
Localization of Leptin Receptor-Like Immunoreactivity in the Corticotropes, Somatotropes, and...
6
Localization of Leptin Receptor-Like Immunoreactivity in the Corticotropes, Somatotropes, and Gonadotropes in the Ovine Anterior Pituitary* JAVED IQBAL, SUELI POMPOLO, ROBERT V. CONSIDINE, AND IAIN J. CLARKE Prince Henry’s Institute of Medical Research (J.I., S.P., I.J.C.), Clayton, Victoria 3168, Australia; and Department of Medicine (R.V.C.), Indiana University, Indianapolis, Indiana 46202 ABSTRACT Leptin is a secreted product of the adipocytes that regulates a variety of functions. The presence of the leptin receptor (LR) has been demonstrated in the endocrine and neuroendocrine tissue, but only limited information is available regarding cell-specific expression in the anterior pituitary gland. We have used double-label immunoflu- orescence histochemistry to study the distribution of LR-like immu- noreactivity (LR-ir) in the corticotropes, somatotropes, and gonado- tropes of the ovine anterior pituitary. LR-ir was found in 34% of cells in the pars distalis and 94% of the cells in the pars tuberalis. In the pars distalis, LR-ir was present in 27% of corticotropes, 69% of so- matotropes, and 29% of gonadotropes. In contrast, 90% of the gona- dotropes in the pars tuberalis were immunopositive for LR. There was no alteration in the number of gonadotropes containing LR-ir during the various phases of the estrous cycle (n 5 3/group) in the pars distalis (luteal phase, 36%; follicular phase, 32%; and estrous phase, 32%). In conclusion, we show that, in the pars distalis, LR-ir is ex- pressed to a greater extent in the somatotropes than in the gonado- tropes or corticotropes. This is in accordance with the documented effects of leptin on pituitary GH secretion. The differential expression of LR-ir between the gonadotropes of the pars distalis and pars tu- beralis probably reflects the different phenotypes of the cells in these two regions. Lower levels of LR-ir expression in gonadotropes and corticotropes of the pars distalis may suggest that leptin does not substantially influence these particular cells, at least in this species. (Endocrinology 141: 1515–1520, 2000) L EPTIN is a hormone secreted from the adipocytes (1) that is involved in energy homeostasis (2– 4) and in regu- lation of neuroendocrine (5– 8) and reproductive functions (9 –12). A deficiency in leptin or the leptin receptor (LR) causes disturbances in feeding behavior (13) and reproduc- tive function (14) in various rodent models, and these effects can be corrected by exogenous leptin administration. Leptin can prevent the reduction in gonadotropin secretion that occurs with starvation (5, 15) and also appears to regulate GH secretion (7, 8, 16, 17). In rodents, leptin may also regulate the hypothalamo-pituitary-adrenal (HPA) axis (18) and the hy- pothalamo-pituitary-thyroid axis (19), but this is not the case in normally fed sheep (20). In various species, imunohistochemical and in situ hybrid- ization studies have demonstrated LR protein and its mes- senger RNA (mRNA) in several tissues including the hypo- thalamus and pituitary gland (21–25). A major site of leptin action is thought to be the hypothalamus where LR expres- sion is seen in various neuropeptidergic neurons (25–27). Apart from a role in the regulation of appetite, mechanisms whereby leptin might influence neuroendocrine functions are not yet clear. Leptin could modulate the expression of neuropeptides that regulate the neuroendocrine system within the hypothalamus, but could also act directly on the anterior pituitary (6, 16, 18, 19, 21). In accordance with the latter possibility, LR mRNA has been localized to GH- secreting and PRL-secreting human pituitary adenomas (21). Although RT-PCR analysis showed presence of LR in the ovine pars distalis, there is no detailed information on lo- calization to particular cell types (24). In fact, there have been no immunohistochemical studies in any species to indicate the presence of LR protein in the anterior pituitary gland. In sheep, an alteration in adiposity generally affects the secretion of GH (28 –30) and gonadotropins (29, 31–36) with no effect on the plasma levels of PRL (29), TSH, or cortisol (36). It seems possible, therefore, that the gonadotropes and/or somatotropes of the pituitary gland are potential targets for leptin. The HPA axis is up-regulated by under- nutrition in rodents (5), but this is not the case in sheep (36, 37). Accordingly, we sought to determine the extent to which somatotropes and gonadotropes express LR-ir in the female sheep pituitary. We also studied, for comparison, LR-ir ex- pression in corticotropes, since the available data suggest that the HPA axis is not regulated by alteration in adiposity, or by leptin administration, in this species. Materials and Methods Tissue collection All procedures were carried out with the prior approval of the Animal Experimentation Ethics Committee of Monash University and Victoria Institute of Animal Science. Study 1. Four pituitaries (including the median eminence/pars tuberalis) were obtained from adult Corriedale ewes of similar age and weight, during the luteal phase of the estrous cycle. The tissue was collected as previously reported (38). Briefly, the sheep were administered 25,000 IU of heparin iv and were killed 5 min later by an overdose of sodium pentobarbital (2.5 ml/kg body weight, iv) (Lethabarb; May & Baker Pty Received September 20, 1999. Address all correspondence and requests for reprints to: Professor Iain J. Clarke, Prince Henry’s Institute of Medical Research, PO Box 5152, Clayton, Victoria 3168, Australia. E-mail: iain.clarke@med. monash. edu.au. * This work was supported by the National Health and Medical Research Council, Australia. 0013-7227/00/$03.00/0 Vol. 141, No. 4 Endocrinology Printed in U.S.A. Copyright © 2000 by The Endocrine Society 1515
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Localization of Leptin Receptor-Like Immunoreactivity in the Corticotropes, Somatotropes, and...

Localization of Leptin Receptor-Like Immunoreactivityin the Corticotropes, Somatotropes, and Gonadotropesin the Ovine Anterior Pituitary*
JAVED IQBAL, SUELI POMPOLO, ROBERT V. CONSIDINE, AND IAIN J. CLARKE
Prince Henry’s Institute of Medical Research (J.I., S.P., I.J.C.), Clayton, Victoria 3168, Australia; andDepartment of Medicine (R.V.C.), Indiana University, Indianapolis, Indiana 46202
ABSTRACTLeptin is a secreted product of the adipocytes that regulates a
variety of functions. The presence of the leptin receptor (LR) has beendemonstrated in the endocrine and neuroendocrine tissue, but onlylimited information is available regarding cell-specific expression inthe anterior pituitary gland. We have used double-label immunoflu-orescence histochemistry to study the distribution of LR-like immu-noreactivity (LR-ir) in the corticotropes, somatotropes, and gonado-tropes of the ovine anterior pituitary. LR-ir was found in 34% of cellsin the pars distalis and 94% of the cells in the pars tuberalis. In thepars distalis, LR-ir was present in 27% of corticotropes, 69% of so-matotropes, and 29% of gonadotropes. In contrast, 90% of the gona-dotropes in the pars tuberalis were immunopositive for LR. There was
no alteration in the number of gonadotropes containing LR-ir duringthe various phases of the estrous cycle (n 5 3/group) in the parsdistalis (luteal phase, 36%; follicular phase, 32%; and estrous phase,32%). In conclusion, we show that, in the pars distalis, LR-ir is ex-pressed to a greater extent in the somatotropes than in the gonado-tropes or corticotropes. This is in accordance with the documentedeffects of leptin on pituitary GH secretion. The differential expressionof LR-ir between the gonadotropes of the pars distalis and pars tu-beralis probably reflects the different phenotypes of the cells in thesetwo regions. Lower levels of LR-ir expression in gonadotropes andcorticotropes of the pars distalis may suggest that leptin does notsubstantially influence these particular cells, at least in this species.(Endocrinology 141: 1515–1520, 2000)
LEPTIN is a hormone secreted from the adipocytes (1) thatis involved in energy homeostasis (2–4) and in regu-
lation of neuroendocrine (5–8) and reproductive functions(9–12). A deficiency in leptin or the leptin receptor (LR)causes disturbances in feeding behavior (13) and reproduc-tive function (14) in various rodent models, and these effectscan be corrected by exogenous leptin administration. Leptincan prevent the reduction in gonadotropin secretion thatoccurs with starvation (5, 15) and also appears to regulate GHsecretion (7, 8, 16, 17). In rodents, leptin may also regulate thehypothalamo-pituitary-adrenal (HPA) axis (18) and the hy-pothalamo-pituitary-thyroid axis (19), but this is not the casein normally fed sheep (20).
In various species, imunohistochemical and in situ hybrid-ization studies have demonstrated LR protein and its mes-senger RNA (mRNA) in several tissues including the hypo-thalamus and pituitary gland (21–25). A major site of leptinaction is thought to be the hypothalamus where LR expres-sion is seen in various neuropeptidergic neurons (25–27).Apart from a role in the regulation of appetite, mechanismswhereby leptin might influence neuroendocrine functionsare not yet clear. Leptin could modulate the expression ofneuropeptides that regulate the neuroendocrine systemwithin the hypothalamus, but could also act directly on theanterior pituitary (6, 16, 18, 19, 21). In accordance with the
latter possibility, LR mRNA has been localized to GH-secreting and PRL-secreting human pituitary adenomas (21).Although RT-PCR analysis showed presence of LR in theovine pars distalis, there is no detailed information on lo-calization to particular cell types (24). In fact, there have beenno immunohistochemical studies in any species to indicatethe presence of LR protein in the anterior pituitary gland.
In sheep, an alteration in adiposity generally affects thesecretion of GH (28–30) and gonadotropins (29, 31–36) withno effect on the plasma levels of PRL (29), TSH, or cortisol(36). It seems possible, therefore, that the gonadotropesand/or somatotropes of the pituitary gland are potentialtargets for leptin. The HPA axis is up-regulated by under-nutrition in rodents (5), but this is not the case in sheep (36,37). Accordingly, we sought to determine the extent to whichsomatotropes and gonadotropes express LR-ir in the femalesheep pituitary. We also studied, for comparison, LR-ir ex-pression in corticotropes, since the available data suggestthat the HPA axis is not regulated by alteration in adiposity,or by leptin administration, in this species.
Materials and MethodsTissue collection
All procedures were carried out with the prior approval of the AnimalExperimentation Ethics Committee of Monash University and VictoriaInstitute of Animal Science.
Study 1. Four pituitaries (including the median eminence/pars tuberalis)were obtained from adult Corriedale ewes of similar age and weight,during the luteal phase of the estrous cycle. The tissue was collected aspreviously reported (38). Briefly, the sheep were administered 25,000 IUof heparin iv and were killed 5 min later by an overdose of sodiumpentobarbital (2.5 ml/kg body weight, iv) (Lethabarb; May & Baker Pty
Received September 20, 1999.Address all correspondence and requests for reprints to: Professor
Iain J. Clarke, Prince Henry’s Institute of Medical Research, PO Box 5152,Clayton, Victoria 3168, Australia. E-mail: iain.clarke@med. monash.edu.au.
* This work was supported by the National Health and MedicalResearch Council, Australia.
0013-7227/00/$03.00/0 Vol. 141, No. 4Endocrinology Printed in U.S.A.Copyright © 2000 by The Endocrine Society
1515

Ltd, Melbourne, Australia). The heads were flushed with 2 liters ofheparinized (12,500 IU/liter) 0.9% saline via carotid arteries followed by2 liters of 4% paraformaldehyde in 0.1 m phosphate buffer (PB, pH 7.4)and 1.5 liters of 4% paraformaldehyde in PB containing 20% sucrose. Thetissues were removed, transferred to a solution containing 4% parafor-maldehyde plus 30% sucrose, and left at 4 C for 5 days. The tissues werethen frozen on powdered dry ice and kept at 220 C until used. Sectionsof 20 mm were cut on a cryostat (CM 1850, Leica Corp., Deerfield, IL) at220 C and thaw-mounted onto gelatin-chrom-coated slides, left over-night at room temperature, and stored at 220 C until processed forimmunohistochemistry. Sections (20 mm) of median eminence/pars tu-beralis tissue were similarly obtained and stored in cryoprotectant so-lution at 220 C until processed for free-floating immunohistochemistry.
Study 2. Analysis of the tissue sections in Study 1 revealed that 90% ofthe gonadotropes expressed LR-ir in the pars tuberalis, whereas only30% of the gonadotropes were double labeled in the pars distalis. Toinvestigate whether expression of LR-ir in gonadotropes varied acrossthe estrous cycle, additional pituitaries and median eminence/pars tu-beralis tissues were collected (as above) during the luteal, follicular, andsurge phases of the estrous cycle (n 5 3/group) and examined for LR-irexpression.
Antibodies
We used a rabbit polyclonal LR antibody raised against a peptidecorresponding to amino acids 874–886 of LR (39), which is common toboth long (OB-Rb) and short (OB-Ra) isoforms of LR. The characteriza-tion and use of the antibody for localization of LR have been reportedpreviously (23, 39). The specificity of LR-ir in the sheep anterior pituitarywas tested by preabsorption of LR antiserum with the peptide used forimmunization at a final concentration of 5, 10, and 20 mg/ml. The useand specificity of GH and LHb and ACTH antibodies for the ovineanterior pituitary have been described previously (38).
Immunohistochemistry
Slide-mounted pituitary tissue sections and free-floating median em-inence/pars tuberalis tissue sections were processed for immunofluo-rescence double labeling according to previously reported protocols (38)with some modification. Briefly, the sections were washed in 0.05 m PBSto remove the fixative and treated with 1% sodium borohydride (Sigma-Aldrich Corp., St. Louis, MO) in 0.01 m PB for 20 min. After washing,the sections were incubated in blocking solution (10% normal goatserum, 1% BSA, 0.1% Triton X-100 in PB) for 30 min and then with LRantiserum (1:500) overnight at room temperature. After washing in PBS,the sections were incubated with Texas Red-conjugated goat antirabbitIgG (1:250, Molecular Probes, Inc., Eugene, OR) in PB for 2 h, washed,and then incubated in blocking solution containing 10% normal donkeyserum for 30 min. Sections were then incubated either with mousemonoclonal anti-ACTH (1:50, Novacastro, Newcastle on Tyne, UK),anti-LHb (1:2500, kindly donated by Dr. J. F. Roser, University of Cal-ifornia, Davis, CA). or anti-GH (1:10, kindly donated by Dr. M. Brandon,University of Melbourne, Australia). After washing, the sections werethen incubated in biotinylated horse antimouse IgG (1:800, DAKO Corp.,Carpinteria, CA) for 1 h, washed, and then incubated with fluoresceinthiocyanate (FITC)-conjugated avidin (1:500, Pierce Chemical Co., Rock-ford, IL) for 1 h and washed again. For the purpose of presentingrepresentative photomicrographs, one section from both pars distalisand pars tuberalis of each animal was also stained with 1% Sudan BlackB (Sigma-Aldrich Corp.) for 10 min to block autofluorescence (40). Inaddition, and to show that some cells did not express LR-ir, represen-tative sections processed for LR-ir only were briefly counterstained with49,69 diamidine-2-phenylindole dihydrochloride (DAPI) for 10 min (1:10,Molecular Probes, Inc., Eugene, OR). This allowed clear identification ofcells that either did or did not contain LR-ir. The slide-mounted pituitarysections were then cover-slipped with anti-fade medium (DAKO Corp.).Free-floating median eminence/pars tuberalis tissue sections weremounted onto gel-chrome-subbed slides and cover slipped with antifademedium (DAKO Corp.). Slides were stored at 4 C in the dark untilanalyzed. In all cases, negative controls (omission of the primary anti-serum) were run simultaneously to confirm the specificity of the im-munostaining and lack of bleed through of the fluorescent markers.
Tissue analysis
Tissue analysis was carried out using a BMX 50 microscope (OlympusCorp., Lake Success, NY) equipped with a mercury light source andTexas Red and FITC band filter systems to visualize the red and greenimmunofluorescence, respectively. The nuclear staining with DAPI wasassessed using the UV light. The expression of LR-ir within each indi-vidual cell labeled either for ACTH, GH, or LHb was visualized byswitching between the Texas Red and FITC filter systems. To estimatethe percentage of cells expressing LR-ir in the pars distalis, 50 cells werevisually counted in each of the five randomly selected area of a repre-sentative section for each animal. Only cells that had discernable nucleiwere counted. There was a clear difference between cells that stainedwith either fluorophor and background, with no cells showing equivocalstaining.
To obtain an estimation of the level of expression of LR-ir in soma-totropes and gonadotropes, 50 cells were visually counted in each of 5randomly selected areas in a representative section for each animal.Since ACTH immunoreactive cells are not as abundant, 10 cells werecounted in each of 10 randomly selected areas. All observations weremade by a single investigator, and each counted cell had a clearlydefined nucleus.
Statistical analysis
The data were not corrected for double-labeling cell count (41). Afterquantification of LR-immunoreactive cells, the data were subjected tostatistical analysis using one-way ANOVA. Our results provide a rel-ative number (mean % 6 sem) of each cell type that is immunopositivefor LR and not the actual number of cells in the pars distalis or parstuberalis.
ResultsStudy 1
No specific LR-ir was seen after the preabsorption of theLR antiserum with antigen peptide or after the omission ofprimary antiserum (Fig. 1). All autofluorescence was com-pletely blocked after counterstaining of the tissue with SudanBlack B (data not shown).
Double labeling of pituitary sections with DAPI and LRantiserum is shown in Fig. 2, A–D. In the pars distalis, LR-irwas found in 34.5 6 1.71% of the cells whereas 94.4 6 1.32%of the cells in the pars tuberalis expressed LR-ir (P , 0.01).
The pattern of distribution of ACTH-, GH-, and LH-se-creting cells was essentially similar to that reported previ-ously (38). LR-ir was observed in some corticotropes, but notothers (Fig. 2, E–H). Similarly, some representative examplesof LR-ir-positive and LR-ir-negative somatotropes (Fig. 2,I–L) and gonadotropes (Fig. 2, M–T) are shown.
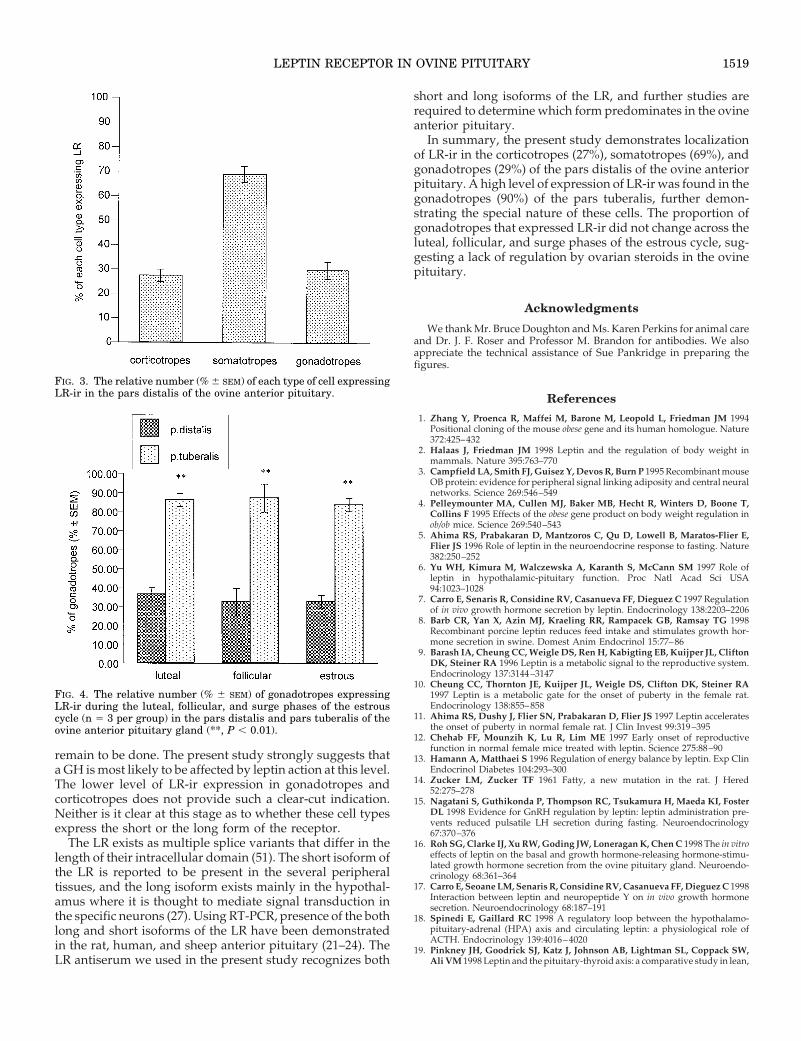
Expression of LR-ir was colocalized in 27.5 6 2.5% ofcorticotropes, 69.0 6 3.3% of somatotropes, and 29.3 6 2.5%of gonadotropes in the pars distalis (Fig. 3). In the parstuberalis, 90.0 6 1.5% of gonadotropes contained LR-ir (Fig.4)(P , 0.01 compared with pars distalis). No somatotropeswere seen in the pars tuberalis, but a few corticotropes wereseen as reported by Clarke et al. (42); these were not immu-nopositive for LR.
Study 2
There was no difference across the estrous cycle in theproportion of gonadotropes that expressed LR-ir in either thepars distalis or pars tuberalis (Fig. 4). As in Study 1, thenumber of gonadotropes that were immunopositive for LRwere significantly (P , 0.01) greater in the pars tuberalis thanin the pars distalis (Fig. 4).
1516 LEPTIN RECEPTOR IN OVINE PITUITARY Endo • 2000Vol 141 • No 4

Discussion
This study demonstrates expression of LR-ir in the corti-cotropes, somatotropes, and gonadotropes in the ovine an-terior pituitary gland. Since no specific immunostaining wasobserved after control procedures and counterstaining withSudan Black B blocked all autofluorescence, the LR-ir ap-pears to be specific. Earlier work has indicated expression ofLR mRNA in the pituitary gland of the sheep (24) and other
species (21, 22). Shimon et al. (21) reported that mRNA for LRmay be found in GH-secreting and PRL-secreting humanpituitary adenomas using RT-PCR, but there are no otherstudies on localization to particular cells in normal pituitarycells.
There is a high level of expression of LR-ir in somatotropes(69%) of the ovine pars distalis and a lower level of expres-sion of LR-ir in corticotropes (27%) and gonadotropes (29%).Since only one third of the gonadotropes were found toexpress LR in the pars distalis of sheep sampled during theluteal phase (Study 1), it remained possible that expressionof LR-ir was regulated by steroids and varied across theestrous cycle. Accordingly, we examined expression of LR-irin gonadotropes across the luteal, follicular, and estrousphases of the estrous cycle but found no cyclic variation. Thisstrongly suggests that the ovarian steroids do not regulateexpression of LR-ir in the gonadotropes.
Only 30% of the gonadotropes express LR-ir in the parsdistalis compared with a high (90%) level of expression in thepars tuberalis gonadotropes. The cells of the pars tuberalisare different in embryonic origin and gene expression, andregulation of the cells is different in the two tissues (43). Thisis exemplified by ontogenetic studies of the distribution ofthe transcription factor PIT-1 in the rat (44, 45) and studieson the expression of arylalkylamine N-acetyltransferase inthe ovine anterior pituitary gland (46). It has also been knownfor some time that the pars tuberalis contains a very highpercentage of gonadotropes (47), but other pituitary celltypes are also present (42, 48). In the ovine, the pars tuberaliscells express a high level of melatonin receptors, whereas thepars distalis cells do not (49, 50). Further, it has been reportedthat variable endocrine conditions do not affect the numberand distribution of gonadotropes in the pars tuberalis of thesheep (48), and this also appears to be true for the parsdistalis. The high level of LR-ir expression in the pars tube-ralis compared with that of the pars distalis could be due tothe close proximity of the former to the neural componentsof the median eminence (hypothalamic neuronal endings)and the primary plexus of the hypophyseal portal system.Thus, it seems most likely that cells of pars tuberalis aredifferentially regulated to those of pars distalis. The resultsof the present study provide further evidence of the differentphenotypes of gonadotropes in the pars tuberalis and parsdistalis of the ovine pituitary.
Studies in vitro and in vivo have suggested that leptin mayact directly on the pituitary gland to affect the secretion ofGH (6–8, 16, 17, 21) and gonadotropins (6, 15), but PRLsecretion is not altered (21). It is well established that leptinregulates the HPA axis in rodents (5, 6, 18) and may affectTSH levels in humans (19), but this is not the case in thesheep, at least with central administration of leptin (20).Collectively, these studies are in accordance with observa-tions that an alteration in adiposity influences the GH (20,28–30) and gonadotropin (LH and FSH) secretion (31–36) insheep, with no effect on PRL (29). Whereas the effect of leptinto regulate GH could reflect, in part, altered secretion ofsomatostatin (30), there could also be direct pituitary regu-lation as indicated by the in vitro studies (see above). Ap-propriate in vivo studies to demonstrate direct pituitary ac-tion of leptin in relation to GH and gonadotropin secretion
FIG. 1. Photomicrographs demonstrating cellular staining for LR-irin the pars distalis. Panel A shows positive staining; panel B showsthe lack of staining after preabsorption of the LR antiserum with theantigen peptide; and panel C shows lack of staining after the omissionof the primary antiserum. Scale bar, 10 mm.
LEPTIN RECEPTOR IN OVINE PITUITARY 1517
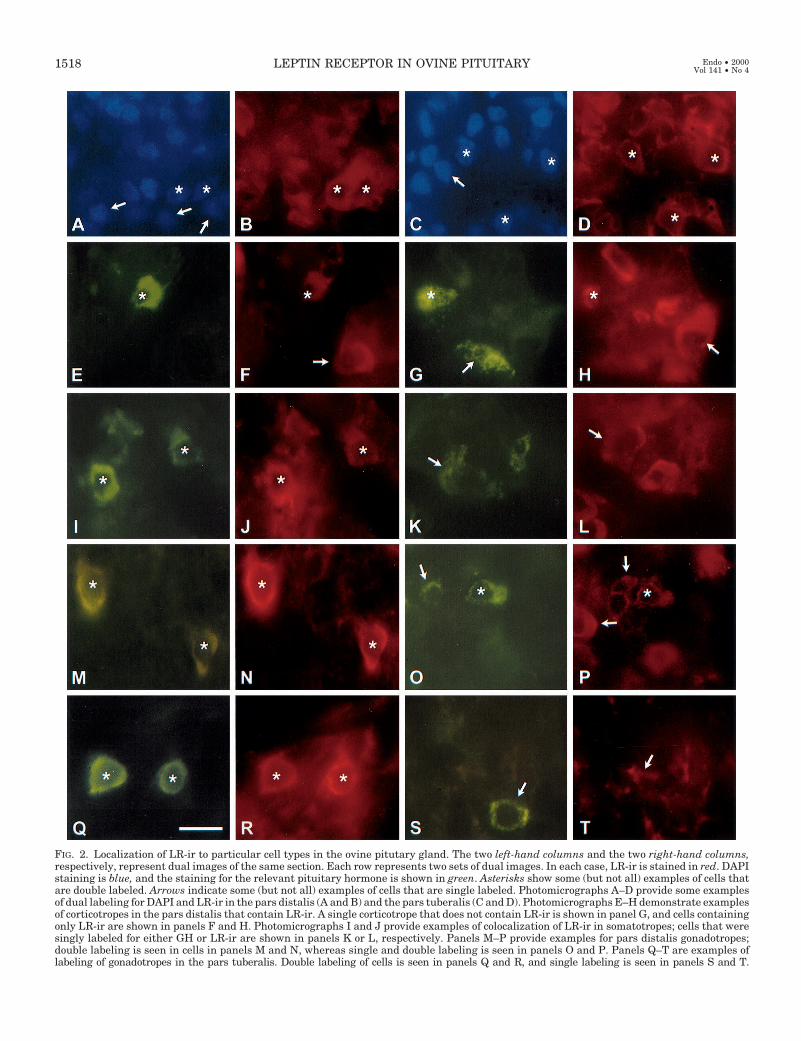
FIG. 2. Localization of LR-ir to particular cell types in the ovine pitutary gland. The two left-hand columns and the two right-hand columns,respectively, represent dual images of the same section. Each row represents two sets of dual images. In each case, LR-ir is stained in red. DAPIstaining is blue, and the staining for the relevant pituitary hormone is shown in green. Asterisks show some (but not all) examples of cells thatare double labeled. Arrows indicate some (but not all) examples of cells that are single labeled. Photomicrographs A–D provide some examplesof dual labeling for DAPI and LR-ir in the pars distalis (A and B) and the pars tuberalis (C and D). Photomicrographs E–H demonstrate examplesof corticotropes in the pars distalis that contain LR-ir. A single corticotrope that does not contain LR-ir is shown in panel G, and cells containingonly LR-ir are shown in panels F and H. Photomicrographs I and J provide examples of colocalization of LR-ir in somatotropes; cells that weresingly labeled for either GH or LR-ir are shown in panels K or L, respectively. Panels M–P provide examples for pars distalis gonadotropes;double labeling is seen in cells in panels M and N, whereas single and double labeling is seen in panels O and P. Panels Q–T are examples oflabeling of gonadotropes in the pars tuberalis. Double labeling of cells is seen in panels Q and R, and single labeling is seen in panels S and T.
1518 LEPTIN RECEPTOR IN OVINE PITUITARY Endo • 2000Vol 141 • No 4
remain to be done. The present study strongly suggests thata GH is most likely to be affected by leptin action at this level.The lower level of LR-ir expression in gonadotropes andcorticotropes does not provide such a clear-cut indication.Neither is it clear at this stage as to whether these cell typesexpress the short or the long form of the receptor.
The LR exists as multiple splice variants that differ in thelength of their intracellular domain (51). The short isoform ofthe LR is reported to be present in the several peripheraltissues, and the long isoform exists mainly in the hypothal-amus where it is thought to mediate signal transduction inthe specific neurons (27). Using RT-PCR, presence of the bothlong and short isoforms of the LR have been demonstratedin the rat, human, and sheep anterior pituitary (21–24). TheLR antiserum we used in the present study recognizes both
short and long isoforms of the LR, and further studies arerequired to determine which form predominates in the ovineanterior pituitary.
In summary, the present study demonstrates localizationof LR-ir in the corticotropes (27%), somatotropes (69%), andgonadotropes (29%) of the pars distalis of the ovine anteriorpituitary. A high level of expression of LR-ir was found in thegonadotropes (90%) of the pars tuberalis, further demon-strating the special nature of these cells. The proportion ofgonadotropes that expressed LR-ir did not change across theluteal, follicular, and surge phases of the estrous cycle, sug-gesting a lack of regulation by ovarian steroids in the ovinepituitary.
Acknowledgments
We thank Mr. Bruce Doughton and Ms. Karen Perkins for animal careand Dr. J. F. Roser and Professor M. Brandon for antibodies. We alsoappreciate the technical assistance of Sue Pankridge in preparing thefigures.
References
1. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM 1994Positional cloning of the mouse obese gene and its human homologue. Nature372:425–432
2. Halaas J, Friedman JM 1998 Leptin and the regulation of body weight inmammals. Nature 395:763–770
3. Campfield LA, Smith FJ, Guisez Y, Devos R, Burn P 1995 Recombinant mouseOB protein: evidence for peripheral signal linking adiposity and central neuralnetworks. Science 269:546–549
4. Pelleymounter MA, Cullen MJ, Baker MB, Hecht R, Winters D, Boone T,Collins F 1995 Effects of the obese gene product on body weight regulation inob/ob mice. Science 269:540–543
5. Ahima RS, Prabakaran D, Mantzoros C, Qu D, Lowell B, Maratos-Flier E,Flier JS 1996 Role of leptin in the neuroendocrine response to fasting. Nature382:250–252
6. Yu WH, Kimura M, Walczewska A, Karanth S, McCann SM 1997 Role ofleptin in hypothalamic-pituitary function. Proc Natl Acad Sci USA94:1023–1028
7. Carro E, Senaris R, Considine RV, Casanueva FF, Dieguez C 1997 Regulationof in vivo growth hormone secretion by leptin. Endocrinology 138:2203–2206
8. Barb CR, Yan X, Azin MJ, Kraeling RR, Rampacek GB, Ramsay TG 1998Recombinant porcine leptin reduces feed intake and stimulates growth hor-mone secretion in swine. Domest Anim Endocrinol 15:77–86
9. Barash IA, Cheung CC, Weigle DS, Ren H, Kabigting EB, Kuijper JL, CliftonDK, Steiner RA 1996 Leptin is a metabolic signal to the reproductive system.Endocrinology 137:3144–3147
10. Cheung CC, Thornton JE, Kuijper JL, Weigle DS, Clifton DK, Steiner RA1997 Leptin is a metabolic gate for the onset of puberty in the female rat.Endocrinology 138:855–858
11. Ahima RS, Dushy J, Flier SN, Prabakaran D, Flier JS 1997 Leptin acceleratesthe onset of puberty in normal female rat. J Clin Invest 99:319–395
12. Chehab FF, Mounzih K, Lu R, Lim ME 1997 Early onset of reproductivefunction in normal female mice treated with leptin. Science 275:88–90
13. Hamann A, Matthaei S 1996 Regulation of energy balance by leptin. Exp ClinEndocrinol Diabetes 104:293–300
14. Zucker LM, Zucker TF 1961 Fatty, a new mutation in the rat. J Hered52:275–278
15. Nagatani S, Guthikonda P, Thompson RC, Tsukamura H, Maeda KI, FosterDL 1998 Evidence for GnRH regulation by leptin: leptin administration pre-vents reduced pulsatile LH secretion during fasting. Neuroendocrinology67:370–376
16. Roh SG, Clarke IJ, Xu RW, Goding JW, Loneragan K, Chen C 1998 The in vitroeffects of leptin on the basal and growth hormone-releasing hormone-stimu-lated growth hormone secretion from the ovine pituitary gland. Neuroendo-crinology 68:361–364
17. Carro E, Seoane LM, Senaris R, Considine RV, Casanueva FF, Dieguez C 1998Interaction between leptin and neuropeptide Y on in vivo growth hormonesecretion. Neuroendocrinology 68:187–191
18. Spinedi E, Gaillard RC 1998 A regulatory loop between the hypothalamo-pituitary-adrenal (HPA) axis and circulating leptin: a physiological role ofACTH. Endocrinology 139:4016–4020
19. Pinkney JH, Goodrick SJ, Katz J, Johnson AB, Lightman SL, Coppack SW,Ali VM 1998 Leptin and the pituitary-thyroid axis: a comparative study in lean,
FIG. 3. The relative number (% 6 SEM) of each type of cell expressingLR-ir in the pars distalis of the ovine anterior pituitary.
FIG. 4. The relative number (% 6 SEM) of gonadotropes expressingLR-ir during the luteal, follicular, and surge phases of the estrouscycle (n 5 3 per group) in the pars distalis and pars tuberalis of theovine anterior pituitary gland (**, P , 0.01).
LEPTIN RECEPTOR IN OVINE PITUITARY 1519

obese, hypothyroid and hyperthyroid subjects. Clin Endocrinol (Oxf)49:583–588
20. Henry BA, Goding JW, Alexander WS, Tilbrook AJ, Canny BJ, Dunshea F,Rao A, Mansell A, Clarke IJ 1999 Central administration of leptin to ovari-ectomized ewes inhibits food intake without affecting the secretion of hor-mones from the pituitary gland: evidence for a dissociation of effects onappetite and neuroendocrine function. Endocrinology 140:1175–1182
21. Shimon I, Yan X, Maggofin DA, Friedman TC, Melmed S 1998 Intact leptinreceptor is selectively expressed in human fetal pituitary and pituitary ade-nomas and signals human fetal pituitary growth hormone secretion. J ClinEndocrinol Metab 83:4059–4064
22. Zamorano PL, Mahesh VB, De Sevilla LM, Chorich LP, Bhat GK, Brann DW1997 Expression and localization of the leptin receptor in the endocrine andneuroendocrine tissues of the rat. Neuroendocrinology 65:223–228
23. Couce ME, Burguera B, Parisi JE, Jensen MD, Lloyd RV 1997 Localization ofleptin receptor in the human brain. Neuroendocrinology 66:145–150
24. Dyer CJ, Simmons JM, Matteri, RL, Keisler DH 1997 Leptin receptor mRNAis expressed in ewe anterior pituitary and adipose tissues and is differentiallyexpressed in hypothalamic regions of well fed and feed restricted ewe. DomestAnim Endocrinol 14:119–128
25. Hakansson ML, Brown H, Gilhardi N, Skoda RC, Miester B 1998 Leptinreceptor immunoreactivity in chemically defined target neurones of the hy-pothalamus. J Neurosci 18:559–572
26. McCowen KC, Chow JC, Smith RJ 1998 Leptin signaling in the hypothalamusof normal rats in vivo. Endocrinology 139:4442–4447
27. Glaum SR, Hara M, Bindokas VP, Lee CC, Polonksy KS, Bell GI, Miller RJ1996 Leptin, the obese gene product, rapidly modulates synaptic transmissionin the hypothalamus. Mol Pharmacol 50:230–235
28. Barker-Gibb ML, Clarke IJ 1996 Increased galanin and neuropeptide Y im-munoreactivity within the hypothalamus of ovariectomized ewes following aprolonged period of reduced body weight correlates with changes in plasmagrowth hormone levels but not gonadotropin levels. Neuroendocrinology64:194–207
29. Thomas GB, Mercer JE, Karalis T, Rao A, Cummins JT, Clarke IJ 1990 Effectsof restricted feeding on the concentration of growth hormone (GH) gonado-tropin and prolactin (PRL) in plasma and on the amounts of messenger ri-bonucleic acid concentration for GH, gonadotropin subunits, and PRL in thepituitary glands of adult ovariectomized ewes. Endocrinology 126:1361–1367
30. Thomas GB, Cummins JT, Francis H. Sudbury AW, McCloud PI, Clarke IJ1991 Effect of restricted feeding on the relationship between hypophysealportal concentrations of growth hormone (GH)-releasing factor and soma-tostatin, and jugular concentrations of GH in ovariectomized ewes. Endocri-nology 128:1151–1158
31. Adam CL, Findlay PA, Kyle CE, Young P, Mercer JG 1997 Effects of chronicfood restriction on pulsatile luteinizing hormone secretion and hypothalamicneuropeptide Y gene expression in castrate male sheep. J Endocrinol152:329–337
32. Pomares CC, Galloway DB, Holmes JHG, Clarke IJ, Tilbrook AJ 1995 Lupinand cowpea supplements for growth, wool production and reproduction inrams. Aust J Exp Agri 35:447–452
33. McShane TM, Petersen SL, McCrone S, Keisler DH 1993 Influence of foodrestriction on neuropeptide-Y, proopiomelanocortin, and luteinizing hor-mone-releasing hormone gene expression in sheep hypothalami. Biol Reprod49:831–839
34. Tatman WR, Judkins MB, Dunn TG, Moss GE 1989 Luteinizing hormone innutrient-restricted ovariectomized ewes. J Anim Sci 68:1097–1102
35. Rhind SM, McMillen S, McKelvey WAC, Rodriguez-Herrejon FF, McNeillyAS 1989 Effects of the body condition of ewes on the secretion of LH and FSHand the pituitary response to gonadotrophin-releasing hormone. J Endocrinol120:497–502
36. Henry BA, Tilbrook AJ, Dunshea FR, Rao A, Blache D, Martin GB, ClarkeIJ 2000 Long-term alterations in adiposity affect the expression of melanin-concentrating hormone and enkephalin but not proopiomelanocortin in thehypothalamus of ovariectomized ewes. Endocrinology 141:1506–1514
37. Henry BA, Dunshea FR, Canny BJ ,Tilbrook AJ, Rao A, Clarke IJ, Expressionof appetite regulating peptides in the hypothalamus of fat and thin sheep inrelation to endocrinology and metabolic parameters. Program of the 81stMeeting of The Endocrine Society, San Diego, CA, 1999, p 375 (AbstractP2–449)
38. Thomas SG, Takahashi M, Neill JD, Clarke IJ 1998 Components of theneuronal exocytotic machinery in the anterior pituitary of the ovariectomizedewe and the effects of estrogen in gonadotropes as studied with confocalmicroscopy. Neuroendocrinology 67:244–259
39. Cao GY, Considine RV, Lynn RB 1997 Leptin in the adrenal medulla of therat. Am J Physiol 273:448–452
40. Romijn HJ, Van Uum JFM, Breedijk I, Emmering J, Radu I, Pool CW 1999Double immunolabeling of neuropeptides in the human hypothalamus asanalyzed by confocal laser scanning fluorescence microscopy. J HistochemCytochem 47:229–235
41. Abercrombie M 1946 Estimation of nuclear population from microtome sec-tion. Anat Rec 94:239–247
42. Clarke IJ, Burman J, Perry RA, Prince KM, Horton RJE 1989 b-Endorphin inhypophyseal portal blood. In: Dyer R, Bicknell J (eds) Brain Opioid Systemsin Reproduction. Oxford University Press, Oxford, UK, pp 135–148
43. Wittkowski WH, Schulze-Bonhage AH, Bockers TM 1992 The pars tuberalisof the hypophysis: a modulator of the pars distalis. Acta Endocrinol (Copenh)126:285–290
44. Lin SC, Li S, Drolet DW, Rosenfeld MG 1994 Pituitary ontogeny of the Snelldwarf mouse reveals Pit-1-independent and Pit-1-dependent origins of thethryotropes. Development 120:515–522
45. Sakai T, Sakamoto S, Ijima K, Matsubara K, Kato Y, Inoue K 1999 Charac-terization of TSH-positive cells in foetal rat pars tuberalis that fail to expressPit-1 factor and thyroid hormone b2 receptors. J Neuroendocrinol 11:187–193
46. Fleming JV, Barrett P, Coon SL, Klein DC, Morgan PJ 1999 Ovine arylalky-lamine N-acetyltransferase in the pineal and pituitary glands: differences infunction and regulation. Endocrinology 140:972–978
47. Mason WT, Waring DW 1986 Patch clamp recording of single ion channelactivation by gonadotrophin-releasing hormone in ovine pituitary gonado-trophs. Neuroendocrinology 43:205–219
48. Skinner DC, Robinson JE 1996 The pars tuberalis of the ewe: no effect ofseason or ovariectomy on the distribution, density or presence of immuno-reactive cells. Cell Tissue Res 284:117–123
49. Morgan PJ, Williams LM, Davidson G, Lawson W, Howell E 1989 Melatoninreceptors in the ovine pars tuberalis: characterization and autoradiographicallocalization. J Neuroendocrinol 1:1–4
50. Skinner DC, Robinson JE 1995 Melatonin-binding sites in the gonadotrophenriched zona tuberalis of ewes. J Reprod Fertil 104:243–250
51. Tartaglia LA, Dembski M, Weng X, Deng N, Culpepper J, Devos R, RichardsGJ, Campfield LA, Clark FT, Deeds MC, Sanker S, Moriarty A, Moore KJ,Smutko JS, Mays GG, Woolf EA, Monroe CA, Tepper RI 1995 Identificationand expression cloning of a leptin receptor OB-R. Cell 83:1263–1271
1520 LEPTIN RECEPTOR IN OVINE PITUITARY Endo • 2000Vol 141 • No 4




















