
Angiotensin receptor-like immunoreactivity in adult brain white matter astrocytes and...
16
Angiotensin Receptor-Like Immunoreactivity in Adult Brain White Matter Astrocytes and Oligodendrocytes DAVID J. FOGARTY AND CARLOS MATUTE* Department of Neurosciences, Faculty of Medicine, University of the Basque Country, Leioa, Spain KEY WORDS AT 1 ; AT 2 ; macroglia; rat; bovine; immunohistochemistry ABSTRACT Most of the physiological effects of brain angiotensins are currently believed to be mediated by angiotensin receptors located principally on neurons. How- ever, numerous studies in vitro have demonstrated the presence of functional angioten- sin receptors on brain astrocytes, raising the possibility that glial cells may also partic- ipate in mediating the effects of the central renin–angiotensin system. Nevertheless, it is uncertain whether these cells in situ express angiotensin receptors, raising questions about the physiological significance of results observed in cell cultures. We have exam- ined the distribution of angiotensin receptor-like immunoreactivity in glial cells in white matter tracts in the adult CNS, using a panel of antisera to the AT 1 and AT 2 angiotensin receptors. Antiserum preadsorption and/or Western blot demonstrated the specificity of the antisera in brain tissue. In immunohistochemical experiments, the AT 1 antisera selectively labeled AT 1 -expressing neurons in the piriform cortex, whereas the AT 2 antiserum stained cells in the trigeminal motor nucleus, these being nuclei known to express AT 1 and AT 2 receptors, respectively. Using double-label immunohistochemistry, we observed AT 1 - and AT 2 -immunoreactive astrocytes and oligodendrocytes in white matter tracts, which include the rat cerebellar white matter, periventricular white matter, and optic nerve, in addition to the bovine corpus callosum and human subcor- tical white matter. In contrast, astrocytes in the gray matter region of the cerebral cortex were not found to be angiotensin receptor-like immunoreactive. These results demon- strate the presence of AT 1 and/or AT 2 angiotensin receptor-like immunoreactivity in brain white matter macroglial cells in situ and support the idea that glial cells may play a more important role in the central renin-angiotensin system than previously thought. GLIA 35:131–146, 2001. © 2001 Wiley-Liss, Inc. INTRODUCTION The renin-angiotensin system (RAS) is, in phyloge- netic terms, one of the oldest hormone systems, sub- serving vital functions such as the regulation of blood pressure and volume and hydroelectrolytic homeosta- sis. The system includes a group of physiologically ac- tive peptides that mediate their effects through specific angiotensin receptors. These receptors are located throughout the body in a wide variety of species, in organs and tissues such as the vasculature, the heart, the kidney, the adrenal gland, and the brain (see re- view by Fitzsimons, 1998). Two principal subtypes of angiotensin receptors, termed the AT 1 and AT 2 recep- tors, have been defined on the basis of electrophysio- Grant sponsor: University of the Basque Country; Grant number: G-14/98; Grant sponsor: Ministry of Education and Culture; Grant number: PM-97/45. *Correspondence to: Carlos Matute, Department of Neurosciences, Faculty of Medicine, University of the Basque Country, E-48940, Leioa, Spain. E-mail: [email protected] Received 13 February 2001; Accepted 22 May 2001 Published online 00 Month 2001 GLIA 35:131–146 (2001) © 2001 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Angiotensin receptor-like immunoreactivity in adult brain white matter astrocytes and...

Angiotensin Receptor-LikeImmunoreactivity in Adult Brain
White Matter Astrocytes andOligodendrocytes
DAVID J. FOGARTY AND CARLOS MATUTE*Department of Neurosciences, Faculty of Medicine, University of the Basque Country,
Leioa, Spain
KEY WORDS AT1; AT2; macroglia; rat; bovine; immunohistochemistry
ABSTRACT Most of the physiological effects of brain angiotensins are currentlybelieved to be mediated by angiotensin receptors located principally on neurons. How-ever, numerous studies in vitro have demonstrated the presence of functional angioten-sin receptors on brain astrocytes, raising the possibility that glial cells may also partic-ipate in mediating the effects of the central renin–angiotensin system. Nevertheless, itis uncertain whether these cells in situ express angiotensin receptors, raising questionsabout the physiological significance of results observed in cell cultures. We have exam-ined the distribution of angiotensin receptor-like immunoreactivity in glial cells in whitematter tracts in the adult CNS, using a panel of antisera to the AT1 and AT2 angiotensinreceptors. Antiserum preadsorption and/or Western blot demonstrated the specificity ofthe antisera in brain tissue. In immunohistochemical experiments, the AT1 antiseraselectively labeled AT1-expressing neurons in the piriform cortex, whereas the AT2antiserum stained cells in the trigeminal motor nucleus, these being nuclei known toexpress AT1 and AT2 receptors, respectively. Using double-label immunohistochemistry,we observed AT1- and AT2-immunoreactive astrocytes and oligodendrocytes in whitematter tracts, which include the rat cerebellar white matter, periventricular whitematter, and optic nerve, in addition to the bovine corpus callosum and human subcor-tical white matter. In contrast, astrocytes in the gray matter region of the cerebral cortexwere not found to be angiotensin receptor-like immunoreactive. These results demon-strate the presence of AT1 and/or AT2 angiotensin receptor-like immunoreactivity inbrain white matter macroglial cells in situ and support the idea that glial cells may playa more important role in the central renin-angiotensin system than previously thought.GLIA 35:131–146, 2001. © 2001 Wiley-Liss, Inc.
INTRODUCTION
The renin-angiotensin system (RAS) is, in phyloge-netic terms, one of the oldest hormone systems, sub-serving vital functions such as the regulation of bloodpressure and volume and hydroelectrolytic homeosta-sis. The system includes a group of physiologically ac-tive peptides that mediate their effects through specificangiotensin receptors. These receptors are locatedthroughout the body in a wide variety of species, inorgans and tissues such as the vasculature, the heart,
the kidney, the adrenal gland, and the brain (see re-view by Fitzsimons, 1998). Two principal subtypes ofangiotensin receptors, termed the AT1 and AT2 recep-tors, have been defined on the basis of electrophysio-
Grant sponsor: University of the Basque Country; Grant number: G-14/98;Grant sponsor: Ministry of Education and Culture; Grant number: PM-97/45.
*Correspondence to: Carlos Matute, Department of Neurosciences, Facultyof Medicine, University of the Basque Country, E-48940, Leioa, Spain.E-mail: [email protected]
Received 13 February 2001; Accepted 22 May 2001
Published online 00 Month 2001
GLIA 35:131–146 (2001)
© 2001 Wiley-Liss, Inc.

logical, pharmacological, and molecular biological stud-ies; the AT1 receptor is antagonized by the imidazolederivative losartan, while the AT2 subtype is antago-nized by PD123,319, a tetrahydroimidazopyridine.Molecular cloning has demonstrated the identity ofthese two receptor subtypes which both belong to theseven transmembrane superfamily of receptors and aretypically coupled to G-proteins (Stroth and Unger,1999).
In recent years, increased attention has focused onthe angiotensin signaling system within the brain (re-viewed by Allen et al., 1998). Intracerebroventricularinjection of angiotensin II results in physiological re-sponses, such as compulsive drinking, increased bloodpressure, and altered cognitive performance (Wrightand Harding, 1997). Indeed, localization studies em-ploying such techniques as autoradiography, in situhybridization, and immunohistochemistry, have shownthe presence of angiotensin receptors in neurons inbrain nuclei that regulate cardiovascular and bodyfluid homeostatic functions (reviewed in Lenkei et al.,1997). However, the expression of brain angiotensinreceptors is not limited to cardiovascular and fluidhomeostatic-related nuclei. Thus, angiotensin receptorexpression has been detected in a variety of other brainregions, including the cerebral cortex, the hippocam-pus, the optic nerve, the colliculi, the olfactory nucleus,and the cerebellum (Phillips et al., 1993; Reagan et al.,1994; Lenkei et al., 1997). These locations suggest thatangiotensins may play novel roles unrelated to thoseclassically associated with the RAS.
Various lines of evidence point to a role for brain glialcells in the central RAS. Thus, the principal source ofbrain angiotensinogen, the angiotensin peptide precur-sor, is the astrocytes (Stornetta et al., 1988; Intebi etal., 1990). Moreover, these cells in vitro express AT1-and AT2-type binding sites (Bottari et al., 1992; Tallantet al., 1996) in addition to a binding site for thehexapeptide angiotensin (3–8) (Greenland et al., 1996).More importantly, the presence of functional angioten-sin receptors has been described on these cells in vitro(Sumners et al., 1994; Tallant and Higson, 1997) and,indeed, in situ (Matute et al., 1994).
However, recent in situ hybridization studies havefailed to report on the presence of AT1 or AT2 mRNAtranscripts in brain glial cells (Lenkei et al., 1996,1998), raising questions about the physiological sig-nificance of glial angiotensin receptor-mediatedevents observed in vitro. In the present study, wehave analyzed the presence of AT1- and AT2-typeimmunoreactivity in adult brain glial cells in situ,using a panel of specific antisera. Our results indi-cate that angiotensin receptor-expressing macroglialcells are located abundantly in brain white mattertracts, rather than in gray matter areas, supportingthe hypothesis that brain glial cells may play animportant role in mediating some of the central ef-fects of angiotensins.
MATERIALS AND METHODS
The research described in this study employing ani-mals was approved by the Animal Care and Use Com-mittee of the University of the Basque Country andconforms to the National Institutes of Health guide-lines.
Western Blot
Adult rats were deeply anesthetized with ether be-fore immediate decapitation, using a specially designedguillotine. Protein extracts were obtained from freshrat brain or bovine brain corpus callosum, which hadbeen cleared of adjacent gray matter, as previouslydescribed (Domercq et al., 1999). In this study, 10–20mg of protein per lane was separated by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE). After electrotransfer to polyvinylidene fluoride(PVDF) membranes (Immobilon-P, Millipore), themembranes were blocked for 2 h at room temperaturewith 5% (w/v) nonfat milk powder in Tris-buffered sa-line (50 mM Tris-HCl, 200 mM NaCl, pH 7.3) contain-ing 0.05% Tween 20, referred to hereafter as blockingsolution. All antisera were purchased from Santa CruzBiotechnology: rabbit anti-AT1 (N10), rabbit anti-AT1(306), and goat anti-AT2 (C18). These antisera werediluted in the same blocking solution to a concentrationof 0.5 mg/ml and incubated with the membranes for 1 hat room temperature. Anti-rabbit or anti-goat second-ary antibodies conjugated to horseradish peroxidase(HRP) (Sigma) were diluted to 1:2,000 in blocking so-lution and applied to the membrane for 30 min at roomtemperature. After extensive washings, bound peroxi-dase was detected using an enhanced chemilumines-cence substrate according to the manufacturer’s in-structions (Super Signal ULTRA, Pierce, Rockford, IL).
Immunohistochemistry
Adult rats anesthetized with chloral hydrate (1 g/kg,i.p.) were transcardially perfused with fixative consist-ing of 4% formaldehyde, freshly generated from para-formaldehyde in 0.1 M phosphate buffer (25 mMNaH2PO4, 75 mM Na2HPO4, pH 7.2). Optic nerves,together with blocks containing the cerebellum andperiventricular white matter, were dissected out andpostfixed in the same fixative for 2 h. Then 3- to 5-mmtissue blocks were cryoprotected in 30% sucrose in 0.1M phosphate buffer overnight at 4°C and frozen at235°C, using Tissue-Tek mounting medium. Frozenblocks were stored at 280°C; 15-mm tissue sectionswere cut using a cryostat, transferred to phosphate-buffered saline (PBS) (2.5 mM NaH2PO4, 7.5 mMNa2HPO4, 142 mM NaCl, pH 7.2–7.4), and processed asdetailed below. Bovine corpus callosum was obtainedfresh from the slaughterhouse and rinsed with 0.1 Mphosphate buffer before immersion fixation, which con-
132 FOGARTY AND MATUTE

sisted of a 2-h incubation in tissue fixative, followed byovernight incubation at 4°C in fresh fixative. Cryopro-tection and tissue sectioning were performed as de-scribed above. Samples of postmortem human neocor-tex including the underlying white matter, and corpuscallosum were kindly made available by Dr. J.J. Meana(Department of Pharmacology, University of theBasque Country). This tissue was from suicide victimsand had been stored at 4°C for approximately 48 hbefore immersion fixation and subsequent processing.
Immunoperoxidase histochemistry was carried outaccording to standard protocols. Briefly, tissues wereincubated with 1% H2O2 for 30 min at room tempera-ture with gentle agitation, in order to inactivate endog-enous peroxidases. After tissue blocking with 2% goator horse serum (for AT1 and AT2 antisera respectively),the sections were incubated overnight with the pri-mary antisera at 4°C (rabbit anti-AT1 N-10, 0.4 mg/ml;rabbit anti-AT1 306, 0.3 mg/ml; goat anti-AT2 C18,0.3 mg/ml). Goat anti-rabbit or horse anti-goat sec-ondary antibodies (1:400, Vector) were then appliedto the sections for 1 h at room temperature, followedby similar incubation with 1:200 avidin– biotin com-plex (Vector). The presence of bound antibodies wasthen revealed using a diaminobenzidine substrate(Boehringer-Mannheim). Control experiments includedomission of primary antisera and yielded no immuno-staining in rat, bovine or human tissues. However, thehorse anti-goat secondary antibodies (Vector) werefound to cross-reacted with bovine tissue, precludingan evaluation of the distribution of AT2 receptor-likeimmunoreactivity in bovine corpus callosum.
For preadsorption experiments, angiotensin receptorantisera were preadsorbed at 4°C overnight in phos-phate buffered saline containing 2% normal serum,with a 10-fold excess (mg/ml) of the corresponding pep-tides (Santa Cruz Biotechnology) or with the noncorre-sponding peptide as an adsorption specificity control.The solution was spun at 13,000 rpm for 3 min and thesupernatant was used as preadsorbed primary anti-serum.
For double-label fluorescent immunohistochemistry,we used a mouse monoclonal anti-glial fibrillary acidicprotein (GFAP) antibody (Boehringer-Mannheim; 200ng/ml). After overnight incubation at 4°C with the anti-receptor antiserum, together with GFAP antibodies,sections were incubated with biotinylated goat anti-rabbit (to label AT1 antisera) or with horse anti-goat (tolabel AT2 antisera) immunoglobulins (1:400; Vector)for 1 h at 37°C. These secondary antibodies were thendetected by incubating sections for 1 h at 37°C with astreptavidin–Texas Red conjugate (1:100; Chemicon).After extensive washings, sections were reblocked with4% normal goat serum and exposed to Alexa 488 goatanti-mouse IgG antibodies (Molecular Probes; 1:200) at37°C for an additional 1 h. Controls included similarsections which had not been exposed to GFAP antibod-ies and demonstrated the absence of cross-reactivitybetween the two detection systems.
Homology Calculations
We used the Genetics Computer Group programsGAP and FASTA (Wisconsin Package Version 10.1,Genetics Computer Group (GCG), Madison, WI) at theEuropean Molecular Biology Laboratory EMBL mirrorsite in the National Biotechnology Centre (CNB) (Ma-drid, Spain), to evaluate the theoretical specificity ofthe angiotensin receptor antisera AT1-N10 and AT2-C18. The AT1-N10 epitope resides on the first 40 aminoacids of the AT1 peptide sequence, whereas the AT2-C18 epitope (KRESMSCRKSSSLREMET) is localizedto the 39 sequence of the AT2 receptor, corresponding toamino acids 343–360. The degree of homology betweenthese epitopes was calculated using the GCG “GAP”programme. In addition, we examined the possibilitythat the AT1 antiserum might be recognizing a similarepitope in the AT2 receptor by calculating the homologybetween the AT1 epitope and the AT2 receptor se-quence. Similar calculations were performed to evalu-ate the possibility that the AT2 antiserum might berecognizing a homologous domain in the AT1 receptor.Finally, with a view to examining the possibility thatthe antisera cross-react with other non-angiotensin re-ceptor peptides, we searched the PIR and SWISSPROTpeptide databases for other molecules that are homol-ogous to the AT1 and AT2 peptide epitopes. These cal-culations were performed using the FASTA pro-gramme, with an amino acid window of two residues.
RESULTSEvidence for the Specificity of Angiotensin
Receptor Antisera
In order to evaluate the specificity of the antiseraemployed in the present study, we examined their per-formance in Western blot. In protein extracts from
Fig. 1. Western blot of protein extracts from the bovine corpuscallosum (bCC), rat cerebellum (rCb), and rat whole brain (rWB)using the AT1 (N-10) antiserum. Proteins were separated by electro-phoresis in 10% SDS-polyacrylamide gels. The antiserum recognizes aprincipal protein species of approximately 50 kilodaltons (kDa), con-sistent with the size of the AT1 receptor. Numbers on the left repre-sent size standards in kDa.
133MACROGLIAL ANGIOTENSIN RECEPTORS IN SITU

bovine and rat tissues, the latter consisting of ratwhole brain and cerebellum homogenates, the AT1 an-tiserum (N-10; 0.5 mg/ml) recognized a principal pro-tein species with a molecular mass of approximately 50kDa (Fig. 1), a size consistent with that of a 7-trans-membrane receptor. Similar results were obtained us-ing another AT1 antiserum (anti-AT1 306; 0.5 mg/ml;not shown). The presence of this protein in the bovinecorpus callosum is indicative of the presence of the AT1receptor in glial cells, as few neuronal somata arepresent in this white matter tract (Matute and Miledi,
1993). In our hands, the antiserum to the AT2 receptor(anti-AT2, C-18, Santa Cruz Biotechnology; ¶2 mg/ml)failed to recognize any protein species in these proteinextracts, suggesting that the corresponding AT2epitope may be sensitive to the denaturing conditionsof Western blot.
We next examined by means of immunohistochemis-try the presence of specific angiotensin receptor-likeimmunoreactivity in the adult brain in situ. In therostral brain, the AT1 antisera selectively labeled thecytoplasm of cells in the piriform cortex (Fig. 2A,B), a
Fig. 2. Evidence for the specific-ity of the angiotensin receptor an-tibodies in labeling angiotensin re-ceptor-expressing cells in the ratbrain in situ. A: The AT1 anti-serum characteristically labelscells in the piriform cortex (arrow-heads) in the rostral brain. B:Higher magnification of a field inmicrophotograph (A) in which spe-cific cytoplasmic AT1 immunola-beling can be observed. Scatteredlabeled cells (arrowheads) can alsobe observed in the adjacent whitematter. The asterisk is below thesame vessel in A and B. C: AT2labeling in the caudal brain ischaracterized by intense labelingof cells of the mesencephalic tri-geminal nucleus (arrowheads) andby specific cytoplasmic labeling ofcells in the motor trigeminal nu-cleus (T). Purkinje cells in the cer-ebellum are also clearly stained(cell layer indicated by asterisk).D: Magnification of the field indi-cated by T in C. The specific cyto-plasmic labeling of these AT2 ex-pressing cells is clearly evident.Scale bars 5 250 mM in A, C; 50mM in B, D.
134 FOGARTY AND MATUTE

Fig. 3. Preadsorption of the angiotensin receptor antisera with thecorresponding peptides. A: AT1 immunostaining in a subset of cere-bellar Purkinje cells above the granule cell layer (GL), in many gran-ule cells and in cells (arrowhead) in the underlying white matter(WM). B: This staining was completely abolished when the antiserumwas preadsorbed with the corresponding AT1 peptide. C: Intense AT2
immunolabeling was observed in cerebellar Purkinje cells, in granulelayer cells and in white matter cells (arrowheads). D: No AT2 immu-nostaining was apparent after preadsorption of the AT2 antiserumwith the corresponding peptide. Scale bars 5 30 mm in A; 15 mm in B,C, D.
135MACROGLIAL ANGIOTENSIN RECEPTORS IN SITU

Figure 4.

typical site of expression of AT1 receptor (Bunnemannet al., 1992; Lenkei et al., 1998). Intense AT2 immuno-labelling was observed more caudally, in the motortrigeminal nucleus (Fig. 2C,D) in which both AT2mRNA and protein have been reported (Lenkei et al.,1996; Reagan et al., 1994). This labeling was absentfrom the nuclei of these cells, supporting its specificity.Curiously, AT2 labeling was also observed in the mes-encephalic trigeminal nucleus. The cerebellum wasalso characteristically labeled by these antisera. Thus,the AT1 antisera densely stained cells in the gray andwhite matter areas of the rat cerebellum (Fig. 3A).Preadsorption of the AT1 (N-10) antiserum (0.4 mg/ml)with a 10-fold concentration (4 mg/ml) of the corre-sponding peptide abolished all AT1 immunostaining(Fig. 3B), indicating that the antiserum exclusivelyrecognizes the AT1 epitope in rat tissue. The AT2 anti-serum also intensely stained many cells in the ratcerebellum in situ (Fig. 3C). Similar preadsorption ex-periments indicated the specificity of this immunoreac-tion. Moreover, preincubation of the antisera with het-erologous peptides (e.g., preincubating the AT1antiserum with the AT2 peptide) did not lead to areduction in immunoreactivity, indicating the specific-ity of the preadsorption process.
Finally, with a view to mathematical evaluation ofthe specificity of the antisera employed, we performedhomology calculations using the GCG programs GAPand FASTA. The degree of homology between the AT1and AT2 peptide sequences in which the correspondingepitopes are located was calculated to be 22% (usingGAP), indicating that it is highly improbable theseantisera will cross-react with the noncorrespondingepitopes. The degree of identity between the AT1 pep-tide in which the corresponding epitope resides and theAT2 receptor was calculated to be 36.1%, whereas thehomology between the AT2 immunizing peptide andthe AT1 receptor sequence was calculated to be 37.5%.These results indicate that the probability that theantisera cross-react with the noncorresponding recep-tor is very low. Finally, in order to evaluate whetherthe antisera are recognizing non-angiotensin receptorepitopes, we searched the entire Swissprot and Pir
databases (including daily updates) for proteins thatshare homology with the angiotensin peptides used togenerate the antisera. These calculations demon-strated that the protein that shares highest homologywith the AT1 peptide is a type 4 chemokine receptor(accession number: P30991; homology 40%). Similarcalculations showed that the peptide with highest ho-mology to the AT2 epitope is an Ig k-chain (accessionnumber: G30601). This homology is 50% identity over14 of 18 amino acids, is so low that it is unlikely thatthe antisera label proteins other than their correspond-ing receptors.
Rat Cerebellar White Matter
We then examined the cellular distribution of AT1and AT2 immunoreactivity in the cerebellum in moredetail. Immunostaining with the AT1 and AT2 antiserawas observed in a subset of cerebellar Purkinje cells(Fig. 3A,C), in accordance with the presence of AT1 andAT2 mRNA in these cells (Lenkei et al., 1997). Moder-ately stained cells scattered throughout the granulecell layer were also observed with both antisera. Athigher magnification, we observed angiotensin recep-tor-like immunoreactivity in numerous cells in the un-derlying white matter, which appeared to be astrocytes(Fig. 3A,C). To verify this hypothesis, we carried outdouble-label fluorescent immunohistochemistry, em-ploying a monoclonal antibody to GFAP, an astrocytemarker. Most AT1
1 cells in the cerebellar white matterwere found to express GFAP, confirming their identifi-cation as astrocytes (Fig. 4A,B). In addition, we fre-quently observed AT1 immunoreactive astrocytes inthe vicinity of blood vessels (Fig. 4A,B).
Immunostaining with the AT2 antiserum was ob-served in Purkinje cells and weak AT2 immunoreactiv-ity was apparent in some Bergmann glial cell processesin the molecular layer. Many of the AT2
1 cells in thecerebellar white matter, like those that were stainedwith the AT1 antiserum, were found to express GFAP.The presence of AT2
1 astrocytes in the vicinity of bloodmicrovessels was similarly observed (Fig. 4C,D). Incontrast to the abundance of angiotensin receptor-ex-pressing astrocytes in the cerebellar white matter, veryfew astrocytes in the overlying gray matter of the gran-ule cell layer were stained with the angiotensin recep-tor antisera (Fig. 4C).
Rat Periventricular White Matter
Staining with the AT1 antiserum was apparent inthe deep layers of the cerebral cortex (Fig. 5A) in ac-cordance with the presence of AT1 mRNA in this region(Bunnemann et al., 1992). In the underlying whitematter, surrounding the lateral ventricles, AT1 immu-noreactivity was also observed. At higher magnifica-tion, the astrocytic morphology of many of these AT1
1
cells could be appreciated (Fig. 5B). AT1 staining was
Fig. 4. Angiotensin receptor-like immunoreactivity in cerebellarwhite matter astrocytes. A: The AT1 antiserum labeled cells with atypical astrocytic morphology (arrows). Some of these cells (smallarrow) were found in the vicinity of blood microvessels (star). B: Thesecells were also found to express the astrocytic marker GFAP. Arrowsin A and B point to the same astrocytes. C: The AT2 antiserumintensely labeled Purkinje cells (thick arrows) and cells in the granulelayer (GL). In addition, some radial processes (small arrows) in themolecular layer (ML) were also stained. In the underlying whitematter (WM), AT2 immunoreactivity was observed in many astrocyte-like cells (long arrows). In the vicinity of blood vessels (star), AT2
1
processes (small arrows) were often observed. D: Double labeling withGFAP revealed the astrocytic nature of AT2-stained cells in the whitematter (symbols indicate the same elements as in C). In the graymatter of the granule cell layer (GL), much fewer AT2
1 astrocytes wereobserved (asterisk). Long, thin arrows in C and D indicate the samecells. The AT2
1 radial fibres in the molecular layer were similarlyconfirmed to be glial in nature, corresponding to Bergmann glialprocesses (small arrows). Scale bars 5 15 mm in A and B; 20 mm in Cand D.
137MACROGLIAL ANGIOTENSIN RECEPTORS IN SITU

Figure 5.

particularly dense in the ependymal cell layer sur-rounding the ventricles, indicating the presence of AT1
receptor in ependymal cells. A similar pattern of im-munoreactivity was observed with the AT2 antiserumin the periventricular white matter (Fig. 5C,D). Onceagain, ependymal cells lining the ventricle were in-tensely AT2 immunoreactive. The astrocytic nature ofmany of these angiotensin receptor-like immunoreac-tive cells was demonstrated by double-label fluores-cent, immunohistochemistry together with anti-GFAPantibodies as a marker of astrocytes (Fig. 6). ManyAT1-stained cell bodies and processes were thus iden-tified as astrocytic. Nevertheless, it was evident thatnot all astrocytic processes were AT1
1. Similarly, theAT2 antiserum stained many astrocyte cell bodies anda subset of astrocytic processes. The diffuse granularnature of neuropilar AT2 staining contrasted with thefibrous distribution of GFAP immunoreactivity (Fig.6C,D), corroborating the specificity of the double-label-ing and the absence of cross-reactivity between GFAPand AT2 receptor immunodetection systems.
Bovine and Human Corpus Callosum
Immunohistochemistry with the AT1 antisera in thebovine and human corpus callosum demonstrated thepresence of numerous lightly stained or unstained cellsamong densely stained cells. The astrocytic morphol-ogy of many of these densely stained cells with immu-noreactive processes was clearly evident at high mag-nification (Fig. 7A,D). In addition, we observednumerous moderately stained cells with a roundedmorphology, either individually scattered throughoutthe tract or grouped together in rows. The latter cellsdo not express GFAP and may thus be classified asinterfascicular oligodendrocytes (Fig. 7B,C). ManyGFAP1 processes were also found to be AT1 immuno-reactive. Similar results were observed in the rat opticnerve with AT1 and AT2 antisera staining astrocyticprocesses in addition to GFAP2 interfascicular oligo-dendrocytes (not shown). However, the AT2 antiserumdid not stain any cells in the human corpus callosum.We were unable to evaluate the presence of AT2 immu-noreactivity in the bovine corpus callosum, as the cor-responding anti-goat secondary antibodies cross-reactwith bovine tissue.
Human Subcortical White Matter
Surprisingly, the AT1 antisera did not label any cellsin this white matter area. In contrast, we observedintense AT2 immunoreactivity in cells and in bloodvessels in the white matter underlying the human ce-rebral cortex (Fig. 8A). Once again, preadsorption ofthe AT2 antiserum abolished all immunoreactivity(Fig. 8B), indicating that in human tissue, the AT2antiserum exclusively recognizes the correspondingAT2 epitope. Many of these AT2-stained cells presenteda clear astrocytic morphology at higher magnification(Fig. 8C,D). AT2
1 immunoreactive astrocytic processeswere observed to extend over distances as long as 80mm from the astrocyte soma towards neighbouringblood vessels (Fig. 8C). Rounded cells without visibleimmunoreactive processes were also AT2
1. Many ofthese cells appear to be oligodendrocytes, althoughtheir precise phenotype was not characterized.
AT1 Receptor-like Immunoreactivity in BrainWhite Matter Versus Gray Matter
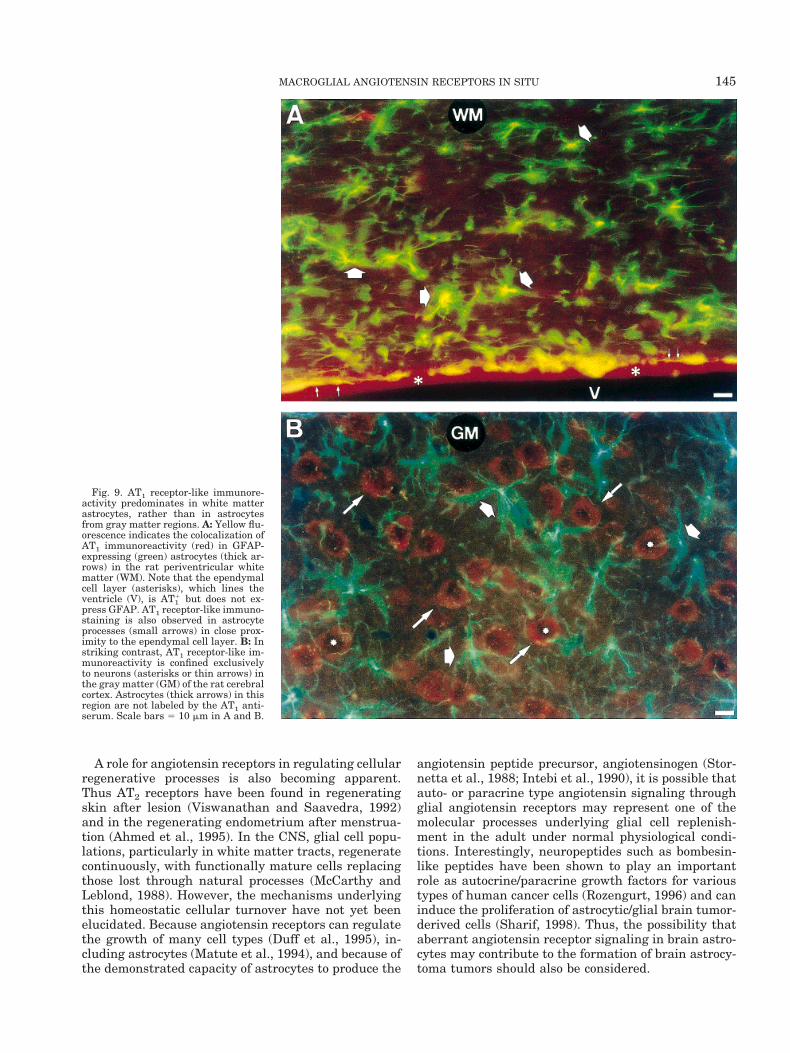
We have shown that angiotensin receptor-like immu-noreactivity colocalizes with GFAP immunoreactivityin many astrocytes in a variety of brain white mattertracts. Thus, AT1 immunostaining in rat periventricu-lar astrocytes is indicated by the overlapping of red(AT1) and green (GFAP) fluorescence, which appears asa yellow fluorescence (Fig. 9A). Ependymal cells clearlydo not express GFAP, and the distinctively red fluores-cence in this cell layer indicates that the colocalizationin astrocytes of angiotensin receptor-like immunoreac-tivity with GFAP is not due to nonspecific cross-reac-tions. However, in the cerebral cortex (Fig. 9B), AT1receptor-like immunoreactivity was located exclusivelyin neurons, as AT1 and GFAP immunofluorescence didnot colocalize (absence of yellow fluorescence). Thisneuronal location of the AT1 receptor in the cerebralcortex is in accordance with the reported presence ofAT1 mRNA in this brain region (Bunnemann et al.,1992). Moreover, this result supports the idea thatbrain angiotensin receptors in gray matter areas arepredominantly neuronal in nature (Lenkei et al., 1997).
DISCUSSIONEvidence for the Specificity of the Antisera
Western blot
The AT1 antisera employed in the present study rec-ognize a protein species with a molecular mass of ;50kDa. This size is consistent with that of a 7-transmem-brane receptor, and in particular with that of the AT1receptor, which is predicted to have a minimum size of41 kDa on the basis of its amino acid composition(Murphy et al., 1991). Indeed, AT1 receptors with amolecular mass of 45 kDa have already been reported
Fig. 5. Angiotensin receptor-like immunoreactivity in cells of therat periventricular white matter. A: The AT1 antiserum labels manycells in this white matter region, in addition to cells of the ependymalcell layer (between arrowheads), which line the lateral ventricle (V).B: Higher magnification of the area indicated by a star in A shows thecharacteristic astrocytic morphology of these immunoreactive cells(arrowheads). C: A similar pattern of immunostaining is observedwith the AT2 antiserum which apparently labels cell somata (arrows).D: The astrocytic morphology of these cells becomes more apparent athigher magnification (arrows). Arrows in C and D point to the samecells. Scale bars 5 80 mm in A and C; 20 mm in B and D.
139MACROGLIAL ANGIOTENSIN RECEPTORS IN SITU

Figure 6.

in rat tissues using a different AT1 antiserum (Paxtonet al., 1993). This protein species was consistently ob-served with the AT1 antisera in protein extracts fromrat whole brain and cerebellum and from the hip-pocampus (latter, not shown). In addition, the antiserarecognize a protein species with a similar molecularmass in bovine extracts, further supporting the speci-ficity of the antisera. Moreover, the specificity of theAT1 (N-10) antiserum has already been demonstratedin endothelial cell lysates (Rose and Audus, 1999). Incontrast, the AT2 antiserum did not recognize proteinsin these extracts, suggesting that the denaturing con-ditions employed in Western blot alter the C-terminalepitope recognized by this antiserum. Nevertheless,preadsorption of the AT1 and AT2 antisera led to acomplete abolition of the corresponding immunoreac-tivity in rat, bovine, and human tissue. This preadsorp-tion process may be considered highly specific becausepreadsorption of each antiserum with heterologous an-tigenic peptides (e.g., preadsorption of the AT1 anti-serum with the AT2 peptide) did not reduce the inten-sity of immunoreactivity.
Immunohistochemistry
By means of immunohistochemistry, we observedthat the antisera selectively labeled sites that includethe piriform cortex (AT1) and the motor trigeminalnucleus of the mesencephalon (AT2). These results areconsistent with the reported presence of the corre-sponding mRNA in these brain nuclei. Surprisingly, wealso observed intense AT2 immunoreactivity in themesencephalic trigeminal nucleus, in which only AT1mRNA has been reported. The lack of homology be-tween the AT2 receptor epitope recognized by the AT2antiserum and the AT1 receptor sequence (37.5% iden-tity with only two adjacent identical residues; GCGGap program) indicates that this labeling is not due tocross-reactivity. However, reports of mismatches in thelocalization of angiotensin receptor mRNA, protein,and binding sites are relatively common (Lenkei et al.,1997). Thus, intense AII receptor labeling has beenreported in the magnocellular neurons of the supraop-tic nucleus and paraventricular nucleus of the hypo-thalamus, nuclei in which AT1 and AT2 binding sitesand receptor mRNA are absent. The significance ofthese findings remain to be elucidated, but they maypoint to the presence of additional, as yet unclonedangiotensin receptor subtypes (Fogarty et al., in prep-aration).
Homology calculations
We performed homology calculations with a view toevaluating the theoretical specificity of the antiseraemployed. Theoretical specificity can be measured interms of homology, as the higher the homology betweentwo peptides, the greater the probability that an anti-serum raised against one of them will crossreact withthe other. Our calculations demonstrated very low ho-mology (22%) between the two peptides in which theAT1 and AT2 antigenic epitopes reside, indicating thatthe antisera are unlikely to cross-react at the noncor-responding epitope. Similarly, low homology was cal-culated for these peptides and the noncorrespondingreceptor (AT1 peptide 3 AT2 receptor 5 36%; AT2 pep-tide 3 AT1 receptor 5 37.5%), indicating that theepitopes used are specific to, and characteristic of, eachreceptor. Interestingly, it has been reported that theAPJ protein, which is highly expressed in the whitematter of the brain, is homologous to angiotensin re-ceptors (O’Dowd et al., 1993). However, neither the AT1nor the AT2 epitopes recognized by the antisera hassignificant homology with this protein (AT1 peptide 3APJ 5 45%; AT2 peptide 3 APJ 5 28%; GCG programGap). Finally, we evaluated whether the antisera maybe recognizing other peptides not belonging to the an-giotensin receptor family, by searching for homologousproteins in the entire Swissprot and Pir databases. Noproteins with homology greater than 50% were found,indicating that the angiotensin receptor peptide se-quences in which the antigenic epitopes reside consti-tute a signature of their corresponding receptor. Over-all, it may be concluded that the antisera specificallyand exclusively recognize their corresponding epitopesin the tissues employed in the present study.
AT1 and AT2 Immunoreactive Glial Cells inBrain White Matter
Previous angiotensin receptor localization studieshave concluded that brain angiotensin receptors arelocated principally in neurons. Thus Lenkei et al.(1998) described the absence of AT1 receptor in glialcells in the lateral hypothalamic area using in situhybridization combined with immunohistochemistry.The same investigators concluded that AT2 is neuronalrather than glial in nature, on the basis of studies withan AT2 riboprobe in the lateral septum (Lenkei et al.,1996). We observed a similar result in the gray matterregion of the cerebral cortex, in which angiotensin re-ceptor-like immunoreactivity was confined exclusivelyto neurons (Fig. 9). In addition, we rarely observedangiotensin receptor-like immunoreactive astrocytes inthe gray matter of the cerebellar granule cell layer.However, Western blot using AT1 antisera clearly sug-gested the presence of AT1 receptor in protein extractsfrom the bovine corpus callosum, a white matter tractthat unites the cerebral hemispheres. This finding isindicative of the presence of AT1 receptors in glial cells,
Fig. 6. AT1 and AT2 immunoreactivity is present in astrocytes inthe rat periventricular white matter surrounding the ventricles (V).A, B: Double-label fluorescent immunohistochemistry with the AT1antiserum and antibodies to GFAP, respectively, reveals the presenceof AT1 immunoreactivity in GFAP1 astrocytes (arrows). Not all astro-cytic processes are AT1
1 (area below large arrow). C, D: Similarly, AT2and GFAP immunoreactivity, respectively, were observed in numer-ous astrocyte somata (arrows) and processes (arrowheads) in thiswhite matter tract. Scale bars 5 20 mm in A and B; 20 mm in C and D.
141MACROGLIAL ANGIOTENSIN RECEPTORS IN SITU

Figure 7.

as few neuronal somata are present in these whitematter tracts (Matute and Miledi, 1993). Moreover,mRNA from the adult bovine and human corpus callo-sum has been found to encode functional AT1 angioten-sin receptors (Matute et al., 1994). In addition, AT1mRNA has been reported in the small cells of severalfiber tracts in the brain (Lenkei et al., 1998) and thepresence of AT2-type binding sites have been observedin the white matter of the cerebellum (Whitebread etal., 1991), although little importance was attributed tothese latter findings. Using double-label fluorescentimmunohistochemistry with AT1 and AT2 specific an-tisera, we clearly demonstrate the presence of AT1 andAT2 immunoreactivity in glial cells, principally astro-cytes and oligodendrocytes, in adult brain white matterareas. Moreover, analysis by reverse transcription-polymerase chain reaction (RT-PCR) and immunocyto-chemistry of purified cultures of astrocytes, oligoden-drocytes, and microglia in vitro confirms these findingsin that angiotensin receptor mRNA and protein wereobserved in astrocytes and oligodendrocytes, but not inmicroglia (D.J.F. and C.M., unpublished results). Over-all, these results indicate that the angiotensin receptorphenotype of glial cells is regionally specific and fur-thermore dependent on the type of brain matter (i.e.,gray or white matter) in which the cells reside.
Possible Functional Significance of GlialAngiotensin Receptor Expression In Situ
The fact that this glial location of angiotensin recep-tor-like immunoreactivity is present in rat, bovine, andhuman tissues suggests functionally important rolesfor glial angiotensin receptors in the adult CNS. Inwould be reasonable to think that glial angiotensinreceptors participate in mediating some of the well-established central effects of angiotensins such as car-diovascular regulation and hydroelectrolytic homeosta-sis (reviewed in Allen et al., 1998). In fact, it has beenshown that activation of astrocyte angiotensin recep-tors in vitro can lead to the liberation of arachidonicacid metabolites such as prostaglandins (Tallant andHigson, 1997) and aldosterone (Matute et al., 1994).These substances can potentially alter neuronal activ-ity, as it has been demonstrated that prostaglandinscan alter the activity of neuronal potassium channels(Zhu et al., 1998) and that aldosterone can upregulate
the expression of brain (Na,K)-ATPases (Grillo et al.,1997). In this way, glial angiotensin receptor activationcould lead to an alteration in the activity of neuronsinvolved in blood pressure regulation and body fluidhomeostasis. In addition, glial aldosterone may stimu-late capillary endothelial sodium channels to regulatethe influx of sodium into the brain, in a manner similarto that which occurs in the kidney (Ardaillou, 1999).
However, this glial location of angiotensin receptorsmay also underly some of the novel functions that haverecently been attributed to the central RAS (see reviewby Wright and Harding, 1997; Stroth and Unger, 1999).We consistently observed angiotensin receptor-like im-munoreactive astrocytes or their processes in the im-mediate vicinity of white matter blood vessels (Figs. 4,8). The capacity of astrocytes to liberate prostaglan-dins, which are important mediators of cerebrovascularcontrol (Busija, 1997), in response to angiotensin recep-tor activation, points to the possibility that angiotensinreceptor-expressing astrocytes play important roles inregulating cerebral blood flow. Moreover, becausewhite matter constitutes a large proportion of the brain(see Bedell and Narayana, 1998), cerebrovascular reg-ulation in the white matter, mediated by angiotensin-induced prostaglandin liberation from astrocytes, couldplay important roles in potentiating blood flow to graymatter regions, under conditions of enhanced meta-bolic requirements due to increased neuronal activity.
Another novel function that has been assigned to thecentral RAS is the regulation of cognitive functionssuch as learning and memory formation (see review byWright and Harding, 1997). Since these functions re-quire the operation of correctly functioning neural net-works, of which cerebral white matter is an essentialcomponent (Filley, 1998), our results raise the possibil-ity that white matter glial angiotensin receptors mayrepresent one of the substrates through which brainangiotensins modulate these cognitive functions.
Glial cells have also been shown to liberate the neu-rosteroid aldosterone upon angiotensin receptor activa-tion (Matute et al., 1994). Glial aldosterone, throughupregulation of neuronal (Na,K)-ATPases, may alsoplay an important neuroprotective role by maintainingthe neuronal membrane potential in pathological con-ditions (Lees, 1991; Grillo et al., 1997). The fact thatthe enzymes responsible for steroid biosynthesis havebeen found in various myelinated tracts in the brain(Gomez-Sanchez et al., 1997) supports this hypothesisand also points to the possibility that angiotensin re-ceptors on oligodendrocytes may regulate the produc-tion of neurosteroids including aldosterone. Indeed,genes encoding steroidogenic enzymes have been re-ported to be expressed in oligodendrocytes (Mellon andDeschepper, 1993). In light of our results, the possibil-ity that glial angiotensin receptors may constitute animportant novel therapeutic target for pathologies in-volving white matter damage such as Alzheimer’s dis-ease (Wallin, 1998), poststroke dementia (Leys et al.,1998), and periventricular leukomalacia (Inder et al.,1999) should be considered.
Fig. 7. AT1 receptor-like immunoreactivity in the bovine and hu-man corpus callosum. A: Intensely labeled process-bearing cells wereobserved in the bovine corpus callosum (arrows), in addition to AT1
1
rounded cells (white arrows), although immunonegative cells withthis morphology were also seen (black arrowheads). B, C: GFAP andAT1 double immunohistochemistry, respectively, show that AT1-ex-pressing rounded cells (small arrows in C) do not express GFAP,indicating that these cells are probably interfascicular oligodendro-cytes. Similarly, GFAP1 astrocyte processes (arrowheads in B) werefound to be AT1
1 (arrowheads in C). D: The AT1 antisera similarlystained process-bearing astrocytes (arrows) and small rounded cells(arrowheads) in the human corpus callosum. Scale bars 5 15 mm in Aand D; 20 mm in B and C.
143MACROGLIAL ANGIOTENSIN RECEPTORS IN SITU
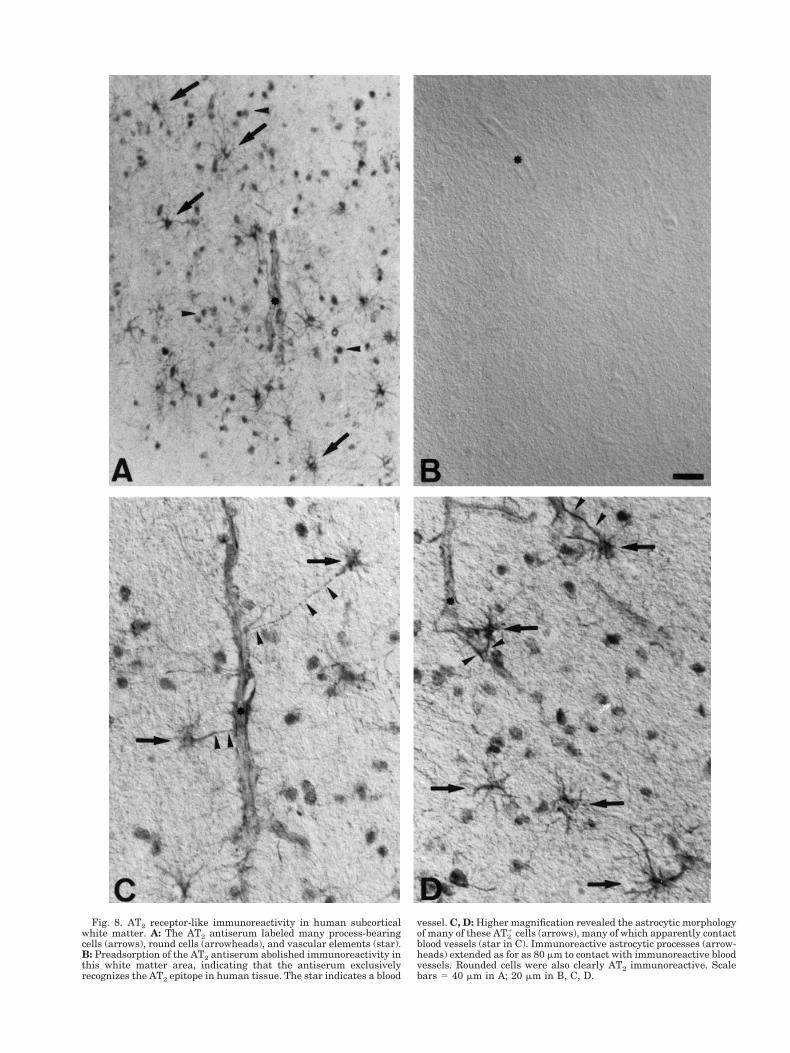
Fig. 8. AT2 receptor-like immunoreactivity in human subcorticalwhite matter. A: The AT2 antiserum labeled many process-bearingcells (arrows), round cells (arrowheads), and vascular elements (star).B: Preadsorption of the AT2 antiserum abolished immunoreactivity inthis white matter area, indicating that the antiserum exclusivelyrecognizes the AT2 epitope in human tissue. The star indicates a blood
vessel. C, D: Higher magnification revealed the astrocytic morphologyof many of these AT2
1 cells (arrows), many of which apparently contactblood vessels (star in C). Immunoreactive astrocytic processes (arrow-heads) extended as for as 80 mm to contact with immunoreactive bloodvessels. Rounded cells were also clearly AT2 immunoreactive. Scalebars 5 40 mm in A; 20 mm in B, C, D.
A role for angiotensin receptors in regulating cellularregenerative processes is also becoming apparent.Thus AT2 receptors have been found in regeneratingskin after lesion (Viswanathan and Saavedra, 1992)and in the regenerating endometrium after menstrua-tion (Ahmed et al., 1995). In the CNS, glial cell popu-lations, particularly in white matter tracts, regeneratecontinuously, with functionally mature cells replacingthose lost through natural processes (McCarthy andLeblond, 1988). However, the mechanisms underlyingthis homeostatic cellular turnover have not yet beenelucidated. Because angiotensin receptors can regulatethe growth of many cell types (Duff et al., 1995), in-cluding astrocytes (Matute et al., 1994), and because ofthe demonstrated capacity of astrocytes to produce the
angiotensin peptide precursor, angiotensinogen (Stor-netta et al., 1988; Intebi et al., 1990), it is possible thatauto- or paracrine type angiotensin signaling throughglial angiotensin receptors may represent one of themolecular processes underlying glial cell replenish-ment in the adult under normal physiological condi-tions. Interestingly, neuropeptides such as bombesin-like peptides have been shown to play an importantrole as autocrine/paracrine growth factors for varioustypes of human cancer cells (Rozengurt, 1996) and caninduce the proliferation of astrocytic/glial brain tumor-derived cells (Sharif, 1998). Thus, the possibility thataberrant angiotensin receptor signaling in brain astro-cytes may contribute to the formation of brain astrocy-toma tumors should also be considered.
Fig. 9. AT1 receptor-like immunore-activity predominates in white matterastrocytes, rather than in astrocytesfrom gray matter regions. A: Yellow flu-orescence indicates the colocalization ofAT1 immunoreactivity (red) in GFAP-expressing (green) astrocytes (thick ar-rows) in the rat periventricular whitematter (WM). Note that the ependymalcell layer (asterisks), which lines theventricle (V), is AT1
1 but does not ex-press GFAP. AT1 receptor-like immuno-staining is also observed in astrocyteprocesses (small arrows) in close prox-imity to the ependymal cell layer. B: Instriking contrast, AT1 receptor-like im-munoreactivity is confined exclusivelyto neurons (asterisks or thin arrows) inthe gray matter (GM) of the rat cerebralcortex. Astrocytes (thick arrows) in thisregion are not labeled by the AT1 anti-serum. Scale bars 5 10 mm in A and B.
145MACROGLIAL ANGIOTENSIN RECEPTORS IN SITU

Our results clearly indicate the presence of AT1 andAT2 immunoreactivity in macroglial cells in variouswhite matter tracts in the adult brain. The identifica-tion of astrocytes and oligodendrocytes as cells thatexpress AT1 and AT2 receptors offers important cluesregarding the physiological roles of these receptors inthe brain.
ACKNOWLEDGMENTS
This work was financed by the University of theBasque Country, grant G-14/98 (fellowship to D.J.F.)and by the Ministry of Education and Culture, grantPM-97/45.
REFERENCES
Ahmed A, Li XF, Shams M, Gregory J, Rollason T, Barnes NM,Newton JR. 1995. Localization of the angiotensin II and its receptorsubtype expression in the human endometrium and identification ofa novel high-affinity angiotensin II binding site. J Clin Invest 96:848–857.
Allen AM, Moeller I, Jenkins TA, Zhuo G, Aldred P, Chai SY, Men-delsohn FAO. 1998. Angiotensin receptors in the nervous system.Brain Res Bull 47:17–28.
Ardaillou R. 1999. Angiotensin II receptors. J Am Soc Nephrol 10:S30–S39.
Bedell BJ, Narayana PA. 1998. Volumetric analysis of white matter,gray matter and CSF using fractional volume analysis. Magn ResonMed 39:961–969.
Bottari SP, Obermuller N, Bogdal Y, Zahs K, Deschepper CF. 1992.Characterization and distribution of angiotensin II binding sites infetal and neonatal astrocytes from different rat brain regions. BrainRes 585:372–376.
Bunnemann B, Naoharu I, Metzger R, Fuxe K, Inagami T, Ganten D.1992. The distribution of angiotensin II AT1 receptor subtypemRNA in the rat brain. Neurosci Lett 142:155–158.
Busija D. 1997. Eicosanoids and cerebrovascular control. In: WelchKMA, Caplar LR, Reis DJ, et al. editors. Primer on cerebrovasculardiseases. San Diego, CA: Academic Press; p 93–95.
Domercq M, Sanchez-Gomez MV, Areso P, Matute C. 1999. Expres-sion of glutamate transporters in rat optic nerve oligodendrocytes.Eur J Neurosci 11:2226–2236.
Duff JL, Marrero MB, Paxton WG, Schieffer B, Bernstein KE, BerkBC. 1995. Angiotensin II signal transduction and the mitogen-activated protein kinase pathway. Cardiovasc Res 30:511–517.
Filley CM. 1998. The behavioral neurology of cerebral white matter.Neurology 50:1535–1540.
Fitzsimons JT. 1998. Angiotensin, thirst and sodium appetite. PhysiolRev 78:583–686.
Gomez-Sanchez CE, Zhou MY, Cozza EN, Morita H, Foecking MF,Gomez-Sanchez EP. 1997. Aldosterone biosynthesis in the ratbrain. Endocrinology 138:3369–3373.
Greenland K, Wyse B, Sernia C. 1996. Identification and character-ization of angiotensin IV binding sites in rat neuron and astrocytecell cultures. J Neuroendocrinol 8:687–693.
Grillo C, Piroli G, Lima A, McEwen BG, De Nicola AF. 1997. Aldoste-rone up-regulates mRNA for the a3 and b1 isoforms of (Na,K)-ATPase in several brain regions from adrenalectomized rats. BrainRes 767:120–127.
Inder TE, Huppi PS, Warfield S, Kikinis R, Zientara GP, Barnes PD,Jolesz F, Volpe JJ. 1999. Periventricular white matter injury in thepremature infant is followed by reduced cerebral cortical gray mat-ter volume at term. Ann Neurol 46:755–760.
Intebi AD, Flaxman MS, Ganong WF, Deschepper CF. 1990. Angio-tensinogen production by rat astroglial cells in vitro and in vivo.Neuroscience 34:545–554.
Lees GJ. 1991. Inhibition of sodium-potassium-ATPase: a potentialubiquitous mechanism contributing to central nervous system neu-ropathology. Brain Res Rev 16:283–300.
Lenkei Z, Palkovits M, Corvol P, Llorens-Cortes C. 1996. Distributionof angiotensin II type-2 receptor (AT2) mRNA expression in theadult rat brain. J Comp Neurol 373:322–339.
Lenkei Z, Palkovits M, Corvol P, Llorens-Cortes C. 1997. Expressionof angiotensin type-1 (AT1) and type-2 (AT2) receptor mRNAs in theadult rat brain: a functional neuroanatomical review. Front Neu-roendocrinol 18:383–439.
Lenkei Z, Palkovits M, Corvol P, Llorens-Cortes C. 1998. Distributionof angiotensin type-1 receptor messenger RNA expression in theadult rat brain. Neuroscience 82:827–841.
Leys D, Henon H, Pasquier F. 1998. White matter changes andpoststroke dementia. Dement Geriatr Cogn Disord 9:25–29.
Matute C, Miledi R. 1993. Neurotransmitter receptors and voltage-dependent calcium channels encoded by mRNA from the adultcorpus callosum. Proc Natl Acad Sci USA 90:3270–3274.
Matute C, Pulakat L, Rio C, Valcarcel C, Miledi R. 1994. Properties ofangiotensin II receptors in glial cells from the adult corpus callo-sum. Proc Natl Acad Sci USA 91:3774–3778.
McCarthy GF, Leblond CP. 1988. Autoradiographic evidence for slowastrocyte turnover and modest oligodendrocyte production in thecorpus callosum of adult mice infused with 3H-thymidine. J CompNeurol 271:589–603.
Mellon SH, Deschepper CF. 1993. Neurosteroid biosynthesis: genesfor adrenal steroidogenic enzymes are expressed in the brain. BrainRes 629:283–292.
Murphy TJ, Alexander RW, Griendling KK, Runge MS, Bernstein KE.1991. Isolation of a cDNA encoding the vascular type-1 angiotensinII receptor. Nature 351:233–236.
O’Dowd BF, Heiber M, Chan A, Heng HHQ, Tsui L-C, Kennedy JL,Shi X, Petronis A, George SR, Nguyen T. 1993. A human gene thatshows identity with the gene encoding the angiotensin receptor islocated on chromosome 11. Gene 136:355–360.
Paxton WG, Runge M, Horaist C, et al. 1993. Immunohistochemicallocalization of rat angiotensin II AT1 receptor. Am J Physiol 264:F989–F995.
Phillips MI, Shen L, Richards EM, Raizada MK. 1993. Immunohisto-chemical mapping of angiotensin AT1 receptors in the brain. RegulPeptide 44:95–107.
Reagan LP, Flanagan-Cato LM, Yee DK, Ma L-Y, Sakai RR, FluhartySJ. 1994. Immunohistochemical mapping of angiotensin type 2(AT2) receptors in rat brain. Brain Res 662:45–59.
Rose JM, Audus KL. 1999. AT1 receptors mediate angiotensin IIuptake and transport by bovine brain microvessel endothelial cellsin primary culture. J Cardiovasc Pharmacol 33:30–35.
Rozengurt E. 1996. Neuropeptide growth factors: signalling pathwaysand role in cancer. In: Pusztai LL, Lewis CE, Yap E, editors. Cellproliferation in cancer: regulatory mechanisms of neoplastic cellgrowth. Oxford: Oxford University Press. p 247–259.
Sharif M. 1998. Mitogenic signaling by substance P and bombesin-likeneuropeptide receptors in astrocytic/glial brain tumor-derived celllines. Int J Oncol 12:273–286.
Stornetta KL, Hawelu-Johnson CL, Guyenet PG, Lynch KR. 1988.Astrocytes synthesize angiotensinogen in brain. Science 242:1444–1446.
Stroth U, Unger T. 1999. The renin-angiotensin system and its recep-tors. J Cardiovasc Pharmacol 33:S21–S28.
Sumners C, Tang W, Paulding W, Raizada MK. 1994. Peptide recep-tors in astroglia: focus on angiotensin II and atrial natriureticpeptide. Glia 11:110–116.
Tallant EA, Higson JT. 1997. Angiotensin II activates distinct signaltransduction pathways in astrocytes isolated from neonatal ratbrain. Glia 19:333–342.
Tallant EA, Diz DI, Ferrario CM. 1996. Identification of AT1 receptorson cultured astrocytes. Adv Exp Med Biol 396:121–129.
Viswanathan M, Saavedra JM. 1992. Expression of angiotensin II AT2receptors in the rat skin during experimental wound healing. Pep-tides 13:783–786.
Wallin A. 1998. The overlap between Alzheimer’s disease and vascu-lar dementia: the role of white matter changes. Dement GeriatrCogn Disord 9:30–35.
Whitebread SE, Taylor V, Bottari SP, et al. 1991. RadioiodinatedCGP42112A: a novel high affinity and highly selective ligand for thecharacterization of angiotensin AT2 receptors. Biochem BiophysRes Commun 181:1365–1371.
Wright JW, Harding JW. 1997. Important roles for angiotensin IIIand IV in the brain renin-angiotensin system. Brain Res Rev 25:96–124.
Zhu M, Gelband CH, Moore JM, Posner P, Sumners C. 1998. Angio-tensin II type 2 receptor stimulation of neuronal delayed rectifierpotassium current involves phospholipase A2 and arachidonic acid.J Neurosci 18:679–686.
146 FOGARTY AND MATUTE




















