
Boophilus microplus: The pattern of bovine immunoglobulin isotype responses to high and low tick...
10
Experimental Parasitology 110 (2005) 12–21 www.elsevier.com/locate/yexpr 0014-4894/$ - see front matter. Published by Elsevier Inc. doi:10.1016/j.exppara.2005.01.006 Boophilus microplus: The pattern of bovine immunoglobulin isotype responses to high and low tick infestations Suely S. Kashino a , Janaína Resende a , Ana Maria S. Sacco c , Clarisse Rocha a , Laila Proença a , Wanessa A. Carvalho a , Alexandre A. Firmino a , Rosevaldo Queiroz a , Magda Benavides c , Laurel J. Gershwin b , Isabel K.F. De Miranda Santos a,d,¤ a Empresa Brasileira de Pesquisa Agropecuária—Embrapa, Brasília DF, Brazil b University of California, Davis, CA, USA c Embrapa Pecuária Sul, Bagé, RS, Brazil d Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brazil Received 19 April 2004; received in revised form 19 January 2005; accepted 20 January 2005 Abstract Cattle present variable levels of resistance to ticks and the immune correlates of these heritable phenotypes must be known in order to develop eVective vaccines. The antibody responses to tick salivary antigens were examined in cattle of tick-susceptible (Hol- stein) and tick-resistant (Nelore) breeds. After heavy infestations, levels of IgG1 and IgG2 antibodies decreased in Holsteins and remained the same in Nelores. Conversely, levels of IgE antibodies increased in Holsteins. DiVerent sizes of tick burdens modulated the IgG1 antibody response in a susceptible breed (Aberdeen): levels were higher than in controls in heavily infested animals, but not in those undergoing intermediary or minimal infestations. The three experimental groups presented similar levels of IgG2 antibodies. Levels of IgE antibodies were higher only in animals undergoing intermediate infestations. These results indicate that tick infesta- tions suppress the IgG antibody response in susceptible breeds, that IgE antibodies are not protective, and that the dose of tick saliva modulates the isotype of host antibody responses. Published by Elsevier Inc. Index Descriptors and Abbreviations: Boophilus microplus (Ixodidae) is a monoxene (one host) hard tick found in subtropical and tropical regions (Mexico, the Caribbean, Central and South America, parts of Africa, Madagascar, Taiwan, Southeast Asia, and Australia). It is found in wild and domestic herbivores, including cattle (Bos taurus taurus, Bos taurus indicus, and crossbreeds); ELISA, enzyme-linked immunosorbent assay; DTT, dithiotreitol; EDTA, ethylenediaminetetraacetic acid; PBS, phosphate buVered saline; IL, interleukin; TMB; 3,3,5,5-tetramethylbenzidine Keywords: Tick; Ixodidae; Boophilus microplus; Saliva; Immunoglobulin isotypes; Bos taurus taurus; Bos taurus indicus 1. Introduction Ticks cause serious economic losses to animal production, in the order of billions of dollars annually (Anonymous, 1995). Together with other hematophagous arthropod vectors of disease, ticks also present one of the most signiWcant problems that face public health. Alter- natives for the control of vectors are urgently needed due to the increasing lack of eYcacy and/or of acceptance of the current methods that rely on chemicals (Pruett, 1999). In the context of animal production, vaccines are an attractive strategy because they are safe for the environ- ment, for the host, and for consumers. The premise for the immunobiological control of ticks is the Wnding that ¤ Corresponding author. E-mail address: [email protected] (I.K.F. De Miranda Santos).
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Boophilus microplus: The pattern of bovine immunoglobulin isotype responses to high and low tick...

Experimental Parasitology 110 (2005) 12–21
www.elsevier.com/locate/yexpr
Boophilus microplus: The pattern of bovine immunoglobulin isotype responses to high and low tick infestations
Suely S. Kashino a, Janaína Resende a, Ana Maria S. Sacco c, Clarisse Rocha a, Laila Proença a, Wanessa A. Carvalho a, Alexandre A. Firmino a, Rosevaldo Queiroz a,
Magda Benavides c, Laurel J. Gershwin b, Isabel K.F. De Miranda Santos a,d,¤
a Empresa Brasileira de Pesquisa Agropecuária—Embrapa, Brasília DF, Brazilb University of California, Davis, CA, USAc Embrapa Pecuária Sul, Bagé, RS, Brazil
d Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brazil
Received 19 April 2004; received in revised form 19 January 2005; accepted 20 January 2005
Abstract
Cattle present variable levels of resistance to ticks and the immune correlates of these heritable phenotypes must be known inorder to develop eVective vaccines. The antibody responses to tick salivary antigens were examined in cattle of tick-susceptible (Hol-stein) and tick-resistant (Nelore) breeds. After heavy infestations, levels of IgG1 and IgG2 antibodies decreased in Holsteins andremained the same in Nelores. Conversely, levels of IgE antibodies increased in Holsteins. DiVerent sizes of tick burdens modulatedthe IgG1 antibody response in a susceptible breed (Aberdeen): levels were higher than in controls in heavily infested animals, but notin those undergoing intermediary or minimal infestations. The three experimental groups presented similar levels of IgG2 antibodies.Levels of IgE antibodies were higher only in animals undergoing intermediate infestations. These results indicate that tick infesta-tions suppress the IgG antibody response in susceptible breeds, that IgE antibodies are not protective, and that the dose of tick salivamodulates the isotype of host antibody responses.Published by Elsevier Inc.
Index Descriptors and Abbreviations: Boophilus microplus (Ixodidae) is a monoxene (one host) hard tick found in subtropical and tropical regions(Mexico, the Caribbean, Central and South America, parts of Africa, Madagascar, Taiwan, Southeast Asia, and Australia). It is found in wild anddomestic herbivores, including cattle (Bos taurus taurus, Bos taurus indicus, and crossbreeds); ELISA, enzyme-linked immunosorbent assay; DTT,dithiotreitol; EDTA, ethylenediaminetetraacetic acid; PBS, phosphate buVered saline; IL, interleukin; TMB; 3,3�,5,5�-tetramethylbenzidine
Keywords: Tick; Ixodidae; Boophilus microplus; Saliva; Immunoglobulin isotypes; Bos taurus taurus; Bos taurus indicus
1. Introduction
Ticks cause serious economic losses to animalproduction, in the order of billions of dollars annually(Anonymous, 1995). Together with other hematophagous
¤ Corresponding author.E-mail address: [email protected] (I.K.F. De Miranda
Santos).
0014-4894/$ - see front matter. Published by Elsevier Inc.doi:10.1016/j.exppara.2005.01.006
arthropod vectors of disease, ticks also present one of themost signiWcant problems that face public health. Alter-natives for the control of vectors are urgently needed dueto the increasing lack of eYcacy and/or of acceptance ofthe current methods that rely on chemicals (Pruett, 1999).In the context of animal production, vaccines are anattractive strategy because they are safe for the environ-ment, for the host, and for consumers. The premise forthe immunobiological control of ticks is the Wnding that

S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21 13
infestations with this parasite stimulate host immuneresponses, which are implicated in their rejection (Pruett,1999; Wikel, 1996). This phenomenon is reXected by adecrease in feeding, in feeding time, and in the reproduc-tive eYciency of the tick.
The outcome of the host/tick interface is much thesame across a given species. In cattle, however, the levelof infestation with ticks can vary according to breed andeven after repeated infestations, susceptible hosts will stillharbour signiWcantly larger numbers of parasites thanresistant breeds (Mattioli and Cassma, 1995; Mattioliet al., 2000; Wambura et al., 1998). It is well establishedthat these distinct phenotypes are heritable, but the hostgenes involved in their expression have not been identi-Wed nor have the immune correlates been fully described.In addition to factors of the adaptive immune response,or exclusive of them, some investigators believe that resis-tance could be mediated by non-adaptive immune factorssuch as grooming activity, skin colour and thickness, areaof skin available for infestation or length of fur (Mattioli,1998; Meltzer, 1996; Norval, 1992).
DiVerences in several aspects of innate and adaptiveimmune responses do indeed occur between tick-resistantand tick-susceptible breeds of cattle. For example, serumlevels of complement (George et al., 1985) and immuno-globulins (Rechav, 1987) as well as lymphoproliferativeresponses to mitogens are described as higher in resistantbreeds and the concentration of skin histamine correlateswith resistance of cattle to ticks (Willadsen et al., 1979).Tick saliva suppresses production of IL-1 by bovine mac-rophages (Ramachandra and Wikel, 1995) and extractsfrom tick salivary glands suppress lymphoproliferation ofconcanavalin A-stimulated bovine peripheral bloodmononuclear cells (Turni et al., 2002), but in all cases sup-pression is less intense in resistant breeds of cattle.Resistant, but not susceptible, bovines mount cutaneousdelayed-type hypersensitivity reactions to antigens of thetick Boophilus microplus (Bechara et al., 2000). Experi-mental evidence also points to a role for the dose of anti-gen in the outcome of the immune response (Constantand Bottomly, 1997; Grakoui et al., 1999; Guery et al.,1996; Menon and Bretscher, 1998). The resulting dose oftick antigens excreted in tick saliva during infestations ofdiVering intensities could, therefore, be a determinant forthe distinct outcomes of anti-tick immune responses seenin diVerent breeds of cattle.
During a blood meal ticks ingest large amounts ofhost serum containing immunoglobulins, which makesantibodies logical candidates for mediating the expres-sion of the distinct phenotypes of tick infestations. Anti-bodies may confer resistance by neutralising the salivarypharmacopoeia that ticks inject into their hosts and,once host blood is ingested, by damaging the parasite’stissues. Furthermore, depending on their characteristics,antibodies can mediate the inXammatory reactions ofthe host (Casadevall and Profski, 2003) that are
detrimental to ticks (Ribeiro, 1995; Wikel, 1996). Thenature of the antibodies elicited in resistant and suscepti-ble breeds in response to tick antigens has not been fullydescribed. We therefore undertook a comparative studyof the tick saliva-speciWc antibody responses of yearlingsof a resistant and a susceptible breed of cattle that wereequally exposed to B. microplus. We also examinedwhether the level of exposure to salivary antigens modu-lates the saliva-speciWc antibody response. Natural infes-tations with B. microplus in a susceptible breed of cattlewere maintained at high, intermediate or low levels bysubmitting animals to tick control measures with diVer-ent degrees of eYcacy for 14 months. Results indicatethat there are signiWcant diVerences in the antibodyresponses to tick salivary antigens in susceptible andresistant breeds of cattle and that, in susceptible breeds,the levels of saliva-speciWc IgG1 antibodies are modu-lated by the level of exposure to ticks.
2. Materials and methods
2.1. Experimental animals and tick infestation protocols
2.1.1. Experimental design for comparison of the anti-tick saliva antibody response in resistant and susceptible cattle
Sera used in the experiment were obtained from non-related yearling bulls of a tick-resistant breed (Nelore, Bostaurus indicus) and of a tick-susceptible breed (Holstein,Bos taurus taurus). Samples were collected at the timepoints indicated in Section 3 and were kept frozen untilassayed together. The animals were managed together atEmbrapa’s experimental farm in the Federal District,Brazil, in a pasture naturally infested with larvae ofB. microplus. The level of infestation of the experimentalpasture was estimated by means of the number ofengorged female adults counted on susceptible hosts (datanot shown). Because susceptible breeds do not tolerateheavy tick infestations, individual Holstein yearlings weretreated with topical acaricides whenever their tick countsexceeded approximately 500 engorged adults per animal.Tick counts on Nelore yearlings ranged from negligible to15 engorged adults per animal. Holstein animals hadundergone tick control with acaricides before the beginningof the experiment. Sera were Wrst collected at the beginningof a season of heavy exposure to ticks (November).
2.1.2. Experimental design for evaluating the eVect of diVerent levels of exposure to tick saliva on the isotype proWle of the antibody response
Sera used in the experiment were obtained from year-ling heifers of a tick-susceptible breed (Aberdeen). Theexperiment was performed at Embrapa’s experimentalfarm in Bagé, Rio Grande do Sul State, Brazil. Animalswere placed in adjacent plots naturally infested withB. microplus and distributed according to elevation and
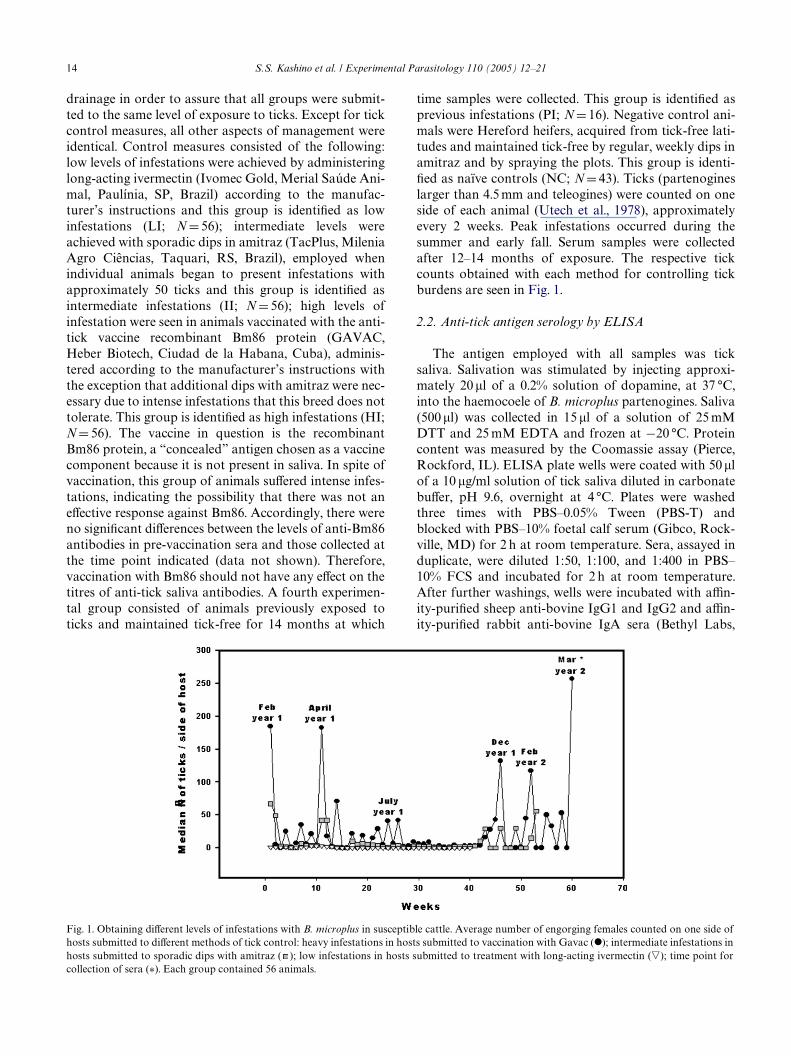
14 S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21
drainage in order to assure that all groups were submit-ted to the same level of exposure to ticks. Except for tickcontrol measures, all other aspects of management wereidentical. Control measures consisted of the following:low levels of infestations were achieved by administeringlong-acting ivermectin (Ivomec Gold, Merial Saúde Ani-mal, Paulínia, SP, Brazil) according to the manufac-turer’s instructions and this group is identiWed as lowinfestations (LI; N D 56); intermediate levels wereachieved with sporadic dips in amitraz (TacPlus, MileniaAgro Ciências, Taquari, RS, Brazil), employed whenindividual animals began to present infestations withapproximately 50 ticks and this group is identiWed asintermediate infestations (II; N D 56); high levels ofinfestation were seen in animals vaccinated with the anti-tick vaccine recombinant Bm86 protein (GAVAC,Heber Biotech, Ciudad de la Habana, Cuba), adminis-tered according to the manufacturer’s instructions withthe exception that additional dips with amitraz were nec-essary due to intense infestations that this breed does nottolerate. This group is identiWed as high infestations (HI;N D 56). The vaccine in question is the recombinantBm86 protein, a “concealed” antigen chosen as a vaccinecomponent because it is not present in saliva. In spite ofvaccination, this group of animals suVered intense infes-tations, indicating the possibility that there was not aneVective response against Bm86. Accordingly, there wereno signiWcant diVerences between the levels of anti-Bm86antibodies in pre-vaccination sera and those collected atthe time point indicated (data not shown). Therefore,vaccination with Bm86 should not have any eVect on thetitres of anti-tick saliva antibodies. A fourth experimen-tal group consisted of animals previously exposed toticks and maintained tick-free for 14 months at which
time samples were collected. This group is identiWed asprevious infestations (PI; N D 16). Negative control ani-mals were Hereford heifers, acquired from tick-free lati-tudes and maintained tick-free by regular, weekly dips inamitraz and by spraying the plots. This group is identi-Wed as naïve controls (NC; N D 43). Ticks (partenogineslarger than 4.5 mm and teleogines) were counted on oneside of each animal (Utech et al., 1978), approximatelyevery 2 weeks. Peak infestations occurred during thesummer and early fall. Serum samples were collectedafter 12–14 months of exposure. The respective tickcounts obtained with each method for controlling tickburdens are seen in Fig. 1.
2.2. Anti-tick antigen serology by ELISA
The antigen employed with all samples was ticksaliva. Salivation was stimulated by injecting approxi-mately 20 �l of a 0.2% solution of dopamine, at 37 °C,into the haemocoele of B. microplus partenogines. Saliva(500 �l) was collected in 15 �l of a solution of 25 mMDTT and 25 mM EDTA and frozen at ¡20 °C. Proteincontent was measured by the Coomassie assay (Pierce,Rockford, IL). ELISA plate wells were coated with 50 �lof a 10 �g/ml solution of tick saliva diluted in carbonatebuVer, pH 9.6, overnight at 4 °C. Plates were washedthree times with PBS–0.05% Tween (PBS-T) andblocked with PBS–10% foetal calf serum (Gibco, Rock-ville, MD) for 2 h at room temperature. Sera, assayed induplicate, were diluted 1:50, 1:100, and 1:400 in PBS–10% FCS and incubated for 2 h at room temperature.After further washings, wells were incubated with aYn-ity-puriWed sheep anti-bovine IgG1 and IgG2 and aYn-ity-puriWed rabbit anti-bovine IgA sera (Bethyl Labs,
Fig. 1. Obtaining diVerent levels of infestations with B. microplus in susceptible cattle. Average number of engorging females counted on one side ofhosts submitted to diVerent methods of tick control: heavy infestations in hosts submitted to vaccination with Gavac (I); intermediate infestations inhosts submitted to sporadic dips with amitraz ( ); low infestations in hosts submitted to treatment with long-acting ivermectin (�); time point for
collection of sera (¤). Each group contained 56 animals.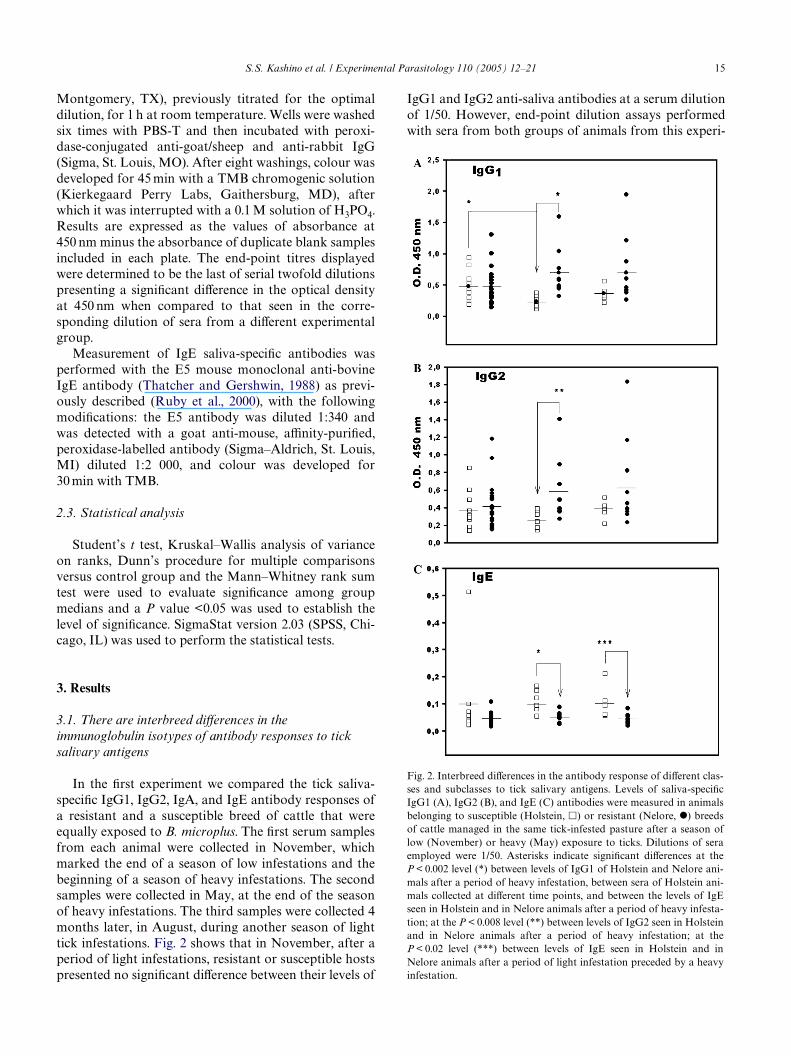
S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21 15
Montgomery, TX), previously titrated for the optimaldilution, for 1 h at room temperature. Wells were washedsix times with PBS-T and then incubated with peroxi-dase-conjugated anti-goat/sheep and anti-rabbit IgG(Sigma, St. Louis, MO). After eight washings, colour wasdeveloped for 45 min with a TMB chromogenic solution(Kierkegaard Perry Labs, Gaithersburg, MD), afterwhich it was interrupted with a 0.1 M solution of H3PO4.Results are expressed as the values of absorbance at450 nm minus the absorbance of duplicate blank samplesincluded in each plate. The end-point titres displayedwere determined to be the last of serial twofold dilutionspresenting a signiWcant diVerence in the optical densityat 450 nm when compared to that seen in the corre-sponding dilution of sera from a diVerent experimentalgroup.
Measurement of IgE saliva-speciWc antibodies wasperformed with the E5 mouse monoclonal anti-bovineIgE antibody (Thatcher and Gershwin, 1988) as previ-ously described (Ruby et al., 2000), with the followingmodiWcations: the E5 antibody was diluted 1:340 andwas detected with a goat anti-mouse, aYnity-puriWed,peroxidase-labelled antibody (Sigma–Aldrich, St. Louis,MI) diluted 1:2 000, and colour was developed for30 min with TMB.
2.3. Statistical analysis
Student’s t test, Kruskal–Wallis analysis of varianceon ranks, Dunn’s procedure for multiple comparisonsversus control group and the Mann–Whitney rank sumtest were used to evaluate signiWcance among groupmedians and a P value <0.05 was used to establish thelevel of signiWcance. SigmaStat version 2.03 (SPSS, Chi-cago, IL) was used to perform the statistical tests.
3. Results
3.1. There are interbreed diVerences in the immunoglobulin isotypes of antibody responses to tick salivary antigens
In the Wrst experiment we compared the tick saliva-speciWc IgG1, IgG2, IgA, and IgE antibody responses ofa resistant and a susceptible breed of cattle that wereequally exposed to B. microplus. The Wrst serum samplesfrom each animal were collected in November, whichmarked the end of a season of low infestations and thebeginning of a season of heavy infestations. The secondsamples were collected in May, at the end of the seasonof heavy infestations. The third samples were collected 4months later, in August, during another season of lighttick infestations. Fig. 2 shows that in November, after aperiod of light infestations, resistant or susceptible hostspresented no signiWcant diVerence between their levels of
IgG1 and IgG2 anti-saliva antibodies at a serum dilutionof 1/50. However, end-point dilution assays performedwith sera from both groups of animals from this experi-
Fig. 2. Interbreed diVerences in the antibody response of diVerent clas-ses and subclasses to tick salivary antigens. Levels of saliva-speciWcIgG1 (A), IgG2 (B), and IgE (C) antibodies were measured in animalsbelonging to susceptible (Holstein, �) or resistant (Nelore, I) breedsof cattle managed in the same tick-infested pasture after a season oflow (November) or heavy (May) exposure to ticks. Dilutions of seraemployed were 1/50. Asterisks indicate signiWcant diVerences at theP < 0.002 level (*) between levels of IgG1 of Holstein and Nelore ani-mals after a period of heavy infestation, between sera of Holstein ani-mals collected at diVerent time points, and between the levels of IgEseen in Holstein and in Nelore animals after a period of heavy infesta-tion; at the P < 0.008 level (**) between levels of IgG2 seen in Holsteinand in Nelore animals after a period of heavy infestation; at theP < 0.02 level (***) between levels of IgE seen in Holstein and inNelore animals after a period of light infestation preceded by a heavyinfestation.
16 S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21
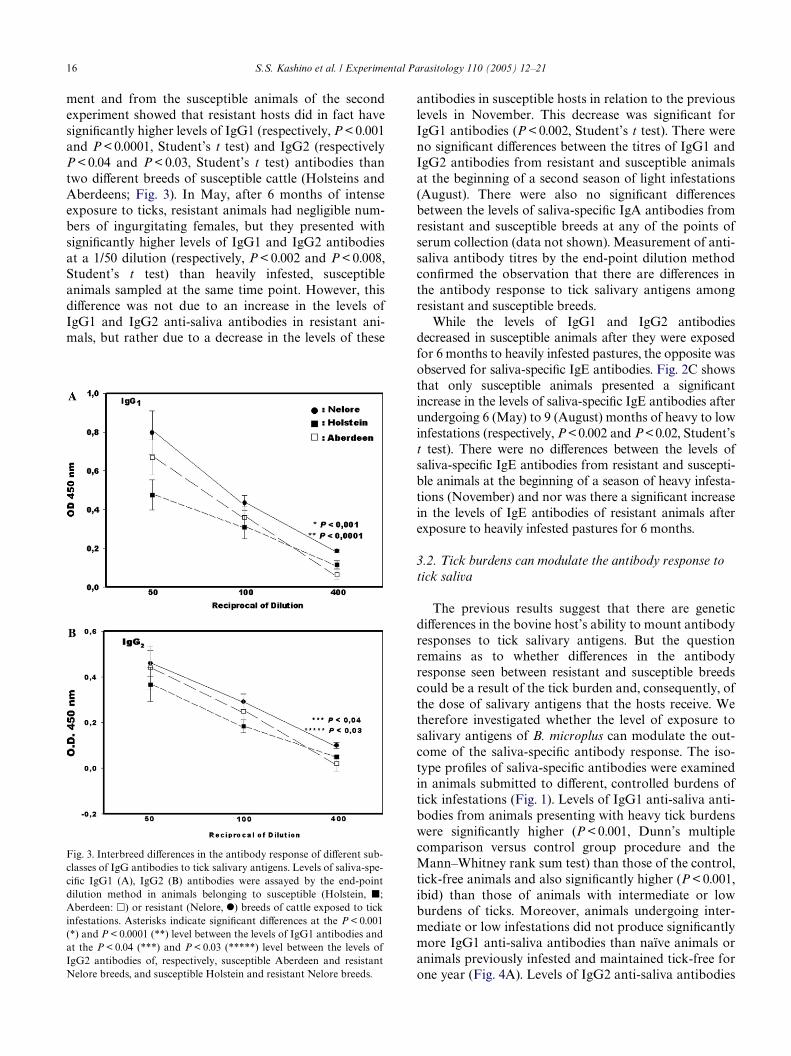
ment and from the susceptible animals of the secondexperiment showed that resistant hosts did in fact havesigniWcantly higher levels of IgG1 (respectively, P < 0.001and P < 0.0001, Student’s t test) and IgG2 (respectivelyP < 0.04 and P < 0.03, Student’s t test) antibodies thantwo diVerent breeds of susceptible cattle (Holsteins andAberdeens; Fig. 3). In May, after 6 months of intenseexposure to ticks, resistant animals had negligible num-bers of ingurgitating females, but they presented withsigniWcantly higher levels of IgG1 and IgG2 antibodiesat a 1/50 dilution (respectively, P < 0.002 and P < 0.008,Student’s t test) than heavily infested, susceptibleanimals sampled at the same time point. However, thisdiVerence was not due to an increase in the levels ofIgG1 and IgG2 anti-saliva antibodies in resistant ani-mals, but rather due to a decrease in the levels of these
Fig. 3. Interbreed diVerences in the antibody response of diVerent sub-classes of IgG antibodies to tick salivary antigens. Levels of saliva-spe-ciWc IgG1 (A), IgG2 (B) antibodies were assayed by the end-pointdilution method in animals belonging to susceptible (Holstein, �;Aberdeen: �) or resistant (Nelore, I) breeds of cattle exposed to tickinfestations. Asterisks indicate signiWcant diVerences at the P < 0.001(*) and P < 0.0001 (**) level between the levels of IgG1 antibodies andat the P < 0.04 (***) and P < 0.03 (*****) level between the levels ofIgG2 antibodies of, respectively, susceptible Aberdeen and resistantNelore breeds, and susceptible Holstein and resistant Nelore breeds.
antibodies in susceptible hosts in relation to the previouslevels in November. This decrease was signiWcant forIgG1 antibodies (P < 0.002, Student’s t test). There wereno signiWcant diVerences between the titres of IgG1 andIgG2 antibodies from resistant and susceptible animalsat the beginning of a second season of light infestations(August). There were also no signiWcant diVerencesbetween the levels of saliva-speciWc IgA antibodies fromresistant and susceptible breeds at any of the points ofserum collection (data not shown). Measurement of anti-saliva antibody titres by the end-point dilution methodconWrmed the observation that there are diVerences inthe antibody response to tick salivary antigens amongresistant and susceptible breeds.
While the levels of IgG1 and IgG2 antibodiesdecreased in susceptible animals after they were exposedfor 6 months to heavily infested pastures, the opposite wasobserved for saliva-speciWc IgE antibodies. Fig. 2C showsthat only susceptible animals presented a signiWcantincrease in the levels of saliva-speciWc IgE antibodies afterundergoing 6 (May) to 9 (August) months of heavy to lowinfestations (respectively, P < 0.002 and P< 0.02, Student’st test). There were no diVerences between the levels ofsaliva-speciWc IgE antibodies from resistant and suscepti-ble animals at the beginning of a season of heavy infesta-tions (November) and nor was there a signiWcant increasein the levels of IgE antibodies of resistant animals afterexposure to heavily infested pastures for 6 months.
3.2. Tick burdens can modulate the antibody response to tick saliva
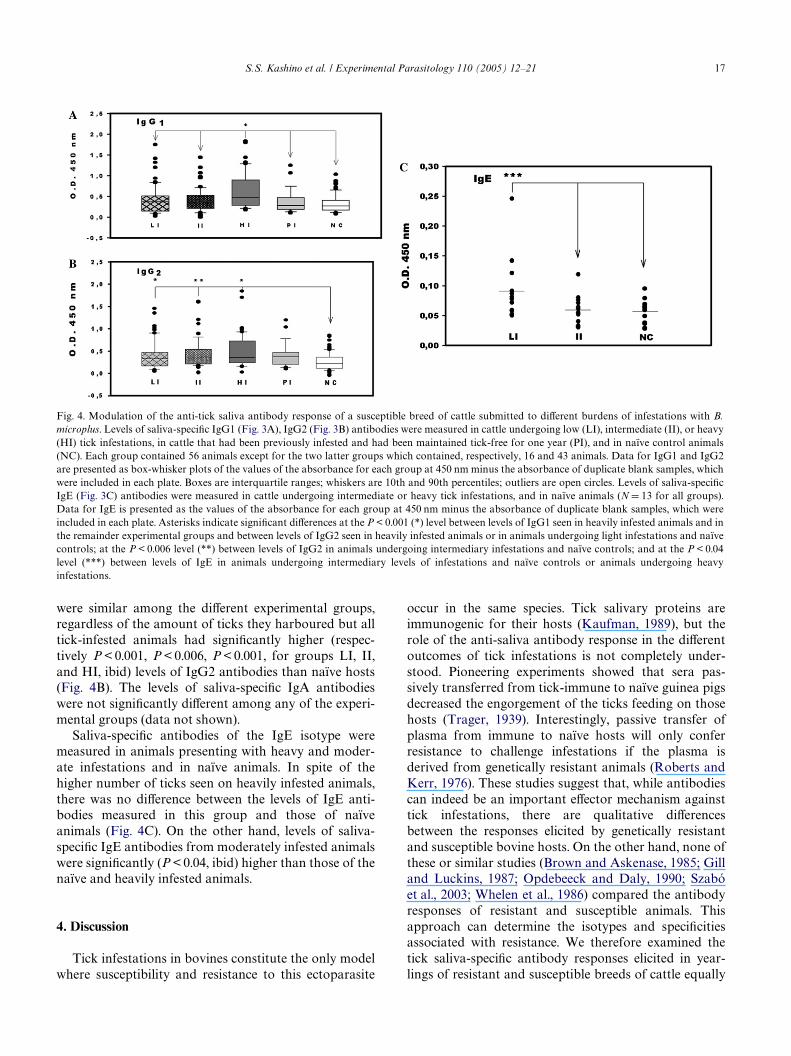
The previous results suggest that there are geneticdiVerences in the bovine host’s ability to mount antibodyresponses to tick salivary antigens. But the questionremains as to whether diVerences in the antibodyresponse seen between resistant and susceptible breedscould be a result of the tick burden and, consequently, ofthe dose of salivary antigens that the hosts receive. Wetherefore investigated whether the level of exposure tosalivary antigens of B. microplus can modulate the out-come of the saliva-speciWc antibody response. The iso-type proWles of saliva-speciWc antibodies were examinedin animals submitted to diVerent, controlled burdens oftick infestations (Fig. 1). Levels of IgG1 anti-saliva anti-bodies from animals presenting with heavy tick burdenswere signiWcantly higher (P < 0.001, Dunn’s multiplecomparison versus control group procedure and theMann–Whitney rank sum test) than those of the control,tick-free animals and also signiWcantly higher (P < 0.001,ibid) than those of animals with intermediate or lowburdens of ticks. Moreover, animals undergoing inter-mediate or low infestations did not produce signiWcantlymore IgG1 anti-saliva antibodies than naïve animals oranimals previously infested and maintained tick-free forone year (Fig. 4A). Levels of IgG2 anti-saliva antibodies
S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21 17
were similar among the diVerent experimental groups,regardless of the amount of ticks they harboured but alltick-infested animals had signiWcantly higher (respec-tively P < 0.001, P < 0.006, P < 0.001, for groups LI, II,and HI, ibid) levels of IgG2 antibodies than naïve hosts(Fig. 4B). The levels of saliva-speciWc IgA antibodieswere not signiWcantly diVerent among any of the experi-mental groups (data not shown).
Saliva-speciWc antibodies of the IgE isotype weremeasured in animals presenting with heavy and moder-ate infestations and in naïve animals. In spite of thehigher number of ticks seen on heavily infested animals,there was no diVerence between the levels of IgE anti-bodies measured in this group and those of naïveanimals (Fig. 4C). On the other hand, levels of saliva-speciWc IgE antibodies from moderately infested animalswere signiWcantly (P < 0.04, ibid) higher than those of thenaïve and heavily infested animals.
4. Discussion
Tick infestations in bovines constitute the only modelwhere susceptibility and resistance to this ectoparasite
occur in the same species. Tick salivary proteins areimmunogenic for their hosts (Kaufman, 1989), but therole of the anti-saliva antibody response in the diVerentoutcomes of tick infestations is not completely under-stood. Pioneering experiments showed that sera pas-sively transferred from tick-immune to naïve guinea pigsdecreased the engorgement of the ticks feeding on thosehosts (Trager, 1939). Interestingly, passive transfer ofplasma from immune to naïve hosts will only conferresistance to challenge infestations if the plasma isderived from genetically resistant animals (Roberts andKerr, 1976). These studies suggest that, while antibodiescan indeed be an important eVector mechanism againsttick infestations, there are qualitative diVerencesbetween the responses elicited by genetically resistantand susceptible bovine hosts. On the other hand, none ofthese or similar studies (Brown and Askenase, 1985; Gilland Luckins, 1987; Opdebeeck and Daly, 1990; Szabóet al., 2003; Whelen et al., 1986) compared the antibodyresponses of resistant and susceptible animals. Thisapproach can determine the isotypes and speciWcitiesassociated with resistance. We therefore examined thetick saliva-speciWc antibody responses elicited in year-lings of resistant and susceptible breeds of cattle equally
Fig. 4. Modulation of the anti-tick saliva antibody response of a susceptible breed of cattle submitted to diVerent burdens of infestations with B.microplus. Levels of saliva-speciWc IgG1 (Fig. 3A), IgG2 (Fig. 3B) antibodies were measured in cattle undergoing low (LI), intermediate (II), or heavy(HI) tick infestations, in cattle that had been previously infested and had been maintained tick-free for one year (PI), and in naïve control animals(NC). Each group contained 56 animals except for the two latter groups which contained, respectively, 16 and 43 animals. Data for IgG1 and IgG2are presented as box-whisker plots of the values of the absorbance for each group at 450 nm minus the absorbance of duplicate blank samples, whichwere included in each plate. Boxes are interquartile ranges; whiskers are 10th and 90th percentiles; outliers are open circles. Levels of saliva-speciWcIgE (Fig. 3C) antibodies were measured in cattle undergoing intermediate or heavy tick infestations, and in naïve animals (N D 13 for all groups).Data for IgE is presented as the values of the absorbance for each group at 450 nm minus the absorbance of duplicate blank samples, which wereincluded in each plate. Asterisks indicate signiWcant diVerences at the P < 0.001 (*) level between levels of IgG1 seen in heavily infested animals and inthe remainder experimental groups and between levels of IgG2 seen in heavily infested animals or in animals undergoing light infestations and naïvecontrols; at the P < 0.006 level (**) between levels of IgG2 in animals undergoing intermediary infestations and naïve controls; and at the P < 0.04level (***) between levels of IgE in animals undergoing intermediary levels of infestations and naïve controls or animals undergoing heavyinfestations.

18 S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21
exposed to B. microplus. To determine if isotype proWlesdiVer in resistant animals because they receive a smallerdose of saliva, we also examined responses in a suscepti-ble breed of cattle exposed to diVerent levels of infesta-tions and, hence, doses of saliva.
Susceptible animals underwent 15- to 100-foldheavier infestations than resistant breeds, which, in turn,had negligible tick counts. End-point titration assaysdetermined that resistant animals presented with signiW-cantly higher levels of IgG1 and IgG2 anti-tick salivaantibodies than two diVerent breeds of susceptible cattle.This diVerence increased after a period of 6 months ofexposure on heavily infested pastures: the tick-resistantbreed exhibited signiWcantly higher levels of IgG1 andIgG2 tick saliva-speciWc antibodies when compared tothose of a susceptible breed managed in the same pas-ture. This diVerence was not due to an increase in the lev-els of saliva-speciWc antibodies in resistant animals, butrather to a decrease in the levels of IgG1 and IgG2saliva-speciWc antibodies in susceptible animals. Theseresults are not explained by interbreed diVerences in thelevels of immunoglobulins (Rechav, 1987) because at thebeginning of the experiment both breeds presented withsimilar levels of saliva-speciWc antibodies. They are inagreement with observations made in Holstein cattleheavily infested with another species of tick, Hyalommamarginatum marginatum (Sahibi et al., 1998). Thedecrease in anti-saliva antibody levels seen in susceptibleanimals that have undergone heavy infestations couldhave several explanations. First, it could result from sup-pression of the immune response by tick saliva. Indeed,cattle exposed to prolonged infestations with Hyalommaasiaticum asiaticum become desensitised to this parasite(Berdyev, A. and Khudainazarova, S.N., cited in Wikeland Allen, 1982). Cattle heavily infested with B. micro-plus show diminished responses to an extraneous antigenwhen compared to controls (Inokuma et al., 1993).Saliva from the tick Rhipicephalus sanguineus impairsthe diVerentiation and maturation of murine dendriticcells (Ferreira, B.R., personal communication), whichinduce immune responses and aVect the growth andfunction of B cells, including in the bovine species(Pinchuk et al., 2003).
Another explanation is that immunoglobulin IgG-binding proteins (IGBPs) present in the tick’s saliva aretargeting and promoting the clearance of this serumcomponent. IGBPs have been described in R. appendi-culatus (Wang and Nuttall, 1994) and salivary glands ofmale B. microplus abundantly express a similar gene (DeMiranda Santos et al., 2004). It is reasonable to speculatethat heavy infestations can deliver a biologically signiW-cant amount of IGBPs due to the large quantity of salivathat susceptible animals receive (Sauer et al., 2000) whentick infestations are not controlled, the case in the pres-ent experiment. It is also possible that IGBP-coatedimmune complexes can negatively modulate host anti-
gen presenting cells. Furthermore, diVerent allotypes ofbovine immunoglobulins may be more or less amenableto the eVects of tick IGBPs, as has been shown for simi-lar proteins from other pathogens (Bastida-Corcueraet al., 1999a,b, 2000). The higher levels of anti-saliva IgGantibodies seen in resistant cattle may, however, be of noconsequence to the tick and simply reXect a more vigor-ous immune response. While our data do not providedeWnite proof for a protective role of IgG anti-salivaantibodies, the presence of an IgG-binding protein in thesaliva of B. microplus is, however, a strong indicationthat antibodies can indeed be deleterious for ticks. Thispoint is further emphasized by recent work showing thatsaliva from the tick Ixodes ricinus inhibits secretion ofIL-10 by activated B cells as well as proliferation of Bcells stimulated with LPS or Borrelia burgdorferi outersurface protein (Hannier et al., 2003, 2004).
Finally, variability among the genes coding for compo-nents of the host immune system could aVect the outcomeof tick infestations. Indeed, heritable polymorphismswithin the regulatory or structural elements of genes of theimmune system are present in the bovine species (Grosseet al., 1999; Heaton et al., 2001). One polymorphic genethat could be especially relevant for the present results isthe Fc receptor (FcRn) �-chain gene (FCGRT), the hapl-otypes of which are associated with diVerent rates of clear-ance of serum IgG in cattle (Laegreid et al., 2002).
It should be pointed out that diVerential recognition oftick salivary antigens by antibodies of resistant and sus-ceptible hosts can account for diVerences in their levels.On the other hand, regardless of the status of their pheno-types for infestations, all hosts apparently receive thesame repertoire of salivary antigens and the hosts thatmaintain higher levels of anti-saliva antibodies may do sobecause they recognize a broader range of antigens. Wetherefore believe that, while the antibody responses toindividual antigens should also be characterized, it is stillnecessary to examine and describe the antibody responseto whole saliva because this represents the outcome in thetrue setting of the parasite–host interface.
In relation to the comparative levels of saliva-speciWcIgE antibodies in resistant and susceptible breeds, theopposite was observed: after a season of heavy infesta-tions, levels were signiWcantly higher in susceptible thanin resistant animals. These results are in agreement withdata from studies in mice infested with the tick I. ricinus.Like most breeds of B. taurus taurus cattle, all strains ofmurine hosts remain susceptible to ticks after repeatedinfestations. Furthermore, tick infestations polarise themurine immune response towards a Th2 phenotype(Christe et al., 1999; Ferreira and Silva, 1999) with agradual increase in total IgE (Christe et al., 1999).
IgE anti-saliva antibodies may be lower in resistantcattle because they have been consumed in supposedlyprotective, IgE-dependent immediate hypersensitivityskin reactions, which have been shown by other

S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21 19
investigators to be more intense in resistant cattle (Wil-ladsen et al., 1979). We and others have performed skintests with extracts of larvae on these and on two othersets of experimental animals and have found no signiW-
cant diVerences in the intensity of the immediatehypersensitivity reactions mounted by these two groups(Bechara et al., 2000). Furthermore, we found no signiW-
cant diVerences in the numbers of mast cells, basophilsor eosinophils present at the site of immediate skinreactions in these same experimental animals. The avail-ability of a much greater quantity of circulating, IgE-crosslinking antigen (tick saliva) in susceptible animals,a consequence of the heavier infestations, may result, ifanything, in a greater consumption of IgE in these hoststhan in resistant animals.
It is noteworthy that, in contrast to what occurs withIgG at the tick–host interface, an IgE-binding proteinhas not been described in ticks. On the other hand, manyspecies of ticks do have histamine-binding proteins (Pae-sen et al., 2000) and, of relevance to this study, salivaryglands of male adults of B. microplus abundantly expressa similar gene (De Miranda Santos et al., 2004). The dis-crepancy between the apparent unimportance of IgE inresistance to ticks and the presence of histamine-bindingproteins in tick saliva could be explained if the release ofhistamine does not depend on cytophilic antibodies fordegranulation of mast cells and basophiles, but, forexample, on the chemokine MCP-1 (Campbell et al.,1999). Indeed, basophile-dependent, cutaneous hyper-sensitivity does not involve antibodies and it may be animportant eVector mechanism in mediating resistance toticks (Falcone et al., 2001).
Dose of antigen determines the class of immunityinduced (Constant and Bottomly, 1997; Grakoui et al.,1999; Guery et al., 1996; Menon and Bretscher, 1998),therefore we next asked, notwithstanding saliva-mediatedimmunosuppression, whether susceptible animals do notdevelop appropriate immune responses because exposureto salivary antigens is high. This situation could favour aTh2 proWle, which is prominent in the tick-susceptiblemurine host (Christe et al., 1999; Ferreira and Silva,1999). We examined the levels of IgG1, IgG2, IgA, andIgE anti-saliva antibodies in Aberdeen cattle, a tick-sus-ceptible breed, exposed to controlled levels of naturalinfestations, and in naïve, tick-free controls. Only heavilyinfested animals produced signiWcantly higher levels ofIgG1 antibodies than naïve, tick-free animals. Regardinganti-saliva IgG2, all experimental groups, including thosethat had been previously infested and were tick-free for 1year, had similar levels of antibodies and these were allsigniWcantly higher than those in naïve animals. Produc-tion and/or clearance of IgG2 antibodies were thus notaVected by the size of the tick burden.
Levels of IgE saliva-speciWc antibodies were signiW-cantly higher in animals undergoing intermediate levelsof tick infestations when compared with naïve controls
and with heavily infested animals. This Wnding, however,does not signify that IgE antibodies are mediating pro-tection for the following reasons: Wrst, the lower level ofinfestation was not maintained by an immune responsebut by acaricides; second, the serum samples from theseanimals had been collected after a recent rise in the levelof exposure to ticks (see Fig. 1) and the lower levels ofIgE antibodies seen in heavily infested animals may bedue to their consumption by the large dose of saliva thatthey had been exposed to; third, in spite of higher IgEantibody levels, tick burdens were heavier in the suscep-tible animals of the previous experiment when comparedwith resistant hosts.
The characteristics of bovine T helper subsets have notbeen Wrmly established. Our results indicate that the bur-den of ticks and, consequently, the dose of antigen inocu-lated into the host modulate the proWles of saliva-speciWcantibodies in a susceptible breed: heavy exposure drivesthe response towards the IgG1 isotype proWle, tentativelyassociated with TH2-type responses in bovines (Estes andBrown, 2002; Mulcahy et al., 1998). These results agreewith a study showing that, contrary to a resistant breedof cattle, a susceptible breed mounted a delayed cutane-ous reactions to tick antigens (Bechara et al., 2000), aresponse associated with a Th1 proWle.
In conclusion, the present results show that there areinterbreed diVerences in the proWles of anti-tick salivaantibodies elicited in cattle. After heavy tick infestationslevels of anti-saliva antibodies of the IgG1 and IgG2 iso-type decrease in susceptible but not in resistant bovines,suggesting that these isotypes may mediate relevanteVector mechanisms. Furthermore, IgE antibodiesincrease only in susceptible hosts, indicating that thisisotype may not have a role in controlling tick infesta-tions. Finally, the results show that the burden of tickinfestations modulates the proWle of IgG1 antibodies, anisotype possibly associated with a TH2 proWle in cattle.In view of these results, factors such as the speciWcityand aYnity of antibodies for individual salivary anti-gens, and genetic polymorphisms and post-translationalmodiWcations of IgG1 and IgG2 that aVect their clear-ance, their binding to IGBPs and their eVector functionsare being examined.
Acknowledgments
This work was supported by the National ResearchCouncil (Conselho Nacional de Desenvolvimento Cient-íWco e Tecnológico—CNPq, Grant PCOPG-52736/99-0),by the Programme for Centres of Excellence (Programade Núcleos de Excelência—PRONEX, Grant 66 1140/1998-9) of the Brazilian Ministry for Science and Technol-ogy, and by the Programme for Technological Develop-ment of Agriculture (Programa de DesenvolvimentoTecnológico da Agropecuária Brasileira—PRODETAB,

20 S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21
Grants 021/98 and 019/01) of the World Bank. Animals ofthe Nelore breed were provided by the Brazilian Associa-tion of Breeders of Zebu Cattle (Associação Brasileira deCriadores de Zebuínos—ABCZ). S.S.K. was a post-doc-toral fellow from CNPq (Grant 301204/2000-4). We thankDr. João Santana da Silva, Dr. Gervasio H. Bechara, andDr. Beatriz R. Ferreira for support and helpful comments.
References
Anonymous, 1995. In: Yearbook Production, 1994, vol. 48. Food andAgriculture Organisation, Rome.
Bastida-Corcuera, F.D., Nielsen, K.H., Corbeil, L.B., 1999a. Binding ofbovine IgG2a and IgG2b allotypes to protein A, protein G, andHaemophilus somnus IgBPs. Veterinary Immunology Immunopa-thology 71, 143–149.
Bastida-Corcuera, F.D., Butler, J.E., Yahiro, S., Corbeil, L.B., 1999b.DiVerential complement activation by bovine IgG2 allotypes. Vet-erinary Immunology Immunopathology 71, 115–123.
Bastida-Corcuera, F., Butler, E., Heyermann, H., Thomford, J.W.,Corbeil, L.B., 2000. Tritrichomonas foetus extracellular cysteineproteinase cleavage of bovine IgG2 allotypes. Journal of Parasitol-ogy 86, 328–332.
Bechara, G.H., Morelli Jr., J., Szabo, M.P., 2000. Skin test and tickimmune status in susceptible and resistant cattle in Brazil. Annalsof the New York Academy of Sciences 916, 570–575.
Brown, S.J., Askenase, P.W., 1985. Amblyomma americanum: require-ment for host Fc receptors in antibody-mediated acquired immuneresistance to ticks. Experimental Parasitology 59, 248–256.
Campbell, E.M., Charo, I.F., Kunkel, S.L., Strieter, R.M., Boring, L.,Gosling, J., Lukacs, N.W., 1999. Monocyte chemoattractant pro-tein-1 mediates cockroach allergen-induced bronchial hyperreactiv-ity in normal but not CCR2¡/¡ mice: the role of mast cells.Journal of Immunology 163, 2160–2167.
Casadevall, A., Profski, L., 2003. Antibody-mediated regulation of cel-lular immunity and the inXammatory response. Trends in Immu-nology 24, 474–478.
Christe, M., Rutti, B., Brossard, M., 1999. InXuence of the genetic back-ground and parasite load of mice on the immune response developedagainst nymphs of Ixodes ricinus. Parasitology Research 85, 557–561.
Constant, S.L., Bottomly, K., 1997. Induction of Th1 and Th2 CD4+ Tcell responses: the alternative approaches. Annual Review ofImmunology 15, 297–322.
De Miranda Santos, I.K., Valenzuela, J.G., Ribeiro, J.M., De Castro,M., Costa, J.N., Costa, A.M., Da Silva, E.R., Neto, O.B., Rocha, C.,DaVre, S., Ferreira, B.R., Da Silva, J.S., Szabo, M.P., Bechara, G.H.,2004. Gene discovery in Boophilus microplus, the cattle tick: thetranscriptomes of ovaries, salivary glands, and hemocytes. Annalsof the New York Academy of Sciences 1026, 242–246.
Estes, D.M., Brown, W.C., 2002. Type 1 and type 2 responses in regula-tion of Ig isotype expression in cattle. Veterinary ImmunologyImmunopathology 90, 1–10.
Falcone, F.H., Pritchard, D.I., Gibbs, B.F., 2001. Do basophils play a rolein immunity against parasites? Trends in Parasitology 17, 126–129.
Ferreira, B.R., Silva, J.S., 1999. Successive tick infestations selectivelypromote a T-helper 2 cytokine proWle in mice. Immunology 96,434–439.
George, J.E., Osburn, R.L., Wikel, S.K., 1985. Acquisition and expres-sion of resistance by Bos indicus and Bos indicus £ Bos taurus calvesto Amblyomma americanum infestation. Journal of Parasitology 71,174–182.
Gill, H.S., Luckins, A.G., 1987. Hyalomma anatolicum: the role ofhumoral factors in the acquisition of host resistance. ExperimentalParasitology 64, 430–437.
Grakoui, A., Donermeyer, D.L., Kanagawa, O., Murphy, K.M., Allen,P.M., 1999. TCR independent pathways mediate the eVects of anti-gen dose and altered peptide ligands on Th cell polarization. Jour-nal of Immunology 162, 1923–1930.
Grosse, W.M., Kappes, S.M., Laegreid, W.W., Keele, J.W., Chitko-McKown, C.G., Heaton, M.P., 1999. Single nucleotide polymor-phism (SNP) discovery and linkage mapping of bovine cytokinegenes. Mammalian Genome 10, 1062–1069.
Guery, J.C., Galbiati, F., Smiroldo, S., Adorini, L., 1996. Selectivedevelopment of T helper (Th)2 cells induced by continuous admin-istration of low dose soluble proteins to normal and beta(2)-micro-globulin-deWcient BALB/c mice. Journal of Experimental Medicine183, 485–497.
Hannier, S., Liversidge, J., Sternberg, J.M., Bowman, A.S., 2003. Ixodesricinus tick salivary gland extract inhibits IL-10 secretion andCD69 expression by mitogen-stimulated murine splenocytes andinduces hyporesponsiveness in B lymphocytes. Parasite Immunol-ogy 25, 27–37.
Hannier, S., Liversidge, J., Sternberg, J.M., Bowman, A.S., 2004. Char-acterization of the B-cell inhibitory protein factor in Ixodes ricinustick saliva: a potential role in enhanced Borrelia burgdoferi trans-mission. Immunology 113, 401–408.
Heaton, M.P., Grosse, W.M., Kappes, S.M., Keele, J.W., Chitko-McK-own, C.G., CardiV, L.V., Braun, A., Little, D.P., Laegreid, W.W.,2001. Estimation of DNA sequence diversity in bovine cytokinegenes. Mammalian Genome 12, 32–37.
Inokuma, H., Kerlin, R.L., Kemp, D.H., Willadsen, P., 1993. EVects ofcattle tick (Boophilus microplus) infestation on the bovine immunesystem. Veterinary Parasitology 47, 107–118.
Kaufman, W.R., 1989. Tick–host interactions: a synthesis of currentconcepts. Parasitology Today 52, 47–56.
Laegreid, W.W., Heaton, M.P., Keen, J.E., Grosse, W.M., Chitko-McK-own, C.G., Smith, T.P.L., Keele, J.W., Bennett, G.L., Besser, T.E.,2002. Association of bovine neonatal Fc receptor �-chain (FCGRT)haplotypes with serum IgG concentration in newborn calves. Mam-malian Genome 13, 704–710.
Mattioli, R.C., Cassma, M., 1995. Comparison of characteristics of lifecycle in female ticks collected on N’Dama and Zebu cattle. TropicalAnimal Health and Production 27, 150–154.
Mattioli, R.C., 1998. Comment on “A possible explanation of theapparent breed-related resistance in cattle to bont tick (Ambly-omma hebraeum) infestations. Veterinary Parasitology 1996, 67,275–279.” Veterinary Parasitology 79, 263–264.
Mattioli, R.C., Pandey, V.S., Murray, M., Fitzpatrick, J.L., 2000. Immu-nogenetic inXuences on tick resistance in African cattle with partic-ular reference to trypanotolerant N’Dama (Bos taurus) andtrypanosusceptible Gobra zebu (Bos indicus) cattle. Acta Tropica75, 263–277.
Meltzer, M.I., 1996. A possible explanation of the apparent breed-related resistance in cattle to bont tick (Amblyomma hebraeum)infestations. Veterinary Parasitology 67, 275–279.
Menon, J.N., Bretscher, P.A., 1998. Parasite dose determines the Th1/Th2 nature of the response to Leishmania major independently ofinfection route and strain of host or parasite. European Journal ofImmunology 28, 4020–4028.
Mulcahy, G., O’Connor, F., McGonigle, S., Dowd, A., Clery, D.G.,Andrews, S.J., Dalton, J.P., 1998. Correlation of speciWc antibodytitre and avidity with protection in cattle immunized against Fasci-ola hepatica. Vaccine 16, 932–939.
Norval, R.H., 1992. Host immunity to infestation with Amblyommahebraeum. Insect Science and Applications 13, 489–494.
Opdebeeck, J.P., Daly, K.E., 1990. Immune responses of infested andvaccinated Hereford cattle to antigens of the cattle tick Boophilusmicroplus. Veterinary Immunology Immunopathology 25, 99–108.
Paesen, G.C., Adams, P.L., Nuttall, P.A., Stuart, D.L., 2000. Tick hista-mine-binding proteins: lipocalins with a second binding cavity. Bio-chimica et Biophysica Acta (BBA) 1482, 92–101.

S.S. Kashino et al. / Experimental Parasitology 110 (2005) 12–21 21
Pinchuk, L.M., Boyd, B.L., Kruger, E.F., Roditi, I., Furger, A., 2003.Bovine dendritic cells generated from monocytes and bone marrowprogenitors regulate immunoglobulin production in peripheralblood B cells. Comparative Immunology, Microbiology and Infec-tious Diseases 26, 233–249.
Pruett, J.H., 1999. Immunological control of arthropod ectoparasites—a review. International Journal of Parasitology 29, 25–32.
Ramachandra, R.N., Wikel, S.K., 1995. EVects of Dermacentor ander-soni (Acari: Ixodidae) salivary gland extracts on Bos indicus and B.taurus lymphocytes and macrophages: in vitro cytokine elaborationand lymphocyte blastogenesis. Journal of Medical Entomology 32,338–345.
Rechav, Y., 1987. Resistance of Brahman and Hereford cattle to Afri-can ticks with reference to serum gammaglobulin levels and bloodcomposition. Experimental and Applied Acarology 3, 219–232.
Ribeiro, J.M.C., 1995. How ticks make a living. Parasitology Today 11,91–93.
Roberts, J.A., Kerr, J.D., 1976. Boophilus microplus: passive transfer ofresistance in cattle. Journal of Parasitology 62, 485–488.
Ruby, K.W., GriYth, R.W., Gershwin, L.J., Kaeberle, M.L., 2000. Hae-mophilus somnus-induced IgE in calves vaccinated with commercialmonovalent H. somnus bacterins. Veterinary Microbiology 76, 373–383.
Sahibi, H., Rhalem, A., Tikki, N., 1998. Comparison of eVects of lowand high tick infestations on acquired cattle tick resistance: Hya-lomma marginatum marginatum. Parasite 5, 69–74.
Sauer, J.R., Essenberg, R.C., Bowman, A.S., 2000. Salivary glands inixodid ticks: control and mechanism of secretion. Journal of InsectPhysiology 46, 1069–1078.
Szabó, M.P.J., Aoki, V.L., Sanches, F.P.S., Aquino, L.P.T.C.T., Garcia,M.V., Machado, R.Z., Bechara, GH., 2003. Antibody and bloodleukocyte response in Rhipicephalus sanguineus (Latreille, 1806)
tick-infested dogs and guinea pigs. Veterinary Parasitology 115,49–59.
Thatcher, E.F., Gershwin, L.J., 1988. Generation and characteriza-tion of murine monoclonal antibodies speciWc for bovine immu-noglobulin E. Veterinary Immunology Immunopathology 18,53–66.
Trager, W., 1939. Acquired immunity to ticks. Journal of Parasitology25, 57–81.
Turni, C., Lee, R.P., Jackson, L.A., 2002. EVect of salivary glandextracts from the tick, Boophilus microplus, on leucocytes fromBrahman and Hereford cattle. Parasite Immunology 24, 355–361.
Utech, K.B.W., Wharton, R.H., Kerr, J.D., 1978. Resistance to Boophi-lus microplus (Canestrini) in diVerent breeds of cattle. AustralianJournal of Agricultural Research 29, 885–895.
Wambura, P.N., Gwakisa, P.S., Silayo, R.S., Rugaimukamu, E.A., 1998.Breed-associated resistance to tick infestations in Bos indicus andtheir crosses with Bos taurus. Veterinary Parasitology 77, 63–70.
Wang, H., Nuttall, P.A., 1994. Excretion of host immunoglobulin intick saliva and detection of IgG-binding proteins in tick haemol-ymph and salivary glands. Parasitology 109, 525–530.
Whelen, A.C., Richardson, L.K., Wikel, S.K., 1986. Dot-ELISA assess-ment of guinea pig antibody responses to repeated Dermacentorandersoni infestations. Journal of Parasitology 72, 155–162.
Wikel, S.K., 1996. Host immunity to ticks. Annual Review of Entomol-ogy 41, 1–22.
Wikel, S.K., Allen, J.R., 1982. Immunological basis of host resistance toticks. In: Obenchain, F.D., Gialun, R. (Eds.), Physiology of Ticks.Pergamon Press, Oxford, p. 172.
Willadsen, P., Wood, G.M., Riding, G.A., 1979. The relation betweenskin histamine concentration, histamine sensitivity and the resis-tance of cattle to the tick, Boophilus microplus. Zeitschrift fur Paras-itenkunde 59, 87–93.












