
Astrocytes in kindling: relevance to epileptogenesis
13
b ELSEVIER Epileps.~ Research 26 11'-~i6~ 1¢~3- 175 EPILEPSY RESEARCH Astrocytes in kindling: relevance to epileptogenesis Moshe Khurgel a., Gwen O. Ivy a.b " Department A,:latomy aml Cell Biology, Faculty ¢{fMedicim', Scarboro,.~h ('amlm~. Unitcr.~it', ~)" T~,ro~Jto, Scarborol~gh, Ont, MiC IA4. b Department t{fPsychology. Scarborough Campus. U~lirersity qfToron;o, Scarbolough. Ore. MIC IA4. Cam~da Received 15 November 1995: accepted 26 February 1996 Abstract Astrogliosis is a prominent feature of epileptic ~bci, and may play a causal role in the development of seizures and the persistance of seizure disorders. We have studied morphological changes in astrocytes with respect to the evolution of seizures using the kindling model of epilepsy. Kindling-induced seizures result in a prominent hypertrophy of astrocytes that is accompanied by a reorganization of astrocytic cytoskeleton. The change in the morphology of astrocytes appears to be seizure-intensity dependent, occurs early in the kindling process, and persists for weeks following the last seizure. In addition to hypertrophy, we have observed an increase in proliferation of astrocytes in hippocampus, amygdala and pirifonn cortex, but no change in the expression of connexin-43 following kindling. Significantly, induction of a localized astrocyte hyoerlrophy prior to initiation of kindling does not result in seizures and does not facilitate kindling. Altogether these data suggest that 'gliosis' is an adaptive response to seizures. Keywords: Seizures; Gliosis: Vimentin; Gap junctions; bFGF: Proliferatkm I. Introduction The possibility of a causal relationship between a disfunction of astrocytes and epileptogenesis has been raised occasionally throughout the history of the study of epilepsy. However, astrocytes have been traditionally viewed as passive 'support cells' in the mammalian brain, and as such, not actively involved in neurotransmission. Thus, apart from a few reports * Corresponding author. Present address: Department of Neuroscience, University of Virginia, Health Sciences Center, P.O. Box 5148, MR4, Charlottesville, VA 22908, USA. Tel.: +1 (804) 924-9111; fax: +1 (804) 982-4380" e-mail: [email protected] advancing an alternative point of view, the develop- ment of hyperactivity and hypersynchronicity charac- teristic of epileptic seizures have been usually thought to represent abnormalities in neuronal connectivity or intrinsic properties of neurons, rather than possi- ble aberrations in the properties of astrocytes. In the past few years the role of astrocytes in neural activity has been receiving increasing atten- tion. Recent findings suggest that certain character- istics of astrocytes are critical for the maintenance of normal neural activity and that a disruption of proper astrocytic function may result in, among other events, development of seizures. We, and several other research groups, have used the 'dndling model of epilepsy to study the nature 0920-12 ! 1/96/$15.00 Copyright © 1996 Elsevier Science B.V. All rights reserved. PI! S0920-1211(96)0005 I-4
-
Upload
bridgewater -
Category
Documents
-
view
3 -
download
0
Transcript of Astrocytes in kindling: relevance to epileptogenesis

b
ELSEVIER Epileps.~ Research 26 11'-~i6~ 1¢~3- 175
EPILEPSY RESEARCH
Astrocytes in kindling: relevance to epileptogenesis
Moshe Khurgel a. , Gwen O. Ivy a.b " Department A,:latomy aml Cell Biology, Faculty ¢{fMedicim', Scarboro,.~h ('amlm~. Unitcr.~it', ~)" T~,ro~Jto, Scarborol~gh, Ont, MiC IA4.
b Department t{fPsychology. Scarborough Campus. U~lirersity qfToron;o, Scarbolough. Ore. MIC IA4. Cam~da
Received 15 November 1995: accepted 26 February 1996
Abstract
Astrogliosis is a prominent feature of epileptic ~bci, and may play a causal role in the development of seizures and the persistance of seizure disorders. We have studied morphological changes in astrocytes with respect to the evolution of seizures using the kindling model of epilepsy. Kindling-induced seizures result in a prominent hypertrophy of astrocytes that is accompanied by a reorganization of astrocytic cytoskeleton. The change in the morphology of astrocytes appears to be seizure-intensity dependent, occurs early in the kindling process, and persists for weeks following the last seizure. In addition to hypertrophy, we have observed an increase in proliferation of astrocytes in hippocampus, amygdala and pirifonn cortex, but no change in the expression of connexin-43 following kindling. Significantly, induction of a localized astrocyte hyoerlrophy prior to initiation of kindling does not result in seizures and does not facilitate kindling. Altogether these data suggest that 'gliosis' is an adaptive response to seizures.
Keywords: Seizures; Gliosis: Vimentin; Gap junctions; bFGF: Proliferatkm
I. Introduction
The possibility of a causal relationship between a disfunction of astrocytes and epileptogenesis has been raised occasionally throughout the history of the study of epilepsy. However, astrocytes have been traditionally viewed as passive 'support cells' in the mammalian brain, and as such, not actively involved in neurotransmission. Thus, apart from a few reports
* Corresponding author. Present address: Department of Neuroscience, University of Virginia, Health Sciences Center, P.O. Box 5148, MR4, Charlottesville, VA 22908, USA. Tel.: +1 (804) 924-9111; fax: +1 (804) 982-4380" e-mail: [email protected]
advancing an alternative point of view, the develop- ment of hyperactivity and hypersynchronicity charac- teristic of epileptic seizures have been usually thought to represent abnormalities in neuronal connectivity or intrinsic properties of neurons, rather than possi- ble aberrations in the properties of astrocytes.
In the past few years the role of astrocytes in neural activity has been receiving increasing atten- tion. Recent findings suggest that certain character- istics of astrocytes are critical for the maintenance of normal neural activity and that a disruption of proper astrocytic function may result in, among other events, development of seizures.
We, and several other research groups, have used the 'dndling model of epilepsy to study the nature
0920-12 ! 1/96/$15.00 Copyright © 1996 Elsevier Science B.V. All rights reserved. PI! S0920-1211(96)0005 I-4

164 M. Khurgel, G.O. It')'~ Epilepsy Research 26(1996) 163-175
and the significance of some of the changes in astrccytes with respect to epileptogenesis. Kindling- induced seizures have been shown to result in hyper- trophy of astrocytes in conjunction with prominent cytoskeletal changes in these cells. These structural changes take place early in the development of kin- dling, intensify during the weeks following the last seizure and persist for long periods of time. The 'gliosis' is confined to the stimulated and seizure- propagating brain regions, such as the piriform cor- tex, amygdala, entorhinal cortex and certain sectors of the hippocampus. Regionally specific patterns of astrocyte hypertrophy can be evoked through kin- dling of different brain sites. In addition to the hypertrophy, there is a limited but significant prolif- eration of these cells.
Reactive astrogliosis is a prominent morphologi- cal feature in seizure foci of human patients and has been observed in many animal models of seizures and of epilepsy. Traditionally, this phenomenon has been regarded as a response of astrocytes to neuronal degeneration, and as such, not directly relevant to the process of epileptogenesis. Using several histological approaches we have been able to demonstrate that certain changes in astrocytes can occur in the ab- sence of neuronal degeneration; probably in response to signals associated with seizure activity. Since hypertrophied astrocytes often appear to constitute the only prominent structural change at the seizure- generating sites, it is conceivable that these cells play a crucial role in the evolution of a seizure-prone state.
In this paper we review selected studies on astro- cytes with the focus on morphological changes in these cells, and discuss the possible relevance of these changes to epilepsy. We also present some of the recent findings from our work with the kindling model which may shed new light on the role of astrocytes in epileptogenesis.
2. General properties of astrocytes in a mature brain
Astrocytes are possibly the most multifaceted and enigmatic cells in the mammalian brain. The extent of participation of astrocytes in neural activity only recently has began to be appreciated due to the
Fig. i. Spatial relationship between astrocytes and other neural elements in the gray matter. Activated vimentin * astrocytes in the Amy/Pi t area of a kindled rat maintaiJi the normal positioning and some of the structural interactions of astrocytes among them- selves, with neurons (N), and blood vessels. Vimentin immunohis- tochemistry, counter-stained with cresyl violet. Scale bar: approx. 15 pm.
rapidly accumulating knowledge about the properties of these cells and the inevitable realization that the comprehension of brain processes will depend on understanding the individual characteristics of and interactions between all cellular elements.
Several classes of astrocytes are now recognized, based on cell morphology and localization in the brain. Among those° the 'typical' gray matter astro- cytes occupy a seemingly strategic position. Their cell bodies are dispersed more or less evenly throughout most regions, while the multiple pro- cesses, which we propose to call 'stellae' (to reflect the star-shaped morphology of these cells and to differentiate these formations from neuronal den- drites and cellular processes of oligodendrocytes and microglia), stretch out to form multiple contacts with other neural elements (Fig. 1) [12,48]. The functional significance of these structural associations is not completely understood: the ensheathing of synapses and neuronal somas is thought to limit the spread of ions and neurotransmitters, the ensheathing of blood capillaries by astrocytic 'end feet' may be important for the maintenance of the blood-brain barrier, while the contacts between astrocytes themselves, which often contain gap junctions, may play a role in spatial redistribution of molecules and, ultimately, communication between these cells.
Physiologically, astrocytes are thought to fulfil several crucial functions. Astrocytes have been hy-

M. Ktmrgel. G.O. h y/Epih'l~sy Research 26 f /996~/63. / 75 165
pothesized for a long time to play a sig~fificant role in ionic homeostasis, in particular the maintenance of proper extracetluar concentrations of K ~ [29,70]. The selective localization of glutamine synthetase [44] and possibly glutamate dehydrogenase [38] in astrocytes designates these cells as key players in the metabolism of glutamate, while the significant pres- ence of monoamine oxidases in astrocytes implicates them in the metabolism of monoamine neurotrans- mitters. In additiom astrocytes have been proposed to be a significant source of trophic factors, the pres- ence of which may influence neuronal function and survival [39]. Astrocytes have been also hypothe- sized to participate in the immune responses in the CNS, possibly through interactions with microglia [191.
Altogether, the spatial positioning of astrocytes in the central nervous system and the metabolic interac- tions with other cell types suggest that astrocytes continuously 'sense' and perhaps even regulate the activity of other neural elements.
3. Reactive astrogliosis
Undoubtedly, the most conspicuous property of astrocytes is their morphological transformation in response to virtually any type. of neural insult. This aspect of astrocytic function has been studied exten- sively (see [15,27,40,43,51,53] for recent reviews), but its functional significance is vague in most types of CNS perturbations.
The most common reaction of astrocytes to injury in CNS is the striking hypertrophy and, occasionally, proliferation of these cells that is refe:~ed to by several synonymous terms such as 'reactive as- trogliosis', 'reactive astrocytosis', 'fibril~ary gliosis' or simply 'gliosis'. The hypertrophy "involves an increase in the size of the soma as well as an increase in the length and width of astrocytic stellae, and is accompanied by a rapid reorganization of the cytoskeleton, with the hallmark accumulation of in- termediate filaments (IFs). The principal protein, which constitutes IFs in astrocytes in mature organ- isms is glial fibrillary acidic protein (GFAP) [16]. Antibodies to GFAP are widely used to assess the 'reactive state' of astrocytes; an increase in the amount of this protein has been shown to coincide
with the hlcrease in immuporeactivity (GFAP-IR) [31,69]. While it has not been shown uneqtfivocaHy that this cytoskeletaB change always represents an actual hypertrophy of astrocytes, indirect evidence from many studies suggests that this is the case.
Another IF protein, vimentin (Vim). which is present in several cell types of mesenchymal origin, is expressed in astrocytes early in development, but appears to be gradually replaced by GFAP during development [10,58]. In a normal adult brain, subtle amounts of vimentin can be detected immunocyto- chemically in white matter astrocytes as well as in the stellae of radial astrocytes in selective brain regions, while most gray matter astrocytes are not immunoreactive with antibodies to vimentin ~see [35]). Under certain pathological conditions astro- cyles re-acquire immunoreactivity to vimentin [56,64], most likely through an upregulation of vi- mentin synthesis [47].
A considerable amount of knowledge exists on the composition and distribution of intermediate fila- ments, but their functional role in astrocytes and other cell types has been elusive. Intermediate fila- ments are known for their exceptional chemical sta- bility: they seem to withstand harsh extracting proce- dures resulting in removal of all other cellular com- ponents, including other components of the cyto- skeleton [4]. This stability suggests a role for IFs in providing structural rigidity to cells. Another distinc- tive characteristic of IFs is their molecular composi- tion: they are made up of different, albeit highly homologous, cell type-specific proteins [63]. This cell type specificity may hold additional clues to the functional significance of IFs, since different cell t,,,,,~ have distinct structural requirements.
GFAP is thought to provide structural stability to astrocytes and their processes [16]. The suppression of GFAP synthesis with an antisense construct has been shown to prevent the formation of stable astro- cytic processes in vitro, while a re-introduction of a fully encoding GFAP cDNA into those cells resulted in re-expression of GFAP, formation of a filamen- tous network, and extention of processes in the presence of neurons [7]. Also, the slow turnover rate of GFAP seems consistent with a structural role (see [16]). On the other hand, very recent studies, utiliz- ing mice with a 'knocked-out' Gfap gene, demon- strated that animals with cells devoid of GFAP de-

166 M. Khurgel, G.O. h'y / Epilepsy Research 26 ( 19961 163-175
velop normally [20,54]. While the morphological data are still scant, the abundance and ultrastructural characteristics of astrocytes as well as their response to certain injuries, such as the needle-wound and scrapie infection, appeared normal [20,54]. There was no apparent overcompensatory expression of vimentin in astrocytes in GFAP- mice [54]. Mice lacking vimentin were also recently generated, with no apparent compensatory upregulation of other in- termediate filament proteins [9].
Altogether, the continuing lack of knowledge about the function of intermediate filaments in nor- mal cells precludes the understanding of the func- tional implications of certain cytoskeletal changes, such as the upregulation of GFAP and vimentin syntheses, in reactive astrocytes. At this point, the only certain conclusion to be drawn is that the morphological transformation signals an alteration in the functional state of astrocytes.
4. Astrocytes in epilepsy
Studies of tissue fi'om patients with various forms of epilepsy and the experimental work on animals reveal that an apparent gliosis is almost always present in brain regions which exhibit epileptiforrn activity [45,61,72]. A number of studies in which an absence of observable changes in astrocytes has been reported [6,49] may have utilized inadequate method- ology, resulting in negative findings.
Since necrotic tissue is usually present in most preparations, hypertrophy of astrocytes has been tra- ditionally viewed as the response of these cells to neuronal degeneration. However, based on his stud- ies of the alumina model of epilepsy, Harris [26] proposed that astrocytes hypertrophy in response to hyperactivity of neurons as opposed to acute neu- ronal degeneration. Also, Meldrum et al. [46] de- scribed gliosis as the only pathological change in the hilus of baboons in which seizures were induced with pentylenetetrazol, while Norenberg and Chu [52] observed morphological alterations in astrocytes as the only structural change in the preictal stage in rats treated with aminophylline.
The earliest mention which links morphological alterations in astrocytes and epilepsy is attributed to Chaslin (1889, cf. [45]). Later, Penfield [55] sug-
gested an active role for astrocytes in epileptogenesis by proposing that the 'scar' tissue, which surrounds the area of cortical damage and with reactive astro- cytes as a prominent component, constitutes the structural basis of post-traumatic epilepsy through 'irritation' of neurons. The concept of physical inter- ference of hypertrophied astrocytes with neuronal activity was also argued by Ward [71], who sug- gested that gliosis caused a mechanical deformation of dendrites, thereby resulting in enduring depolar- izations, which in turn gave rise to high frequency neuronal discharge, possibly evoking seizure activ- ity.
Pollen and Trachtenberg [59] advanced an alterna- tive hypothesis that the decisive factor in the devel- opment of post-traumatic focal epilepsy was a break- down of the ability of astrocytes to control extracel- lular K + levels, whereby a build up in K + woulo result in hyperactivity of neurons. This hypothesis, while provocative, has riot received much experimen- tal support [30,42]. Instead, morphologically modi- fied astrocytes may directly contribute to the process of epileptogenesis through resulting changes in ex- traceilular space or changes in the metabolic proper- ties of these cells. Indeed, astrocytes at the epilepto- genic foci have been shown to present an altered enzymatic profile. Du et al. [13] recently have pre- sented evidence that reactive astrocytes in the stimu- lation-induced status epilepticus model display ele- vated immunoreactivity with antibodies to kynure- nine aminotrasferase and other enzymes. Similarly, Tiffany-Castiglioni et al. [67] have shown an in- crease in glutamine synthetase activity following FeCl2/pentylenetetrazol-induced seizures. There may also be alterations in the levels of carbonic anhydrase, which is thought to be vital for the propea" anion and acid-base balance in the brain [23].
5. Astrocytes in kindling
The kindling model of epilepsy is particularly well suited to study the involvement of astrocytes in epilepsy because (1) the development of seizures is subject to experimental control, and therefore allows evaluation of changes in astrocytes at different stages of epileptogenesis, (2) this model is widely accepted as a valuable tool for testing epileptogenesis follow-
M . Khm,~,c l , (7. O. h ' y / E t> ih ,psy R e . + c a r ¢ ' h 2 6 ~ ] 9 9 5 ~ 1 6 3 - ] 75 1 6 7
ing exper imemal interventions, and thereR~re al lows evaluation of strategies designed to directly tes~ the significance o f gliosis in epilepsy.
Brotchi and colleagues [61 were the first to use the
kindling model to address the involvement of astro- cytes in epileptogenesis. Previous work by that group has shown that epileptic loci contain a distinct sub- type of reactive astrocytes, which has been referred to as "activated' astrocytes [5]. Activated astrocytes apparently contain elevated levels of certain dehy-
drogenases, bu¢ ~d~erwise are indistinguishable f lom "reactive + celts, ~he +'+ + +ader reactin~ ~o a mechanical trauma of CNS, when examined with either CajaFs gold-suNima~e method or amiserum to GFAP [14]. (+o~sequen~Iy, activmi~>.+~ of astrocy~es was proposed to be closely rela~ed +to the development of an epilep- togenic focus: however, no e n z y m a t i c a l y or morpho-
logicalb' activated asIrocytes were found in the brains of kindled rats [6]. |rl contrast, we observed a strik- ing hypertrophy of astrocytes in the kindling model
+.
C
w
• ' " '" + . ~ g ' + + 7 ' . . f .
. - . .
i <D:
D " " " . . . . + • ,+ . . - ; '++ ~+, + • _~ -'p + , ~ + + . , . ' + .
+.++- ~ . / '~ . ,.,~ , . . + ; ,+
% . m " ~ + . ' o .
~ p , ; ' < - : . ' f ' ~ t +.1 e' + - J / + ;;9++" " + L o • I ? ~ '
• ; " '+, ~ + " + ' .:~+M" ,+ -+-. + .
~ , v "~' :S ' ~ ~ , ~ . " ~ j : " • --? ,~:_)P ,, + ..,, +<.
- ~ +'+" .+t- ~ ' "+ ~ " "; . . . , ~,-++ -.,Y
~,~+ t"'°Ja~t% ¢+" ~'~" +'+- ..... ' "" " ~ " J. ~ ", " • ~" " + : ~'~L" +'A ' p • .,+ "+ +
, ............- .... , :-'. - ,.. , . : L + :
>
' ".. " ' --.:++' ~-L + "" . b ". ;+2 ~, . .' _ ." ~ , " +. - -+~ - .'0 ,+ +:~:"+ ~ .~" . +" " "i • - + , ' ~ k,
• ...+-~-:.. - ,. -~. .... ..-**~ .. . . . . . _~, . ~'-- ,.~ -_
• . .+ • t-~- , +':.. " 3 " ~ . . ~.. , ,' . o. ++ ," ++ ++ ." .
• + "+ ~, , 0 ,:~-'+ . . + . •, • ~,+ • . . - - ~ ,," : + .~,.~. -. + ~ . ~- 7/ + ~,.+% +:= + , : : ~ , . , . ~ ++ iv ,
• ~ , , -+ , + . . . + . ~ + + " , "++~ ~ . ".~iL r + . + +++ . - . + "~/: +,.+j P . ..., ~- + .~' .+- ++, g-e.+ . • ' "* " " "~ +" -- " '~ .'~ "+ .... " :. " ,-:','.-. iV;.- . @ " .++ ". .
• -,t., ++ . • ,, , .,...;,v+.~,,+~+~+z++,+~-,....+.-,,,v.+'~..~. _ _ _ - ' 7 ..... 3' " + " "".."- :":'r+ +++':+ " " ++ "- .'*l:,~ ~,p ++ .+ +', ' "+'.++ , ~'+ " " ' . ." ,"+.: -,~ .', it,..,+ . ] + ¢
z , ' . , - . + ' ~ . . + , , ' ] / , , + ' , . , + . " , . ~ , + , , ' ~ , + + . : : + - - . + ' ~ P , + + , ~ ' + ~ - . , -
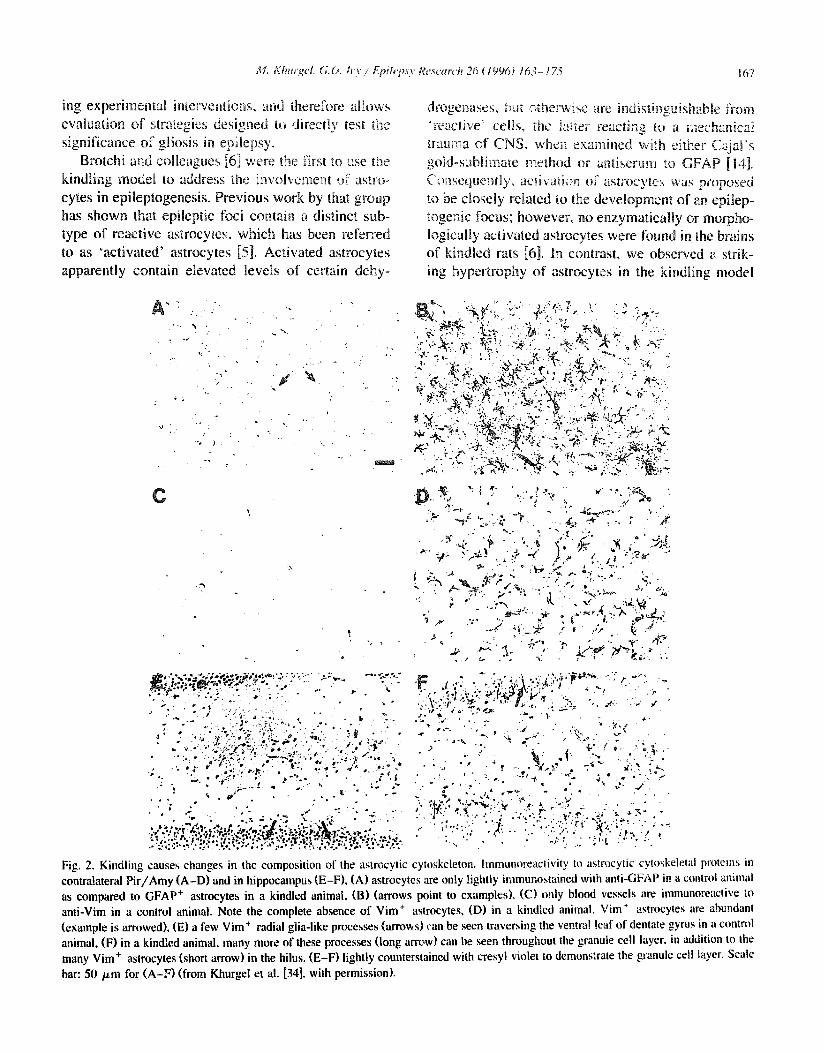
Fig. 2, Kindling causes changes in the composition of the astrocytic cytoskeleton, hnmunoreactivity to astrocytic cytoskeletal pl'oteiils ill conlralateral P i t /Amy (A-D) and in hippocampus (E-F), (A) astrocytes are only lightly immunostained with anti-GFAP in a control animal as compared to GFAP + astrocytes in a kindled animal, (B) (arrows point to examples), (C) only blood vessels are immunoreactive to anti-Vim in a control animal. Note the complete absence of Vim + astrocytes, (D) in a kindled animal, Vim + astrocytes are abundant (example is at'rowed), (E) a few Vim + radial glia-like processes (arrows) +.'an be seen traversing the ventral leaf of dentate gyrus in a control animal, (F) in a kindled animal, many mote of these processes (long am>w) can be seen throughout the granule cell layer, in addition to the many Vim + astrocytes (short arrow) in the hilus. (E-F) lightly counterstained with cresyl violet to demonstrate the granule cell layer. Scale bar: 50 p,m for (A-F) (from Khurgel et al. [34], with permission),

168 M. Khurgel, G.O. It'y / Epilepsy Research 26 f 1996) 163-17.5
of epilepsy. Kindling-induced seizures produced long lasting structural changes in astrocytes within brain regions that participate in kindling-evoked seizures, most notably hippocampus (Hipp), piriform cortex (Pir) and amygdaloid nuclei (Amy) [34-37,60].
In addition to the increase in GFAP-IR (Fig. 2a and b" Fig. 3), a strong de novo immunoreactivity to vimentin (Vim-!R) was found [35] (Fig. 2c-f). While vimentin might be present under certain conditions in activated microglia [21], in kindled brains the newly expressed vimentin was localized to astro- cytes: all Vim + cells were also GFAP + [35]. Also, double-labelling with lectin revealed no Vim + mi- croglia, in agreement with our finding that microglia were not morphologically activated in kindling [37]. The fact that activated astrocytes in an epileptogenic focus re-express vimentin is potentially significant for the pathological assessment of gliosis in epileptic patients. Since traditional methods of evaluating gliosis are usually not satisfactory [45], de novo expression of vimentin in astrocytes may prove help- ful in differentiating between reactive and otherwise apparently hypertrophied astrocytes.
The increase in GFAP-IR in kindling is accompa- nied by increases in GFAP mRNA [65] and total GFAP [24,68], as well as an increase in the volume fraction of astrocytic stellae [28], suggesting that the
immunocytochemical results reflect a true hypertro- phy of these cells.
Morphological changes in astrocytes can be ob- served in the early stages of epileptogenesis and may be seizure-intensity dependent, with a higher inten- sity of Vim-lR and a more widespread activation of astrocytes occurring in fi l ly kindled animals, com- pared to p,-,viz,!iy kindled ones (i.e. those with only stage 1 or 2 seizurcs~ [36]. Howcver, in some cases, changes in GFAP are similar in partially a,,,,! in fidJy kindled anm,als [24,37]. Levels of GFAP are stil! significantly elevated between l and 2 weeks [25,68], and up to a month [34] following the last seizure, but seem to subside by two months following the com- pletion of kindling in most areas [25]. The long-term persistence of Vim-IR has not been addressed yet.
Regional variation in activation of astrocytes likely reflects differences in the spread of seizures due to the choice of a particular kindling paradigm. In most experiments, astrocytic reaction occurs bilaterally [24,34,35], but activation confined to the stimulated side only has also been observed [37,65] (see Fig. 3). Astrocytes in Amy and Pir are activated in most preparations: this phenomenon has been observed in amygdala kindled rats in our laboratory [34,35,37,60], in hippocampus kindled rats [24] and, recently, in septum kindled rats [36], whereas activation of astro-
% increace GFAP-IR
2O
~'t ~ Ipsllateral ~ Contralateral
15
T 10
PlR ENT AMY HIL DG CA1
Fig. 3. The effect of kindling on GFAP-immunoreactivity (IR). An increase in GFAP-IR (expressed as percent difference in GFAP-IR [optical density units] between kindled animals and unstimulated controls) is evident in all measured regions, and is significant, ( ' ) p < 9.05, ( * * ) p < 0.01, on the ~timulated side of kindled animals in p i r i fe~ cortex (PIR), ent'arhinai c~rtex (FNT), has,medial amygdaloid nucleus (AMY), hilar region of hinpoeampus (H!L) and molecular layers of dentate gyms (DG) (from Khurgel et al. [36] with permission).

M. Ktmrgel. G.O. h 3' / Epilepsy Re~ear~'h 26 ~ 1996 ~ / 6 3 - 1 7 5 ~ 6 9
cytes in other brain regions may depend on the placement of the stimulating electrode.
Not all proteins are upregulated in astrocytes in kindling: Hanson etal . [24] have observed a decrease in the levels of Sl00 in Pit and Amy, and no change in the l o r d s of glutamine synthetase 24 h following hippocampal kindling, while Tiffany-Castiglioni et al. [67] have reported a significant decrease in glu- tamine synthetase activity in the ipsilateral neocortex 4 weeks following amygdala k i n d l i n g .
It is important to stress that the changes in astro- cytes described above have been observed in the absence of overt neuronal degeneration [28,37,65]. [n addition, we have been able to demonstrate that activation of astrocytes in kindting occurs in the absence of microglial activation, which otherwise would be indicative of neural trauma [37].
6. Ce|l pra|iferation
Based on their finding of unchanged levels of glutamine synthetase following kindling, Hanson et
a!. [24] concluded that a change in the number of u ~ H ~ l q e ~ v . a_i ~ c ~ l JaJo j as~rocytes is "'-~: " "" '~ . . . . . . . . . . . , ~-nsv,, .~tudy by ~ ' ' - -
et at. [ i l l failed ~o uncover significant differences in the number of GFAP" profiles in selective areas of the hippocampus in kindled rats.
We used a direct test of cell proliferation by injecting the animMs with bromodeoxyuridine (BrdU), an analog of thymidine [22], 7 days after completion of amygadMa kindling and 2 h prior to eulhanasia. Frozen brain sections were reacted with anti-BrdU (1:206~ Boehringer Mannheim), anti- GFAP or anti-Vim [35] or processed for lee,in his,o- chemistry [37,66]; several double-labelling combina- tions were also performed.
Some of the resul,s are shown in Fig. 4. Com- pared to the low levels of cell proliferation in normal brains (Fig. 4a and c), there was a dramatic increase in the number of proliferating cells following kin- dlh~g, mainly in the P i r / A m y region (Fig. 4b) and in Hipp (Fig 4d). It should be noted that since the animals were 'pulsed' with BrdU on one occasion only, the current evidence of kindling-induced cell proliferation is probably an underestimation of the
l "" : " " " " "
, " : : : : .... % , . • i " -:. ' .:, : . . . . •
• ~ . , : . x ' . , , . . ,~ ~ ; ; . : . . e . " . . ",. o ."O,
' , " ' . , , ' , *M " " . " . ' - . , e , . 7 . ~ V ' -' , ' . :
", : , " : ~'~ • . , . : 2 . , : '."" ".-, : , . ' . : . " " ' : . ~ W ' " , . ' ;".~ ; ,. . . . . . . . . ; . . , . ~:'~ " . . ' . . . . ,.,~,~',~.,"'""~" ' . _ ' " ' '~ , . • . ' , " . . . . , ' . ' . ' . . ; r ~ : . - : ! : , . ' : ' , • . - . , . . " " , . . . . . . . . . . . . ' . . ~ .. :,:.. ':.:,-~~/...g.~,,.: ~ / : . . ' : ! . . . . . . . . .
, : • , : . , : " - - i • - ~" t , .~-:,-' ,- .~,' , ':2 ~ ; - - "
° , ~ . % . • , . . - . o . , . ~ . . .
' - d ~ ; : ? t . ' ' . ' -~ ":.~,,"~,:. ~. "~" ~ , - . ' . : . ' , , ~ , • . i , , . : ':
. . . ~ . . . . " . ~ . . . . . o . . ~ " . t t , $ . . : . , ° ° : . X ° . . • % , . ° . .
- b : ' " ' : " : - ~ ° " • v ° . ~ - , t . . - •
" ' J , " a . " • • t ,
i o " - ' ¢ , , + % - e " " • ~ ' " ' ~ •
• , , L . .~ . ~ , 0 • , l f , ~ . ~ , . . . . . e
Fig. 4. Kindling induces cell proliferation• Occasional proliferating cells (arrows) are labeled with BrdU in the Amy/Pir region CA) or hippocampal hilus (C) of unstimulated control animals. Following kindhng, there is all increase in the number of BrdU + cells in Amy/Pir (B} and hippocampus (D) and other brain regions• Note the large increase in the number of proliferating cells in the subgranular layer of the hilus between control rats Cstraight arrows in (C) and kindled rats (D). Many cell pairs Ccurved arrow in CC)) can be seen in the Amy/Pir and hilus of kindled rats, suggesting that BrdU + cells are indeed proliferating cells. Open arrow in CA) and CB) outlines the pyramidal cell layer of the piriform cortex (pir). Scale bars: approx. 120 tzm for (A), CB); 60 v,m for (C), (D).
170 M. Khurgel. G.O. h'y / Epih, psy Research 26 (1996) 163-17.5
number of new cells. Some of the BrdU + cells were also GFAP + or Vim +, but none were lectin +, while others could not be identified with the methods used here (Khurgel et al., in preparatiot~). It is conceivable that in addition to cell division in astrocytes, seizure-associated factors induce proliferation of pre- cursor cells which later differentiate into astrocytes. On the other hand, the dramatic increase in the number of BrdU + cells in the subgranular layers of dentate gyrus (Fig. 4d), may be indicative of an increase in proliferation of granule ceils, which con- tinue dividing in low numbers in adult rats [32]. The latter is a plausible explanation, especially since we have previously reported an increase in Vim + radial gila-like processes in DG foiiowing kindling [35] (Fig. 2f).
Incorporation of BrdU into DNA may be indica- tive of an extensive DNA repair in cells presumably damaged by the seizures in addition to (or instead of) DNA replication/cell division. This possibility re- mains to be explored. However, the presence of adjacent BrdU + 'cell pairs" (curved arrow, Fig. 4c) in normal and kindled animals suggests that BrdU + cells are indeed proliferating cells.
Altogether, these findings show that kindling-in- duced seizures result in increased cell division and present an intriguing possibility that proliferation of astrocytes can be upregulated with elevated neural activity, in the absence of extensive neural damage.
7. Expression of connexin-43
Changes in the astrocytic network, partially repre- sented by astrocyte hypertrophy and proliferation, potentially have important implications for epilepto- genesis. A related significant change could be an alteration in coupling between astrocytes.
Increased numbers of cellular connections, both desmosomes and gap junctions, between astrocytes seem to be present in alumina-induced epileptic foci [26]. Naus et al. [50] have shown an increase in the expression of connexin-43 (Cx43), the main protein in astrocytie gap junctions, in tissue excised from epileptic patients. It has been argued that astrocytes cultured from such tissue exhibit repeated oscilla- tions in response to a pulse of glutamate [8], suggest- ing an increase in excitability of, and perhaps an
increased coupling between these cells. An augmen- tation in coupling between astrocytes could con- tribute to epileptogenesis by participating in hyper- synchronization of neuronal firing. This argument is
C
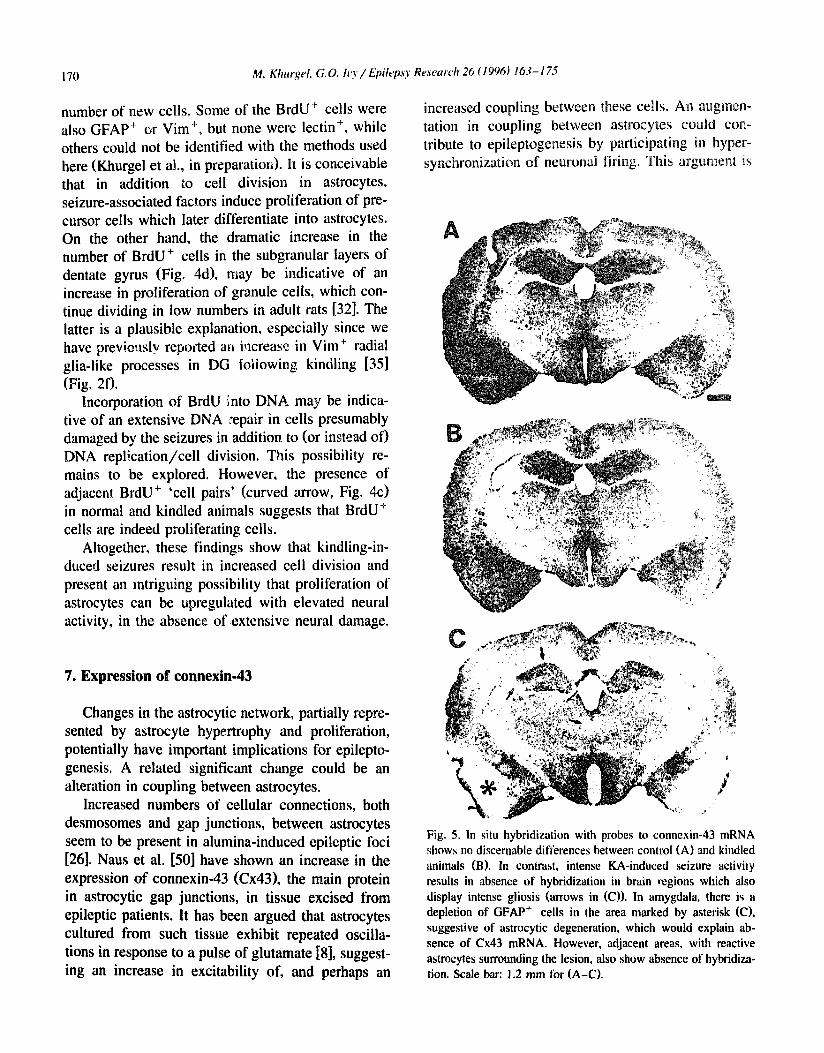
Fig. 5. In situ hybridization with probes to connexin-43 mRNA shows no discernable differences between control (A) and kindled animals (B). In contrast, intense KA-induced seizure activity results in absence of hybridization in brain regions which also display intense gliosis (arrows in (C)). In amygdala, there is a depletion of GFAP + cells in the area marked by asterisk (C), suggestive of astrocytic degeneration, which would explain ab- sence of Cx43 mRNA. However, adjacent areas, with reactive astrocytes surrounding the lesion, also show absence of hybridiza- tion. Scale bar: 1.2 mm for (A-C).

M. Khu rgel, G. 0 h 3/EpikTzS3' Re.search 26 ~ t 996 ~ 163-175 t 71
supported by the data from a recent study in wt:ich astrocyteso isolated from hyperexcitable tissue, have been shown to display a fast f~uorescence recovery after photobleach, and therefi~re thought to be exle~- sively coupled [41 ].
We and our colleagues have conducted a pilot study aimed at assessing the expression of connexin- 43 in kindling. Frozen brain sections from kindled rats were processed for in situ hybridization using Cx43 cRNA probes as described elsewhere [2]. The sections from kindled animals were compared to those from implanted, but unstimulated control and a positive control, in which I h of status epilepricus was induced with systemically-administered kainic acid (KA). The examples of respective patterns of hybridization are presented in Fig. 5. No aifferences in Cx43 mRNA distribution were apparent between the unstimulated control and kindled animals. How- ever, a clear depletion in Cx43 mRNA was observed in areas ot i nfense gliosis ip the KA-treated animal. Specifically, regions where a marked neuronal de- generation is known to take place, such as midline thalamic nuclei, nucleus reuniens, hippocampal areas CAI and CA3, neocortex, and the Amy/Pi t area all show an absence of hybridized probe (arrows, Fig. 5c). Since a prolound astrocyte hypertrophy is pre- sent in these areas, the lack of hybridization suggests a shut-down of Cx43 synthesis in reactive astrocytes in the KA model. At the same time, the absence of a significant difference in hybridization between con- trol and kindled animals suggests that kindling- activated astrocytes do not significantly upregu|ate or downregulate their expression of Cx43. It is possi- ble, of course, that the method employed here is not sensitive enough to detect subtle changes in levels of this mRNA. Thus, additional work is required for precise quantification of potential changes in expres- sion of Cx43. It is also conceivable that there are changes in physiological coupling between astro- cytes independent of levels of Cx43. This possibility awaits experimental confirmation.
8. Recent advances
The evidence is accumulating that astrocytes un- dergo a variety of changes during or following kin- dling-mediated epileptogenesis (see Table 1.)
Table I
Summary of cha~lges m aslrocy'les il~ kindbng
Observed change Ref.
Prolifcralion of astrocyles ? I Khurgel and I',y} GFAP-IR ~ [I 1,35,37] Levels of GFAP ~ [24,25,68] Levels of GFAP mRNA ~ [65.68] De novo Vim-|R {35.37] Hypertrophy of aslrocytes [28] Cx43 mRNA n.c. (Khurgel and ivy) GS n.c./11 [24,67] SI {H} 11 [24] Glial muscarinic recept. ~1 [i}
no. No observed change. GS Glulamirm synthetase.
What, then, is the significance of these changes with respect to development of seizures? Is it possi- ble that some of these structural changes reflect a functional transformation of astrocytes, which in turn leads to inadequate control of the extracellular envi- ronment, similar to the deficiency in potassium buffering or other metabolic abnormalities proposed for 'glial scar' astrocytes?
Recently, we have been attempting to address these questions using novel experimental approaches. We ha,'-, decided to evaluate the significance of altered astrocytk morphology hi epileptogenesis ei- ther by activating astrocytes or by selectively remov- ing these cells from the seizure-initiating site prior to commencement of kindling.
Seizures have been shown to result in increased levels of various growth factors in the brain regions conveying the seizure activity [3,17,! 8,62]. Among the upregulated proteins is the basic fibroblast growth factor I.bFGF) [ 18,62], which has been shown to be a potent mitogen and stellation factor for astrocytes [57]. It is conceivable that, among its other functions, bFGF serves as a signal for astrocyte hypertrophy following perturbations of CNS.
We have injected bFGF (Sigma; 0.25 /xg in 5 /xl artificial cerebrospinal fluid) into Amy of naive rats in an attempt to mimic the seizure- or trauma-media- ted induction of gliosis. Injection of bFGF results in increased GFAP-IR and Vim-IR, and apparent hyper- trophy of astrocytes throughout the Amy nuclei as early as 48 h post injection (Fig, 6a and b). The

172 M . K h u r g e l , G . O . h ,3 ' / E p i l e p s y R e s e a r c h 2 6 ( 1 9 9 6 ) 1 6 3 - 1 7 5
A
, . . , .
, : : .~ ~ ; . , . . . , , - ~ . ~ , ~ .~,¢.,~ ~ '~ . .~ ' . . ' . , , ~e , ~ ,~. . . . . . . . . . . . . . . ~ : - ~. . . . . . . . . , , - ~ . ~ :, ~ ~ . ~-:. .~., ,~, ~ : ~ . ~ " . ' i ~ ' ~ . , ~ . . ~, .., '~':;.z7 ' ,
-,~,_. , , ,~. - , " - ~ . - ~ ' ~ : ::~ - : - ~ . ~ - - :~...~.~.~. , ,., . . . . . : . ,~ ~ . . ~. ~ • " ~ . . . ~ I - ~s . ' ~ ; / " k ~ " ' ~ * , • ~ .
..... - ~ . . . . V . ; ~ ~.- . . . . ~.
"~."" --,-'~ .. • ,. ' . ~ :" -'~-,.~..K. ! , ' - ~ 7",....'- - "~"' "7:- ~ ' .' ~ --?.. " , " , • ~ , , . : ' , . . . . - . . ~ ..~ ~ - ~ : . , ~ . ,. ~ - • p : . - . , = , , : , - - ~ - ~ , ,*~.,.
.~. ,. . . . . . t ~.-'-.~%. -a~ ".~": "'. " ,:',, • '; "~',~ ' ' ~ "~" ~ " ::- ' - ~ ' ~ d~. ~ ' ~ ~ ¢ ' v ~ . . , .-~. ~ ' . x ~ . ~ -. •
~:~,~"~'J i . ,~. ~ ~v~!:.~: : . ~ .f/g~. ~" :" .~ ~;~:'~ "~" "¢~': ~ :~t~ ~'- -, ~ " " ~-~i~; e ¢ ) : ~ , ~ . l , , ~ ";"',y :~ ~,,~, '. '.~.'<" " "'::'.,~, ~: : ~ .,~'~" :-~ _ . , ' ' : ' ~ . ~"~'~"~"':: ~ : ~-::~:~, ":'..,.~_,':'" _ F~,~ ,~.',t~ ~ ~ ~ . ~i/~'~.~" ~ ~ ~ ! 2 ~ - ) ' : ) ' ~ ~ " ' f ' ' ~ ~ , .~ ~ .~,~, ~ i ~ . ~ , ~ . ~ , ~ . , , . x ! .:~ ,.,~ ~.=,.~':: ' ..~ ' ~" ~"~" " " c . . . . '~ " ' " '" ''~''; ~"
• ~ , ~ , - ~ . . . . . , . . . . . . . . . ~ ~ ' . ~ - ? ~ : " ; " ~ " . , . , ,
Fig. 6. Experimental manipulat ion o f astrocytes: (A and C) G F A P * as t rocytes in the amygda la o f naive rats, (B) injection o f b F G F results
in increased GFAP- IR and apparent hyper t rophy o f astrocytes, (D) injection o f a -aminoad ipa te resul ts in the ablation o f as t rocytes in the
area o f i~jection (aAA), whi le sparing the neurons. Sect ions in (C and D) were counters ta ined wi th c resyl violet. Scale bar: approx. 6 0 / x m
for ( A - D ) .
intensity of astroglial reaction is similar to the one observed in kindling. We are currently investigating the effects of this treatment on development of seizures. The results obtained so far show that the animals, treated with bFGF, do not display any overt behavioral seizures. Also, monitoring of animals im- planted with recording electrodes following similar injections of bFGF reveals an absence of epilepti- form activity (Khurgel et al., in preparation). The properties of bFGF-activated astrocytes with respect to the 'type' of induced gliosis remain to be deter- mined. Nevertheless, these preliminary data argue against the hypothesis that gliosis is in itself epilep- togenic.
Another promising approach that we are pursuing is the examination of neuronal function and develop- ment of seizures in the absence of astrocytes. We have recently characterized in vivo the selective astroglial toxicity of a-aminoadipate [33]. Iajections of (~-aminoadipate into Amy of rats result in selec- tive ablation of astrocytes, without affecting neuronal density [33] (Fig. 6c and d). The consequences of
this treatment on the development of seizures are also currently under investigation.
9. Summary and conclusions
The role of astrocytes in epilepsy has been a long standing, albeit not very prominent, line of inquiry in the field of epileptology. The kindling model, which is widely used to explore various aspects of seizure development, has been utilized to discover extensive changes in astrocytes during epileptogenesis. The proliferation, hypertrophy and cytoskeletal changes in astrocytes in kindling are reminiscent of the clas- sical 'gliosis', observed in post-traumatic epilepsy and in long-term epilepsies with various aetiologies. However, there may exist e~sential differences be- tween those phenomena. In particular, there is a potential diversity in metabolic coupling and enzy- matic profiles of reactive astrocytes in different per- turbations of the CNS, which remain to be explored.
Circumstantial evidence links the changes in as- trocytes to the development of an epileptic state, but

M. Khurget, G.O. b'y / EpihT~sy Research 26 ( t 990) t63- / 75 173
direct tests of the contribt|~ion of astrocytes to epflep- togenesis have been lacking until recently. New find- ings suggest that the presence of reactive astrocytes is not necessarily epileptogenic. The research in progress may yield additional answers with respect to the invoDement of these ceils in seizure disorders.
Acknowledgements
The authors would like to thank S.C. Barsoum and A.C. Koo for their assistance with some of the above studies, Drs. D.J. Belliveau and C.C. Naus for performing the Cx43 mRNA assays, and Drs, R.J. Racine and N.W. Milgram for their invaluable con- tributions to this research. We also thank Drs. N.W. Milgram and O. Steward for discussions and helpful comments on this manuscript. Much of the above work has been supported by NSERC grants to G.O.L and, in part, by the Epilepsy Canada 1995 Canadian National Studentship award to M.K. The presenta- tion of this work at WONOEP III was generously supported in part by Ciba-Geigy Ltd, Canada, the Optimist Club of Ajax, Ontario, and a travel grant from the Department of Anatomy and Cell Biology. University of Toronto to M.K.
References
[I] Beldhuis, H.J., Everts, H.G., Van Der Zee, E.P,., Luiten, P.G. and Bohus, B., Amygdala kindling-induced seizures selec- tively i..pair spatial memory. 2. Efl~ects on hippocampal neuronal and glial muscarinic acetylcholine receptor, Hip- pocampus, 2 (1992) 411-419.
[2] Belliveau, DJ. and Naus, C.C., Cellular localization of gap junction mRNAs in developing rat brain, Dev. Neurosci., 17 (1995) 81-96.
[3] Bengzon, J., SiiderstriJm, S., Kokaia, Z., Kokaia, M., Em- fors, P., Persson, H., Ebendal, T. and Lindvaii, O., Widespread increase of nerve growth factor protein in the rat forebrain after kindling-induced seizures, Brain Res., 587 (1992) 338-342.
[4] Bershadsky, A.D. and Vasiliev, J.M., Cytoskeleton. Plenum Press, i~rew York, 1988.
[5] Brotchi, J., Scuvee-Moreau, J. and Dresse, A., Corrrlation entre le caract~:re (~pileptog~ne d'un foyer i~sionnei et la presence dans celui-ci d"astrocytes attires" (pr~sentant une activit~ tr~sintense des d~shydrog~nases et de l'alpha- glucan-pbosphorylase). Etude chez I'homme et l'animak Acta Neurol. Belg., 78 (1978) 267-278.
[6] brotchi, 1., Tanaka, T. and Le'~i¢l. V., Lack ~ activai~d a~m~cvte~ in the kindIine phenomenon. E.W~. NeuroL, 5g (19781 n ]9-125.
[7] Chen, W.-J. and Liem, R.K.H.. Ree×pression of glial fibrib lary acidic pr~ein re~ct~es the ability of asm~cytoma cells to fi~rm processc;, in response l~ neurons, d. Cell Biol,, ~27 119941 813-823.
[8] CornelbBell, A.H. and Williamson, A., Hyperexeitability of neurons and astrocytes in epileptic human cortex. In: S. Fedoroffo B.H.J. luurlink and R. Doucetle (Eds.). Birdogy and Pathology of Astroo,te-Neuron Interactions, Plenum Press, New York, t993, pp. 51-65.
[9] Colucci-Guyon, E., Porlier, M.M,. Dunia, [., Panlin, D., Pournin, S. and Babinet, C., Mice lacking vimentin develop and reproduce without an obvious phenotype, Cell, 79 (1994) 679-694.
[10] Dahl, D., Rueger, D.G., Bignami, A., Vimen|in, the 57 000 molecular weight protein of fibroblast filaments, is the major cytoskeletal component in immature glia, Eur. J. Cell Biol.. 24 11981) 191-196.
[11] Dalby, N.O., Rondouin, G. and Lemer-Natoh, M., Increase in GAP-43 and GFAP immunoreactivity in the ra~ hippocam- pus subsequent to pefforant path kindling, J. Neurosci. Res., 41 (1995) 613-619.
[12] De Robertis, E. and Gernschenfeld, H.M., Submicroscopic morphology and function of glial cells, Rec. Neurobiol.. 3 (1961) ~--65.
[13] Du, F., Wiilimnson, J., Bertram, E., Lothmam E., Oknno, E. aud Schwarcz, R., Kynurenine pathway enzymes in a rat model of chronic epilepsy: immunohistochemical study of activated glial cells, Neurosciem'e., 55 (1993) 975-989.
[14] Duchesne, P.Y., Gheuens, J., Brotchi. J. and Gerebtzoff, M.A., Normal and reactive astrocytes: a comparative study by immunohistochemistry and by a classical histological technique, Cell. Mol. Biol., 24 (1979)237-239.
[15] Eddleston. M. and Mucke, L.. Molecular profile of reactive astrocytes: implications tot their role in neurologic disease, Neurosci., 54 (1993) 15-36.
[16] Eng, L.F. and Lee, Y.L., Intermediate filaments in astrocytes. In: H. Kettenmann and B.R. Ransom (Eds.), Neuroglia, Oxlord Univ. Press, New York, 1995, pp. 650-667.
[17] Gall, C.M. and lsackson, P.J., Limbic seizures increase neu- ronal production of messenger RNA for nerve growth factor, Science, 245 (1989) 758-761.
[18] Gall, C.M., Berscbauer, R. and lsaekson, P.J., Seizures in- crease basic fibroblast growth factor mRNA in adult fore- brain neurons and glia, Mol. Brain Res., 21 (1994) 190-205.
[19] Gehrmann, J., Matsumoto, Y. and Kreutzberg, G.W., Mi- croglia: intrinsic immunoeffector cell of the brain, Brain Res. Rer., 20 (1995) 269-287.
[20] Gomi, H., Yokoyama, T., Fujimoto, K., lkeda, T., Katoh, A., Itoh, T. and ltohara, S., Mice devoid of die glial fibrillary acidic protein develop normally and are susceptible to scrapie prions, Neuron, 14 (1995) 29-41.
[21] Graeber, M.B., Streit, W.J. and Kreutzberg, G.W., The mi- croglial cytoskeleton: vimentin is localized within activated cells in situ, J. Neuroqvtol., 17 (1988) 573-580.

174 M. Khurgel. G.O. h'y / Epilepo' Reseatz'h 26 (1996) 163-17,5
[22] Gratzner, H.G., Monoclom.l antibody to 5-Bromo- and 5- lododeoxyuridine: a new reagent tbr detection of DNA repli- cation, Science, 218 (1982) 474-475.
[23] Grisar, T., Guillame, D., Bureau, M. and Heeren-Bureau, M., Astroeytes contribution to the epilepsies: molecular aspects. In: M.D. Norenberg, L. Hertz, A. Schousboe (Eds.), The Biochemical Pathology of Astrocytes, Alan R. Liss. New York. 1988, pp. 487-501.
[24] Hansen, A., J0rgensen, O.S, Bolwig, T.G. and Barry, D.I.. Hippocampal kindling alters the concentration of glial fibril- lary acidic protein and other marker proteins in the rat brain, Brain Res., 531 (1990) 307-311.
[25] Hansen, A., Jorgensen, O.S., Bolwig, T.G. and Barry, D.I., Hippocampal kindling in the rat is associated with time-de- pendent increases in the concentration of glial fibrillary acidic protein, J. Neurochem., 57 (1991) 1716-1720.
[26] Harris, A.B., Cortical neuroglia in experimental epilepsy, E.rp. Neurol., 49 ( 1975 ) 69 ! -715.
[27] Hatten, M.E., Liem, R.K.H., Shelanski, M.L. and Mason. C.A., Astroglia in CNS injury, Glia, 4 (1991) 233-243.
[28] Hawrylak, N., Chang, F.-L.F. and Greenough, W.T., Astro- eyfic and synaptic response to kindling in hippocampal sub- field CAl. II. Synaptogenesis and astrocytic process in- creases to in vivo kindling, Brahl Res., 603 (1993) 309-316.
[29] Hertz, L., Possible role of neuroglia: a potassium-mediated neuronal-neuroglial-neuronal impulse transmission system, Nature, 206 (1965) 1091 - 1094.
[30] Hertz, L., An intense potassium uptake into astrocytes, its further enhancement by high concentrations of potassium, and its possible involvement in potassium homeostasis at the cellular level, Brain Res., 145 (1978) 202-208.
[31] Hozumi, I., Chiu, F.-C. and N,~rton, W.T., Biochemical and inununocytochemical changes in glial fibrillary acidic protein alter stab wounds, Brahl Res., 524 (1990) 64-7 I.
[32] Kaplan, M.S. and Hinds, J,W., Neurogenesis in the adult rat: electron microscopic analysis of light radioautographs, Sci- ence, 197 (1977) 1092-1094.
[33] Khurgel, M., Koo, A.C. and Ivy, G.O., Evidence of a localized ablation of astrocytes by intracerebral injection of L-a-aminoadipate. Gila, in press.
[34] Khurgel, M., Racine, R.J., Milgram, N.W. and Ivy, G.O., Progressive changes in the pattern and intensity of astrocyte hypertrophy in the brains of amygdala kindled rats, Soc. Neurosci. Abstr., 16 (1990) 342.
[35] Khurgel, M., Racine, R.J. and Ivy, G.O., Kindling causes changes in the composition of the astrocytic cytoskeleton, Brain Res., 592 (1992) 338-342.
[36] Khurgel, M., Racine, R,J. and Ivy, G.O., Seizure intensity- mediated astrogliosis in septum kindled rats, Epilepsia, 36 (Suppl. 4) (1995) 20.
[37] Khurgel, M., Switzer, R.C., Teskey, G.C., Spiller, A.E., Racine, R.J. and Ivy, G.O., Activation of astrocytes during epileptogenesis in the absence of neuronal degeneration, NeurobioL Dis., 2 (1995) 23-35.
[38] Kuglcr, P., Enzymes involved in glutamatergic and GABAer- gic neurotransmission, Int. Rev. Cytol., 147 (1993) 285-336.
[39] Labourdette, G. and Sensenhrenner, M., Growth factors and
their receptors in the central nervous system. In: H, Ketten- mann and B.R. Ransom (Eds.), Neurt:,¢lm, Oxford Univ. Press, New York, 1995, pp. 441-459.
[40] Landis, D.M.D.. The early reactions of non-neur~ma] cells to brain injury, Ann. Rec. Neurosci., 17 (1994) 133-151.
[41] Lee, S.H.. Magge, S., Spencer, D.D., Sontheimer, H. and Cornell-Bell, A.H., Human epileptic astrocytes exhibit in- creased gap junction coupling, Glia, 15 (1995) 195-202.
[42] Lewis, D.V., Mutsuga. N., Schuette, W.H. and Van Buren, J., Potassium clearance and reactive gliosis in the alumina gel lesion, Epih, psia, 18 (1977} 499-506.
[43] Malhotra, S.K., Aldskogius, H. and Shnitka, T.K., Biochemi- cal diversity among reactive astrocytes, Dendron, I (1992} 55-68.
[44] Martinez-Hernandez, A., Bell, K.P. and Norenberg, M.D., Glutamine synthetase: glial localization in brain, St'ieme, 195 (1977) 1356-1358.
[45] Meldrum, B.S. and Bruton, C.J., Epilepsy. In: J.H. Adams and L.W. Duchen (Eds.), GreenfieM's Neurripathology, 5th Ed,, Oxford Univ. Press, New York, 1992, pp. 1246-1283,
[46] Meldrum, B.S., Papy, J.J., Toure, M.F. and Brierly, J.B., Four models for studying cerebral lesions secondary to epileptic seizures, Ade. Neurol,, 10 (1975)147-161.
[47] Mickuki, S.A. and Oblinger, M.M., Vimentin mRNA expres- sion increases after corticospinal axotomy in the adult ham- ster, Metab. Brain Dis., 6 (1991) 33-49.
[48] Montgomery, D.L., Astrocytes: form, functions, and roles in disease, Vet. PathoL, 31 (1994) 145-167.
[49] Munoz, D,G., MeNab, B., George, D.H. and Johnson, D., Absence of gliosis in the brains of epileptic fowl, Can. J. Neurol. Sci.. 15 (IORR} 409-412.
[50] Naus, C.C., Bechberger, J.F. and Paul, D.L.. Gap junction gene expression in human seizure disorder, Eap. Netavl., 111 (1991) 198-203.
[51] Norenberg, M.D., Astrocytc responses to CNS injury, J. Neuropath. Exp. Neurol., 53 (t994) 213-220.
[52] Norenberg, M.D. and Chu, N.-S., Aminopbylline-indueed preictal alterations in cortical astrocytes, Exp. Neurol., 54 (1977) 340-351.
[53] Norton, W.T., Aquino, D.A., Hozumi, I., Chiu, F.-C. and Brosnan, C.F., Quantitative aspects of reactive gliosis: a review, Neurochem. Res., 17 (1992) 877-885.
[54] Pekny, M., Leveer,, P., Pekna, M., Eliasson, C., Berthold, C.H., Westermark, B. and Betsholtz, C., Mice lacking glial fibrillary acidic protein display astrocytes devoid of interme- diate filaments, but develop and reproduce normally, EMBO J., 14 (1995) 1590-1598.
[55] Penfield, W., The mechanism of eieatricial contraction, Brain, 50 (1927) 499-517.
[56] Petito, C.K., MorgeUo, S., Felix, J.C. and Lesser, M.L., The two patterns of rea'five gliosis in postischemic rat brain, J. Cereb. Blood Flow Mdab., 10 (i990) 850-859.
[57] Pettman, B., Weibel, M., Sensenbrenner, M. and Labour- dette, Purification of two astroglial growth factors from bovine brain, FEBS, 189 (1985) 102-108.
[58] Pixley, S.K.R. and De Vellis, L, Trar~sition between imma- ture radial glia and mature astrocytes studied with a mono-

M. Ktmrgel. (1.0. I* y / Epilel~sy R~'searc, 26 (1996) 16,¢ f75 I75
c]onzfl antibody to vimmltin. Del. Bra,;n Re.,., 15 {1984} 201 - 2{19.
[391 Pt;k" ~, D.A. al~d Ttaci~.c,bc~, M.C., Ncmoglk~: e~iosi, a,,d I ,.~..,-1253. focal epilepsy, Scie,ce, 1~7 (t970} ~'~'
[6(I] Racine, R.J., Ivy. G.O. and Milgram, N.W., Kindling: c[ini cal relevance and anatonfica~ substrate, in: T.G. Bo[wig and MR. Trimble (Eds.), The Clini;'¢d Relet'am'e of Kindling, John Wiley and Sons Ltd., Chichester, ]989, pp. ]5--34.
[6|] Ribak, C.E., Cuntempola~y methods in neumcymlogy "a..'vd their application to the study of epilepsy. |n: A.V. Delgado- Escueta, A.A. Ward, Jr., D.M. Woodbury and R.J. Porter (Eds.), Basic Mechanisms o f the Epilepsws, Adv. Neurol.. VoL 44, Raven Press, New York, 1986, pp. 739-764.
[62] Riva, M.A., Gale, K. and Mocchetti, I., Basic fibrob]ast g;owth factor mRNA increases in specific brain regions tbllt,'wing convulsive seizures. Mol. Brain Re.s, 15 {~992) 311-318.
[63] Robson, R.M.. Intermediate filaments, Curt. Opin. Cell Biol., I { 19891 36-43.
[64] Schiffer, D., Giordana, M.]'., Migheli, A., Giaccone, G., Pezzmta, S. ao.d Mauro, A., Glial fibrillary acidic protein anad vimentin in the experimental gila| reaction of the rat brain, Brain Res., 374 (1986~ llO-118.
[65] Steward, O., Torre, E.R., Tomasulo, R. and Lothman, E..
Neuronal acfivhy up-regulates asm',.g[ia] ge,'qe expression, Proc. N:ttt. Ac~id. Sci.. 8g (199]) 6{419-6923.
cells using the ]ectiu from Griffm~ ,simplic~fi~lia (GSA [-B~ k J. 11~.~ hem. (_~vto¢ hem., 38 ( [ 990 ~ 1683- t 686.
[67] Tiffany-Castiglioni, E.C., Peterson, S.L. and Casdglioni, AA.. Alterations in glutamine synthetase activity by FeCle-in- duced focal and kindled amygdaloid seizures. J. Nel,'osci.
[68] Torre. E.R., Lothman, E. and S|eward, O.. Gliai response to neuronal activity: GFAP-mRNA and protein levels are tran- siendy increased in the hippocampus after seizures, I-3ram Res., 63t (~993) 256-264.
[69] Vijayam V.K., Lee, Y.-L., Eng, L.F.. Increase m glial fibrib lary acidic protein t'bllowing neural trauma, MoL Chem. Netwopath., ] 3 (1990) |07- I 18.
[70] W'a]z, W., Role of ghal cells in the regumation of the brain ion microenvironment, Prog. ,ve,r~ btol.. 33 ( 1989} 309-333.
[7]] Ward. A.A.. The epileptic neurone. EFih'psia. 2 ~lq61~ 70 8O.
[72] Ward. A.A., Gila and epilepsy. In: E. Schoffeniels, G. Frank. L. Hertz and D.B. Tower (Eds.), Dynamic Properde~ efGlia Cells. Pergamon Press. Oxford, 1978, pp. 413-427.




















