
Copulation activates Fos-like immunoreactivity in the male quail forebrain
17
BEHAVIOURAL BRAIN RESEARCH ELSEVIER BehavIOural Brain Research 85 (1997) 143-159 Research Report Copulation activates Fos-like immunoreactivity in the male quail forebrain Simone L. Meddle a.\ Verdun M. King a, Brian K. Follett a.2, John C. Wingfield b, Marilyn Ramenofsky b, Agnes Foidart c, Jacques Balthazart c.* a BBSRC Group on Photoperiodism and Reproduction. School of Biological Sciences, University of Bristol, Bristol BS8 1 UG, UK b Department of Zoology. University of Washington, Seattle, WA 98105, USA C Laboratory of Biochemistry, University of Liege, 17 place Delcour (Bat. L1). Liege B-4020. Belgium Received 26 April 1996; received in revised form 26 August 1996; accepted 26 August 1996 Abstract It has been demonstrated using Fos immunocytochemistry that copulation activates specific cell populations in the mammalian brain. Prior to this study. no similar work has been carried out in birds. In mammals, Fos has identified brain circuits activated by genital (penile)/somatosensory and by olfactory/vomeronasal stimuli. Such inputs, of course, should play little or no role in birds (no penis. little or no role for olfaction) and a differential responsiveness could therefore be expected. Male Japanese quail (Coturnix japonica) were allowed to interact freely with adult females and the presence of active sexual behavior, including cloacal contact movements, was confirmed in each case. Control subjects were exposed to a domestic chick (same size as an adult quail) and no sexual behavior was observed. Copulation induced the appearance of Fos-like immunoreactive (FLI) cells in the preoptic area, the hyperstriatum ventrale, parts of the archistriatum. and the nucleus intercollicularis. Induction of FLI cells was observed throughout the rostral to caudal extent of the preoptic region of males from the level of the tractus septomesencephalicus to the level of the anterior commissure, and in the rostral part of the hypothalamus to the level of the supraoptic decussation. The FLI cells did not lie directly adjacent to the third ventricle, but were located 500-1000 11m from the ventricle wall at the level of the lateral edge of the medial preoptic nucleus or, in more caudal sections, in a position ventrolateral to the bed nucleus striae terminalis. It is unlikely that the Fos induction in males resulted from copulation-induced endocrine changes because copulation did not affect plasma levels of luteinizing hormone or testosterone. It is concluded that the responses were due to copulation- associated somatosensory inputs and/or to stimuli originating from the female. Keywords Sexual behavior: Neuronal activation, Preoptic area: Nucleus intercolhculans: Coturnix japonica: Immediate early gene 1. Introduction The anterior hypothalamic preoptic area (POA) plays a major role in the control of many aspects of repro- duction including male sexual behavior and control of gonadotrophin release in birds, as in mammals (reviews: in [7,49,52]). The association of the POA with sexual behavior was established through various studies includ- ing hormone implantation into discrete brain nuclei, * Corresponding author Fax: (32) (43) 665971: E-mail: jbalthazart@ulg ac be lPresent address: Department of PhYSIOlogy. Umversity of Edmburgh Medical School, Edmburgh EH8 9AG. UK 2Present address: UniverSity of Warwick, Coventry CV4 7AL UK. 0166-4328/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved PII SOI66-4328(96)00169-6 lesions, electrical stimulation, and analysis of hormone binding sites (reviews: [5,10,25,49,52,55]), In Japanese quail (Colurnix japonica) , a sexually dimorphic nucleus has been identified in the POA: the medial preoptic nucleus (POM) is significantly larger in males than in females [56,77], Behavioral studies have shown that the POM is a necessary and sufficient site for testosterone to act so as to activate male copulation. Lesions of this nucleus block the activation of the behavior by testoster- one while stereotaxic implantation of testosterone in the POM of castrated males restores full levels of behavioral activity [16,17]. Testosterone must also be aromatized within the POM to activate the behavior: implantation of aromatase inhibitors in the POA block the behavioral effects of the steroid only if they are implanted within
Transcript of Copulation activates Fos-like immunoreactivity in the male quail forebrain

BEHAVIOURAL BRAIN
RESEARCH ELSEVIER BehavIOural Brain Research 85 (1997) 143-159
Research Report
Copulation activates Fos-like immunoreactivity in the male quail forebrain
Simone L. Meddle a.\ Verdun M. King a, Brian K. Follett a.2, John C. Wingfield b,
Marilyn Ramenofsky b, Agnes Foidart c, Jacques Balthazart c.*
a BBSRC Group on Photoperiodism and Reproduction. School of Biological Sciences, University of Bristol, Bristol BS8 1 UG, UK b Department of Zoology. University of Washington, Seattle, WA 98105, USA
C Laboratory of Biochemistry, University of Liege, 17 place Delcour (Bat. L1). Liege B-4020. Belgium
Received 26 April 1996; received in revised form 26 August 1996; accepted 26 August 1996
Abstract
It has been demonstrated using Fos immunocytochemistry that copulation activates specific cell populations in the mammalian brain. Prior to this study. no similar work has been carried out in birds. In mammals, Fos has identified brain circuits activated by genital (penile)/somatosensory and by olfactory/vomeronasal stimuli. Such inputs, of course, should play little or no role in birds (no penis. little or no role for olfaction) and a differential responsiveness could therefore be expected. Male Japanese quail (Coturnix japonica) were allowed to interact freely with adult females and the presence of active sexual behavior, including cloacal contact movements, was confirmed in each case. Control subjects were exposed to a domestic chick (same size as an adult quail) and no sexual behavior was observed. Copulation induced the appearance of Fos-like immunoreactive (FLI) cells in the preoptic area, the hyperstriatum ventrale, parts of the archistriatum. and the nucleus intercollicularis. Induction of FLI cells was observed throughout the rostral to caudal extent of the preoptic region of males from the level of the tractus septomesencephalicus to the level of the anterior commissure, and in the rostral part of the hypothalamus to the level of the supraoptic decussation. The FLI cells did not lie directly adjacent to the third ventricle, but were located 500-1000 11m from the ventricle wall at the level of the lateral edge of the medial preoptic nucleus or, in more caudal sections, in a position ventrolateral to the bed nucleus striae terminalis. It is unlikely that the Fos induction in males resulted from copulation-induced endocrine changes because copulation did not affect plasma levels of luteinizing hormone or testosterone. It is concluded that the responses were due to copulationassociated somatosensory inputs and/or to stimuli originating from the female.
Keywords Sexual behavior: Neuronal activation, Preoptic area: Nucleus intercolhculans: Coturnix japonica: Immediate early gene
1. Introduction
The anterior hypothalamic preoptic area (POA) plays a major role in the control of many aspects of reproduction including male sexual behavior and control of gonadotrophin release in birds, as in mammals (reviews: in [7,49,52]). The association of the POA with sexual behavior was established through various studies including hormone implantation into discrete brain nuclei,
* Corresponding author Fax: (32) (43) 665971: E-mail: jbalthazart@ulg ac be lPresent address: Department of PhYSIOlogy. Umversity of Edmburgh Medical School, Edmburgh EH8 9AG. UK 2Present address: UniverSity of Warwick, Coventry CV4 7AL UK.
0166-4328/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved PII SOI66-4328(96)00169-6
lesions, electrical stimulation, and analysis of hormone binding sites (reviews: [5,10,25,49,52,55]), In Japanese quail (Colurnix japonica) , a sexually dimorphic nucleus has been identified in the POA: the medial preoptic nucleus (POM) is significantly larger in males than in females [56,77], Behavioral studies have shown that the POM is a necessary and sufficient site for testosterone to act so as to activate male copulation. Lesions of this nucleus block the activation of the behavior by testosterone while stereotaxic implantation of testosterone in the POM of castrated males restores full levels of behavioral activity [16,17]. Testosterone must also be aromatized within the POM to activate the behavior: implantation of aromatase inhibitors in the POA block the behavioral effects of the steroid only if they are implanted within

144 S.L. Meddle et at. / Behavioural Brain Research 85 (1997) 143-159
this nucleus [9,15]. These studies have identified the POM as an area implicated in the control of quail reproductive behavior. However, further progress in understanding the underlying neural mechanisms requires the elucidation of the specific cell groups involved.
In mammals, the detection of immediate early genes (e.g., c-fos) has provided a powerful technique to assess individual cell activity. Induction of Fos-like immunoreactivity (FLI) has, for example, been demonstrated in appropriate nociceptive and sensory motor systems in rodents following injury and seizures, noxious stimuli, dehydration [28,39,68,74] and even phase shifts in the circadian system [6,65]. The use of immunocytochemical techniques to locate FLI in mammals has also yielded valuable information regarding the identification of neuronal populations that are activated following male copulatory behavior. For example, in gonadally intact male rats sacrificed 1 h after copulation, significant increases in FLI were observed in the medial amygdala (mAMYG), the bed nucleus of the stria terminalis (BNST), the central tegmental field (CTF) and the medial POA [18,19,24,67,80]. All these brain regions had previously been associated with the display of male sexual behavior in rodents through lesioning, electrical stimulation and steroid implantation studies [52]. Similar studies carried out in male golden hamsters [44,86] and gerbils [87] have shown similar distributions of FLI after copulation.
In the rat, FLI observed within the medial POA and the BNST following copulation was significantly reduced following lesions in the mAMYG and CTF suggesting that activation within the medial POA and BNST is driven by afferent inputs from these regions [18]. These findings imply that stimuli associated with copulation and intromission with a sexually receptive female activate a limbic-hypothalamic circuit via genital/ somatosensory and olfactory/vomeronasal stimuli, relayed presumably by the CTF and mAMYG, respectively. In agreement with this interpretation, female hamster vaginal secretion alone can induce FLI in a pattern similar to that achieved following copulation [29]. In addition, testosterone levels rise in male mammals following exposure to receptive females [35,46,64] and these endocrine changes may contribute to the FLI expression in the mammalian brain [30].
The primary aim of the present study was to investigate the brain circuits associated with copulation in quail using FLI as a marker of cell activation. Quail, like most birds, are devoid of an intromittent organ and are reputed to rely little or not at all on olfactory information during sexual interactions. In male mammals, most of the cells expressing FLI after copulation are supposed to be activated either by olfactory / vomeronal or by genital/somatosensory inputs. It might therefore be expected that the Fos response in quail
would be substantially different from that described in mammals.
2. Materials and methods
2.1. Subjects
Male and female Japanese quail (Coturnix japonica) hatched from eggs obtained at the breeding colony (strain S7) established in the BBSRC Group on Photoperiodism and Reproduction at Bristol were raised under short photoperiods (8:16 h light/dark) for the first 3 weeks and then transferred to long days (20:4 h light/dark) where they stayed for a period of 10 weeks. Four weeks prior to experimentation, subjects were transferred to brooding cages (20 x 26 x 36 cm) and separated visually, but not auditorily, from one another. At that time, all subjects were sexually mature. Food and water were available ad libitum.
2.2. Experimental design
Sixteen male quail (13 weeks of age and sexually naive) were used to investigate the effects of copulation on FLI in the brain. Experimental (EXP) stimuli were provided by 8 laying female quail of the same age as the males. These females were also maintained on 20 h light/4 h dark and housed separately before experimentation. In order to control for the general arousal related to the introduction of a female congener, a control stimulus (CTL) was specifically required which would represent another bird introduced within the male home cage, but would not cause dramatic behavioral stimulation. Eight domestic l-week-old hen chicks were chosen as this non-provocative control stimulus because they were the same size as a female quail, although their markings and behavior were completely different. The chicks showed passive non-aggressive behavior and offered no challenge to the male quail. Chicks were purchased commercially and housed together under constant light and a temperature of 35°C before use as a stimulus.
Eight male quail were tested in each of these two conditions (EXP or CTL). One experimental and one control bird were always observed at the same time and their brains processed in parallel. Immediately before the behavioral test, a blood sample was collected from each male for luteinizing hormone (LH) and testosterone measurement. The stimulus animal (female quail or young chick) was then introduced in the residence cage of the experimental subject and birds allowed to freely interact for 1.5-2 h. As soon as the stimuli were introduced into the cage, male sexual behavior was recorded visually from behind a screen for 1 h. Latency to first neck grab (min) and total frequency of crowing, neck

s.L. Meddle et ai/Behavioural Brain Research 85 (1997) 143-159 145
grabs, strutting, mount attempts and cloacal contact movements were recorded (see [2,40] for description). Quail were left undisturbed for 30-60 min and a second blood sample taken immediately prior to sacrifice. The interval of 90-120 min between the beginning of sexual interactions and sacrifice was selected because previous experiments indicated that this is the period when maximum induction of Fos is observed after injection of hypertonic saline in quail and other avian species [50,5 L 73]. Following fixation, the testes were removed and their paired mass recorded.
2.3. Blood sampling and hormone assays
Blood samples (ca. 300 Ill) were collected by venepuncture, centrifuged and the plasmas stored at - 20De. Levels of LH were measured in a single assay by a micromodification of the radioimmunoassay originally devised by Follett et al. [33] and the results expressed in terms of ng/ml against a chicken LH standard (fraction IRC2). Testosterone was measured in triplicate using a commercial antiserum (Biogenesis, Bournemouth, Dorset; 8680-6004) by a protocol similar to the one used in the LH assay and previously described and validated [57].
2.4. Perfusion and Fos immunocytochemistry
Birds were deeply anesthetized with an overdose of sodium pentobarbital (250 mg/kg, i.m.) and then perfused intracardially with 30 ml of 0.9% physiological saline containing 750 IU heparin followed by 300 ml of modified Zamboni fixative (1.8% paraformaldehyde and 7.5 ml saturated picric acid in 0.01 M phosphatebuffered saline (PBS) at pH 7.2-7.4). Brains were dissected out of the skull, postfixed at 4°C overnight, washed in PBS and embedded in gelatin.
Coronal section (60 11m thick) were cut on a vibratome from the lobus parolfactorius to the level of the third nerve and processed for Fos immunocytochemistry as free-floating sections in PBS buffer as described previously [51]. Briefly, sections were washed in PBS containing 0.2% Triton X-100 (PBST) and incubated in 0.3% hydrogen peroxide in PBS and in 2% normal goat serum in PBST. They were then left for 70 h in the primary antiserum diluted I :5000 in 2% normal goat serum in PBST. The antibody-antigen complex was localized using the avidin-biotin complex (ABC) method performed with a Vector Elite kit (Vector Laboratories PLC, Cambridge, UK). The peroxidase was finally visualized with diaminobenzidine. Sections were mounted on gelatin-coated slides, dehydrated and mounted in DPX mountant (BDH, Poole, UK).
The primary antibody (9/3 Fos) was raised by the BBSRC's central immunology facility at Babraham (Cambridge, UK) against a synthetic polypeptide from
the C-terminal of chicken c-fos (sequence taken from [34]; see [51,73] for detail). Previous studies have shown that when control sections are incubated with primary antiserum preabsorbed with I Ilg/ml of the synthetic chicken Fos peptide or non-immune rabbit serum, no staining is observed [51,73].
2.5. Analysis of data
Sections were observed under the microscope with x 10-20 objectives and labeled nuclei counted directly at this magnification. When large number of FLI cells were detected in a given section, it was first drawn with the help of a camera lucida and numbers of positive nuclei counted on these drawings. Counts were made on both sides of the brain with the observer being blind to the treatment received by the animals. No correction for double counting was performed because objects that were counted (nuclei) are small compared with section thickness and double counting was very unlikely. Graphs displaying the mean numbers of FLI cells in selected brain areas were prepared by first re-aligning the sections from different subjects in the rostral to caudal order with the help of anatomical landmarks, such as the anterior commissure, the posterior commissure or the para ventricular organ. The first section in which a given landmark appeared when moving in a rostral to caudal direction was taken as the reference point. Mean numbers of FLI cells at the different levels were then calculated and plotted. Cell counts were performed either in every successive section where the structure under consideration was small, or every second or third section when wider areas had to be considered.
Data were analyzed by one- or two-way analysis of variance (ANOVA) with the experimental groups as the independent factor and the successive sections in a given area as the repeated factor. These were followed when appropriate by post-hoc Fisher protected least significant difference tests. Student t-tests, x2-tests and Mann-Whitney V-tests were also used where appropriate to compare pairs of results. Data presented in all graphs are means ± SEM. Differences were considered significant at an ex-level of P:::::; 0.05.
3. Results
3.1. Behavioral interactions and hormone plasma levels
Males exposed to a domestic chick usually appeared startled at first and two of them even tried to run away. After some habituation, most birds then ignored the chick. Male quail never showed sexual or aggressive behavior toward the chick, but they continued to crow. Chicks did, on occasion, approach the male and attempt to be brooded by crouching beneath the male and

146 s.L. Meddle et al. / Behavioural Brain Research 85 ( 1997) 143-159
pushing up at its wing. The quail did not react to this behavior.
By contrast, when presented to a female quail, all males showed head grabs, mounts or attempts to mount, and then cloacal contact movements (Fig. 1). Neck grab usually occurred in less than 1 min (see Fig. 1 for quantitative analysis). The female initially pecked at or retreated from the male, but later crouched (or halfcrouched) and allowed the male to mount her. Strutting behavior was also observed in all males following the introduction of a female. The mean frequency of crowing was reduced in the presence of females, but this effect did not reach statistical significance.
The plasma levels of LH were similar in the two groups of males before exposure to the stimuli and were not altered by the experimental procedures (Before: EXP, 5.52± 1.77; CTL, 4.95± 1.09. After: EXP, 6.53± 1.69; CTL, 5.34±0.96 ng/ml, means±SEM). Similarly, no group difference and no effect of the treatments was detected for testosterone levels (Before: EXP, 3.58±0.68; CTL, 3.15±0.67. After: EXP, 2.57±1.06; CTL: 5.58±2.13ng/ml, means±SEM). This lack of experimental effects was confirmed statistically by two-way ANOVAs with the treatments as the independent variable and the two estimates of LH and testosterone (before and after the test) as the repeated variable. These analyses revealed no overall difference between the groups (LH, F1.14 = 0.229, P = 0.639; testosterone, F1.14 = 1.097, P=0.312), no effect of the repeated
200~--~~----------.----------------,
150
100
so
o
40
30
20
10
o Strut
60
20
~ ~
*** Neck
***
Cloacal Contact Movements
Crow
Fig. l. Bar graphs showing the mean frequency (± SEM) of the behaviors displayed by male quail when presented to a female conspecific (EXP) or to a young domestic hen chick as a control (CTL). The latency between introduction of the stimulus and the first neck grab is also shown. Eight males were tested in each condition and data were compared by bilateral Mann-Whitney U-test whose results are shown at the top of the bars as follows: *** P < 0.00l.
variable (LH, F1,14=0.704, P=0.415; testosterone, F1,14=0.282, P=0.603) and no interaction (LH, F1,14 =0.134, P= 0,720; testosterone, F1.14 = 1.654, P= 0.219).
3.2. Effect of copulatory behavior on Fos-like immunoreactivity
FLI cells were observed in several discrete brain areas (Figs. 2 and 3) and the dense immunoreactivity was always nuclear. Numerous positive cells were observed in the rostral forebrain running along the lateral edges of the ventricles at the level of the hyper striatum ventrale (Fig.2A).
At slightly more caudal levels, a group of FLI cells extended laterally in the hyperstriatum ventrale above the lamina hyperstriatica (Figs. 2B-D and 3C). Dense populations of FLI cells were also present throughout the preoptic region and anterior hypothalamus, as well as in slightly more dorsal position in the paleostriatum ventrale, the nucleus accumbens (Figs. 2B,C and 3A), and more caudally at the dorsal tips of the bed nucleus of the stria terminalis (Fig. 2C,D). At a similar level in the brain, numerous FLI cells could be seen in the lateral and medial septum. Positive cells were also detected in the archistriatum mainly intermingled with the most lateral aspects of the anterior commissure and of the occipitomesencephalic tract. This cell group extended into the nucleus taeniae (Figs. 2C and 3B). Large numbers of cells were present in the hypothalamic region between the level of the supraoptic decussation to the most caudal aspects (Figs.2D-F and 3D,E). They overlapped with the paraventricular, ventromedial and mammillary nuclei. In these sections, FLI material was also observed in many cells of the thalamus including the habenular region. In the mesencephalon, many FLI cells were located in the intercollicular nucleus throughout its rostral to caudal extension and at more medial levels within the central gray (Figs. 2E,F and 3F,G).
Patterns of FLI distribution from both treatment groups were plotted on pre drawn brain atlases. Visual inspection of these drawings suggested that the number of FLI cells may have been affected by the experimental treatments in a number of specific brain regions; these were then submitted to a quantitative analysis.
3.3. Preoptic area-anterior hypothalamus
In the preoptic area and anterior hypothalamus, a symmetrical cluster of FLI nuclei was consistently observed on both sides of the midline under or just rostral to the tractus septomesencephalicus (TSM) and, at more posterior levels, on both sides of the third ventricle. These cells extended from the rostral edge of the TSM to the level of the bed nucleus striae terminalis,

S.L Meddle et ai/Behavioural Brain Research 85 ( 1997) 143-159 147
A ~-- HA /~
.~ o
~ ~PVl Ac / AId
Alv
B E
F
Fig. 2. Schematic drawings of coronal sections in the male quail brain Illustrating the anatomical localization of FLI Immunoreactive cells (dots) that were detected in subjects exposed to a female congener. Sections A-F are arranged in a rostral to caudal order. The boxes indicate the areas where quantitative analyses were carried out. AA, archistriatum anterior; Ac. nucleus accumbens; Ald. archistriatum intermedium, pars dorsalis; Alv, archistriatum intermedium. pars ventralis; AM, nucleus antenor medialis hypothalami; CA, commissura anterior; Cb, cerebellum; CP, commissura posterior; CPa. commissura pallii; DSM, decussatio supramamillaris; E, ectostriatum; FPL, fasciculus prosencephali lateralis (lateral forebrain bundle); GCt, substantia grisea centralis; HA, hyperstriatum accessorium; Hp, hippocampus; HV. hyperstriatum ventrale; ICo. nucleus mtercollicularis; IH, nucleus inferior hypothalami; LHY. nucleus lateralis hypothalami; ME, eminentia mediana; MLd. nucleus mesencephahcus lateralis, pars dorsalis; ML, nucleus mamillans laterahs; MM, nucleus mamillaris medialis; N. neostriatum; nST. nucleus striae terminalis; OM. tractus occipitomesencephalicus; OV, nucleus ovoidalis; PHN. nucleus periventricularis hypothalami; POM. nucleus preopticus medialis; PVN. nucleus paraventriculans; PVO. organum paraventriculare; PVT. paleostriatum ventrale; Ru, nucleus ruber; SCNm. nucleus suprachiasmaticus. pars medialis; SL nucleus septalis lateral is; SM, nucleus septahs medialis; Tn. nucleus taeniae; TSM, tractus septomesencephalicus; Tu. nucleus tuberis.
just rostral to the supraoptic decussation. They were not directly adjacent to the ventricle, but, instead, were located throughout the sections at 500-1 000 ~m from the midline (Fig. 4).
The number of FLI cells in this group was counted in every section and the sections from different subjects were re-aligned in the rostral to caudal direction taking the first section where the anterior commissure appeared as a landmark. The mean numbers of positive cells per section computed in this way for the two experimental groups are presented in Fig. 5 and, as can be observed, larger numbers of FLI cells were observed in the experimental birds in sections located on both the rostral and caudal sides of the anterior commissure. The difference between the two experimental groups
extended throughout the preoptic area and anterior hypothalamus.
A two-way ANOVA of the data relative to sections 6-25 (a few missing data points at both ends of the cell group prevented inclusion of the other sections in the analysis) with the two experimental groups as independent variable and the consecutive sections as repeated measure, showed that the overall number of FLI cells differed significantly between the two experimental groups (Fl. 14 = 8.037, P = 0.0132) and between successive sections (FI9 ,266 = 1.846, P=0.0185). There was also a very strong interaction between these two variables (F19,266 = 8.085, Ps;O.OOOl). Visual inspection of these results as plotted in Fig. 5 revealed the origin of this interaction: there appeared to be no experimental effect
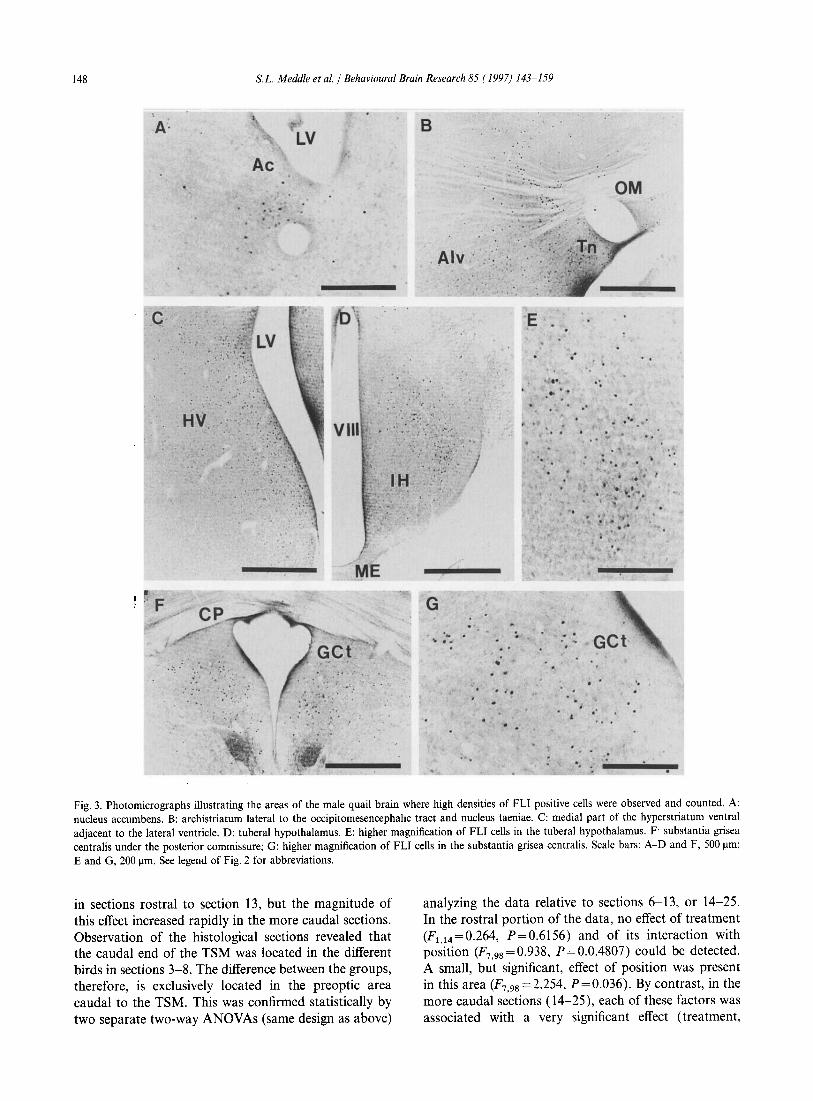
148 s.L. Meddle et al. / Behavioural Brain Research 85 ( 1997) 143-159
-. ·.c .... ,.: .. : ..... , .
. ::; : .... j. ~:. LV .. . .~.' .' :':;' ,
. ", .>~::\.,I.I~ . : .. ' • '··:.~i
, ~ .... ~ .. ,,: .
.':, .,
B
.: .. : . ~",:.'; '~=.. .. ~ .. ....!; .. ~,w'
... :·· .. ~.<~:;;;'.~~H~: .
.. '. / ~·;.r.!!;'· ~:. . :'
. Alv: .,' .. ::; .... ;';. '.~" .. , . . ... ::~ .. ~ ~
< I
3
G
'.. . ... #
",
. •
" . .. ", .:. . .-
.. , .... ~ . . > . ,
'" :" ..... «,O'{.~ :,.":
" . ... ' 'lttf i...
' .. .. '.:.t ",
.. Jt • fit .• ~
.. . ,
. "
j .' 4 " .
. .' '.
.' ..
OM
, .
" "
Fig. 3. Photomicrographs illustrating the areas of the male quail brain where high densities of FLI positive cells were observed and counted. A: nucleus accumbens. B: archistriatum lateral to the occipitomesencephahc tract and nucleus taeniae. C: medial part of the hyperstriatum ventral adjacent to the lateral ventricle. D: tuberal hypothalamus. E: higher magnification of FLI cells in the tuberal hypothalamus. F substantia grisea centralis under the posterior commissure; G: higher magnification of FLI cells in the substantia grisea centralis. Scale bars: A-D and F, 500 J.lm; E and G. 200 J.lID. See legend of Fig. 2 for abbreviations.
in sections rostral to section 13, but the magnitude of this effect increased rapidly in the more caudal sections. Observation of the histological sections revealed that the caudal end of the TSM was located in the different birds in sections 3-8. The difference between the groups, therefore, is exclusively located in the preoptic area caudal to the TSM. This was confirmed statistically by two separate two-way ANOVAs (same design as above)
analyzing the data relative to sections 6-13, or 14-25. In the rostral portion of the data, no effect of treatment (F1,14=0.264, P=0.6156) and of its interaction with position (F7 ,98 = 0.938, P = 0.0.4807) could be detected. A small, but significant, effect of position was present in this area (F7 ,98 =2.254, P=0.036). By contrast, in the more caudal sections (14-25), each of these factors was associated with a very significant effect (treatment,

A
, ..
o
G
s.L. Meddle et at. / Behavioural Brain Research 85 ( 1997) 143-159
B
" "'~',:, ;.~ ;; .' ... '" .
'0 ~.:
0' ; 't J
.f' ,'"
'",
"'\ . ~ ;t::. , I
,-' ~Y
...... ..
E
H • .. •
.. ... - ' t, .. t
. ,,-.. .. .. , -,-,' - '\""
.,. " '" :! • ..:a. i • ''''-'i'c''' ....... " . I.'f .••. "'.. ~ ',-, '~':-.,,-~'
.. "f6~'V'~f'.' . ~. ,~,~, ,I ' •
CA
- "
•• .... ...
.. .. •
','
.,-
0' • ..
•
c
• !,
"
F
t
.. .. ... At . .
..
-, . . 'i ". ..
. ..... .:. ' .. i ...
, . }
" ,
~'!.i.:~ " . "', .. ~.
. I ~ .
~
, ... '. • .. .'
. ,,-... !:~~ ;t. "-~ "!I' " ~
149
•
•
Fig. 4, Photomicrographs illustrating the induction of FLI in the preoptic area/anterior hypothalamus of male quail which copulated with a female conspecific. A-C Illustrate 3 levels of the preoptic area-anterior hypothalamus of male quail that were exposed to a young chick as a controL D-F show similar levels in birds that were exposed to a female quaiL G. H and I show higher magmfications of the main group of FLI cells m D, E and F, respectively, A and D are taken roughly III the middle of the preoptic area III the rostral to caudal axis (medial part of the nucleus preoptic medialis (POM», Band E illustrate the caudal part of the preoptic area under the antenor commissure (caudal end of POM) and C and F show the anterior hypothalamus at the level of the bed nucleus of the stna tennmalis (BNST) VIII. third ventricle. MagnificatIOn is the same in A-F and G-L Scale bars: F, 500 flm; I, 100 flm.
150 s.L. Meddle et al. / Behavioural Brain Research 85 ( 1997) 143-159
Preoptic area· anterior hypothalamus 80
5 "- 60 ~Oi: ,,'" ~+I
~~ "'5
40
.... = 00
~~ ~~ = 20 Z
Nucleus Intercollicularis 400.---------~----------------------__.
5. 300 MLd -
10 15 20
Section number
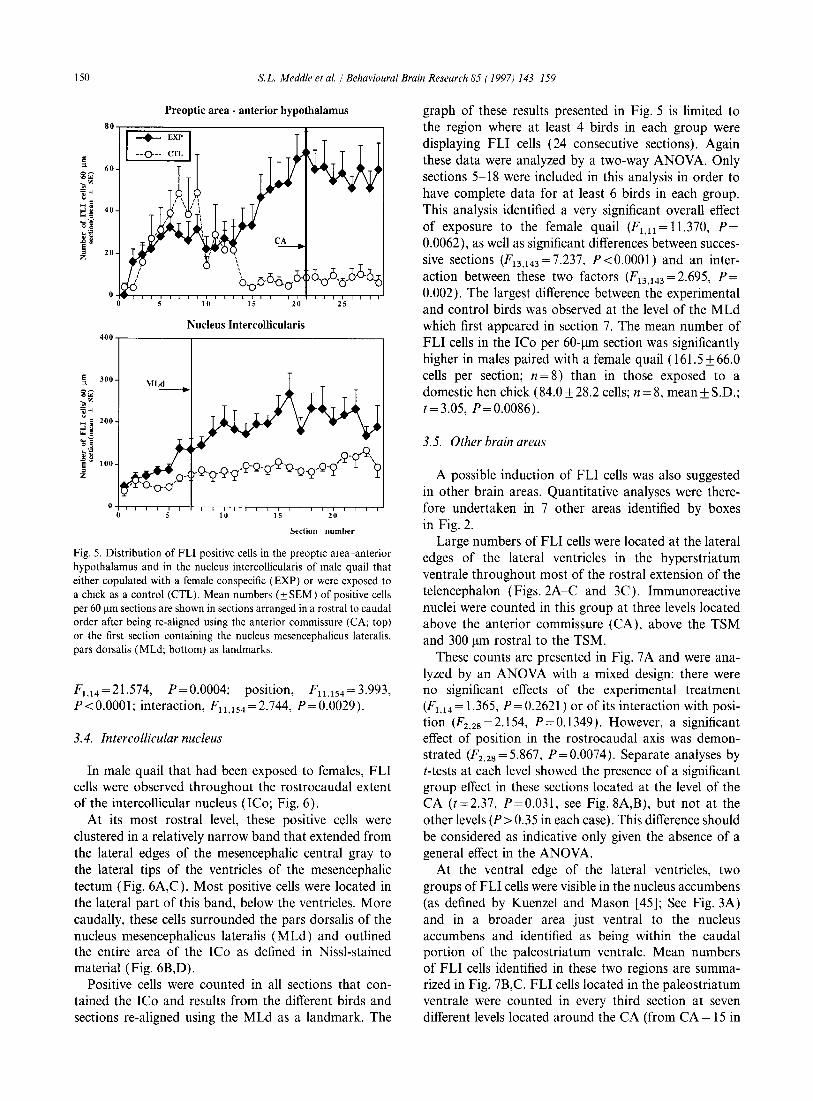
Fig. 5. Distribution of FLI positive cells in the preoptic area-anterior hypothalamus and in the nucleus intercollicularis of male quail that either copulated with a female conspecific (EXP) or were exposed to a chick as a control (CTL). Mean numbers (±SEM) of positive cells per 60 Ilm sections are shown in sections arranged in a rostral to caudal order after being re-aligned using the anterior commissure (CA; top) or the first section containing the nucleus mesencephalicus lateralis, pars dorsalis (MLd; bottom) as landmarks.
F1,14=21.574, P=0.0004; posItion, Fll .154 = 3.993, P<O.OOOl; interaction, Fll .154 =2.744, P=0.0029).
3.4. Intercollicular nucleus
In male quail that had been exposed to females, FLI cells were observed throughout the rostrocaudal extent of the intercollicular nucleus (ICo; Fig. 6).
At its most rostral level, these positive cells were clustered in a relatively narrow band that extended from the lateral edges of the mesencephalic central gray to the lateral tips of the ventricles of the mesencephalic tectum (Fig. 6A,C). Most positive cells were located in the lateral part of this band, below the ventricles. More caudally, these cells surrounded the pars dorsalis of the nucleus mesencephalicus lateralis (MLd) and outlined the entire area of the ICo as defined in Nissl-stained material (Fig. 6B,D).
Positive cells were counted in all sections that contained the ICo and results from the different birds and sections re-aligned using the MLd as a landmark. The
graph of these results presented in Fig. 5 is limited to the region where at least 4 birds in each group were displaying FLI cells (24 consecutive sections). Again these data were analyzed by a two-way ANOV A. Only sections 5-18 were included in this analysis in order to have complete data for at least 6 birds in each group. This analysis identified a very significant overall effect of exposure to the female quail (Fl,ll = 11.370, P= 0.0062), as well as significant differences between successive sections (F13,143=7.237, P<O.OOOI) and an interaction between these two factors (F13.143 = 2.695, P= 0.002). The largest difference between the experimental and control birds was observed at the level of the MLd which first appeared in section 7. The mean number of FLI cells in the ICo per 60-llm section was significantly higher in males paired with a female quail (161.5 ± 66.0 cells per section; n = 8) than in those exposed to a domestic hen chick (84.0 ± 28.2 cells; n = 8, mean ± S.D.; t=3.05, P=0.0086).
3.5. Other brain areas
A possible induction of FLI cells was also suggested in other brain areas. Quantitative analyses were therefore undertaken in 7 other areas identified by boxes in Fig. 2.
Large numbers of FLI cells were located at the lateral edges of the lateral ventricles in the hyperstriatum ventrale throughout most of the rostral extension of the telencephalon (Figs.2A-C and 3C). Immunoreactive nuclei were counted in this group at three levels located above the anterior commissure (CA), above the TSM and 300 11m rostral to the TSM.
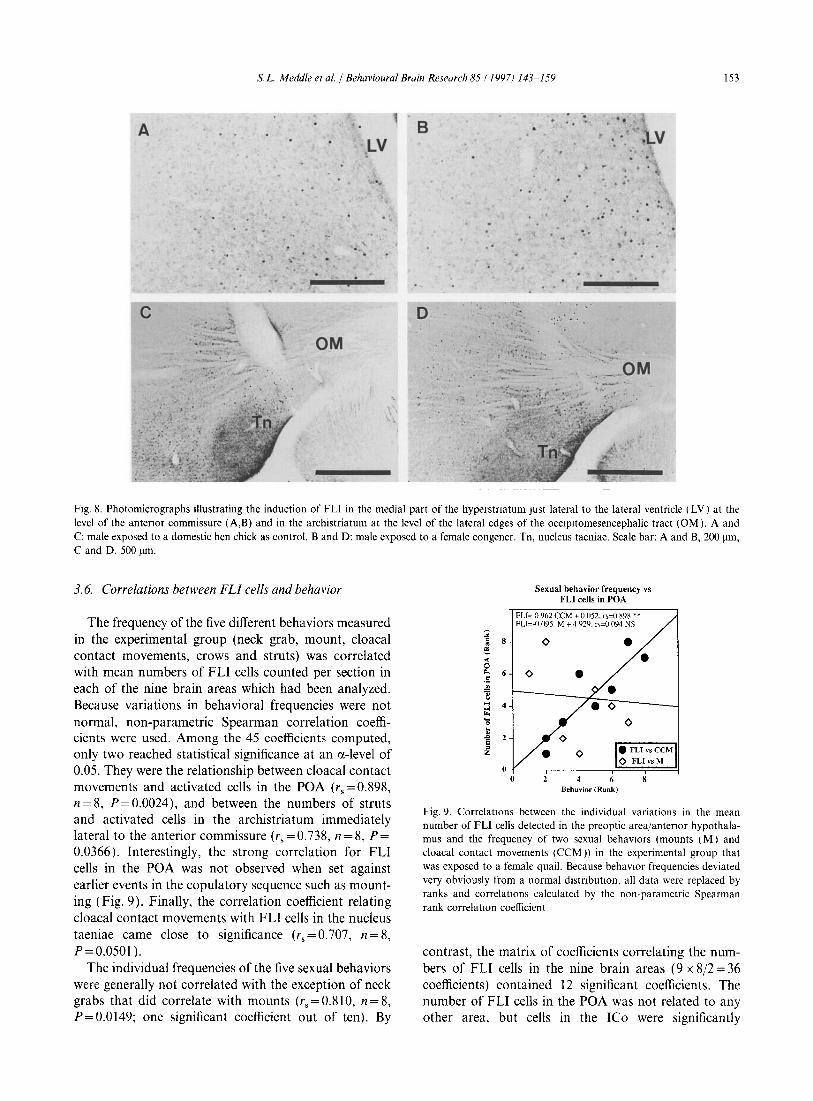
These counts are presented in Fig. 7 A and were analyzed by an ANOV A with a mixed design: there were no significant effects of the experimental treatment (F1.14 = 1.365, P = 0.2621) or of its interaction with position (F2.2s=2.154, P=0.1349). However, a significant effect of position in the rostrocaudal axis was demonstrated (F2 •2S =5.867, P=0.0074). Separate analyses by t-tests at each level showed the presence of a significant group effect in these sections located at the level of the CA (t=2.37, P=0.031, see Fig. 8A,B), but not at the other levels (P>0.35 in each case). This difference should be considered as indicative only given the absence of a general effect in the ANOVA.
At the ventral edge of the lateral ventricles, two groups of FLI cells were visible in the nucleus accumbens (as defined by Kuenzel and Mason [45]; See Fig.3A) and in a broader area just ventral to the nucleus accumbens and identified as being within the caudal portion of the paleostriatum ventrale. Mean numbers of FLI cells identified in these two regions are summarized in Fig. 7B,C. FLI cells located in the paleostriatum ventrale were counted in every third section at seven different levels located around the CA (from CA - 15 in
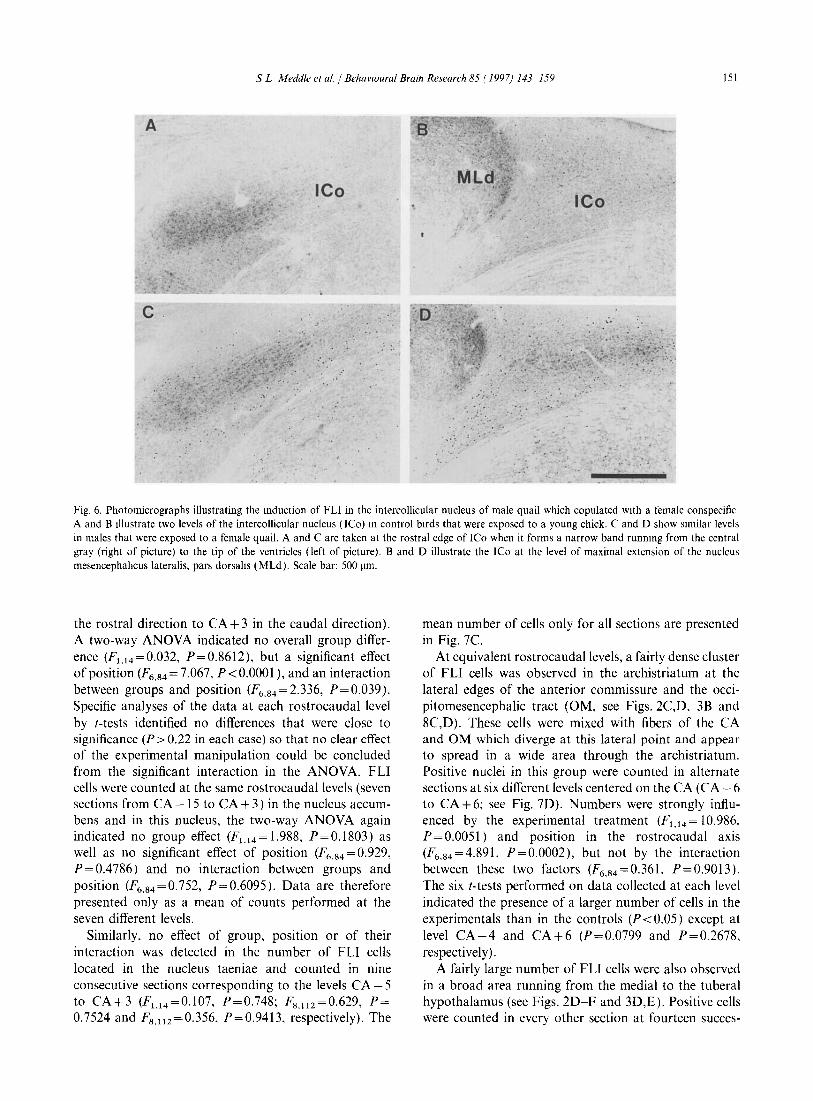
S L Meddle el at. / BehavIOural Brain Research 85 ( 1997) 143-159 151
A
leo
c.
'., ".
. ,', ,"
............ , ..... '-ri "',' '." ... ':~;"'*' .:D' .. ' .... :.i:~.:::;~\}:,
M{"
·lio·. Ir:· .
... , ... . '. ',:"
>~'1 • ,'" .r
".
. :, ....
:.
," • ~. • ~# .,
. ',.,:,. . '. ~~ . ,' .
,', ; ..... . ~.
.... :
Fig. 6. Photomicrographs illustrating the mduction of FLI in the intercollicular nucleus of male quail which copulated with a female conspecific A and B Illustrate two levels of the intercollicular nucleus (lCo) m control birds that were exposed to a young chick. C and D show similar levels in males that were exposed to a female quail. A and C are taken at the rostral edge of ICo when it forms a narrow band runnmg from the central gray (right of picture) to the tip of the ventricles (left of picture). Band D illustrate the ICo at the level of maximal extension of the nucleus mesencephahcus lateralis. pars dorsahs (MLd). Scale bar: 500 !lm.
the rostral direction to CA + 3 in the caudal direction). A two-way ANOVA indicated no overall group difference (Fl.14=0.032, P=0.8612), but a significant effect of position (F6 •84 = 7.067, P <0.0001), and an interaction between groups and position (F6•84 =2.336, P=0.039). Specific analyses of the data at each rostrocaudal level by [-tests identified no differences that were close to significance (P> 0.22 in each case) so that no clear effect of the experimental manipulation could be concluded from the significant interaction in the ANOVA. FLI cells were counted at the same rostrocaudallevels (seven sections from CA - 15 to CA + 3 ) in the nucleus accumbens and in this nucleus, the two-way ANOVA again indicated no group effect (F1.l4 = 1.988, P = 0.1803) as well as no significant effect of position (F6 •84 = 0.929, P=0.4786) and no interaction between groups and position (F6 •84 =0.752, P=0.6095). Data are therefore presented only as a mean of counts performed at the seven different levels.
Similarly, no effect of group, position or of their interaction was detected in the number of FLI cells located in the nucleus taeniae and counted in nine consecutive sections corresponding to the levels CA - 5 to CA+3 (F1.14=0.107, P=0.748; F8 .1l2 =0.629, P= 0.7524 and F8 .1l2 =0.356, P=0.9413, respectively). The
mean number of cells only for all sections are presented in Fig. 7C.
At equivalent rostrocaudallevels, a fairly dense cluster of FLI cells was observed in the archistriatum at the lateral edges of the anterior commissure and the occipitomesencephalic tract (OM, see Figs.2C,D, 3B and 8C,D). These cells were mixed with fibers of the CA and OM which diverge at this lateral point and appear to spread in a wide area through the archistriatum. Positive nuclei in this group were counted in alternate sections at six different levels centered on the CA (CA - 6 to CA + 6; see Fig. 7D). Numbers were strongly influenced by the experimental treatment (F1,14 = 10.986, P = 0.0051) and position in the rostrocaudal axis (F6•84 =4.891, P=0.0002), but not by the interaction between these two factors (F6•84 =0.361, P=0.90l3). The six [-tests performed on data collected at each level indicated the presence of a larger number of cells in the experimentals than in the controls (P < 0.05) except at level CA-4 and CA+6 (p=0.0799 and P=0.2678, respectively).
A fairly large number of FLI cells were also observed in a broad area running from the medial to the tuberal hypothalamus (see Figs. 2D-F and 3D,E). Positive cells were counted in every other section at fourteen succes-

152 s.L. Meddle et al. / Behavioural Brain Research 85 ( 1997) 143-159
A
c
E
Hyperstriatum 800~----________ ~~ __________________ -,
fiOO
400
200
30
20
10
r···················-1····· ... ............... ?
-+- EXP
-0--' CTL
Ant. TSM CA
N. Accumbens and N. Taeniae • EXP
o CTL
Accumbens Taeniae
Medial and Tuberal Hypothalamus 200~----________________ ~~ _________ ~
--<>-- EXP
-0--' CTL
ISO
100
50
·14 ·12 ·10 .8 ., ·4 ·2 pvo +2 +4 +6 +8 +10 +12
B Paleotriatum ventrale posterior 80~------------------------~------~-,
'0 40 .. 11/ ~
e i. 20
D
-+- EXP
-0--' CTL
T 1 __ ,t --1,---1 ///h....... .' ... y..... 1
CA·I' CA·12 CA.9 CA·6 CA.3 CA CA+3
Archistriatum 125 ~--------------"""':"':""""""':--------------,
..... c
100
.. 50
il e = z 25
F
-+- EXP
-0--' CTL
. . ...... o·······?·······?·······1 .. ~ ....... r 1 g ...... .
CA.6 CA·4 CA·2 CA
Substantia grisea centralis 150...------------....::-----------------,
.!!l
'B 100
::3 ~ ... c ..
11/
~ 50
= Z
-+- EXP
-0--. CTL
r····y-···y····y-···y····y···· .,.-
CP CP+I CP+2 CP+3 CP.4 CP+5 CP+6 CP+' CP+8 CP+9
FIg. 7. Quantitative analysis of FLI positive cells in several brain regions of male quail which copulated with a female conspecific (EXP) or were exposed to a chick as a control (CTL). Mean numbers (± SEM) of posItive cells per 60-llm sections are shown in sectIOns arranged in a rostral to caudal order after being re-alIgned using the anterior commissure (CA), the posterior commissure (CP) or the paraventricular organ (PVO) as landmarks. Because no rostrocaudal difference was detected In the nucleus accumbens and nucleus taeniae, means across all levels are presented.
sive levels (aligned using the paraventricular organ (PVO) as a landmark, Fig.7E). More FLI cells were usually present in the experimental group of quail than in controls, but large interindividual variability meant that the difference between the two groups did not reach statistical significance (F1,14 = 1.466, P = 0.246). A sig· nificant effect of position in the rostrocaudal axis was observed (F13.182 = 12.888, P < 0.0001) and the inter· action between groups and position came close to signi· ficance (F13,18Z=1.762, P=0.0521). Student's (·tests between experimental and control values at each level failed to confirm this significant difference.
Finally, a population of FLI cells was counted in the substantia grisea centralis (central gray, GCt; Fig. 3F,G) in ten successive sections caudal to the posterior commissure (CP; Fig.7F). No significant effect of experimental treatment was detected (Fl,14 = 0.516, P=0.4844) despite the fact that numerical values at most levels were higher in the experimental group of quail. Significant effects of position and interaction were present (F9,126 = 3.270, P=0.0016 and F9.126 = 2.595, P = 0.0089, respectively). Student's (·tests failed, however, to confirm significant group differences at any of these levels.
s.L. Meddle et al. / Behavioural Brain Research 85 ( 1997) 143-159 153
A . . 'LV ~
.~ ...
-I " . ~ "t • . ....... ::.~ • -
, . . ,. t ; $ "
OM
B •
.
o·
'."
..
. .
-. ..
J>
. °t.' 1. I.' : .• t'll'tv
...... ~. \ . :~\
• t'
> ' . : .. ~, . .. .
•• t . .
Fig. 8. Photomicrographs illustrating the induction of FLI in the medial part of the hyperstnatum Just lateral to the lateral ventricle (LV) at the level of the antenor commissure (A,B) and in the archistriatum at the level of the lateral edges of the occlpltomesencephalic tract (OM). A and C: male exposed to a domestic hen chick as control. Band D: male exposed to a female congener. Tn, nucleus taeniae. Scale bar: A and B, 200 11m, C and D. 500 11m.
3.6. Correlations between FLI cells and behavior
The frequency of the five different behaviors measured in the experimental group (neck grab, mount, cloacal contact movements, crows and struts) was correlated with mean numbers of FLI cells counted per section in each of the nine brain areas which had been analyzed. Because variations in behavioral frequencies were not normal, non-parametric Spearman correlation coefficients were used. Among the 45 coefficients computed, only two reached statistical significance at an ex-level of 0.05. They were the relationship between cloacal contact movements and activated cells in the POA (rs = 0.898, n = 8, P = 0.0024), and between the numbers of struts and activated cells in the archistriatum immediately lateral to the anterior commissure (rs=0.738, n=8, P= 0.0366). Interestingly, the strong correlation for FLI cells in the POA was not observed when set against earlier events in the copulatory sequence such as mounting (Fig. 9). Finally, the correlation coefficient relating cloacal contact movements with FLI cells in the nucleus taeniae came close to significance (rs = 0.707, n = 8, P=0.0501 ).
The individual frequencies of the five sexual behaviors were generally not correlated with the exception of neck grabs that did correlate with mounts (rs=0.81O, n=8, P=0.0149; one significant coefficient out of ten). By
2 = 8 .. ~ ...: 0
"" 6 .5 ~
~ :l ... 'E ~ 2 ;: = Z
0 0
Sexual behavior frequency vs FLI cells in POA
FLI~ () 962 CCM + 0 052. ,,~O 898 ** FLI~-(1.095 M + 4 929. ,,~O 094 NS
<> • • <> •
<> <>
• <> • FLIvsCCM
<> FLI vs M
6 Behavior (Rank)
Fig. 9. CorrelatIOns between the individual variations in the mean number of FLI cells detected in the preoptic area/antenor hypothalamus and the frequency of two sexual behaVIOrs (mounts (M) and cloacal contact movements (CCM)) in the experimental group that was exposed to a female quail. Because behaVIOr frequencies deviated very obviously from a normal distnbutlOn. all data were replaced by ranks and correlatIOns calculated by the non-parametric Spearman rank correlatIOn coefficient
contrast, the matrix of coefficients correlating the numbers of FLI cells in the nine brain areas (9 x 8/2 = 36 coefficients) contained 12 significant coefficients. The number of FLI cells in the POA was not related to any other area, but cells in the leo were significantly

154 s.L. Meddle ef at. / BehavIOural Brain Research 85 ( 1997) 143-159
correlated with cells in seven out of the eight other areas (correlation with nucleus taeniae only was not significant). In addition, correlations of FLI cell numbers were observed between the hyperstriatal area at the edge of lateral ventricles on the one hand, and the ventral paleostriatum or the archistriatum lateral to the CA on the other, between the central gray and the paleostriatum ventrale, nucleus accumbens and medial hypothalamus, and finally between the paleostriatum ventrale and the medial hypothalamus
4. Discussion
4.1. Behavioral and hormonal results
As expected based on previous studies in this and other laboratories [2,14,20], male quail presented with a sexually mature female displayed vigorous sexual behavior and each male achieved more than ten apparently successful copulations during the I-h observation period. A domestic chick of the same size as a female quail failed to induce any sort of copulatory display or aggression, and proved valuable as the control stimulus for general arousal. By contrast, the crow vocalization occurred in both situations, and even tended to occur with lower frequencies in the presence of a female compared to a chick. This is consistent with previous reports demonstrating that crowing is a contact call preferentially produced when no congener is in sight [27,36,37]. This provides additional evidence suggesting that the chick was not perceived as a conspecific.
Although the presence of a female quail elicited major behavioral changes, in agreement with an earlier study [26], no alterations in LH or testosterone plasma levels were detected. It is possible that, under long days, male quail are maximally stimulated and any additional stimulatory effect of a female is not significant. Endocrine responses to the presence of a female have been demonstrated in starlings (Sturn us vulgaris), white-crowned sparrows (Zonotrichia leucophrys) and ring doves (Streptopelia risoria) (see [7,84] for review). The presence or absence of these responses appears to be a speciesspecific characteristics that may relate to the social system and reproductive strategy (e.g., monogamous vs. polygynous species) [82,83]. Regardless of this possible species-specificity, it must be pointed out that the lack of changes in the plasma LH and testosterone levels of male quail during the present experiment strongly indicates that the activation of brain FLI cells does not depend on a change in the endocrine environment caused by the pairing with a female. The role of specific inputs and/or of performing specific behaviors must therefore be considered.
4.2. Immunocytochemical specificity
A strong and reliable induction of FLI cells was detected in several brain regions of males that had been allowed to copulate with females by comparison with the subjects exposed to the control procedure. Previous work demonstrates that this immunoreactive signal is specific of the Fos or of Fos-related antigens. No immunoreactivity is observed when the primary antibody is replaced by non-immune rabbit serum or pre incubated with the synthetic 20-amino acid peptide from the C-terminus of chicken Fos [51]. This primary antibody also gives a specific band in Western blots at the expected size (about 50 kDa) of the chicken Fos protein [34.51,73]. Furthermore, previous physiological experiments devoted to the central control of osmoregulation or to effects of varying photoperiods have identified FLI cells in relevant brain areas with antibody 9/3 used in the present study [51,73]. Finally, the anatomical nature of the FLI induction observed here leaves little doubt concerning the specificity of the immunoreactive signal. The immunoreactive material was observed exclusively in nuclear location as expected for the Fos antigen. It was also neuroanatomically discrete and present only in brain areas that were previously demonstrated to be more or less directly implicated in the control of reproductive behavior.
4.3. FLI induction in the PDA
Interaction with sexually mature females induced differential expression of FLI in four brain areas of the male: the preoptic area/anterior hypothalamus, the nucleus intercollicularis, part of the archistriatum just lateral to the anterior commissure, and, possibly, part of the hyper striatum ventrale just lateral to the lateral ventricles at the level of the anterior commissure. The most prominent induction of Fos cells occurred in the preoptic area-anterior hypothalamus and this finding substantiates earlier work using other techniques which implicates the quail's POA in copulatory behavior [15-17,55]. It is particularly interesting that Fos induction within the POA increased in magnitude from the rostral to the caudal part of the region, so that the maximal effect was seen in sections located at the level of the anterior commissure (Fig. 5). This is the part of the POA that has been linked most closely to the steroidinduced facilitation of copulation (review: [55]). The finding of Fos activation by copulation in this area therefore reinforces the notion that the caudal part of the POA constitutes a key center of integration in copulation. The fact that levels of FLI were not significantly different between CTL and EXP birds in the anterior POA emphasizes the neuroanatomical specificity of the difference observed in the caudal POA and provides an excellent control for possible non-specific

s.L. Meddle et aI/BehavIOural Bralll Research 85 ( 1997) 143-/59 155
effects. Furthermore, few FLI cells were observed in the caudal POA and anterior hypothalamus of birds exposed to the control situation which indicates that induction of FLI in the EXP group does not reflect some nonspecific stimulation related to handling or sensory stimulations connected to the interaction with a live moving object. The observed neuronal activation therefore appears to relate specifically to the exposure to the female congener and/or to the performance of copulatory behavior. Support for the latter interpretation comes from the observation that the number of FLI cells in the POA of experimental subjects was significantly correlated with the frequency of the cloacal contact movements they performed during the observation period, but not with the frequency of other reproductive behavior, such as neck grabs, mounts, struts or crows (Fig. 9). This could be taken as an indication that it is actually the copulatory behavior sensu stricto (cloacal contact and ejaculation) that induced the neuronal activation and not the mere interaction with a female. Similar results have been observed in a number of mammalian species where it is clearly proven that ejaculation markedly increases Fos expression in the POA [18,24]. A more limited, but nevertheless significant, increase in FLI cells is, however, observed in the POA of mammals after intromissions without ejaculations or even mounts only [18,24,85]. In quail, evidence for a specific association between cloacal contact movements and FLI induction in the POA is only correlative at this point and experiments directly testing the role of cloacal contacts movements on FLI expression should now be carried out by providing males with restricted access to females as has been done in the mammalian studies cited above.
4.4. FLI induction in the ICo
The mesencephalic nucleus intercollicularis (lCo) has been implicated in mediating vocal behavior in birds. In Japanese quail, specifically, electrical stimulation of the ICo elicits a wide variety of natural calls with the medial region of the ICo being the most sensitive to stimulation [4,62]. ICo lesions reduce (but do not completely suppress) crowing [71,72], a typical androgen-dependent male vocalization. Castration completely abolishes crowing which is restored by testosterone [1,3,14,70]. The neural site(s) of testosterone action on crowing probably include the ICo and receptors for both estrogen and androgen are common throughout the nucleus [11,12,79]. Experiments trying to demonstrate directly this implication of the ICo by implanting steroids in the nucleus have met with variable success [59,78], but a recent study indicates by bilateral lesioning of the ICo eliminates crowing in male quail and that testosterone implants in the mesencephalon at the level of the ICo
activate this behavior [88]. Taken together, all these data argue for the ICo being critical for crowing.
At first sight, therefore, it is not unexpected that presentation to a sexually mature female resulted in marked neuronal activation throughout the ICo, as demonstrated by induction of FLI cells. However, the presence of the female did not activate crowing, but actually decreased its mean frequency even though not significantly. Consequently, the behavioral observations provide no obvious interpretation for the Fos induction in the ICo, but two plausible explanations can be offered. It may, first, be a consequence of suppressing crowing which, in this scenario, becomes an active inhibitory process within the ICo. GABA receptors (benzodiazepine binding) exist in the quail's ICo [23] and it is possible that the activation of these inhibitory networks would then be reflected in FLI induction. Alternatively, it could be speculated that ICo activation is a consequence of POA stimulation. The quail's ICo receives sparse afferent projections from the POM [8], so that, in theory, FLI induction in the ICo could come about from activation in the POA. Additional studies would be required to distinguish between these two hypotheses.
4.5. FLI induction in the archistriatum and hyperstriatum
In male mammals, Fos induction is observed in parts of the amygdala following sexual interaction and copulation with a female [18,19,24,44,54,86]. The nucleus taeniae in birds is a part of the archistriatum and is usually considered as the homolog of the mammalian amygdala [48,89]: no Fos induction was detected in this nucleus in the present experiments. This may be a direct consequence of birds not relying on olfactory information in general [75], and during courtship/copulation in particular (see, however, [13]). In contrast, olfactory signals are of paramount importance for these behaviors in rodents and it is suggested that the induction of Fos in the mammalian amygdala is related to the processing of olfactory/vomeronasal information originating from the female [18,24,29,53]. It needs to be noted though that Fos induction in the mammalian amygdala essentially concerns its medial part [86]. It has been claimed that the nucleus taeniae of birds is the specific homolog of the mammalian medial amygdala based both on the similarity of anatomical relationships and on the high density of steroid receptors present in both structures [11,12,76,79,89]. Recent neurochemical studies rather point to a homology between nucleus taeniae and the central amygdala based on the high density of aromatase-immunoreactive cells present in these structures [31,32,41]). The precise homologies between birds and mammals therefore remains questionable and there is possibly an additional anatomical reason for not detecting FLI induction in this nucleus of quail.
It was interesting to note that Fos induction occurred

156 S L. Meddle et al. / Behavioural Brain Research 85 ( 1997) 143-159
in a neighboring part of the archistriatum just lateral to the anterior commissure and occipitomesencephalic tract. The specific homology of this brain region with the amygdaloid complex in mammals is uncertain but it is usually accepted that the caudal and medial avian archistriatum constitute the 'limbic' component of the telencephalon that projects to the hypothalamus, and therefore represent the avian counterpart of the mammalian amygdala. The anterior archistriatum is, by contrast, associated mainly with somatic sensorimotor systems [21,89]. This anterior region is also said to be the only part of the archistriatum that receives fibers from the anterior commissure. The inducible FLI cells were in an area that apparently cuts across this boundary and encompasses parts of the anterior and part of the medial archistriatum. The exact significance of this induction remains to be established, but it might be related to activation of either the limbic system or the sensorimotor systems involved in the performance of copulatory behavior or both. In birds, lesions of the archistriatum produce deficits in avoidance behavior while electrical stimulation of this region affects a number of dependent variables more or less directly related to reproduction, such as avoidance, fear and aggressive behavior or ovulation [42,47,58,60]. It is therefore not entirely unexpected to find an induction of FLI cells in this region.
Specific induction of Fos was also possibly identified in the medial hyper striatum ventrale just lateral to the lateral ventricles. This region has a shape reminiscent of the auditory field L, but it is too rostral to correspond to this sensory area [45,81]. The localization of the induced cells matches fairly well though with an area identified as the intermediate and medial part of the hyperstriatum ventrale (IMHV) that has been associated with filial imprinting in young chicks [38]. The IMHV receives connections from most sensory systems and is also connected directly or indirectly to major outlets of the telencephalon. Through such connections, it may control various aspects of behavioral activity including posture and locomotion or agonistic behavior [38]. It has not been, to our knowledge, associated with the control of copUlation, but the present data suggest that this hypothesis should be tested.
4.6. Comparisons with mammals
Using Fos induction as a neural label has yielded useful information on the neuronal populations that are activated following copulation in mammals. Fos induction occurs in the preoptic area and anterior hypothalamus of several mammalian species as well as quail [18,19,44,54,67,86]. The detailed anatomy of the activated cells appears, however, to be slightly different. In mammals, Fos expression occurs throughout the medial preoptic nucleus of males after ejaculation while the
Fos-positive cells in quail were restricted to a lateral region and never reached the walls of the third ventricle. This area of the POA corresponds to the lateral aspects of the sexually dimorphic POM and/or to that part of the POA immediately lateral to this nucleus. Using immunocytochemistry of aromatase, it is suggested that the quail's POM is homologous to the rat's medial preoptic nucleus [55]. Thus, Fos induction is more restricted and located more laterally within the POA of birds compared with mammals. At more caudal levels, Fos induction has been observed in the BNST of rodents [18,19,44,86] while in quail, the induction is seen more laterally, adjacent to, but only overlapping partly with the BNST.
This difference may relate to the absence in quail of an intromittent organ (penis). Somatosensory feedback from the genital region might, therefore, not be as prominent during copulation in birds as it is in mammals. In rats, lesions of the central tegmental field significantly reduce Fos expression in the POA when associated with lesions of the medial amygdala [18]. Because lesions of the amygdala alone do not affect the number of Fos-immunoreactive cells detected in the POA, these data support the notion that somatosensory feedback from the genital area normally reaching the POA through the central tegmental field is important for the activation of the POA in mammals (although they obviously interact with olfactory information traveling through the amygdala). As mentioned above, it is also clear that interactions with females that do not include ejaculation are less effective in inducing Fos in the rat's POA than full sexual behavior including ejaculation which also points to the importance of genital inputs [18,24]. It is therefore plausible that the more localized signal observed in quail compared with the rat relates to species differences in their copulatory patterns.
In addition, olfactory-vomeronasal information from the female contributes to inducing Fos in the mammalian amygdala, BNST and POA [18,29,53]. No increase in FLI cells was observed in the quail's homolog of the amygdala (nucleus taeniae, caudal and medial archistriatum) with the possible exception of a localized induction at the lateral edges of the anterior commissure-occipitomesencephalic tract that may overlap with the medial archistriatum. In addition, FLI induction in the POA and BNST region was more restricted and concerned more lateral aspects of these areas in quail than in rats. This also may reflect a species difference in the use of olfactory cues during sexual interactions.
In contrast, a strong neuronal activation was detected in the quail's intercollicular nucleus (a part of the avian torus semicircularis) that is a key component of the vocal control pathway in birds [60,62]. The specific homolog of the leo in the mammalian brain is not known ([21]; see, however, [63] for a detailed discussion of this topic), but it is widely accepted that the torus

S L Meddle et al. / Behavioural Brain Research 85 ( 1997) 143-159 157
semicircularis is homologous to the mammalian inferior colliculus [43,63]. No Fos induction has, to our knowledge, been described after copulation in the inferior colliculus of the rat's brain. Neuronal activation, as identified by increased Fos expression or deoxyglucose uptake has been reported in the auditory pathways of mammals including the inferior colliculus after acoustic stimulation or aUdiogenic seizure [22,61,66,69]. It must be noted, however, that in birds, the auditory part of the torus semicircularis is the nucleus mesencephalicus lateralis, pars dorsalis (MLd), a structure located in the middle of the Ico where no FLI cells were detected after copulation (see Fig. 6). The Fos induction seen in mammals after acoustic stimulation therefore does not relate anatomically to the induction seen after copulation in the avian ICo. This activation of the ICo appears specific to the present study. It may reflect the important role played by vocal behavior in the social life of the quail even if changes in vocalizations were not specifically observed during the interactions with females.
Acknowledgement
This work was supported in part by grants from the NIH (MH 50388), the Belgian FRFC (Nbr. 2.9003.91), the French Community of Belgium (Action concertee 93/98-171), the EC Human Capital Program (CT94-0472) and the University of Liege (Fonds Speciaux pour la Recherche) to J.B. It was also supported by the AFRC (now BBSRC) and by a John Simon Guggenheim Memorial Foundation Fellowship, a Benjamin Meaker Foundation Fellowship from the University of Bristol, and a grant from the Underwood Fund (AFRC) to J.C.W. We thank Dr. P.J. Sharp for the gift of the avian Fos antibody.
References
[I] Adkins. E.K., Hormonal baSIS of sexual differentiatIOn in the Japanese quaiL 1. Compo Physiol Psychol. 89 (1975) 61-71.
[2] Adkins. E.K and Adler. N.T.. Hormonal control of behavior in the Japanese quail, 1. Compo Physiol. Psychol., 81 (1972) 27-36.
[3] Adkins. E.K .. Boop, J.J .. Koutmk, D.L.. Morns, J.B. and PniewSki, E.E . Further eVidence that androgen aromatization is essential for the activation of copulation in male quail, Physiol. Behav .. 24 (1980) 441-446.
[4] Armitage. S.E. and Seller. T.J., Midbrain regions involved in call production of Japanese quail, Experientia. 37 (1981) 847-848.
[5] Arnold. A.P. and Gorski. R.A .. Gonadal steroid mduction of structural sex differences in the central nervous system. Annu. Rev. Neurosci .. 7 (1984) 413-442.
[6] Aronm. N .. Sagar. S.M .. Sharp. F.R. and Schwartz, W.J .. Light regulates expression of a Fos-related protein m the rat suprachiasmatic nuclei. Proc. Natl Acad Sci. USA. 87 (1990) 5959-5962.
[7] Balthazart, J .. Hormonal correlates of behaVIOr. In D.S. Farner, J.R. Kmg and K.e. Parkes (Eds.). Avian Biology, Academic Press, New York. 1983, pp. 221 365.
[8] Balthazart. J., Dupiereux, V .. Aste, N .• VigliettJ-Panzica. e.. Barrese, M. and Panzica. G.e.. Afferent and efferent connections of the sexually dimorphiC medial preoptic nucleus of the male quail revealed by in vitro transport of Dil, Cell Tissue Res .. 276 (1994) 455-475.
[9] Balthazart. J .• Evrard, L. and Surlemont, e., Effects of the nonsterOidal aromatase inhibitor. R 76713 on testosterone-induced sexual behavior m the Japanese quail (Coturlllx coturlllx japonica). Horm. Behav. 24 (1990) 510-531.
[!O] Balthazart. J. and Foidart, A .. Neural bases of behaVIOral sex differences in quail. In M. Haug, R.E. Whalen, e. Aron and K.L. Olsen (Eds.), The Development of Sex Differences and Similarities III Behaviour, Kluwer Academic Publishers, Dordrecht, 1993. pp.51-75.
[11] Balthazart, 1.. Foidart. A .. Wilson, E.M. and Ball, G.F., Immunocytochemical localization of androgen receptors in the male songbird and quail brain, I Compo Neurol, 317 (1992) 407-420.
[12] Balthazart, J., Gahr, M. and Surlemont, e., Distribution of estrogen receptors in the brain of the Japanese quail: an immunocytochemical study. Brain Res., 501 (1989) 205-214.
[13] Balthazart, J. and Schoffeniels, E., Pheromones are involved m the control of sexual behaviour in birds, Natunnssenschaften, 66 (1979) 55-56.
[14] Balthazart. J., Schumacher, M. and Ottinger, M.A., Sexual differences m the Japanese quail: behavior. morphology and intracellular metabolism of testosterone, Gen Compo Endocrinol., 51 (1983) 191-207.
[15] Balthazart. J. and Surlemont, e.. Androgen and estrogen action in the preoptic area and activation of copulatory behavior in quail. Physiol. Behav., 48 (1990) 599-609.
[16] Balthazart. J. and Surlemont, C. Copulatory behavior is controlled by the sexually dimorphic nucleus of the quail POA. Brain Res. Bull .. 25 (1990) 7-14.
[17] Balthazart. J .. Surlemont, C and Harada, N .. Aromatase as a cellular marker of testosterone action m the preoptic area. PhyslOl. Behav., 51 (1992) 395-409.
[18] Baum, M.J. and Everitt, B.J., Increased expression of c-fos m the medial preoptic area after mating in male rats. role of afferent mputs from the medial amygdala and midbram central tegmental field. NeurOSCience. 50 (1992) 627-646.
[19] Baum, M.J. and Wersinger. S.R., Equivalent levels of matingmduced neural c-fos Immunoreactivity in castrated male rats given androgen, estrogen. or no steroid replacement. BIOI. Reprod.. 48 (1993) 1341-1347
[20] Beach, F A. and Inman. N.G . Effects of castration and androgen replacement on mating in Japanese quail, Proc Natl. A cad. Sci USA, 54 (1965) 1426-1431.
[21] Breazile. J.E. and Kuenzel, W.J., Systema nervosum centrale. In J.J. Baumel, A.S. King. J.E. Breazlle. H.E. Evans and J C Vanden Berge (Eds.), Handhook of AVian Anatomy: Nomllla Anatomicum Avium, Nuttall Ornithological Club. Cambndge. MA. 1993. pp.493-554.
[22] Brown, M C and LIU. T.S .. Fos-hke Immunoreactivity in central auditory neurons of the mouse. 1. Compo Neurol., 357 (1995) 85-97.
[23] Canonaco, M., Tavolaro, R., Cerra. M.e. and Franzoni. M.F , Distribution of benzodiazepine binding sites in the brain of the male Japanese quail and its correlation to a hormonal control: quantitative autoradIOgraphy study. Neuroendocrinology. 55 (1992) 35-43.
[24] Coolen, L.M .. The Neural OrganizatIOn of Sexual Behavior in the Male Rat. A Functional Neuroanatomlcal Fos-Study, ThesIs, Univ. Nljmegen. 1995.
[25] Crews, D. and Silver, R., Reproductive physiology and behaVIOr mteractions in nonmammalian vertebrates. In N. Adler. D. Pfaff and R.W. Goy (Eds), Handhook oj Behavioral NeurohlOlogy, Vol 7. Plenum Publishmg Co. New York. 1985. pp. 101-182.

158 s.L. Meddle et ai/BehavIOural Brain Research 85 (1997) 143-159
[26] Delville, Y., Sulon, J., Hendrick, J.e. and Balthazart, J., Effect of the presence of females on the pituitary-testicular activity in male Japanese quail (Cotumix coturmx japomca), Gen. Compo Endocrinol., 55 (1984) 295-305.
[27] Domjan, M. and Nash, S., Stimulus control of social behaviour in male Japanese quail, Anim. Behav, 36 (1988) 1006-1015.
[28] Dragunov, M. and Robertson, H.A., Generalized seizures mduce c-fos protem(s) m mammalian neurons, Neurosci Lett, 82 (1987) 157-161.
[29] Fiber, J.M., Adames, P. and Swann, J.M., Pheromones induce C-fos in limbic areas regulating male hamster mating behaviour, Neuroreport, 4 (1993) 871-874.
[30] Fiber, J.M. and Swann, J.M., Testosterone differentially regulates pheromone induced Fos expression in limbic regions of male and female Synan hamsters, Abstracts 27th Conference on Reproductive Behavior, 1995, p. 18.
[31] Foidart, A., Harada, N. and Balthazart, J., Aromatase-immunoreactive cells are present in mouse brain areas that are known to express high levels of aromatase activity, Cell Tissue Res., 280 (1995) 561-574.
[32] Foidart, A., Reid, J.. Absil, P., Yoshimura, N., Harada, N. and Balthazart, J., Critical re-examination of the distribution of aromatase-immunoreactive cells in the quail forebrain using antibodies raised against human placental aromatase and agamst the recombinant quail, mouse or human enzyme, J. Chem Neuroanat.,8 (1995) 267-282.
[33] Follett, B.K., Scanes, e.G. and Cunningham, F.J., A radioimmunoassay for avian luteinizing hormone, J. Endocrinol, 52 (1972) 359-398.
[34] Fujiwara, K.T.. Ashida, K., Nishima, H .. Iba, H., Miyajima, N., Nishizawa, M. and Kawai, S., The chicken c-fos gene: cloning and nucleotide sequence analysis, J. Virol., 61 (1987) 4012-4018.
[35] Graham, J.M. and Desjardins, e., Classical conditioning: induction of lutemizmg hormone and testosterone secretion in anticipation of sexual activity, Science, 210 (1980) 1039-1041.
[36] Guyomarc'h, e. and Guyomarc'h, J.e., La stimulation du developpement sexuel des femelles de caille japonaise, Cotumix coturnix japonica, par des chants de males: mise en evidence de penode privilegiees dans Ie nycthemere, C R Acad. Sci. Paris, Serie III, 295 (1982) 37-40.
[37] Guyomarc'h, J.e. and Thibout, E., Rythmes et cycles dans remission du chant chez la callie japonaise (Ccj), Rev Compo Anim., 3 (1969) 37-49.
[38] Horn, G., Memory, Imprinting, and the Brain, Clarendon Press, Oxford, 1985, pp. 1-315.
[39] Hunt, S.P .. Pini, A. and Evan, G., Induction ofa c-fos like protem in spinal cord neurons following sensory stimulation, Nature, 328 (1987) 632-634.
[40] Hutchison, R.E., Hormonal differentiation of sexual behavior in Japanese quail, Horm. Behav., 11 (1978) 363-387
[41] Jakab, R.L., Horvath, T.L., Leranth, e., Harada, N. and Naftolin, F., Aromatase immunoreactivity in the rat brain: gonadectomy-sensitive hypothalamic neurons and an unresponsive 'limbic ring' of the lateral septum-bed nucleus-amygdala complex, J. Steroid Biochem. Mol. Bioi., 44 (1993) 481-498.
[42] Juhasz, L.P. and Van Tienhoven, A., Effect of electrical stimulation of the telencephalon on ovulation and ovipositIOn in the hen, Am. J. Physiol., 207 (1964) 286-290.
[43] Karten, H.J., The organization of the ascending auditory pathway in the pigeon ( Columba 11Vla). I. Diencephalic projections of the infenor colliculus (nucleus mesencephali lateralis, pars dorsahs). Brain Res, 6 (1967) 409-427.
[44] Kollack, S.S and Newman, S.W., Mating behavior induces selective expression of Fos protein within the chemosensory pathways of the male Syrian hamster brain, NeurOSCI. Lett., 143 (1992) 223-228.
[45] Kuenzel, W.I. and Masson, M., A stereotaxic atlas of the bram
of the chick (Gallus domesticus), The Johns Hopkins University Press, Baltimore. MD, 1988, pp. 1-166.
[46] Macrides, F., Bartke, A., Fernandez, F. and D'Angelo, W., Effects of exposure to vaginal odour and receptive females on plasma testosterone in the male hamster, Neuroendocrinology, 15 (1974) 355-364.
[47] Maley, M.J., Electrical stimulation of agonistic behavior in the mallard, Behaviour, 34 (1969) 138-160.
[48] Martinez-Vargas, M.e., Stumpf, W.E. and Sar, M., Anatomical distribution of estrogen target cells m the avian CNS: a comparison with the mammahan CNS, J. Compo Neurol., 167 (1976) 83-104.
[49] McEwen, B.S., Neural gonadal steroid actions, Science, 211 (1981) 1303-1311.
[50] Meddle, S.L., Photoneuroendocrine Control of Reproduction in Japanese Quail: the Use of the Immediate Early Gene (c-fos) as a Marker of Cell ACllvation, PhD thesis, Univ. of Bristol. UK, Bristol, 1995.
[51] Meddle, S.L. and Follett, B.K., Photoperiodic activatIOn of foslike immunoreactive protein in neurones withm the tuberal hypothalamus of Japanese quail. J. Comp Physiol (A). 176 (1995) 79-89.
[52] Meisel. R.L. and Sachs, B.D, The physiology of male sexual behavIOr. In E. Knobil and J.D. Neill (Eds.), The Physiology of ReproductIOn, Raven Press, New York, Vol. 2, 1994. pp. 3-105.
[53] Moffatt, e.A., Ball, G.F. and Nelson, R.I., The effects of photoperiod on olfactory c-fos expression in prairie voles, Microtus ochrogaster, Bralll Res., 677 (1995) 82-88.
[54] Oboh, A.M .. Paredes, R.G. and Baum, M.I., A sex comparison of increments in FOS immunoreactivity in forebrain neurons of gonadectomized, testosterone-treated rats after mounting an estrous female, Be/wI'. Neural Bioi, 63 (1995) 66-73.
[55] Panzica, G.e., Viglietti-Panzica. e. and Balthazart, J., The sexually dimorphic medial preoptic nucleus of quail: a key brain area mediating steroid action on male sexual behavior, Front. Neuroendocrinol, 17 (1996) 51-125.
[56] Panzica, G.e., Viglietti-Panzica, e., Ca1cagm, M., Anselmettl, G.e., Schumacher, M. and Balthazart, J., Sexual differentiation and hormonal control of the sexually dimorphic preoptic medial nucleus in quail. Brain Res .. 416 (1987) 59-68.
[57] Parkinson, T.J. and Follett, B.K., Thyroidectomy abolishes seasonal testicular cycles of Soay rams. Proc. R Soc. Lond. B , 259 (1995) 1-6.
[58] Philhps. R.E., Wildness in the mallard duck. Effects of bram lesions and stimulation on escape behaVIOr and reproductIOn, J Compo Neurol., 122 (1964) 139-155.
[59] Phillips. R.E. and Barfield, R.I .. Effects of testosterone implants m midbrain vocal areas of capons, Brain Res., 122 (1977) 378-381.
[60] Phillips, R.E. and Youngren, O.M., Bram stimulatIOn and speciestypical behaviour: activities evoked by electncal stimulation of the brains of chickens (Gallus gallus), Alllm Be/wI', 19 (1971) 757-779.
[61] Pierson, M. and Snyder-Keller, A., Development of frequencyselective domains in inferior cohculus of normal and neonatally noise-exposed rats, Brain Res., 636 (1994) 55-67.
[62] Potash, L.M., Vocalizations elicited by electrical brain stimulation in Coturnix coturmx japonica, Behaviour, 31 (1970) 149-167.
[63] Puelles, L., Robles, e., Martinez-de-Ia-Torre, M. and Martinez, S., New subdivisIOn schema for the aVian torus semiclrcularis: neurochemical maps m the chick, J Compo Neurol.. 340 (1994) 98-125.
[64] Purvis. K. and Haynes, N B .. Short term effects of copulatIOn, human chorionic gonadotrophin injection and non-tactile association with a female on testosterone levels in the male rat, J Endocnnol, 60 (1974) 429-439.
[65] Rea, M.A., Light increases Fos-related protein immunoreactivity in the rat SCN, Bralll Res. Bull., 23 (1989) 577-581

S.L Meddle et al I Behavioural Brain Research 85 ( 1997) 143-159 159
[66] Reimer, K., Simultaneous demonstration of Fos-like immunoreactivity and 2-deoxy-glucose uptake in the inferior colliculus of the mouse. Brain Res., 616 (1993) 339-343.
[67] Robertson, G.S., Pfaus. J.G., Atkmson, L.1.. Matsumura, H., Phillips. A.G. and Fiblger, H.e., Sexual behaVIOr increases c-fos expression m the forebrain of the male rat, Brain Res, 564 (1991) 352-357
[68] Sagar, S.M., Sharp, FR. and Curran. T., Expression of c-fos protein in the brain' metabolic mapping at the cellular level, SCIence. 240 (1988) 1328-1331.
[69] Sato, K .. Houtani, T., Ueyama, M., Ikeda, M., Yamashita, T., Kumazawa, T and Sugimoto, T . Identification of brainstem sites with neuronal Fos protein mduced by acoustic stimulation with pure tones. Acta Otolarynol, Supp!., 500 ( 1993) 18-22.
[70] Schumacher, M. and Balthazart, 1.. The effects of testosterone and ItS metabolItes on sexual behavior and morphology in male and female Japanese quail, PhyslOl. Behav, 30 ( 1983) 335-339.
[71] Seller, T.1 , Midbrain vocalization centers m birds, Trends NeurOSel., 4 (1981) 301-303
[72] Seller, T.1 , Control of sound production m birds. In B. Lewis (Ed.), Bwacoustlcs: A CO/llparatll't' Approach, Academic Press, New York. 1983. pp. 93-124
[73] Sharp, P.1 , LI, Q., Talbot, R T , Barker, P., Huskisson, N. and Lea. R W., Identification of hypothalamic nuclei involved in osmoregulation usmg fos immunocytochemistry in the domestic hen (Gallus domestzcus), Ring dove (Streptopelia nsoria), Japanese quail (ColUmlx japonica) and Zebra finch (Taenopygia guttata), Cell Tissue Res, 282 (1995) 351-361
[74] Sharp, R.F., Sagar, S.M , Hicks, K, Lowenstem, 0 and Hlsanaga. K .. C-fo, mRNA. fos and fos related antigen induction by hypertoillc saline and stres" J. Neurosci., II (1991) 2321-2331.
[75] Sturkie, P.O., Avian PhYSIOlogy, Spnnger Verlag, New York, 1986, pp. 1516.
[76] Thompson, R., Goodson, J., RusclO. M. and Adkms-Regan. E, Damage to the telencephalic nucleus taeniae disrupts sexual approach behavior m male Japanese quail, Soc Neurosci. Ahstr, 21 (1995) 1463.
[77] Viglietti-PanzlCa, e., Panzica, G.e., Fiori, M.G., Calcaglll, M., AnselmettI. G.C and Balthazart, 1., A sexually dimorphic nucleus m the quail preoptic area, Neurose; Lett .• 64 ( 1986) 129-134.
[78] Watson. J.T., Neuroanatol/llcal Lo('ali~ation oj Endocrine Control of Reproducllve BehaVIOr 1II the Japanese Quail (Coturnix lapOIll('(l), PhD TheSIS, Cornell Uillversity, Ithaca, NY. 1989.
[79] Watson, J.T. and Adkms-Regan, E .. Neuroanatomical 10calIza-
tion of sex steroid-concentratmg cells m the Japanese quail (Coturnix lapolllca): AutoradIOgraphy With [3H]-testosterone, [3H]estradiol, and [3H ]-dihydrotestosterone, Neuroendocrlllology, 49 (1989) 51-64.
[80] Wersinger, S.R., Baum, M.1. and Erskine, M.S., Mating-induced FOS-like immunoreactIvity in the rat forebrain: a sex comparison and a dimorphic effect of pelvic nerve transection. J NeuroendoCYInol. 5 (1993) 557 -568.
[81] Wild, J.M .. Karten. H.1. and Frost, B.1 .. Connections of the auditory forebrain in the pigeon (Columha livia) , J. Camp Neurol, 337 ( 1993) 32-62.
[82] Wmgfield. J.e., InterrelatIOnships of androgens, aggression, and mating systems. In M Wada, S. Ishii and e.G. Scanes (Eds.), Endocrlllology of Bzrds Molecular to BehaVIOral, Japan SCientific Societies Press and Springer Verlag, Tokyo and Berlin, 1990, pp. 185 205.
[83] Wingfield, J.e., Hegner, R.E., Dufty, A.M. and Ball, G.F , The 'challenge hypotheSIS': theoretical implications for patterns of testosterone secretion, mating systems and breedings strategies, Am. Nat., 136 (1990) 829-846.
[84] Wingfield, J.e., Whaling, e.S. and Marler, P., Commulllcation m vertebrate aggression and reproduction: the role of hormones. In E Knobll and J.D. Neill (Eds.), The PhYSIOlogy of ReproductIOn, 2nd edn., Raven Press, New York, 1994, pp. 303-341.
[85] Witt, D M. and Insel, T R., Increased Fos expressIOn m oxytocm neurons following masculme sexual behaVIOr. J. Neuroendocrlllol. 6 (1994) 13-18.
[86] Wood, R.l. and Newman, S.W., Mating activates androgen receptor-contaming neurons m chemosensory pathways of the male Syrian hamster brain, Bram Res., 614 (1993) 65-77.
[87] Yahr, P., Neural CirCUitry for the Hormonal-Control of Male Sexual BehavIOr. In P.E. Micevych and R.P Hammer, Jr. (Eds.), NeurohlOloglcal Effect" oj Sex Steroid Hormones, Cambridge University Press, Cambndge, 1995, pp. 40- 56.
[88] Yasaki, Y., Matsushima, T. and Aoki, K., The role of the mesencephalIc call region m the Japanese qUail chick. In M. Burrows. T Matheson, P.L. Newland and H. Schuppe (Eds.), Nervous systems and hehaviour Proceedings of the 4th International Congress of Neuroet/zology, Georg Thieme verlag, Stuttgart, New York. 1995,319 pp
[89] Zeier, H. and Karten, H.1., The archlstnatum of the pigeon: organization of afferent and efferent connectIOns, Brain Res. 31 (1971) 313-326




















