Scienze Naturali” of San Giuliano. An incomplete juvenileleft mandiblewith twomolariform teeth, possibly pm4 andm1, andtwo fragments of a unworn molariform tooth have been docu-mented at San Demetrio ne’Vestini. The ?m1 consist of 9 plates, itsdimensions are length 125mm andwidth 45 mm (D’Erasmo,1931).All these remains probably came from the same formation ofMiddle Pleistocene age, a fluvial-lacustrine deposit of the sectorBAQ4 (Fig. 1), containing levels of tephra and volcanic minerals inthe sand fraction (Bertini and Bosi, 1993). The specimens from SanDemetrio, originally preserved in the IstitutoTecnico of Chieti, werelost during the Second World War. The mandible and the
Fig. 4. Mammuthus meridionalis (Nesti, 1825), Campo di Pile: left tusk (SBABp 318). Comparistusk of Mammuthus meridionals (Nesti, 1825) found at Madonna della Strada (Scoppito; dat
molariform tooth were identified as “Elephas meridionalis”(D’Erasmo, 1931); successively, Ambrosetti (pers. com. in Bertiniand Bosi, 1993) referred the remains to a M. trogontherii or P. anti-quus juvenile individual. However, the features of the teeth suggestthat they belong to M. trogontherii, as supported by the strati-graphic context and consistently with their morphology anddimensions (Agostini et al., 2001).
3. Campo di Pile: geology and Paleontology
Campo di Pile is located in the industrial area west of L’Aquila.The geological context is represented by a fluvial succession orga-nized in repeated alternations of sand and silt with discontinuouslenses of matrix-supported gravel. The succession dips 15e20� to
on among intra- and extra-alveolar measurements of tusks from Campo di Pile and thea from Maccagno, 1962) and at Colle Mancino (Genzano; data from Maccagno, 1965).

S. Agostini et al. / Quaternary International 276-277 (2012) 42e5246
the south and appears clearly tilted. The main outcroppings of theCampo di Pile succession are visible only in artificial cuts, alongroads, and also during occasional excavations for buildingfoundations.
The discoveries of elephant remains occurred in two steps. Thefirst fossil-bearing layer was excavated in June 2009, two tusks, twoupper penultimate (M2), and two non-erupted upper last (M3)molariform teeth, as well as some isolated plates belong to the M3,were buried here to a depth of about 2 m in an area of 4 m2. Thetusks were placed orthogonally to the fluvial current, whosedirection has been reconstructed by means of cross-stratification(foreset lamination) and convolute lamination (Fig. 2).
Successively, in October 2011, new industrial building excava-tions unearthed a fragment of right tusk, belonging to a secondindividual, about 50 m away from the first finding, in a similarcontext of facies. Furthermore, a large fragment of a horn-core ofa large bovid was found 1 m above the tusk.
A few other remains (a horse metatarsal and two deer teeth)were scattered in the area between the two mammoth finds. Thesebones are too few and poorly preserved for any firm identification,albeit the proportions of the incomplete metatarsal suggest itmight belong to a middle-sized, moderately slender stenonoidhorse (Equus sp. cf. ? Equus stenonis). The dimensions of the teethindicate a large-sized deer. Preliminary analysis performed on sandsamples has not provided any small mammal remains.
3.1. Geological setting
The L’Aquila Basin is one of the largest depressions of tectonicorigin in the central Apennines of the Abruzzo region, now drainedby the Aterno River to which tributaries flow, such as the RaioStream on the right hand of the hydrographic basin and the RaialeStream on the left hand. During the Early and Middle Pleistocene,L’Aquila basin was the site of a complex fluvial-lacustrine system,indicated by several allostratigraphic units that divide it intodistinct sub-basins (Fig.1). The system of sub-basins was crossed byslightly incised valleys and depressed areas with lakes and highmarshes, environments that do not always allow clear correlationsamong the depositional units. The main Quaternary climatechanges are recorded by palaeosols, repeated erosive processes andpollen data from the lacustrine facies (GE.MI.NA., 1963; Magaldiand Tallini, 2000; Galli et al., 2010; Magri et al., 2010).
In the central sector of the basin, affected by distensile tectonicprocess, the morphostructural evolution drove the progressive
Table 1- Dental variables of penultimate and ultimate molariform teeth (M2 and M3) of Mamm
Site Specimen Tooth Length OcclusalLength
Width Ow
Campo di Pile SBAAp 320 M2 206 197 90 8Campo di Pile SBAAp 321 M2 197 195 92 8Redicicoli MPR E2 M2 201 177 94 8Monte Tenda MCSNV-10688 M2 206 c.120 90 7Campo di Pilea SBAAp 322 M3 >215 e c.93 e
Farneta IGF 12434 M3 301 215 115 1Pietrafitta CET 80 M3 318 190 103 9Pietrafitta CET 1119 M3 245 133 84 7Madonna della Strada sn right M3 255 210 98 9Madonna della Strada sn left M3 258 240 95 9Leffe MSNB 1171 M3 243 e e e
Leffe MSNB ‘1877’ M3 e 150 e 9Torrente Crostolo MSNRE-CRO3 M3 >243 171 110 1Rio Pradella MCICS 7644 M3 235 120 90 8Colle Marino right M3 259 248 108 1Colle Marino left M3 258 245 109 1
a Tooth partially reconstructed by using isolate plates.
westward migration of the lacustrine depocentre and the basinextension (Bosi and Bertini, 1970; Bagnaia et al., 1989; Bertini et al.,1989; Bertini and Bosi, 1993; Cavinato et al., 1994; Galli et al., 2010;Messina et al., 2010; Agostini et al., in press).
The Campo di Pile elephantine-bearing site is situated in theScoppito, Sassa, Roio and L’Aquila west sector, where two fluvio-lacustrine units crop out. The succession of the Early Pleistoceneis characterized by sands and silt typical of lacustrine and fluvialenvironments and are pertinent to the stratigraphic unit called“Madonna della Strada” (MdS). In particular, the MdS unit, for someauthors a complex of units (Bosi and Messina, 1991) consists ofa facies association. At the bottom sediments deposited in lacus-trine and relative shore environments with repeated lignite andorganic silt levels are present, while at the top alluvial facies typicalof a braided river system, alluvial fan and marsh facies occur. Thecomplete skeleton ofM. meridionalis from Scoppito (Madonna dellaStrada, AQ) was recovered in the sandy lacustrine sediments of thisunit (Maccagno, 1962; Magri et al., 2010; Mancini et al., 2011).
The second unit, dated to the early Middle Pleistocene and herenamed “Pagliare di Sassa” (PdS), lies in unconformity to the MdSunit. The PdS unit is mainly constituted of sands and silt withresedimented soils at the base. The PdS shows alluvial fan faciescharacterized by dense flows and progradational geometries(foreset and topset) interfingered with lacustrine marsh and fluvialfacies. The homonymous fossil-bearing site of Pagliare di Sassa wasdiscovered in the PdS stratigraphic unit (Palombo et al., 2010).
At the Campo di Pile site, the southern mammoth fossil remainswere found near the top of the outcropping stratigraphic section,formed by an alternation of yellow sand and grey silt, depositedalong a fluvial route in a beach context (Fl, Fsm and Fm facies modelafter Miall, 1996) (Fig. 3). The fluvial, coarse-grained sedimentscontaining the mammoth remains overlap lacustrine clay strata.Thin layers of high marsh facies (organic-matter-rich mud and claywith Planorbis) are both laterally heterophic and overlie thefossiliferous layers. The Campo di Pile stratigraphic section is cut byan erosional surface below the deposition of alluvial gravel sedi-ments characterized by “cut and fill” and lateral bar geometries.This latter depositional unit is generally referred to the late MiddlePleistocene (Coltorti and Dramis, 2006; Tallini et al., 2011).
A possible constraint on the age of the Campo di Pile depositsmay be the presence of a palaeosol without tephra components,lying just above the fossil-bearing levels, which in turn is overlainby more than 7 m of clay and organic-matter-rich mud. Magaldiand Tallini (2000) recognized several palaeosols in the L’Aquila-
uthus meridionalis from selected Italian localities.
cclusalidth
Height Numberof plates
Platesin use
Enamelthickness
Lamellarfrequency
Hypsodontyindex (H/W)
0.5 94 9 9 3.1 4.5 c. 1.040.1 91 9 9 3.1 4.5 c. 0.994 123 9 9 2.6 6 1.36 115 10 7 3.0 6 1.28
118 >11 0 e 5 c. 1.2704 e 14 12 2.9 5 1.173 131 14 10 3.3 5 1.322 130 14 8 2.6 6 1.52 e >6 5 3.5 5 e
3 e >7 7 3.5 5 e
e 14 0 e 5.5 e
0 e 12 6 e 5.5 e
00 170 >11 8 3.0 5 1.53 158 14 6 2.4 6 1.7601 141 >11 11 3.7 5 1.300.5 146 >11 11 3.7 5 1.33

S. Agostini et al. / Quaternary International 276-277 (2012) 42e52 47
Scoppito Basin, some of which are characterized by the absence oftephra and are present near the top of MdS Unit. Accordingly, thehypothesis that the palaeosol at Campo di Pile may be correlatedwith MdS soils cannot be ruled out. Nevertheless, the hypothesisneeds sounder support due to the high tectonic activity charac-terizing the area, which displaced the MdS Unit (Tallini et al., 2011).
Fig. 5. Mammuthus meridionalis (Nesti, 1825) from Pile (L’Aquila Basin, central Italy):natural section of a large fragment of tusk showing Schreger lines and angles.
3.2. The southern-mammoth from Campo di Pile
The mammoth remains found at Campo di Pile belong to twoindividuals. The remains found during the first survey in the Campodi Pile area (two tusks and the molariform teeth) belong to a singleindividual. The tusks are in a poor state of preservation, coveredwith cemented sediment, rich in iron-manganese oxides and rarepyrite framboids, as a result of an intense syn-diageneticgeochemical process related to alteration and pedological phasesuccessive to the burial. The pervasive oxidation of the tusks hasgiven them a red-brown color. The condition of the M2 wasacceptable; the incomplete M3 was not fully formed, the cementwas present only at the base, sand and fine gravel filling theremaining space between the plates. The other molar, M3, wasfound as isolated plates in an imbricate disposition.
3.2.1. TusksTusks are quite slender, less massive and curved than are usually
found in M. meridionalis adult males (Fig. 4). They show a doublecurvature (spiraling) not much more accentuated than in somePalaeoloxodon (see e.g. German specimens from Neumark Nord,Palombo et al., 2010; and from Greece, Tsoukala et al., 2011), andless than in M. meridionalis adult males. The third tusk, a largefragment found in the same area, does not substantially differ fromthe others.
To give further support to the identification of Campo di Pileelephants, the Schreger line pattern has been examined on the flatsurfaces of natural transversal sections of broken portions of tusks.Each surface was photographed in high-resolution, and the imageswere processed with the software CV9000, which allowedmeasurement of Schreger angles, with a precision of about 95%(Palombo and Villa, 2001, 2007). The whole spectrum of the angleswas measured, including ‘outer’ (near the dentine-cementumjunction) and ‘inner’ angles (near the proximal-distal tusk axis)(Table 1).
Fig. 6. Mammuthus meridionalis (Nesti, 1825) from Pile (L’Aquila Basin, central Italy): upperocclusal (A, B) and lateral (A0 , B0) view.
The Schreger pattern, a unique characteristic of proboscideanivory and visible in cross-sections of tusks (Obermayer, 1881),consists of two intersecting sets of spiral lines that radiate clock-wise and counterclockwise from the tusk’s longitudinal axis. Theintersections of Schreger lines form two types of Schreger angles econcave angles opening towards themedial (inner) area of the tusk,and convex angles opening towards the lateral (outer) area of thetusk e whose width increases from the tusk nerve/pulp cavity(inner angles) towards the dentine-cementum junction (outerangles) (Espinoza and Mann, 1993; Fisher et al., 1998; Palombo andVilla, 2001; Trapani and Fisher, 2003). This pattern is the visualmanifestation of the arrangement of the dentine tubules along thelongitudinal axis of the tusk.
The Schreger line pattern of Campo di Pile tusks clearly indicatesthat the tusk belongs to a Mammuthus representative (Fig. 5). Onthe natural cross-sections of tusks found at Campo di Pile, thevalues of the Schreger angles, including those near the dentine-cementum junction are, on average, less than 90� and thereforenarrower than in extant elephants and fossil palaeoloxodonts andconsistent with the average value reported for the inner Schreger
left (SBAAp 321) (A) and right (SBAAp 320) (B) penultimate molariform teeth (M2) in

Table
2-Den
talv
ariables
ofpen
ultim
atean
dultim
atemolariform
teeth(M
2an
dM3)
ofMam
muthu
smeridiona
lisfrom
thepre-Jaram
illoEa
rlyPleistoc
eneItaliansitesof
Upper
Valdarno(O
livolaþ
Tassofaunal
units),a
ndlatest
Early
Pleistoc
eneRus
sian
localityof
Syny
ayaBalka
(Tam
irpen
insu
la).
Toothlength
Width
(W)
Heigh
t(H
)Plates
Enam
elthickn
ess
Lamellarfreq
uen
cyHyp
sodon
tyindex
(H/W
)
NMea
nRan
geN
Mea
nRan
geN
Mea
nRan
geN
Mea
nRan
geN
Mea
nRan
geN
Mea
nRan
geN
Mea
nRan
ge
M3e
Upper
Valdarno
(Oliv
olaþ
TassoFU
)21
269
226e
343
2910
183
e12
327
131
101e
160
3313
11e14
333.2
2.6e
3.9
365
4.0e
6.0
131.3
1.10
e1.60
M3e
Sinya
yaBalka
1424
122
5e31
714
106
93e11
711
146
127e
162
1014
12e16
143.14
2.5e
3.5
145.3
4.5e
6.0
101.38
1.13
e1.56
M2e
Upper
Valdarno
(Oliv
olaþ
TassoFU
)13
212
175e
243
1589
80e10
311
120
103e
148
179
8e10
172.70
2.2e
3.4
65.00
4.5e
6.0
111.40
1.00
e1.60
M2e
Sinya
yaBalka
1822
919
4e25
517
9990
e11
215
143
125e
166
189
8e11
182.80
2.5e
3.5
185.6
4.5e
6.0
141.48
1.17
e1.75
S. Agostini et al. / Quaternary International 276-277 (2012) 42e5248
angles of M. meridionalis, M. trogontherii and most Mammuthusprimigenius. According to Fisher et al. (1998), the values of Schregerangles in Mammuthus tusks range from 62� to 105� with a mean of87.1�. The values of the ‘outer’ Schreger angles of M. primigenius,measured by Trapani and Fisher (2003) at the dentine-cementumjunction, range from about 70� to 100�, while single maximumvalues of 115� and 125� were found by Ábelova (2008) close to thedentine-cementum junction, around 0e2 cm from the tusk surface.In the sample of M. primigenius (49 tusks) examined by Palomboand Villa (2001, 2007), the values of Schreger outer angles rangefrom 65� to 90� with a mean of 71.2�, while the values of innerSchreger angles range from 33� to 55�, with a mean of 41�, showingan increase from the center of the tusk to the dentine-cementumjunction of about 57%. Similar values characterize the ‘outer’Schreger angles of M. meridionalis (67�e85�) and M. trogontherii(68�e77�) (Palombo and Villa, 2001, 2007).
Although only a few inner Schreger angles of the Campo diPile tusks have been measured because of the poor preservationstatus shown by the innermost portion of all analyzed sections(extending to about 20 mm from the central axis of the tusk), thevalues of the measured angles, clearly “V” shaped, do notsignificantly differ each other, ranging from about 33� to 45�,with an average value of 40�. The values of angles of the sectionband located between about 20 to about 40 mm from the centreof the tusk, range from about 39� to 57� (average value ¼ 51�),while those located in the band between about 40 and 60 mmrange from about 58� to 95� (average value ¼ 70�). The range ofthe outermost angles, located close to dentine-cementum junc-tion, is from about 85� to 95�, with an average value of 89�. Therange of variability of Schreger angles increases from the centerof the tusk to the dentine-cementum junction, and their averagevalue shows an increase of about 120%. All in all, the valuesmeasured on the natural section of Campo di Pile tusks fall withinthe range of Mammuthus. Taking into account the position of theanalyzed sections (about at the middle point of the tusk length),the prevalence of V shaped angles, the nearly constant bendradius shown by the Schreger lines in the inner portion of thesection, and the Schreger lines slightly bending and divergingfrom the radius near the dentine-cementum junction, arecoherent with the features commonly observed in adultmammoth specimens (Palombo and Villa, 2001; Àbelova, 2008),consistently with the age of about 28e30 years inferred for theCampo di Pile individual on the basis of the worn stage of itspenultimate molariform teeth.
3.2.2. Molariform teethThe penultimate upper molariform teeth (M2) are not particu-
larly high-crowned probably because of their quite advanced wear(Fig. 6; Table 1). Both have nine worn plates, three of which aresupported by the first root. The occlusal surface is ovate witha faintly convex lateral side. On the occlusal surface, in the lessworn plates, the enamel occlusal figures show a quite large ovalcentral loop, and two smaller lateral and medial loops, while in theplates at an intermediate wear stage, the central expansion isrounded and definitely less wide than the lateral and medial parts.The enamel is thin, crinkled, and regularly folded, but the folds donot extend to the lateral and medial borders of the enamel figure.All in all, the morphology and dimensions of the molars fall withinthe wide range of late Villafranchian M. meridionalis of the Italianpeninsula (Palombo and Ferretti, 2005). Conversely, molariformteeth from Italian early Galerian sites (e.g. Redicicoli), are moreadvanced in having a higher lamellar frequency and hypsodontyindex, as shown by specimens from the French site of Saint Prest(around 1.1e1.0 Ma) (¼ M. meridionalis depereti in Guerin et al.,2003) and by the rich Russian sample from the Sinyaya Balka

S. Agostini et al. / Quaternary International 276-277 (2012) 42e52 49
(Taman peninsula, southern Russia) dated to around 1.0 Ma(Dubrovo and Palombo, unpublished data) (Tables 1, 2).
Only a few plates of ultimate molariform teeth (M3) are fusedto each other, and a number of plates have been found isolated,making it difficult to reconstruct the actual shape of these teeth. Itis worth noting that the ovoidal outline of the shape is consistentwith the common shape of M. meridionalis upper molars, as wellas the number and development of plate tubercules, the centralmaintaining a nearly columnar shape from the roots to the top ofthe plate, while those of the lingual and labial sides show a nearlytriangular shape (Fig. 7). The hypsodonty index inferred for thereconstructed M3 is, as expected, higher than those obtained forthe worn M2, albeit it could be overestimated because thecalculated width does not count the cementum thickness(Table 1).
Since previous studies have demonstrated an interest in theanalysis of enamel structure in inferring the evolutionary level ofMammuthus representatives (see Ferretti, 2003a,b, and referencestherein), an isolated plate of the last molar was sectioned forenamel microstructure analysis under the scanning electronmicroscope. Small portions of dental tissue were removed andcubic samples were sawn with a circular diamond saw formineralogical sampling in order to have horizontal and sagittalsections of enamel, which were then examined at low vacuum bymeans of a Quantas 250 SEM at different magnifications. Theenamel pattern of the Campo di Pile mammoth looks like thatthose already described by Ferretti (2003a, 2003b in Mammuthusrepresentatives. In particular, the enamel of molariform teethfrom Campo di Pile is characterized by two main arrangements ofenamel prisms (3-D enamel and radial enamel), by an enameltype lacking a prismatic organization (prismless enamel) and bythe presence of decussating prism layers forming Hunter-Schreger bands. In the 3-D enamel type (sensu Pfretzschener,
Fig. 7. Mammuthus meridionalis (Nesti, 1825) from Pile (L’Aquila Basin, central Italy):ultimate, unworn still uncomplete molariform tooth (M3, SBAAp 322) reconstructed byadding three isolated plates to the 8 already fused plates. (A) ¼ occlusal view;(B) ¼ lateral view.
1994), prisms, although departing parallel to each, suddenlyform bundles interweaving in all three spatial directions. In theradial enamel, prisms are parallel to each other, and directedradially from the enamel-dentine junction (EDJ). Some prismsangle at about 60� to the occlusal surface and show a ginko-leafor a key�hole section (radial enamel subtype A of Ferretti, 2003a)(Fig. 8). Some others are sub-parallel to the occlusal and normalto the enamel outer surface and show an irregular key-holetransverse section (radial enamel subtype B), while others areparallel to the occlusal surface and show a key-hole sub-circularsection (radial enamel subtype B) (Fig. 9). The prismlessenamel, lacking organized prisms, is characterized by a matrix ofapatite crystallites parallel to each other, but diversely oriented.Moreover, slight prism decussating in layers led to the presenceof variously undulated Hunter-Schreger bands (HBS). Theseenamel types characterize the three different layers that can bedetected in sagittal sections (parallel to the tooth occlusal-basaland mesio-distal axes) of the enamel band. From the enamel-cement junction (ECJ) to the EDJ, three different layers can bedetected: outer, middle and inner enamel layer (Fig. 10). Theoutermost layer, representing less than 20% of the total enamelthickness, shows a rather complex pattern with the outermostthin zone with a prismless enamel type passing to a radialenamel subtype C, while the inner part shows a radial enamel ofsubtype C. The transition to the middle layer is marked by a sharpchange in the inclination of the prisms (Fig. 11). In this layer, withthe thickness accounting for about 65% of the total thickness ofthe enamel band, the radial enamel of the outer zone passes toa decussed enamel with a large, slightly undulated HSB rising tothe occlusal surface with an average angle of about 90�. Themiddle layer gradually passes to the inner layer, the thinnest ofabout 15% of the total enamel thickness, consisting of 3-D enameltype. The enamel pattern shown by teeth found at Campo di Pileis consistent with the pattern commonly found in M. meridionalis.Moreover, the relative thickness of the layers indicates a non-advanced Mammuthus representative (see Ferretti, 2003a,2003b).
Fig. 8. Mammuthus meridionalis (Nesti, 1825) from Pile (L’Aquila Basin, central Italy):Horizontal section of middle enamel layer showing the ginko-leaf cross-section ofprisms.

Fig. 11. Mammuthus meridionalis (Nesti, 1825) from Pile (L’Aquila Basin, central Italy):Sagittal section of the enamel showing the change in prism bending at the transitionfrom middle to outer enamel.
Fig. 9. Mammuthus meridionalis (Nesti, 1825) from Pile (L’Aquila Basin, central Italy):Horizontal section of outer enamel layer with prisms showing arch-shaped sectionsand frequent fusion with adjacent prisms.
S. Agostini et al. / Quaternary International 276-277 (2012) 42e5250
4. Remarks
The southern mammoth remains found at Campo di Pile inJune 2009, likely belong to a 28e30 year old female. Themorphology and dimensions of the penultimate molariformteeth fall within the range of variation of those of M. meridionalismeridionalis from Upper Valdarno. Based on the stratigraphicevidence available thus far, the hypothesis that the Campo di PileM. meridionalis remains might be younger than those of Scoppito,possibly because they were found in the uppermost levels of MdS
Fig. 10. Mammuthus meridionalis (Nesti, 1825) from Pile (L’Aquila Basin, central Italy):Sagittal section showing the three enamel layer, the Hunter-Schreger bands charac-terizing the middle layer, and the irregularity of the outer enamel surface at thecontact with the cementum.
Unit, seems to be reasonable (Fig. 12). Therefore, assuming thatthe age of the Campo di Pile findings may be slightly less thanthose of M. meridionalis “vestinus” from Madonna della Strada-Scoppito, it seems to confirm once more that, on one hand,some dental characteristics firmly differentiate this “subspecies”from the Upper Valdarno populations, and on the other, thatderived populations of southern mammoth only occurred in Italyby the end of the Early Pleistocene (e.g. Oriolo, Rio Pradella)(Ferretti, 1999; Palombo and Ferretti, 2005). On average, themorphology and dimensions of the M. meridionalis teeth fromItalian pre-Jaramillo localities (e.g. Mugello, Tuscany; Pietrafitta,Perugia, Leffe lower level, Bergamo; and may be Torrente Cros-tolo, Reggio Emilia) confirm that the evolutionary levels of themolariform teeth of southern mammoth samples dating fromaround 1.5e1.2 Ma do not substantially differ from that of UpperValdarno specimens (see Palombo and Ferretti, 2005 and refer-ences therein) (Table 1). Following Azzaroli (1977), “M. mer-idionalis vestinus” representatives would have been present inthe Italian peninsula from around 1.5 to 0.8 Ma. They would havediffered from the typical form from Upper Valdarno in havinga larger overall size, a more advanced cranial morphology asdocumented by the Farneta specimens, showing a shorter andhigher skull than those from Upper Valdarno, with the vertexmore caudally displaced, an extremely concave forehead, andlonger tusk sheaths. Nevertheless, only the latter feature can beobserved in the incomplete skull from Madonna della Strada.Unfortunately, the skulls of M. meridionalis found in the Pie-trafitta faunal assemblage (Perugia), included in the same faunalunit as the Farneta and Madonna della Strada ones (Palombo,2009; Magri et al., 2010), cannot give further support to theM. m. vestinus diagnosis because they are crushed and deformed.Accordingly, in view of the small number of preserved skulls ofM. meridionalis, the high variability of elephant skulls and thechance that the peculiar features of the very large skull fromFarneta might be related to the huge dimensions of its tusks, thevalidity of the subspecies has to be considered with caution.
Ongoing research at the Campo di Pile site may contribute tobetter defining the age of the site and the actual composition of the

Fig. 12. Biochronological scheme showing the possible chronological setting of the localities of the L’Aquila Basin yielding elephants remains.
S. Agostini et al. / Quaternary International 276-277 (2012) 42e52 51
accompanying fauna, thus enhancing knowledge of the Quaternaryfauna and the paleoenvironmental evolution of the L’Aquila Basin.
Acknowledgements
We are grateful to the Editors of this special issue of QuaternaryInternational for inviting our contribution. We would also like tothank D. Cosentino and a second anonymous reviewer for theiruseful comments. We thank S. Caramiello, A. Berardinelli, S. Letta,who prepared the fossil material, and Marco Albano for his help inSEM analysis. The English version was revised by Steve Haire.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.quaint.2012.05.013.
References
Ábelova, M., 2008. Schreger pattern analysis of Mammuthus primigenius tusk:analytical approach and utility. Bulletin of Geosciences 83 (2), 225e232.
Agostini, S., Di Canzio, E., Rossi, M.A., 2001. Abruzzo (Italy): the Plio- PleistoceneProboscidean e Bearing Sites. In: The World of Elephants InternationalCongress, Rome, 2001: pp. 163e166.
Agostini, S., Rossi, M.A., Tallini, M., in press. Geologia e paleontologia del Quaternarionel territorio aquilano. In: Atti del Convegno “Nei dintorni di L’Aquila. Ricerchearcheologiche nel territorio dei Vestini Cismontani prima e dopo il terremoto del6 aprile 2009”, Roma, 12e13 Febbraio 2010, École française de Rome.
Ambrosetti, P., Azzaroli, A., Bonadonna, F.P., Follieri, M., 1972. A scheme of Pleisto-cene chronology for the Tyrrhenian side of central Italy. Bollettino della SocietàGeologica Italiana 91, 169e184.
Azzaroli, A., 1977. Evolutionary patterns of Villafranchian elephants in central Italy.In: Atti dell’Accademia Nazionale dei Lincei, Memorie Classe Scienze Fisiche,vol. 14(8), pp. 149e168.
Bagnaia,R.,D’Epifanio,A., SylosLabbini, S.,1989.Aquila and subequanbasin:anexampleof Quaternary evolution in central Apennines, Italy. Quaternaria Nova 1, 1e23.
Bertini, T., Bosi, C., 1993. La Tettonica quaternaria della Conca di Fossa (L’Aquila). IlQuaternario 6 (2), 293e314.
Bertini, T., Bosi, C., Galadini, F., 1989. La conca di Fossa e S. Demetrio dei Vestini. In:Elementi di tettonica pliocenico quaternaria ed indizi di sismicità olocenica nel-l’Appennino laziale-abruzzese. Guida all’escursione della S.G.I. Esa Grafica, Roma.
Bonanni, T., 1872. La Provincia del 2� Abruzzo ulteriore, con la descrizione fisico-topografica-geologica-mineralogica. Aquila, Grossi.
Bosi, C., Bertini, T., 1970. La geologia della media valle dell’Aterno. Memorie SocietàGeologica Italiana 9, 719e777.
Bosi, C., Messina, P., 1991. Ipotesi di correlazione fra successioni morfo-litostratigrafiche plio-pleistoceniche nell’Appennino laziale-abruzzese. StudiGeologici Camerti v.s. 1991/2, CROP 11, 257e263.

S. Agostini et al. / Quaternary International 276-277 (2012) 42e5252
Cavinato, G.P., Cosentino, D., De Rita, D., Funicello, R., Parotto, M., 1994. Tecto-nicesedimentary evolution of intrapenninic basin and correlations with thevolcano-tectonic activity in Central Italy. Memorie Descrittive Carta Geologicad’Italia XLIX, 63e76.
Coltorti, M., Dramis, F., 2006. Stratigrafia dei depositi continentali. In: Centamore, E.,Crescenti, U., Dramis, F. (Eds.), Note illustrative della Carta Geologica d’Italia allascala 1:50.000, foglio 359 “L’Aquila”. APAT-Regione Abruzzo, Rome, pp. 83e93.
D’Erasmo, G., 1931. L’Elephas Meridionalis nell’Abruzzo e nella Lucania. In: Atti dellaRealeAccademiadelle ScienzefisicheematematichediNapoli, vol.18(8), pp.1e32.
Espinoza, E.O., Mann, M.J., 1993. The history and significance of the Schreger Patternin Proboscidean ivory characterization. Journal of the American Institute forConservation 32, 241e248.
Esu, D., Girotti, O., Kotsakis, T., 1992. Molluschi e vertebrati di alcuni bacini con-tinentali dell’Appennino centrale: indicazioni biostratigrafiche e paleo-ecologiche. Studi Geologici Camerti v.s. 1992/2 CROP 11, 295e299.
Esu, D., Girotti, O., Kotsakis, T., 1993. Palaeobiogeographical observations on Villa-franchian continental molluscs of Italy. Scripta Geologica 2, 101e119.
Ferretti, M.P., 1999. Mammuthus meridionalis (Mammalia, Proboscidea, Ele-phantidae) from the “Sabbie Gialle’” of Oriolo (Cava La Salita, Faenza, NorthernItaly) and other European late population of southern mammoth. EclogaeGeologicae Helvetiae 92, 503e515.
Ferretti, M.P., 2003a. Structure and evolution of mammoth molar enamel. ActaPalaeontologica Polonica 48 (3), 383e396.
Ferretti, M.P., 2003b. Functional aspects of the enamel evolution in Mammuthus(Proboscidea, Elephantidae). Deinsea 9, 111e116.
Fisher, D.C., Trapani, J., Shoshani, J.,Woodford,M.S.,1998. Schregerangles inMammothand Mastodon tusk dentin. Current Research in the Pleistocene 15, 105e106.
Galli, P., Giaccio, B., Messina, P., 2010. The 2009 central Italy earthquake seenthrough 0.5 Myr-long tectonic history of the L’Aquila fault system. QuaternaryScience Reviews 29, 3768e3789.
GE.MI.NA, 1963. Ligniti e Torbe dell’Italia Continentale. Geomineraria Nazionale,Torino, pp. 1e319.
Guerin, C., Dewolf, Y., Lautridou, J.-P., 2003. Revision of a famous site: Saint-Prest(Chartres, France). Geobios 36 (1), 55e82.
Kotsakis, T., 1988. Biostratigraphy of Plio-Pleistocene Arvicolids (rodents) of Italy.Modern Geology 13, 163e175.
Maccagno, A.M., 1958. Relazione sulla tecnica di scavo, restauro e montaggio del-l’elefante fossile rinvenuto presso l’Aquila, vol. 2. Annuario delle Istituzionid’Alta Cultura dell’Aquila, pp. 1e14.
Maccagno, A.M., 1962. L’Elephas meridionalis Nesti di Contrada “Madonna dellaStrada” Scoppito (L’Aquila). In: Atti Accademia Scienze Fisiche e Naturali,Napoli, vol. 4, pp. 1e129.
Maccagno, A.M., 1965. Nuovi ritrovamenti di resti elefantini nel Villafranchianodella Conca aquilana. Bollettino della Società dei Naturalisti in Napoli 74, 1e22.
Magaldi, D., Tallini, M., 2000. A micromorphological index of soil development forthe Quaternary geology research. Catena 41, 261e276.
Magri, D., Di Rita, F., Palombo, M.R., 2010. An Early Pleistocene interglacial recordfrom an intermontane basin of central Italy (Scoppito, L’Aquila). QuaternaryInternational 225, 106e113.
Maini, P.S., 1956. Sopra una zanna di Elephas Antiquus Italicus rinvenuta a S.EusanioForconese. Annali dell’Istituto Superiore di Scienze e Lettere S.Chiara 6, RealeMonastero S.Chiara, Napoli, pp. 1e12.
Mancini, M., Cavuoto, G., Pandolfi, L., Petronio, C., Salari, L., Sardella, R., 2011.Coupling basin infill history and mammal biochronology in a Pleistocene
intramontane basin: the case of western L’Aquila Basin (central Apennines,Italy). Quaternary International, 1e16. doi:10.1016/jquaint.2011.03.020.
Marini, P.G., 1967. Il lago pleistocenico della conca de L’Aquila. Ed. CET, Lanciano.1e64.
Messina, P., Galli, P., Giaccio, B., Falcucci, E., Galadini, F., Gori, S., Peronace, E.,Scardia, G., Sposato, A., 2010. Evoluzione geologica e tettonica quaternariadell’area epicentrale del terremoto aquilano del 2009 (Bacino di Paganica-Barisciano, Appennino centrale). Abstract GNGTS 2010. Sessione 1.1, 73e74.
Miall, A.D., 1996. The Geology of Fluvial Deposits. In: Sedimentary Facies, BasinAnalysis and Petroleum Geology. Springer-Verlag, Berlin.
Mozzetti, F., 1893. Sulle ossa d’un elefante scoperte nelle Pagliare di Sassa. Mono-grafia inedita. In: Bollettino della Società di Storia Patria Anton LudovicoAntinori negli Abruzzi. Santini Editore, Aquila, pp. 143e170.
Nesti, F., 1825. Sulla nuova specie di elefante fossile del Valdarno all’IllustrissimoSig. Dott. Prof. Ottaviano Targioni Tozzetti (Lettere sopra alcune ossa fossili delValdarno non per anco descritte). Nuovo Giornale di Lettere 11 (24), 195e216.
Obermayer, F., 1881. Beitrag zur kenntnis des zahnbeines vom elefanten, nilpferd,walross und narwal, vol. 1. Verlag des Vereines der Weiner Handels-Akademic.102e113.
Palombo, M.R., Ferretti, M.P., 2005. Elephant fossil record from Italy: knowledge,problems, and perspectives. Quaternary International 126-128, 107e136.
Palombo, M.R., Villa, P., 2001. Schreger lines as support in the Elephantinae iden-tification. In: Cavaretta, G., Gioia, P., Mussi, M., Palombo, M.R. (Eds.), The Worldof Elephants. Consiglio Nazionale Richerche, Roma, pp. 656e660.
Palombo, M.R., Villa, P., 2007. L’utilità dell’analisi delle linee di Schreger nello studiodei Proboscidati. In: Fiore, I., Malerba, G., Chilardi, S. (Eds.), Atti del 3� CongressoInternazionale di Archeozoologia. Collana del Bollettino di Paletnologia Italiana,Roma, Studi di Paletnologia 2 (2005), pp. 35e44.
Palombo, M.R., Mussi, M., Agostini, S., Barbieri, M., Di Canzio, E., Di Rita, F., Fiore, I.,Iacumin, P., Magri, D., Speranza, F., 2010. Human peopling of Italian intra-montane basins. The early Middle Pleistocene site of Pagliare di Sassa (L’Aquila,central Italy). Quaternary International 223-224, 170e178.
Palombo, M.R., 2009. Biochronology of terrestrial mammals and Quaternarysubdivisions: a case study of large mammals from the Italian peninsula. IlQuaternario. Italian Journal of Geosciences 22 (2), 291e306.
Pfretzschener, H.U., 1994. Biomechanik der Schmelzmikrostruktur in dem Back-enzähnen von Grossäugern. Paläontographica, Abteilung A 233 (1e3), 1e88.
Sala, B., Masini, F., 2007. Late Pliocene and Pleistocene small mammal chronology inthe Italian peninsula. Quaternary International 160, 4e16.
Tallini, M., Amoroso, S., Angelino, A., Azzara, R.M., Bergamaschi, F., Bordoni, P.,Cara, F., Cavinato, G.P., Cavuoto, G., Cogliano, R., Cultrera, G., De Caterini, G., DelMonaco, F., Di Eusebio, F., Di Filippo, M., Di Fiore, V., Di Giulio, G., Di Nezza, M.,Fodarella, A., Iavarone, M., Leoni, G., Marsan, P., Milana, G., Monaco, P.,Pucillo, S., Rapolla, A., Riccio, G., Sanò, T., Scotto di vettimo, P., Taddei, B.,Tarallo, D., Totani, G., Zaffiro, P., Bertrand, E., Duval, A., Régnier, J., Marcucci, S.,2011. La Microzonazione sismica delle macroaree, Macroarea 1, L’Aquila centro.In: Castenetto, S., Naso, G. (Eds.), La Microzonazione sismica per la ricostruzionedell’Area Aquilana. 3. Regione Abruzzo-Protezione Civile, pp. 3e60.
Trapani, J., Fisher, D.C., 2003. Discriminating proboscidean taxa using features of theSchreger pattern in tusk dentin. Journal of Archaeological Science 30, 429e438.
Tsoukala, E., Mol, D., Pappa, S., Vlachos, E., van Logchem, W., Vaxevanopoulos, M.,Reumer, J., 2011. Elephas antiquus in Greece: new finds and a reappraisal ofolder material (Mammalia, Proboscidea, Elephantidae). Quaternary Interna-tional 245 (2), 339e349.




















