
Heterogeneity of growth hormone immunoreactivity in lymphoid tissues and changes during ontogeny in...
10
General and Comparative Endocrinology 144 (2005) 28–37 www.elsevier.com/locate/ygcen 0016-6480/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2005.04.007 Heterogeneity of growth hormone immunoreactivity in lymphoid tissues and changes during ontogeny in domestic fowl M. Luna a,¤ , N. Barraza a , L. Berumen a , M. Carranza a , E. Pedernera b , S. Harvey c , C. Arámburo a a Department of Cellular and Molecular Neurobiology, Institute of Neurobiology, Campus UNAM-UAQ Juriquilla, Universidad Nacional Autónoma de México, A.P. 1-1141, Querétaro, 76001, Mexico b Department of Embriology, Facultad de Medicina, Universidad Nacional Autónoma de México, Ciudad Universitaria, México, DF 04510, Mexico c Department of Physiology, University of Alberta, Edmonton, Alta., Canada T6G 2H7 Received 16 December 2004; revised 12 April 2005; accepted 18 April 2005 Available online 3 June 2005 Abstract Growth hormone (GH) expression is not conWned to the pituitary and occurs in many extrapituitary tissues. Here, we describe the presence of GH-like moieties in chicken lymphoid tissues and particularly in the bursa of Fabricius. GH-immunoreactivity (GH-IR), determined by ELISA, was found in thymus, spleen, and in bursa of young chickens, but at concentrations <1% of those in the pituitary gland. Although the GH concentration in the spleen and bursa was approximately 0.82 and 0.23% of that in the pituitary at 9-weeks of age, because of their greater mass, the total GH content in the spleen, bursa, and in thymus were 236, 5.18, and 31.5%, respectively, of that in the pituitary gland. This GH-IR was associated with several proteins of diVerent molecular size, as in the pituitary gland, when analyzed by SDS–PAGE under reducing conditions. While most of the GH-IR in the pituitary was associated with the 26 kDa monomer (40%), the putatively glycosylated 29 kDa variant (16%), the 52 kDa dimer (14%) and the 15 kDa submonomeric isoform (16%), GH-IR in the lymphoid tissues was primarily associated (27–36%) with a 17kDa moiety, although bands of 14, 26, 29, 32, 37, 40, and 52 kDa were also identi Wed in these tissues. The heterogeneity pattern and relative abundance of bursal GH-IR bands were deter- mined during development between embryonic day 13 (ED13) and 9-weeks of age. The relative proportion of the 17 kDa GH-like band was higher (45–58%) in posthatched birds than in the 15 and 18-day old embryos (21 and 19%, respectively). The 26kDa isoform was minimally present in embryos (<4% of total GH-IR) but in posthatched chicks it increased to 12–20%. Conversely, while GH-IR of 37, 40, and 45 kDa were abundantly present in embryonic bursa (»30% at ED13 and »52–55% at ED15 and ED18, respectively), in neona- tal chicks and juveniles they accounted for less than 5%. These ontogenic changes were comparable to those previously reported for sim- ilar GH-IR proteins in the chicken testis during development. In summary, these results demonstrate age-related and tissue-speciWc changes in the content and composition of GH in immune tissues of the chicken, in which GH is likely to be an autocrine or paracrine regulator. 2005 Elsevier Inc. All rights reserved. Keywords: Chicken growth hormone; Spleen; Thymus; Bursa of Fabricius; GH heterogeneity; Immune GH 1. Introduction It is now well established that growth hormone (GH) is expressed in immune tissues. GH mRNA identical to that in the pituitary gland has been demonstrated in rat (Rohn and Weigent, 1995) and human (Hattori et al., 1999; Liu et al., 1997; Melen et al., 1997; Palmetshofer et al., 1995; Wu et al., 1996; Yang et al., 1999) leukocytes, in canine (Lantinga van Leeuwen et al., 2000) and bovine (Chen et al., 1997) lymphoid tissues and in the thymus, spleen and bursa of Fabricius of chickens (Render et al., 1995a). * Corresponding author. Fax: +52 44 22 3810 04. E-mail address: [email protected] (M. Luna).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Heterogeneity of growth hormone immunoreactivity in lymphoid tissues and changes during ontogeny in...

General and Comparative Endocrinology 144 (2005) 28–37
www.elsevier.com/locate/ygcen
Heterogeneity of growth hormone immunoreactivity in lymphoid tissues and changes during ontogeny in domestic fowl
M. Luna a,¤, N. Barraza a, L. Berumen a, M. Carranza a, E. Pedernera b, S. Harvey c, C. Arámburo a
a Department of Cellular and Molecular Neurobiology, Institute of Neurobiology, Campus UNAM-UAQ Juriquilla, Universidad Nacional Autónoma de México, A.P. 1-1141, Querétaro, 76001, Mexico
b Department of Embriology, Facultad de Medicina, Universidad Nacional Autónoma de México, Ciudad Universitaria, México, DF 04510, Mexicoc Department of Physiology, University of Alberta, Edmonton, Alta., Canada T6G 2H7
Received 16 December 2004; revised 12 April 2005; accepted 18 April 2005Available online 3 June 2005
Abstract
Growth hormone (GH) expression is not conWned to the pituitary and occurs in many extrapituitary tissues. Here, we describe thepresence of GH-like moieties in chicken lymphoid tissues and particularly in the bursa of Fabricius. GH-immunoreactivity (GH-IR),determined by ELISA, was found in thymus, spleen, and in bursa of young chickens, but at concentrations <1% of those in the pituitarygland. Although the GH concentration in the spleen and bursa was approximately 0.82 and 0.23% of that in the pituitary at 9-weeks ofage, because of their greater mass, the total GH content in the spleen, bursa, and in thymus were 236, 5.18, and 31.5%, respectively, ofthat in the pituitary gland. This GH-IR was associated with several proteins of diVerent molecular size, as in the pituitary gland, whenanalyzed by SDS–PAGE under reducing conditions. While most of the GH-IR in the pituitary was associated with the 26 kDamonomer (40%), the putatively glycosylated 29 kDa variant (16%), the 52 kDa dimer (14%) and the 15 kDa submonomeric isoform(16%), GH-IR in the lymphoid tissues was primarily associated (27–36%) with a 17 kDa moiety, although bands of 14, 26, 29, 32, 37, 40,and 52 kDa were also identiWed in these tissues. The heterogeneity pattern and relative abundance of bursal GH-IR bands were deter-mined during development between embryonic day 13 (ED13) and 9-weeks of age. The relative proportion of the 17 kDa GH-like bandwas higher (45–58%) in posthatched birds than in the 15 and 18-day old embryos (21 and 19%, respectively). The 26 kDa isoform wasminimally present in embryos (<4% of total GH-IR) but in posthatched chicks it increased to 12–20%. Conversely, while GH-IR of 37,40, and 45 kDa were abundantly present in embryonic bursa (»30% at ED13 and »52–55% at ED15 and ED18, respectively), in neona-tal chicks and juveniles they accounted for less than 5%. These ontogenic changes were comparable to those previously reported for sim-ilar GH-IR proteins in the chicken testis during development. In summary, these results demonstrate age-related and tissue-speciWcchanges in the content and composition of GH in immune tissues of the chicken, in which GH is likely to be an autocrine or paracrineregulator. 2005 Elsevier Inc. All rights reserved.
Keywords: Chicken growth hormone; Spleen; Thymus; Bursa of Fabricius; GH heterogeneity; Immune GH
1. Introduction
It is now well established that growth hormone (GH) isexpressed in immune tissues. GH mRNA identical to that
* Corresponding author. Fax: +52 44 22 3810 04.E-mail address: [email protected] (M. Luna).
0016-6480/$ - see front matter 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.ygcen.2005.04.007
in the pituitary gland has been demonstrated in rat (Rohnand Weigent, 1995) and human (Hattori et al., 1999; Liuet al., 1997; Melen et al., 1997; Palmetshofer et al., 1995;Wu et al., 1996; Yang et al., 1999) leukocytes, in canine(Lantinga van Leeuwen et al., 2000) and bovine (Chenet al., 1997) lymphoid tissues and in the thymus, spleenand bursa of Fabricius of chickens (Render et al., 1995a).

M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37 29
As in the pituitary, (22 kDa) GH is similarly present innumerous mammalian lymphoid tissues and lymphoidcells (Weigent et al., 1988; Weigent and Blalock, 1989).Other GH variants of larger (>300,000; Weigent et al.,1988) and smaller (20 kDa, Hattori et al., 1999; Lantingavan Leeuwen et al., 2000; Palmetshofer et al., 1995)molecular size are, however, also present in human andrat immune cells, and a 14 kDa variant may also betranslated (Palmetshofer et al., 1995). This heterogeneityis likely to reXect diVerences in GH bioactivity (Arám-buro et al., 1989c, 1990, 2001b) and likely provides amechanism for subtle modulation of GH action withinthe immune system.
Whilst monomer GH is present in immune tissues ofjuvenile chickens (Render et al., 1995a), the possiblepresence of other GH variants is uncertain. This possi-bility has therefore been examined in the present study,in which ontogenic changes in the relative abundanceof immune GH moieties has also been quantitativelydetermined in the bursa of Fabricius, since ontogenicchanges occur in the relative abundance of GHmoieties in the chicken pituitary (Arámburo et al.,2000) and other extrapituitary (testis) tissues (Lunaet al., 2004).
2. Materials and methods
2.1. Tissues
Broiler fowl bursas were collected during embryogen-esis (embryonic day (ED) 13 and 15 of the 21 dayincubation period), after hatch (day 1), and before(weeks 4–9) puberty. The tissues were immediately fro-zen in dry ice and stored at ¡70 °C until used for assay.For comparison, thymus, spleen, and pituitary tissueswere also collected from some birds.
2.2. Tissue extracts
Frozen tissues were homogenized (1 g/10 ml) in anEDTA-free protease inhibitor cocktail (Mini-Complete,Roche Mannheim, Germany), containing 1 mM PMSF,pH 9.0. The homogenates were then agitated on a mag-netic stirrer for 2 h at 4 °C and centrifuged (12,000 rpm at4 °C for 15 min). The supernatants were collected and theprotein content determined by the Bradford micro-method (Bio-Rad, Hercules, CA).
2.3. ELISA
The presence of GH-immunoreactivity in the tissueextracts were investigated using an indirect enzyme-linked immunosorbant assay (ELISA), similar to thatdescribed by Martínez-Coria et al. (2002). BrieXy, 96-well microtiter plates (Immulon 2HB, Chantully, VA)
were coated overnight at 4 °C, with 12 ng recombinantchicken GH (rcGH) in 100�l in 0.1 M carbonate buVer,pH 10.3. The plates were extensively washed with TPBS(0.01 M sodium phosphate, 0.15 mM NaCl, 0.05% (w/v)Tween-20) for Wve times using an automatic microplateimmunowasher (Bio-Rad, Mod. 1250, Hercules, CA).This procedure was performed after each incubationstep. Tissue extracts or serial dilutions of rcGH (5120–5 ng/ml) in TPBS with 1% (w/v) non-fat dried milk werethen incubated for 16 h with 100 �l primary antibodyCAP-1 (at a Wnal concentration of 1:100,000) raised inrabbits against native pituitary GH (Arámburo et al.,1989b). This antibody is speciWc for GH and has nocross reactivity (<0.001%) with any other pituitary hor-mones (Arámburo et al., 1989b). The samples and stan-dards (100 �l) were then added to the coated wells andincubated for a further 2 h at room temperature. Horse-radish peroxidase-anti-rabbit IgG conjugate (Bio-Rad)was then added (at a dilution of 1:3,000 in 5% (w/v) non-fat dried milk, in 0.1 M TPBS, pH 7.0) and incubated for2 h at room temperature. Bound secondary antibodieswere then developed by reaction with 2,2�-amino-di-[3-ethylbenzothiazoline sulfate] substrate (Roche Mann-heim, Germany). The plates were read 1 h later, in anautomatic ELISA Microplate Reader (Bio-Rad), at awavelength of 405 nm. The assay has a sensitivity of 2 ng/well (20 ng/ml) and inter-assay and intra-assay coeY-cients of variation of <4% (Martínez-Coria et al., 2002).
2.4. Western blotting
GH-immunoreactivity in tissue extracts were alsoinvestigated by one-dimensional sodium dodecyl sul-phate–polyacrylamide gel electrophoresis (SDS–PAGE)followed by Western blotting. Samples (containing 30�gprotein) were analyzed by in 1.0 mm thick, 6 cm long,15% gels; using the buVer system of Laemmli (1970) in amini-Protean II cell (Bio-Rad). Samples were electro-phoresed, under reducing conditions (in the presence of5% (w/v) 2-mercaptoethanol) and under non-reducingconditions. After electrophoresis, the proteins wereequilibrated in transfer buVer (25 mM Tris–HCl,192 mM glycine, and 20% (v/v) methanol, pH 8.3) for30–60 min and electrotransferred (at 200 mA for 60 min)to nitrocellulose membranes (Bio-Rad). After transfer,the membranes were washed with 30 mM Tris, 500 mMNaCl, pH 7.5 (TBS) for 5 min, then blocked with 5% (w/v) non-fat dried milk (Bio-Rad) in TBS, for 2 h at roomtemperature. After washing the membranes with TPBSfor 15 min, they were incubated overnight at room tem-perature with rabbit-anti-chicken GH antisera (CAP-1,Arámburo et al., 1989a,b, 1990), diluted 1:10,000–1:100,000 in TPBS (1% (w/v) non-fat dried milk in 0.05%(v/v) Tween-20 and TBS). The membranes were thenrinsed three times (for 15 min) in TTBS and antibodybinding was visualized using anti-rabbit IgG horseradish
30 M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37
peroxidase conjugate (Bio-Rad), diluted 1:3000 in1% (w/v) non-fat dried milk in TTBS for 2 h.Immunoreactive bands were developed by incubation ofthe membranes in ECL chemiluminescent reagent(Amersham Pharmacia, Buckinghamshire, UK) for1 min and exposed to Kodak Biomax ML Wlm. Mono-meric GH runs as a 22 kDa band under non-reducingconditions and as a 26 kDa band under reducing condi-tions due to the unfolding of the protein provoked bythe increase in the Stoke’s radium after disulWde reduc-tion by 2-mercaptoethanol (Arámburo et al., 1989a).
2.5. Densitometric analysis
The number, amount, and relative proportion ofimmunoreactive GH moieties was determined by densi-tometric analysis after digitalization of the luminogramsin a HP scanner, using IP LabGel 2.2 software (Scanalyt-ics, Fairfax, VA, USA), as previously described (Arám-buro et al., 2000; Martínez-Coria et al., 2002). This assayhas a sensitivity <2 ng for the GH standard and an inter-assay coeYcient of variation of 8.5%.
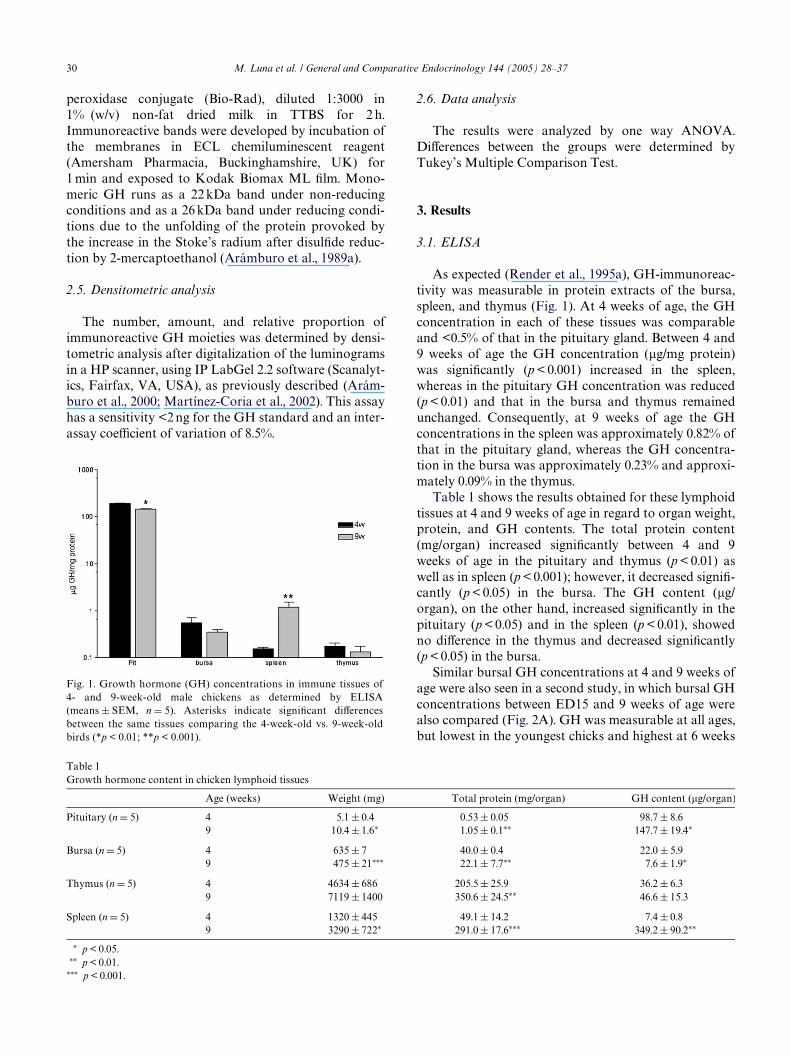
Fig. 1. Growth hormone (GH) concentrations in immune tissues of4- and 9-week-old male chickens as determined by ELISA(means § SEM, n D 5). Asterisks indicate signiWcant diVerencesbetween the same tissues comparing the 4-week-old vs. 9-week-oldbirds (*p < 0.01; **p < 0.001).
2.6. Data analysis
The results were analyzed by one way ANOVA.DiVerences between the groups were determined byTukey’s Multiple Comparison Test.
3. Results
3.1. ELISA
As expected (Render et al., 1995a), GH-immunoreac-tivity was measurable in protein extracts of the bursa,spleen, and thymus (Fig. 1). At 4 weeks of age, the GHconcentration in each of these tissues was comparableand <0.5% of that in the pituitary gland. Between 4 and9 weeks of age the GH concentration (�g/mg protein)was signiWcantly (p < 0.001) increased in the spleen,whereas in the pituitary GH concentration was reduced(p < 0.01) and that in the bursa and thymus remainedunchanged. Consequently, at 9 weeks of age the GHconcentrations in the spleen was approximately 0.82% ofthat in the pituitary gland, whereas the GH concentra-tion in the bursa was approximately 0.23% and approxi-mately 0.09% in the thymus.
Table 1 shows the results obtained for these lymphoidtissues at 4 and 9 weeks of age in regard to organ weight,protein, and GH contents. The total protein content(mg/organ) increased signiWcantly between 4 and 9weeks of age in the pituitary and thymus (p < 0.01) aswell as in spleen (p < 0.001); however, it decreased signiW-cantly (p < 0.05) in the bursa. The GH content (�g/organ), on the other hand, increased signiWcantly in thepituitary (p < 0.05) and in the spleen (p < 0.01), showedno diVerence in the thymus and decreased signiWcantly(p < 0.05) in the bursa.
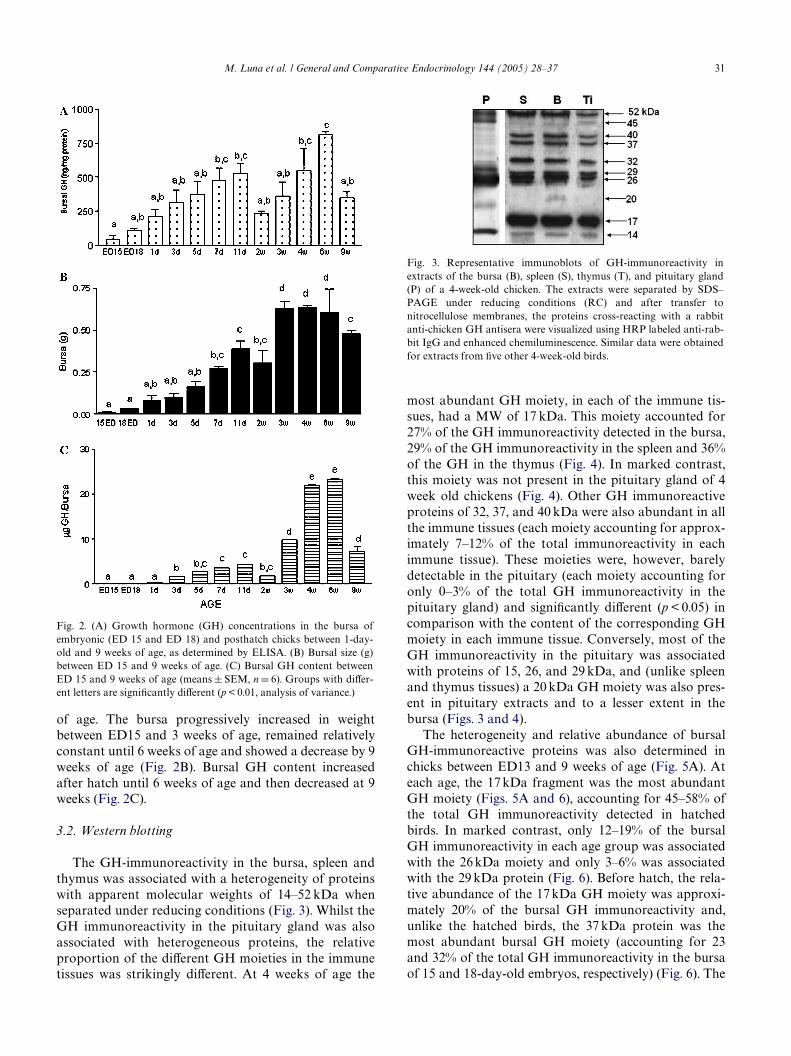
Similar bursal GH concentrations at 4 and 9 weeks ofage were also seen in a second study, in which bursal GHconcentrations between ED15 and 9 weeks of age werealso compared (Fig. 2A). GH was measurable at all ages,but lowest in the youngest chicks and highest at 6 weeks
Table 1Growth hormone content in chicken lymphoid tissues
¤ p < 0.05.¤¤ p < 0.01.
¤¤¤ p < 0.001.
Age (weeks) Weight (mg) Total protein (mg/organ) GH content (�g/organ)
Pituitary (n D 5) 4 5.1 § 0.4 0.53 § 0.05 98.7 § 8.69 10.4 § 1.6¤ 1.05 § 0.1¤¤ 147.7 § 19.4¤
Bursa (n D 5) 4 635 § 7 40.0 § 0.4 22.0 § 5.99 475 § 21¤¤¤ 22.1 § 7.7¤¤ 7.6 § 1.9¤
Thymus (n D 5) 4 4634 § 686 205.5 § 25.9 36.2 § 6.39 7119 § 1400 350.6 § 24.5¤¤ 46.6 § 15.3
Spleen (n D 5) 4 1320 § 445 49.1 § 14.2 7.4 § 0.89 3290 § 722¤ 291.0 § 17.6¤¤¤ 349.2 § 90.2¤¤
ative Endocrinology 144 (2005) 28–37 31
of age. The bursa progressively increased in weightbetween ED15 and 3 weeks of age, remained relativelyconstant until 6 weeks of age and showed a decrease by 9weeks of age (Fig. 2B). Bursal GH content increasedafter hatch until 6 weeks of age and then decreased at 9weeks (Fig. 2C).
3.2. Western blotting
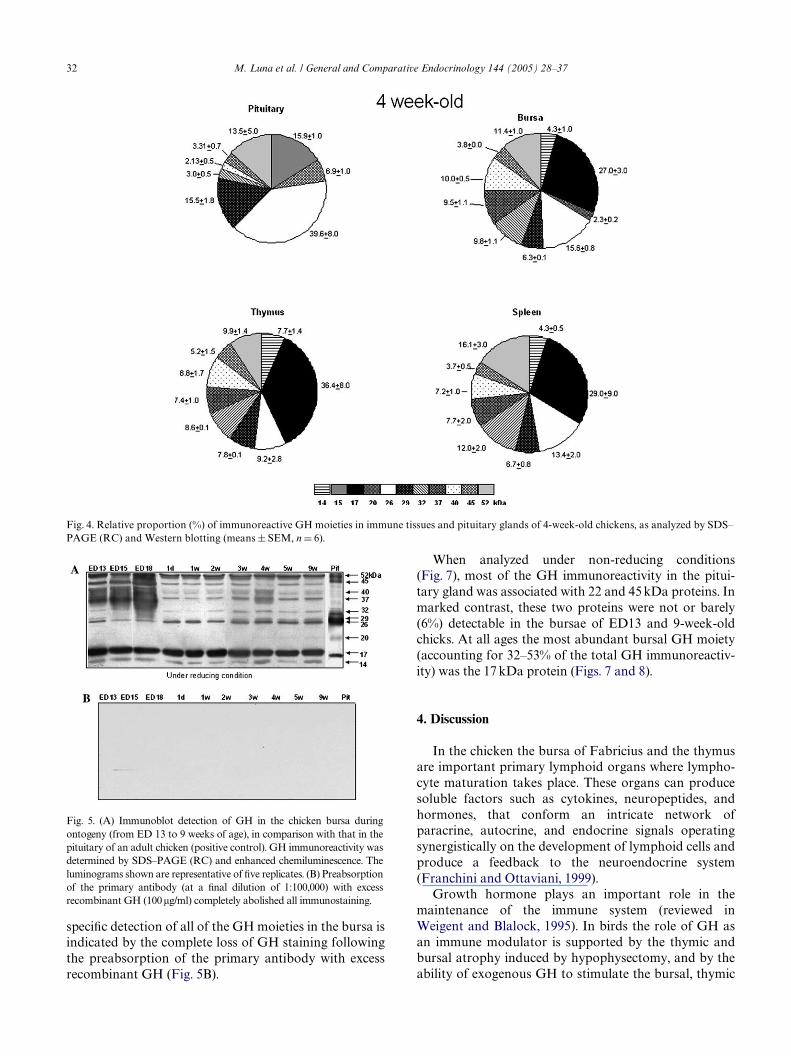
The GH-immunoreactivity in the bursa, spleen andthymus was associated with a heterogeneity of proteinswith apparent molecular weights of 14–52 kDa whenseparated under reducing conditions (Fig. 3). Whilst theGH immunoreactivity in the pituitary gland was alsoassociated with heterogeneous proteins, the relativeproportion of the diVerent GH moieties in the immunetissues was strikingly diVerent. At 4 weeks of age the
Fig. 2. (A) Growth hormone (GH) concentrations in the bursa ofembryonic (ED 15 and ED 18) and posthatch chicks between 1-day-old and 9 weeks of age, as determined by ELISA. (B) Bursal size (g)between ED 15 and 9 weeks of age. (C) Bursal GH content betweenED 15 and 9 weeks of age (means § SEM, n D 6). Groups with diVer-ent letters are signiWcantly diVerent (p < 0.01, analysis of variance.)
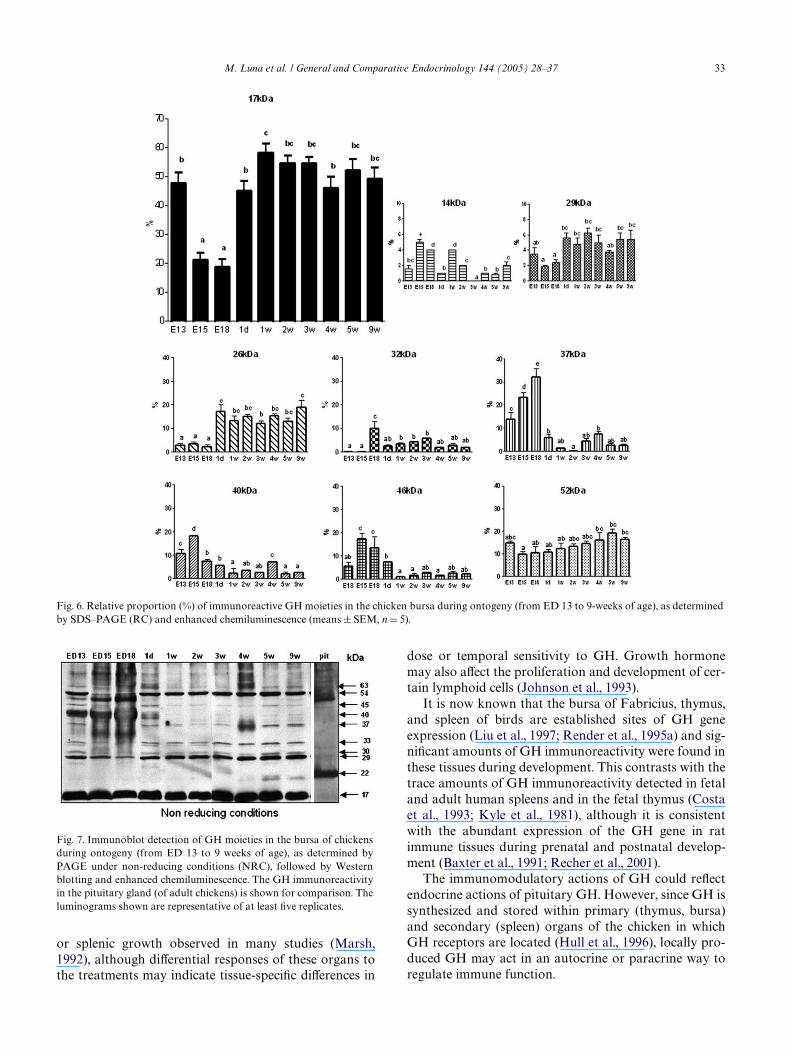
most abundant GH moiety, in each of the immune tis-sues, had a MW of 17 kDa. This moiety accounted for27% of the GH immunoreactivity detected in the bursa,29% of the GH immunoreactivity in the spleen and 36%of the GH in the thymus (Fig. 4). In marked contrast,this moiety was not present in the pituitary gland of 4week old chickens (Fig. 4). Other GH immunoreactiveproteins of 32, 37, and 40 kDa were also abundant in allthe immune tissues (each moiety accounting for approx-imately 7–12% of the total immunoreactivity in eachimmune tissue). These moieties were, however, barelydetectable in the pituitary (each moiety accounting foronly 0–3% of the total GH immunoreactivity in thepituitary gland) and signiWcantly diVerent (p < 0.05) incomparison with the content of the corresponding GHmoiety in each immune tissue. Conversely, most of theGH immunoreactivity in the pituitary was associatedwith proteins of 15, 26, and 29 kDa, and (unlike spleenand thymus tissues) a 20 kDa GH moiety was also pres-ent in pituitary extracts and to a lesser extent in thebursa (Figs. 3 and 4).
The heterogeneity and relative abundance of bursalGH-immunoreactive proteins was also determined inchicks between ED13 and 9 weeks of age (Fig. 5A). Ateach age, the 17 kDa fragment was the most abundantGH moiety (Figs. 5A and 6), accounting for 45–58% ofthe total GH immunoreactivity detected in hatchedbirds. In marked contrast, only 12–19% of the bursalGH immunoreactivity in each age group was associatedwith the 26 kDa moiety and only 3–6% was associatedwith the 29 kDa protein (Fig. 6). Before hatch, the rela-tive abundance of the 17 kDa GH moiety was approxi-mately 20% of the bursal GH immunoreactivity and,unlike the hatched birds, the 37 kDa protein was themost abundant bursal GH moiety (accounting for 23and 32% of the total GH immunoreactivity in the bursaof 15 and 18-day-old embryos, respectively) (Fig. 6). The
Fig. 3. Representative immunoblots of GH-immunoreactivity inextracts of the bursa (B), spleen (S), thymus (T), and pituitary gland(P) of a 4-week-old chicken. The extracts were separated by SDS–PAGE under reducing conditions (RC) and after transfer tonitrocellulose membranes, the proteins cross-reacting with a rabbitanti-chicken GH antisera were visualized using HRP labeled anti-rab-bit IgG and enhanced chemiluminescence. Similar data were obtainedfor extracts from Wve other 4-week-old birds.
M. Luna et al. / General and Compar
32 M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37
speciWc detection of all of the GH moieties in the bursa isindicated by the complete loss of GH staining followingthe preabsorption of the primary antibody with excessrecombinant GH (Fig. 5B).
Fig. 5. (A) Immunoblot detection of GH in the chicken bursa duringontogeny (from ED 13 to 9 weeks of age), in comparison with that in thepituitary of an adult chicken (positive control). GH immunoreactivity wasdetermined by SDS–PAGE (RC) and enhanced chemiluminescence. Theluminograms shown are representative of Wve replicates. (B) Preabsorptionof the primary antibody (at a Wnal dilution of 1:100,000) with excessrecombinant GH (100�g/ml) completely abolished all immunostaining.
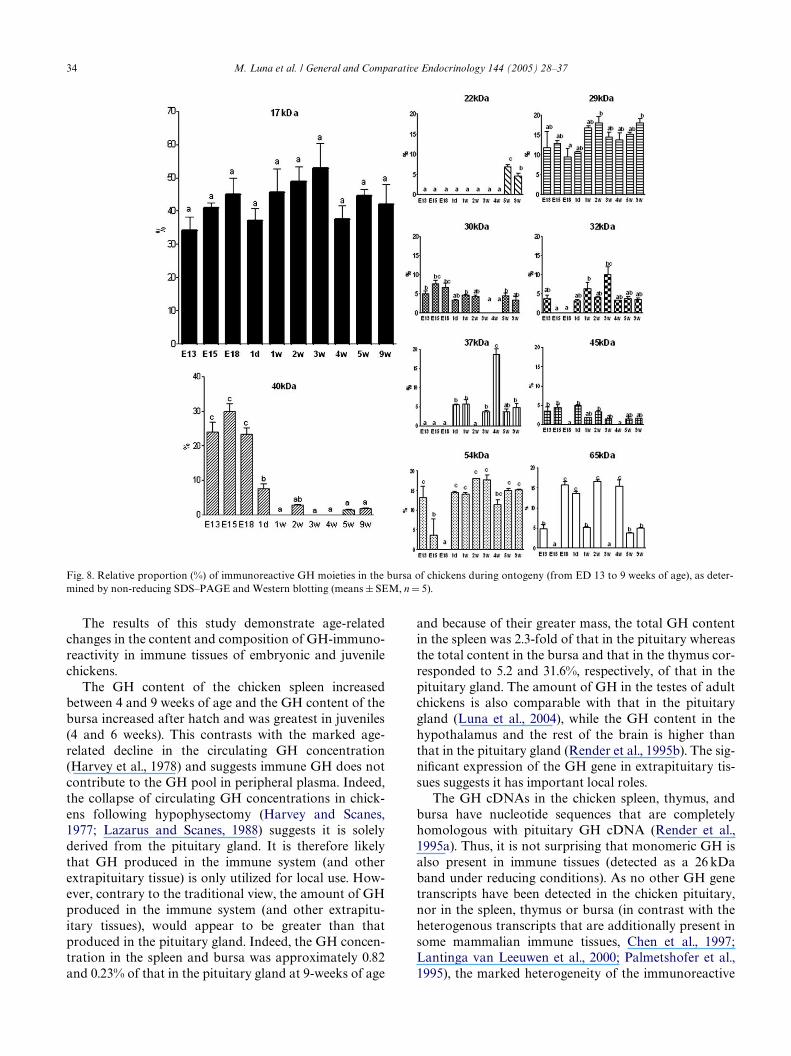
When analyzed under non-reducing conditions(Fig. 7), most of the GH immunoreactivity in the pitui-tary gland was associated with 22 and 45 kDa proteins. Inmarked contrast, these two proteins were not or barely(6%) detectable in the bursae of ED13 and 9-week-oldchicks. At all ages the most abundant bursal GH moiety(accounting for 32–53% of the total GH immunoreactiv-ity) was the 17 kDa protein (Figs. 7 and 8).
4. Discussion
In the chicken the bursa of Fabricius and the thymusare important primary lymphoid organs where lympho-cyte maturation takes place. These organs can producesoluble factors such as cytokines, neuropeptides, andhormones, that conform an intricate network ofparacrine, autocrine, and endocrine signals operatingsynergistically on the development of lymphoid cells andproduce a feedback to the neuroendocrine system(Franchini and Ottaviani, 1999).
Growth hormone plays an important role in themaintenance of the immune system (reviewed inWeigent and Blalock, 1995). In birds the role of GH asan immune modulator is supported by the thymic andbursal atrophy induced by hypophysectomy, and by theability of exogenous GH to stimulate the bursal, thymic
Fig. 4. Relative proportion (%) of immunoreactive GH moieties in immune tissues and pituitary glands of 4-week-old chickens, as analyzed by SDS–PAGE (RC) and Western blotting (means § SEM, n D 6).
M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37 33
or splenic growth observed in many studies (Marsh,1992), although diVerential responses of these organs tothe treatments may indicate tissue-speciWc diVerences in
Fig. 7. Immunoblot detection of GH moieties in the bursa of chickensduring ontogeny (from ED 13 to 9 weeks of age), as determined byPAGE under non-reducing conditions (NRC), followed by Westernblotting and enhanced chemiluminescence. The GH immunoreactivityin the pituitary gland (of adult chickens) is shown for comparison. Theluminograms shown are representative of at least Wve replicates.
dose or temporal sensitivity to GH. Growth hormonemay also aVect the proliferation and development of cer-tain lymphoid cells (Johnson et al., 1993).
It is now known that the bursa of Fabricius, thymus,and spleen of birds are established sites of GH geneexpression (Liu et al., 1997; Render et al., 1995a) and sig-niWcant amounts of GH immunoreactivity were found inthese tissues during development. This contrasts with thetrace amounts of GH immunoreactivity detected in fetaland adult human spleens and in the fetal thymus (Costaet al., 1993; Kyle et al., 1981), although it is consistentwith the abundant expression of the GH gene in ratimmune tissues during prenatal and postnatal develop-ment (Baxter et al., 1991; Recher et al., 2001).
The immunomodulatory actions of GH could reXectendocrine actions of pituitary GH. However, since GH issynthesized and stored within primary (thymus, bursa)and secondary (spleen) organs of the chicken in whichGH receptors are located (Hull et al., 1996), locally pro-duced GH may act in an autocrine or paracrine way toregulate immune function.
Fig. 6. Relative proportion (%) of immunoreactive GH moieties in the chicken bursa during ontogeny (from ED 13 to 9-weeks of age), as determinedby SDS–PAGE (RC) and enhanced chemiluminescence (means § SEM, n D 5).
34 M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37
The results of this study demonstrate age-relatedchanges in the content and composition of GH-immuno-reactivity in immune tissues of embryonic and juvenilechickens.
The GH content of the chicken spleen increasedbetween 4 and 9 weeks of age and the GH content of thebursa increased after hatch and was greatest in juveniles(4 and 6 weeks). This contrasts with the marked age-related decline in the circulating GH concentration(Harvey et al., 1978) and suggests immune GH does notcontribute to the GH pool in peripheral plasma. Indeed,the collapse of circulating GH concentrations in chick-ens following hypophysectomy (Harvey and Scanes,1977; Lazarus and Scanes, 1988) suggests it is solelyderived from the pituitary gland. It is therefore likelythat GH produced in the immune system (and otherextrapituitary tissue) is only utilized for local use. How-ever, contrary to the traditional view, the amount of GHproduced in the immune system (and other extrapitu-itary tissues), would appear to be greater than thatproduced in the pituitary gland. Indeed, the GH concen-tration in the spleen and bursa was approximately 0.82and 0.23% of that in the pituitary gland at 9-weeks of age
and because of their greater mass, the total GH contentin the spleen was 2.3-fold of that in the pituitary whereasthe total content in the bursa and that in the thymus cor-responded to 5.2 and 31.6%, respectively, of that in thepituitary gland. The amount of GH in the testes of adultchickens is also comparable with that in the pituitarygland (Luna et al., 2004), while the GH content in thehypothalamus and the rest of the brain is higher thanthat in the pituitary gland (Render et al., 1995b). The sig-niWcant expression of the GH gene in extrapituitary tis-sues suggests it has important local roles.
The GH cDNAs in the chicken spleen, thymus, andbursa have nucleotide sequences that are completelyhomologous with pituitary GH cDNA (Render et al.,1995a). Thus, it is not surprising that monomeric GH isalso present in immune tissues (detected as a 26 kDaband under reducing conditions). As no other GH genetranscripts have been detected in the chicken pituitary,nor in the spleen, thymus or bursa (in contrast with theheterogenous transcripts that are additionally present insome mammalian immune tissues, Chen et al., 1997;Lantinga van Leeuwen et al., 2000; Palmetshofer et al.,1995), the marked heterogeneity of the immunoreactive
Fig. 8. Relative proportion (%) of immunoreactive GH moieties in the bursa of chickens during ontogeny (from ED 13 to 9 weeks of age), as deter-mined by non-reducing SDS–PAGE and Western blotting (means § SEM, n D 5).

M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37 35
GH proteins detected in these immune tissues is likely toresult from posttranslational diVerences in the process-ing of the GH mRNA. It has been shown that GHfunctional diversity can be explained by its molecularheterogeneity as some GH variants express particularbioactivities (Arámburo et al., 1989c, 1990, 2001b;Martínez-Coria et al., 2002). This is also the case forPRL where posttranslational processing of the matureprotein results in variants that have speciWc activities(Clapp et al., 1993; Corbacho et al., 2000a,b, 2002;González et al., 2004). The possibility of regulating theexpression and modiWcation of these proteins in atissue-speciWc manner would provide a mechanism toamplify the physiological capabilities of these peptidemessengers.
The eight proteins with GH immunoreactivity in thechicken pituitary, ranging in MW size between 15 and52 kDa, are thus likely to include translational variants(e.g., glycosylated and phosphorylated forms), proteo-lytic cleavage fragments (e.g., the 15 kDa variant), pro-tein bound forms and oligomers (including dimer andtrimer forms) (Arámburo et al., 1991, 2001a). It is ofinterest that seven of these proteins (the 15 kDa variantexcepted) are present in the thymus, spleen and bursa,although the 26 kDa monomer is not the most abundantGH moiety in these immune tissues. The absence of the15 kDa fragment in these tissues suggests the full-lengthmonomer undergo tissue-speciWc processing and thethrombin-like protease that cleaves monomer GHbetween Arg133 and Gly134. (Arámburo et al., 2001b)may be absent or deWcient in immune tissues. The15 kDa fragment is, similarly, barely detectable inextracts of the chicken testis during development (Lunaet al., 2004) and suggests this variant is primarily pro-duced in the pituitary gland.
In further contrast with the pituitary, proteins of 14and 32 kDa were additionally present, in all the immunetissues, in which most of the GH immunoreactivity wasassociated with a 17 kDa variant that was only present intrace amounts in pituitary extracts. It is of interest thatthe 17 kDa moiety is also the most abundant GH variantin testicular extracts, in which 14 and 32 kDa forms aresimilarly present (Luna et al., 2004). Processing of GHmRNA or the coded monomeric protein would thusappear to diVer between pituitary and extrapituitary tis-sues. It may also diVer during ontogeny in each tissue.
The bursa grows rapidly during embryogenesis andearly weeks after hatching and then decreases in size.Bursal development and involution varies greatly indiVerent chicken strains (Milicévic et al., 1986). In thisstudy, the bursal weight was maximal between 3 and 6weeks of age and by 9 weeks of age it started todecrease. Both the GH concentration and contentdecreased concomitantly at this age. It is known thatthe bursa is one of the most important sites of apoptoticcell death in the lymphoid system, showing rates several
fold higher than in the thymus (Paramithiotis et al.,1995). Recently, it has been shown that GH of retinalorigin in chick embryos may act as an antiapoptoticfactor in the retinal ganglion cells (Sanders et al., 2005).It would thus be interesting to speculate that locallysynthesized GH in the bursa may be involved in themodulation of apoptosis contributing to regulate thegrowth and involution of this organ.
In the bursa, the 26 kDa GH moiety was minimallypresent in the embryo (<4% of total GHimmunoreactivity), although it accounted for approxi-mately 12–20% of total GH immunoreactivity inhatched birds, neonates and juveniles. Conversely,while GH proteins of 37, 40, and 45 kDa were abun-dantly present in the bursa of embryonic chicks (collec-tively accounting for 30% of bursal GHimmunoreactivity at ED 13 and >55% of bursal GHimmunoreactivity at ED 15 and ED 18); these proteinswere of lesser abundance neonatally (collectivelyaccounting for <18% of bursal immunoreactivity at 1day and <5% of bursal immunoreactivity at 1 and 2weeks of age) and in juveniles (collectively accountingfor <15% of bursal GH immunoreactivity between 3and 9 weeks of age). These ontogenic changes werecomparable to those previously seen in the relativeabundance of the same GH proteins in the chick testisduring ontogeny (Luna et al., 2004). Similar age-relatedchanges were also observed in the relative abundanceof the GH moieties detected under non-reducing condi-tions, in which the 17 kDa form was the most abundantbursal GH variant and the 40 kDa form was primarilyassociated with embryonic tissues.
The production and release of diVerent GH moietiesin the pituitary gland is known to be diVerentially regu-lated by diVerent GH secretagogues (Martínez-Coriaet al., 2002). It is therefore likely that they are also diVer-entially regulated in a tissue-speciWc manner duringontogeny. Moreover, as GH variants in the chicken pitu-itary gland diVer in biological activity and potency(Arámburo et al., 1989c, 1990, 2001b), tissue- and age-related diVerences in the relative abundance of the GHmoieties in immune tissues during development mightprovide a mechanism to subtly modulate the action ofGH in immune function.
The GH receptor is a member of the cytokine-recep-tor-super family and GH has numerous cytokine-likeroles in activating immune cells during immune chal-lenge and in inducing the proliferation and diVerentia-tion of stromal epithelial cells (see Clark, 1997). Indeed,the blockade of GH expression or GH action in immunecells dramatically impairs immune function (Weigentet al., 1991; Sabharwal and Varma, 1996), demonstratingimmune GH acts as a physiologically important auto-crine or paracrine immune regulator. Immune GH issimilarly likely to act as an immune regulator in birds, inwhich the GH receptor is present in the thymus, spleen

36 M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37
and bursa and in circulating leukocytes (Hull et al., 1996,2001). The increased morbidity and mortality of sex-linked dwarf chickens (Marsh et al., 1991; Marsh, 1983,1992); that are resistant to GH action because of a GHreceptor dysfunction (Burnside et al., 1991; Hull et al.,1993) supports this view.
In summary, these results demonstrate age-relatedand tissue-speciWc changes in the content and composi-tion of GH in immune tissues of the chicken, in whichGH is likely to be an autocrine or paracrine regulator.
Acknowledgments
We thank Leopoldo González-Santos and E. NydiaHernández-Ríos (digital analysis), Pilar Galarza-Barios(librarian); Martín García-Servín (vivarium), AlbertoLara, and Omar González-Hernández (computing) andGerardo Courtois for excellent technical assistance. Thiswork was supported by grants from DGAPA-UNAM(PAPIIT IN22602 and IN242704), CONACyT (31817-Nand 41218-Q), Pilgrim’s Pride of Mexico and NSERC ofCanada.
References
Arámburo, C., Carranza, M., Sánchez, R., Perera, G., 1989a. Partial bio-chemical and biological characterization of puriWed chicken growthhormone (cGH). Isolation of cGH charge variants and evidence thatcGH is phosphorylated. Gen. Comp. Endocrinol. 76, 330–339.
Arámburo, C., Sánchez, R., Fenton, B., Perera, G., Valverde-Rodrí-guez, C., 1989b. Desarrollo de un radioinmunoensayo homólogo yespecíWco para la determinación de hormona de crecimiento depollo (cGH). Vet. Mex. 20, 397–405.
Arámburo, C., Campbell, R.M., Scanes, C.G., 1989c. Heterogeneity ofchicken growth hormone (cGH). IdentiWcation of lipolytic andnon-lipolytic variants. Life Sci. 45, 2201–2207.
Arámburo, C., Montiel, J.L., Perera, G., Navarrete, S., Sánchez, R.,1990. Molecular isoforms of chicken growth hormone (cGH):diVerent bioactivities of cGH charge variants. Gen. Comp. Endo-crinol. 80, 59–67.
Arámburo, C., Navarrete, S., Montiel, J.L., Sánchez, R., Berghman,L.R., 1991. PuriWcation and electrophoretic analysis of glycosylatedchicken growth hormone (G-cGH): evidence of G-cGH isoforms.Gen. Comp. Endocrinol. 84, 135–146.
Arámburo, C., Luna, M., Carranza, M., Reyes, M., Martínez-Coria, H.,Scanes, C.G., 2000. Growth hormone size variants: changes in thepituitary during development of the chicken. Proc. Soc. Exp. Biol.Med. 223, 67–74.
Arámburo, C., Carranza, M., Martínez-Coria, H., Reyes, M., Berumen,L., López, J., Pascacio, H., Huerta, L., Luna, M., 2001a. Molecularand functional heterogeneity of growth hormone. In: Dawson, A.,Chaturvedi, C.M. (Eds.), Avian Endocrinology. Narosa PublishingHouse, New Delhi, pp. 273–286.
Arámburo, C., Carranza, M., Reyes, M., Luna, M., Martínez-Coria, H.,Berumen, L., Scanes, C.G., 2001b. Characterization of a bioactive15 kDa fragment produced by proteolytic cleavage of chickengrowth hormone. Endocrine 15, 231–240.
Baxter, J.B., Blalock, E., Weigent, D.A., 1991. Expression of immunoreactivegrowth hormone in leukocytes in vivo. J. Neuroimmunol. 33, 43–54.
Burnside, J., Liou, S.S., Cogburn, L.A., 1991. Molecular cloning of thechicken growth hormone receptor complementary deoxyribonu-cleic acid: mutation of the gene in sex-linked dwarf chickens. Endo-crinology 128, 3138–3192.
Chen, H.-T., Schuler, L.A., Schultz, R.D., 1997. Growth hormone andPit-1 expression in bovine fetal lymphoid cells. Domest. Anim.Endocrinol. 14, 399–407.
Clapp, C., Martial, J.A., Guzman, R.C., Rentier, Delure, R., Weiner,R.I., 1993. The 16-kilodalton N-terminal fragment of human pro-lactin is a potent inhibitor of angiogenesis. Endocrinology 133,1292–1299.
Clark, R., 1997. The somatogenic hormones and insulin-like growthfactor-1: stimulators of lymphopoiesis and immune function.Endocrine Rev. 18, 157–178.
Corbacho, A.M., Macotela, Y., Nava, G., Torner, L., Duenas, Z., Noris,G., Morales, M.A., Martinez de la Escalera, G., Clapp, C., 2000a.Human umbilical vein endothelial cells express multiple prolactinisoforms. J. Endocrinol. 166, 53–62.
Corbacho, A.M., Nava, G., Eiserich, J.P., Noris, G., Macotela, Y., Stru-man, I., Martinez de la Escalera, G., Clapp, C., 2000b. Proteolyticcleavage confers nitric oxide synthase inducing activity upon pro-lactin. J. Biol. Chem. 275, 13183–13186.
Corbacho, A.M., Martinez de la Escalera, G., Clapp, C., 2002. Roles ofprolactin and related members of the prolactin/growth hormone/placental lactogen family in angiogenesis. J. Endocrinol. 173, 219–238.
Costa, A., Zoppetti, G., Benedetto, C., Bertino, E., Marozio, L., Fabris,C., Arisio, R., Giraudi, G.F., Testori, O., Ariano, M., et al., 1993.Immunolike growth hormone substance in tissues from humanembryos/fetuses and adults. J. Endocrinol. Investig. 16, 625–633.
González, C., Corbacho, A.M., Eiserich, J.P., Garcia, C., Lopez-Bar-rera, F., Morales-Tlalpan, V., Barajas-Espinosa, A., Diaz-Muñoz,M., Rubio, R., Lin, S.-H., Martinez de la Escalera, G., Clapp, C.,2004. 16K-prolactin inhibits activation of endotelial nitric oxidesynthase, intracellular calcium mobilization, and endothelium-dependent vasorelaxation. Endocrinology 145, 5714–5722.
Franchini, A., Ottaviani, E., 1999. Immunoreactive POMC-derivedpeptides and cytokines in the chicken thymus and bursa of Fabri-cius microenvironments: age-related changes. J. Neuroendocrinol.11, 685–692.
Harvey, S., Scanes, C.G., 1977. PuriWcation and radioimmunoassay ofchicken growth hormone. J. Endocrinol . 73, 321–329.
Harvey, S., Scanes, C.G., Chadwick, A., Bolton, N.J., 1978. InXuence offasting, glucose and insulin on the levels of growth hormone andprolactin in the plasma of the domestic Xow (Gallus domesticus). J.Endocrinol. 76, 501–506.
Hattori, N., Kitagawa, K., Inagaki, C., 1999. Human lymphocytesexpress hGH-N gene transcripts of 22 kDa, 20 kDa andminor forms of GH, but not hGH-V gene. Eur. J. Endocrinol. 141,413–418.
Hull, K.L., Fraser, R.A., Marsh, J.A., Harvey, S., 1993. Growth hor-mone receptor gene expression in sex-linked dwarf Leghorn chick-ens: evidence against a gene deletion. J. Endocrinol. 137, 91–98.
Hull, K.L., Thiagarajah, A., Harvey, S., 1996. Cellular localization ofgrowth hormone receptors/binding proteins in immune tissues. CellTissue Res. 286, 69–80.
Hull, K.L., Nette, F., Harvey, S., 2001. Bidirectional communica-tion between the immune and neuroendocrine system: role ofgrowth hormone. In: Dawson, A., Chaturvedi, C.M. (Eds.),Avian Endocrinology. Narosa Publishing House, New Delhi,pp. 437–443.
Johnson, B.E., Scanes, C.G., King, D.B., Marsh, J.A., 1993. EVects ofhypophysectomy and growth hormone on immune development inthe domestic fowl. Dev. Comp. Immunol. 17, 331–339.
Kyle, C.V., Evans, M.C., Odell, W.D., 1981. Growth hormone-likematerial in normal humans tissues. J. Clin. Endocrinol. Metab. 53,1138–1144.

M. Luna et al. / General and Comparative Endocrinology 144 (2005) 28–37 37
Lantinga van Leeuwen, I.S., Teske, E., van Garderen, E., Mol, J.A.,2000. Growth hormone gene expression in normal lymph nodesand lymphomas of the dog. Anticancer Res. 20, 2371–2376.
Lazarus, D.D., Scanes, C.G., 1988. Acute eVects of hypophysectomyand administration of pancreatic and thyroid hormones on circu-lating concentrations of somatomedin-C in young chickens: rela-tionship between growth hormone and somatomedin-C. Domest.Anim. Endocrinol. 5, 283–289.
Luna, M., Huerta, L., Berumen, L., Martínez-Coria, H., Harvey, S.,Arámburo, C., 2004. Growth hormone in the male reproductivetract of the chicken: heterogeneity and changes during ontogenyand maturation. Gen. Comp. Endocrinol. 137, 37–49.
Liu, J., Zheng, D., Deng, J., Xiao, S., Liu, S., Shi, Y., 1997. Immunoreac-tive growth hormone in human peripheral T lymphocytes: encod-ing sequence of cDNA identical to that of the pituitary humangrowth hormone. Chin. Med. J. 110, 362–365.
Marsh, J.A., 1983. Assessment of antibody production in sex-linkeddwarf and autosomal dwarf chickens. Develop. Comp. Immunol. 7,535–542.
Marsh, J.A., Johnson, B.E., SaWeh, B., Kendall, M., Dardenne, M.,Scanes, C.G., 1991. Assessment of thymulin activity in Cornell SLDand K chickens. Poult. Sci. 70, 24 (Abst.).
Marsh, J.A., 1992. Neuroendocrine-immune interactions in the avianspecies—a review. Poult. Sci. Rev. 4, 129–167.
Martínez-Coria, H., López-Rosales, L.J., Carranza, M., Berumen, L.,Luna, M., Arámburo, C., 2002. DiVerential secretion of chickengrowth hormone variants after growth hormone-releasing hor-mone stimulation in vitro. Endocrine 17, 91–102.
Melen, L., Hennen, G., Dullaart, R.P., Heinen, E., Igout, A., 1997. Bothpituitary and placental growth hormone transcripts are expressedin human peripheral blood mononuclear cells (PBMC). Clin. Exp.Immunol. 110, 223–340.
Milicévic, Z., Vujic, D., Isakovic, K., Micic, M., Milicevic, N., 1986.Involution of bursa of Fabricius in male and female chickens: alight microscopic histoquantitative study. Poult. Sci. 65, 2318–2323.
Palmetshofer, A., Zechner, D., Luger, T.A., Barta, A., 1995. Splicingvariants of the human growth hormone mRNA: detection in pitui-tary, mononuclear cells and dermal Wbroblasts. Mol. Cell. Endocri-nol. 113, 225–234.
Paramithiotis, E., Jacobsen, K.A., RatcliVe, M.J.H., 1995. Loss of surg-ace immunoglobulin expression precedes B cell death by apoptosisin the bursa of Fabricius. J. Exp. Med. 181, 105–113.
Recher, S., Raccurt, M., Lambert, A., Lobie, P.E., Mertani, H.C.,Morel, G., 2001. Prenatal and adult growth hormone gene expres-sion in rat lymphoid organs. J. Histochem. Cytochem. 49, 347–354.
Render, C.L., Hull, K.L., Harvey, S., 1995a. Expression of the growthhormone gene in immune tissues. Endocrine 3, 729–735.
Render, C.L., Hull, K.L., Harvey, S., 1995b. Neural expression of thepituitary GH gene. J. Endocrinol. 147, 413–422.
Rohn, W.M., Weigent, D.A., 1995. Cloning and nucleotide sequencingof rat lymphocyte growth hormone cDNA. Neuroimmunomodula-tion 2, 108–114.
Sabharwal, P., Varma, S., 1996. Growth hormone synthesized andsecreted by human thymocytes acts via insulin-like growth factor Ias an autocrine and paracrine growth factor. J. Clin. Endocrinol.Metab. 81, 2663–2669.
Sanders, E.J., Parker, E., Arámburo, C., Harvey, S. 2005. Retinalgrowth hormone is an anti-apoptotic factor in embryonic retinalganglion cell diVerentiation. Exp. Eye Res. (in press).
Weigent, D.A., Baxter, J.B., Wear, W.E., Sith, L.R., Bost, K.L., Blalock,J.E., 1988. Production of immunoreactive growth hormone bymononuclear leukocytes. FASEB J. 2, 2812–2818.
Weigent, D.A., Blalock, J.E., 1989. Expression of growth hormone bylymphocytes. Int. Rev. Immunol. 4, 193–211.
Weigent, D.A., Blalock, J.E., LeBoeuf, R., 1991. An antisense oligo-deoxynucleotide to growth hormone messenger ribonucleic acidinhibits lymphocyte proliferation. Endocrinology 128, 2053–2057.
Weigent, D.A., Blalock, J.E., 1995. Growth hormone eVects on theimmune system. In: Shiverick, K.T., Rosenbloom, A.L. (Eds.),Growth Hormone: Basic and Clinical Aspects. CRC Press, BocaRaton, FL, pp. 141–152.
Wu, H., Devi, R., Malarkey, W.B., 1996. Localization of growth hor-mone messenger ribonucleic acid in the human immune system—aclinical research study. J. Clin. Endocrinol. Metab. 81, 1278–1282.
Yang, Y., Guo, L., Ma, L., Liu, X., 1999. Expression of growth hor-mone and insulin-like growth factor in the immune system of chil-dren. Horm. Metab. Res. 31, 380–384.




















