
Nitric Oxide Donors Suppress Chemokine Production by Keratinocytes in Vitro and in Vivo
Upload
independentCategory
view
2download
0
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r. com/ l oca te / sc r
Stem Cell Research (2009) 2, 56–67
REGULAR ARTICLE
Human neural stem cells and astrocytes, but notneurons, suppress an allogeneic lymphocyte responseElisabet Åkesson a,b, Nina Wolmer-Solberg c, Madeleine Cederarv c,Scott Falci d, Jenny Odeberg a,b,c,⁎
a Division of Neurodegeneration, Department of Neurobiology, Care Sciences and Society, Karolinska Institutet, Novum,S-141 86 Stockholm, Swedenb Stockholms Sjukhem, Stockholm, Swedenc Center for Molecular Medicine, Department of Medicine, Karolinska Institutet, Karolinska University Hospital, Solna,S-171 76 Stockholm, Swedend Craig Hospital, Englewood, Denver, CO 80110, USA
Received 21 January 2008; received in revised form 4 June 2008; accepted 28 June 2008
Abstract Transplantation of human neural stem cells (NSCs) and their derivatives is a promising future treatment forneurodegenerative disease and traumatic nervous system lesions. An important issue is what kind of immunological reaction thecellular transplant and host interaction will result in. Previously, we reported that human NSCs, despite expressing MHC class Iand class II molecules, do not trigger an allogeneic T cell response. Here, the immunocompetence of human NSCs, as well asdifferentiated neural cells, was further studied. Astrocytes expressed both MHC class I and class II molecules to a degreeequivalent to that of the NSCs, whereas neurons expressed only MHC class I molecules. Neither the NSCs nor the differentiatedcells triggered an allogeneic lymphocyte response. Instead, these potential donor NSCs and astrocytes, but not the neurons,exhibited a suppressive effect on an allogeneic immune response. The suppressive effect mediated by NSCs most likely involvescell–cell interaction. When the immunogenicity of human NSCs was tested in an acute spinal cord injury model in rodent, axenogeneic rejection response was triggered. Thus, human NSCs and their derived astrocytes do not initiate, but insteadsuppress, an allogeneic response, while they cannot block a graft rejection in a xenogeneic setting.
© 2008 Elsevier B.V. All rights reserved.Introduction
Neural stem cells (NSCs) have emerged as a promisingtherapeutic tool for patients with neurodegenerative dis-eases and traumatic injuries, such as spinal cord injury (SCI).
⁎ Corresponding author. Division of Neurodegeneration, Departmentof Neurobiology, Care Sciences and Society, Karolinska Institutet,Novum, S-141 86 Stockholm, Sweden. Fax: +46 8 5858 3880.
E-mail address: [email protected] (J. Odeberg).
1873-5061/$ – see front matter © 2008 Elsevier B.V. All rights reserveddoi:10.1016/j.scr.2008.06.002
In experimental models cell therapeutic strategies haveresulted in rescue, repair, and/or replacement of injuredneural cells and tissues. A crucial step toward successfulclinical application of neural cell therapy is to unravel howimmunocompetent in vitro expanded human NSCs are,whether a host rejection response may be induced, andwhether these cells present immunomodulatory and therebyeven neuroprotective effects after nervous system lesions.
Interestingly, NSCs expanded in vitro display dissimilaritiesin the major histocompatibility complex (MHC) expressionpattern, most likely owing to different culture conditions and
.

Figure 1 Cell populations included in the study. (A) A represen-tative micrograph of a neurosphere derived from human forebraincontaining NSCs. NSCs were stained for nestin (red). (B) Neuronalcultures stained for β-tubulin III (green). Inset: β-tubulin III (green)and nestin (red). (C) Astrocyte cultures were stained for GFAP(green). Inset:GFAP (green) andnestin (red). In all sections,Hoechstnuclear stain was applied (blue). Scale bars, 25 μm.
57Human NSCs and astrocytes suppress lymphocyte response
species origin. Newly isolated cells from the human centralnervous system (CNS) express low levels of MHC class I and classII expression (Åkesson et al., 2000). While we have previouslyshown that both MHC class I and class II are induced upon invitro culture of human NSCs (Odeberg et al., 2005), Al Nimer etal. (2004) reported only MHC class II expression, despite similarculture conditions and cell source. The MHC class I expressionseems to be positively regulated by leukemia inhibitory factor(LIF) (Wright et al., 2003), which might explain the relativelyhigh expression of MHC on our NSCs, since they are culturedwith ciliary neurotrophic factor (CNTF) which belongs to thesame cytokine family (the interleukin 6 family) as LIF. Theexpression of MHC class I and class II on NSCs is induced and/orfurther increased by IFN-γ treatment (Odeberg et al., 2005;Poltavtseva et al., 2002;Wright et al., 2003). In contrast,mouseNSCs were reported not to express either MHC class I or class II(Hori et al., 2007; Mammolenti et al., 2004), while rat NSCsexpress only MHC class I (McLaren et al., 2001). The presence ofMHC class I and class II molecules on human NSCs implies a riskfor recognition by alloreactive T cells after transplantation,thus indicating a potential risk for immunological rejection dueto MHC incompatibility and subsequent requirement ofimmunosuppressive treatment to avoid rejection.
The disadvantages of using immunosuppressive treatmentinclude an increased risk for opportunistic infections (Garridoet al., 2006), toxic side effects (Rezzani, 2006), and potentialnegative effects on donor cells. Recently, cyclosporin was foundto reduce cell proliferation and affect differentiation to favorastrocytogenesis of rat neural stem cells in vitro (Guo et al.,2007).However, cyclosporin has also been successfully provenasa neuroprotective drug in SCI (Alonso et al., 1995; Curtis et al.,1993; Diaz-Ruiz et al., 2005; Ibarra et al., 2007) and by blockingthe activity of romatase, a calcineurin-independent enzyme(Sosa et al., 2005; Wei et al., 2004). In contrast, cyclosporintreatment after transplantation of NSCs to the brain lacked anassociation with improved function or increased graft survival(Modo et al., 2002; Schwarz et al., 2006).
Despite the expression of MHC on human NSCs, they are notrecognized by alloreactive lymphocytes in vitro (Odeberg etal., 2005). However, the in vivo situation offersthe transplanted human NSCs a more complex milieu, such asexposure to a variety of pro- and anti-inflammatory cytokines(Ishii et al., 2006; Nakamura et al., 2003) following aneurotrauma that may change their immunocompetence.Another issue to consider is the maturation and differentiationthat occur within the host that may also affect the immuno-genicity and possible immunomodulatory potential.
In this study, the expression of immunological proteins,the immunogenicity, and the possible immunomodulatorypotential of human NSCs versus more differentiated neuralcell populations were investigated to gain further insight ofrelevance for the development of neural cell therapies.
Results
Expression of MHC molecules on NSCs, astrocytes,and neurons
Here, we further investigated the expression of MHC class Iand II molecules on NSCs and more differentiated neuronaland astrocyte cultures. The NSCs in the neurosphere cultures

58 E. Åkesson et al.
expressed CD133 (N70%) (data not shown) and nestin (N90%,Fig. 1A) to a high degree. After 7 days of culture underdifferentiation conditions, the majority of the cells in theneuronal cultures were immunoreactive (IR) for both NeuNand β-tubulin III (78±12 and 59±25%, respectively; Fig. 1B).Fewer than 10% of the cells in the neuronal cultures were IRfor glial fibrillary acidic protein (GFAP) or CD133, and 20–30% of the cells were IR for nestin (Fig. 1B, inset). Themajority of the cells in the astrocyte cultures were IR for GFAP(70±14%; Fig. 1C), and only a minority expressed β-tubulin III(13±55%). In addition, amajority of theGFAP-expressing cellswas also positive for nestin at this time point (Fig. 1C, inset).
Seven days after the onset of the differentiation, weanalyzed the cells for the expression of MHC class I and classII molecules on the cell surface. While differentiation intoastrocytic phenotypes did not alter the number of cellsexpressing these molecules, a significant reduction wasdemonstrated on cells in the neuronal cultures. Both thepercentage of cells IR for MHC class I and class II and the level
Figure 2 The expression of MHC class I and class II molecules. NSCsurface expression of MHC by flow-cytometric analysis. The percentaas the mean fluorescence intensity (MFI; B and D, respectively), is shANOVA. (E) The histograms show one representative example of MHC cor neuronal cultures (isotype shown as black histograms).
of expression (mean fluorescence intensity; MFI) weresignificantly reduced upon differentiation into neurons(Fig. 2). There were no differences between cases in theexpression patterns of MHC molecules due to the firsttrimester gestational ages included in the study (data notshown), as also previously reported (Piao et al., 2006).
Phenotypic characterization of the MHC-expressingcells
To investigate further what type of cells in the human NSC,astrocyte, and neuron cultures expressed MHC molecules ontheir cell surface, we double stained the cells for MHC andthe cellular markers GFAP, β-tubulin III, and CD133 prior toanalysis by flow cytometry. The results from these experi-ments clearly show that NSCs (CD133 IR cells) and astrocytes(GFAP IR cells) do indeed express MHC class I and class IImolecules on the cell surface (Fig. 3). The majority of theneurons (β-tubulin III IR cells) were also expressing MHC class I
s, neuronal, and astrocyte cultures were examined for the cell-ge of cells expressing (A) MHC class I and (C) MHC class II, as wellown. Mean values±SEM (n=5 different donors/group). ⁎Pb0.05,lass I or class II molecule expression on cells from NSC, astrocyte,

Figure 3 Characterization of the MHC-expressing cells. Two-color flow-cytometric analysis of MHC expression on NSCs or cells fromneuronal or astrocyte cultures. Each dot plot represents 10,000 cells. The percentages of cells expressing MHC class I or II and one ofthe cell-specific markers CD133, GFAP, and β-tubulin III are presented.
59Human NSCs and astrocytes suppress lymphocyte response
molecules (Fig. 3), but lacking MHC class II molecules. A smallpopulation of the cells in the neuron cultures was β-tubulin IIInegative, butMHCclass Imolecule positive (Fig. 3). In addition,a small percentage of MHC class II IR cells was observed in theneuron cultures that were coexpressing CD133 (Fig. 3), mostlikely representing immature cells (NSCs) still present in theneuronal cultures. Thus, human in vitro expanded NSCs,astrocytes, and neurons do express MHC class I molecules,while MHC class II is present on NSCs and astrocytes.
The majority of NSCs and astrocytes, but notneurons, express the adhesion markers lymphocytefunction-associated antigen-3 (LFA-3) andintercellular adhesion molecule-1 (ICAM-1)
To analyze further the potential of NSCs, astrocytes, andneurons to elicit an immune response after transplantation, weexamined the expression of the adhesion markers LFA-3 andICAM-1 and the costimulatorymolecules CD80, CD86, and CD40using flow cytometry. Neither astrocytes nor neurons wereCD80 or CD40 positive, but sparse CD86 expression was foundon a small population of both neurons and astrocytes (11±8 and
4±2%, respectively). Neither of the costimulatory moleculeswas detected on NSCs (data not shown).
In contrast to the costimulatory molecules, the majority ofNSCs and astrocytes expressed the adhesion molecules LFA-3and ICAM-1 on their cell surfaces (Fig. 4). Upon differentiationinto neurons the percentage of cells IR for the adhesionmarkers was significantly reduced (Fig. 4A). Interestingly,astrocytes expressed enhanced levels (MFI) of ICAM-1 com-pared to the NSCs (Fig. 4B).
Neither NSCs, astrocytes, nor neurons trigger anallogeneic lymphocyte reaction
We have previously shown that despite high expression of bothMHC class I and class II molecules on NSCs, they were unable toelicit a proliferative immune response (Odeberg et al., 2005).Therefore, we were also interested in investigating theimmunogenicity of human NSC-derived astrocytes and neurons.A proliferative responsewas not demonstratedwhen allogeneiclymphocyteswere stimulatedwith NSCs, neurons, or astrocytesin mixed lymphocyte culture (MLC) assays (Fig. 5A). Theimmunogenicity of the cells was not affected by treatmentwith

Figure 4 The presence of the adhesion molecules ICAM-1 andLFA-3 on NSCs, astrocytes, and neurons. Cells were examined forthe expression of ICAM-1 and LFA-3 by flow cytometry, and (A) thepercentage of cells and (B) the MFI are shown. The valuesrepresent means±SEM (n≥4 different donors/group). §Pb0.05,compared to both NSC and astrocytes (§1, ICAM-1, and §2, LFA-3).⁎Pb0.05.
Figure 5 Neither NSCs, neurons, nor astrocytes trigger alymphocyte proliferative response. (A) Cells from neuronal andastrocyte cultures as well as NSCs were cultured together withallogeneic PBMCs. Differentiated cells were also treated withIFN-γ. The proliferative lymphocyte response was measured ascounts per minute (cpm). As positive control PBMCs werestimulated with noncompatible allogeneic PBMCs (allo) and asnegative control, PBMCswere stimulatedwith themselves (auto).The values are means±SEM (PBMCs and neural cells have nN3different donors/group). (B) The surface expression of MHC class Iand class II molecules before (filled curve) and after IFN-γ (blackline) treatment is shown (one representative example). §Pb0.05(ANOVA) compared to all other groups presented in graph.
60 E. Åkesson et al.
the inflammatory cytokine IFN-γ (Fig. 5A), despite increasedlevels of MHC class I and class II molecule expression induced bythe IFN-γ treatment (Fig. 5B). Thus, neither NSCs, neurons, norastrocytes triggered a proliferative Tcell response.
NSCs and astrocytes suppress lymphocyteproliferation induced by alloantigen
To characterize whether our NSCs and differentiated cells hadthe ability to modulate a human allogeneic immune response,we stimulated peripheral blood mononuclear cells (PBMCs)with alloantigen in the presence or absence of NSCs ordifferentiated cells (neurons or astrocytes). NSCs clearlyinhibited lymphocyte proliferation induced by alloantigen andat a NSC:PBMC ratio of 1:1 a complete suppression was seen(Fig. 6A). Astrocytes, but not neurons, did also interfere withalloantigen stimulation of lymphocytes (Fig. 6B) at equivalentcell ratios. The capacity of NSCs to suppress a lymphocyte (Tcell) response was further investigated by stimulation oflymphocytes with the T cell mitogens phytohemagglutinin(PHA) and concanavalin A (Con A) in the presence or absence ofNSCs. Interestingly, NSCs did not interfere with either PHA orConA stimulation at the ratios 1:10 and 1:1. However, when thecell concentration of NSCs was increased even further (5:1),PHA but not Con A activation was suppressed (Fig. 6C).
NSCs can mediate their inhibitory effect via severaldifferent mechanisms, such as release of soluble factor orcell–cell interactions. Soluble factors can be either releasedcontinuously from NSCs or induced upon the cell–cell interac-tion and be secreted by either NSCs or lymphocytes. To analyzewhether NSCs release soluble factors that suppress analloantigen stimulation of lymphocytes, we stimulated PBMCswith alloantigens in the presence or absence of supernatantfrom NSC cultures, which did not affect lymphocyte prolifera-tion (Fig. 6D). In addition, we analyzed the supernatant from

Figure 6 Lymphocyte stimulation by alloantigens is suppressed by human NSCs and astrocytes. PBMCs were stimulated withalloantigens (allo) in the presence or absence of (A) NSCs or (B) cells from neuronal or astrocyte cultures. The cell concentration ratios(neural cells:PBMCs) testedwere 1:10 and 1:1. As a negative control PBMCswere also stimulated with themselves (auto). The values aremeans±SEM (n=6/group). In (C), human PBMCswere stimulatedwith themitogens PHA and Con A, in the presence or absence of NSCs atdifferent cell concentrations (NSCs:PBMCs ratio 1:10, 1:1, and 5:1) (mean values±SEM (n=6/group)). (D) PBMCs were also stimulatedwith alloantigen in the presence or absence of supernatant from NSC cultures (means±SEM (n=6/group)). In all individual experimentsrun, PBMCs and neural cells were derived from at least three different donors/cases. ⁎Difference between groups, pb0.05 (ANOVA).
61Human NSCs and astrocytes suppress lymphocyte response
alloantigen stimulation in the presence or absence of NSCs,after 72 h incubation, for the presence of IL-2, IFN-γ, IL-10, andTGF-β, of which the latter two are known to suppress T cellproliferation. The supernatants from lymphocytes stimulatedwith alloantigen in the presence of NSCs did not displayelevated levels of either IL-10 or TGF-β (data not shown).Furthermore, they contained levels of IL-2 and IFN-γ similarto those of lymphocytes stimulated with alloantigens inthe absence of NSCs (data not shown). Furthermore, humanPBMCs were stimulated with incompatible PBMC populationswith or without conditioned medium from NSC cultures toevaluate whether the suppressive factor(s) were soluble.No suppression was observed of an allogeneic responseunder these conditions (Fig 6 D).Thus, these results in-dicate that the suppressive effect mediated by NSCs onthe lymphocyte proliferative response is dependent on cell–cell contact rather than on the investigated soluble factorsalone.
Human NSCs trigger a rejection response in axenograft model
With low and even suppressive immune actions by the humanNSCs in the in vitro allo-experiments, we investigated theimmunogenicity of these cells further in vivo, in a xeno-transplantation model in the SCI rodent. Six differentexperimental groups were included, with five animals/group. One animal presented weight loss early after surgeryand was thereby excluded from the study. The survival ofhuman neural cells deriving from the transplanted neuro-spheres was analyzed by immunohistochemistry usinghuman-specific HSP27 antibodies (Fig. 7). All the immuno-suppressed cyclosporin A (CSA)-treated rodents includednumerous HSP27-IR cells in their spinal cord parenchyma at 6weeks posttransplantation. However, in the saline-treatedgroups only three rodents presented human cells at this timepoint and they were all grafted with the P0 human NSCs,
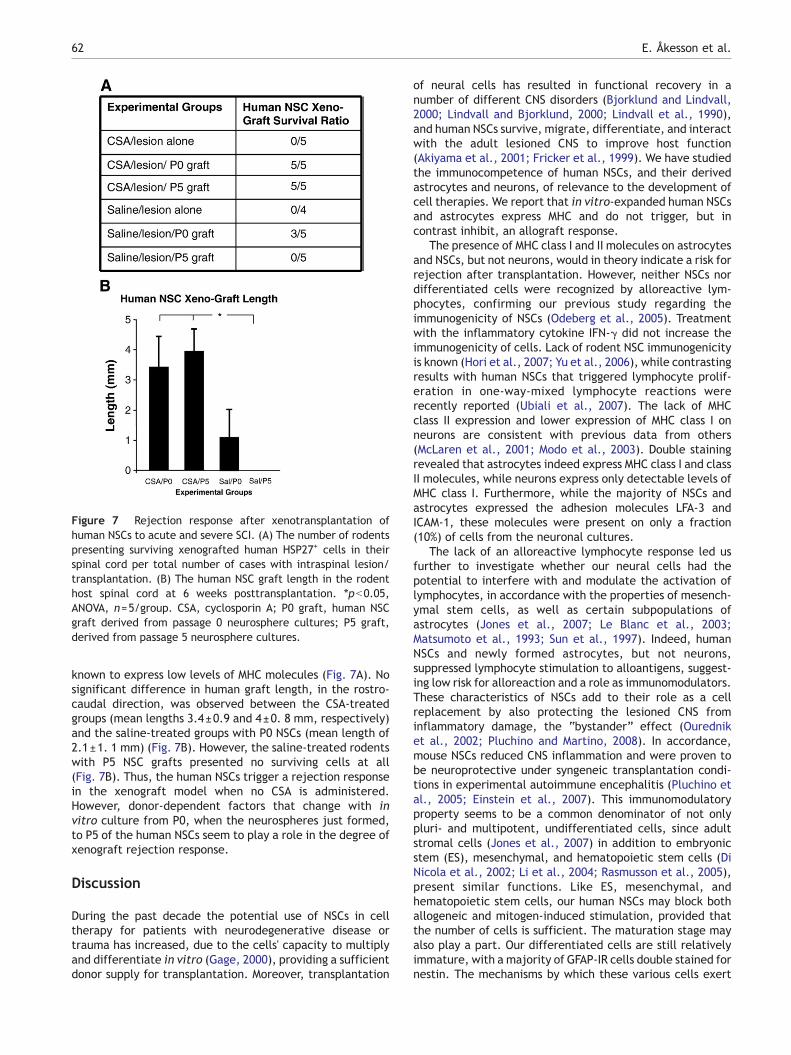
Figure 7 Rejection response after xenotransplantation ofhuman NSCs to acute and severe SCI. (A) The number of rodentspresenting surviving xenografted human HSP27+ cells in theirspinal cord per total number of cases with intraspinal lesion/transplantation. (B) The human NSC graft length in the rodenthost spinal cord at 6 weeks posttransplantation. ⁎pb0.05,ANOVA, n=5/group. CSA, cyclosporin A; P0 graft, human NSCgraft derived from passage 0 neurosphere cultures; P5 graft,derived from passage 5 neurosphere cultures.
62 E. Åkesson et al.
known to express low levels of MHC molecules (Fig. 7A). Nosignificant difference in human graft length, in the rostro-caudal direction, was observed between the CSA-treatedgroups (mean lengths 3.4±0.9 and 4±0. 8 mm, respectively)and the saline-treated groups with P0 NSCs (mean length of2.1±1. 1 mm) (Fig. 7B). However, the saline-treated rodentswith P5 NSC grafts presented no surviving cells at all(Fig. 7B). Thus, the human NSCs trigger a rejection responsein the xenograft model when no CSA is administered.However, donor-dependent factors that change with invitro culture from P0, when the neurospheres just formed,to P5 of the human NSCs seem to play a role in the degree ofxenograft rejection response.
Discussion
During the past decade the potential use of NSCs in celltherapy for patients with neurodegenerative disease ortrauma has increased, due to the cells' capacity to multiplyand differentiate in vitro (Gage, 2000), providing a sufficientdonor supply for transplantation. Moreover, transplantation
of neural cells has resulted in functional recovery in anumber of different CNS disorders (Bjorklund and Lindvall,2000; Lindvall and Bjorklund, 2000; Lindvall et al., 1990),and human NSCs survive, migrate, differentiate, and interactwith the adult lesioned CNS to improve host function(Akiyama et al., 2001; Fricker et al., 1999). We have studiedthe immunocompetence of human NSCs, and their derivedastrocytes and neurons, of relevance to the development ofcell therapies. We report that in vitro-expanded human NSCsand astrocytes express MHC and do not trigger, but incontrast inhibit, an allograft response.
The presence of MHC class I and II molecules on astrocytesand NSCs, but not neurons, would in theory indicate a risk forrejection after transplantation. However, neither NSCs nordifferentiated cells were recognized by alloreactive lym-phocytes, confirming our previous study regarding theimmunogenicity of NSCs (Odeberg et al., 2005). Treatmentwith the inflammatory cytokine IFN-γ did not increase theimmunogenicity of cells. Lack of rodent NSC immunogenicityis known (Hori et al., 2007; Yu et al., 2006), while contrastingresults with human NSCs that triggered lymphocyte prolif-eration in one-way-mixed lymphocyte reactions wererecently reported (Ubiali et al., 2007). The lack of MHCclass II expression and lower expression of MHC class I onneurons are consistent with previous data from others(McLaren et al., 2001; Modo et al., 2003). Double stainingrevealed that astrocytes indeed express MHC class I and classII molecules, while neurons express only detectable levels ofMHC class I. Furthermore, while the majority of NSCs andastrocytes expressed the adhesion molecules LFA-3 andICAM-1, these molecules were present on only a fraction(10%) of cells from the neuronal cultures.
The lack of an alloreactive lymphocyte response led usfurther to investigate whether our neural cells had thepotential to interfere with and modulate the activation oflymphocytes, in accordance with the properties of mesench-ymal stem cells, as well as certain subpopulations ofastrocytes (Jones et al., 2007; Le Blanc et al., 2003;Matsumoto et al., 1993; Sun et al., 1997). Indeed, humanNSCs and newly formed astrocytes, but not neurons,suppressed lymphocyte stimulation to alloantigens, suggest-ing low risk for alloreaction and a role as immunomodulators.These characteristics of NSCs add to their role as a cellreplacement by also protecting the lesioned CNS frominflammatory damage, the “bystander” effect (Ouredniket al., 2002; Pluchino and Martino, 2008). In accordance,mouse NSCs reduced CNS inflammation and were proven tobe neuroprotective under syngeneic transplantation condi-tions in experimental autoimmune encephalitis (Pluchino etal., 2005; Einstein et al., 2007). This immunomodulatoryproperty seems to be a common denominator of not onlypluri- and multipotent, undifferentiated cells, since adultstromal cells (Jones et al., 2007) in addition to embryonicstem (ES), mesenchymal, and hematopoietic stem cells (DiNicola et al., 2002; Li et al., 2004; Rasmusson et al., 2005),present similar functions. Like ES, mesenchymal, andhematopoietic stem cells, our human NSCs may block bothallogeneic and mitogen-induced stimulation, provided thatthe number of cells is sufficient. The maturation stage mayalso play a part. Our differentiated cells are still relativelyimmature, with a majority of GFAP-IR cells double stained fornestin. The mechanisms by which these various cells exert

63Human NSCs and astrocytes suppress lymphocyte response
their immunoregulatory functions must be further evaluatedand may be diverse.
Thus, our study indicates that human NSCs possessfeatures that counteract an immune response, providingprotection in the allogeneic situation, by inhibition of alymphocyte response. This suppressive effect may depend ona cell–cell interaction between lymphocytes and humanNSCs since supernatant from NSC cultures did not inhibitlymphocyte proliferation. Furthermore, the inhibition wasnot due to increased production of IL-10 and TGF-β, furthersupporting that the effect mediated by NSCs on thelymphocyte proliferative response is dependent on cell–cell contact rather than on these soluble factors.
The inhibition of an inflammatory response by neuralprecursor cells in vivo may in fact act both in the CNS (Yu etal., 2006) and as a peripheral immunosuppression elicitedfrom grafted neural precursor cells in the lymph nodes andspleen, counteracting experimental autoimmune encephali-tis (Ben-Hur, 2007; Einstein et al., 2006). A challenge is nowto understand and regulate this immunomodulation by thedonor cells and to investigate during what time window theseproperties are most effective in the injured nervous systemto reduce the need for immunosuppressive drugs and toachieve neuroprotection.
In vitro studies allow human allograft models to bescrutinized, while investigating a complex and full hostresponse in vivo may only be performed in the xenograftsituation. The two are significantly diverse host–graftmodels. A majority of in vivo xenograft models, includinghuman fetal neural donor cells (non-in vitro-expanded),demonstrate rejection of the human donor cells (Barker etal., 2000; Finsen et al., 1991; Mason et al., 1986) andsimilarly human NSCs grafted to a traumatic brain injuryrequired immunosuppressive treatment (Al Nimer et al.,2004; Ortega et al., 1992; Wennersten et al., 2006). We herereport that our human NSCs, unable to trigger an alloreac-tion, induce a xenoreaction resulting in graft rejection.However, donor-dependent factors that change with in vitroculture from P0 to P5 of the human NSCs seem to play a rolein the strength of the xenograft response. We know that MHCclass I and II molecules substantially increase between thosein vitro passages and may explain the lack of surviving humancells in the case of P5 NSC xenografts treated with salinealone. This implies that in vitro-expanded long-termcultured human NSCs may exhibit an increased risk forrejection in an allotransplantation. Therefore, the knowl-edge of how to reduce/regulate these donor-dependentfactors after long-term in vitro expansion is required, to gainsufficient cell numbers for cryostorage, safety tests, andclinical application.
In the few studies investigating the immunologicalresponse to transplanted NSCs in an allogeneic situation inanimal species divergent results have been achieved andtherefore are difficult to interpret for the human clinicalsetting. Hori et al. (2007) demonstrated that allogeneicmouse CNS progenitor cells transplanted beneath the kidneycapsule were not rejected, suggesting that the cells lackimmunogenicity, supporting our in vitro data. In contrast,transplantation of allogeneic rat NSCs into traumatized brainwas associated with increased infiltration of T cells to thesite of transplantation compared to lesion alone (Zheng etal., 2007). The different results may reflect NSC species
differences, as well as a difference in MHC expression, and/or that the immune response is more alert/prone to becomeactivated in traumatized compared to normal animals. If NSCexpression of MHC is the major factor for an acceptance orrejection response, human NSCs should be more prone totriggering a strong immune response and become rejected,since they are reported to have both MHC class I and class II(Åkesson et al., 2000; Al Nimer et al., 2004; Odeberg et al.,2005; Poltavtseva et al., 2002; Wright et al., 2003), whereasmouse and rat NSCs lack or express only low levels of MHCclass I molecules (Hori et al., 2007; Mammolenti et al., 2004;McLaren et al., 2001).
In summary, human NSCs, neurons, and astrocytes allexhibit relatively low immunogenicity. Furthermore, NSCsand astrocytes have the potential to modulate an immuneresponse, which may favor successful cell therapy forpatients with neurodegenerative diseases, traumatic inju-ries, or stroke in the future. However, further research isrequired to fully understand and regulate the interactionbetween transplanted NSCs and the host immune system.
Materials and methods
Human forebrain tissue and peripheral bloodmononuclear cells
After obtaining informed consent from the pregnant womenand approval by the Regional Human Ethics Committee,Stockholm, we collected subcortical forebrain tissue fromhuman abortion material of 5–12 weeks of gestation, aspreviously described (Odeberg et al., 2005; Piao et al.,2006).
After approval by the Regional Human Ethics Committee,Stockholm, human PBMCs were purified from buffy coatsfrom healthy blood donors (provided by the KarolinskaUniversity Hospital blood center), using Lymphoprep (Axis-Shield PoC AS) according to the manufacturer's instructions.
Establishment of human neurosphere cultures
The samples of human tissues were dissociated mechanicallyinto single-cell suspensions employing a glass–Teflon homo-genizer, following which neurospheres were established andpropagated as previously described (Åkesson et al., 2007;Odeberg et al., 2005, 2006, 2007; Piao et al., 2006). Theneurospheres were passaged every 7–10 days, using TrypLe(Invitrogen, Life Technologies). Herein, we refer to culturedhuman neural cells expanded as free-floating neurospheresas NSCs.
Differentiation of NSCs
The neurospheres were dissociated into a single-cell suspen-sion and suspended either in Neurobasal medium supplemen-ted with B27 (Invitrogen Life Technologies) or in DMEM:F12supplemented with 0.6% glucose, 5 mM Hepes, 1% N2supplement, and 10% fetal calf serum (FCS). Neurobasalmedium promotes NSC differentiation primarily to neurons,whereas the medium containing 10% FCS promotes thedifferentiation primarily to astrocytes. Cells were seededonto poly-d-lysine (Sigma, 0. 1 mg /ml)-coated wells (6-

64 E. Åkesson et al.
wells/plate; Nunc) at a cell concentration of 240×103 cells/well. Cells were also seeded onto poly-d-lysine-coatedcircular glass coverslips in 24-well plates (Nunc) at a cellconcentration of 60,000 cells/ml (neuronal cultures) or30,000 cells/ml (astrocyte cultures). After 7 days ofdifferentiation, immunocytochemical, flow-cytometric, orimmunological assays were performed as described below.
Flow-cytometric analysis
A fluorescence-activated cell sorter (FACSort; Becton–Dick-inson) was used to analyze the NSCs, neurons, or astrocytesfor the expression of human leukocyte antigen (HLA) class I(clone W6/32, Dakopatts), HLA-DR (clone L243), CD80, CD86,LFA-3, and ICAM-1 PE (all purchased from BD Bioscience) andCD133 (Miltenyi Biotec). The isotype controls used were asfollows: IgG2a PE or FITC, IgG1 FITC, IgG1, and IgG2b(Dakopatts). The cells were stained with the respectiveantibodies at 4 °C for 30 min, washed, and resuspended inphosphate-buffered saline (PBS). Only the viable cells,propidium iodide negative, were analyzed using CELLQuestsoftware (Becton–Dickinson Immunocytometry Systems).
For intracellular staining, cellswere fixedandpermeabilizedwithCytofix/Cytoperm (BDBiosciences) and stainedwith one ofthe polyclonal antibodies GFAP (rabbit; Dakopatts), nestin(Chemicon), and β-tubulin III (Biosite) or with the respectiveisotype control. Following rinsing, the cellswere incubatedwiththe appropriate secondary antibody (either Alexa Fluor 488(1:600; Molecular Probes) or Cy3 (1:1200; Jackson ImmunoR-esearch Laboratories)) and analyzed as above.
For cell samples double stained for cell-surfacemarkers andintracellular markers, cells were first stained for the cell-surface molecule, thereafter fixed/permeabilized, and thenstained for the intracellular molecule. In these experimentsboth single anddouble stainswereperformed to correct for therisk of false double labeling.
Mixed lymphocyte cultures
NSC, astrocyte, or neuronal cultures were harvested (N90% ofthe cells were viable as demonstrated with trypan blue),followed by dilution in RPMI medium with 10% FCS. The cellswere irradiated (20 Gy; Gammacell 2000, Molsgard, Denmark),and 5×104 cells were cocultured with PBMCs (105 cells), in200 μl medium in 96-well plates, and incubated at 37 °C and 5%CO2. As a positive control, the PBMCswere also stimulatedwithalloantigen, i.e., irradiatedPBMCs fromanother donor, amixedlymphocyte reaction culture. At 5 days poststimulation, thecells were pulsed with 1 μCi [methyl-3H]thymidine (AmershamLife Science). The following day, the cells were harvested ontofilters, using a plate harvester (Harvester 996; Tomtec,Hamden, CT, USA) according to the manufacturer's instruc-tions, and counted in an automated radioactivity counter (1450MicroBeta Trilux, Wallac, Sweden AB). The stimulated prolif-eration was calculated as the mean counts per minute oftriplicate samples.
To study the effects of NSCs, neurons, and astrocytes onlymphocyte reactivity in MLCs, PBMCs (105 cells) werestimulated with an equal number of irradiated PBMCs fromanother donor. A third part of irradiated NSCs or differentiatedcells was added at the beginning of the experiment, at a cellconcentration of 1×104 cells/well (1:10 dilution). In the dose–
response experiments, neural cells (NSCs or differentiatedcells) were added at a neural cell:PBMC ratio of 1:10 and 1:1.These experiments were performed with human NSCs andPBMCs from at least three different donors.
MLC in the presence of supernatant from irradiated NSCsPBMCs were stimulated with allogeneic PBMCs (as describedabove) in the presence or absence of supernatant fromirradiated NSCs. The supernatant was prepared as follows:NSCs were irradiated and 0.5×106 cells/ml were incubatedfor 6 days. Thereafter the supernatant was harvested,cleared by centrifugation, and then added to the MLCassay. The plates were harvested as described above.
IFN-γ treatmentNSC, neuron, or astrocyte cultures were treated with eitherIFN-γ (R&D Systems; 200 U/ml) or vehicle (RPMI with 10%FCS), as a control, for 3 days prior to further experiments.
Mitogen stimulation
PBMCs (5×104 cells/well) were stimulated with PHA (5 μg/ml)or Con A (20 μg/ml) (both from Sigma) in triplicates, in thepresence or absence of irradiated NSCs at the following NSC:PBMC ratios: 1:10, 1:1, and 5:1. The cultures were pulsed with1 μCi [methyl-3H]thymidine (Amersham Life Science) at 2 dayspoststimulation. The day after, cells were harvested ontofilters as described above.
Experimental animals and surgical procedure
Thirty female Harlan Sprague–Dawley rats, 170–190 g (B-land,Denmark) were included in the study. The experimentalprocedure was approved by The Experimental Animal EthicsCommittee of Southern Stockholm, Huddinge Tingsrätt. Thefollowing experimental groups were included in the study: (1)CSA-treated rodents with a spinal cord lesion alone, (2) CSA-treated rodentswith a spinal cord lesion and passage (P) 0, NSCgraft, (3) CSA-treated rodents with a spinal cord lesion and P5NSC graft, (4) saline-treated rodents with a spinal cord lesionalone, (5) saline-treated rodents with a spinal cord lesion andP0 NSC graft, and (6) saline-treated rodents with a spinal cordlesion and P5 NSC graft. For a detailed description of thesurgical procedure, see the supplementary materials andmethods. At 42 days postsurgery the animals were reanesthe-tized and perfusion fixed through the ascending aorta with100 ml Ca2+-free Tyrode's solution, followed by 400 mlphosphate-buffered 4% paraformaldehyde (Merck, Darmstadt,Germany). The tissue was postfixed for 90 min and cryopre-served in 20% sucrose in 0. 1 M PBS at 4°C for at least 48 h.
Immunocytochemistry
Human neurospheres were fixed and like the rodent spinalcords cryosectioned at 5 or 10 μm thickness, respectively, aspreviously described (Piao et al., 2006; Åkesson et al., 2007).The differentiated cells present on the glass coverslips werefixed with buffered 4% paraformaldehyde 4, for 15 min atroom temperature, and thereafter both sections and cellswere preincubated with 1.5% normal goat serum for 30 minprior to incubation with the primary antibodies. Thefollowing antibodies were used: nestin (1:200, mouse, or

65Human NSCs and astrocytes suppress lymphocyte response
1:400, rabbit; both from Chemicon), GFAP (1:500; Dako-patts), β-tubulin III (1:800, mouse; Chemicon), NeuN (1:100,mouse; Chemicon), and human specific heat shock protein 27(HSP27, 1:1000; Nordic Biosite).
The staining procedure was performed as previouslydescribed (Odeberg et al., 2005; Piao et al., 2006).Visualization of the primary antibodies was achieved byincubation with secondary antibodies conjugated to AlexaFluor 488 (Molecular Probes; diluted 1:400) or Cy3(Jackson ImmunoResearch Laboratories; diluted 1:1800).The cell nuclei were stained with Hoechst (1 μg/ml;Molecular Probes) prior to the coverslips being mountedwith PVA-DABCO (9.6% polyvinyl alcohol, 24% glycerol, and2.5% 1,4-diazabicyclo (2.2.2) octane in 67 mM Tris–Cl,pH 8.0).
The images were examined in a fluorescence micro-scope (Zeiss; Axiophot) and documented using a CCDcamera (Hamamatsu; ORCA-ER, C4742-95) together withthe Openlab software for Macintosh (Improvision). Thenumber and/or percentage of cells expressing the differ-ent antigens analyzed was calculated by counting threefields per glass (objective 20×). The microscopy of theintraspinal transplantation was performed blindly withcoded slides. The human graft length was estimated bymultiplying the number of host spinal cord cross sections(with human HSP27+ cells present) with the sectioninterval of 0. 4 mm.
Quantification of cytokines
Supernatants from PBMCs stimulated with alloantigens for 72 hin the presence of NSCs (1:10 NSC:PBMC ratio) were collected,cleared by centrifugation, and frozen (−70 °C) for cytokineanalysis. Flow Cytomix (Bender MedSystems GmbH) was usedto measure the levels of IFN-γ, TGF-β, IL-10, and IL-2, and thedata were analyzed with FlowCytomixPro 2.2 software.
Statistical analyses
Analysis of possible differences in the size of the cellpopulations, degree of fluorescence intensity, proliferationresponse, and human NSC graft length between various groupswas performed by multifactorial analysis of variance (ANOVA).Sheffe's test was employed as post hoc test. p valuesb0.05were considered significant. The mean values±SEM arepresented in the figures and text. The statistical programStatview 4.12, for Macintosh computers, was used forstatistical analysis (Abacus Concepts, Berkeley, CA, USA).
Acknowledgments
We thank Eva-Britt Samuelsson and Lena Holmberg forexcellent technical support. We express our sincereappreciation to Cecilia Götherström and Berit Sundbergfor their helpful assistance in the MLC plate-readingprocess. This study was supported financially by grantsfrom the Craig Hospital Foundation, Denver, Colorado,USA; the Stockholms Sjukhem Foundation; The ResearchFunds of Karolinska Institutet; The Swedish Brain Founda-tion; the M. Bergvalls Foundation; and the Swedish Societyof Medicine.
Appendix A. Supplementary data
Supplementary data associatedwith this article can be found,in the online version, at doi:10.1016/j.scr.2008.06.002.
References
Åkesson, E., Markling, L., Kjaeldgaard, A., Falci, S., Ringden, O.,2000. MHC antigen expression in human first trimester spinalcord with implications for clinical transplantation procedures.J. Neuroimmunol. 111, 210–214.
Åkesson, E., Piao, J.H., Samuelsson, E.B.,Holmberg, L., Kjaeldgaard,A.,Falci, S., Sundstrom, E., Seiger, A., 2007. Long-term culture andneuronal survival after intraspinal transplantation of human spinalcord-derived neurospheres. Physiol. Behav 92, 60–66.
Akiyama, Y., Honmou, O., Kato, T., Uede, T., Hashi, K., Kocsis, J.D.,2001. Transplantation of clonal neural precursor cells derivedfrom adult human brain establishes functional peripheral myelinin the rat spinal cord. Exp. Neurol. 167, 27–39.
Al Nimer, F., Wennersten, A., Holmin, S., Meijer, X., Wahlberg, L.,Mathiesen, T., 2004. MHC expression after human neural stemcell transplantation to brain contused rats. Neuroreport 15,1871–1875.
Alonso, G., Ridet, J.L., Oestreicher, A.B., Gispen, W.H., Privat, A.,1995. B-50 (GAP-43) immunoreactivity is rarely detected withinintact catecholaminergic and serotonergic axons innervating thebrain and spinal cord of the adult rat, but is associated with theseaxons following lesion. Exp. Neurol. 134, 35–48.
Barker, R.A., Ratcliffe, E., McLaughlin, M., Richards, A., Dunnett,S.B., 2000. A role for complement in the rejection of porcineventral mesencephalic xenografts in a rat model of Parkinson'sdisease. J. Neurosci. 20, 3415–3424.
Ben-Hur, T., 2007. Immunomodulation by neural stem cells. J. Neurol.Sci. 265, 102–104.
Bjorklund, A., Lindvall, O., 2000. Cell replacement therapies forcentral nervous system disorders. Nat. Neurosci. 3, 537–544.
Curtis, R., Green, D., Lindsay, R.M., Wilkin, G.P., 1993. Up-regulationof GAP-43 and growth of axons in rat spinal cord after compressioninjury. J. Neurocytol. 22, 51–64.
Di Nicola, M., Carlo-Stella, C., Magni, M., Milanesi, M., Longoni, P.D.,Matteucci, P., Grisanti, S., Gianni, A.M., 2002. Human bonemarrow stromal cells suppress T-lymphocyte proliferationinduced by cellular or nonspecific mitogenic stimuli. Blood 99,3838–3843.
Diaz-Ruiz, A., Vergara, P., Perez-Severiano, F., Segovia, J., Guizar-Sahagun, G., Ibarra, A., Rios, C., 2005. Cyclosporin-A inhibitsconstitutive nitric oxide synthase activity and neuronal andendothelial nitric oxide synthase expressions after spinal cordinjury in rats. Neurochem. Res. 30, 245–251.
Einstein, O., Grigoriadis, N., Mizrachi-Kol, R., Reinhartz, E.,Polyzoidou, E., Lavon, I., Milonas, I., Karussis, D., Abramsky,O., Ben-Hur, T., 2006. Transplanted neural precursor cells reducebrain inflammation to attenuate chronic experimental autoim-mune encephalomyelitis. Exp. Neurol. 198, 275–284.
Einstein, O., Fainstein, N., Vaknin, I., Mizrachi-Kol, R., Reihartz, E.,Grigoriadis, N., Lavon, I., Baniyash, M., Lassmann, H., Ben-Hur, T.,2007. Neural precursors attenuate autoimmune encephalomyelitisby peripheral immunosuppression. Ann. Neurol. 61, 209–218.
Finsen, B.R., Sorensen, T., Castellano, B., Pedersen, E.B., Zimmer,J., 1991. Leukocyte infiltration and glial reactions in xenograftsof mouse brain tissue undergoing rejection in the adult rat brain:a light and electron microscopical immunocytochemical study.J. Neuroimmunol. 32, 159–183.
Fricker, R.A., Carpenter, M.K., Winkler, C., Greco, C., Gates, M.A.,Bjorklund, A., 1999. Site-specific migration and neuronaldifferentiation of human neural progenitor cells after transplan-tation in the adult rat brain. J. Neurosci. 19, 5990–6005.

66 E. Åkesson et al.
Gage, F.H., 2000. Mammalian neural stem cells. Science 287,1433–1438.
Garrido, R.S., Aguado, J.M., Diaz-Pedroche, C., Len, O., Montejo,M., Moreno, A., Gurgui, M., Torre-Cisneros, J., Pareja, F.,Segovia, J., Garcia, M., Lumbreras, C., 2006. A review of criticalperiods for opportunistic infection in the new transplantationera. Transplantation 82, 1457–1462.
Guo, J., Zeng, Y., Liang, Y., Wang, L., Su, H., Wu, W., 2007.Cyclosporine affects the proliferation and differentiation ofneural stem cells in culture. Neuroreport 18, 863–868.
Hori, J., Ng, T.F., Shatos, M., Klassen, H., Streilein, J.W., Young, M.J., 2007. Neural progenitor cells lack immunogenicity and resistdestruction as allografts. Ocul. Immunol. Inflammation 15,261–273.
Ibarra, A., Hernandez, E., Lomeli, J., Pineda, D., Buenrostro, M.,Martinon, S., Garcia, E., Flores, N., Guizar-Sahagun, G., Correa,D., Madrazo, I., 2007. Cyclosporin-A enhances non-functionalaxonal growing after complete spinal cord transection. Brain Res.1149, 200–209.
Ishii, K., Nakamura, M., Dai, H., Finn, T.P., Okano, H., Toyama, Y.,Bregman, B.S., 2006. Neutralization of ciliary neurotrophicfactor reduces astrocyte production from transplanted neuralstem cells and promotes regeneration of corticospinal tractfibers in spinal cord injury. J. Neurosci. Res. 84, 1669–1681.
Jones, S., Horwood, N., Cope, A., Dazzi, F., 2007. The antiproliferativeeffect of mesenchymal stem cells is a fundamental property sharedby all stromal cells. J. Immunol. 179, 2824–2831.
Le Blanc, K., Tammik, C., Rosendahl, K., Zetterberg, E., Ringden,O., 2003. HLA expression and immunologic properties ofdifferentiated and undifferentiated mesenchymal stem cells.Exp. Hematol. 31, 890–896.
Li, L., Baroja, M.L., Majumdar, A., Chadwick, K., Rouleau, A.,Gallacher, L., Ferber, I., Lebkowski, J., Martin, T., Madrenas, J.,Bhatia, M., 2004. Human embryonic stem cells possess immune-privileged properties. Stem Cells 22, 448–456.
Lindvall, O., Rehncrona, S., Brundin, P., Gustavii, B., Astedt, B.,Widner, H., Lindholm, T., Bjorklund, A., Leenders, K.L.,Rothwell, J.C., et al., 1990. Neural transplantation in Parkinson'sdisease: the Swedish experience. Prog. Brain Res. 82, 729–734.
Lindvall, O., Bjorklund, A., 2000. First step towards cell therapy forHuntington's disease. Lancet 356, 1945–1946.
Mammolenti, M., Gajavelli, S., Tsoulfas, P., Levy, R., 2004. Absenceof major histocompatibility complex class I on neural stem cellsdoes not permit natural killer cell killing and prevents recogni-tion by alloreactive cytotoxic T lymphocytes in vitro. Stem Cells22, 1101–1110.
Mason, D.W., Charlton, H.M., Jones, A.J., Lavy, C.B., Puklavec, M.,Simmonds, S.J., 1986. The fate of allogeneic and xenogeneicneuronal tissue transplanted into the third ventricle of rodents.Neuroscience 19, 685–694.
Matsumoto, Y., Hanawa, H., Tsuchida, M., Abo, T., 1993. In situinactivation of infiltrating T cells in the central nervous systemwith autoimmune encephalomyelitis: the role of astrocytes.Immunology 79, 381–390.
McLaren, F.H., Svendsen, C.N., Van der Meide, P., Joly, E., 2001.Analysis of neural stem cells by flow cytometry: cellulardifferentiation modifies patterns of MHC expression. J. Neu-roimmunol. 112, 35–46.
Modo, M., Rezaie, P., Heuschling, P., Patel, S., Male, D.K., Hodges,H., 2002. Transplantation of neural stem cells in a rat model ofstroke: assessment of short-term graft survival and acute hostimmunological response. Brain Res. 958, 70–82.
Modo, M., Mellodew, K., Rezaie, P., 2003. In vitro expression of majorhistocompatibility class I and class II antigens by conditionallyimmortalized murine neural stem cells. Neurosci. Lett. 337,85–88.
Nakamura,M., Houghtling, R.A., MacArthur, L., Bayer, B.M., Bregman,B.S., 2003. Differences in cytokine gene expression profile
between acute and secondary injury in adult rat spinal cord.Exp. Neurol. 184, 313–325.
Odeberg, J., Piao, J.H., Samuelsson, E.B., Falci, S., Åkesson, E.,2005. Low immunogenicity of in vitro-expanded human neuralcells despite high MHC expression. J. Neuroimmunol. 161,1–11.
Odeberg, J., Wolmer, N., Falci, S., Westgren, M., Seiger, A.,Soderberg-Naucler, C., 2006. Human cytomegalovirus inhibitsneuronal differentiation and induces apoptosis in human neuralprecursor cells. J. Virol. 80, 8929–8939.
Odeberg, J., Wolmer, N., Falci, S., Westgren, M., Sundstrom, E.,Seiger, A., Soderberg-Naucler, C., 2007. Late human cytome-galovirus (HCMV) proteins inhibit differentiation of humanneural precursor cells into astrocytes. J. Neurosci. Res. 85,583–593.
Ortega, J.D., Sagen, J., Pappas, G.D., 1992. Short-term immuno-suppression enhances long-term survival of bovine chromaffincell xenografts in rat CNS. Cell Transplant. 1, 33–41.
Ourednik, J., Ourednik, V., Lynch, W.P., Schachner, M., Snyder, E.Y.,2002. Neural stem cells display an inherent mechanism forrescuing dysfunctional neurons. Nat. Biotechnol. 20, 1103–1110.
Piao, J.H., Odeberg, J., Samuelsson, E.B., Kjaeldgaard, A., Falci, S.,Seiger, A., Sundstrom, E., Åkesson, E., 2006. Cellular composi-tion of long-term human spinal cord- and forebrain-derivedneurosphere cultures. J. Neurosci. Res. 84, 471–482.
Pluchino, S., Martino, G., 2008. Neural stem cell-mediatedimmunomodulation: repairing the haemorrhagic brain. Brain131, 604–605.
Pluchino, S., Zanotti, L., Rossi, B., Brambilla, E., Ottoboni, L.,Salani, G., Martinello, M., Cattalini, A., Bergami, A., Furlan, R.,Comi, G., Constantin, G., Martino, G., 2005. Neurosphere-derived multipotent precursors promote neuroprotection by animmunomodulatory mechanism. Nature 436, 266–271.
Poltavtseva, R.A., Marey, M.V., Aleksandrova, M.A., Revishchin, A.V.,Korochkin, L.I., Sukhikh, G.T., 2002. Evaluation of progenitor cellcultures from human embryos for neurotransplantation. BrainRes. Dev. Brain Res. 134, 149–154.
Rasmusson, I., Ringden, O., Sundberg, B., Le Blanc, K., 2005.Mesenchymal stem cells inhibit lymphocyte proliferation bymitogens and alloantigens by different mechanisms. Exp. CellRes. 305, 33–41.
Rezzani, R., 2006. Exploring cyclosporine A side effects and theprotective role played by antioxidants: the morphological andimmunohistochemical studies. Histol. Histopathol. 21,301–316.
Schwarz, S.C., Wittlinger, J., Schober, R., Storch, A., Schwarz, J.,2006. Transplantation of human neural precursor cells in the 6-OHDA lesioned rats: effect of immunosuppression with cyclos-porine A. Parkinsonism Relat. Disord. 12, 302–308.
Sosa, I., Reyes, O., Kuffler, D.P., 2005. Immunosuppressants:neuroprotection and promoting neurological recovery followingperipheral nerve and spinal cord lesions. Exp. Neurol. 195,7–15.
Sun, D., Coleclough, C., Whitaker, J.N., 1997. Nonactivated astrocytesdownregulate T cell receptor expression and reduce antigen-specific proliferation and cytokine production of myelin basicprotein (MBP)-reactive Tcells. J. Neuroimmunol. 78, 69–78.
Ubiali, F., Nava, S., Nessi, V., Frigerio, S., Parati, E., Bernasconi, P.,Mantegazza, R., Baggi, F., 2007. Allorecognition of human neuralstem cells by peripheral blood lymphocytes despite low expres-sion of MHC molecules: role of TGF-{beta} in modulatingproliferation. Int. Immunol. 19, 1063–1074.
Wei, L., Steiner, J.P., Hamilton, G.S., Wu, Y.Q., 2004. Synthesisand neurotrophic activity of nonimmunosuppressant cyclos-porin A derivatives. Bioorg. Med. Chem. Lett. 14,4549–4551.
Wennersten, A., Holmin, S., Al Nimer, F., Meijer, X., Wahlberg, L.U.,Mathiesen, T., 2006. Sustained survival of xenografted human

67Human NSCs and astrocytes suppress lymphocyte response
neural stem/progenitor cells in experimental brain traumadespite discontinuation of immunosuppression. Exp. Neurol.199, 339–347.
Wright, L.S., Li, J., Caldwell, M.A., Wallace, K., Johnson, J.A.,Svendsen, C.N., 2003. Gene expression in human neural stem cells:effects of leukemia inhibitory factor. J. Neurochem. 86, 179–195.
Yu, J.J., Sun, X., Yuan, X., Lee, J.W., Snyder, E.Y., Yu, J.S., 2006.Immunomodulatory neural stem cells for brain tumour therapy.Expert Opin. Biol. Ther. 6, 1255–1262.
Zheng, X.S., Yang, X.F., Liu, W.G., Pan, D.S., Hu, W.W., Li, G., 2007.Transplantation of neural stem cells into the traumatized braininduces lymphocyte infiltration. Brain Inj. 21, 275–278.
Copyright © 2022 FDOKUMEN




















