
Expression of glutamate transporters in rat optic nerve oligodendrocytes
11
Expression of glutamate transporters in rat optic nerve oligodendrocytes Marı ´a Domercq, Marı ´a Victoria Sa ´ nchez-Go ´mez, Pilar Areso and Carlos Matute Departamento de Neurociencias, Facultad de Medicina y Odontologı ´a, Universidad del Paı ´s Vasco, 48940 Leioa, Vizcaya, Spain Keywords: axon–glia signalling, cell culture, glial transporters, glutamine synthetase Abstract To investigate the role of glutamate transport in non-synaptic glia, we characterized the expression of three major glutamate transporters (EAAC1, GLAST and GLT-1) in rat optic nerve in situ using reverse transcription-polymerase chain reaction in combination with Western blot and immunochemistry with specific antibodies. GLAST was localized to interfascicular oligodendrocytes, whereas a subpopulation of cells, probably immature oligodendrocyte cells, expressed EAAC1. In contrast, astrocytes, expressed only GLT-1, consistent with the idea that this is the major glutamate transporter in this cell type. In addition, we observed that glutamine synthetase, a key enzyme in glutamate metabolism, was localized in oligodendrocytes in situ. To examine the properties of these glutamate transporters, we conducted uptake experiments in glial cultures. The kinetics of sodium-dependent glutamate uptake in cultured oligodendrocytes from the perinatal rat optic nerve were markedly different from those observed in type- 1 astrocytes from the newborn rat cerebral cortex, with higher affinity and lower V max . In both cell types, glutamate transport was inhibited by L-trans-pyrrolidine-2,4-dicarboxylate (t-PDC). In contrast, dihydrokainate exhibited significantly more uptake inhibition in oligodendrocytes than in type-1 astrocytes. These results provide evidence for the expression of functional sodium-dependent glutamate transporters in optic nerve oligodendrocytes, and suggest that this cell type may play a role in the glutamate–glutamine cycle. Introduction Glutamate is the major excitatory neurotransmitter in the mammalian CNS, and its uptake is essential to maintain a low concentration of extracellular glutamate and to prevent excitotoxicity (Choi, 1992; Rothstein et al., 1996). Clearance of glutamate is carried out by high- affinity, sodium-dependent glutamate transporters localized in neuronal and glial cell membranes. The molecular properties and function of these transporters have recently been elucidated (for a review see Kanai et al., 1997). At least five glutamate transporters have been cloned: GLAST (Storck et al., 1992), GLT-1 (Pines et al., 1992), EAAC1 (Kanai & Hediger, 1992), EAAT4 (Fairman et al., 1995) and EAAT5 (Arriza et al., 1997). Localization studies indicate that EAAC1 is present in neurons throughout the brain (Kanai & Hediger, 1992). GLAST was found to be abundantly expressed in cerebellar Bergmann glia (Storck et al., 1992), while GLT-1 was observed in astroglia throughout the brain (Danbolt et al., 1992). Finally, EAAT4 is expressed in cerebellar Purkinje cells (Fairman et al., 1995) and EAAT5 is located in cells of the retina (Arriza et al., 1997). Based on these studies, EAAC1 is regarded as the major neuronal glutamate transporter, while GLAST and GLT-1 are viewed as the principal glial glutamate transporters (Rothstein et al., 1994). Glutamate transporters are also located in non-synaptic regions (Lehre et al., 1995; Choi & Chiu, 1997), but their properties as well as the processes which they subserve are presently unclear. In the optic nerve, it has been proposed that neurons may signal to glia along their axons via a calcium-independent mechanism, due to reversal of glutamate transporters present at nodes of Ranvier (Kriegler & Chiu, 1993). This feature, together with the fact that glutamate receptor activation can lead to oligodendroglial excitotoxicity (Yoshioka et al., 1996; Matute et al., 1997; Matute, 1998; McDonald et al., 1998), strongly suggests that the extracellular glutamate concentration is regulated by transporters in order to prevent oligodendrocyte cell death and demyelination. There is little information, however, about the type of transporters mediating glutamate uptake in axonal tracts of the CNS. In the present study, we analysed the expression of EAAC1, GLAST and GLT-1, the most widely distributed glutamate transporters, in the adult optic nerve using reverse transcription- polymerase chain reaction (RT-PCR), Western blot analysis and immunochemistry. The results of these experiments led us to analyse glutamate uptake in cultured oligodendroglial cells and to compare it with that in cultured astrocytes. Taken together, our results strongly suggest that oligodendrocytes contribute to the maintenance of low levels of extracellular glutamate. In addition, we found that glutamine synthetase, a key enzyme in glutamate metabolism, was expressed in oligodendrocytes from the optic nerve in situ. Materials and methods Materials Culture reagents were purchased from Sigma (St. Louis, MO, USA) unless otherwise stated. L-[ 3 H]-Glutamate (specific activity 46 Ci/ mmol) was from Amersham Life Sciences (Buckinghamshire, UK) and growth factors were from Alomone (Jerusalem, Israel). Drugs L Correspondence: C. Matute, as above. E-mail: [email protected] Received 12 August 1998, revised 28 January 1999, accepted 11 February 1999 European Journal of Neuroscience, Vol. 11, pp. 2226–2236, 1999 Ó European Neuroscience Association
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Expression of glutamate transporters in rat optic nerve oligodendrocytes

Expression of glutamate transporters in rat optic nerveoligodendrocytes
MarõÂa Domercq, MarõÂa Victoria SaÂnchez-GoÂmez, Pilar Areso and Carlos MatuteDepartamento de Neurociencias, Facultad de Medicina y OdontologõÂa, Universidad del PaõÂs Vasco, 48940 Leioa, Vizcaya,Spain
Keywords: axon±glia signalling, cell culture, glial transporters, glutamine synthetase
Abstract
To investigate the role of glutamate transport in non-synaptic glia, we characterized the expression of three major glutamatetransporters (EAAC1, GLAST and GLT-1) in rat optic nerve in situ using reverse transcription-polymerase chain reaction incombination with Western blot and immunochemistry with speci®c antibodies. GLAST was localized to interfascicularoligodendrocytes, whereas a subpopulation of cells, probably immature oligodendrocyte cells, expressed EAAC1. In contrast,astrocytes, expressed only GLT-1, consistent with the idea that this is the major glutamate transporter in this cell type. In addition, weobserved that glutamine synthetase, a key enzyme in glutamate metabolism, was localized in oligodendrocytes in situ. To examinethe properties of these glutamate transporters, we conducted uptake experiments in glial cultures. The kinetics of sodium-dependentglutamate uptake in cultured oligodendrocytes from the perinatal rat optic nerve were markedly different from those observed in type-1 astrocytes from the newborn rat cerebral cortex, with higher af®nity and lower Vmax. In both cell types, glutamate transport wasinhibited by L-trans-pyrrolidine-2,4-dicarboxylate (t-PDC). In contrast, dihydrokainate exhibited signi®cantly more uptake inhibition inoligodendrocytes than in type-1 astrocytes. These results provide evidence for the expression of functional sodium-dependentglutamate transporters in optic nerve oligodendrocytes, and suggest that this cell type may play a role in the glutamate±glutaminecycle.
Introduction
Glutamate is the major excitatory neurotransmitter in the mammalian
CNS, and its uptake is essential to maintain a low concentration of
extracellular glutamate and to prevent excitotoxicity (Choi, 1992;
Rothstein et al., 1996). Clearance of glutamate is carried out by high-
af®nity, sodium-dependent glutamate transporters localized in
neuronal and glial cell membranes. The molecular properties and
function of these transporters have recently been elucidated (for a
review see Kanai et al., 1997). At least ®ve glutamate transporters
have been cloned: GLAST (Storck et al., 1992), GLT-1 (Pines et al.,
1992), EAAC1 (Kanai & Hediger, 1992), EAAT4 (Fairman et al.,
1995) and EAAT5 (Arriza et al., 1997). Localization studies indicate
that EAAC1 is present in neurons throughout the brain (Kanai &
Hediger, 1992). GLAST was found to be abundantly expressed in
cerebellar Bergmann glia (Storck et al., 1992), while GLT-1 was
observed in astroglia throughout the brain (Danbolt et al., 1992).
Finally, EAAT4 is expressed in cerebellar Purkinje cells (Fairman
et al., 1995) and EAAT5 is located in cells of the retina (Arriza et al.,
1997). Based on these studies, EAAC1 is regarded as the major
neuronal glutamate transporter, while GLAST and GLT-1 are viewed
as the principal glial glutamate transporters (Rothstein et al., 1994).
Glutamate transporters are also located in non-synaptic regions
(Lehre et al., 1995; Choi & Chiu, 1997), but their properties as well as
the processes which they subserve are presently unclear. In the optic
nerve, it has been proposed that neurons may signal to glia along their
axons via a calcium-independent mechanism, due to reversal of
glutamate transporters present at nodes of Ranvier (Kriegler & Chiu,
1993). This feature, together with the fact that glutamate receptor
activation can lead to oligodendroglial excitotoxicity (Yoshioka et al.,
1996; Matute et al., 1997; Matute, 1998; McDonald et al., 1998),
strongly suggests that the extracellular glutamate concentration is
regulated by transporters in order to prevent oligodendrocyte cell
death and demyelination. There is little information, however, about
the type of transporters mediating glutamate uptake in axonal tracts of
the CNS.
In the present study, we analysed the expression of EAAC1,
GLAST and GLT-1, the most widely distributed glutamate
transporters, in the adult optic nerve using reverse transcription-
polymerase chain reaction (RT-PCR), Western blot analysis and
immunochemistry. The results of these experiments led us to analyse
glutamate uptake in cultured oligodendroglial cells and to compare it
with that in cultured astrocytes. Taken together, our results strongly
suggest that oligodendrocytes contribute to the maintenance of low
levels of extracellular glutamate. In addition, we found that glutamine
synthetase, a key enzyme in glutamate metabolism, was expressed in
oligodendrocytes from the optic nerve in situ.
Materials and methods
Materials
Culture reagents were purchased from Sigma (St. Louis, MO, USA)
unless otherwise stated. L-[3H]-Glutamate (speci®c activity 46 Ci/
mmol) was from Amersham Life Sciences (Buckinghamshire, UK)
and growth factors were from Alomone (Jerusalem, Israel). Drugs
L
Correspondence: C. Matute, as above. E-mail: [email protected]
Received 12 August 1998, revised 28 January 1999, accepted 11 February1999
European Journal of Neuroscience, Vol. 11, pp. 2226±2236, 1999 Ó European Neuroscience Association

acting on glutamate transporters were obtained from Tocris Cookson
(Bristol, UK) except for glutamate (Sigma).
Cerebral cortical cultures
Primary astrocyte cultures were established from the cerebral cortex
of newborn Sprague±Dawley rats and prepared according to the
procedure described by McCarthy & De Vellis (1980) with minor
modi®cations. Brie¯y, cerebral cortices were dissected from 1-day-
old rats, incubated at 37 °C for 15 min in calcium- and magnesium-
free Hank's balanced salt solution, and dissociated mechanically.
Cells were then seeded into a 75-cm2 tissue culture ¯ask precoated
with poly-D-lysine and grown in Iscove's modi®ed Dulbecco's
medium (Gibco BRL, Paisley, UK) supplemented with 10% foetal
bovine serum, 50 U/mL penicillin, 50 mg/mL streptomycin and
2.5 mg/mL amphotericin B. Cells were cultured at 37 °C in a
humidi®ed 95% air/5% CO2 incubator for 13±15 days. When cells
reached con¯uence (days 10±12), the ¯asks were shaken for 1 h
(37 °C, 200 r.p.m.) to detach microglial cells. The decanted medium
was poured into Petri dishes for bacterial cultures, to which microglia
selectively adhere. Microglia were subsequently cultured in the
astrocyte-cultured medium described above. Flasks were once again
shaken for an additional 14 h (37 °C, 200 r.p.m.) to remove O-2A
progenitor cells. The remaining cell layer, formed by type-1
astrocytes (typically > 95% of the cells), was trypsinized and the
dissociated cells were plated onto poly-D-lysine-precoated glass
coverslips (12 mm) at a density of 5 3 103 cells/well for immunocyto-
chemistry and 5 3 104 cells/well for L-[3H]-glutamate uptake experi-
ments, cultured as above, and used 3±5 days later.
Optic nerve cultures
Oligodendrocyte cultures were obtained from optic nerves of 12-day-
old Sprague±Dawley rats using the procedure described by Barres
et al. (1992). Nerves were dissected out, minced and incubated at
37 °C for 40 min with 1.25 mg/mL collagenase, 0.04 mg/mL DNase,
0.125% trypsin/0.02% EDTA in Ca2+- and Mg2+-free Hank's
balanced salt solution. The enzymatic reaction was then stopped
with 10% foetal bovine serum. The cells were centrifuged at 220 g,
and the pellet was mechanically dissociated in 10% foetal bovine
serum in Dulbecco's modi®ed Eagle's medium (Gibco BRL). The
cells were pelleted at 220 g for 10 min, resuspended in a chemically
de®ned medium (Barres et al., 1992), and seeded onto poly-D-lysine-
coated glass coverslips (12 mm diameter) at 5 3 103 and 25 3 103
cells/well for immunocytochemistry and L-[3H]-glutamate uptake
experiments, respectively. Cultures were maintained at 37 °C and 5%
CO2 for 3±7 days before use, and fresh medium was added every day.
RNA isolation and RT-PCR analysis
Total RNA was extracted from adult rat whole brain and optic nerve
samples, and from type-1 astrocyte cultures and oligodendrocyte
cultures using the guanidinium/phenol/chloroform method
(Chomczynski & Sacchi, 1987). Rat brain tissues were dissected
out from deeply anaesthetized animals and snap frozen in liquid
nitrogen. Reverse transcription into cDNA was carried out using
random hexamer primers and avian myeloblastosis virus-reverse
transcriptase as described by the supplier (USB, OH, USA).
Oligonucleotide primers used for PCR ampli®cation are described
in Table 1. Each pair of primers ef®ciently detected the corresponding
RNA transcript by RT-PCR in samples of rat brain. The reaction
mixture was heated to 95 °C for 2 min, cooled to 80 °C, and then
1.25 U Taq DNA polymerase (Dynazyme; Finnzymes, Espoo,
Finland) was added at this temperature. PCR ampli®cation was
carried out in thin-walled tubes for a total of 40 cycles as follows:
95 °C for 50 s, the average of the melting temperature of the
corresponding oligonucleotide pair + 5 °C for 50 s, 72 °C for 50 s, for
two cycles; subsequently the annealing temperature was lowered by
1 °C every second cycle until it was 9 °C lower than the starting
annealing temperature. Finally, 22 more cycles were run at the ®nal
annealing temperature. Ampli®ed products were analysed by
electrophoresis in 1.8% agarose gels and viewed with ethidium
bromide staining. A FX174 HaeIII digest was used as a size standard.
The speci®city of the ampli®ed PCR products was assessed by DNA
sequencing by using the dideoxy chain termination method (Sanger
et al., 1977). Standard protocols of the manufacturer for Taq DNA
polymerase-initiated cycle sequencing reactions with ¯uorescently
labelled dideoxynucleotide terminators (Applied Biosystems, Foster
City, CA, USA) were used. The sequencing reactions were analysed
using a 377 automated DNA sequencer (Applied Biosystems).
Western blot analysis
The speci®city of the antibodies and the expression of glutamate
transporters in rat optic nerve were assessed by immunoblotting using
adult rat hippocampus and cerebellum as controls. Af®nity-puri®ed
polyclonal antibodies to C-terminal EAAC1, N-terminal GLAST and
C-terminal GLT-1 epitopes were generously provided by Dr J.D.
Rothstein (Rothstein et al., 1994, 1996). Tissue was collected from
fresh rat brains, dissected and homogenized in 20 mM Tris±HCl
(pH 7.4), 10% sucrose, 1 mM EDTA, 5 mM EGTA, 1 mM phenyl-
methylsulphonic ¯uoride, 10 mM benzamidine, 20 mg/mL leupeptin,
and 0.1 mM pepstatin with a dounce homogenizer (Heidolph,
Germany). Following centrifugation of the homogenate (10 000 g
for 30 min), the supernatants were diluted 1 : 1 with an electrophoresis
sample buffer (Laemmli, 1970) and then boiled for 5 min. Proteins
were separated by sodium dodecyl sulphate±polyacrylamide gel
electrophoresis (SDS±PAGE, 7.5% polyacrylamide gels) and then
transferred to polyvinylidene ¯uoride membranes (Immobilon P,
Millipore) with 5% methanol in 10 mM 3-cyclohexylamino-1-
propanesulphonic acid (pH 10.5) by means of a semidry electroblotter
(Sartorius AG, Goettingen, Germany). Non-speci®c binding was
blocked for 1 h at room temperature with 10% non-fat milk and 2%
bovine serum albumin diluted in 50 mM Tris±HCl, 200 mM NaCl and
0.1% Tween-20 (pH 7.4). Blots were incubated for 48 h at 4 °C with
af®nity-puri®ed antibodies to EAAC1 (0.6 mg/mL), GLAST (0.2 mg/
mL) and GLT-1 (0.034 mg/mL) in the same buffer containing 1%
non-fat milk. After washing, blots were ®nally incubated with
horseradish peroxidase-conjugated antirabbit antibody (1 : 1000,
Sigma). Immunoreactive proteins were visualized with an enhanced
chemiluminescence substrate (Super Signal ULTRA; Pierce,
Rockford, IL, USA).
Immunochemistry
Expression of glutamate transporters was examined by immunohis-
tochemistry using the antibodies described above. All the results with
the antibody to N-GLAST were corroborated with antibodies A1 (to
amino acid residues 1±25) generously provided by Dr N.C. Danbolt
(Lehre et al., 1995). Rat optic nerve samples were dissected after
perfusing animals with 4% paraformaldehyde in 0.1 M phosphate
buffer (pH 7.4), post®xed for 2 h in the same solution, cryoprotected
in 30% sucrose and mounted with Tissue-Tek (Sakura Finetek USA,
Torrance, CA, USA). Cryostat sections (20 mm thick) were processed
for immunohistochemistry in free-¯oating sections as previously
described (Rothstein et al., 1994). Sections were preincubated (1 h)
with 4% normal goat serum diluted in 0.1% Triton X-100 and 0.02%
sodium azide in phosphate-buffered saline (PBS), and were then
incubated in the af®nity-puri®ed antitransporter protein antibody for
RGlutamate uptake in oligodendroglia 2227
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

48 h at 4 °C at a concentration of 66 ng/mL for EAAC1, 16 ng/mL for
GLAST and 148 ng/mL for GLT-1 in PBS containing 2% normal goat
serum, 0.1% Triton X-100 and 0.02% sodium azide. Immuno-
detection was carried out using biotinylated secondary antibodies
(goat antirabbit IgG; Vector Laboratories, Burlingame, CA, USA)
followed by incubation with an avidin±biotin±peroxidase complex
(Vector Laboratories) as indicated by the supplier, and developed
using a standard diaminobenzidine reaction. Antibodies to glutamine
synthetase (Transduction Laboratories, Lexington, KY, USA) and to
glutathione-S-transferase Yp subunit (GST-Yp; Biotrin Int., Dublin,
Ireland) were used at concentrations of 0.8 mg/mL and 1 : 5000,
respectively. Negative controls in all immunostaining experiments
included the omission of the primary antibody, incubation with
control, non-immune IgGs, and overnight antibody preabsorption
with a 10- and 100-fold excess of the corresponding synthetic
EAAC1, GLAST and GLT-1 peptides, and yielded no labelling.
Double immuno¯uorescent labelling experiments were carried out
at 4 °C. Sections were incubated with antibodies to glutamate
transporters GLAST or GLT-1, and with mouse monoclonal
antibodies to the following glial markers, glial ®brillary acidic
protein (anti-GFAP; 0.2 mg/mL; Boehringer Mannheim, Mannheim,
Germany) and 2¢,3¢-cyclic nucleotide 3¢-phosphodiesterase (anti-
CNPase; 1 : 100; Sigma). Primary antibody binding was viewed with
a ¯uorescein isothiocyanate (FITC)-conjugated goat antirabbit IgG
(AlexaTM 488; 1 : 200; Molecular Probes, Eugene, OR, USA) and
with biotinylated antibodies to mouse IgGs (1 : 200; Vector
Laboratories) followed by a streptavidin±tetramethyl rhodamine
isothiocyanate-conjugate (1 : 100; Chemicon, Temecula, CA, USA).
To characterize the glial cell types present in our cultures,
immuno¯uorescent labelling experiments were performed with
antibodies to GFAP (20 mg/mL; Dako DiagnoÂsticos S.A., Barcelona,
Spain) and galactocerebroside C (anti-Gal C, 5 mg/mL; Boehringer
Mannheim). Microglia were labelled with FITC-conjugated Griffonia
simplicifolia B4-isolectin (20 mg/mL, Vector Laboratories), as
previously described (Gottlieb & Matute, 1997). Cells were washed
with PBS, ®xed in 4% paraformaldehyde in PBS for 20 min, and then
processed for immunocytochemistry as described above.
L-[3H]-Glutamate uptake experiments
Sister cultures of those used for immunocytochemistry were assayed
for L-[3H]-glutamate transport. Uptake was assessed at 37 °C in
300 mL saline solution containing (in mM): NaCl, 140; KCl, 5; MgCl2,
2; CaCl2, 2; HEPES, 10; glucose, 4.5 g/L (pH 7.4). Cells were
equilibrated for 5 min in the absence or presence of uptake inhibitors,
and then incubated for 10 min with 100 nM L-[3H]-glutamate and
unlabelled glutamate (0±1000 mM). Experiments using uptake in-
hibitors were performed with 100 nM L-[3H]-glutamate. In all studies,
uptake was stopped with two washes in ice-cold PBS, followed
immediately by cell lysis in 0.1 N NaOH/0.1% Triton X-100. Aliquots
were taken for scintillation counting and for protein assays (Lowry
et al., 1951) using bovine serum albumin standards. Sodium-
dependent uptake was calculated to be the difference between the
amount of radioactivity in the presence of sodium and the amount
observed in the choline-containing buffer (sodium chloride was
substituted for equimolar concentration of choline chloride).
Preliminary studies showed that uptake was linear over at least
30 min (data not shown).
Data analysis
Eadie±Hofstee transformations of the concentration-dependence of
uptake were ®t by linear regression analysis. Statistical comparisons
were performed by repeated-measures analysis of variance (ANOVA)
with a Fisher's protected least signi®cant difference (PLSD) post hoc
analysis or by a Student's two-tailed t-test. Data are represented as the
mean 6 SEM of at least three independent experiments performed in
triplicate.
L
TABLE 1. Oligonucleotide primers and PCR conditions
Annealing MgCl2 ProductSubtype Upstream primer (5¢±3¢) Downstream primer (5¢±3¢) temperature (°C) (mM) size (bp)
EAAC1 TGGCCACTGTCCTGAGTGGG CTAGAACTGCGAGGTCTGAG 68 2.5 719GLAST CCTCTCCTCTACTTCCTGGTA CTACATCTTGGTTTCGCTGT 55 1 633GLT-1 CTTGATTTACTTTGTAGTGA AAAGGAAGCCTGTTTAGAGC 59 3.5 805
Oligonucleotide sequences were used as primers in PCR experiments. Reaction parameters were empirically determined.
FIG. 1. RT-PCR with speci®c primers for the glutamate transporters EAAC1, GLAST and GLT-1 in rat brain, optic nerve, and astrocyte and oligodendrocytecultures derived from cerebral cortex and optic nerve, respectively. RT-PCR conditions were suitable to ef®ciently amplify the corresponding transcripts in wholerat brain samples used as a positive control (rwb). The three genes are expressed in the rat optic nerve (ron), and in astrocyte (ast) and oligodendrocyte cultures(olig). Molecular standards in base pairs correspond to the FX174 HaeIII digest. The numbers on the left indicate size in base pairs.
2228 M. Domercq et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

Results
Expression of glutamate transporters in situ
We ®rst examined the expression of RNAs encoding the glutamate
transporters EAAC1, GLAST and GLT-1 in rat optic nerve. Primers
were designed to amplify fragments from a C-terminal region of each
transporter (Table 1). The speci®c primer pairs used in these
experiments ef®ciently ampli®ed the corresponding glutamate
transporter transcripts in rat whole brain samples (Fig. 1), as shown
by gel electrophoresis and subsequent sequencing analysis. In
contrast, negative controls in which the cDNA was substituted with
water yielded no amplicons (data not shown). Analysis using RT-
PCR of RNA extracted from the adult rat optic nerve resulted in
fragments of the expected size using the primers speci®c for the
transporters EAAC1, GLAST and GLT-1 (Fig. 1).
Because the optic nerve is devoid of neuronal cell bodies, and
axons are not known to contain RNA, it is most likely that the PCR
ampli®ed products arise from RNA present in glial cells. To con®rm
this idea and to determine the phenotype of the cells expressing the
three glutamate transporters examined, we carried out Western blot
analysis and immunohistochemical experiments in rat optic nerve
sections. The speci®city of the antibodies has been assessed
previously (Rothstein et al., 1994, 1996) and was corroborated by
Western blot analysis of control samples. Thus, antibodies to EAAC1
recognized a » 69-kDa protein in hippocampus homogenates, antisera
to GLAST recognized a » 65-kDa protein in cerebellum homo-
genates, and antisera to GLT-1 recognized a » 72-kDa protein in
hippocampus homogenates (Fig. 2). The larger molecular bands may
correspond to dimers and/or larger aggregates, as shown previously
(Haugeto et al., 1996). In rat optic nerve homogenates, a high level of
GLAST and GLT-1 expression was detected, whereas EAAC1 was
expressed at lower levels (Fig. 2). In all instances, the apparent size of
the transporters was similar in the optic nerve and the brain areas
examined. However, the levels of expression were higher in the latter.
Immunohistochemistry with antibodies to EAAC1, GLAST and
GLT-1 strongly stained morphologically different types of glial cells
in rat optic nerve (Fig. 3). EAAC1 was detected in relatively few cells
that had no particular orientation, with round or elongated shape, and
a variable number (between one and ®ve) of branches (Fig. 3A and
B). A representative sample of EAAC1+ cells was drawn using a
camera lucida (see Fig. 5 below). GLAST was expressed in small
rounded cells with no visible processes frequently grouped forming
short rows of cells orientated parallel to axonal ®bres (Fig. 3C), a
feature that corresponds to interfascicular oligodendrocytes (Peters
et al., 1991). Finally, the expression of GLT-1 was detected in cells
randomly distributed among neuronal ®bres. These cells exhibited
stellate morphology with small somata and a large number of radial
processes (Fig. 3D), typical features of the ®brous astrocytes reported
to be present in the white matter (Peters et al., 1991).
Double-labelling experiments con®rmed the expression of GLAST
in CNPase-stained oligodendrocytes and GLT-1 mainly in GFAP+
astrocytic processes (Fig. 4). The EAAC1+ cells were not labelled
with astroglial (GFAP), oligodendroglial (CNPase) or microglial
(Griffonia simplicifolia B4-isolectin) markers, and their morphology
was comparable to that of a representative sample of GST-Yp+ cells
(Fig. 5), a marker of immature oligodendrocytes (Gensert &
Goldman, 1996).
Glutamine synthetase in rat optic nerve
To test the possibility that glial cells in the optic nerve metabolize the
glutamate taken up into these cells, we analysed the expression of
glutamine synthetase in the optic nerve using immunohistochemistry.
Our results, in accordance with previous data (Tansey et al., 1991),
showed that optic nerve glutamine synthetase was expressed in
oligodendrocytes and ®brous astrocytes (Fig. 6). Glutamate synthe-
tase in the latter cell type was particularly abundant in the optic
chiasm (Fig. 6B).
Glutamate transport assays in cultured glial cells
We next investigated the kinetics of sodium-dependent glutamate
transport in oligodendrocyte cultures obtained from optic nerves of
12-day-old rats. After 3 days in vitro, cultures consisted almost
exclusively of Gal C+ oligodendrocytes, as previously described
(Matute et al., 1997). In addition, we used astrocyte cultures from
newborn rat cerebral cortex as a positive control and to compare the
kinetics of glutamate transporters in both cell types.
Kinetic analysis showed signi®cant differences in sodium-
dependent glutamate uptake by oligodendrocytes versus type-1
R
FIG. 2. Western blot analysis of the glutamate transporter subtypes expressed in the rat optic nerve. Samples (25 mg of protein per lane) were subjected to SDS±PAGE, and immunoblotted with antibodies raised against synthetic peptides corresponding to EAAC1, GLAST and GLT-1 (Rothstein et al., 1994, 1996). Thelevels of EAAC1, GLAST and GLT-1 were lower in the rat optic nerve than in the hippocampus (EAAC1 and GLT-1) and cerebellum (GLAST), the regions usedas positive controls. hpc, hippocampus; cb, cerebellum; on, optic nerve from rat brain.
Glutamate uptake in oligodendroglia 2229
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

astrocytes. The apparent glutamate Km and the maximal velocity
(Vmax) values in oligodendrocytes were 5.2 6 0.8 mM and
1.28 6 0.14 nmol/min/mg protein, much lower than those in type-1
astrocytes, 91 6 4.5 mM and 25.6 6 1.04 nmol/min/mg protein, re-
spectively (Fig. 7A and B). In contrast, total glutamate uptake by
microglial cultures (95% pure or greater) derived from mixed glial
cultures from newborn rat cerebral cortex was lower than 0.05% of
that measured in type-1 astrocyte and oligodendrocyte cultures.
The relative ef®cacy of glutamate transporter inhibitors and the
ionic dependence of glutamate transport in the two types of glial
cultures are shown in Fig. 7C. We observed that in both astrocyte and
oligodendrocyte cultures, L-[3H]-glutamate uptake was almost
completely abolished by preincubation with L-trans-pyrrolidine-2,4-
dicarboxylate (t-PDC, 500 mM), a transportable inhibitor of glutamate
uptake, indicating that the observed ef¯ux is mostly due to a
transporter-mediated mechanism. In contrast, dihydrokainate (DHK,
1 mM), a non-transportable inhibitor of the glutamate transporter
GLT-1, inhibited glutamate uptake to only a small degree in
astrocytes (23.5%; P < 0.05), while it almost completely blocked it
in oligodendrocytes (84.8%). Substitution of extracellular sodium
with choline inhibited the glutamate response by over 95% in both
astrocytes and oligodendrocytes. Taken together, these data indicate
that oligodendrocytes and type-1 astrocytes exhibit high-af®nity
sodium-dependent glutamate transport, although its ef®ciency
appears to be different in the two cell types.
Expression of glutamate transporter subtypes in vitro
RT-PCR of RNA extracted from cultured astrocytes and oligo-
dendrocytes resulted in the ampli®cation of EAAC1, GLAST and
GLT-1 (Fig. 1). Although the cultures used in these experiments were
highly enriched in either type-1 astrocytes or oligodendrocytes, the
high sensitivity of the PCR technique does not provide information as
to whether the ampli®ed transcripts are expressed by the main cell
types present in those cultures or by contaminant cells. To address
this problem, we analysed these cultures using immunochemistry
with speci®c antibodies to each of the three glutamate transporters
(Rothstein et al., 1994, 1996). Type-1 astrocytes were immuno-
labelled with antibodies to GLAST and GLT-1, but not EAAC1
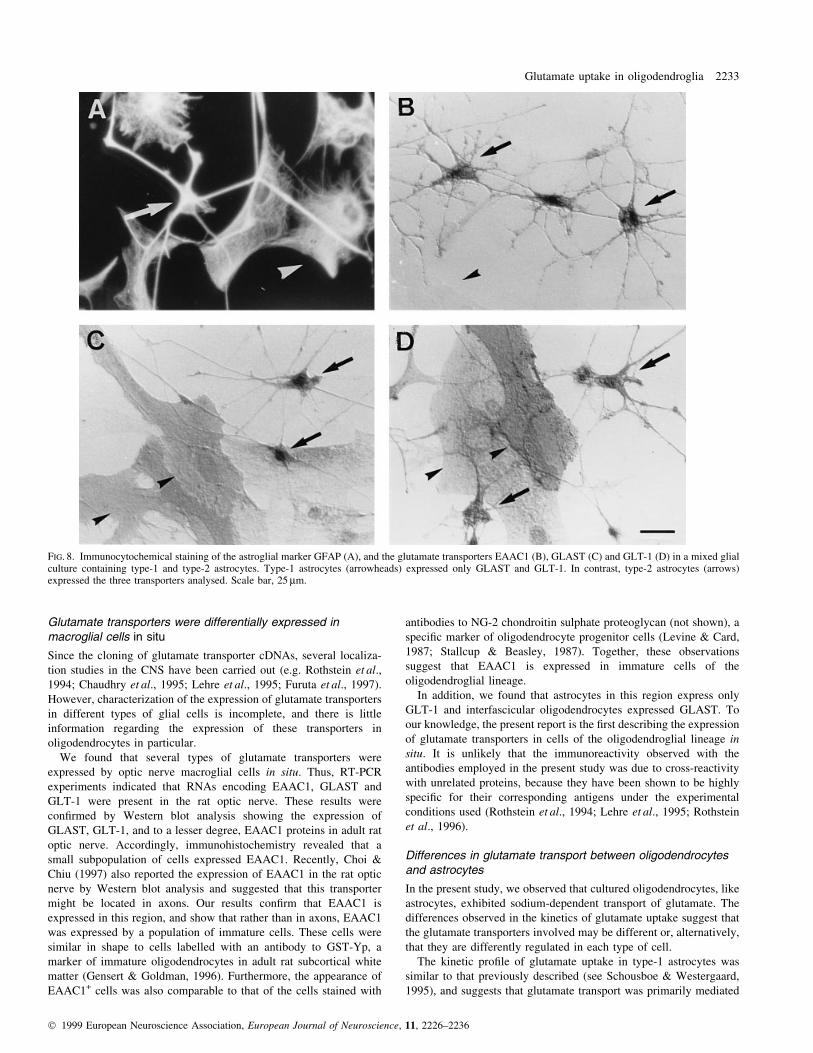
(Fig. 8). In contrast, the three transporters were expressed in a very
small subpopulation of cells present in the astrocyte cultures, which
were GFAP+, phase-dark and stellate in morphology, and therefore
identi®ed as type-2 astrocytes (Fig. 8).
L
FIG. 3. Immunohistochemical staining in the rat optic nerve with antibodies to EAAC1 (A and B), GLAST (C) and GLT-1 (D). EAAC1 was expressed in a smallsubpopulation of cells. GLAST was present in interfascicular oligodendrocytes, whereas GLT-1 was restricted to ®brous astrocytes. Scale bar, 50 mm (A,C and D),10 mm (B).
2230 M. Domercq et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

In optic nerve oligodendrocyte cultures, the three transporters
analysed were present at all the stages examined (3 h to 3
days after seeding), as shown in Fig. 9. The subcellular
distribution of each glutamate transporter was different. EAAC1
was located in the soma and proximal and distal processes,
while GLAST was present in the soma and proximal parts of the
oligodendroglial branches, and GLT-1 was restricted to the
soma.
R
FIG. 4. Immuno¯uorescent double-labelling of CNPase and GLAST (A and A¢, and B and B¢, respectively), and GFAP and GLT-1 (C and D, respectively) inlongitudinal sections of the adult rat optic nerve. Each pair of photographs was taken from the same ®eld. The GLAST transporter was present in CNPase+
oligodendrocytes (small arrows) and GLT-1 was mainly located in GFAP+ astrocytic processes (large arrows). Scale bar, 50 mm.
FIG. 5. Camera lucida drawings of a representative group of EAAC1+ and GST-Yp+ cells. Notice the morphological similarity between both groups of cells.Although some of the cells appear quite complex in morphology, they are still much simpler than multibranched astrocytes or microglial cells, and can be clearlydistinguished from interfascicular oligodendrocytes. Scale bar, 50 mm.
Glutamate uptake in oligodendroglia 2231
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

No immunolabelling with antibodies to EAAC1, GLAST or GLT-1
was observed in any of the microglial cultures assayed (not shown), a
characteristic that correlates well with the lack of signi®cant
glutamate uptake observed in these cells.
Discussion
The main ®nding of this work is that oligodendroglia from rat optic
nerve express glutamate transporters both in situ and in vitro.
Cultured oligodendroglia in vitro exhibit sodium-dependent gluta-
mate transport with a higher af®nity and lower ef®ciency, compared
to type-1 astrocytes.
L
FIG. 6. Immunohistochemical staining in the rat optic nerve (A) and opticchiasm (B) with antibodies to glutamine synthetase. The enzyme was localizedmainly in interfascicular oligodendrocytes (arrowheads) and in a subpopula-tion of ®brous astrocytes (arrows). Scale bar, 50 mm.
FIG. 7. Properties of glutamate transport in cultured type-1 astrocytes andoligodendrocytes. (A and B) Eadie±Hofstee plots of concentration depen-dence of sodium-dependent L-[3H]-glutamate uptake in type-1 astrocytes (A)and oligodendrocytes (B). Uptake velocity (in nmol/min/mg protein) wasmeasured using 100 nM radiolabelled glutamate and increasing concentra-tions of unlabelled glutamate (0.1±1000 mM). For astrocytes: Vmax = 25.6 n-mol/min/mg protein; Km = 91 mM; r2 = 0.99. For oligodendrocytes:Vmax = 1.28 nmol/min/mg protein; Km = 5.8 mM; r2 = 0.94. Symbols representmeans 6 SEM of at least three independent experiments performed in tripli-cate. (C) The pharmacological pro®le and ionic dependence of L-[3H]-gluta-mate uptake. Uptake of 100 nM L-[3H]-glutamate was assessed in thepresence of the indicated amount of inhibitor and expressed as a percentageof the uptake rate in the corresponding control culture type. Transport inboth cell types was inhibited by t-PDC (500 mM) and when sodium was sub-stituted by choline. In contrast, DHK (1 mM) exhibited signi®cantly more in-hibition of transport in oligodendrocyte cultures than in type-1 astrocytecultures (84.8% versus 23.5%, respectively; P < 0.01 by two-tailed Student'st-test). Bars represent means 6 SEM (n = 3) of at least three independent ex-periments performed in triplicate. *P < 0.05 by ANOVA with Fisher's PLSDpost hoc analysis.
2232 M. Domercq et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236
Glutamate transporters were differentially expressed inmacroglial cells in situ
Since the cloning of glutamate transporter cDNAs, several localiza-
tion studies in the CNS have been carried out (e.g. Rothstein et al.,
1994; Chaudhry et al., 1995; Lehre et al., 1995; Furuta et al., 1997).
However, characterization of the expression of glutamate transporters
in different types of glial cells is incomplete, and there is little
information regarding the expression of these transporters in
oligodendrocytes in particular.
We found that several types of glutamate transporters were
expressed by optic nerve macroglial cells in situ. Thus, RT-PCR
experiments indicated that RNAs encoding EAAC1, GLAST and
GLT-1 were present in the rat optic nerve. These results were
con®rmed by Western blot analysis showing the expression of
GLAST, GLT-1, and to a lesser degree, EAAC1 proteins in adult rat
optic nerve. Accordingly, immunohistochemistry revealed that a
small subpopulation of cells expressed EAAC1. Recently, Choi &
Chiu (1997) also reported the expression of EAAC1 in the rat optic
nerve by Western blot analysis and suggested that this transporter
might be located in axons. Our results con®rm that EAAC1 is
expressed in this region, and show that rather than in axons, EAAC1
was expressed by a population of immature cells. These cells were
similar in shape to cells labelled with an antibody to GST-Yp, a
marker of immature oligodendrocytes in adult rat subcortical white
matter (Gensert & Goldman, 1996). Furthermore, the appearance of
EAAC1+ cells was also comparable to that of the cells stained with
antibodies to NG-2 chondroitin sulphate proteoglycan (not shown), a
speci®c marker of oligodendrocyte progenitor cells (Levine & Card,
1987; Stallcup & Beasley, 1987). Together, these observations
suggest that EAAC1 is expressed in immature cells of the
oligodendroglial lineage.
In addition, we found that astrocytes in this region express only
GLT-1 and interfascicular oligodendrocytes expressed GLAST. To
our knowledge, the present report is the ®rst describing the expression
of glutamate transporters in cells of the oligodendroglial lineage in
situ. It is unlikely that the immunoreactivity observed with the
antibodies employed in the present study was due to cross-reactivity
with unrelated proteins, because they have been shown to be highly
speci®c for their corresponding antigens under the experimental
conditions used (Rothstein et al., 1994; Lehre et al., 1995; Rothstein
et al., 1996).
Differences in glutamate transport between oligodendrocytesand astrocytes
In the present study, we observed that cultured oligodendrocytes, like
astrocytes, exhibited sodium-dependent transport of glutamate. The
differences observed in the kinetics of glutamate uptake suggest that
the glutamate transporters involved may be different or, alternatively,
that they are differently regulated in each type of cell.
The kinetic pro®le of glutamate uptake in type-1 astrocytes was
similar to that previously described (see Schousboe & Westergaard,
1995), and suggests that glutamate transport was primarily mediated
R
FIG. 8. Immunocytochemical staining of the astroglial marker GFAP (A), and the glutamate transporters EAAC1 (B), GLAST (C) and GLT-1 (D) in a mixed glialculture containing type-1 and type-2 astrocytes. Type-1 astrocytes (arrowheads) expressed only GLAST and GLT-1. In contrast, type-2 astrocytes (arrows)expressed the three transporters analysed. Scale bar, 25 mm.
Glutamate uptake in oligodendroglia 2233
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

by GLAST in this cell type. This idea is supported by the fact that
glutamate transport had a Km of 91 6 4.5 mM, which is similar to that
observed in studies of GLAST expression in Xenopus oocytes (77 mM;
Storck et al., 1992), and that DHK (1 mM), a selective and potent
blocker of GLT-1-mediated transport (Arriza et al., 1994), only
induced a slight inhibition of L-glutamate uptake. Accordingly,
GLAST was found to be expressed in cultured astrocytes. It appears
then, that GLT-1, which we also observed in astrocytes, contributes to
glutamate transport to a minor degree in astrocytes in vitro.
Consistent with this idea, DHK had only negligible effects on
astrocyte cultures (Garlin et al., 1995; Swanson et al., 1997). Our
results correlate well with previous ®ndings demonstrating the
presence of GLAST mRNA and GLT-1 mRNA in cultured astrocytes
from different regions of rat brain including the cerebral cortex
(Kondo et al., 1995), and with data indicating that undifferentiated
astrocytes express GLAST and that under certain culture conditions
they may also express GLT-1 (Swanson et al., 1997).
In oligodendrocytes, the Km of glutamate uptake was 5.2 6 0.8 mM,
suggesting that a different type of glutamate transporter is involved.
Because expression studies in liposomes/HeLa cells have indicated
that the Km for L-glutamate uptake mediated by GLT-1 is 2 mM (Kanai
& Hediger, 1992), and DHK was an ef®cient and selective inhibitor
of this transporter subtype, it is likely that glutamate uptake in
oligodendrocytes in vitro is mediated primarily by this transporter.
However, DHK did not completely block glutamate uptake, not
excluding the minor participation of other glutamate transporter
subtypes. In addition to GLT-1, we also observed that EAAC1 and
GLAST were expressed in immature and mature oligodendroglial
cells, a ®nding that is consistent with a previous report (Kondo et al.,
1995) and recent preliminary evidence (Wang et al., 1997). There-
fore, it appears that these two transporters also participate in
glutamate clearance by oligodendrocytes, although our L-glutamate
uptake assays suggest that their overall contribution may be minor.
Overall, the kinetic characteristics of glutamate uptake in cultured
oligodendrocytes were more similar to those described in neurons
than in astrocytes. Whether these differences are related to the
glutamate transporters expressed or to the cell phenotypes remains
the subject of ongoing studies.
The expression of glutamate transporters in oligodendroglia from
adult optic nerve in situ was different than that in oligodendrocyte
cultures from the same area, where we observed expression of
EAAC1, GLAST and GLT-1 at all stages examined. Because
glutamate transporter expression is developmentally regulated
(Ullesvang et al., 1997), the differences could be due to the
differences between the age of the animals used (P12 rats for
oligodendrocytes cultures versus adult rats for immunohisto-
chemistry). It is also conceivable that the expression of GLT-1 is
regulated differently in vitro than in vivo, where oligodendrocytes
contact axons and astrocytes and have a different extracellular milieu.
The latter is critical regarding the glutamate concentration bathing the
oligodendrocytes, which in our cultures was » 10±15 mM (data not
shown), whereas in white matter tracts this concentration may be
signi®cantly lower (Miele et al., 1996). This higher concentration of
glutamate in vitro may account for the increased expression of
L
FIG. 9. Immunocytochemical staining of the oligodendroglial marker Gal C (A), and the glutamate transporters EAAC1 (B), GLAST (C) and GLT-1 (D) indifferentiated oligodendrocyte cultures at 3 days in vitro. The vast majority of the cells in these cultures expressed the oligodendrocyte marker Gal C and the threetransporters analysed. Scale bar, 25 mm.
2234 M. Domercq et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

glutamate transporter types in these cultured cells. Recently, the
expression of GLT-1 has been reported in neuronal hippocampal
microcultures (Mennerick et al., 1998), whereas neuronal labelling in
situ was not reported in earlier studies, suggesting that GLT-1
expression may be subject to regulation by culture environment.
Functional signi®cance of glutamate transporters in rat opticnerve
The current understanding of glutamate signalling in the CNS
indicates that the extracellular glutamate concentration must be
spatially and temporally regulated to terminate glutamate receptor
activation and to protect neurons from excitotoxicity, hence necessi-
tating the proper functioning of glutamate carriers at and around
synapses (Choi, 1992; Rothstein et al., 1996; Tanaka et al., 1997).
However, the role of glutamate transport in non-synaptic areas is not
presently understood.
Oligodendrocytes, like neurons, are also highly vulnerable to
glutamate, and the associated toxicity is caused by sustained activation
of AMPA and kainate receptors (Yoshioka et al., 1996; Matute et al.,
1997; Matute, 1998; McDonald et al., 1998). It is therefore
conceivable that the glutamate transporters expressed by astrocytes
and oligodendrocytes in the white matter maintain the extracellular
glutamate concentration below levels that are potentially toxic to
oligodendrocytes. In axonal tracts, glutamate can be released into the
extracellular space from astrocytes (Martin, 1992; Levi & Gallo,
1995), and from the axons after propagation of action potentials
(Weinreich & Hammerschlag, 1975) by reversal of glutamate
transporters at the nodes of Ranvier (Kriegler & Chiu, 1993). Thus,
impaired glutamate clearance in white matter may affect the viability
of oligodendrocytes. Because oligodendrocytes are the primary
damaged cell type in multiple sclerosis (Prineas & McDonald,
1997), it is possible that aberrant functioning of glutamate transporters
contributes to this leukoencephalopathy. Interestingly, a recently
mapped multiple sclerosis susceptibility locus involves the gene
encoding GLAST (Becker et al., 1997).
In the mammalian CNS, glutamate released from neurons is taken
up by astrocytes and rapidly converted to glutamine due to the
presence of glutamine synthetase in such cells. Glutamine is then
released and taken up into neurons where it undergoes amidation to
form glutamate and ammonia (Benjamin & Quastel, 1972, 1975).
Neuron terminals appear to be completely dependent on this cycle to
replenish the neurotransmitter pool of glutamate (Laake et al., 1995).
Recent studies have found that glutamine synthetase colocalizes with
GLAST transporter in MuÈller glial cell processes ensheathing
glutamatergic synapses (Derouiche & Rauen, 1995). The fact that
oligodendrocytes in the optic nerve express glutamine synthetase and
GLAST suggests that this cell type may participate in the glutamate±
glutamine cycle and provide axons with glutamine, as a source for
glutamate. Indeed, glutaminase immunoreactivity has been described
in axons and adaxonal glia in the cray®sh giant nerve ®bre (McKinnon
et al., 1995).
In summary, we have demonstrated that oligodendrocytes express
glutamate transporters in vitro and in situ. These results support the
idea that glutamate serves as an axon-to-glial signal and provide
evidence that the glutamate±glutamine cycle may modulate the
concentration of periaxonal glutamate in a manner similar to that
described for perisynaptic regions.
Acknowledgements
We are grateful to D.J. Fogarty for reviewing the manuscript, and to Dr J.D.Rothstein (Johns Hopkins University, Baltimore) and Dr N.C. Danbolt (Oslo
University, Norway) for the gift of polyclonal antibodies to the glutamatetransporters examined. This work was supported by grants from the GobiernoVasco (94/106, 96/17) and the Ministerio de EducacioÂn y Cultura (PB94-1377). M.D. held a fellowship from the Gobierno Vasco.
Abbreviations
CNPase, 2¢,3¢-cyclic nucleotide 3¢-phosphodiesterase; DHK, dihydrokainate;FITC, ¯uorescein isothiocyanate; Gal C, galactocerebroside C; GFAP, glial®brillary acidic protein; GST-Yp, glutathione-S-transferase Yp subunit;PBS, phosphate-buffered saline; RT-PCR, reverse transcription-polymerasechain reaction; SDS±PAGE, sodium dodecyl sulphate±polyacrylamide gelelectrophoresis; t-PDC, L-trans-pyrrolidine-2,4-dicarboxylate.
References
Arriza, J.L., Eliasof, S., Kavanaugh, M.P. & Amara, S.G. (1997) Excitatoryamino acid transporter 5, a retinal glutamate transporter coupled to achloride conductance. Proc. Natl Acad. Sci. USA, 94, 4155±4160.
Arriza, J.L., Fairman, W.A., Wadiche, J.I., Murdoch, G.H., Kavanaugh, M.P.& Amara, S.G. (1994) Functional comparisons of three glutamatetransporter subtypes cloned from human motor cortex. J. Neurosci., 14,5559±5569.
Barres, B.A., Hart, I.K., Coles, H.S.R., Burne, J.F., Voyvodic, J.T.,Richardson, W.D. & Raff, M.C. (1992) Cell death and control of cellsurvival in the oligodendrocyte lineage. Cell, 70, 31±46.
Becker, K.G., Mattson, D.H., Powers, J.M., Gado, A.M. & Biddison, W.E.(1997) Analysis of a sequenced cDNA library from multiple sclerosislesions. J. Neuroimmunol., 77, 27±38.
Benjamin, A.M. & Quastel, J.H. (1972) Localizations of amino acids in brainslices from the rat. Biochem. J., 128, 631±646.
Benjamin, A.M. & Quastel, J.H. (1975) Metabolism of amino acids andammonia in rat brain cortex slices in vitro: a possible role of ammonia inbrain function. J. Neurochem., 25, 197±206.
Chaudhry, F.A., Lehre, K.P., van Lookeren Campagne, M., Ottersen, O.P.,Danbolt, N.C. & Storm-Mathisen, J. (1995) Glutamate transporters in glialplasma membranes: highly differentiated localizations revealed byquantitative ultrastructural immunocytochemistry. Neuron, 15, 711±720.
Choi, D.W. (1992) Excitotoxic cell death. J. Neurobiol., 23, 1261±1276.Choi, I. & Chiu, Y. (1997) Expression of high-af®nity neuronal and glial
glutamate transporters in the rat optic nerve. Glia, 20, 184±192.Chomczynski, P. & Sacchi, N. (1987) Single-step method of RNA isolation by
acid guanidinium thiocyanate-phenol-chloroform extraction. Anal.
Biochem., 162, 156±159.Danbolt, N.C., Storm-Mathisen, J. & Kanner, B.I. (1992) A [Na+ + K+]
coupled L-glutamate transporter puri®ed from rat brain is located in glialcell processes. Neuroscience, 51, 295±310.
Derouiche, A. & Rauen, T. (1995) Coincidence of L-glutamate/L-aspartatetransporter (GLAST) and glutamine synthetase (GS) immunoreactions inretinal glia: evidence for coupling of GLAST and GS in transmitterclearance. J. Neurosci. Res., 42, 131±143.
Fairman, W.A., Vanderberg, R.J., Arriza, J.L., Kavanaugh, M.P. & Amara,S.G. (1995) An excitatory amino-acid transporter with properties of aligand-gated channel. Nature, 375, 599±603.
Furuta, A., Rothstein, J.D. & Martin, L.J. (1997) Glutamate transporter proteinsubtypes are expressed differentially during rat CNS development. J.
Neurosci., 17, 8363±8375.Garlin, A.B., Sinor, A.D., Sinor, J.D., Jee, S.H., Grinspan, J.B. & Robinson,
M.B. (1995) Pharmacology of sodium-dependent high-af®nity L-[3H]-glutamate transport in glial cultures. J. Neurochem., 64, 2572±2580.
Gensert, J.M. & Goldman, J.E. (1996) In vivo characterization of endogenousproliferating cells in adult rat subcortical white matter. Glia, 17, 39±51.
Gottlieb, M. & Matute, C. (1997) Expression of ionotropic glutamate receptorsubunits in glial cells of the hippocampal CA1 area following transientforebrain ischemia. J. Cereb. Blood Flow Metabol., 17, 290±300.
Haugeto, .O., Ullesvang, K., Levy, L.M., Chaudry, F.A., HonoreÂ, T., Nielsen,M., Lehre, K.P. & Danbolt, N.C. (1996) Brain glutamate transporterproteins form homomultimers. J. Biol. Chem., 271, 27 715±27 722.
Kanai, Y. & Hediger, M.A. (1992) Primary structure and functionalcharacterization of a high-af®nity glutamate transporter. Nature, 360,467±471.
Kanai, Y., Trotti, D., Nussberger, S. & Hediger, M.A. (1997) The high af®nityglutamate transporter family: structure, function, and physiological
RGlutamate uptake in oligodendroglia 2235
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236

relevance. In Reith, M.E.A. (ed.), Neurotransmitter Transporters: Structure,Function, and Regulation. Humana, Totowa, pp. 171±213.
Kondo, K., Hashimoto, H., Kitanaka, J., Sawada, M., Suzumara, A.,Marunouchi, T. & Baba, A. (1995) Expression of glutamate transportersin cultured glial cells. Neurosci. Lett., 188, 140±142.
Kriegler, S. & Chiu, S.Y. (1993) Calcium signaling of glial cells alongmammalian axons. J. Neurosci., 13, 4229±4245.
Laake, J.H., Slyngstad, T.A., Haug, F.M.S. & Ottersen, O.P. (1995) Glutaminefrom glial cells is essential for the maintenance of the nerve terminal pool ofglutamate: immunogold evidence from hippocampal slice culture. J.Neurochem., 65, 871±881.
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature, 227, 680±685.
Lehre, K.P., Levy, L.M., Ottersen, O.P., Storm-Mathisen, J. & Danbolt, N.C.(1995) Differential expression of two glial glutamate transporters in the ratbrain: quantitative and immunocytochemical observations. J. Neurosci., 15,1835±1853.
Levi, G. & Gallo, V. (1995) Release of neuroactive amino acids from glia. InKettenmann, H. & Ransom, B.R. (eds), Neuroglia. Oxford University Press,New York, pp. 815±826.
Levine, J.M. & Card, J.P. (1987) Light and EM localization of a cell surfaceantigen (NG-2) in the rat cerebellum. J. Neurosci., 7, 2711±2720.
Lowry, O.H., Rosebrough, N.H., Farr, A.L. & Randall, R.J. (1951) Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem., 193, 265±275.
Martin, D.L. (1992) Synthesis and release of neuroactive substances by glialcells. Glia, 5, 81±94.
Matute, C. (1998) Characteristics of acute and chronic kainate excitotoxicdamage to the optic nerve. Proc. Natl Acad. Sci. USA, 95, 10 229±10 234.
Matute, C., SaÂnchez-GoÂmez, M.V., MartõÂnez-MillaÂn, L. & Miledi, R. (1997)Glutamate receptor-mediated toxicity in optic nerve oligodendrocytes. Proc.Natl Acad. Sci. USA, 94, 8830±8835.
McCarthy, K.D. & De Vellis, J. (1980) Preparation of separate astroglial andoligodendroglial cell cultures from rat cerebral tissue. J. Cell. Biol., 85,890±902.
McDonald, J.W., Althomsoms, S.P., Hyre, K.L., Choi, D.W. & Goldberg,M.P. (1998) Oligodendrocytes from forebrain are highly vulnerable toAMPA/kainate receptor-mediated excitotoxicity. Nature Med., 4, 291±297.
McKinnon, E., Hargittal, P., Grossfeld, R.M. & Lieberman, E.M. (1995)Glutamine cycle enzymes in the cray®sh giant nerve ®ber: implications foraxon-to-glia signaling. Glia, 14, 198±208.
Mennerick, S., Dhond, R.P., Benz, A., Xu, W., Rothstein, J.D., Danbolt, N.C.,Isenberg, K.E. & Zorumski, C.F. (1998) Neuronal expression of theglutamate transporter GLT-1 in hippocampal microcultures. J. Neurosci.,18, 4490±4499.
Miele, M., Berners, M., Boutelle, M.G., Kusakabe, H. & Fillenz, M. (1996)The determination of the extracellular concentration of brain glutamateusing quantitative microdialysis. Brain Res., 707, 131±133.
Peters, A., Palay, S.L. & Webster, H.D. (1991) The neuroglial cells. In Peters,
A., Palay, S.L. & Webster, H.D. (eds), The Fine Structure of the NervousSystem. Oxford University Press, New York, pp. 273±311.
Pines, G., Danbolt, N.C., Bjoras, M., Zhang, Y., Bendahan, A., Eide, L.,Koepsell, H., Storm-Mathisen, J., Seeberg, E. & Kanner, B.I. (1992)Cloning and expression of a rat brain L-glutamate transporter. Nature, 360,464±467.
Prineas, J.W. & McDonald, W.I. (1997) Demyelinating diseases. In Graham,D.I. & Lantos, P.L. (eds), Green®eld's Neuropathology. Oxford UniversityPress, New York, pp. 813±896.
Rothstein, J.D., Dykes-Hoberg, M., Pardo, C., Bristol, L.A., Jin, L., Kuncl,R.W., Kanai, Y., Hediger, M.A., Wang, Y., Schielke, J.P. & Welty, D.F.(1996) Knockout of glutamate transporters reveals a major role forastroglial transport in excitotoxicity and clearance of glutamate. Neuron,16, 675±686.
Rothstein, J.D., Martin, L., Levey, A.I., Dykes-Hoberg, M., Jin, L., Wu, D.,Nash, N. & Kuncl, R.W. (1994) Localization of neuronal and glialglutamate transporters. Neuron, 13, 713±725.
Sanger, F., Nicklen, S. & Coulson, A.R. (1977) DNA sequencing with chain-terminating inhibitors. Proc. Natl Acad. Sci. USA, 74, 5463±5467.
Schousboe, A. & Westergaard, N. (1995) Transport of neuroactive aminoacids in astrocytes. In Kettenmann, H. & Ransom, B.R. (eds), Neuroglia.Oxford University Press, New York, pp. 246±258.
Stallcup, W.B. & Beasley, L. (1987) Bipotential glial precursor cells of theoptic nerve express the NG-2 proteoglycan. J. Neurosci., 7, 2737±2744.
Storck, T., Schulte, S., Hofmann, K. & Stoffel, W. (1992) Structure, expression,and functional analysis of a Na+-dependent glutamate/aspartate transporterfrom rat brain. Proc. Natl Acad. Sci. USA, 89, 10 955±10 959.
Swanson, R.A., Liu, J., Miller, J.W., Rothstein, J.D., Farell, K., Stein, B.A. &Longuemare, M.C. (1997) Neuronal regulation of glutamate transportersubtype expression in astrocytes. J. Neurosci., 1, 932±940.
Tanaka, K., Watase, K., Manabe, T., Yamada, K., Watanabe, M., Takahashi,K., Iwama, H., Nishikawa, T., Ichihara, N., Kikuchi, T., Okuyama, S.,Kawashima, N., Hori, S., Takimoto, M. & Wada, K. (1997) Epilepsy andexacerbation of brain injury in mice lacking the glutamate transporter GLT-1. Science, 276, 1699±1702.
Tansey, F.A., Farooq, M. & Cammer, W. (1991) Glutamine synthetase inoligodendrocytes and astrocytes: new biochemical andimmunocytochemical evidence. J. Neurochem., 56, 266±272.
Ullesvang, K., Lehre, K.P., Storm-Mathisen, J. & Danbolt, N.C. (1997)Differential developmental expression of the two rat brain glutamatetransporter proteins GLAST and GLT-1. Eur. J. Neurosci., 9, 1646±1655.
Wang, J., Gan, X.D., Chung, J.H., Volpe, J.J. & Rosenberg, P.A. (1997)Strong expression of the neuronal glutamate transporter EAAC1 in ratoligodendrocytes in culture. Soc. Neurosci. Abstr., 23, 1916.
Weinreich, D. & Hammerschlag, R. (1975) Nerve impulse-enhanced release ofamino acids from non-synaptic regions of peripheral and central nervetrunks of bullfrog. Brain Res., 84, 137±142.
Yoshioka, A., Bacskai, B. & Pleasure, D. (1996) Pathophysiology ofoligodendroglial excitotoxicity. J. Neurosci. Res., 46, 427±437.
L2236 M. Domercq et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 2226±2236




















