
Central nervous system of Rhipicephalus sanguineus ticks (Acari: Ixodidae): an ultrastructural study
9
ORIGINAL PAPER Central nervous system of Rhipicephalus sanguineus ticks (Acari: Ixodidae): an ultrastructural study Gislaine Cristina Roma & Pablo Henrique Nunes & Patrícia Rosa de Oliveira & Rafael Neodini Remédio & Gervásio Henrique Bechara & Maria Izabel Camargo-Mathias Received: 12 March 2012 / Accepted: 3 May 2012 # Springer-Verlag 2012 Abstract This study performed the ultrastructural descrip- tion of the synganglion of Rhipicephalus sanguineus males and females, aiming to contribute to the understanding of the cellular organization of this organ. The results show that the central nervous system of these individuals consists of a mass of fused nerves, named synganglion, from where nerves emerge towards several parts of the body. It is sur- rounded by the neural lamella, a uniform and acellular layer, constituted by repeated layers of homogeneous and finely granular material. The perineurium is just below, composed of glial cells, which extensions invaginate throughout the nervous tissue. The synganglion is internally divided into an outer cortex, which contains the cellular bodies of the neural cells and an inner neuropile. The neural cells can be classified into two types according to cell size, cytoplasm–nucleus rela- tion, and neurosecretory activity. Type I cells are oval or spherical and present a large nucleus occupying most part of the cytoplasm, which contains few organelles. Type 2 cells are polygonal, present a great cytoplasm volume, and their nuclei are located in the cell periphery. The cytoplasm of these cells contains a well-developed rough endoplasmic reticulum, Golgi regions, mitochondria, and several neurosecretory gran- ules. The subperineurium and the tracheal ramifications are found between the cortex and the neuropile. The latter is formed mainly by neural fibers, tracheal elements, and glial cells. The results obtained show that R. sanguineus males' and females' nervous tissue present an ultrastructural organization similar to the one described in the literature for other tick species. Introduction The ultrastructure of the central nervous system of ticks was already described to Argasidae (Coons et al. 1974; El Shoura 1986) and Ixodidae (Binnington and Obenchain 1982; Ivanov 1983). According to these authors, this system consists of a mass of fused neural cells named synganglion, a set of peripheral nerves, as well as neuroendocrine organs. This structure represents a remarkable evolutive phenome- non, once this organ does not present internal segmentation due to the high level of condensation of the ganglia which form it (Roshdy and Marzouk 1984), which would also occur externally, once these ectoparasites do not present body divisions having fused head, thorax, and abdomen (Storer and Usinger 2000). The tick nervous system is characterized as a vital organ for the biological success of this group as it presents a great diversity of functions, such as control of all the metabolic processes which occur in the organism of these ectopara- sites. Thus, morphology, cellular structure, and functional organization of this tissue require intensive investigation to provide information that can help in the understanding of G. C. Roma (*) : P. R. de Oliveira : R. N. Remédio : M. I. Camargo-Mathias Departamento de Biologia, Instituto de Biociências, Universidade Estadual Paulista “Júlio de Mesquita Filho”, UNESP, Avenida 24 A, 1515, 13506-900 Rio Claro, SP, Brazil e-mail: [email protected] P. H. Nunes Departamento de Medicina Veterinária Preventiva e Saúde Animal, Faculdade de Medicina Veterinária e Zootecnia, USP, Avenida Prof. Orlando Marques de Paiva, 87, 05508-270 São Paulo, SP, Brazil G. H. Bechara Departamento de Patologia Veterinária, Faculdade de Ciências Agrárias e Veterinárias, UNESP, Via de acesso Prof. Paulo Castellane, s/n, 14884-900 Jaboticabal, SP, Brazil Parasitol Res DOI 10.1007/s00436-012-2962-2
Transcript of Central nervous system of Rhipicephalus sanguineus ticks (Acari: Ixodidae): an ultrastructural study

ORIGINAL PAPER
Central nervous system of Rhipicephalus sanguineus ticks(Acari: Ixodidae): an ultrastructural study
Gislaine Cristina Roma & Pablo Henrique Nunes &
Patrícia Rosa de Oliveira & Rafael Neodini Remédio &
Gervásio Henrique Bechara &
Maria Izabel Camargo-Mathias
Received: 12 March 2012 /Accepted: 3 May 2012# Springer-Verlag 2012
Abstract This study performed the ultrastructural descrip-tion of the synganglion of Rhipicephalus sanguineus malesand females, aiming to contribute to the understanding ofthe cellular organization of this organ. The results show thatthe central nervous system of these individuals consists of amass of fused nerves, named synganglion, from wherenerves emerge towards several parts of the body. It is sur-rounded by the neural lamella, a uniform and acellular layer,constituted by repeated layers of homogeneous and finelygranular material. The perineurium is just below, composedof glial cells, which extensions invaginate throughout thenervous tissue. The synganglion is internally divided into anouter cortex, which contains the cellular bodies of the neuralcells and an inner neuropile. The neural cells can be classifiedinto two types according to cell size, cytoplasm–nucleus rela-tion, and neurosecretory activity. Type I cells are oval orspherical and present a large nucleus occupying most part ofthe cytoplasm, which contains few organelles. Type 2 cells are
polygonal, present a great cytoplasm volume, and their nucleiare located in the cell periphery. The cytoplasm of these cellscontains a well-developed rough endoplasmic reticulum,Golgi regions, mitochondria, and several neurosecretory gran-ules. The subperineurium and the tracheal ramifications arefound between the cortex and the neuropile. The latter isformed mainly by neural fibers, tracheal elements, and glialcells. The results obtained show that R. sanguineusmales' andfemales' nervous tissue present an ultrastructural organizationsimilar to the one described in the literature for other tickspecies.
Introduction
The ultrastructure of the central nervous system of ticks wasalready described to Argasidae (Coons et al. 1974; ElShoura 1986) and Ixodidae (Binnington and Obenchain1982; Ivanov 1983). According to these authors, this systemconsists of a mass of fused neural cells named synganglion,a set of peripheral nerves, as well as neuroendocrine organs.This structure represents a remarkable evolutive phenome-non, once this organ does not present internal segmentationdue to the high level of condensation of the ganglia whichform it (Roshdy and Marzouk 1984), which would alsooccur externally, once these ectoparasites do not presentbody divisions having fused head, thorax, and abdomen(Storer and Usinger 2000).
The tick nervous system is characterized as a vital organfor the biological success of this group as it presents a greatdiversity of functions, such as control of all the metabolicprocesses which occur in the organism of these ectopara-sites. Thus, morphology, cellular structure, and functionalorganization of this tissue require intensive investigation toprovide information that can help in the understanding of
G. C. Roma (*) : P. R. de Oliveira : R. N. Remédio :M. I. Camargo-MathiasDepartamento de Biologia, Instituto de Biociências, UniversidadeEstadual Paulista “Júlio de Mesquita Filho”, UNESP,Avenida 24 A, 1515,13506-900 Rio Claro, SP, Brazile-mail: [email protected]
P. H. NunesDepartamento de Medicina Veterinária Preventiva e SaúdeAnimal, Faculdade de Medicina Veterinária e Zootecnia, USP,Avenida Prof. Orlando Marques de Paiva, 87,05508-270 São Paulo, SP, Brazil
G. H. BecharaDepartamento de Patologia Veterinária, Faculdade de CiênciasAgrárias e Veterinárias, UNESP,Via de acesso Prof. Paulo Castellane, s/n,14884-900 Jaboticabal, SP, Brazil
Parasitol ResDOI 10.1007/s00436-012-2962-2

the general physiology of Acari, an important group ofanimals of great medical and veterinary importance (Roshdyand Marzouk 1984).
Ticks are characterized as an important group of patho-gen vectors in the Arthropoda phylum, being only compa-rable to mosquitoes (Culicidae) (Marcondes 2009). They areresponsible for the transmission of pathogens which affectdomestic animals and human beings, including bacteria,helminths, protozoa, and viruses (Jongejan and Uilenberg2004).
Rhipicephalus sanguineus tick (Latreille 1806), popular-ly known as the brown dog tick, is cosmopolitan of tropicaland temperate regions (Walker 1994; Paz et al. 2008) pre-senting a wide geographic distribution in the Americas,Europe, Africa, Asia, and Australia (Ribeiro et al. 1996).Over the last years, this species has drawn the attention ofpublic health organizations for having become an urbanplague (Paz et al. 2008), fact confirmed by the great expan-sion of the veterinary pharmaceutical industry, concernedabout creating new formulations and acaricide substances.Until the early 80s, the manufacturing of commercial acar-icides with specific action for the treatment of R. sanguineusin dogs was scarce. Currently, there are tens of products inthe market, with different formulations and applications,representing significant profit for the pharmaceutical indus-tries in the veterinary line (Labruna 2004).
Several studies have been carried out on ticks aiming tofind out an efficient method to control these ectoparasites.However, most of these studies refer to tests on the efficacyof synthetic or natural methods (Taylor 2001; Mencke et al.2003; Farias et al. 2007, 2009).
In this sense, this study aimed to describe, through trans-mission electron microscopy (TEM), the ultrastructure ofthe central nervous system (synganglion) of R. sanguineusmales and females. As this system is characterized as thetarget of several acaricides with neurotoxic action, this studywill certainly serve as a basis for future research which willneed this knowledge to better understand the cellular orga-nization of this organ as well as its physiology in order toimprove or find new methods of tick control.
Material and methods
R. sanguineus ticks
Unfed adults (males and females) of R. sanguineus weresupplied by the tick colonymaintained at the Brazilian Centralof Studies on Ticks Morphology (BCSTM) at the BiosciencesInstitute of São Paulo State University, Rio Claro, SP, Brazil.The ticks were kept under controlled conditions (28±1°C,80 % relative humidity, and 12 h photoperiod) in a BiologicalOxygen Demand incubator and blood fed on New Zealand
White rabbits. Details on feeding and maintenance of R.sanguineus ticks on the hosts are given by Bechara et al.(1995). This study was approved by the Ethics Committee inthe Animal Use, CEUA, UNESP, Rio Claro, SP, Brazil, Pro-tocol no. 4093.
Transmission electron microscopy (TEM)
R. sanguineus males and females were dissected on Petridishes containing phosphate-buffered saline (PBS) solution(NaCl 0.13 M, Na2HPO4 0.017 M, KH2PO4 0.02 M, pH7.2). Synganglia were removed, fixed in 2.5 % glutaralde-hyde fixative solution in 0.1 M sodium cacodylate buffer(pH 7.2) for 2 h. Two 15-min washes in cacodylate bufferfollowed this process. Postfixation was performed in 1 %osmium tetroxide in 0.1 M in the same buffer (pH 7.2) for2 h in darkness, followed by another two 15-min washes inthe same buffer. To increase the contrast, material wasimmersed in a solution of 2 % uranyl acetate in 10 %acetone, for 4 h in darkness. Dehydration was performedin a graded acetone series (50, 70, 90, 95, and 100 %) for5 min each. Then, the material was embedded in Epon resindiluted in acetone (1:1) for 12 h, following pure Epon resin,and incubated at 60°C for 24 h. Semithin sections wereobtained with Sorvall-Porter Blum MT2-B ultramicrotomeand stained with azur II (1 %) and methylene blue (1 %).
Grids containing ultrathin sections were contrasted withuranyl acetate and lead citrate (Reynolds 1963) for 45 minand 10 min, respectively. Then, they were analyzed andphotographed in a Philips CM 100 TEM from the BiologyDepartment at the Biosciences Institute, UNESP, campus ofRio Claro, SP, Brazil.
Results
The central nervous system of R. sanguineus males andfemales consists of a mass of fused nerves, named syngan-glion, from where nerves emerge towards several parts ofthe body. The esophagus runs through the synganglion(Figs. 1a and 3l) and divides it into two regions: (1) supra-esophageal, the smaller of the two regions, consisting of aprotocerebrum, a single dorsal ganglion located anterodor-sally to the esophagus and (2) subesophageal, which con-sists in most part of the synganglion, being located in theventral and posterior regions of the esophagus.
The nervous tissue of these ectoparasites consists of anouter cortex and an inner neuropile, both surrounded by aneural lamella or neurilemma, a uniform and acellular layer.This structure is enclosed by a cavity named periganglionicsinus, which continues to the aorta and heart of the ticks(Figs. 1a–b, 2a–c, and 3a).
Parasitol Res

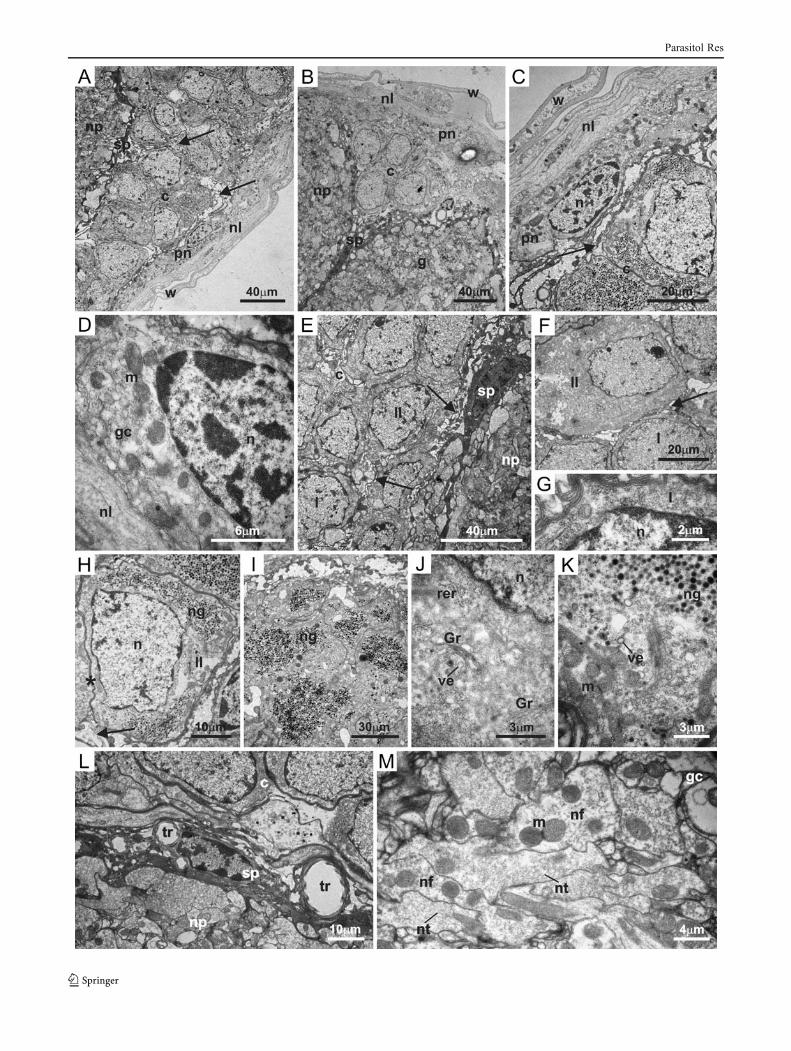
No differences between the ultrastructure of R. sangui-neus males' and females' synganglia were found in thisstudy. Thus, this organ will be described in the same wayfor the ticks here analyzed. Ultrastructurally, the neurallamella consists of repeated layers of homogeneous andfinely granular material which encloses the synganglion aswell as the esophageal channel and the peripheral nerveswhich emerge from this organ (Figs. 1a–b, 2a–c, and 3a–c).The periganglionic membrane or perineurium is just below,formed by a series of glial cells, which extensions invagi-nate throughout the nervous system (Figs. 1a–b, 2a–d, and3a–c). These cells are characterized by their elongated nu-cleus as well as by their irregular membrane (Figs. 1b, 2c–d,
and 3b–c). Several mitochondria are found in the cytoplasm(Figs. 1b, 2d and 3c).
The synganglion is internally covered by an outer corticalregion named cortex which contains neural cell bodies orneurons (Figs. 1a–b; 2a–c, e; and 3a). Two cell types can bedistinguished in the cortex of R. sanguineus males andfemales according to the size of the cell, cytoplasm–nucleusrelation, and neurosecretory activity (Figs. 1b; 2e—f; and3a, d). Type I cells are most frequently found in all theextensions of the cortex. These cells are spherical or ovaland present a large nucleus compared to the cytoplasm,which contains few organelles, such as mitochondria(Figs. 1b; 2e–g; and 3a, d–e). The nuclei of type I cells
Fig. 1 Schematic drawing ofthe central nervous system(synganglion) of R. sanguineusticks. a General view and bdetail of synganglion of malesand females. nl neural lamella,p perineurium, c cortex, spsubperineurium, np neuropile, gganglion, es esophagus, gc glialcells, e extensions of glial cells,n nucleus, m mitochondria, Itype I neural cells, II type IIneural cells, rer roughendoplasmic reticulum, GrGolgi region, ng neurosecretorygranules, tr tracheal elements,nf neural fibers, nt neurotubules
Parasitol Res
Parasitol Res

present condensed chromatin accumulation adjacent to thenuclear envelope (Figs. 1b and 3e).
Neural cells type II are polygonal and are distributedamong type I cells. They present a great volume of cyto-plasm in comparison with the other cell type as well asnucleus located in the periphery of the cell and with dispersechromatin (Figs. 1b; 2e–f, h; and 3a, d, and f). The cyto-plasm of these cells contains a greater number of organellesin comparison with the type I cell, with well-developedrough endoplasmic reticulum, Golgi regions, vesicles,mitochondria, and several neurosecretory granules (Figs. 1b;2 h–k; and 3f–h).
The cortex cells can be in direct contact with each otherthrough the junction of the cellular membranes and/or sep-arated by the extensions of the perineurium glial cells(Figs. 1b; 2a, e–f, h; and 3d, f). The subperineurium is foundbetween the cortex and the neuropile and presents the sameultrastructural characteristics of the perineurium; however,branches of the tracheas are found here (Figs. 1a–b; 2a–b, e,l; and 3a, i).
The neuropile, inner structure of the synganglion, isconstituted by ramifications of the neural cells, also calledneural fibers, by the tracheal elements which penetrate theneuropile through the subperineurium and glial cells(Figs. 1a–b; 2a–b, e, l–m; and 3a, i, j–k). The neural fibersare the most common elements found in the neuropile.These structures are mainly formed by mitochondria, neuro-filaments, and neurotubules surrounded by the extensions ofthe glial cells (Figs. 1b; 2 l–m; and 3j–k).
Discussion
Over the last five decades, acaricides with differentactive ingredients have been used; however, differentmechanisms of resistance have already been developedby the ticks as strategies to survive. The selection ofresistant tick strains mainly occurs due to the incorrectuse of acaricides (Häuserman et al. 1992). This is arelevant fact, once it is an irreversible process. For thisreason and for the problems caused by the contamina-tion of nontarget organisms and the environment, thesearch for less toxic and environment-impacting acari-cide products has been intensified. However, to makethis happen, morphophysiological studies on the differ-ent tick organs are necessary to better study the biologyand physiology of these specimens; once, without thisknowledge, it is not possible to develop viable alterna-tives to control ticks. In this sense, this study performsthe ultrastructural description of the central nervoussystem (synganglion) of R. sanguineus males andfemales, since this organ is the target of neurotoxicacaricides.
As described for ticks in general (Obenchain and Oliver1976; Roshdy and Marzouk 1984; Binnington 1986; ElShoura 1989; Sonenshine 1991; Oliver et al. 1992; Prullageet al. 1992; Lees and Bowman 2007), the central nervoussystem of R. sanguineus males and females consists in amass of fused nerves named synganglion, from wherenerves emerge towards different parts of the body.
The synganglion of R. sanguineus is found inside acavity named periganglionic sinus, which is continuous tothe aorta and heart. This location allows the synganglion tosecrete and/or continuously absorb several essential substan-ces from the hemolymph in order to maintain metabolism.This data corroborates Coons and Alberti (1999) on theirstudies about the internal morphology of ticks. In this study,the ultrastructural analysis did not show differences betweenthe central nervous systems of R. sanguineus males andfemales, confirming preliminary light microscopy data byRoma et al. (2012).
According to Coons et al. (1974), Ivanov (1983), ElShoura (1986), Sonenshine (1991), and Coons and Alberti(1999), the synganglion of Argasidae and Ixodidae tickswould be surrounded by the neural lamella, constituted byseveral layers of finely granular homogeneous material. Theresults obtained here corroborate these authors' findings,suggesting that this structure would have the function tosupport the tissues located in its interior, in addition toresisting to the hydrostatic pressure and acts as a selectivepermeability barrier for several nutrients and ions from thehemolymph.
The perineurium or periganglionic membrane is just be-low the neural lamella, and it is formed by glial cells, which
�Fig. 2 Electron micrograph of synganglion of males of Rhipicephalussanguineus ticks. a–b General view of the synganglion showing wallof the blood sinus (w), neural lamella (nl), perineurium (pn), extensionsof glial cells (arrow) into the cortex (c), subperineurium (sp), neuropile(np), and ganglion (g). c General view and d detail of perineurium (pn)formed by glial cells (gc) exhibiting mitochondria (m) and elongatednucleus (n), besides extensions of glial cells (arrow). e General view ofcortex (c) showing the extensions of glial cells (arrow) formingextracellular spaces between glial cells and nervous cells of thecortex. Observe types I (I) and II (II) neural cells. f General viewof types I (I) and II (II) neural cells, showing these cells areseparated by extensions of glial cells (arrow). g Detail of type I(I) neural cell; observe nucleus (n) and few organelles. h Type II(II) neural cell exhibiting nucleus (n) and neurosecretory granules(ng); observe the direct contact between cell membranes of adja-cent neural cells (asterisk) or separated by extensions of glialcells (arrow). i Detail of great amount of neurosecretory granules(ng) in the cytoplasm of type II neural cells. j–k Detail of thecytoplasm of type II neural cells showing rough endoplasmicreticulum (rer), Golgi region (Gr), vesicles (ve), mitochondria(m), nucleus (n), and neurosecretory granules (ng). l Detail ofsubperineurium (sp) between cortex (c) and neuropile (np); ob-serve tracheal elements (tr). m Detail of neuropile exhibiting theneural fibers (nf), neurotubules (nt), mitochondria (m), and glialcells (gc)
Parasitol Res

Parasitol Res

extensions penetrate throughout the nervous tissue. Theseinvaginations form extracellular spaces among the cortexcells. These results corroborate the findings of Coons et al.(1974) and Binnington and Obenchain (1982) for Argasarboreus and Boophilus microplus, respectively. Accordingto these authors, these extracellular spaces would functionas a reservoir of cations which would be used during theregulation of the ionic concentration of the cortex cells,which would certainly be occurring here, once several mi-tochondria were found in the glial cells of the perineuriumof R. sanguineus males and females. These organelleswould probably be involved in the production of the neces-sary energy for this process to occur (through the synthesisof ATP). However, these spaces were not observed in thenervous tissue of Amblyomma americanum larvae (ElShoura 1989).
In this study, it is also suggested that the extensionsof the glial cells would increase the contact area be-tween these cells and those found in the cortex, allow-ing the exchange of substances between them.According to Sonenshine (1991), the glial cells wouldplay a fundamental role in the metabolic regulations ofthe neural activities. Thus, it is suggested that thesecells in R. sanguineus males and females would beinvolved in the transportation of elements from thehemolymph (absorbed via neural lamella, according toMarzouk et al. 2001) to the cortex cells, a functionwhich is essential for the maintenance of the physiolog-ical processes which are inherent to this organ. Thus,the elements synthetized by the cortex neural cellswould be secreted in the periganglionic sinus throughthe transportation to the perineurium and posteriorly tothe neural lamella, which due to its selective permeability
(Marzouk et al. 2001) would eliminate the neurosecretiondirectly in the hemolymph, controlling the general me-tabolism of all the organs and tissues of the ticks.
Coons and Alberti (1999) reported that deposits of gly-cogen would be found in the glial cell cytoplasm ofengorged tick females and used during the oviposition ofthese ectoparasites. However, this element was not found inthe glial cells of R. sanguineus females, probably due to thefact that these females were fasting, not having started theengorgement phase.
The synganglion of R. sanguineus males and females,as of ticks in general, is covered by an outer regionnamed cortex which contains neural cell bodies, calledneurons by some authors (Sonenshine 1991; Coons andAlberti 1999). Two types of cells can be identified inthis region, corroborating the studies of Coons et al.(1974), Ivanov (1983), and El Shoura (1986) for otherspecies of ticks.
Type I neural cells are oval or spherical and presenta large nucleus occupying most part of the cytoplasm.According to these data, these cells would be similar tothe motor neurons described by El Shoura (1989) andCoons and Alberti (1999) for other species of ticks andwould be involved in the transmission of impulses fromthe central nervous system to the target organs. In thecytoplasm of these cells, both in R. sanguineus malesand females, few organelles were observed, such asmitochondria, suggesting that type I neural cells wouldnot be related to the synthesis of neurosecretory ele-ments, and the organelles would be involved in themaintenance of the cell's metabolism. The hypothesisthat type I cells would not be neurosecretory are con-firmed in the results by Coons et al. (1974) and Ivanov(1983) for other species of ticks. However, El Shoura(1986) reported that Ornithodoros erraticus females andA. americanum larvae would play an important role in thesynthesis of neurosecretory granules. Moreover, accordingto Sonenshine (1991), some authors believe that theneurosecretory activity capacity of the neural cells wouldbe a phase of the cellular cycle which is not limitedto a certain type of cell.
According to Coons and Alberti (1999), type II neuralcells would be distributed among type I cells, formingsmall groups, the neurosecretory centers. They are charac-terized by a great volume of cytoplasm and nucleuslocated in the periphery of the cell. These data corrob-orate the ones found for the synganglion of R. sangui-neus males and females. A greater number of organelleswere observed in the cytoplasm of these cells in com-parison with type I cell, such as, well-developed roughendoplasmic reticulum and Golgi regions, vesicles, mi-tochondria, and several deposits of neurosecretory gran-ules. These results suggest that type II cells would be
�Fig. 3 Electron micrograph of synganglion of females of Rhipicephalussanguineus ticks. a–b General view of the synganglion showing neurallamella (nl), perineurium (pn), cortex (c) with types I (I) and II (II) neuralcells, subperineurium (sp), and neuropile (np). Observe in b the exten-sions (arrow) of glial cells (gc) of perineurium into the cortex, besides thegreat amount of neurosecretory granules (ng) in type II (II) neural cells. cDetail of neural lamella (nl) and perineurium (pn), showing the glial cell(gc) with many mitochondria (m), elongated nucleus (n), and extensions(arrow). d General view of types I (I) and II (II) neural cells, showingthese cells are separated by extensions of glial cells (arrow). e Detail oftype I (I) neural cells exhibiting nucleus (n) and few organelles, such asmitochondria (m). f Type II (II) neural cell exhibiting nucleus (n), neuro-secretory granules (ng), mitochondria (m), Golgi region (Gr), and roughendoplasmic reticulum (rer). Observe the direct contact between cellmembranes of adjacent neural cells (asterisk) or separated by extensionsof glial cells (arrow). g–hDetail of the cytoplasm of the type II (II) neuralcell showing neurosecretory granules (ng), mitochondria (m), Golgiregion (Gr), vesicles (ve), besides well-developed rough endoplasmicreticulum (rer). i Detail of subperineurium (sp) between cortex (c) andneuropile (np); observe tracheal elements (tr). jGeneral view and k detailof neuropile exhibiting the neural fibers (nf), neurotubules (nt), andmitochondria (m); observe the glial cells (gc) among the neural fibers.l Esophagus (es) into the neuropile (np)
Parasitol Res

secretory, corroborating descriptions by Ivanov (1983)and El Shoura (1986) while studying the synganglionof Argasidae and Ixodidae.
El Shoura (1989) described a third type of neural cells(type III) in A. americanum larvae, which would have alarge nucleus and narrow cytoplasm with few organelles;however, the function of these cells would not be knownyet. It should be stressed that this type of cell was notidentified in this study.
Considering the results here obtained, it can be suggestedthat the communication between the neural cells of the cortex(types I and II) would happen through the direct contactbetween cells, once in some regions of the membrane ofadjacent cells would be fused. These membranes could beacting as a barrier of selective permeability, controlling thetransportations of material in both directions (intra- and extra-cellular). On the other hand, the union of the membranes ofadjacent cells could also be related with the provision ofgreater cohesion to this tissue. However, in some regions,the neural cells are separated by the extensions of the perineu-rium glial cells, which, as described above, would control allthe metabolism of the tick central nervous system.
Between the cortex and the neuropile of R. sanguineussynganglion, as described for ticks in general (Coons andAlberti 1999), is the subperineurium, which presents thesame ultrastructural characteristics of the perineurium.However, branches of tracheas were found in this region,which would be providing the necessary oxygen supply tothis tissue of high energetic consumption, considering thefunctions performed.
The neuropile is the most complex region of thecentral nervous system of the ticks (El Shoura 1989).In R. sanguineus males and females, as well as in otherticks’ species (Sonenshine 1991; Coons and Alberti1999), it is formed by the ramifications of types I andII nervous cells by the tracheal elements and glial cells.Most neural fibers are surrounded by the extensions ofglial cells, which can come from two sources: (1) thecells located in the neuropile itself or (2) the extensionsof the glial cells of the subperineurium.
Thus, considering data here described, it can be concludedthat the central nervous system of R. sanguineus males andfemales presents an ultrastructural organization similar to theone described in literature for other species of ticks.Moreover,the importance of this kind of study for the improvement anddiscovery of new efficient methods to control ticks withoutharming hosts and the environment is emphasized here.
Acknowledgments The authors are grateful to Fundação de Amparoà Pesquisa do Estado de São Paulo—FAPESP (grant nos. 2010/51942-8, 2011/06865-8, and 2011/10427-6) for financial support.
Conflict of interest The authors declare that there are no conflicts ofinterest.
References
Bechara GH, Szabó MPJ, Ferreira BR, Garcia MV (1995) Rhipicepha-lus sanguineus tick in Brazil: feeding and reproductive aspectsunder laboratorial conditions. Braz J Vet Parasitol 4:61–66
Binnington KC (1986) Ultrastructure of the tick neuroendocrine sys-tem. In: Sauer JR, Hair JA (eds) Morphology, physiology andbehavioural biology of ticks. Ellis Horwood, Chichester, pp 152–164
Binnington KC, Obenchain FD (1982) Structure and function of thecirculatory, nervous, and neuroendocrine system of ticks. In:Obenchain FD, Galun R (eds) Physiology of ticks. Pergamon,Oxford, pp 351–398
Coons LB, Alberti G (1999) The acari-ticks. In: Harrison FW, Foelix R(eds) Microscopic anatomy of invertebrates. Chelicerate Arthro-poda, Wiley, pp 267–514
Coons LB, Roshdy MA, Axtell RC (1974) Fine structure of the centralnervous system of Dermacentor variabilis, Amblyomma ameri-canum and Argas arboreus (Ixodidea). J Parasitol 60:687–698
El Shoura SM (1986) Fine structure of the synganglion of Ornitho-doros (Pavlovskyella) erraticus (Ixodoidea: Argasidae). Acarolo-gia 27:295–302
El Shoura SM (1989) Ultrastructure of the synganglion in the larvaltick Amblyomma americanum (Ixodoidea: Ixodidae). Exp ApplAcarol 7:239–244
Farias MPO, Sousa DP, Arruda AC, Arruda MSP, Wanderley AG,Alves LC, Faustino MAG (2007) Eficácia in vitro do óleo daCarapa guianensis Aubl. (andiroba) no controle de Boophilusmicroplus (Acari: Ixodidae). Rev Bras Plantas Med 9:68–71
Farias MPO, Sousa DP, Arruda AC, Wanderley AG, Teixeira WC,Alves LC, Faustino MAG (2009) Potencial acaricida do óleo deandiroba Carapa guianensis Aubl. sobre fêmeas adultas ingurgi-tadas de Anocentor nitens Neumann, 1897 e Rhipicephalus san-guineus Latreille, 1806. Arq Bras Med Vet Zootec 61:877–882
Haüserman W, Friedel T, Hess EA, Strong MB (1992) A new activeingredient for a new approach to protect cattle against ticks. In:Proceedings of XIX International Congress of Entomology, pp 138
Ivanov VP (1983) Central nervous system. In: Balashov YuS (ed) AnAtlas of Ixodid Tick Ultrastructure, Entomol Soc Am Spec Publ,pp 175-190
Jongejan F, Uilenberg G (2004) The global importance of ticks. Para-sitology 129(Suppl 1):S3–S14
Labruna MB (2004) Biologia-ecologia de Rhipicephalus sanguineus(Acari: Ixodidae). Rev Bras Parasitol Vet 13:123–124
Lees K, Bowman AS (2007) Tick neurobiology: recent advances andthe post-genomic era. Invert Neurosci 7:183–198
Marcondes CB (2009) Doenças transmitidas e causadas por artrópodes.Editora Atheneu, São Paulo
Marzouk AS, Mohamed FSA, Omar NR (2001) Fine structure of thesynganglion of unfed female Rhipicephalus (Rhipicephalus) san-guineus (Latreille) (Ixodoidea: Ixodidae). J Egypt Soc Parasitol31:1–12
Mencke N, Volp P, Volfova V, Stanneck D (2003) Repellent efficacy ofa combination containing imidacloroprid and permethrin againstsand flies (Phlebotomus papatasi) on dogs. Parasitol Res 90:108–111
Obenchain FD, Oliver JH (1976) Peripheral nervous system of theticks, Amblyomma tuberculatumMarx and Argas radiatus Railliet(Acari: Ixodoidea). J Parasitol 62:811–818
Oliver JH, Zhu XX, Vogel GN, Dotson EM (1992) Role of syngan-glion in oogenesis of the tick Ornithodoros parkeri (Acari: Arga-sidae). J Parasitol 78:93–98
Paz GF, Labruna MB, Leite RC (2008) Ritmo de queda de Rhipice-phalus sanguineus (Acari: Ixodidae) de cães artificialmente infes-tados. Rev Bras Parasitol Vet 17:139–144
Parasitol Res

Prullage JB, Pound JM, Meola SM (1992) Synganglion morphologyand neurosecretory centres of adult Amblyomma americanum (L.)(Acari: Ixodidae). J Med Entomol 29:1023–1034
Reynolds ES (1963) The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J Cell Biol 17:208–212
Ribeiro AL, Faccini JLH, Daemon E (1996) Estudo das variaçõesmorfológicas de Rhipicephalus sanguineus (Latreille, 1806) (Ac-ari: Ixodidae) no Brasil. Rev Univ Fed Rural Rio J 18:25–33
Roma GC, Nunes PH, Remedio RN, Camargo-Mathias MI (2012)Synganglion histology in different stages of Rhipicephalus san-guineus ticks (Acari: Ixodidae). Parasitol Res. doi:10.1007/s00436-011-2785-6
Roshdy MA, Marzouk AS (1984) The subgenus Persicargas (Ixodoi-dea: Argasidae: Argas): A. (P.) Arboreus central nervous system. JParasit 70:774–787
Sonenshine DE (1991) Biology of ticks. Oxford University Press, NewYork
Storer TI, Usinger RL (2000) Zoologia Geral. Companhia EditoraNacional, São Paulo
Taylor MA (2001) Recent developments in ectoparasiticides. Vet J161:253–268
Walker A (1994) The arthropods of humans and domestic ani-mals. A guide to preliminary identification. Chapman andHall, London
Parasitol Res




















