
Egg wax from the cattle tick Rhipicephalus (Boophilus) microplus inhibits Pseudomonas aeruginosa...
11
Ticks and Tick-borne Diseases 4 (2013) 366–376 Contents lists available at ScienceDirect Ticks and Tick-borne Diseases j ourna l ho me p age: www.elsevier.com/locate/ttbdis Egg wax from the cattle tick Rhipicephalus (Boophilus) microplus inhibits Pseudomonas aeruginosa biofilm Karine R. Zimmer a , Alexandre J. Macedo a,b , Gianlucca G. Nicastro c , Regina L. Baldini c , Carlos Termignoni a,∗ a Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul, Av. Bento Gonc ¸ alves 9500, 91501-970, Porto Alegre, RS, Brazil b Faculdade de Farmácia, Universidade Federal do Rio Grande do Sul, Av. Ipiranga 2752, 90610-000, Porto Alegre, RS, Brazil c Departamento de Bioquímica, Instituto de Química, Universidade de São Paulo, Av. Prof. Lineu Prestes 748, 05508-000, São Paulo, SP, Brazil a r t i c l e i n f o Article history: Received 5 September 2012 Accepted 10 January 2013 Available online 11 April 2013 Keywords: Biofilm Pseudomonas aeruginosa Rhipicephalus (Boophilus) microplus Egg wax Protective mechanisms a b s t r a c t Rhipicephalus (Boophilus) microplus is constantly challenged during its life cycle by microorganisms present in their hosts or in the environment. Tick eggs may be especially vulnerable to environmental conditions because they are exposed to a rich and diverse microflora in the soil. Despite being oviposited in such hostile sites, tick eggs remain viable, suggesting that the egg surface has defense mechanisms against opportunistic and/or pathogenic organisms. R. microplus engorged females deposit a superficial wax layer onto their eggs during oviposition. This egg wax is essential for preventing desiccation as well as acting as a barrier against attack by microorganisms. In this study, we report the detection of anti-biofilm activity of R. microplus egg wax against Pseudomonas aeruginosa PA14. Genes involved in the functions of production and maintenance of the biofilm extracellular matrix, pelA and cdrA, respectively, were markedly downregulated by a tick egg-wax extract. Moreover, this extract strongly inhibited fliC gene expression. Instead of a compact extracellular matrix, P. aeruginosa PA14 treated with egg-wax extract produces a fragile one. Also, the colony morphology of cells treated with egg-wax extract appears much paler and brownish, instead of the bright purple characteristic of normal colonies. Swarming motility was also inhibited by treatment with the egg-wax extract. The inhibition of P. aeruginosa biofilm does not seem to depend on inhibition of the quorum sensing system since mRNA levels of the 3 regulators of this system were not inhibited by egg-wax extract. © 2013 Elsevier GmbH. All rights reserved. Introduction Arthropods have a variety of defense mechanisms against microbial infection. In this context, a number of active molecules have been identified in tick species, such as antimicrobial com- pounds (Pavis et al., 1994; Potterat et al., 1997; Johns et al., 1998; Fogac ¸ a et al., 1999; Lai et al., 2004; Arrieta et al., 2006; Esteves et al., 2009; Kopácek et al., 2010; Oldiges et al., 2012; Yu et al., 2012), lysozymes (Kopácek et al., 1999; Grunclová et al., 2003), pro- tease inhibitors (Fogac ¸ a et al., 2004), and lectins (Grubhoffer et al., 2005). Considering the environmental conditions in the soil where tick eggs are deposited, one might expect the eggs to be vulnerable to microbial attack (Booth, 1992; Esteves et al., 2009). However, eggs remain viable in the ground, with no signs of bacterial coloniza- tion. All ticks possess a specialized gland, called Géné’s organ that ∗ Corresponding author. Tel.: +55 51 33086082; fax: +55 51 33087309. E-mail addresses: [email protected], [email protected] (C. Termignoni). produces a wax that is deposited on the egg surface during oviposi- tion. In addition to water-proofing the egg, the wax acts as a barrier to attack by invading microorganisms (Booth, 1992). Pseudomonas aeruginosa is an opportunistic pathogen found in a variety of soils (Kaszab et al., 2011). Also, P. aeruginosa has been extensively utilized as a model organism for the study of biofilms (Friedman and Kolter, 2004a). Among the virulence factors that account for the pathogenicity of this bacterium is biofilm forma- tion. Biofilm formation is a complex and well-orchestrated process involving different sequential steps: (i) planktonic cells encounter a biotic or abiotic surface; (ii) initial attachment (a reversible process); (iii) formation of microcolonies; and (iv) biofilm matura- tion and development. During biofilm formation, an extracellular polysaccharide (EPS) is produced, and it acts as a glue to keep microbial cells together, providing structural support and facili- tating microbial attachment. Although the exact composition of the matrix is uncertain, extracellular polysaccharides, protein, and nucleic acids have shown to be present (Flemming and Wingender, 2010). Microorganisms living in biofilms exhibit high resistance to antibiotics and host immune defenses, and so they are difficult to eradicate (Wang et al., 2008). 1877-959X/$ – see front matter © 2013 Elsevier GmbH. All rights reserved. http://dx.doi.org/10.1016/j.ttbdis.2013.01.005
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Egg wax from the cattle tick Rhipicephalus (Boophilus) microplus inhibits Pseudomonas aeruginosa...

EP
K
a
b
c
a
ARAA
KBPREP
I
mhpFe2t2
emrt
c
1h
Ticks and Tick-borne Diseases 4 (2013) 366– 376
Contents lists available at ScienceDirect
Ticks and Tick-borne Diseases
j ourna l ho me p age: www.elsev ier .com/ locate / t tbd is
gg wax from the cattle tick Rhipicephalus (Boophilus) microplus inhibitsseudomonas aeruginosa biofilm
arine R. Zimmera, Alexandre J. Macedoa,b, Gianlucca G. Nicastroc, Regina L. Baldini c, Carlos Termignonia,∗
Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul, Av. Bento Gonc alves 9500, 91501-970, Porto Alegre, RS, BrazilFaculdade de Farmácia, Universidade Federal do Rio Grande do Sul, Av. Ipiranga 2752, 90610-000, Porto Alegre, RS, BrazilDepartamento de Bioquímica, Instituto de Química, Universidade de São Paulo, Av. Prof. Lineu Prestes 748, 05508-000, São Paulo, SP, Brazil
r t i c l e i n f o
rticle history:eceived 5 September 2012ccepted 10 January 2013vailable online 11 April 2013
eywords:iofilmseudomonas aeruginosahipicephalus (Boophilus) microplusgg waxrotective mechanisms
a b s t r a c t
Rhipicephalus (Boophilus) microplus is constantly challenged during its life cycle by microorganismspresent in their hosts or in the environment. Tick eggs may be especially vulnerable to environmentalconditions because they are exposed to a rich and diverse microflora in the soil. Despite being ovipositedin such hostile sites, tick eggs remain viable, suggesting that the egg surface has defense mechanismsagainst opportunistic and/or pathogenic organisms. R. microplus engorged females deposit a superficialwax layer onto their eggs during oviposition. This egg wax is essential for preventing desiccation as well asacting as a barrier against attack by microorganisms. In this study, we report the detection of anti-biofilmactivity of R. microplus egg wax against Pseudomonas aeruginosa PA14. Genes involved in the functionsof production and maintenance of the biofilm extracellular matrix, pelA and cdrA, respectively, weremarkedly downregulated by a tick egg-wax extract. Moreover, this extract strongly inhibited fliC gene
expression. Instead of a compact extracellular matrix, P. aeruginosa PA14 treated with egg-wax extractproduces a fragile one. Also, the colony morphology of cells treated with egg-wax extract appears muchpaler and brownish, instead of the bright purple characteristic of normal colonies. Swarming motilitywas also inhibited by treatment with the egg-wax extract. The inhibition of P. aeruginosa biofilm doesnot seem to depend on inhibition of the quorum sensing system since mRNA levels of the 3 regulators ofthis system were not inhibited by egg-wax extract.ntroduction
Arthropods have a variety of defense mechanisms againsticrobial infection. In this context, a number of active molecules
ave been identified in tick species, such as antimicrobial com-ounds (Pavis et al., 1994; Potterat et al., 1997; Johns et al., 1998;ogac a et al., 1999; Lai et al., 2004; Arrieta et al., 2006; Estevest al., 2009; Kopácek et al., 2010; Oldiges et al., 2012; Yu et al.,012), lysozymes (Kopácek et al., 1999; Grunclová et al., 2003), pro-ease inhibitors (Fogac a et al., 2004), and lectins (Grubhoffer et al.,005).
Considering the environmental conditions in the soil where tickggs are deposited, one might expect the eggs to be vulnerable to
icrobial attack (Booth, 1992; Esteves et al., 2009). However, eggsemain viable in the ground, with no signs of bacterial coloniza-ion. All ticks possess a specialized gland, called Géné’s organ that
∗ Corresponding author. Tel.: +55 51 33086082; fax: +55 51 33087309.E-mail addresses: [email protected],
[email protected] (C. Termignoni).
877-959X/$ – see front matter © 2013 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.ttbdis.2013.01.005
© 2013 Elsevier GmbH. All rights reserved.
produces a wax that is deposited on the egg surface during oviposi-tion. In addition to water-proofing the egg, the wax acts as a barrierto attack by invading microorganisms (Booth, 1992).
Pseudomonas aeruginosa is an opportunistic pathogen found ina variety of soils (Kaszab et al., 2011). Also, P. aeruginosa has beenextensively utilized as a model organism for the study of biofilms(Friedman and Kolter, 2004a). Among the virulence factors thataccount for the pathogenicity of this bacterium is biofilm forma-tion. Biofilm formation is a complex and well-orchestrated processinvolving different sequential steps: (i) planktonic cells encountera biotic or abiotic surface; (ii) initial attachment (a reversibleprocess); (iii) formation of microcolonies; and (iv) biofilm matura-tion and development. During biofilm formation, an extracellularpolysaccharide (EPS) is produced, and it acts as a glue to keepmicrobial cells together, providing structural support and facili-tating microbial attachment. Although the exact composition ofthe matrix is uncertain, extracellular polysaccharides, protein, and
nucleic acids have shown to be present (Flemming and Wingender,2010). Microorganisms living in biofilms exhibit high resistance toantibiotics and host immune defenses, and so they are difficult toeradicate (Wang et al., 2008).
ck-bor
ttpldlvb
malad(ErH
mbea
aamweee
M
A
tGrshpb
E
catmwmp(eswatFu
K.R. Zimmer et al. / Ticks and Ti
Many bacterial virulence factors, including biofilm forma-ion, are controlled by a system of bacterial communication,he quorum sensing system (QS; Hengge, 2009). P. aeruginosaossesses 2 N-acylhomoserine lactone (AHL)-regulated systems:
asR-lasI and rhlR-rhlI, with the cognate signal molecules N-(3-oxo-odecanoyl)-L-homoserine lactone and N-butyryl-L-homoserine
actone, respectively. A third cell-signaling system, mvfR (multipleirulence factor R), regulates Pseudomonas quinolone signal (PQS)iosynthesis (Zhu et al., 2004).
A second messenger, bis-(3′-5′)-cyclic dimeric guanosineonophosphate (c-di-GMP), controls a transition between a motile
nd a biofilm lifestyle (Hengge, 2009). c-di-GMP is a positive regu-ator of biofilm formation, EPS production, and pellicle formation,nd a negative regulator of swarming motility. Regulation of c-i-GMP levels involves 2 types of enzyme: diguanylate cyclasesDGCs), with GGDEF motifs, and phosphodiesterases (PDEs), withAL motifs, responsible for synthesis and breakdown of c-di-GMP,espectively (Römling and Amikam, 2006; Cotter and Stibitz, 2007;engge, 2009).
The discovery of compounds capable of inhibiting biofilm for-ation is an issue of extreme interest because it is estimated that
iofilms affect at least 65% of human bacterial infections (Costertont al., 2003). Also, biofilm-related infections are found in plants andnimals (Olson et al., 2002).
In the current work, we report the detection of anti-biofilmctivity of Rhipicephalus microplus cattle tick egg-wax against P.eruginosa PA14. We show that genes involved in production andaintenance of the biofilm extracellular matrix, pelA and cdrA,ere repressed by egg-wax extract. We also demonstrated that this
xtract inhibits fliC gene expression. The structure and other prop-rties of the molecule responsible for this activity will be describedlsewhere.
aterials and methods
nimals and eggs
R. microplus were obtained from the colony maintained athe Faculdade de Veterinária at the Universidade Federal do Riorande do Sul, Porto Alegre, Brazil. Engorged females fed to
epletion were rinsed with water, dried with tissue paper, andtored in the incubator for oviposition (28 ◦C and high relativeumidity). The eggs were collected at the end of oviposition androcessed for extraction of the egg wax as described immediatelyelow.
xtraction of the tick egg wax
We adapted the method of Arrieta et al. (2006). Briefly, we addedhloroform and methanol (2:1, v/v) to eggs (10 ml/g) under mildgitation for 60 min. The supernatant was transferred to a glassube. Thereafter, Milli-Q water was added to the eggs (5 ml/g) and
aintained under mild agitation again for 60 min. This supernatantas collected and mixed with the first supernatant. The solventixture was centrifuged for 10 min at 1000 rpm to separate the
hases. The solvents were evaporated under a nitrogen streamchloroform:methanol extract) or lyophilized (water:methanolxtract), and the extracts were then weighed. The samples weretored at −80 ◦C until use. The extracts were prepared in Milli-Qater at the following concentrations in order to be used in the
ssays: 0.3125, 0.625, 1.25, 2.5, and 5.0 mg/ml. In some assays, onlyhe extract at the lowest active concentration (2.5 mg/ml) was used.or control samples, the extract was replaced by an equivalent vol-me of Milli-Q water.
ne Diseases 4 (2013) 366– 376 367
Bacterial strain and growth conditions
In all assays, P. aeruginosa strain PA14 (Rahme et al., 1995) wasgrown in Luria-Bertani broth (LB; peptone 10 g/l, yeast extract 5 g/l,and NaCl 10 g/l) at 37 ◦C on a rotary shaker (150 rpm) (IncubatorShaker Series Excella E25, New Brunswick Scientific) and adjustedto an optical density (OD) at 600 nm equivalent to 108 CFU/ml.
Anti-biofilm assay
Standard sterile 96-well polystyrene flat bottom microtiterplates (Costar 3599 Corning, Inc. NY, USA) were used to evaluatebiofilm formation according to a protocol adapted from Trentinet al. (2011). In the biofilm formation inhibition assay (anti-biofilm assay), 100 �l of diluted bacterial suspension (equivalent to108 CFU/ml), 50 �l of LB broth and 100 �l of extract were added tothe wells and incubated at 37 ◦C for 24 h. In preformed biofilm assay(biofilm eradication assay), 100 �l of diluted bacterial suspension(equivalent to 108 CFU/ml) and 100 �l of LB broth were added towells and incubated at 37 ◦C for 24 h. Then the wells were washed 3times with 200 �l of sterile saline 0.9% and filled with 100 �l of egg-wax extract and 100 �l of LB broth and incubated again at 37 ◦C for24 h. Controls (untreated) were prepared with Milli-Q water replac-ing the extract. Following this incubation time (anti-biofilm andbiofilm eradication assays), the content of wells was removed andwashed 3 times with 200 �l of sterile saline 0.9%. The remainingattached bacteria were heat-fixed by exposing them to hot air at60 ◦C for 1 h. The adherent biofilm layer formed was stained with200 �l of 0.4% crystal violet for 15 min at room temperature. Excessstain was removed by placing plates under running tap water untilthe flowing water appeared colorless. The dye bound to the cellswas solubilized with 99.5% DMSO (Sigma, St. Louis, MO, USA), andthe absorbance (570 nm) was measured (Spectramax M2e Multi-mode Microplate Reader, Molecular Devices, Sunnyvale, CA, USA).
Biofilm formation was also evaluated in glass tubes by inoc-ulating a bacterial suspension in each tube (2 ml, equivalent to108 CFU/ml), LB broth (1 ml), and tick egg-wax extract (2 ml at aconcentration of 2.5 mg/ml). The tubes were then kept at 37 ◦Cfor 24 h. They were then washed with sterile 0.9% NaCl andstained with 0.4% crystal violet. The dye bound to the cells wasthen solubilized with DMSO. Absorbance at 570 nm was measuredon a Spectramax M2e Multimode Microplate Reader (MolecularDevices, Sunnyvale, CA, USA).
Antibacterial assay
The bacterial growth assay was evaluated by the difference ofinitial (t = 0) and final (t = 24 h) absorbance at 600 nm (Musk et al.,2005). Gentamicin sulphate (6.5 �g/ml; Sigma–Aldrich Co.®, USA)added to LB broth served as a growth inhibition control in all assays.
Scanning electron microscopy (SEM)
P. aeruginosa PA14 biofilm was grown in sterile 96-wellpolystyrene flat bottom microtiter plates (Costar 3599 Corning, Inc.NY, USA) holding a coverslip using LB broth and Milli-Q water inthe control sample (untreated) and LB broth and extract (at a con-centration of 2.5 mg/ml) in the treatment group for 24 h at 37 ◦C.The samples were washed with sterile 0.9% NaCl to remove plank-tonic cells. Then the biofilm was fixed with 2.5% glutaraldehyde(prepared in 100 mM cacodylate buffer pH 7.2) for 3 h and washed
with 100 mM cacodylate buffer (pH 7.2). The samples were thendehydrated with increasing concentrations of acetone, dried to theCO2 critical point using a Bal-Tec 030 apparatus (CPD 030 Balzers,Liechtenstein), and placed on a mounting base. Finally, the samples
368 K.R. Zimmer et al. / Ticks and Tick-bor
Table 1Primers used in qRT-PCR.
Primer name Sequence
rhlR up CCACACGATTCCCTTCACCrhlR low TCGCTCCAGACCACCATTTlasR up CGCGAAAGCAGCACGAGTTlasR low TTCCGCTTCCACGCTGAGGmvfR up GCCATCCCGCCGTCGTTCTmvfR low TCCGCGTTGTCCTGCTTGATpelA up CACTTCTATTCGGGGACCAApelA low TGTAGTCGCTCATCCACAGCPA14 61200 up ACAGCCAGTTCAACGACCTCPA14 61200 low TGAAATACTCGCTGCCATTGfliC L GCAACAGGCCGAACTGACfliC R GGTCTCGTAGGCGTTGGArsmA up GGGTCGGAGAGACCCTGArsmA low GTGCACGGCGACTTCCTTrsmZ up ACACGCAACCCCGAAGGArsmZ low GTATTACCCGCCCACTCTpilA-LEFT TGCCGGCATATCAGGATTpilA-RIGHT CGTTACCGGCTTGAGCATpslE up GTCGACAGCCTGGAGAAATCpslE down AACAGCATCAGCAGGGTTTTcupC L ATGGCAAGCTGCTGAACCcupC R TAAGCAGCCCGCAGATTCcupD1 L ACTATCGCCGCATTTCTCcupD1 R ACGGTGTCGAGTATCACGoprF L ACAGCGTTCGCAACATGA
wm
Rr
aewswtTep
P
wbtaad
(aCwoF
C
t
in all directions in the wells-containing media, whereas bacteria
oprF R TAGGTGCCACGAACGTCAPA14 58690 up TGACGTCATCAACCTGGTGTPA14 58690 down CGGCTGAAGAAGGTCTTGTC
ere coated with gold and examined under a scanning electronicroscope (model JEOL Instrument JSM-6060) at 10 kV.
NA extraction and quantitative real-time polymerase chaineaction (qRT-PCR)
Static cultures were grown as mentioned above in anti-biofilmssay and utilized for RNA extraction. In this assay, we used thextract at a concentration of 2.5 mg/ml. Total RNA was extractedith Trizol (Invitrogen), treated with DNase I, and used for cDNA
ynthesis with Improm II (Promega) and random primers. cDNAas amplified with specific primers using SYBR Green PCR Mas-
er Mix in the 7300 Real-Time PCR System (Applied Biosystems).he threshold cycle data analysis (Livak and Schmittgen, 2001) wasmployed. nadB was used for normalization of cDNA amount. Therimer sequences used throughout this study are listed in Table 1.
. aeruginosa PA14 colony morphology
Bacterial cells from cultures (cultured in 10 g/l tryptone) treatedith tick egg-wax extract (2.5 mg/ml), as described in the anti-
iofilm assay, were spotted (15 �l) on Congo-Red plates (10 g/lryptone, 40 �g/ml Congo-Red, 20 �g/ml Coomassie Brilliant Blue,nd 1% agar). Plates were incubated at 37 ◦C for 6 days for visu-lization of the colony morphology (adapted from the methodeveloped by Friedman and Kolter, 2004a).
Colony morphology was also evaluated spotting bacterial cells5 �l) from cultures (cultured in 10 g/l tryptone) as described in thenti-biofilm assay in Congo-Red plates (10 g/l tryptone, 40 �g/mlongo-Red, 20 �g/ml Coomassie Brilliant Blue, and 1% agar). Platesere incubated at room temperature for 6 days for visualization
f colony morphology (adapted from the method described byriedman and Kolter, 2004a).
ongo-Red assay
Pellicle morphology and the ability of P. aeruginosa PA14o absorb the dye present in the medium were evaluated as
ne Diseases 4 (2013) 366– 376
follows: Bacteria (2 ml/108 CFU/ml), 1 ml of LB broth supplementedwith Congo-Red, and 2 ml of tick egg-wax extract (2.5 mg/ml)were placed in glass tubes and left at room temperature for 6days. The pellicles were removed with a bacteriological loop andplaced in water (adapted from the method developed by Friedmanand Kolter, 2004a). Control samples (untreated) were included, inwhich the extract was replaced by Milli-Q water.
Matrix weight
The P. aeruginosa PA14 matrix from the assay mentioned above(Congo-Red assay) was collected, washed with water, lyophilizedin pre-weighed plastic tubes, and its weight was recorded.
Pyocyanin and LasB elastase measurements/proteolytic activity inculture supernatants
Pyocyanin, LasB elastase, and proteolytic activity assays werecarried out using the culture supernatants. Bacterial suspensions ofP. aeruginosa PA14 (8 ml/108 CFU/ml) were cultured with LB broth(4 ml) and treated with tick egg-wax extract (8 ml) for 24 h at 37 ◦C.The culture supernatants were then collected by centrifugation.The amount of pyocyanin in culture supernatants was measuredspectrophotometrically at 695 nm (Fothergill et al., 2007).
LasB activity in the culture supernatants was determined by theelastin Congo-Red (ECR) assay, adapted from Tron et al. (2004).Briefly, 10 mg of ECR beads were incubated with 0.1 ml super-natant in 0.9 ml 10 mM sodium phosphate buffer, pH 7.0, rotatingovernight at 37 ◦C. Insoluble ECR was precipitated by centrifu-gation, and the absorbance of the supernatant was measured at495 nm.
Proteolytic activity was measured on casein agar plates. Thesupernatant (50 �l) was added to wells in the plates. The plateswere incubated for 24 h at 37 ◦C (Kong et al., 2005). The zone ofcasein hydrolysis was indicated by a clear zone in the plates. Thecontrol sample consisted of Milli-Q water instead of extract.
Sensitivity to oxidative stress
Aliquots from bacterial cultures of P. aeruginosa PA14 (100 �l)cultivated in LB broth (50 �l) for 24 h in the presence of tick egg-wax extract (100 �l) were spread uniformly onto LB agar plates.The control sample contained Milli-Q water instead of the extract.Then aluminum cylinders with a diameter of 6 mm were placed intriplicate on each plate and loaded with 100 �l 3% H2O2. The areaof growth inhibition was measured following incubation at 37 ◦C(Fonseca et al., 2004).
Swimming motility
We utilized the method of Malapaka et al. (2007). Motilityagar was prepared (10 g/l tryptone, 0.5 g/l NaCl, and 0.3 g/l agar),and 100 �l was added to each well of a standard sterile 96-wellpolystyrene flat bottom microtiter plates (Costar 3599 Corning, Inc.NY, USA). Egg-wax extract (100 �l) was added to each well andthen mixed to ensure an even distribution within the well; theculture medium was allowed to solidify overnight at room tem-perature. Then 10 �l of bacterial cells, treated or untreated, wereinoculated in the wells. The offset-inoculation method was used todetect the inhibition of motility. With this method, bacteria spread
whose motility is inhibited grow only in the immediate vicinity ofthe inoculation zone. The plates were then incubated for 15 h at37 ◦C. Absorbance at 550 nm was measured on a Spectramax M2e,Multimode Microplate Reader.
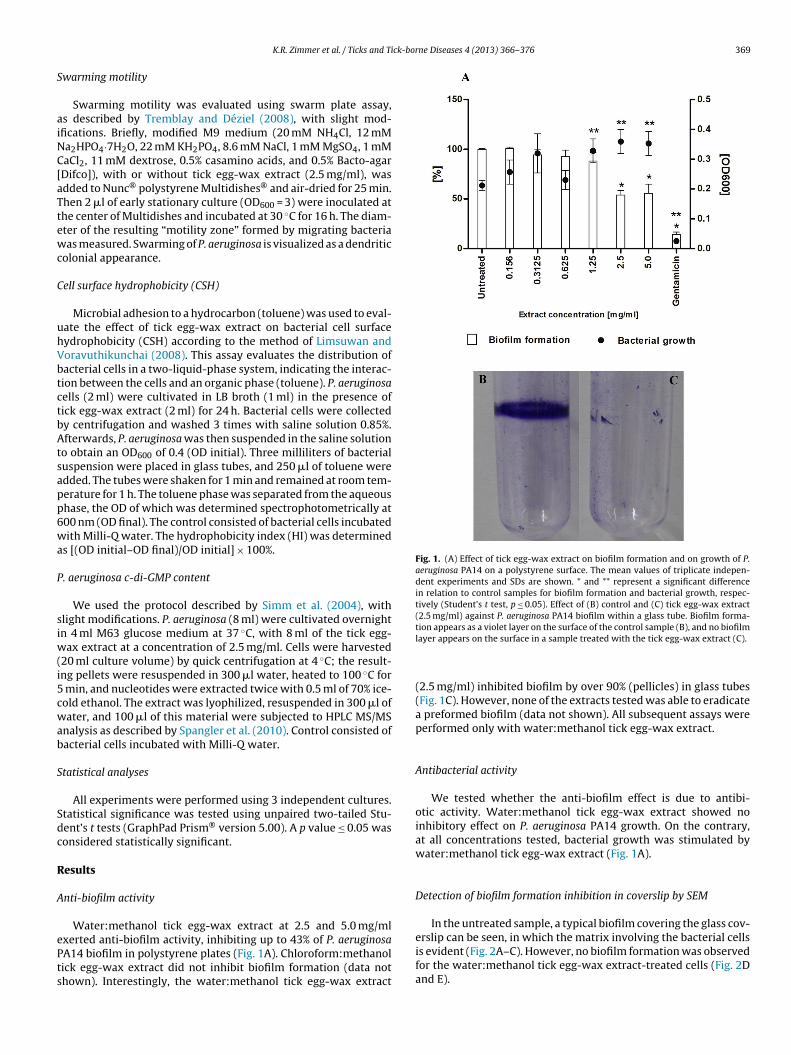
ck-borne Diseases 4 (2013) 366– 376 369
S
aiNC[aTtewc
C
uhVbtctbAtsapp6wa
P
siw(i5cwab
S
Sdc
R
A
ePts
Fig. 1. (A) Effect of tick egg-wax extract on biofilm formation and on growth of P.aeruginosa PA14 on a polystyrene surface. The mean values of triplicate indepen-dent experiments and SDs are shown. * and ** represent a significant differencein relation to control samples for biofilm formation and bacterial growth, respec-tively (Student’s t test, p ≤ 0.05). Effect of (B) control and (C) tick egg-wax extract(2.5 mg/ml) against P. aeruginosa PA14 biofilm within a glass tube. Biofilm forma-tion appears as a violet layer on the surface of the control sample (B), and no biofilm
K.R. Zimmer et al. / Ticks and Ti
warming motility
Swarming motility was evaluated using swarm plate assay,s described by Tremblay and Déziel (2008), with slight mod-fications. Briefly, modified M9 medium (20 mM NH4Cl, 12 mMa2HPO4·7H2O, 22 mM KH2PO4, 8.6 mM NaCl, 1 mM MgSO4, 1 mMaCl2, 11 mM dextrose, 0.5% casamino acids, and 0.5% Bacto-agarDifco]), with or without tick egg-wax extract (2.5 mg/ml), wasdded to Nunc® polystyrene Multidishes® and air-dried for 25 min.hen 2 �l of early stationary culture (OD600 = 3) were inoculated athe center of Multidishes and incubated at 30 ◦C for 16 h. The diam-ter of the resulting “motility zone” formed by migrating bacteriaas measured. Swarming of P. aeruginosa is visualized as a dendritic
olonial appearance.
ell surface hydrophobicity (CSH)
Microbial adhesion to a hydrocarbon (toluene) was used to eval-ate the effect of tick egg-wax extract on bacterial cell surfaceydrophobicity (CSH) according to the method of Limsuwan andoravuthikunchai (2008). This assay evaluates the distribution ofacterial cells in a two-liquid-phase system, indicating the interac-ion between the cells and an organic phase (toluene). P. aeruginosaells (2 ml) were cultivated in LB broth (1 ml) in the presence ofick egg-wax extract (2 ml) for 24 h. Bacterial cells were collectedy centrifugation and washed 3 times with saline solution 0.85%.fterwards, P. aeruginosa was then suspended in the saline solution
o obtain an OD600 of 0.4 (OD initial). Three milliliters of bacterialuspension were placed in glass tubes, and 250 �l of toluene weredded. The tubes were shaken for 1 min and remained at room tem-erature for 1 h. The toluene phase was separated from the aqueoushase, the OD of which was determined spectrophotometrically at00 nm (OD final). The control consisted of bacterial cells incubatedith Milli-Q water. The hydrophobicity index (HI) was determined
s [(OD initial–OD final)/OD initial] × 100%.
. aeruginosa c-di-GMP content
We used the protocol described by Simm et al. (2004), withlight modifications. P. aeruginosa (8 ml) were cultivated overnightn 4 ml M63 glucose medium at 37 ◦C, with 8 ml of the tick egg-
ax extract at a concentration of 2.5 mg/ml. Cells were harvested20 ml culture volume) by quick centrifugation at 4 ◦C; the result-ng pellets were resuspended in 300 �l water, heated to 100 ◦C for
min, and nucleotides were extracted twice with 0.5 ml of 70% ice-old ethanol. The extract was lyophilized, resuspended in 300 �l ofater, and 100 �l of this material were subjected to HPLC MS/MS
nalysis as described by Spangler et al. (2010). Control consisted ofacterial cells incubated with Milli-Q water.
tatistical analyses
All experiments were performed using 3 independent cultures.tatistical significance was tested using unpaired two-tailed Stu-ent’s t tests (GraphPad Prism® version 5.00). A p value ≤ 0.05 wasonsidered statistically significant.
esults
nti-biofilm activity
Water:methanol tick egg-wax extract at 2.5 and 5.0 mg/ml
xerted anti-biofilm activity, inhibiting up to 43% of P. aeruginosaA14 biofilm in polystyrene plates (Fig. 1A). Chloroform:methanolick egg-wax extract did not inhibit biofilm formation (data nothown). Interestingly, the water:methanol tick egg-wax extractlayer appears on the surface in a sample treated with the tick egg-wax extract (C).
(2.5 mg/ml) inhibited biofilm by over 90% (pellicles) in glass tubes(Fig. 1C). However, none of the extracts tested was able to eradicatea preformed biofilm (data not shown). All subsequent assays wereperformed only with water:methanol tick egg-wax extract.
Antibacterial activity
We tested whether the anti-biofilm effect is due to antibi-otic activity. Water:methanol tick egg-wax extract showed noinhibitory effect on P. aeruginosa PA14 growth. On the contrary,at all concentrations tested, bacterial growth was stimulated bywater:methanol tick egg-wax extract (Fig. 1A).
Detection of biofilm formation inhibition in coverslip by SEM
In the untreated sample, a typical biofilm covering the glass cov-erslip can be seen, in which the matrix involving the bacterial cells
is evident (Fig. 2A–C). However, no biofilm formation was observedfor the water:methanol tick egg-wax extract-treated cells (Fig. 2Dand E).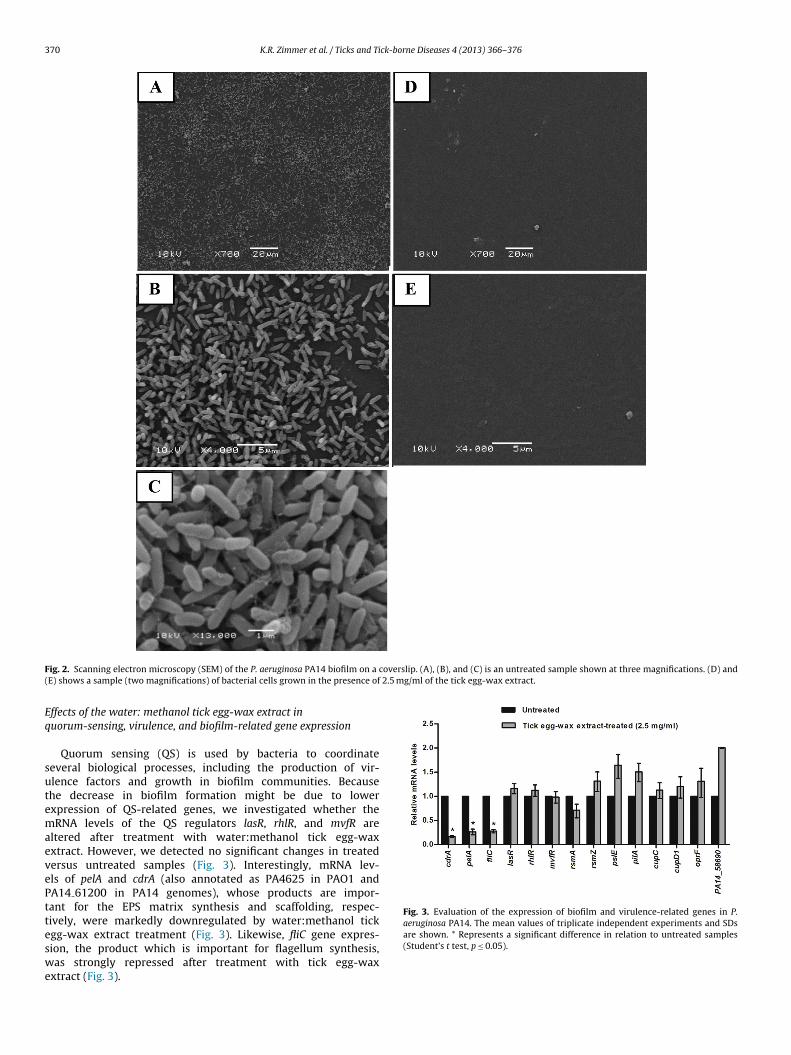
370 K.R. Zimmer et al. / Ticks and Tick-borne Diseases 4 (2013) 366– 376
F overslip. (A), (B), and (C) is an untreated sample shown at three magnifications. (D) and( 2.5 mg/ml of the tick egg-wax extract.
Eq
sutemaevePtteswe
Fig. 3. Evaluation of the expression of biofilm and virulence-related genes in P.aeruginosa PA14. The mean values of triplicate independent experiments and SDs
ig. 2. Scanning electron microscopy (SEM) of the P. aeruginosa PA14 biofilm on a cE) shows a sample (two magnifications) of bacterial cells grown in the presence of
ffects of the water: methanol tick egg-wax extract inuorum-sensing, virulence, and biofilm-related gene expression
Quorum sensing (QS) is used by bacteria to coordinateeveral biological processes, including the production of vir-lence factors and growth in biofilm communities. Becausehe decrease in biofilm formation might be due to lowerxpression of QS-related genes, we investigated whether theRNA levels of the QS regulators lasR, rhlR, and mvfR are
ltered after treatment with water:methanol tick egg-waxxtract. However, we detected no significant changes in treatedersus untreated samples (Fig. 3). Interestingly, mRNA lev-ls of pelA and cdrA (also annotated as PA4625 in PAO1 andA14 61200 in PA14 genomes), whose products are impor-ant for the EPS matrix synthesis and scaffolding, respec-ively, were markedly downregulated by water:methanol tick
gg-wax extract treatment (Fig. 3). Likewise, fliC gene expres-ion, the product which is important for flagellum synthesis,as strongly repressed after treatment with tick egg-waxxtract (Fig. 3).
are shown. * Represents a significant difference in relation to untreated samples(Student’s t test, p ≤ 0.05).
K.R. Zimmer et al. / Ticks and Tick-borne Diseases 4 (2013) 366– 376 371
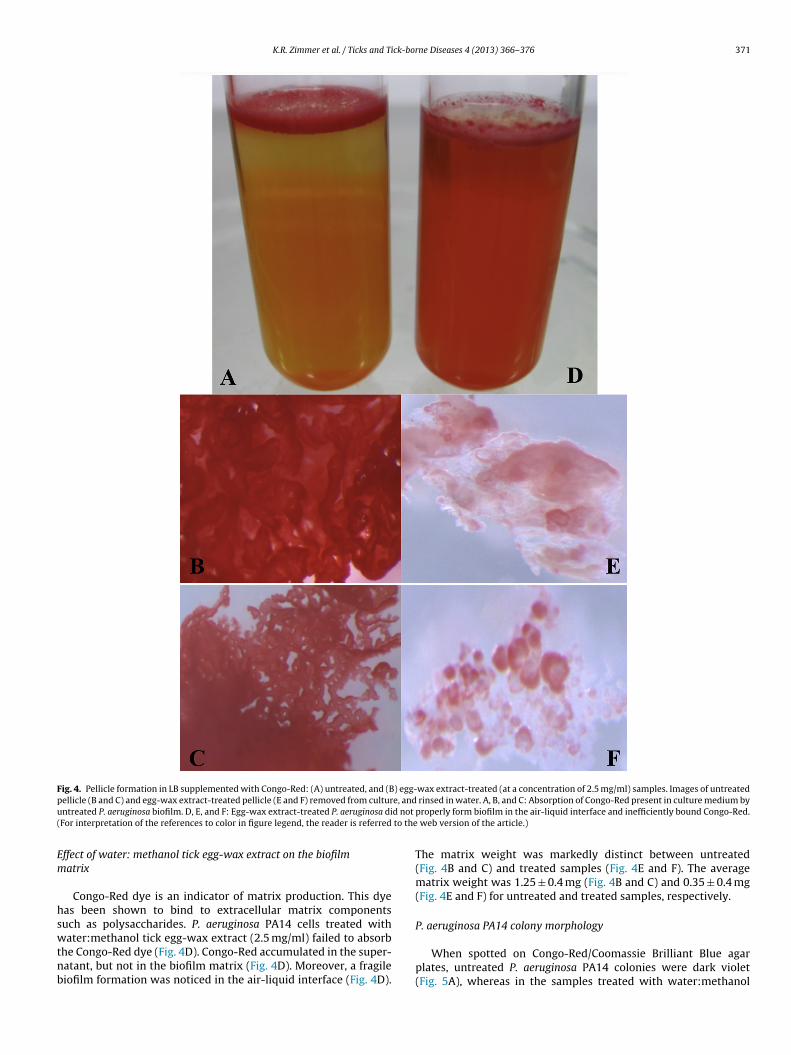
Fig. 4. Pellicle formation in LB supplemented with Congo-Red: (A) untreated, and (B) egg-wax extract-treated (at a concentration of 2.5 mg/ml) samples. Images of untreatedp e, andu d not
( to the
Em
hswtnb
ellicle (B and C) and egg-wax extract-treated pellicle (E and F) removed from culturntreated P. aeruginosa biofilm. D, E, and F: Egg-wax extract-treated P. aeruginosa diFor interpretation of the references to color in figure legend, the reader is referred
ffect of water: methanol tick egg-wax extract on the biofilmatrix
Congo-Red dye is an indicator of matrix production. This dyeas been shown to bind to extracellular matrix componentsuch as polysaccharides. P. aeruginosa PA14 cells treated with
ater:methanol tick egg-wax extract (2.5 mg/ml) failed to absorbhe Congo-Red dye (Fig. 4D). Congo-Red accumulated in the super-atant, but not in the biofilm matrix (Fig. 4D). Moreover, a fragileiofilm formation was noticed in the air-liquid interface (Fig. 4D).
rinsed in water. A, B, and C: Absorption of Congo-Red present in culture medium byproperly form biofilm in the air-liquid interface and inefficiently bound Congo-Red.
web version of the article.)
The matrix weight was markedly distinct between untreated(Fig. 4B and C) and treated samples (Fig. 4E and F). The averagematrix weight was 1.25 ± 0.4 mg (Fig. 4B and C) and 0.35 ± 0.4 mg(Fig. 4E and F) for untreated and treated samples, respectively.
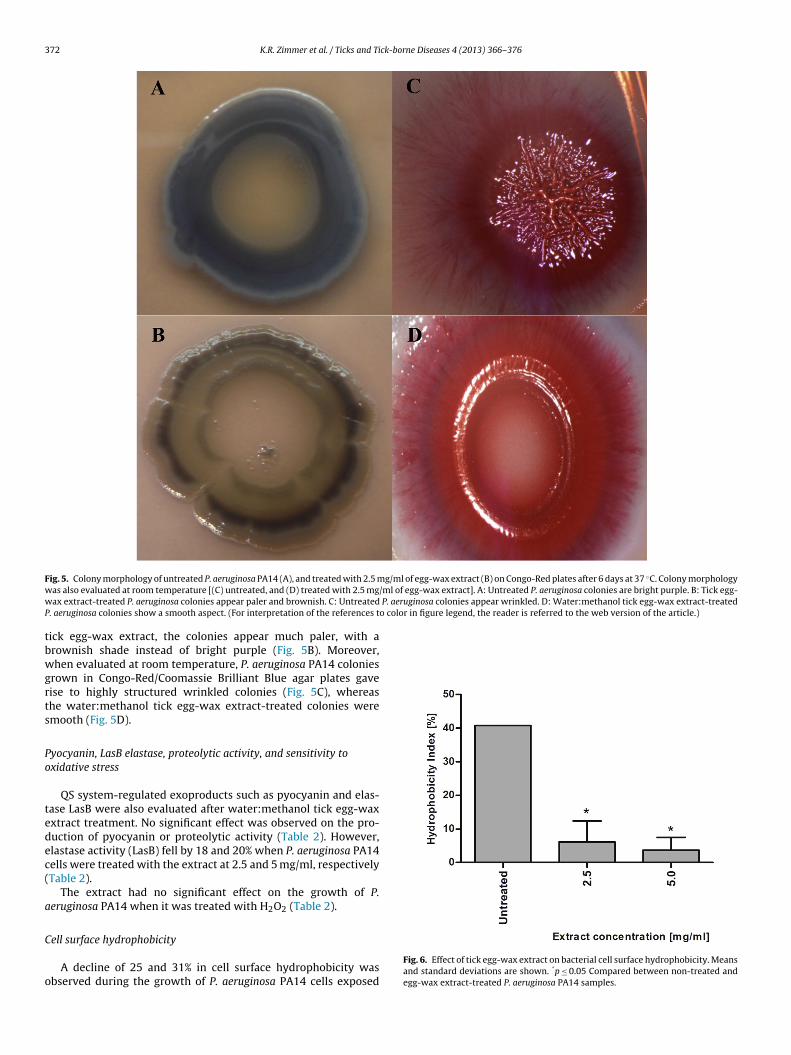
P. aeruginosa PA14 colony morphology
When spotted on Congo-Red/Coomassie Brilliant Blue agarplates, untreated P. aeruginosa PA14 colonies were dark violet(Fig. 5A), whereas in the samples treated with water:methanol
372 K.R. Zimmer et al. / Ticks and Tick-borne Diseases 4 (2013) 366– 376
F g/ml of egg-wax extract (B) on Congo-Red plates after 6 days at 37 ◦C. Colony morphologyw /ml of egg-wax extract]. A: Untreated P. aeruginosa colonies are bright purple. B: Tick egg-w P. aeruginosa colonies appear wrinkled. D: Water:methanol tick egg-wax extract-treatedP o color in figure legend, the reader is referred to the web version of the article.)
tbwgrts
Po
tedec(
a
C
o
ig. 5. Colony morphology of untreated P. aeruginosa PA14 (A), and treated with 2.5 mas also evaluated at room temperature [(C) untreated, and (D) treated with 2.5 mgax extract-treated P. aeruginosa colonies appear paler and brownish. C: Untreated
. aeruginosa colonies show a smooth aspect. (For interpretation of the references t
ick egg-wax extract, the colonies appear much paler, with arownish shade instead of bright purple (Fig. 5B). Moreover,hen evaluated at room temperature, P. aeruginosa PA14 colonies
rown in Congo-Red/Coomassie Brilliant Blue agar plates gaveise to highly structured wrinkled colonies (Fig. 5C), whereashe water:methanol tick egg-wax extract-treated colonies weremooth (Fig. 5D).
yocyanin, LasB elastase, proteolytic activity, and sensitivity toxidative stress
QS system-regulated exoproducts such as pyocyanin and elas-ase LasB were also evaluated after water:methanol tick egg-waxxtract treatment. No significant effect was observed on the pro-uction of pyocyanin or proteolytic activity (Table 2). However,lastase activity (LasB) fell by 18 and 20% when P. aeruginosa PA14ells were treated with the extract at 2.5 and 5 mg/ml, respectivelyTable 2).
The extract had no significant effect on the growth of P.eruginosa PA14 when it was treated with H2O2 (Table 2).
ell surface hydrophobicity
A decline of 25 and 31% in cell surface hydrophobicity wasbserved during the growth of P. aeruginosa PA14 cells exposed
Fig. 6. Effect of tick egg-wax extract on bacterial cell surface hydrophobicity. Meansand standard deviations are shown. *p ≤ 0.05 Compared between non-treated andegg-wax extract-treated P. aeruginosa PA14 samples.

K.R. Zimmer et al. / Ticks and Tick-borne Diseases 4 (2013) 366– 376 373
Table 2Effect of water:methanol tick egg-wax extract on pyocyanin production, elastase activity (LasB), proteolytic activity, and sensitivity of P. aeruginosa PA14 to oxidative stress.The mean values of triplicate independent experiments and SDs are shown.
Egg-wax extract(mg/ml)
Pyocyanin production(Abs at 695 nm)
LasB activity(Abs at 495 nm)
Proteolytic activity(mm casein hydrolysis)
Oxidative stress (mm bacterialgrowth inhibition zone)
0 0.18 ± 0.02 1.52 ± 0.003 27 ± 0.11 23 ± 0.190.156 0.20 ± 0.003 1.70 ± 0.07 22 ± 0.07 24 ± 0.090.3125 0.17 ± 0.007 1.48 ± 0.04 25 ± 0.33 20 ± 0.130.625 0.20 ± 0.004 1.66 ± 0.34 26 ± 0.11 25 ± 0.131.25 0.18 ± 0.002 1.38 ± 0.20 27 ± 0.10 22 ± 0.222.5 0.17 ± 0.003 1.28 ± 0.16* 26 ± 0.05 20 ± 0.155.0 0.19 ± 0.001 1.23 ± 0.06* 25 ± 0.34 23 ± 0.13
*p value ≤ 0.05 compared between untreated P. aeruginosa PA14 (0) and test samples (egg-wax extract-treated PA14).
F 2.5 mc he ino
tr
B
omwstiuiwi
c
Gtetcte
repression of bacterial genes related to the production of biofilmmatrix (pelA and cdrA) and flagellum synthesis (fliC).
ig. 7. Swarming motility of (A) P. aeruginosa PA14 untreated and (B) treated witholonial appearance while bacteria treated with tick egg-wax extract remained in t
o 2.5 and 5.0 mg/ml of the water:methanol tick egg-wax extract,espectively (Fig. 6).
acterial motility (swimming and swarming)
Because bacterial motility is involved in the initial stagesf biofilm formation, we evaluated swimming and swarmingotilities in P. aeruginosa PA14. Bacterial cells treated withater:methanol tick egg-wax extract (2.5 mg/ml) swam at the
ame rate as untreated samples (data not shown). However,he same concentration of water:methanol tick egg-wax extractnhibited swarming motility (Fig. 7). As can be seen in Fig. 7,ntreated P. aeruginosa migrate from the inoculation center, form-
ng branched fractal patterns. In contrast, P. aeruginosa treatedith tick egg-wax extract are not able to migrate from the site of
noculation.
-di-GMP levels
pelA and cdrA genes are upregulated in an elevated c-di-
MP background (Starkey et al., 2009), and we show in Fig. 3hat those genes were downregulated by water:methanol tickgg-wax extract. Although we hypothesized that water:methanolick egg-wax extract might reduce the intracellular levels of-di-GMP, there was no significant difference in c-di-GMPotal levels between control and treatment with egg-waxxtract (Fig. 8).
g/ml egg-wax extract. Swarming in untreated P. aeruginosa is seen as a dendriticculation point.
Discussion
The results presented here support the hypothesis that the eggwax of R. microplus contains material that inhibits P. aeruginosabiofilm formation on the eggs. This anti-biofilm effect involves the
Fig. 8. Quantification of c-di-GMP of untreated and egg-wax extract-treated P.aeruginosa PA14 samples by HPLC tandem mass spectrometry. The mean valuesof triplicate independent experiments and SDs are shown.

3 ck-bor
ibbtitbtiais
ebdCEalatIrda
a(tltiTrt2mwcstns
frePi(mi
ominCtn
tGm
74 K.R. Zimmer et al. / Ticks and Ti
Friedman and Kolter (2004b) showed that pel has a critical rolen biofilm formation, because pel mutants did not form matureiofilms on surfaces such as polystyrene, polyvinylchloride, andorosilicate glass. Our results demonstrate a complete inhibition ofhe biofilm on glass surfaces (Fig. 1C); however, on polystyrene, thenhibition observed was about 40% of that after treatment of bac-erial cells with the tick egg-wax extract (Fig. 1A). In pel mutants,iofilm can be damaged by the washing procedure to remove plank-onic cells (Friedman and Kolter, 2004a, 2004b), as also observedn our study. We noticed that, when treated with egg-wax extract,
biofilm was present on glass surface, although the gentle wash-ng procedure performed completely detached the biofilm from theurface.
The inhibition of P. aeruginosa PA14 biofilm matrix by egg-waxxtract was also verified by colony morphology and the ability ofacterial cells to bind the Congo-Red and Coomassie Brilliant Blueye, both of which are indicators of matrix production (Fig. 5).ongo-Red is used to observe colony morphology because it detectsPS production, and EPS is an essential biofilm component (Uedand Wood, 2009). This dye has been shown to bind to extracellu-ar matrix components in several microorganisms, among whichre Salmonella enterica serovar Typhimurium, Salmonella Enteri-idis, and Escherichia coli (Zogaj et al., 2001; Solano et al., 2002).n our study, the Congo-Red assay clearly showed that bacte-ial cells treated with egg-wax extract were less able to bind theye (Fig. 4D). Moreover, when egg-wax extract was present, P.eruginosa PA14 produced considerably less matrix (Fig. 4E and F).
Matrix production is a crucial step in biofilm developmentnd occurs when cells exhibit a high degree of autoaggregationFriedman and Kolter, 2004a). The ability to autoaggregate leadso various macroscopic phenotypes, including production of pel-icles at the air–liquid interface of standing liquid cultures andhe production of highly structured colonies on agar plates. Pels the major polysaccharide of P. aeruginosa PA14 biofilm matrix.he pel locus contains 7 genes (pelA-G), the products of which areequired for matrix formation and which are therefore critical forhe autoaggregative properties of the strain (Friedman and Kolter,004a; Ryder et al., 2007). pel mutants do not form pellicles orature solid-surface-associated biofilms. In addition, whereas theild-type strain gives rise to wrinkled colonies under some growth
onditions, the P. aeruginosa PA14 pel mutant colonies are flat andmooth (Friedman and Kolter, 2004a, 2004b). Our results (extract-reated bacterial cells form smooth colonies in agar plates and doot develop proper pellicles; Fig. 4) are in accordance with the lattertudies.
Bacterial adhesion is a complex process that is affected by manyactors, including the physicochemical characteristics of bacte-ia as cell surface hydrophobicity (Simões et al., 2010). Egg-waxxtract significantly decreases the cell surface hydrophobicity of. aeruginosa PA14 (Fig. 6). A decrease in bacterial hydrophobicitys generally suggested as a cause of bacterial adhesion inhibitionThenmozhi et al., 2009). This fact, together with the inhibition of
atrix components and synthesis of flagella explain the decreasen bacterial adhesion and thus in biofilm formation.
The Las, Rhl, and MvfR QS systems control the productionf secreted virulence factors, including proteases and secondaryetabolites, and are important for biofilm formation. However,
nhibition of P. aeruginosa biofilm by tick egg-wax extract doesot seem to be directly related to QS system inhibition (Fig. 3).orroborating the hypothesis of non-interference in the QS sys-em, virulence factors (pyocyanin, and proteolytic activity) wereot affected by treatment with egg-wax extract (Table 2).
Because tick egg-wax extract did not affect genes related tohe P. aeruginosa QS system, we evaluated c-di-GMP levels. Inram-negative bacteria, production of c-di-GMP by DGCs is theain trigger for production of extracellular polysaccharides and for
ne Diseases 4 (2013) 366– 376
biofilm formation (Hengge, 2009). Treatment with the tick egg-waxextract does not alter total c-di-GMP level. This finding suggeststhat tick egg-wax extract might be involved in degrading c-di-GMP-binding proteins or competing with them, since c-di-GMP levelswere not altered (Fig. 8) while pelA and cdrA were downregulated.Lory et al. (2009) showed that c-di-GMP controls polysaccharidesynthesis primarily through the action of c-di-GMP-binding pro-teins. During biosynthesis of Pel and alginate, c-di-GMP binds tospecific receptors such as PelD and Alg44. PelD mutants were unableto bind c-di-GMP (Lory et al., 2009).
We also observed that the tick egg-wax extract inhibited fliCgene expression (Fig. 3). Motility enables P. aeruginosa to colonizedifferent environments, to attach to surfaces, and to form biofilms.Also, motility is associated with the pathogenesis of this bacterium(Yeung et al., 2009). Extracellular appendages including flagella andpili probably are the bacterial structures that first establish contactwith the adhesion surface in the initial stages of biofilm forma-tion. It is possible that the flagellum itself may form an adhesivebond with the adhesion surface (Simões et al., 2010). It is assumedthat the primary functions of flagella in biofilm formation involvetransport and initial contacts between bacterial cell and adhesionsurface (Sauer and Camper, 2001). Indeed, P. aeruginosa mutantsdefective in flagella-mediated motility and mutants defective inbiogenesis of the polar-localized type IV pili are unable to start thebiofilm process on abiotic surfaces (O’Toole and Kolter, 1998). Therole of flagella in biofilm formation is not limited to the initial phase(Houry et al., 2010). It has been suggested that flagellum-mediatedmotility is required in the biofilm growth of Listeria monocytogenes(Renier et al., 2011).
Several studies have indicated that motility and the presenceof flagella in uropathogenic Escherichia coli (UPEC) have a role inpathogenesis (Lane et al., 2005; Haugen et al., 2007; Schwan, 2008).Recently, Hidalgo et al. (2011) evaluated the effect of cranberriesand purified cranberry-derived proanthocyanidins (cPACs) display-ing activity against UPEC. In that study, the authors showed thatcranberry compounds inhibit UPEC motility via repression of thefliC gene, suggesting that flagellum-mediated motility might be akey mechanism by which cPACs prevent urinary tract infections.
Flagella are also required for bacterial motility. Swarming,considered as a model of bacterial social behavior, is a flagellum-dependent movement observed in viscous environments such assemisolid agar surfaces (Tremblay and Déziel, 2008). Here, wealso showed that fliC gene expression and swarming motility wereinhibited by tick egg-wax extract.
Borlee et al. (2010) characterized one of the components of thebiofilm matrix in P. aeruginosa, namely CdrA. It is a large non-fimbrial �-helical adhesin that contributes to biofilm structuralintegrity. We demonstrated that tick egg-wax extract repressescdrA gene expression (Fig. 3). The cdrAB locus comprises a two-partner secretion (TPS) system with a large adhesin (CdrA) andits transporter (CdrB). CdrA probably binds directly to the man-nose moiety of Psl (Flemming and Wingender, 2010), and it wassuggested that CdrA is a multivalent adhesin able to recognize mul-tiple carbohydrates (Borlee et al., 2010). cdrA and cdrB mutants aredefective in biofilm initiation (1 and 2 h) as well as final biofilm for-mation (24 h), suggesting that both gene products influence biofilmformation (Ueda and Wood, 2009).
Work attempting to identify the anti-biofilm molecule respon-sible for this activity identified boophiline (N-(3-sulfooxy-25-cholest-5-en-26-oyl)-L-isoleucine) as an anti-biofilm present inegg wax. Since boophiline represses fliC and cdrA and does notrepress pelA, egg wax has at least a second anti-biofilm molecule
(Zimmer et al., 2013).In summary, the cattle tick R. microplus has mechanisms toprotect its eggs against P. aeruginosa PA14 biofilm. We showedthat R. microplus egg-wax extract inhibits 3 critical pathways for

ck-bor
ttsBtd
A
lKsSeeE
R
A
B
B
C
C
E
F
F
F
F
F
F
F
G
G
H
H
H
H
J
K.R. Zimmer et al. / Ticks and Ti
he establishment of P. aeruginosa biofilm: one responsible forhe synthesis of flagella (fliC) and 2 responsible for the synthe-is of structural components of the biofilm matrix (pelA and cdrA).iofilm-inhibiting substances are a matter of great interest becausehey make biofilms more fragile and susceptible to host immuneefenses and antibiotics.
cknowledgments
We are deeply grateful to Dr. Reuben Kaufman for his excel-ent critical review, comments, and suggestions on the manuscript..R.Z. was a recipient of a CAPES PhD studentship. This study wasupported by Fundac ão de Apoio a Pesquisa do Rio Grande doul (FAPERGS), Conselho Nacional de Desenvolvimento Científico
Tecnológico (CNPq), Instituto Nacional de Ciência e Tecnologiam Entomologia Molecular, and Fundac ão de Apoio a Pesquisa dostado de São Paulo (FAPESP).
eferences
rrieta, M.C., Leskiw, B.K., Kaufman, W.R., 2006. Antimicrobial activity in the eggwax of the African cattle tick Amblyomma hebraeum (Acari: Ixodidae). Exp. Appl.Acarol. 39, 297–313.
ooth, T.F., 1992. Observation on the composition and biosynthesis of egg wax lipidsin the cattle tick, Boophilus microplus. Exp. Appl. Acarol. 14, 137–149.
orlee, B.R., Goldamn, A.D., Murakami, K., Samudrala, R., Wozniak, D.J., Parsek, M.R.,2010. Pseudomonas aeruginosa uses a cyclic-di-GMP-regulated adhesin to rein-force the biofilm extracellular matrix. Mol. Microb. 75, 827–842.
osterton, W., Veeh, R., Shirtliff, M., Pasmore, M., Post, C., Erlich, G., 2003. The appli-cation of biofilm science to the study and control of chronic bacterial infections.J. Clin. Invest. 112, 1466–1477.
otter, P.A., Stibitz, S., 2007. c-di-GMP-mediated regulation of virulence and biofilmformation. Curr. Opin. Microbiol. 10, 17–23.
steves, E., Fogac a, A.C., Maldonado, R., Silva, F.D., Manso, P.P., Pelajo-Machado,M., Valle, D., Daffre, S., 2009. Antimicrobial activity in the tick Rhipicephalus(Boophilus) microplus eggs: cellular localization and temporal expression ofmicroplusin during oogenesis and embryogenesis. Dev. Comp. Immunol. 33,913–919.
lemming, H.C., Wingender, J., 2010. The biofilm matrix. Nat. Rev. Microbiol. 8,623–633.
ogac a, A.C., da Silva Jr., P.I., Miranda, M.T., Bianchi, A.G., Miranda, A., Ribolla, P.E.,Daffre, S., 1999. Antimicrobial activity of a bovine hemoglobin fragment in thetick Boophilus microplus. J. Biol. Chem. 274, 25330–25334.
ogac a, A.C., Lorenzini, D.M., Kaku, L.M., Esteves, E., Bulet, P., Daffre, S., 2004.Cysteine-rich antimicrobial peptides of the cattle tick Boophilus microplus: iso-lation, structural characterization and tissue expression profile. Dev. Comp.Immunol. 28, 191–200.
onseca, A.P., Extremina, C., Fonseca, A.F., Sousa, J.C., 2004. Effect of subinhibitoryconcentration of piperacillin/tazobactam on Pseudomonas aeruginosa. J. Med.Microbiol. 53, 903–910.
othergill, J.L., Panagea, S., Hart, C.A., Walshaw, M.J., Pitt, T.L., Winstanley, C., 2007.Widespread pyocyanin over-production among isolates of a cystic fibrosis epi-demic strain. BMC Microbiol. 23, 7–45.
riedman, L., Kolter, R., 2004a. Two genetic loci produce distinct carbohydrate-rich structural components of the Pseudomonas aeruginosa biofilm matrix. J.Bacteriol. 186, 4457–4465.
riedman, L., Kolter, R., 2004b. Genes involved in matrix formation in Pseudomonasaeruginosa PA14 biofilms. Mol. Microb. 51, 675–690.
rubhoffer, L., Golovchenko, M., Vancová, M., Zacharovová-Slavícková, K., Rudenko,N., Oliver Jr., J.H., 2005. Lyme borreliosis: insights into tick-/host-borrelia rela-tions. Folia Parasitol. 52, 279–294.
runclová, L., Fouquier, H., Hypsa, V., Kopácek, P., 2003. Lysozyme from the gut ofthe soft tick Ornithodoros moubata: the sequence, phylogeny and post-feedingregulation. Dev. Comp. Immunol. 27, 651–660.
augen, B.J., Pellett, S., Redford, P., Hamilton, H.L., Roesch, P.L., Welch, R.A., 2007.In vivo gene expression analysis identifies genes required for enhanced col-
onization of the mouse urinary tract by uropathogenic Escherichia coli strainCFT073 dsdA. Infect. Immun. 75, 278–289.
engge, R., 2009. Principles of c-di-GMP signaling in bacteria. Nat. Rev. Microbiol.7, 263–273.
idalgo, G., Chan, M., Tufenkji, N., 2011. Inhibition of Escherichia coli CFT073 fliCexpression and motility by cranberry materials. Appl. Environ. Microbiol. 77,6852–6857.
oury, A., Briandet, R., Aymerich, S., Gohar, M., 2010. Involvement of motility andflagella in Bacillus cereus biofilm formation. Microbiology 156, 1009–1018.
ohns, R., Sonenshine, D.E., Hynes, W.L., 1998. Control of bacterial infections in thehard tick Dermacentor variabilis (Acari: Ixodidae): evidence for the existence ofantimicrobial proteins in tick hemolymph. J. Med. Entomol. 35, 458–464.
ne Diseases 4 (2013) 366– 376 375
Kaszab, E., Szoboszlay, S., Dobolyi, C., Hahn, J., Pék, N., Kriszt, B., 2011. Antibioticresistance profiles and virulence markers of Pseudomonas aeruginosa strainsisolated from composts. Bioresour. Technol. 102, 1543–1548.
Kong, K.F., Jayawardena, S.R., Indulkar, S.D., Del Puerto, A., Kohl, C.L., Høiby, N.,Mathee, K., 2005. Pseudomonas aeruginosa AmpR is a global transcriptionalfactor that regulates expression of AmpC and PoxB beta-lactamases, proteases,quorum sensing, and other virulence factors. Antimicrob. Agents Chemother.49, 4567–4575.
Kopácek, P., Vogt, R., Jindrak, L., Weise, C., Safarik, I., 1999. Purification and charac-terization of the lysozyme from the gut of the soft tick Ornithodoros moubata.Insect Biochem. Mol. Biol. 29, 989–997.
Kopácek, P., Hajdusek, O., Buresová, V., Daffre, S., 2010. Tick innate immunity. Adv.Exp. Med. Biol. 708, 137–162.
Lai, R., Takeuchi, H., Lomas, L.O., Jonczy, J., Rigden, D.J., Rees, H.H., Turner, P.C., 2004.A new type of antimicrobial protein with multiple histidines from the hard tick,Amblyomma hebraeum. FASEB J. 18, 1444–1449.
Lane, M.C., Lockatell, V., Monterosso, G., Lamphier, D., Weinert, J., Hebel, J.R., Johnson,D.E., Mobley, H.L., 2005. Role of motility in the colonization of uropathogenicEscherichia coli in the urinary tract. Infect. Immun. 73, 7644–7656.
Limsuwan, S., Voravuthikunchai, S.P., 2008. Boesenbergia pandurata (Roxb.) Schltr.,Eleutherine americana Merr. and Rhodomyrtus tomentosa (Aiton) Hassk. asantibiofilm producing and antiquorum sensing in Streptococcus pyogenes. FEMSImmunol. Med. Microbiol. 53, 429–436.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25,402–408.
Lory, S., Merighi, M., Hyodo, M., 2009. Multiple activities of c-di-GMP in Pseudomonasaeruginosa. Nucleic Acids Symp. Ser. 53, 51–52.
Malapaka, V.R., Barrese, A.A., Tripp, B.C., Tripp, B.C., 2007. High-throughputscreening for antimicrobial compounds using a 96-well format bacterial motilityabsorbance assay. J. Biomol. Screen. 12, 849–854.
Musk, D.J., Banko, D.A., Hergenrother, P.J., 2005. Iron salts perturb biofilm forma-tion and disrupt existing biofilms of Pseudomonas aeruginosa. Chem. Biol. 12,789–796.
Oldiges, D.P., Parizi, L.F., Zimmer, K.R., Lorenzini, D.M., Seixas, A., Masuda, A., da SilvaVaz Jr., I., Termignoni, C., 2012. A Rhipicephalus (Boophilus) microplus cathepsinwith dual peptidase and antimicrobial activity. Int. J. Parasitol. 42, 635–645.
Olson, M., Howard, C., Morck, D., Buret, A., Read, R., 2002. Biofilm bacteria: formationand comparative susceptibility to antibiotics. Can. J. Vet. Res. 66, 86–92.
O’Toole, G.A., Kolter, R., 1998. Flagellar and twitching motility are necessary forPseudomonas aeruginosa biofilm development. Mol. Microbiol. 30, 295–304.
Pavis, C., Mauleon, H., Barre, N., Maibeche, M., 1994. Dermal gland secretion of trop-ical bont tick, Amblyomma variegatum (Acarina: Ixodidae): biological activity onpredators and pathogens. J. Chem. Ecol. 20, 1495–1503.
Potterat, O., Hostettmann, K., Höltzel, A., Jung, G., Diehl, P.A., Petrini, O., 1997.Boophiline, an antimicrobial sterol amide from the cattle tick Boophilus
microplus. Helv. Chim. Acta 80, 2066–2072.Rahme, L.G., Stevens, E.J., Wolfort, S.F., Shao, J., Tompkins, R.G., Ausubel, F.M., 1995.
Common virulence factors for bacterial pathogenicity in plants and animals.Science 268, 1899–1902.
Renier, S., Hébraud, M., Desvaux, M., 2011. Molecular biology of surface colonizationby Listeria monocytogenes: an additional facet of an opportunistic Gram-positivefoodborne pathogen. Environ. Microbiol. 13, 835–850.
Römling, U., Amikam, D., 2006. Cyclic di-GMP as a second messenger. Curr. Opin.Microbiol. 9, 218–228.
Ryder, C., Byrd, M., Wozniak, D.J., 2007. Role of polysaccharides in Pseudomonasaeruginosa biofilm development. Curr. Opin. Microbiol. 10, 644–648.
Sauer, K., Camper, A.K., 2001. Characterization of phenotypic changes in Pseu-domonas putida in response to surface-associated growth. J. Bacteriol. 183,6579–6589.
Schwan, W.R., 2008. Flagella allow uropathogenic Escherichia coli ascension intomurine kidneys. Int. J. Med. Microbiol. 298, 441–447.
Simm, R., Morr, M., Kader, A., Nimtz, M., Römling, U., 2004. GGDEF and EAL domainsinversely regulate cyclic di-GMP levels and transition from sessility to motility.Mol. Microbiol. 53, 1123–1134.
Simões, M., Simões, L.C., Vieira, M.J., 2010. A review of current and emergent biofilmcontrol strategies. LWT – Food Sci. Technol. 43, 573–583.
Solano, C., Garcia, B., Valle, J., Berasain, C., Ghigo, J.M., Gamazo, C., Lasa, I., 2002.Genetic analysis of Salmonella enteritidis biofilm formation: critical role of cel-lulose. Mol. Microbiol. 43, 793–808.
Spangler, C., Böhm, A., Jenal, U., Seifert, R., Kaever, V., 2010. A liquidchromatography-coupled tandem mass spectrometry method for quantitationof cyclic di-guanosine monophosphate. J. Microbiol. Methods 81, 226–231.
Starkey, M., Hickman, J.H., Zhang, N., De Long, S., Hinz, A., Palacios, S., Manoil, C.,Kirisits, J., Starner, T.D., Wozniak, D.J., Harwood, C.S., Parsek, M.R., 2009. Pseu-domonas aeruginosa rugose small-colony variants have adaptations that likelypromote persistence in the cystic fibrosis lung. J. Bacteriol. 191, 3492–3503.
Thenmozhi, R., Nithyanand, P., Rathna, J., Pandian, S.K., 2009. Antibiofilm activity ofcoral-associated bacteria against different clinical M serotypes of Streptococcuspyogenes. FEMS Immunol. Med. Microbiol. 57, 284–294.
Tremblay, J., Déziel, E., 2008. Improving the reproducibility of Pseudomonas
aeruginosa swarming motility assays. J. Basic Microbiol. 48, 509–515.Trentin, D.S., Giordani, R.B., Zimmer, K.R., Da Silva, A.G., Da Silva, M.V., Correia, M.T.,Baumvol, I.J., Macedo, A.J., 2011. Potential of medicinal plants from the Braziliansemi-arid region (Caatinga) against Staphylococcus epidermidis planktonic andbiofilm lifestyles. J. Ethnopharmacol. 137, 327–335.

3 ck-bor
T
U
W
Y
Y
76 K.R. Zimmer et al. / Ticks and Ti
ron, E.A., Wilke, H.L., Petermann, S.R., Rust, L., 2004. Pseudomonas aeruginosa fromcanine otitis externa exhibit a quorum sensing deficiency. Vet. Microbiol. 99,121–129.
eda, A., Wood, T.K., 2009. Connecting quorum sensing, c-di-GMP, Pel polysac-charide, and biofilm formation in Pseudomonas aeruginosa through tyrosinephosphatase TpbA (PA3885). PLoS. Pathog. 5, 1–15.
ang, E.W., Agostini, G., Olomu, O., Runco, D., Jung, J.Y., Chole, R.A., 2008. Gentianviolet and ferric ammonium citrate disrupt Pseudomonas aeruginosa biofilms.Laryngoscope 118, 2050–2056.
eung, A.T., Torfs, E.C., Jamshidi, F., Bains, M., Wiegand, I., Hancock, R.E., Overhage, J.,2009. Swarming of Pseudomonas aeruginosa is controlled by a broad spectrumof transcriptional regulators, including MetR. J. Bacteriol. 191, 5592–5602.
u, Z., Thomson, E.L.S., Liu, J., Dennis, J.J., Jacobs, R.L., Kaufman, W.R., 2012.Antimicrobial activity in the egg of the Amblyomma hebraeum (Acari: Ixodidae)
ne Diseases 4 (2013) 366– 376
is associated with free fatty acids C16:1 and C18:2. Exp. Appl. Acarol. 58,453–470.
Zhu, H., Bandara, R., Conibear, T.C., Thuruthyil, S.J., Rice, S.A., Kjelleberg, S., Givskov,M., Willcox, M.D., 2004. Pseudomonas aeruginosa with lasI quorum-sensing defi-ciency during corneal infection. Invest. Ophthalmol. Vis. Sci. 45, 1897–1903.
Zimmer, K.R., Macedo, A.J., Giordani, R.B., Conceic ão, J.M., Nicastro, G.G., Boechat, A.L.,Baldini, R.L., Abraham, W.R., Termignoni, C., 2013. A steroidal molecule presentin the egg wax of the tick Rhipicephalus (Boophilus) microplus inhibits bacte-rial biofilms. Environ Microbiol, doi:10.1111/1462-2920.12082 [E-pub ahead of
print].Zogaj, X., Nimtz, M., Rohde, M., Bokranz, W., Römling, U., 2001. The multicellularmorphotypes of Salmonella typhimurium and Escherichia coli produce cellu-lose as the second component of the extracellular matrix. Mol. Microbiol. 39,1452–1463.




















