
Observations concerning the mechanics of ovulation in the fowl

1 23
Parasitology ResearchFounded as Zeitschrift fürParasitenkunde ISSN 0932-0113Volume 109Number 4 Parasitol Res (2011) 109:1113-1123DOI 10.1007/s00436-011-2354-z
Efficacy of abamectin against the fowltick, Argas (Persicargas) persicus (Oken,1818) (Ixodoidea: Argasidae)
Ashraf A. Montasser, Aleya S. Marzouk,Sherif H. El-Alfy & Asmaa A. Baioumy

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ORIGINAL PAPER
Efficacy of abamectin against the fowl tick, Argas(Persicargas) persicus (Oken, 1818) (Ixodoidea: Argasidae)
Ashraf A. Montasser & Aleya S. Marzouk &
Sherif H. El-Alfy & Asmaa A. Baioumy
Received: 29 December 2010 /Accepted: 15 March 2011 /Published online: 6 May 2011# Springer-Verlag 2011
Abstract Abamectin, in aqueous solutions of dose rates0.0625, 0.125, 0.25, 0.5, and 1 mL/L, was sprayed ondifferent feeding stages of the tick Argas (P.) persicus. Theresults revealed a marked increase of immobile and deadmale or female ticks following a single treatment with theabove doses, particularly the higher ones, and during the5 weeks after treatment. Abamectin seriously decreased thepercentage of fed ticks as well as that of oviposition andhatching. A decrease in the amount of emitted coxal fluidwas observed also following treatment. Although theamount of ingested blood increased following abamectintreatment, digestion remained similar. The study alsorevealed that spraying a dose of 0.5 mL/L of abamectin infowl shelters, i.e., floor, walls, ceiling, etc., is sufficient toeradicate A. persicus population.
Introduction
Ticks are important ectoparasites of domestic animals(Roberts and Janovy 2005). The fowl tick, Argas persicus,is of considerable veterinary importance as a parasite ofpoultry and wild birds causing weakness and anemia (Walland Shearer 1997), reduction in egg production and growth,in addition to their death (Quilez et al. 2005). This tick isthe vector of fowl spirochaetosis (Wall and Shearer 1997;
Shah et al. 2006). Recently, the bacteria Staphylococcusaureus, Salmonella pullorum, and Escherichia coli wereisolated from this species (Shah et al. 2006).
The avermectins are a family of macrocyclic lactonesproduced by the soil organism Streptomyces avermitilis,which were discovered in the mid-1970s as a direct resultof a screening effort for natural products with anthelminticproperties (Lasota and Dybas 1990). Abamectin is amixture of avermectins containing >80% avermectin B1aand <20% avermectin B1b (Meister 1992). Abamectinexhibited immense efficiency in the control of ectoparasitesinfesting cattle in Germany (Heinze-Mutz et al. 1993), inaddition to extraordinary activity against insect and mitepests of a worldwide range of agronomic, vegetable, andornamental crops (Yee et al. 2001; Kumar and Poehling2007; Kavallieratos et al. 2009; Civolani et al. 2010).
The effect of avermectins against ticks using differentroutes of application was considerably studied (Davey etal. 2010; Lohmeyer et al. 2009; Sajid et al. 2009; Marzouket al. 2004; Montasser 2004). However, studies concernedwith the effect of abamectin against ticks are scarce. Theonly reports available are those using pour-on preparationagainst Rhipicephalus appendiculatus and Rhipicephalusdecoloratus (Van Der Merwe et al. 2005) and subcutane-ous injection against Boophilus microplus (Pereira 2009).The aim of the present work was to study the mainbiological parameters affecting reproduction and develop-ment after the direct application of abamectin in a sprayformulation on adult A. persicus itself away from itspoultry hosts since an important aspect of acaricideresearch has always been to be safe toward non-targetorganisms.
A. A. Montasser (*) :A. S. Marzouk : S. H. El-Alfy :A. A. BaioumyZoology Department, Faculty of Science, Ain Shams University,Cairo, Egypte-mail: [email protected]
Parasitol Res (2011) 109:1113–1123DOI 10.1007/s00436-011-2354-z
Author's personal copy

Materials and methods
Tick origin
A. persicus, used in the present work, originated from afowl house in Abo-Rawash area, 6th October Governorate,Egypt, and was maintained in an incubator adjusted to 28±1°C and 75±5% relative humidity in the Zoology Depart-ment laboratories, Faculty of Science, Ain Shams Univer-sity. Ticks were identified and grouped into fed, partiallyfed, and unfed males, females, and nymphs. The domesticpigeon, Columba livia domestica, was used as a host.Unfed nymphs were reared to produce virgin males andfemales, whereas some of the unfed ticks were placed tofeed for 1–2 h on the pigeon auxiliary region and otherswere transferred to glass vials for experiments on the unfedgroup.
Abamectin treatment
Ticks were treated using the spray method of Cetin et al.(2009). Briefly, a single treatment of abamectin (Vertemic1.8% EC, Syngenta Co., Switzerland, expiry date July2009, a formulation containing 80% Avermectin B1a and20% Avermectin B1b) was applied through a hand triggersprayer (0.5-L capacity). Ten milliliters was sprayed onticks in a Petri dish containing filter paper, about 20–25 cmfrom the surface. Ticks were then transferred to clean glassvials with small pieces of filter paper and then placed in theincubator.
The used doses of abamectin were 0.0625, 0.125, 0.25,0.5, and 1 mL/L distilled water. In order to control allticks, doses were applied to unfed, partially fed, and fullyfed adults. Unfed ticks were allowed to feed after 1 day oftreatment (the least applicable period from treatment tofeeding). Fully fed ticks were treated after 2 h fromfeeding (the least applicable period from feeding totreatment).
Biological studies
Mobility and mortality
They were determined by the percent of mobile, immobile,and dead ticks during the 5 weeks following treatment.Specimens were pressed several times to check theirmovement.
Sexual behavior
Directly after feeding, each pair of male and female tickswas isolated in a Petri dish, 3 cm in diameter, with at least1 cm between them and allowed to mate in the incubator in
dark conditions within 1 h. Every 5 min, specimens werechecked for their sexual response in which specimens thatassume a copulatory position (Khalil et al. 1981) weredescribed as positively active and vice versa.
Amounts of ingested blood and emitted coxal fluid
The amount of ingested blood by males or females wasdetermined by subtracting the weight before feeding from theweight directly after feeding, while the amount of emitted coxalfluid was determined by subtracting the weight after 2 h offeeding from the weight directly after feeding (Marzouk et al.2004). The effect of abamectin on sexual behavior, amount ofingested blood, and emitted coxal fluid were only investigatedin the unfed group because in the partially or fully fed groups,these parameters took place before treatment.
Blood digestion
Blood digestion was determined by comparing the weightsof ticks between successive weeks from zero (directly afterfeeding), first, second, third, fourth, and fifth weeks afterfeeding (Marzouk et al. 2004). The weights of laid eggswere added to the weights of their females.
Oviposition, hatchability, and fertility
Oviposition was represented by oviposition percent (numberof oviposited females/total number of females×100), ovipo-sition period (days from feeding to egg laying), and numberof eggs per ovipositing female. Hatchability was representedby the hatching period (days from onset of eggs to theirhatch) and hatching percent (number of hatched eggs/totalnumber of eggs×100). Fertility is the weight of egg massdivided by the weight of replete female (Khalil et al. 1984)and the weight of one egg by dividing the weight of eggmass by the total number of eggs.
Statistical method
Experiments were repeated three times, each including 15–20 pairs of males and females. Means and standard errorswere calculated and the relationships were tested by theStudent's t test (Fisher 1948).
Results
Mobility and mortality
Abamectin obviously causes paralysis or death in A.persicus adults particularly with the increase of the sprayeddose and/or the time after treatment. Percent of mobile,
1114 Parasitol Res (2011) 109:1113–1123
Author's personal copy
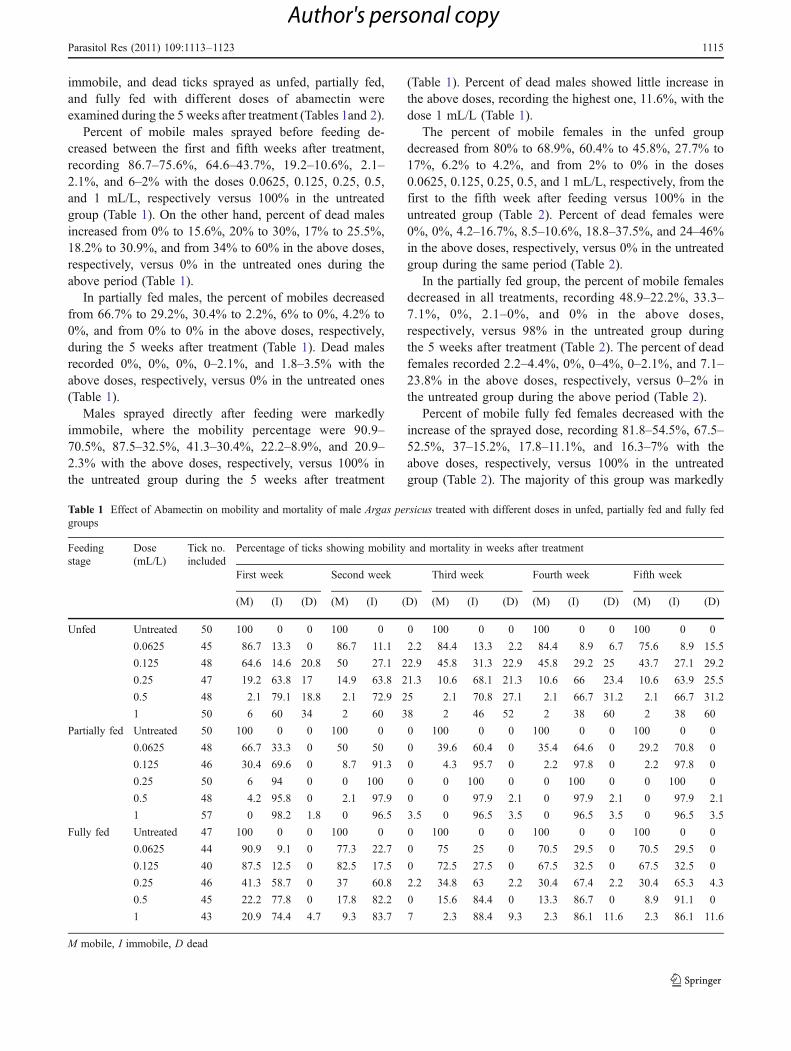
immobile, and dead ticks sprayed as unfed, partially fed,and fully fed with different doses of abamectin wereexamined during the 5 weeks after treatment (Tables 1and 2).
Percent of mobile males sprayed before feeding de-creased between the first and fifth weeks after treatment,recording 86.7–75.6%, 64.6–43.7%, 19.2–10.6%, 2.1–2.1%, and 6–2% with the doses 0.0625, 0.125, 0.25, 0.5,and 1 mL/L, respectively versus 100% in the untreatedgroup (Table 1). On the other hand, percent of dead malesincreased from 0% to 15.6%, 20% to 30%, 17% to 25.5%,18.2% to 30.9%, and from 34% to 60% in the above doses,respectively, versus 0% in the untreated ones during theabove period (Table 1).
In partially fed males, the percent of mobiles decreasedfrom 66.7% to 29.2%, 30.4% to 2.2%, 6% to 0%, 4.2% to0%, and from 0% to 0% in the above doses, respectively,during the 5 weeks after treatment (Table 1). Dead malesrecorded 0%, 0%, 0%, 0–2.1%, and 1.8–3.5% with theabove doses, respectively, versus 0% in the untreated ones(Table 1).
Males sprayed directly after feeding were markedlyimmobile, where the mobility percentage were 90.9–70.5%, 87.5–32.5%, 41.3–30.4%, 22.2–8.9%, and 20.9–2.3% with the above doses, respectively, versus 100% inthe untreated group during the 5 weeks after treatment
(Table 1). Percent of dead males showed little increase inthe above doses, recording the highest one, 11.6%, with thedose 1 mL/L (Table 1).
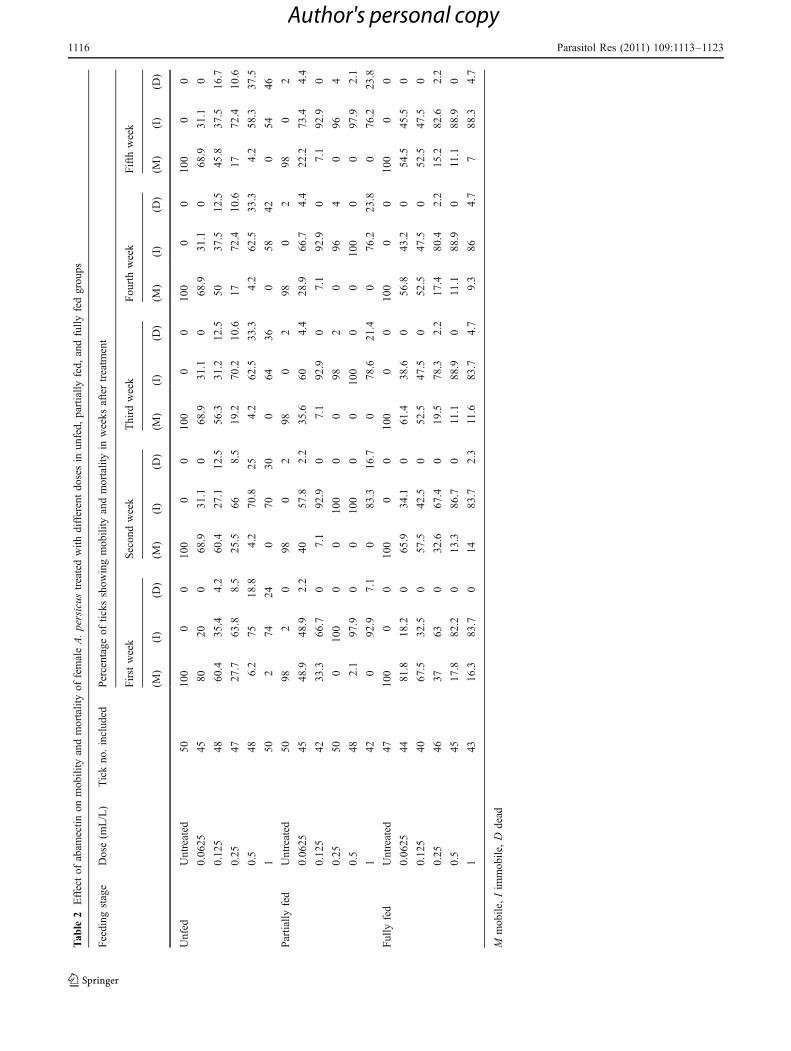
The percent of mobile females in the unfed groupdecreased from 80% to 68.9%, 60.4% to 45.8%, 27.7% to17%, 6.2% to 4.2%, and from 2% to 0% in the doses0.0625, 0.125, 0.25, 0.5, and 1 mL/L, respectively, from thefirst to the fifth week after feeding versus 100% in theuntreated group (Table 2). Percent of dead females were0%, 0%, 4.2–16.7%, 8.5–10.6%, 18.8–37.5%, and 24–46%in the above doses, respectively, versus 0% in the untreatedgroup during the same period (Table 2).
In the partially fed group, the percent of mobile femalesdecreased in all treatments, recording 48.9–22.2%, 33.3–7.1%, 0%, 2.1–0%, and 0% in the above doses,respectively, versus 98% in the untreated group duringthe 5 weeks after treatment (Table 2). The percent of deadfemales recorded 2.2–4.4%, 0%, 0–4%, 0–2.1%, and 7.1–23.8% in the above doses, respectively, versus 0–2% inthe untreated group during the above period (Table 2).
Percent of mobile fully fed females decreased with theincrease of the sprayed dose, recording 81.8–54.5%, 67.5–52.5%, 37–15.2%, 17.8–11.1%, and 16.3–7% with theabove doses, respectively, versus 100% in the untreatedgroup (Table 2). The majority of this group was markedly
Table 1 Effect of Abamectin on mobility and mortality of male Argas persicus treated with different doses in unfed, partially fed and fully fedgroups
Feedingstage
Dose(mL/L)
Tick no.included
Percentage of ticks showing mobility and mortality in weeks after treatment
First week Second week Third week Fourth week Fifth week
(M) (I) (D) (M) (I) (D) (M) (I) (D) (M) (I) (D) (M) (I) (D)
Unfed Untreated 50 100 0 0 100 0 0 100 0 0 100 0 0 100 0 0
0.0625 45 86.7 13.3 0 86.7 11.1 2.2 84.4 13.3 2.2 84.4 8.9 6.7 75.6 8.9 15.5
0.125 48 64.6 14.6 20.8 50 27.1 22.9 45.8 31.3 22.9 45.8 29.2 25 43.7 27.1 29.2
0.25 47 19.2 63.8 17 14.9 63.8 21.3 10.6 68.1 21.3 10.6 66 23.4 10.6 63.9 25.5
0.5 48 2.1 79.1 18.8 2.1 72.9 25 2.1 70.8 27.1 2.1 66.7 31.2 2.1 66.7 31.2
1 50 6 60 34 2 60 38 2 46 52 2 38 60 2 38 60
Partially fed Untreated 50 100 0 0 100 0 0 100 0 0 100 0 0 100 0 0
0.0625 48 66.7 33.3 0 50 50 0 39.6 60.4 0 35.4 64.6 0 29.2 70.8 0
0.125 46 30.4 69.6 0 8.7 91.3 0 4.3 95.7 0 2.2 97.8 0 2.2 97.8 0
0.25 50 6 94 0 0 100 0 0 100 0 0 100 0 0 100 0
0.5 48 4.2 95.8 0 2.1 97.9 0 0 97.9 2.1 0 97.9 2.1 0 97.9 2.1
1 57 0 98.2 1.8 0 96.5 3.5 0 96.5 3.5 0 96.5 3.5 0 96.5 3.5
Fully fed Untreated 47 100 0 0 100 0 0 100 0 0 100 0 0 100 0 0
0.0625 44 90.9 9.1 0 77.3 22.7 0 75 25 0 70.5 29.5 0 70.5 29.5 0
0.125 40 87.5 12.5 0 82.5 17.5 0 72.5 27.5 0 67.5 32.5 0 67.5 32.5 0
0.25 46 41.3 58.7 0 37 60.8 2.2 34.8 63 2.2 30.4 67.4 2.2 30.4 65.3 4.3
0.5 45 22.2 77.8 0 17.8 82.2 0 15.6 84.4 0 13.3 86.7 0 8.9 91.1 0
1 43 20.9 74.4 4.7 9.3 83.7 7 2.3 88.4 9.3 2.3 86.1 11.6 2.3 86.1 11.6
M mobile, I immobile, D dead
Parasitol Res (2011) 109:1113–1123 1115
Author's personal copy
Tab
le2
Effectof
abam
ectin
onmob
ility
andmortalityof
femaleA.persicus
treatedwith
differentdo
sesin
unfed,
partially
fed,
andfully
fedgrou
ps
Feeding
stage
Dose(m
L/L)
Tickno
.includ
edPercentageof
ticks
show
ingmob
ility
andmortalityin
weeks
aftertreatm
ent
Firstweek
Secon
dweek
Third
week
Fou
rthweek
Fifth
week
(M)
(I)
(D)
(M)
(I)
(D)
(M)
(I)
(D)
(M)
(I)
(D)
(M)
(I)
(D)
Unfed
Untreated
5010
00
010
00
010
00
010
00
010
00
0
0.06
2545
8020
068
.931
.10
68.9
31.1
068
.931
.10
68.9
31.1
0
0.12
548
60.4
35.4
4.2
60.4
27.1
12.5
56.3
31.2
12.5
5037
.512
.545
.837
.516
.7
0.25
4727
.763
.88.5
25.5
668.5
19.2
70.2
10.6
1772
.410
.617
72.4
10.6
0.5
486.2
7518
.84.2
70.8
254.2
62.5
33.3
4.2
62.5
33.3
4.2
58.3
37.5
150
274
240
7030
064
360
5842
054
46
Partially
fed
Untreated
5098
20
980
298
02
980
298
02
0.06
2545
48.9
48.9
2.2
4057
.82.2
35.6
604.4
28.9
66.7
4.4
22.2
73.4
4.4
0.12
542
33.3
66.7
07.1
92.9
07.1
92.9
07.1
92.9
07.1
92.9
0
0.25
500
100
00
100
00
982
096
40
964
0.5
482.1
97.9
00
100
00
100
00
100
00
97.9
2.1
142
092
.97.1
083
.316
.70
78.6
21.4
076
.223
.80
76.2
23.8
Fully
fed
Untreated
4710
00
010
00
010
00
010
00
010
00
0
0.06
2544
81.8
18.2
065
.934
.10
61.4
38.6
056
.843
.20
54.5
45.5
0
0.12
540
67.5
32.5
057
.542
.50
52.5
47.5
052
.547
.50
52.5
47.5
0
0.25
4637
630
32.6
67.4
019
.578
.32.2
17.4
80.4
2.2
15.2
82.6
2.2
0.5
4517
.882
.20
13.3
86.7
011.1
88.9
011.1
88.9
011.1
88.9
0
143
16.3
83.7
014
83.7
2.3
11.6
83.7
4.7
9.3
864.7
788
.34.7
Mmob
ile,Iim
mob
ile,D
dead
1116 Parasitol Res (2011) 109:1113–1123
Author's personal copy
paralyzed where the percent of dead females was rare in alldoses, recording its highest value (4.7%) with the dose1 mL/L (Table 2).
Generally, the average percentage of immobile anddead ticks collectively in the above tested male andfemale groups during the 5 weeks following abamectintreatment at the dose of 0.5 mL/L was >93% (Tables 1and 2).
Sexual behavior
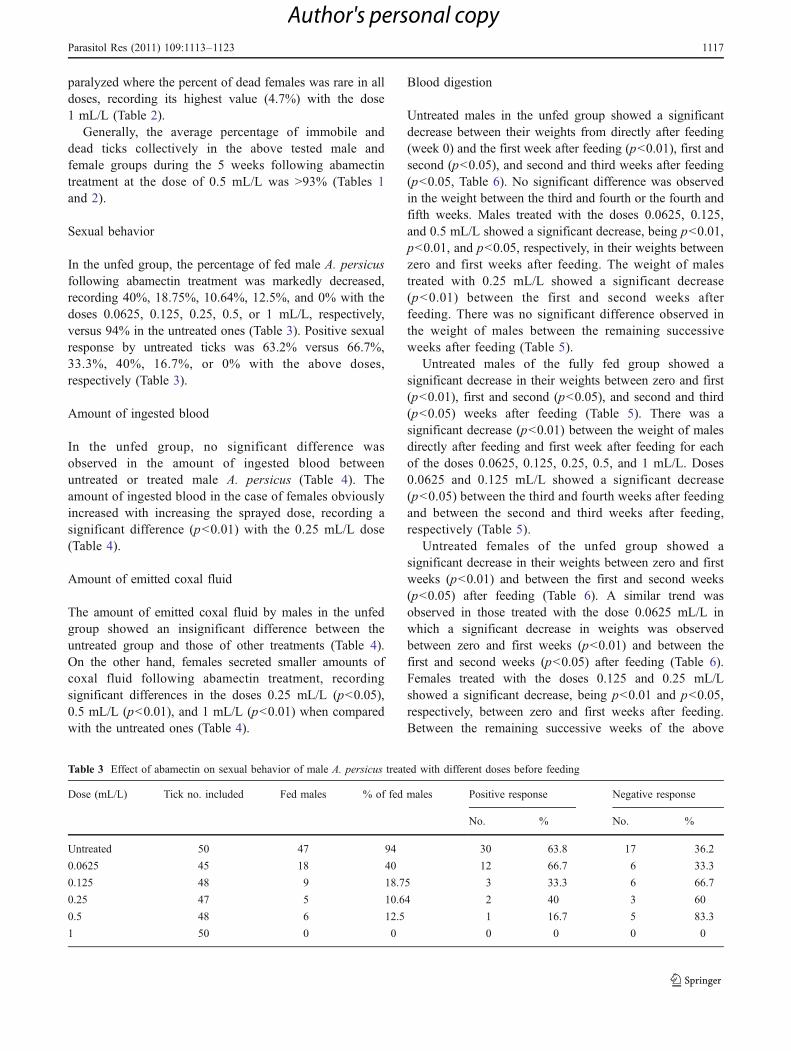
In the unfed group, the percentage of fed male A. persicusfollowing abamectin treatment was markedly decreased,recording 40%, 18.75%, 10.64%, 12.5%, and 0% with thedoses 0.0625, 0.125, 0.25, 0.5, or 1 mL/L, respectively,versus 94% in the untreated ones (Table 3). Positive sexualresponse by untreated ticks was 63.2% versus 66.7%,33.3%, 40%, 16.7%, or 0% with the above doses,respectively (Table 3).
Amount of ingested blood
In the unfed group, no significant difference wasobserved in the amount of ingested blood betweenuntreated or treated male A. persicus (Table 4). Theamount of ingested blood in the case of females obviouslyincreased with increasing the sprayed dose, recording asignificant difference (p<0.01) with the 0.25 mL/L dose(Table 4).
Amount of emitted coxal fluid
The amount of emitted coxal fluid by males in the unfedgroup showed an insignificant difference between theuntreated group and those of other treatments (Table 4).On the other hand, females secreted smaller amounts ofcoxal fluid following abamectin treatment, recordingsignificant differences in the doses 0.25 mL/L (p<0.05),0.5 mL/L (p<0.01), and 1 mL/L (p<0.01) when comparedwith the untreated ones (Table 4).
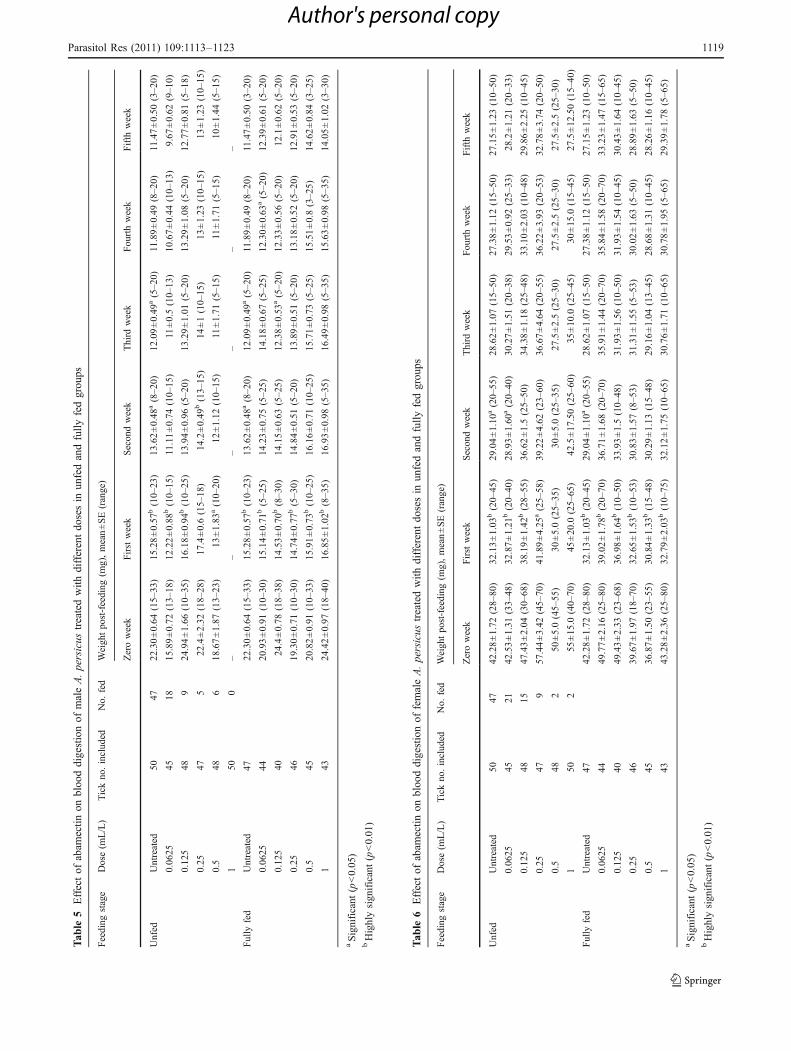
Blood digestion
Untreated males in the unfed group showed a significantdecrease between their weights from directly after feeding(week 0) and the first week after feeding (p<0.01), first andsecond (p<0.05), and second and third weeks after feeding(p<0.05, Table 6). No significant difference was observedin the weight between the third and fourth or the fourth andfifth weeks. Males treated with the doses 0.0625, 0.125,and 0.5 mL/L showed a significant decrease, being p<0.01,p<0.01, and p<0.05, respectively, in their weights betweenzero and first weeks after feeding. The weight of malestreated with 0.25 mL/L showed a significant decrease(p<0.01) between the first and second weeks afterfeeding. There was no significant difference observed inthe weight of males between the remaining successiveweeks after feeding (Table 5).
Untreated males of the fully fed group showed asignificant decrease in their weights between zero and first(p<0.01), first and second (p<0.05), and second and third(p<0.05) weeks after feeding (Table 5). There was asignificant decrease (p<0.01) between the weight of malesdirectly after feeding and first week after feeding for eachof the doses 0.0625, 0.125, 0.25, 0.5, and 1 mL/L. Doses0.0625 and 0.125 mL/L showed a significant decrease(p<0.05) between the third and fourth weeks after feedingand between the second and third weeks after feeding,respectively (Table 5).
Untreated females of the unfed group showed asignificant decrease in their weights between zero and firstweeks (p<0.01) and between the first and second weeks(p<0.05) after feeding (Table 6). A similar trend wasobserved in those treated with the dose 0.0625 mL/L inwhich a significant decrease in weights was observedbetween zero and first weeks (p<0.01) and between thefirst and second weeks (p<0.05) after feeding (Table 6).Females treated with the doses 0.125 and 0.25 mL/Lshowed a significant decrease, being p<0.01 and p<0.05,respectively, between zero and first weeks after feeding.Between the remaining successive weeks of the above
Table 3 Effect of abamectin on sexual behavior of male A. persicus treated with different doses before feeding
Dose (mL/L) Tick no. included Fed males % of fed males Positive response Negative response
No. % No. %
Untreated 50 47 94 30 63.8 17 36.2
0.0625 45 18 40 12 66.7 6 33.3
0.125 48 9 18.75 3 33.3 6 66.7
0.25 47 5 10.64 2 40 3 60
0.5 48 6 12.5 1 16.7 5 83.3
1 50 0 0 0 0 0 0
Parasitol Res (2011) 109:1113–1123 1117
Author's personal copy

doses, the female weights showed no significant difference.Females treated with the doses 0.5 and 1 mL/L showed aslight blood digestion, as observed from the insignificantdegradation in their weights between successive weeks afterfeeding (Table 6).
Blood digestion was more or less similar in the untreatedand abamectin-treated females in the fully fed group whereboth showed a significant decrease (p<0.01) in theirweights between zero and first weeks after feeding(Table 6). The weight of females in the untreated grouprevealed a significant decrease (p<0.05) between the firstand second weeks. Both untreated and treated females didnot show any significant differences in their weightsbetween the remaining successive weeks after feeding(Table 6).
Oviposition, hatchability, and fertility
Percent of oviposited females in the unfed group decreasedwith increasing the sprayed dose, recording 34% inuntreated ticks versus 19% and 13.3% in those treated withthe doses 0.0625 and 0.125 mL/L, respectively, and zero inthe remaining doses. Abamectin causes a delay in theoviposition period and a significant decrease in fertility andweight of one egg in the case of the 0.125 mL/L dose. Nosignificant difference was observed between the untreatedand the abamectin-sprayed ticks in the case of number ofeggs, weight of egg mass, hatching period, and percentage(Table 7).
Percent of oviposited females in the fully fed groupdecreased with increasing the sprayed dose (Table 7). Itrecorded 9.1% and 7.5% in those treated with the doses0.0625 and 0.125 mL/L, respectively, and 0% in the
remaining doses versus 34% in untreated ticks. However,the oviposition period was markedly decreased with the0.125-mL/L dose, recording 9.7 versus 15.2 days in theuntreated group. Abamectin caused a minor decrease inhatching percent and delay in hatching period. Nosignificant difference was observed in the number of eggs,fertility, and weight of one egg between females of theuntreated and those treated with the doses 0.0625 or0.125 mL/L (Table 7).
Discussion
Adult A. persicus spends most of its life away from thehost, where it stays about 1 h only on the host duringfeeding. Therefore, the present study is directed to applyabamectin against ticks at different feeding stages in theirhabitats and away from their host, i.e., in the vicinity of afowl shelter. The spray method of drug administration usedin the present study is characterized by lesser time-consuming, simple application where there is no need forextra training and the equipment are easily portable.Spraying method of abamectin was widely applied againstmites (Humeres and Morse 2005; Venzon et al. 2008; Kohet al. 2009; Lin et al. 2009).
The present study showed that the maximum percent ofmobile male and female A. persicus was observed duringthe first week after feeding following abamectin treatmentwith a single dose of 0.0625, 0.125, 0.25, 0.5, or 1 mL/Ldistilled water and gradually decreased until the fifth weekwith increasing the sprayed dose, while the percent ofimmobile and dead ones were increased with increasing theabove doses from the first to the fifth weeks after feeding.
Table 4 Effect of abamectin on the ability of feeding and amounts of ingested blood and emitted coxal fluid by adult A. persicus treated withdifferent doses before feeding
Dose (mL/L) Sex Tick no. included No. fed % fed Ingested blood (mg), mean±SE (range) Emitted coxal fluid (mg), mean±SE (range)
Untreated Male 50 47 94 14.80±0.63 (9–25) 3.81±0.50 (0–13)
Female 50 47 94 32.32±1.67 (17–69) 6.57±0.75 (0–15)
0.0625 Male 45 18 40 12.5±1.15 (8–12)a 3±0.65 (0–5)
Female 45 21 46.7 32.33±1.37 (22–37) 7.13±0.80 (5–12)
0.125 Male 48 9 18.8 17.22±1.63 (2–27) 4.72±0.94 (0–15)
Female 48 15 31.3 36.43±1.92 (21–56) 5.81±0.94 (0–13)
0.25 Male 47 5 10.6 16.4±2.32 (12–22) 3.2±1.96 (0–8)
Female 47 9 19.1 43.44±3.42 (31–56)b 5±0.5 (2–20)a
0.5 Male 48 6 12.5 12.67±1.87 (7–17) 4.33±0.42 (3–5)
Female 48 2 4.2 39.5±4.50 (35–44) 4.5±0.4 (10–15)b
1 Male 50 0 0 – –
Female 50 2 4 42.5±10.50 (32–53) 4.2±0 (4.2–4.2)b
a Significant (p<0.05)b Highly significant (p<0.01)
1118 Parasitol Res (2011) 109:1113–1123
Author's personal copy
Tab
le5
Effectof
abam
ectin
onbloo
ddigestionof
maleA.persicus
treatedwith
differentdo
sesin
unfedandfully
fedgrou
ps
Feeding
stage
Dose(m
L/L)
Tickno.included
No.
fed
Weightpost-feeding
(mg),mean±SE(range)
Zeroweek
Firstweek
Secondweek
Third
week
Fourthweek
Fifth
week
Unfed
Untreated
5047
22.30±0.64
(15–
33)
15.28±0.57
b(10–
23)
13.62±0.48
a(8–2
0)12.09±0.49
a(5–2
0)11.89±0.49
(8–2
0)11.47±0.50
(3–2
0)
0.0625
4518
15.89±0.72
(13–
18)
12.22±0.88
b(10–
15)
11.11±0.74
(10–15)
11±0.5(10–13)
10.67±0.44
(10–
13)
9.67
±0.62
(9–1
0)
0.125
489
24.94±1.66
(10–
35)
16.18±0.94
b(10–
25)
13.94±0.96
(5–2
0)13.29±1.01
(5–2
0)13.29±1.08
(5–2
0)12.77±0.81
(5–1
8)
0.25
475
22.4±2.32
(18–
28)
17.4±0.6(15–18)
14.2±0.49
b(13–15)
14±1(10–15)
13±1.23
(10–
15)
13±1.23
(10–15)
0.5
486
18.67±1.87
(13–
23)
13±1.83
a(10–20)
12±1.12
(10–15)
11±1.71
(5–1
5)11
±1.71
(5–1
5)10
±1.44
(5–1
5)
150
0–
––
––
–
Fully
fed
Untreated
4722.30±0.64
(15–
33)
15.28±0.57
b(10–
23)
13.62±0.48
a(8–2
0)12.09±0.49
a(5–2
0)11.89±0.49
(8–2
0)11.47±0.50
(3–2
0)
0.0625
4420.93±0.91
(10–
30)
15.14±0.71
b(5–2
5)14.23±0.75
(5–2
5)14.18±0.67
(5–2
5)12.30±0.63
a(5–2
0)12.39±0.61
(5–2
0)
0.125
4024.4±0.78
(18–
38)
14.53±0.70
b(8–3
0)14.15±0.63
(5–2
5)12.38±0.53
a(5–2
0)12.33±0.56
(5–2
0)12.1±0.62
(5–2
0)
0.25
4619.30±0.71
(10–
30)
14.74±0.77
b(5–3
0)14.84±0.51
(5–2
0)13.89±0.51
(5–2
0)13.18±0.52
(5–2
0)12.91±0.53
(5–2
0)
0.5
4520.82±0.91
(10–
33)
15.91±0.73
b(10–
25)
16.16±0.71
(10–25)
15.71±0.73
(5–2
5)15.51±0.8(3–2
5)14.62±0.84
(3–2
5)
143
24.42±0.97
(18–
40)
16.85±1.02
b(8–3
5)16.93±0.98
(5–3
5)16.49±0.98
(5–3
5)15.63±0.98
(5–3
5)14.05±1.02
(3–3
0)
aSignificant
(p<0.05)
bHighlysign
ificant(p<0.01)
Tab
le6
Effectof
abam
ectin
onbloo
ddigestionof
femaleA.persicus
treatedwith
differentdo
sesin
unfedandfully
fedgrou
ps
Feeding
stage
Dose(m
L/L)
Tickno.included
No.
fed
Weightpost-feeding
(mg),mean±SE(range)
Zeroweek
Firstweek
Secondweek
Third
week
Fourthweek
Fifth
week
Unfed
Untreated
5047
42.28±1.72
(28–
80)
32.13±1.03
b(20–
45)
29.04±1.10
a(20–55)
28.62±1.07
(15–
50)
27.38±1.12
(15–
50)
27.15±1.23
(10–50)
0.0625
4521
42.53±1.31
(33–
48)
32.87±1.21
b(20–
40)
28.93±1.60
a(20–40)
30.27±1.51
(20–
38)
29.53±0.92
(25–
33)
28.2±1.21
(20–33)
0.125
4815
47.43±2.04
(30–
68)
38.19±1.42
b(28–
55)
36.62±1.5(25–50)
34.38±1.18
(25–
48)
33.10±2.03
(10–
48)
29.86±2.25
(10–45)
0.25
479
57.44±3.42
(45–
70)
41.89±4.25
a(25–
58)
39.22±4.62
(23–60)
36.67±4.64
(20–
55)
36.22±3.93
(20–
53)
32.78±3.74
(20–50)
0.5
482
50±5.0(45–
55)
30±5.0(25–
35)
30±5.0(25–35)
27.5±2.5(25–
30)
27.5±2.5(25–
30)
27.5±2.5(25–30)
150
255
±15.0
(40–
70)
45±20.0
(25–65)
42.5±17.50(25–60)
35±10.0
(25–
45)
30±15.0
(15–
45)
27.5±12.50(15–40)
Fully
fed
Untreated
4742.28±1.72
(28–
80)
32.13±1.03
b(20–
45)
29.04±1.10
a(20–55)
28.62±1.07
(15–
50)
27.38±1.12
(15–
50)
27.15±1.23
(10–50)
0.0625
4449.77±2.16
(25–
80)
39.02±1.78
b(20–
70)
36.71±1.68
(20–70)
35.91±1.44
(20–
70)
35.84±1.58
(20–
70)
33.23±1.47
(15–65)
0.125
4049.43±2.33
(23–
68)
36.98±1.64
b(10–
50)
33.93±1.5(10–48)
31.93±1.56
(10–
50)
31.93±1.54
(10–
45)
30.43±1.64
(10–45)
0.25
4639.67±1.97
(18–
70)
32.65±1.53
b(10–
53)
30.83±1.57
(8–5
3)31.31±1.55
(5–5
3)30.02±1.63
(5–5
0)28.89±1.63
(5–5
0)
0.5
4536.87±1.50
(23–
55)
30.84±1.33
b(15–
48)
30.29±1.13
(15–48)
29.16±1.04
(13–
45)
28.68±1.31
(10–
45)
28.26±1.16
(10–45)
143
43.28±2.36
(25–
80)
32.79±2.03
b(10–
75)
32.12±1.75
(10–65)
30.76±1.71
(10–
65)
30.78±1.95
(5–6
5)29.39±1.78
(5–6
5)
aSignificant
(p<0.05
)bHighlysign
ificant(p<0.01
)
Parasitol Res (2011) 109:1113–1123 1119
Author's personal copy

Tab
le7
Effectof
abam
ectin
onov
iposition
,hatchability,
andfertility
ofA.persicus
treatedwith
differentdo
sesin
unfedandfully
fedgrou
ps
Feeding
stage
Dose
(mL/)L
Tickno
.includ
edNo.
fed
No.
ofov
iposited
females
Percent
ofov
iposited
females
Ovipo
sitio
nperiod
(days),
mean±SE
(range)
No.
ofeggs,
mean±SE
(range)
Hatching
percent,
%±SE(range)
Hatching
period
(days),
mean±SE
(range)
Meanweigh
tof
replete
female(m
g),
mean±SE
(range)
Mean
eggmass(m
g),
mean±SE
(range)
Fertility
eggmass/female
weigh
t,mean±SE
(range)
Weigh
tof
oneegg(m
g),
mean±SE
(range)
Unfed
Untreated
5047
1634
15.19±1.59
(6–28)
62.38±7.64
(18–12
3)80
.47±5.53
(19–99
)13
±1.05
(9–2
4)51
.13±4.22
(30–80
)10
.63±1.51
(5–2
5)0.20
±0.02
(0.09–0.33
)0.18
±0.01
(0.08–0.28
)
0.06
2545
214
1915
.75±5.42
(10–32
)54
.25±15
.61
(20–85
)76
.90±12
.22
(42–96
)11.25±1.65
(9–1
6)45
.25±3.07
(38–53
)10
±3.54
(5–2
0)0.22
±0.08
(0.094–0
.444
)0.19
±0.04
(0.12–0.26
)
0.12
548
152
13.3
22±12
.0(10–34
)72
.5±22
.50
(50–95
)47
.37±47
.38
(0–95)
10.2±1.73
(7–1
5)64
±6.0a
(58–70
)7.5±2.5
(5–1
0)0.12
±0.03
b
(0.09–0.14
)0.10
±0.00
3b
(0.1–0.105
)
0.25
479
00
––
––
––
––
0.5
482
00
––
––
––
––
150
20
0–
––
––
––
–
Fully
fed
Untreated
4716
3415
.19±1.59
(6–28)
62.38±7.64
(18–12
3)80
.47±5.53
(19–99
)13
±1.05
(9–2
4)51
.13±4.22
(30–80
)10
.63±1.51
(5–2
5)0.20
±0.02
(0.09–0.33
)0.18
±0.01
(0.08–0.28
)
0.06
2544
49.1
15.25±3.61
(6–21)
84.75±20
.14
(26–111)
80.18±4.85
(73–95
)15
.25±2.59
(8–1
9)67
.5±7.22
a
(55–80
)10
±2.04
(5–1
5)0.15
±0.03
(0.06–0.19
)0.13
±0.03
(0.09–0.19
)
0.12
540
37.5
9.67
±2.19
a
(7–14)
62.67±25
.28
(9–9
8)61
.48±26
.89
(22–10
9)20
.67±10
.17
(10–41
)52
±8.72
(38–68
)10
±2.89
(5–1
5)0.19
±0.05
(0.13–0.3)
0.18
±0.03
(0.14–0.23
)
0.25
460
0–
––
––
––
–
0.5
450
0–
––
––
––
–
143
00
––
––
––
––
aSignificant
(p<0.05)
bHighlysign
ificant(p<0.01)
1120 Parasitol Res (2011) 109:1113–1123
Author's personal copy

Similarly, Van Der Merwe et al. (2005) noticed that pour-onpreparation of 0.02% abamectin was effective against R.appendiculatus and R. decoloratus naturally infestingcattle. Borges et al. (2008) observed more than 95%efficacy from 21 to 56 days post-treatment and 100% from16 to 22 days post-treatment when they subcutaneouslyinjected a formulation containing both ivermectin andabamectin at the dose of 250 μg/kg into naturally andartificially infected bovines, respectively, with R. microplus.Pereira (2009) applied a subcutaneous injection of abamectinat the dose of 200 μg/kg into calves artificially infested withB. microplus and found that the average count of ticks from3 to 28 days after treatment was 18.41 versus 56.51 in thecontrol group. The current observations also correspond tothose presented by Marzouk et al. (2004) and Montasser(2004) who used ivermectin and doramectin, respectively, asa subcutaneous injection against A. persicus. The effectobserved in the present study strongly confirms that the useof a macrocyclic lactone leads to paralysis in A. persicus and,consequently, to death. This is probably due to theirdocumented role in blocking the synaptic transmissionbetween neurons or between neurons and muscle cells bystimulating the release of γ-amino butyric acid (GABA)(Turner and Schaeffer 1989). In mites infesting animals,100% efficacy was observed in naturally infested cattlewith Sarcoptes scabiei var. bovis after 7 days and inartificially infected ones with Psoroptes ovis after 21 dayspost-treatment following the application of a subcutaneousinjection of abamectin at the dose of 200 μg/kg (Heinze-Mutz et al. 1993). As blood-sucking insects, mosquitoes(Aedes aegypti, Culex quinquefasciatus, and Anophelesquadrimaculatus) have shown high toxicity of a topicalapplication of abamectin, as observed by Pridgeon et al.(2008).
The percentage of fed ticks in the present study wasextremely decreased following abamectin spraying, partic-ularly with the higher doses. This may be partly attributedto the drug drastic effect on mobility affecting the ticks’ability to move to get their meals after spraying. Similarresults have been recorded by Benz and Cox (1989) inwhich abamectin injection at the dose of 200 μg/kgsignificantly reduced the number of engorged female B.microplus from treated cattle for at least 21 days.
The percentage of positive male sexual response andamount of emitted coxal fluid by male or female A. persicussprayed during their unfed period showed a markeddecrease than that recorded from the untreated group,particularly with the higher doses. These observations aresimilar to others reported by Marzouk et al. (2004) andMontasser (2004) who found drastic effects on sexualbehavior and amount of emitted coxal fluid of the presenttick following the use of 400-μg/kg subcutaneous injectionsof ivermectin and doramectin, respectively. The early
mating behavior of A. persicus (Abou El-Kheir 1995) asin other soft ticks (Schlein and Gunders 1981; Mohamed etal. 1990) was found to be regulated by a volatilepheromone emitted from the female coxal opening andassociated with the coxal fluid. Release of the female sexpheromone is related to feeding, which provides thestimulus via hormonal regulation (Sonenshine 1985).Therefore, diminished male sexual response encounteredin the present study may be attributed partly to thedecreased emitted coxal fluid or partly to the disruption ofthe digestive and absorptive processes. The diminishedmobility of the male caused by the drug may also affect itsability to reach the receptive female. The spraying methodmay also be responsible for confusion and suppression ofthe effect of the possibly diminished volatile sex phero-mone found in the notably decreased coxal fluid. Recently,Xu et al. (2010) observed that abamectin-treated femalemoth, Plutella xylostella, produced less sex pheromone andexhibited lower mating percentages with males.
The amount of blood ingested by ticks sprayed thenallowed to feed was higher either in males or females of alltreatments, with a significant increase in the weight offemales treated with 0.25 mL/L than the untreated group.However, blood digestion of male and female ticks sprayedbefore or after feeding revealed a more or less similarpattern as that observed in untreated ones. They exhibited asignificant decrease in their weights between zero and firstweeks after feeding followed by minor significant differ-ences between the successive 5 weeks. Similar results wereobtained in the case of A. persicus fed on pigeonssubcutaneously injected with ivermectin (Marzouk et al.2004) or doramectin (Montasser 2004). However, ivermec-tin was recorded to cause a reduction in weights ofengorged ixodid ticks, which may be due to their differentmode of feeding and route of drug administration (Lancasteret al. 1982; Wilson et al. 1991; Miller et al. 1997). Theincrease in the amount of ingested blood in treated A.persicus eliminates the fact that abamectin may act as afeeding inhibitor but rather exerts its effect on the muscle ornerve cells of the weakened treated ticks, resulting in thedifficulty of withdrawal of their mouthparts and allowingthem to engulf larger amounts of blood.
In the present study, abamectin caused a significantdecrease in the percentage of ovipositing females, reachingzero in higher doses. It also caused a delay in ovipositionperiod and a decrease in fertility and weight of egg.Following abamectin treatment, similar effects on theseparameters were previously observed in the tick B. microplus(Benz and Cox 1989) as well as in other pests such as themite, Tetranychus urticae (Ismail et al. 2007); the sweetpotato whitefly, Bemisia tabaci (Kumar and Poehling 2007);the moth, Marmara gulosa (Grafton-Cardwell et al. 2008);and the liver fluke, Fasciola hepatica (Martin et al. 2009). In
Parasitol Res (2011) 109:1113–1123 1121
Author's personal copy

ticks, the actual volume or weight of condensed blood mealexerts its effects on reproductive development (Diehl et al.1982). However, at suboptimal levels, oogenesis may not beinitiated or, if it begins, will not be sustained. A further generalreduction in the number of eggs produced may be due toprolongation of the period between feeding and oviposition.This suggests that abamectin may also apply its effect throughcertain routes, possibly the nervous system and specificallyvia the neuroendocrine or endocrine mechanism essential inthe regulation of oocyte differentiation and/or vitellogenesis(Pound and Oliver 1979).
The mode of abamectin action in ticks remains hesitant.Zhou et al. (2008) isolated DNA subunit of GABA-activated chloride channel from the moth, P. xylostella,and suggested that it contained the binding site ofabamectin in a dose-dependent manner. Zhu et al. (2010)noticed a significant increase of GABA content inabamectin-resistant strains of the mite Tetranychuscinnabarinus and assumed the increase to be associatedwith resistance against abamectin. Kwon et al. (2010)observed a glutamate-gated chloride channel gene in T.urticae and suggested its relation to abamectin resistance.In ticks, GABA and glutamate are considered as putativeinhibitory or excitatory neuromuscular transmitters, respec-tively (Binnington and Obenchain 1982). Consequently, wemay suggest that abamectin, in a dose-dependent manner,stimulates the release of any of the above neuromusculartransmitters or others in A. persicus, which binds to receptorsites in target cells. This binding results in an increased flowof chloride ions into the nerve and muscle cells, thereforeblocking further functions.
Acknowledgments The authors wish to express their deep gratitudeand appreciation to Dr. Sherif H. El-Alfy, Department of Zoology,Faculty of Science, Ain Shams University, Egypt for his great interestand critical review of the manuscript.
References
Abou El-Kheir, S. M. (1995): Studies on the sex pheromone in thetick Argas (Persicargas) persicus (Oken). Unpublished Ph.D.thesis, Banha University, Egypt
Benz GW, Cox JL (1989) Biochemistry. In: Campbell WC (ed)Ivermectin and abamectin. Springer, New York, pp 230–234
Binnington KC, Obenchain FD (1982) Structure and function of thecirculatory, nervous and neuroendocrine systems of ticks. In:Obenchain FD, Galun R (eds) Physiology of ticks. Pergamon,Oxford, pp 351–398
Borges FA, Silva HC, Buzzulini C, Soares VE, Santos E, Oliveira GP,Costa AJ (2008) Endectocide activity of new long-actionformulation containing 2.25% ivermectin+1.25% abamectin incattle. Vet Parasitol 155:299–307
Cetin H, Cilek JE, Oz E, Aydin L, Deveci O, Yanikoglu A (2009)Comparative efficacy of spinosad with conventional acaricides
against hard and soft tick populations from Antalya, Turkey. VetParasitol 163(1–2):101–104
Civolani S, Cassanelli S, Rivi M, Manicardi GC, Peretto R, Chicca M,Pasqualini E, Leis M (2010) Survey of susceptibility toabamectin of Pear psylla (Hemiptera: Psyllidae) in northernItaly. J Econ Entomol 103(3):816–822
Davey RB, Pound JM, Miller JA, Klavons JA (2010) Therapeutic andpersistent efficacy of a long-acting (LA) formulation ofivermectin against Rhipicephalus (Boophilus) microplus (Acari:Ixodidae) and sera concentration through time in treated cattle.Vet Parasitol 169(1–2):149–156
Diehl PA, Aeschlimann A, Obenchain FD (1982) Tick reproduction:oogenesis and oviposition. In: Obenchain FD, Galun R (eds)Physiology of ticks. Pergamon, Oxford, pp 277–350
Fisher RA (1948) Statistical methods for research workers, 10th edn.Hafner, New York
Grafton-Cardwell EE, Montez GM, Reagan CA, Dunn RA, Ouyang Y(2008) Response of citrus peelminer Marmara gulosa Guillénand Davis (Lepidoptera: Gracillariidae) stages to various insecti-cides. Pest Manag Sci 64(11):1143–1150
Heinze-Mutz EM, Barth D, Cramer LG, Gross SJ, Visser M (1993)Efficacy of abamectin against ectoparasites of cattle. Vet Rec 132(18):455–457
Humeres EC, Morse JG (2005) Baseline susceptibility of persea mite(Acari: Tetranychidae) to abamectin and milbemectin in avocadogroves in Southern California. Exp Appl Acarol 36(1–2):51–59
Ismail MS, Soliman MF, El-Naggar MH, Ghallab MM (2007)Acaricidal activity of spinosad and abamectin against two-spotted spider mites. Exp Appl Acarol 43(2):129–135
Kavallieratos NG, Athanassiou CG, Vayias BJ, Mihail SB, TomanovićZ (2009) Insecticidal efficacy of abamectin against three stored-product insect pests: influence of dose rate, temperature,commodity, and exposure interval. J Econ Entomol 102(3):1352–1359
Khalil GM, Nada SA, Sonenshine DE (1981) Sex pheromoneregulation of mating behavior in the camel tick Hyalommadromedarii (Ixodoidea: Ixodidae). J Parasitol 67:70–76
Khalil GM, Sonenshine DE, Hanafy HA, Abdelmonem AE (1984)Juvenile hormone I effects on the camel tick, Hyalommadromedarii (Acari: Ixodidae). J Med Entomol 21(5):561–566
Koh S, Ahn J, Im A, Tung C, Lee SH, Lee J (2009) Monitoring ofacaricide resistance of Tetranychus urticae (Acari: Tetranychidae)from Korean apple orchards. J Asia Pacific Entomol 12:15–21
Kumar P, Poehling HM (2007) Effects of azadirachtin, abamectin, andspinosad on sweetpotato whitefly (Homoptera: Aleyrodidae) ontomato plants under laboratory and greenhouse conditions in thehumid tropics. J Econ Entomol 100(2):411–420
Kwon DH, Yoon KS, Clark JM, Lee SH (2010) A point mutation in aglutamate-gated chloride channel confers abamectin resistance inthe two-spotted spider mite, Tetranychus urticae Koch. InsectMol Biol 19(4):583–591
Lancaster JL, Simco JS, Kilgore RL (1982) Systematic efficacy ofivermectin MK-933 against the lone star tick. J Econ Entomol75:242–244
Lasota JA, Dybas RA (1990) Abamectin as a pesticide for agriculturaluse. Acta Leiden 59(1–2):217–225
Lin H, Chuan-Hua X, Jin-JunW, Ming L, Wen-Cai L, Zhin-Mo Z (2009)Resistance selection and biochemical mechanism of resistance totwo acaricides in Tetranychus cinnabarinus (Boiduval). PesticideBiochem Physiol 93:47–52
Lohmeyer KH, Miller JA, Pound JM, Oehler DD (2009) Efficacy ofeprinomectin and doramectin against Amblyomma americanum(Acari: Ixodidae) on cattle. J Econ Entomol 102(2):809–814
Martin P, Chambers M, Hennessy D (2009) Efficacy against Fasciolahepatica and the pharmacokinetics of triclabendazole adminis-tered by oral and topical routes. Aust Vet J 87(5):200–203
1122 Parasitol Res (2011) 109:1113–1123
Author's personal copy

Marzouk AS, Swelim HH, Montasser AA, Gadallah AI (2004)Biological effects of ivermectin on the fowl tick Argas(Persicargas) persicus (Oken) (Ixodoidea: Argasidae). Egypt JHosp Med 17:93–105
Meister RT (1992) Farm chemicals handbook '92. Meister Publishing,Willoughby
Miller JA, Garris GI, Oehler DD (1997) Control of lone star ticks oncattle with ivermectin. J Agric Entomol 14(2):199–204
Mohamed FS, Khalil GM, Marzouk AS, Roshdy MA (1990) Sexpheromone regulation of mating behaviour in the tick Ornithodoros(Ornithodoros) savignyi (Audouin) (Acari: Argasidae). J MedEntomol 27:288–294
Montasser AA (2004) Doramectin effects on the soft tick, Argas(Persicargas) persicus (Ixodoidea: Argasidae). J Egypt Ger SocZool 43(D):147–157
Pereira JR (2009) The efficiency of avermectins (abamectin, doramectinand ivermectin) in the control of Boophilus microplus, in artificiallyinfested bovines kept in field conditions. Vet Parasitol 162(1–2):116–119
Pound JM, Oliver JH (1979) Juvenile hormone: evidence of its role inthe reproduction of ticks. Science 206:355–357
Pridgeon JW, Pereira RM, Becnal JJ, Allan SA, Clark GG, Linthicum KJ(2008) Susceptibility of Aedes aegypti, Culex quinquefasciatus Say,and Anopheles quadrimaculatus Say to 19 pesticides with differentmodes of action. J Med Entomol 45(1):82–87
Quilez J, Sanchez-Acedo C, Avendano C, Cacho ED, Lopez-Bernad F(2005) Efficacy of two peroxygen-based disinfectants forinactivation of Cryptosporidium parvum oocysts. Appl EnvironMicrobiol 71:2479–2483
Roberts LS, Janovy J (2005) Foundations of parasitology, 7th edn.McGraw Hill, Boston, pp 644–646
Sajid MS, Iqbal Z, Khan MN, Muhammad G (2009) In vitro and in vivoefficacies of ivermectin and cypermethrin against the cattle tickHyalomma anatolicum anatolicum (Acari: Ixodidae). Parasitol Res105(4):1133–1138
Schlein Y, Gunders AE (1981) Pheromone of Ornithodoros spp.(Argasidae) in the coxal fluid of female ticks. Parasitology82:467–471
Shah AH, Khan MN, Iqbal Z, Safid MS, Akhtar MS (2006) Someepidemiological aspects and vector role of tick infestation onlayers in the Faisalabad district (Pakistan). World Poult Sci J62:145–157
Sonenshine DE (1985) Pheromones and other semiochemicals of theAcari. Annu Rev Entomol 30:1–28
Turner MJ, Schaeffer JM (1989) Biochemistry. In: Campbell WC (ed)Ivermectin and abamectin. Springer, New York, pp 73–89
Van Der Merwe JS, Smit FJ, Durand AM, Krüger LP, Michael LM(2005) Acaricide efficiency of amitraz/cypermethrin and aba-mectin pour-on preparations in game. Onderstepoort J Vet Res 72(4):309–314
Venzon M, Rosado MC, Molina-Rugama AJ, Duarte VS, Dias R,Pallini A (2008) Acaricidal efficacy of neem against Polyphago-tarsonemus latus (Banks) (Acari: Tarsonemidae). Crop Protect 27(3/5):869–872
Wall R, Shearer D (1997) Veterinary entomology. Chapman and Hall,London, pp 97–140, 132–133
Wilson KJ, Hair JA, Sauer TR, Weeks DL (1991) Effect ofivermectin on the volume of blood ingested by two species ofticks (Acari: Ixodidae) feeding on cattle. J Med Entomol 28(3):465–468
Xu Z, Cao GC, Dong SL (2010) Changes of sex pheromonecommunication systems associated with Tebufenozide andabamectin resistance in Diamondback moth, Plutella xylostella(L.). J Chem Ecol 36(5):526–534
Yee WL, Phillips PA, Faber BA (2001) Effects of aerial sprayvolume, coverage, and abamectin on Scirtothrips perseae(Thysanoptera: Thripidae). J Econ Entomol 94(5):1090–1096
Zhou XM, Wu QJ, Zhang YJ, Bai LY, Huang XY (2008) Cloningand characterization of a GABA receptor from Plutellaxylostella (Lepidoptera: Plutellidae). J Econ Entomol 101(6):1888–1896
Zhu XJ, Lu WC, Feng YN, He L (2010) High γ-aminobutyric acidcontent, a novel component associated with resistance toabamectin in Tetranychus cinnabarinus (Boisduval). J InsectPhysiol 56(12):1895–1900
Parasitol Res (2011) 109:1113–1123 1123
Author's personal copy
Copyright © 2022 FDOKUMEN




















