Selection of an ivermectin-resistant strain of Rhipicephalus microplus (Acari: Ixodidae) in Brazil

0
d
www.elsevier.com/locate/vetpar
Veterinary Parasitology 129 (2005) 299–311
Morphological, histological, and ultrastructural studies
of the ovary of the cattle-tick Boophilus microplus
(Canestrini, 1887) (Acari: Ixodidae)
Kelly Cristina Saito a,*, Gervasio Henrique Bechara b, Erika Takagi Nunes a,Patricia Rosa de Oliveira a, Sandra Eloisi Denardi a,
Maria Izabel Camargo Mathias a
a Departamento de Biologia, I.B., Universidade Estadual Paulista Julio de Mesquita Filho, Av. 24 A, No. 1515 Cx,
Postal 199, CEP 13506-900, Rio Claro, SP, Brasilb Departamento de Patologia Veterinaria, FCAV, UNESP, Jaboticabal, SP, Brasil
Accepted 4 September 2004
Abstract
This study presents the morphology of the ovary, as well as the dynamics of the vitellogenesis process in oocytes of the cattle-
tick Boophilus microplus. The ovary of these individuals is of the panoistic type; therefore, it lacks nurse cells. This organ
consists of a single tubular structure, continuous, and composed of a lumen delimitated by a wall of small epithelial cells with
rounded nuclei. In this tick species, the oocytes were classified into six stages varying from I to VI and according to: cytoplasm
appearance and presence of the germ vesicle, yolk granules, and chorion. Oocytes of various sizes and at different developmental
stages remain attached to the ovary through a cellular pedicel until completing stage V. Afterwards, they are liberated into the
lumen and from there to the exterior. Some oocytes (classified as type VI) showed an atypical appearance indicating that some of
the cellular components would be undergoing a degenerative process and/or reabsorption.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Boophilus microplus; Cattle-tick; Ovary; Vitellogenesis; Histology; Ultrastructure
1. Introduction
The economic importance of ticks is widely
acknowledge and is related to their feeding habits.
When feeding, many species of ticks transmit diseases
to man and other animals. The diseases transmitted by
* Corresponding author. Tel.: +55 19 35340009.
304-4017/$ – see front matter # 2004 Elsevier B.V. All rights reserved
oi:10.1016/j.vetpar.2004.09.020
ticks are caused by protozoa, viruses, Rickettsias, and
spirochetes and cause the appearance of dermatosis
and other more severe infections (Rey, 1973). Among
the species belonging to the family Ixodidae is the tick
Boophilus microplus, known as the southern cattle-
tick, which is of great veterinary importance for
transmitting the disease known as cattle babesiosis or
cattle fever. This disease causes severe economical
.

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311300
losses to the cattle industry in many countries (Arthur,
1996).
Numerous studies are currently under way aiming
at finding an efficient control strategy that would
minimize the damages caused by these parasites. A
new control perspective uses immunological methods
consisting of the identification, isolation, and synth-
esis of proteins that protect the tissues and organs of
the tick, mainly those of the reproductive system,
aiming to obtain an anti-cattle-tick vaccine (Tellam
et al., 1992; Willadsen, 1997).
According to Sonenshine (1994), the female
reproductive system of ticks generally consists of a
large U-shaped ovary located at the posterior region of
the body, with a pair of oviducts, an uterus, a muscular
connection tube, vagina, and genital opening. After
fertilization, the ovary increases in size considerably
exhibiting eggs at all developmental stages (Rey,
1973). The ovary is an organ with a lumen delimitated
by a wall of small epithelial cells of rounded nuclei, in
which the oocytes at the different developmental
stages are attached by means of a pedicel (Sonenshine,
1994). Denardi et al. (2004) studied the ovaries of the
lone star tick Amblyomma cajennense and suggested
that the oocytes are developed simultaneously but
asynchronously; therefore, it is possible to observe
throughout the ovaries of these individuals oocytes
from the initial developmental stages to the most
mature ones.
In view of the data mentioned above and due to the
existence of few and out-of-date studies focusing on the
internal organs of the cattle-tick B. microplus, the
present work was aimed at collecting new data on the
morphological, histological, and ultrastructural features
of the female reproductive system of B. microplus, as
well as to attempt to establish the dynamics of the
vitellogenesis process in these individuals. The final aim
is to provide information that would contribute to the
future control of this parasite.
2. Material and methods
About 20 females of the cattle-tick B. microplus
were collected from tick colonies maintained in
controlled conditions (28 8C, 80% humidity and 12 h
photoperiod) at the Department of Animal Pathology,
Veterinary College, UNESP, Jaboticabal, SP, Brazil.
Equipment from the Histology and Electron Micro-
scopy Laboratories of the Biology Department at the
Biosciences Institute, UNESP, Rio Claro, SP, Brazil,
was utilized throughout the study.
Individuals were maintained in the refrigerator
(4 8C) for thermal shock anesthesia and dissected in
saline solution.
2.1. Morphology
Individuals were anesthetized and the ovaries were
removed and schematically drawn with the aid of
camera lucida coupled to a Zeiss stereomicroscope.
2.2. Scanning Electron Microscopy (SEM)
The ovaries were removed, fixed in Karnovsky for
24 h, and dehydrated in a graded 70–100% ethanol
and acetone series. The material was processed by
cryptal point drying, sputtered with gold, and
examined and photographed under a SEM Phillips
505.
2.3. Histology
The ovaries were fixed in 4% paraformaldehyde.
The material was then dehydrated in ethanol,
embedded in JB4 resin during 24 h at 4 8C and then
transferred to plastic moulds previously filled with
resin containing a catalyzer. After resin polymeriza-
tion, the material was sectioned using a Sorvall JB4
microtome (Bio Rad) and stained with hematoxiline
and eosin, following routine histological procedures.
2.4. Histochemistry
Histochemical tests were applied in order to detect
the presence of the following compounds: proteins
(Bromophenol Blue); lipids (Nile Blue); polysacchar-
ides (simultaneous staining with PAS/ Alcian Blue);
acids (Toluidine Blue); and calcium (Von Kossa).
2.5. Transmission Electron Microscopy (TEM)
The material was fixed in 2.5% glutaraldehyde,
postfixed in 1% OsO4, and embedded in Epon
Araldite. The material was then included in pure
Epon resin and polymerized at 60 8C for 72 h.

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311 301
Ultrathin sections were contrasted with acetate and
lead citrate. Afterwards, screens containing ultrathin
sections of the material were examined and photo-
graphed in a TEM Phillips MC 100.
3. Results
3.1. Ultramorphology
The application of scanning electron microscopy
techniques (SEM) allowed us to verify that the ovary
of the southern cattle tick consists of a single tubular
organ, in the shape of a horseshoe, to which numerous
rounded oocytes of various sizes and at various
developmental stages are attached (Fig. 1). This
morphological variation of the oocytes is related to the
fact that their development does not proceed
uniformly, in other words, the different developmental
stages are processed simultaneously but asynchro-
nously throughout the ovary (Fig. 1).
3.2. Histology
The ovary of the tick B. microplus is composed of a
lumen delimitated by a wall of small epithelial cells
with rounded nuclei, to which the oocytes in the
different developmental stages are attached through a
pedicel (Figs. 2 and 3A,B). Once mature, the oocytes
are liberated inside the lumen and from there to the
exterior (Fig. 2).
The pedicels, which attach the oocytes to the
ovarian wall, are composed of cells that resemble
those that compose the epithelial wall (Fig. 3A,C).
Since the differences found in the oocytes were
significant, we classified them following the criteria
previously established for the oocytes of A. cajennense
by Denardi et al. (2004). The classification suggested
for the oocytes found in the ovary of B. microplus is as
follows.
3.3. Oocyte I or primary oocyte
Characterized by its small size and frequently
rounded shape. The plasmic membrane is relatively
thin and delimitates a homogeneous cytoplasmic
content containing a germ vesicle with an evident
nucleolus (Figs. 2, 3B, 4A,D and 5A).
3.3.1. Oocyte II or pre-vitellogenic oocyte
It is larger when compared to the previous stage. A
fine granulation is observed dispersed throughout the
cytoplasm and the germ vesicle with a nucleolus still
evident (Figs. 2, 3A,D, 4A and 5A,B).
3.3.2. Oocyte III or vitellogenic oocyte
Its volume is considerably increased in relation to
the previous stages. The cytoplasm contains a coarser
granulation and the germ vesicle appears located at the
pole, next to the ovarian wall. Surrounding the oocyte,
at the external surface of the plasmic membrane, starts
the deposition of the chorion (Figs. 2, 3A,E, 4C,E and
5B).
3.3.3. Oocyte IV
Large yolk granules appear at the central region of
the cytoplasm, arising from smaller granules at the
peripheral region. The presence of a germ vesicle is
still evidenced. The thick chorion appears completely
deposited (Figs. 2; 3 C; 4C, E).
3.3.4. Oocyte V
This type of oocyte is well developed and
surrounded by a thick chorion. The cytoplasm
appears completely filled by numerous and large yolk
granules; the germ vesicle is rarely observed (Figs. 2,
3A and 4C,F).
3.3.5. Oocyte VI
Possesses an abnormal morpho-histological aspect
characterized by a cytoplasmic disorganization and
numerous folds at the chorion, rendering it an irregular
contour (Figs. 1, 3A, 4B,E and 5C). They were
observed in all individuals examinated.
4. Histochemistry
4.1. Bromophenol Blue test for the detection of
proteins (according to Junqueira and Junqueira, 1983).
This technique revealed the presence of proteins in
the plasmic membrane and inside all the oocytes.
Nevertheless, the distribution of this material through-
out the cytoplasm was not uniform: in stages I and II,
the proteins appeared dispersed homogeneously; in
stage III the proteins appeared as a fine granulation
K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311302
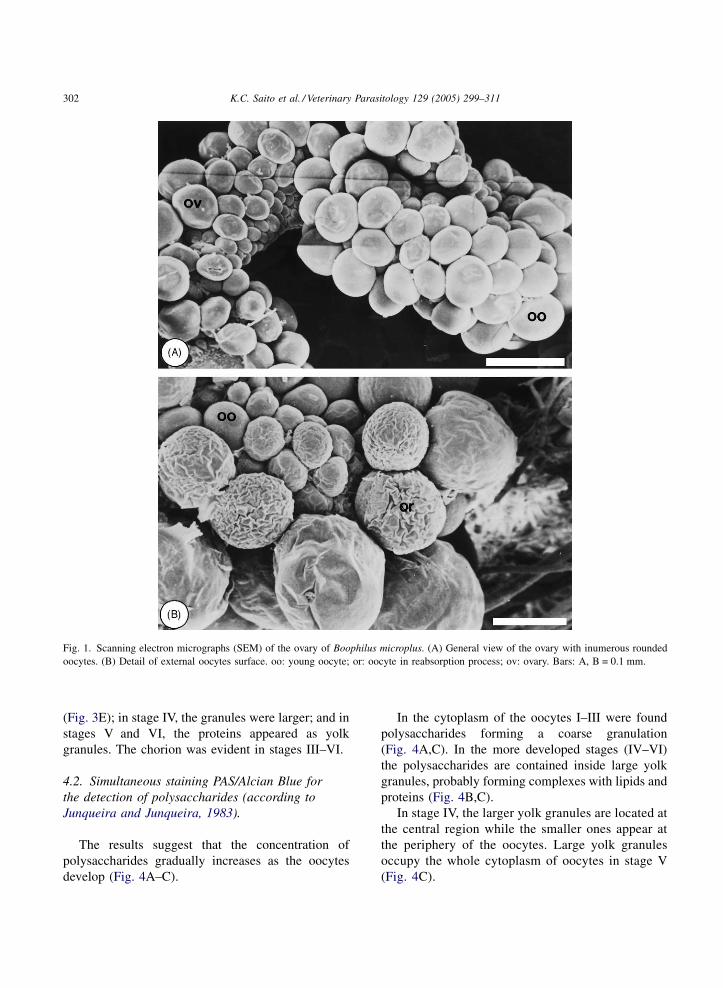
Fig. 1. Scanning electron micrographs (SEM) of the ovary of Boophilus microplus. (A) General view of the ovary with inumerous rounded
oocytes. (B) Detail of external oocytes surface. oo: young oocyte; or: oocyte in reabsorption process; ov: ovary. Bars: A, B = 0.1 mm.
(Fig. 3E); in stage IV, the granules were larger; and in
stages V and VI, the proteins appeared as yolk
granules. The chorion was evident in stages III–VI.
4.2. Simultaneous staining PAS/Alcian Blue for
the detection of polysaccharides (according to
Junqueira and Junqueira, 1983).
The results suggest that the concentration of
polysaccharides gradually increases as the oocytes
develop (Fig. 4A–C).
In the cytoplasm of the oocytes I–III were found
polysaccharides forming a coarse granulation
(Fig. 4A,C). In the more developed stages (IV–VI)
the polysaccharides are contained inside large yolk
granules, probably forming complexes with lipids and
proteins (Fig. 4B,C).
In stage IV, the larger yolk granules are located at
the central region while the smaller ones appear at
the periphery of the oocytes. Large yolk granules
occupy the whole cytoplasm of oocytes in stage V
(Fig. 4C).

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311 303
Fig. 2. Diagrammatic summary of oogenesis in ovarian cross-section of Boophilus microplus. I: oocyte I; II: oocyte II; III: oocyte III; IV: oocyte
IV; V: oocyte V; ec: ovary ephitelium; lu: lumen; ovd: oviduct; gv: germinal vesicle.
4.3. Nile Blue test for the detection of lipids
(according to Junqueira and Junqueira, 1983).
This histochemical test revealed the presence of
lipids in the plasmic and nuclear membranes and also
inside the oocytes (Fig. 4D–G). The concentration of
this material stored in the cytoplasm of the oocytes
varied according to the various developmental
stages.
The presence of a high concentration of lipids
in the cytoplasm of all oocytes was verified.
However, in the stages III–VI, lipids were only
present at the spaces between the yolk granules
(Fig. 4D–G).
The chorion reacted positively to this test, thus
indicating the presence of lipids in its composition
(Fig. 4E,G).
4.4. Von Kossa test for the detection of calcium
(according to Junqueira and Junqueira, 1983).
The results revealed that calcium is present in the
oocytes III–VI, distributed throughout their cytoplasm
(Fig. 5A–C).
K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311304
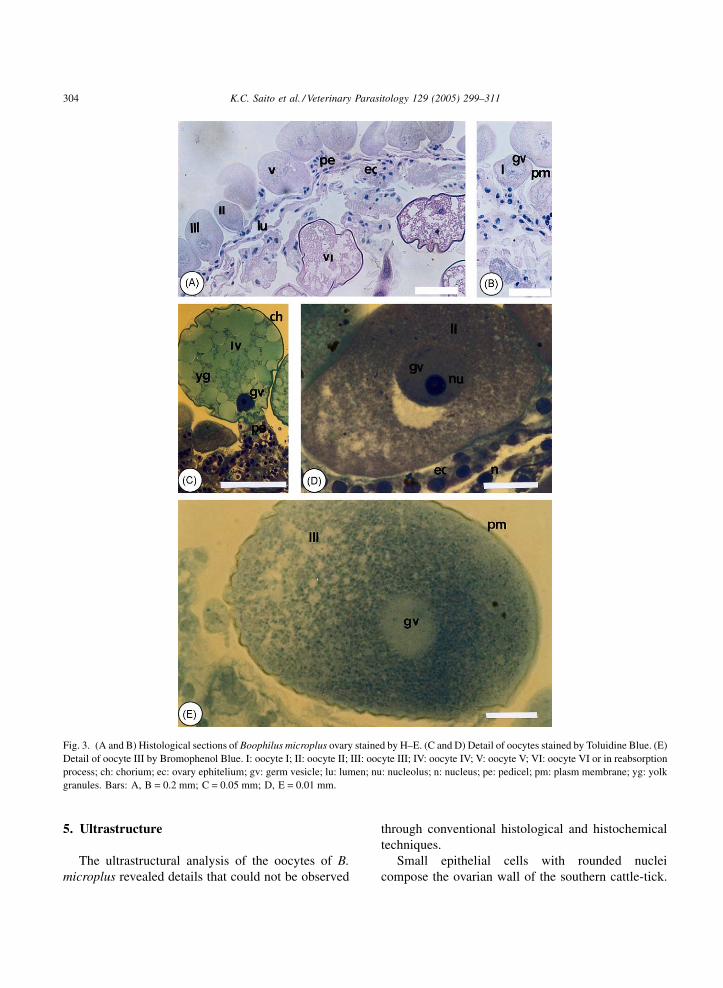
Fig. 3. (A and B) Histological sections of Boophilus microplus ovary stained by H–E. (C and D) Detail of oocytes stained by Toluidine Blue. (E)
Detail of oocyte III by Bromophenol Blue. I: oocyte I; II: oocyte II; III: oocyte III; IV: oocyte IV; V: oocyte V; VI: oocyte VI or in reabsorption
process; ch: chorium; ec: ovary ephitelium; gv: germ vesicle; lu: lumen; nu: nucleolus; n: nucleus; pe: pedicel; pm: plasm membrane; yg: yolk
granules. Bars: A, B = 0.2 mm; C = 0.05 mm; D, E = 0.01 mm.
5. Ultrastructure
The ultrastructural analysis of the oocytes of B.
microplus revealed details that could not be observed
through conventional histological and histochemical
techniques.
Small epithelial cells with rounded nuclei
compose the ovarian wall of the southern cattle-tick.

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311 305
Fig. 4. Histological sections of Boophilus microplus ovary. (A–C) Stained by PAS/Alcian Blue. (D–G) Stained by Nile Blue. I: oocyte I; II:
oocyte II; III: oocyte III; IV: oocyte IV; V: oocyte V; VI: oocyte VI; ch: chorium; ec: ovary ephitelium; gv: germ vesicle; lu: lumen; n: nucleolus;
pe: pedicel; yg: yolk granules. Bars: A, B, E, and G = 0.2 mm; C and F = 0.01 mm; D = 0.05 mm.
The oocytes at the different developmental stages are
attached to this wall through a cellular pedicel (Fig. 6J).
The cells that compose the wall and the pedicels
presented a varied morphology from rounded to oval
with a rounded nucleus surrounded by a fine layer of
cytoplasm, in which we noted the presence of vacuoles
and granular endoplasmic reticulum (Fig. 6J).
The basal lamina that appears in these cells was
differentiated into two layers: a more external and
thinner layer of fibrillar aspect and a more internal and
thicker layer (Fig. 6C). This basal lamina support the
more external cells that formed the pedicels and on the
oocytes, being absent at the point of contact between
the oocytes and the pedicel cells, in which we verified
a specialization of the membrane of these two cellular
types in the shape of interdigitations (Fig. 6J).
The cytoplasm of oocytes at the initial develop-
mental stages (I and II) shelters the germ vesicle and a

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311306
Fig. 5. Histological sections of Boophilus microplus ovary stained by Von Kossa. I: Oocyte I; II: oocyte II; III: oocyte III; VI: oocyte VI; ca:
calcium spheres; ec: ovary ephitelium; lu: lumen. Bars: A, B = 0.2 mm; C = 0.01 mm.

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311 307
Fig. 6. Transmission electron micrographs of Boophilus microplus ovary. (A) Oocyte I. (B) The perinuclear region of oocyte II. (C) Peripheric
region of the oocyte III. (D) Chorium formation of oocyte IV. (E) The chorium of oocyte V completely deposited. (F) The cytoplasm of oocyte V
presented large yolk granules. (G and H) Myelinic bodies and autophagic vacuoles presents in oocyte VI. (I) Chorium of oocyte VI showing
myelinic bodies inside them. (J) Interface oocyte/pedicel region. av: autophagic vacuoles; bl: basal lamina; ch: chorium; g: glycogen; ger:
granular endoplasmic reticulum; gv: germ vesicle, l: lipid granule; m: mitochondria; mb: myelinic bodies; nu: nucleolus; o: oocyte, p: protein
granule; pe: pedicel; pm: plasmic membrane; sv: secretion vesicles; i: interdigitation between two pedicel cells; n: nucleus. Bars: A = 10 mm; B,
C and G = 2 mm; D, F, H and I = 1 mm; E and J = 5 mm.

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311308
small amount of material stored as a fine granulation
that renders a homogeneous appearance to the
cytoplasm (Fig. 6A,B).
During stage I, the chromatin of the germ vesicle is
poorly condensed and a large amount of granular
endoplasmic reticulum, glycogen, and ribosomes
appear free in the cytoplasm (Fig. 6A).
The oocytes in stage II, also known as pre-
vitellogenic oocytes, presented in their cytoplasm a
large amount of granular endoplasmic reticulum,
vesicles of varied morphologies, glycogen, and RNA
precursors that were found at the perinuclear region
together with numerous mitochondria (Fig. 6B).
In the cytoplasm of oocytes in stage III a larger
amount of glycogen, granular endoplasmic reticulum,
polyribosomes, and lipid droplets of varying shapes
and sizes were noted (Fig. 6C).
At the periphery of oocytes in stage III, in the
region next to the basal lamina, small secretion
vesicles being incorporated by the cell were observed
(Fig. 6C). The plasmic membrane presented numerous
infoldings towards the basal lamina and, in the spaces
formed between these structures, the export of vesicles
filled with an electrondense material was verified; this
material is deposited and then polymerizes for the
formation of the chorion.
As the oocytes progress in their development (stages
IVand V), the infoldings of the plasmic membrane tend
to decrease and a larger amount of yolk granules is
observed in the cytoplasm, resulting from the fusion of
small secretion vesicles (Fig. 6D–F).
In oocytes V, the chorion appeared completely
formed with micropores that probably facilitate the
oxygenation of the internal structures of the future egg
(Fig. 6E). During this same stage, the cytoplasm of the
oocyte presented large yolk granules (Fig. 6F).
The oocytes in stage VI presented in their
cytoplasm strong indications that some of their
components were undergoing degeneration and/or
reabsorption. We noted the presence of myelinic
bodies as well as mitochondria and endoplasmic
reticulum with altered morphology inside autophagic
vacuoles (Fig. 6G,H).
The chorion of the degenerating oocytes presented
characteristics that differed from the oocytes in
previous developmental stages, showing a few
myelinic bodies inside them (Fig. 6I) as well as a
disorganized cytoplasmic yolk.
6. Discussion
The ultramorphological analysis revealed that the
ovary of the cattle-tick consists of a single tubular
structure, continuous, and in the shape of a horseshoe,
located at the posterior third of the animal’s body. This
data corroborate the results obtained by Sonenshine
(1994) in the species Dermatocentor andersoni and D.
variabilis, by Denardi et al. (2004) in A. cajennense, and
by Oliveira et al. (2004) in Rhipicephalus sanguineus.
The application of histological techniques allowed
the classification of the ovary of B. microplus as
belonging to the panoistic type, in which all the cells
present, except the ones that compose the ovarian wall,
are oogonies that would give rise to the future oocytes.
Nurse cells are absent in this type of ovary. This
classification for B. microplus is the same described
for A. cajennense in the study made by Denardi et al.
(2004) and for R. sanguineus collected in Brazil and
Argentina (Oliveira et al., 2004).
Morphologically, the ovary of B. microplus is not
segmented into different developmental zones as
occur in the ovaries of most arthropods.
The morphological variation that exists between
the oocytes found in the ovary of this species was not
only restricted to the external aspects but also to their
internal composition. These differences were a
consequence of the different developmental stages
displayed by the oocytes, thus suggesting a simulta-
neous but asynchronous development of these cells,
which was also observed in the ovary of A. cajennense,
as reported by Denardi et al. (2004).
It was demonstrated that in the cattle-tick, the
oogenesis proceeds in a distal-proximal direction,
with the less developed oocytes occupying the
distal region while the more developed oocytes (or
vitellogenic) occupy the proximal region. This
configuration was also observed by Wagner-Jevseenko
(1958) in other species of ticks and by Fontanetti and
Cunha (1993) in millipedes.
The significant morphological and histochemical
differences related to the developmental stage and the
amount of yolk found in the oocytes of B. microplus
allowed the classification of the oocytes into stages
that varied from I to VI. This classification was based
on the one proposed by Denardi et al. (2004) for the
development of oocytes of the lone star tick A.
cajennense, in which the author described the

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311 309
occurrence of oocytes in five different developmental
stages. We described an additional stage for the
oocytes of B. microplus, where were observed features
of degeneration. The main characteristics considered
for this classification were the aspect of the cytoplasm,
the presence of a germ vesicle, the presence and size of
yolk granules, and the presence of the chorion.
The histological study revealed that the oocytes in
stages I–IV, and rarely some of stage V are attached to
the ovarian wall through a pedicel. This structure
results from the proliferation of epithelial cells of the
ovarian wall during the development of the oocytes.
This phenomenon was described by Till (1961) and
Balashov (1983) for other species of ticks.
During the first studies performed with A.
cajennense, El Said (1992) suggested that the pedicel
releases the mature oocytes into the ovarian lumen due
to the continuous growth pressure they exert against
the ovarian wall. In addition to this function, Balashov
(1983) suggested that the pedicels would play the role
of the nurse cells, present in ovaries of the meroistic
type and thus absent in B. microplus, which aid in the
incorporation of proteins and lipids that would later be
incorporated into the oocytes.
The ultrastructural study revealed a specialization
of the plasmic membranes at the interface between
the pedicel cells and the oocytes, where they present
infoldings that interdigitate, thus increasing the con-
tact surface between these two types of cells. This
observation corroborates the idea that the pedicels
are actually aiding in the production of elements
that would later be incorporated into the oocytes of
B. microplus.
The basal lamina was observed leaning on the
plasmic membranes of the oocytes and pedicels, except
at the point of contact between these two cellular
types. The basal lamina was divided into two different
regions: a more external layer, thinner and with a
fibrillar aspect, and a more internal layer, thicker and of
granular appearance. This description of the basal
lamina coincides with the observations of Denardi et al.
(2004) in A. cajennense and of Silva (1999) in locust of
the genus Panulirus. In B. microplus, the basal lamina
does not penetrate the interface region between the
pedicel and the oocyte, the region of intense contact
between these two cellular types.
According to Reddy and Locke (1990), the basal
lamina’s main function would be to serve as a barrier
between the elements of the hemolymph and of the
oocytes, thus monitoring the traffic of macromole-
cules in both directions.
At the perinuclear region of the oocytes in the
developmental stages I and II, we observed an intense
traffic of material from the nucleus to the cytoplasm
through the nuclear pores, probably rRNA, with a
consequent increase in the amount of ribosomes as
well as the deposition of electrondense material at this
region.
The characteristics described above for oocytes I
and II were also described for A. cajennense by Denardi
et al. (2004) and for R. sanguineus by Oliveira et al.
(2004), thus suggesting the occurrence of an endogen-
ous synthetic activity of only structural proteins during
the first stage and later, during the second stage, a
production of vitellogenic proteins, including the
exogenous incorporation of yolk elements.
Oocytes at the end of stage II and at the beginning of
stage III presented a significant increase in the number
of organelles accumulated in the cytoplasm, which
might be related to the preparation of these cells for the
next stage, during which these organelles would be a
prerequisite for the endogenous production and/or the
exogenous incorporation of yolk (Balashov, 1972).
The abundance of rough endoplasmic reticulum in
oocytes III of B. microplus suggests that the presence
of this organelle indicates that endogenous synthesis
of yolk is occurring in the oocyte. In addition to
the endoplasmic reticulum, some authors relate the
participation of the Golgi complex in endogenous
vitellogenesis. Nevertheless, this organelle was not
observed in the oocytes of the cattle-tick, probably
indicating a small participation of these cells in the
vitellogenesis process. Norevang (1968), while
reviewing the ultrastructural aspects of oogenesis in
diverse animal groups, suggested that in most oocytes
the Golgi complex is not a prominent element and few
comments have been made with regard to their
function in oocyte physiology in general.
Consequently, we suggest that the vitellogenesis
process in B. microplus occurs through the endogen-
ous production of lipids and proteins until reaching
developmental stage III; afterwards, there are exo-
genous elements that are incorporated from the
hemolymph. The hypothesis of exogenous production
of yolk results from the observation of small yolk
vesicles located preferentially at the peripheral region

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311310
of the oocytes while larger yolk granules concentrate
at the central region, thus indicating that the material
might be incorporated through pinocytosis.
When the vitellogenesis process is completed
during stage V, the infoldings observed frequently
on the plasmic membrane of the oocytes of B.
microplus decrease in size and number until totally
disappearing, thus confirming that such specialization
would be related to the incorporation of materials from
an exogenous source in order to contribute to yolk
formation inside the oocytes.
Our results indicate that the yolk granules of
oocytes of the cattle-tick are only composed of
glycoproteic elements, with lipids being only present
in the cytoplasm between the granules. This observa-
tion disagrees with the results observed by Ramamurty
(1968) in insects and by Oliveira et al. (2004) in
R. sanguineus, species in which the yolk granules are
composed of lipids, proteins, and carbohydrates.
In oocytes at developmental stage V of B.
microplus, the large yolk granules are distributed
throughout the whole cytoplasm. Nevertheless, at the
peripheral region it is still possible to observe a thinner
granulation, which indicates the incorporation of
extra-ovarian materials via pinocytosis vesicles that
Denardi et al. (2004) described as originated from the
hemolymph in A. cajennense.
As the oocyte completes its development, it becomes
surrounded by the chorion, towhich numerous functions
are attributed, such as preservation of the egg with the
consequent preservation of the species (King, 1960);
protection against mechanical stimuli, desiccation, and
predation; and allowing gas exchange for the oxygena-
tion of the embryo (Hinton, 1982).
According to Camargo-Mathias (1993), in the ant
Neoponera villosa the chorion is structurally formed
by the endochorion (more internal and electrondense)
and by the exochorion (more external and less
electrondense), which are normally composed of
proteins, lipids, and carbohydrates synthesized by the
follicular cells in the case of meroistic polytrophic
ovaries (King, 1960).
Numerous authors believe that the chorion is
produced by the oocyte itself, originating from
vesicles of the endoplasmic reticulum and of the
Golgi, which fuse to the plasmic membrane of the
more developed oocytes and discharge their contents
to the extracellular space in which it polymerizes.
The ultrastructural study revealed that, in B.
microplus, the chorion is not composed of different
layers and, due to the absence of follicular cells, it is
composed of an electron-dense material produced by
the oocyte itself. This material later accumulates at the
space between the plasmic membrane and the basal
lamina; its composition in the cattle-tick consists
basically of compounds of a lipoproteic nature.
Inside the oocytes, mainly those in stages III–V, in
addition to the yolk granules we found calcium
spheres. This element was also observed by Denardi
et al. (2004) in the more developed oocytes of A.
cajennense and in millipedes by Kubrakiewicz (1989),
that suggested that this element originated from
systems of intracellular mineralization.
According to Camargo-Mathias et al. (1998), in
millipedes the accumulation of this mineral in the
shape of spherocrystals could represent a mechanism
of homeostasis or some sort of special reserve, which
would later be used in the calcification of the
exoskeleton of the millipede embryo (Pettit, 1970).
The actual function of this element present in the
cytoplasm of the oocytes of B. microplus cannot yet be
confirmed.
Throughout the ovary of B. microplus we observed a
few oocytes with an abnormal morpho-histological
appearance, characterized by a cytoplasmic disorgani-
zation and many folds on the chorion. The ultrastructure
of these oocytes revealed strong indications that some
of the cellular components would be undergoing a
degenerative process and/or reabsorption, since we
observed myelinic bodies as well as mitochondria and
endoplasmic reticulum inside autophagic vacuoles.
Thus, these oocytes appear to be undergoing
autolysis without being phagocytized, or without the
participation of hemolymph cells, phenomenon that was
also described by Crayon (1941) in his analyses with
oocytes of pagurid crabs undergoing reabsorption.
Oliveira et al. (2004) showed that in R. sanguineus
a process of reabsorption in some oocytes probably
occurs as a mechanism to recover certain nutrients.
Lusis (1963) even suggested that this process occurs
due to a series of physiological, ecological, and
behavioral factors, such as lack of food by a
determined period, which would interrupt the deposi-
tion of yolk and would promote the lysis of the
existing granules with a consequent rupture of the
cytoplasm (Bell and Bohm, 1975).

K.C. Saito et al. / Veterinary Parasitology 129 (2005) 299–311 311
The causes for the appearance of oocytes undergoing
reabsorption in the tick B. microplus remain unknown.
The findings described here regarding the mor-
phological, histological, and ultrastructural aspects of
the female reproductive system of the cattle-tick
B. microplus may contribute to the future control of
this parasite, since the survival of the species depends
on this system.
Acknowledgments
This work has been supported by FAPESP
(Fundacao de Amparo a Pesquisa do Estado de Sao
Paulo). The authors wish to tank Ronaldo Del Vecchio,
Antonio Teruyoshi Yabuki, Monika Iamonte, Gerson
de Melo Souza and Cristiane Marcia Mileo for their
technical support. Part of this work has been facilitated
through the International Consortium of Ticks and
Tick-borne Diseases (ICTTD-2) supported by the
INCO-DEV program of the European Union under
contract number ICA4-CT-2000-30006.
References
Arthur, D.R., 1996. Ticks: a monograph of the Ixodoidea: on the
genera Dermatocentor, Anocentor, Cosmiomma. In: Boophilus e
Margaphorus, Cambridge University Press, London, p. 215.
Balashov, Y.S., 1972. A translation of bloodsucking ticks (Ixodidae)
vectors of diseases of man and animals. Misc. Publ. Entomol.
Soc. Am. 8 (5), 159–376.
Balashov, Yu.S., 1983. In: Raikhel A.S., Hoogstraal H. (Eds.), An
Atlas of Ixodid Tick Ultrastructure. Entomological Society of
America, 289 pp. (Special Publication).
Bell, W.J., Bohm, M.K., 1975. Oosorption in insects. Boil. Rev. 50,
373–396.
Camargo-Mathias, M.I., 1993. 157f. Histoquımica e ultra-estrutura
dos ovarios de operarias e rainhas de formigas Neoponera villosa
(Hymenoptera: Ponerinae). Dissertacao (Doutorado em Zoolo-
gia)- Instituto de Biociencias, Universidade Estadual Paulista,
Rio Claro.
Camargo-Mathias, M.I., Fontanetti, C.S., Mico-Balagues, E., 1998.
Histochemical studies of Rhinocricus padbergi Verhoeff ovaries
(Diplopoda, Spirobolida, Rhinocricidae). Cytobios, Cambridge
169–184.
Crayon, J., 1941. Morphologie et structure de I’appareil genital
femelle chez quelques pagures. Bull. Soc. Zool. France. LXVI,
95–122.
Denardi, S.E., Bechara, G.H., Oliveira, P.R., Nunes, E.T., Saito,
K.C., Camargo-Mathias, M.I., 2004. Morphological character-
ization of the ovary and vitellogenesis dynamics in the tick
Amblyomma cajennense (Acari: Ixodidae). Vet. Parasitol. 125,
379–395.
El Said, A., 1992. A contribution to the anatomy and histology of the
female reproductive system of Amblyomma cajennense (Acar-
ina: Ixodidae).. J. Egypt. Soc. Parasitol., Cairo. 22, 391–400.
Fontanetti, C.S., Cunha, M.A.S., 1993. Morfologia ovariana e
desenvolvimento dos ovocitos de Rhinocricus padberg Verhoeff
(Diplopoda, Spirobolida, Rhino cricidae). Ver. Brasil. Biol. 53
(1), 7–12.
Hinton, H.E., 1982. Biology of Insect Egg Shells. Pergamon,
Oxford.
Junqueira, L.C.U., Junqueira, L.M.M., 1983. Tecnicas basicas de
citologia e histologia. Livraria Editora Santos, 48–81.
King, R.C., 1960. Oogenesis in adult Drosophila melanogaster IX
Studies on the citochemistry and ultrastructure of developing
oocytes. Growth 24, 265–323.
Kubrakiewicz, J., 1989. Deposition of calcium salts in oocytes and
ovarian somatic tissue of millipedes. Tissue Cell 21, 443–
446.
Lusis, O., 1963. The histology and histochemistry of development
and resorption in the terminal oocytes of the desert locust
Schistocerca gregaria. Quart. J. Micro. Sci. 104, 57–68.
Norevang, A., 1968. Electron microscopic morphology of oogen-
esis. Int. Rev. Cytol. 23, 114–186.
Oliveira, P.R., Bechara, G.H., Denardi, S.E., Saito, K.C., Nunes,
E.T., Camargo-Mathias, M.I., 2004. Morphological character-
ization of the ovary and oocyte vitellogenesis of the tick
Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae).
Exp. Parasitol., in press.
Pettit, J., 1970. Sur la nature et l’accumulation de substances
minerals dans les oocytes des Polydesmus complanatus (Myr-
iapoda Diplopoda). Compte Rendu Hebd des Seances de l’.
Acad. Sci. 27, 2107–2110.
Ramamurty, P.S., 1968. Origin and distribution of glycogen during
vitellogenesis of the scorpion fly Panorpa communis. J. Insect.
Physiol. 14, 1325–1330.
Reddy, J.T., Locke, M., 1990. The size limited penetration of gold
particles through insect basal laminae. J. Insect Physiol. 36 (6),
397.
Rey, L., 1973. Parasitologia. Guanabara Koogan 633–641.
Silva, J.R.F., 1999. Estudo morfologico em ovarios de lagostas do
genero Panulirus. White 1847 (Crustacea: Decapoda: Palinur-
idae). Dissertacao (Doutor em Zoologia), Instituto de Biocien-
cias da Universidade de Sao Paulo, 187 pp.
Sonenshine, D.E., 1994. Biology of ticks V. 1. Ed.. Oxford Uni-
versity Press, New York, pp. 280–303.
Tellam, R.L., Smith, D., Kemp, D.H., 1992. Vaccination against
ticks. In: Yong, W.K. (Ed.), Animal Parasite Control Using
Biotechnology. CRC Press, Boca Raton, pp. 303–331.
Till, W.M., 1961. A contribution to the anatomy and histology of the
brown ear tick Rhipicephalus appendiculatus Neumann. Mem.
Entomol. Soc. South Africa 6, 1–124.
Wagner-Jevseenko, O., 1958. Fortpflanzung bei Ornithodorus mou-
bata und genitale Uebertragung von Borrelia duttoni. Acta
Trop., Basel. 15, 119–168.
Willadsen, P., 1997. Novel vaccines for ectoparasites. Vet. Parasit.
71, 209–222.
Copyright © 2022 FDOKUMEN