
Active gels as a description of the actin‐myosin cytoskeleton

Ž .Brain Research 824 1999 204–217
Research report
Axonal cytoskeleton changes in experimental optic neuritis
Bing Zhu a,), G.R. Wayne Moore b, Thomas J. Zwimpfer c, Lorne F. Kastrukoff e, Jason K. Dyer d,John D. Steeves c,d, Donald W. Paty e, Max S. Cynader a
a Department of Ophthalmology, VancouÕer Hospital and Health Science Center, UniÕersity of British Columbia, 2550 Willow Street, VancouÕer, BritishColumbia, Canada V5Z 3N9
b Department of Pathology and Laboratory Medicine, VancouÕer Hospital and Health Science Center, UniÕersity of British Columbia, 910 W10th AÕenue,VancouÕer, British Columbia, Canada V5Z 4E3
c Department of Surgery, VancouÕer Hospital and Health Science Center, UniÕersity of British Columbia, 700 W10th AÕenue, VancouÕer, BritishColumbia, Canada V5Z 4E5
d Department of Zoology, VancouÕer Hospital and Health Science Center, UniÕersity of British Columbia, 6270 UniÕersity BouleÕard, VancouÕer, BritishColumbia, Canada V6T 1Z4
e Department of Medicine, VancouÕer Hospital and Health Science Center, UniÕersity of British Columbia, 2211 Westbrook mall, VancouÕer, BritishColumbia, Canada V6T 2B5
Accepted 26 January 1999
Abstract
Ž . Ž .Axonal loss and degeneration in multiple sclerosis MS and experimental allergic encephalomyelitis EAE have been suggested bybrain imaging, pathological and axonal transport studies. Further elucidation of the processes and mechanisms of axonal degeneration indemyelinating diseases is therefore of potential importance in order to alleviate the permanent disabilities of MS patients. However,detailed studies in this area are impeded by the small number of reliable models in which the onset and location of demyelination can be
Ž .well-controlled. In this study, microinjection of polyclonal rabbit anti-galactocerebroside anti-Gal C antibody and guinea pigcomplement was used to induce local demyelination in the rat optic nerve. We found that treatment with appropriate volumes of theantibody and complement could induce local demyelination with minimal pressure- or trauma-induced damage. Local changes in
Ž . Ž . Ž . Ž .neurofilaments NFs and microtubules MTs were examined with both immunohistochemistry IHC and electron microscopy EM . Onday 1 after microinjection, we observed moderate NF and MT disassembly in the local demyelinated area, although in most cases, noapparent inflammatory cell infiltration was seen. The NF and MT changes became more apparent on days 3, 5, 7 after microinjection,along with gradually increased inflammatory cell infiltration. These results suggested that acute demyelination itself may induce localcytoskeleton changes in the demyelinated axons, and that the ensuing local inflammation may further enhance the axonal damage. Whenthe lesions were stained with specific antibodies for T lymphocytes, macrophages, and astrocytes, we found that most of the cells weremacrophages, suggesting that macrophages may play a greater role in inflammation-related axonal degeneration and axonal loss. Theseresults were confirmed and further characterized on the ultrastructural level. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Multiple sclerosis; Axonal degeneration; Neurofilament protein; Microtubule; Macrophage; Anti-galactocerebroside
1. Introduction
In contrast to the intensive immunopathological andŽ .neuroimaging studies in multiple sclerosis MS , the neu-
ronal or axonal changes in this disease are only occasion-ally reported and only recently have received major atten-tion. The available findings can generally be categorizedinto four general areas. Firstly, proton magnetic resonance
) Corresponding author. Fax: q1-604-875-4663; E-mail:[email protected]
spectroscopy shows a significant reduction in the concen-Ž .tration of N-acetylaspartate NAA in the chronically le-
sioned areas and the degree of NAA reduction correlatesw xwith the disease progression 1,12,13,43 . Since NAA is
only found in neurons, its concentration reduction isthought to represent neuronal or axonal damage or loss inMS. Secondly, EM examinations have found that the
w xdemyelinated axons in MS patients become thinner 11,50 ,and the axonal density is decreased in the demyelinated
w xareas with an expanded extracellular space 2 . Degenerat-ing swollen axons may also be seen in the active demyeli-
w xnated areas 57 . Axonal loss has been estimated to range
0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 99 01191-9

( )B. Zhu et al.rBrain Research 824 1999 204–217 205
from 10–20% in early MS, and up to 80% in the destruc-w xtive lesions of acute MS 36 . Axonal transection may be a
w xconsistent feature of the MS lesions 69 . The brain’s grossappearance in MS shows secondary tract degeneration and
w xbrain atrophy with the dilation of cerebral ventricles 36 .Thirdly, immunostaining of amyloid precursor protein, asensitive marker of axonal damage, showed its expression
w xin acute MS lesions 20 . Finally, ophthalmoscopic exami-nation has suggested that there are subtle signs of retinal
w xnerve fiber atrophy 23 , after even a single attack of opticw xneuritis 41 . Although these findings are important and
suggestive, the great variabilities in lesion location, sever-ity, activity, duration, and times of relapses have madefurther research somewhat difficult.
In the animal models of MS, axonal degeneration hasalso been reported to be correlated with both the severityof symptoms in acute experimental allergic en-
Ž . w xcephalomyelitis EAE 49 and with the disease durationw xand times of relapses in chronic EAE 3 . Findings have
included reduced axonal density and axonal diameters,massive axonal swellings or spheroids, lucent axoplasm,
Ž .disarrayed neurofilaments NFs and relatively fewer mi-Ž . w xcrotubules MTs 27,28,49,52,53,66 . Axonal transport
studies in the affected tracts show an increase in fastaxonal transport and varied changes in slow axonal trans-
w xport 34,48,54,67,72,73 , which also suggest axonal cyto-skeleton changes in demyelinated axons.
It thus appears that axonal degeneration, loss and dys-function occur in the demyelinated areas of both MS andanimal models. There is also evidence suggesting thataxonal cytoskeleton changes may play a major role in theaxonal pathology. However, important questions have notyet been answered, these include: How soon do the axonalcytoskeletonal changes occur after demyelination, and howdo these changes evolve? How extensive are the changesfollowing acute demyelination? Are they distributed fromthe lesion distally to the axonal terminals as in Walleriandegeneration, or are they local changes? What are thepossible mechanisms which induce the changes? Sinceneuronal andror axonal changes are thought to be oneexplanation for the permanent disabilities in MS patients,the answers to the above questions should provide insightsinto the possible strategies for both prevention and treat-ment of the symptoms due to neuronal changes in MS.
In this study, we have used microinjection of polyclonalŽ .rabbit anti-galactocerebroside anti-Gal C antibody and
guinea pig complement into the rat optic nerves to inducelocal demyelination. This demyelination model has severaladvantages. The time and location of the occurrence ofdemyelination are easily controlled. The axons of rat opticnerves comprise a well-defined pathway with a singledirection, and the retinal ganglion cells, as well as thepostsynaptic neurons, i.e., lateral geniculate nuclei areavailable for further studies. To minimize injury inducedby microinjection, we have tested various injection vol-umes as well as different surgical access methods.
We have examined the extent and time course of axonalNF and MT changes in the acute phase of demyelination in
Ž .this model using both immunohistochemistry IHC andŽ .electron microscopy EM . We found that the axonal NF
and MT changes occurred only locally in the acutelydemyelinated area, and in most instances, began beforeapparent infiltration of inflammatory cells. These changeswere, however, greatly enhanced as local inflammationdeveloped. A moderate local increase in microglia wasobserved on day 1 after microinjection, while later, theintensive cell infiltration consisted mainly of macrophages.The results suggest two distinct mechanisms underlyingaxonal damage following demyelination, one being inflam-mation-independent, and the other resulting from inflam-matory response, in which macrophages may play a signif-icant role.
2. Material and methods
2.1. Antibodies and reagents
Ž .Polyclonal anti-Gal C antibody 1 mgrml was raisedin rabbits, and partially purified to a predominance of the
w xIgG subclass as described in a recent paper 17 . GuineaŽpig complement was obtained from Gibco Grand Island,
. Ž .NY . Luxol fast blue LFB was obtained from BDHŽ .laboratory supplies Poole, England . Cresyl violet acetate
Ž .CV , monoclonal mouse anti-NF 68 antibody, monoclonalmouse anti-a-tubulin antibody, purified rabbit IgG, normal
Ž .goat serum, anti-mouse IgG whole molecule biotin con-X Ž .jugate, 3,3 -diaminobenzidine DAB were obtained from
Ž .Sigma St. Louis, MO . Monoclonal mouse anti-myelinŽ .basic protein MBP antibody was obtained from
Ž .Boehringer Mannheim Laval, Quebec . Monoclonal mouseŽ .anti-rat macrophages ED2 , and monoclonal mouse anti-rat
Ž .CD3 were from Serotec Oxford, England . PolyclonalŽ .rabbit anti-cow glial fibrillary acidic protein GFAP was
Ž .from DAKO Glostrup, Denmark . The ABC staining kitand biotinylated anti-mouse IgG, anti-mouse IgM, anti-rabbit IgG were obtained from Vector LaboratoriesŽ .Burlingame, CA . All other reagents were of analyticalgrade or better and obtained from commercial sources.
2.2. Animals
Ž .A total of 59 adult male Wistar rats 300–400 g wereobtained from the Animal Care Center, University ofBritish Columbia. Twenty rats divided into four groupswere used to determine the proper microinjection volume.The 10 optic nerves of each five rats were injected with
Ž3.0 ml, 1.2 ml, 0.8 ml or 0.4 ml normal rabbit IgG 1. Ž . Ž .mgrml 60% and guinea pig complement 40% , respec-
tively. After finding a 0.4 ml injection volume to beoptimal, an additional five rats were microinjected with 0.4
Ž . Ž .ml normal rabbit IgG 1 mgrml 60% and guinea pig

( )B. Zhu et al.rBrain Research 824 1999 204–217206
Ž .complement 40% to form a control totalling 20 opticnerves. An additional 22 rats were used to examine thetime course of demyelination, axonal cytoskeletal changes,and the types of inflammatory cells using IHC. Each opticnerve was microinjected with 0.4 ml anti-Gal C antibodyŽ . Ž .60% and complement 40% , and 4, 4, 4, and 10 ratswere sacrificed on post-injection days 1, 3, 5, 7, respec-tively. The remaining 12 rats were used in an EM study.For this, four optic nerves were obtained in each of thefollowing six groups: non-injected; 7 days after normalrabbit IgG and complement microinjection; 1, 3, 5, 7 daysafter anti-Gal C antibody and complement microinjection.
2.3. Surgery and microinjection
Ž .The rats were anesthetized with ketamine 100 mgrkgŽ .and xylazine 10 mgrkg by i.p. injection. The hair cover-
ing the skull was shaved. After the rats were placed in thestereotaxic frame, a midline incision about 3 cm long wasmade along the scalp. Layers of fasciae, fat tissue, and thesuperior rectus were dissected along the margo supraor-bitalis until the optic nerves were exposed. The dura andarachnoid matter around the optic nerves were then incisedlongitudinally. Glass micropipettes, with the tip externaldiameters of about 24 mm were filled with the requiredvolumes of antibody and complement. The pipettes wereinserted into the rat optic nerve as superficial as possible atabout 2 mm behind the optic disc, and the fluid was slowlypressure-injected into the nerve over about 30 s. Each opticnerve was injected only once.
2.4. Tissue preparation and histological examination
ŽThe rats were sacrificed with 1.0 ml Euthanyl MTC.Pharmaceutical by i.p. injection, and were perfused tran-
scardially with about 50 ml 0.1 M phosphate bufferedŽ . Ž .saline PBS containing 0.9% NaCl pH7.4 . The opticŽ .nerves from the disc to optic chiasm were immediately
removed and frozen with 2-methylbutane at y808C for 10min. The tissues were stored in a y808C freezer forseveral hours until sections were prepared. A total of 45 8mm thick serial sections were cut longitudinally along eachoptic nerve on a Reichert-Jung 2800 Frigocut cryostat. Theslices were stored in a y208C freezer for several daysbefore staining.
The 1st, 16th, 31st sections were used to screen fordemyelination with LFB staining with CV counterstaining.The sections were first stained in LFB solution at 608C for3 h. Then, they were washed in distilled water. Differentia-tion was performed with rapid immersion in 0.05% aque-ous lithium carbonate and several changes of 70% alcohol.After distilled water washes, the sections were counter-stained with 1% CV for 1 min. They were washed indistilled water again, and dehydrated in a graded series ofalcohols and then cleaned in xylene. Finally, the sectionswere coverslipped. If demyelination was found on a given
section, anti-MBP, anti-NF and anti-a-tubulin IHC wascarried out separately on the next three serial sections.
2.5. IHC
The slides were first air-dried for 30 min, and then wereimmersed in acetone for 10 min. After three 10-min washesin 0.1 M PBS, the sections were incubated with theprimary antibodies with 5% normal goat serum at 48Covernight. The monoclonal anti-MBP, NF, and a-tubulinantibodies were diluted to 1:400, 1:400, and 1:1000 inPBS, respectively. The monoclonal macrophage, CD3 andGFAP antibodies were diluted to 1:100 in PBS. After threewashes in PBS on the next day, the sections were incu-
Ž .bated with the appropriate secondary antibody 1:1000 for1 h. They were then washed in PBS three times again, andthen incubated with avidin–biotin–peroxidase complexŽ .ABC, avidin and biotin were at 1:1000 for 45 min. Aftertwo washes in PBS and one wash in Tris-buffered solutionŽ .TBS , the sections were reacted for peroxidase enzyme
Ž .activity using filtered DAB 0.3 mgrml in TBS andŽ .H O 0.03% for 5–10 min. The slides were then washed2 2
in distilled water, gradually dehydrated in a graded seriesof alcohols, cleared in xylenes, and coverslipped.
2.6. EM
Each microinjected rat was sacrificed on day 7 with 1.0ml Euthanyl by i.p. injection, and perfused transcardiallywith about 100 ml Karnovsky’s fixation solution contain-ing 2.5% glutaldehyde. The proximal parts of the opticnerves were cut into four to five segments, each with alength of about 1 mm. They were postfixed with 2%osmium tetroxide for 1 h, dehydrated with increasingconcentrations of ethanol, cleared in propylene oxide, andembedded in Epon. Cross-sections 1 mm thick were pre-pared on a LKB ultratome, and stained with 1% Toluidineblue in 1% Borax for 30 s on a hot plate. If demyelinationwas seen on these sections, 90 nm thick sections wereprepared on a Reichert-Jung Ultracut E microtome, andexamined with a Zeiss 10 electron microscope.
3. Results
3.1. Defining the proper injection Õolume
First, we compared the histological changes with LFBand CV staining 7 days after injection of 0.4 ml to 3 ml
Ž . Ž .normal rabbit IgG 60% and complement 40% . With the3.0 ml injection volume, we invariably found by lightmicroscope small cavitation in the proximal part of theoptic nerve, which signified local damage. When the injec-tion volume was decreased to 1.2 ml, there were stilloccasional cavity-like damages. When the volume of 0.8ml was injected, no cavity-like changes were seen. How-ever, with NF and a-tubulin IHC, the areas of local NF
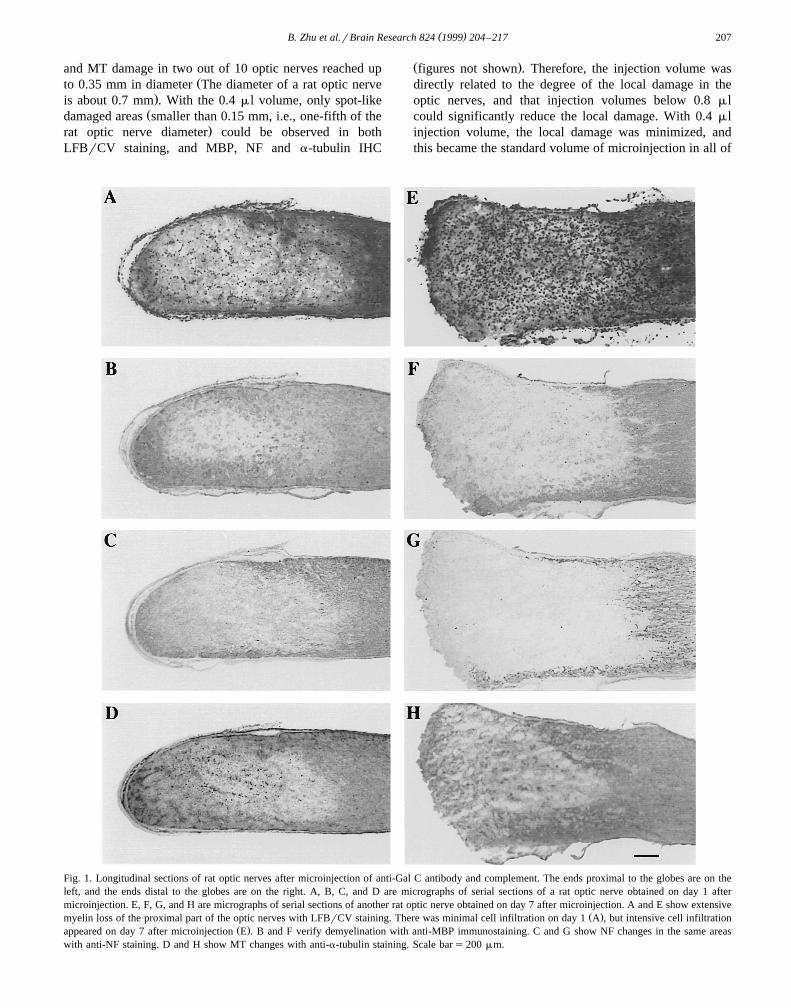
( )B. Zhu et al.rBrain Research 824 1999 204–217 207
and MT damage in two out of 10 optic nerves reached upŽto 0.35 mm in diameter The diameter of a rat optic nerve
.is about 0.7 mm . With the 0.4 ml volume, only spot-likeŽdamaged areas smaller than 0.15 mm, i.e., one-fifth of the
.rat optic nerve diameter could be observed in bothLFBrCV staining, and MBP, NF and a-tubulin IHC
Ž .figures not shown . Therefore, the injection volume wasdirectly related to the degree of the local damage in theoptic nerves, and that injection volumes below 0.8 mlcould significantly reduce the local damage. With 0.4 mlinjection volume, the local damage was minimized, andthis became the standard volume of microinjection in all of
Fig. 1. Longitudinal sections of rat optic nerves after microinjection of anti-Gal C antibody and complement. The ends proximal to the globes are on theleft, and the ends distal to the globes are on the right. A, B, C, and D are micrographs of serial sections of a rat optic nerve obtained on day 1 aftermicroinjection. E, F, G, and H are micrographs of serial sections of another rat optic nerve obtained on day 7 after microinjection. A and E show extensive
Ž .myelin loss of the proximal part of the optic nerves with LFBrCV staining. There was minimal cell infiltration on day 1 A , but intensive cell infiltrationŽ .appeared on day 7 after microinjection E . B and F verify demyelination with anti-MBP immunostaining. C and G show NF changes in the same areas
with anti-NF staining. D and H show MT changes with anti-a-tubulin staining. Scale bars200 mm.

( )B. Zhu et al.rBrain Research 824 1999 204–217208
the following experiments. The 20 rat optic nerves whichwere injected with 0.4 ml of normal rabbit IgG andcomplement and displayed very local myelin, NF and MTdisruption became the control group for the followingdemyelination studies. We also examined four non-injectedrat optic nerves and four rat optic nerves obtained 7 daysafter microinjection of normal rabbit IgG and guinea pigcomplement by EM. Almost all of the axons in the normalrat optic nerves were myelinated. No apparent demyelina-tion or axonal cytoskeleton changes were found in thecontrol injection nerves, and only a few macrophages and
Žmicroglia were found in very localized areas figures not.shown .
3.2. Time course study of demyelination induction
After defining the proper injection volume, we injected20 rat optic nerves with 0.4 ml polyclonal rabbit anti-Gal
Ž . Ž .C antibody 60% and guinea pig complement 40% .Histological and IHC examinations were carried out ontissue sections obtained 7 days after microinjection. WithLFBrCV staining, optic nerves showed various degrees of
Ž .demyelination and very marked cell infiltration Fig. 1ETo minimize the effect of direct local injury caused by themicroinjection, we selected seven optic nerves with exten-sive demyelination areas and four with large demyelination
Ž .areas as defined in Table 1 as qqq and qq . ‘Large’demyelination areas involved the entire or almost entirewidth of the nerve. ‘Extensive’ demyelination areas weredefined as large demyelination areas extending longitudi-nally over 1 mm or more. We verified the demyelination inserial sections of these 11 optic nerves using IHC with
Ž .monoclonal anti-MBP antibody Fig. 1F . Using the samemethods, 5r8, 3r8, 4r8 of rat optic nerves, which were
Ž .obtained on day 1 Fig. 1A and B , day 3, and day 5 aftermicroinjection, respectively were found to have large orextensive demyelination areas. Despite our best efforts tostabilize and define optimal experimental conditions, thedegree of demyelination induced by the same treatmentwas quite variable. As Table 1 shows, only about half ofthe injected nerves showed a substantial zone of demyeli-nation which transversed or nearly transversed the full-
width of the optic nerve. Only those animals which showedeither ‘large’ or ‘extensive’ demyelination were used forobservations of cytoskeletal or inflammatory alterations.
We also examined 16 optic nerves by EM over the fourtime points, i.e., days 1, 3, 5, and 7 after anti-Gal C andcomplement microinjection. 3r4, 2r4, 2r4, and 3r4 ofthe injected optic nerves obtained on days 1, 3, 5, 7,respectively were found to have large areas of demyelina-tion with toluidine blue staining. The demyelinated areasof those optic nerves were used in the EM studies. On day
Ž1, the myelin was greatly disrupted in the lesion areas Fig..8A . Especially notable was vesicular breakdown of
myelin. On days 3, 5, 7, more completely demyelinatedŽ .axons could be seen Fig. 8B, Fig. 9 . Although the
vesicular disruption of myelin could still be seen on day 3,it was rare on days 5 or 7. The myelin debris was largelycleared by day 7 after microinjection. From day 5, therewas evidence of gliosis. On day 7, axons frequently showedthin myelin sheaths, probably representing remyelination,
Žbut partial demyelination could not be excluded Fig. 2B,.Fig. 8B, Fig. 9 . Wallerian degeneration of axons was only
occasionally observed.
3.3. Time course study for axonal NF and MT changes
We performed IHC with monoclonal anti-NF and anti-a-tubulin antibodies on sections of selected demyelinatedoptic nerves, and found a well-defined time-course ofchanges in NFs and MTs. On day 1 after microinjection,the NF immunostaining showed evenly reduced reactivity
Ž .within the demyelinated area Fig. 1C . By 3 and 5 daysafter injection, the NF immunostaining largely appeared assparse NF reactivity, with occasionally ovoid staining
Žwhich represent axonal swelling at the distal margin away.from the globe of the demyelinated zone of the nerve
Ž .figures not shown . In 7 days after injection, NF reactivityŽ .almost disappeared in the extensive qqq demyeli-
nated lesions with numerous axonal swellings appearing atŽ .the distal margin of the demyelinated areas Fig. 1G . In
Ž .large qq demyelinated lesions, 3r4 showed sparse NFimmunoreactivity by day 7. The MT immunostainingshowed decreased reactivity within the extensively de-
Table 1Time course studies of the demyelination and the MT, NF changes after microinjection of anti-Gal C antibody and complement
Days after No. of injected No. of nerves with No. of nerves with No. of nerves with No. of nervesa b cinjection optic nerves demyelination cell infiltration NF changes with
MT changesqq qqq " q qq q qq qqq
1 8 3 2 5 0 0 5 0 0 53 8 2 1 0 2 1 0 3 0 35 8 3 1 0 1 3 0 2 2 47 20 4 7 0 0 11 0 3 8 11
a Ž . Ž .Degree of demyelination: qq large, crossing the width of optic nerves; qqq extensive, large demyelination areas longitudinally exceeding 1 mm.b Ž . Ž . Ž .Degree of cell infiltration: " minimal; q some; qq numerous.c Ž . Ž . Ž .NF staining pattern changes: q decreased reactivity; qq sparse, with a few axonal spheroids at the distal margin; qqq almost disappeared inthe central demyelinated areas, with numerous axonal spheroids at the distal margin.
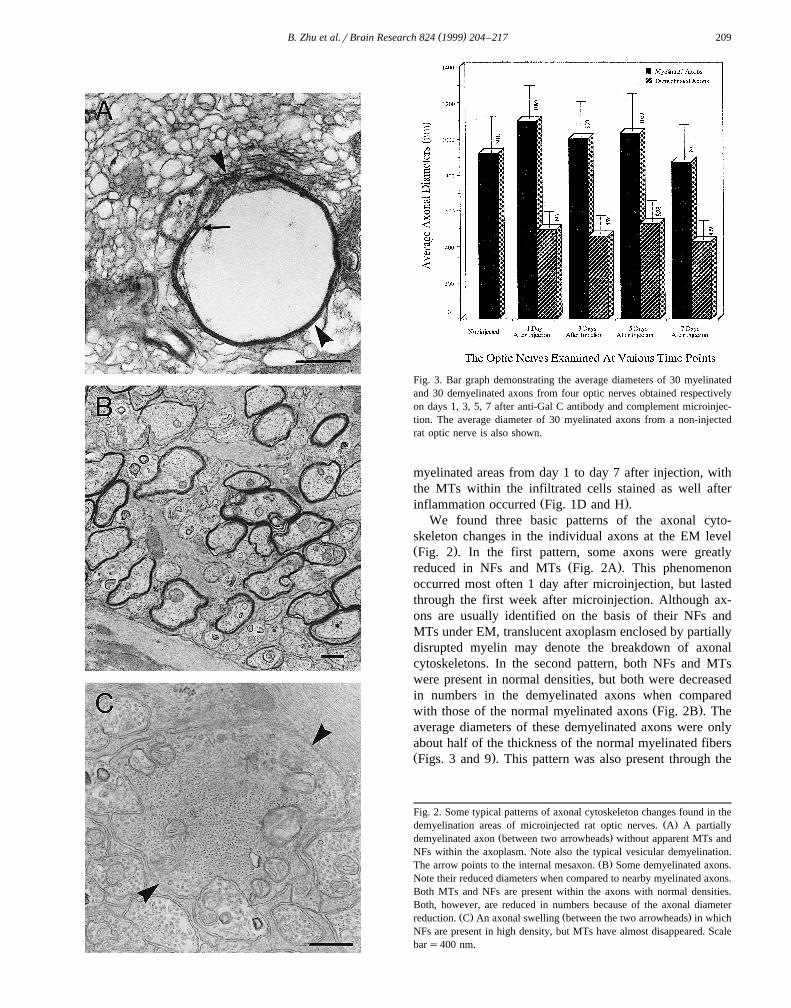
( )B. Zhu et al.rBrain Research 824 1999 204–217 209
Fig. 3. Bar graph demonstrating the average diameters of 30 myelinatedand 30 demyelinated axons from four optic nerves obtained respectivelyon days 1, 3, 5, 7 after anti-Gal C antibody and complement microinjec-tion. The average diameter of 30 myelinated axons from a non-injectedrat optic nerve is also shown.
myelinated areas from day 1 to day 7 after injection, withthe MTs within the infiltrated cells stained as well after
Ž .inflammation occurred Fig. 1D and H .We found three basic patterns of the axonal cyto-
skeleton changes in the individual axons at the EM levelŽ .Fig. 2 . In the first pattern, some axons were greatly
Ž .reduced in NFs and MTs Fig. 2A . This phenomenonoccurred most often 1 day after microinjection, but lastedthrough the first week after microinjection. Although ax-ons are usually identified on the basis of their NFs andMTs under EM, translucent axoplasm enclosed by partiallydisrupted myelin may denote the breakdown of axonalcytoskeletons. In the second pattern, both NFs and MTswere present in normal densities, but both were decreasedin numbers in the demyelinated axons when compared
Ž .with those of the normal myelinated axons Fig. 2B . Theaverage diameters of these demyelinated axons were onlyabout half of the thickness of the normal myelinated fibersŽ .Figs. 3 and 9 . This pattern was also present through the
Fig. 2. Some typical patterns of axonal cytoskeleton changes found in theŽ .demyelination areas of microinjected rat optic nerves. A A partially
Ž .demyelinated axon between two arrowheads without apparent MTs andNFs within the axoplasm. Note also the typical vesicular demyelination.
Ž .The arrow points to the internal mesaxon. B Some demyelinated axons.Note their reduced diameters when compared to nearby myelinated axons.Both MTs and NFs are present within the axons with normal densities.Both, however, are reduced in numbers because of the axonal diameter
Ž . Ž .reduction. C An axonal swelling between the two arrowheads in whichNFs are present in high density, but MTs have almost disappeared. Scalebars400 nm.
( )B. Zhu et al.rBrain Research 824 1999 204–217210
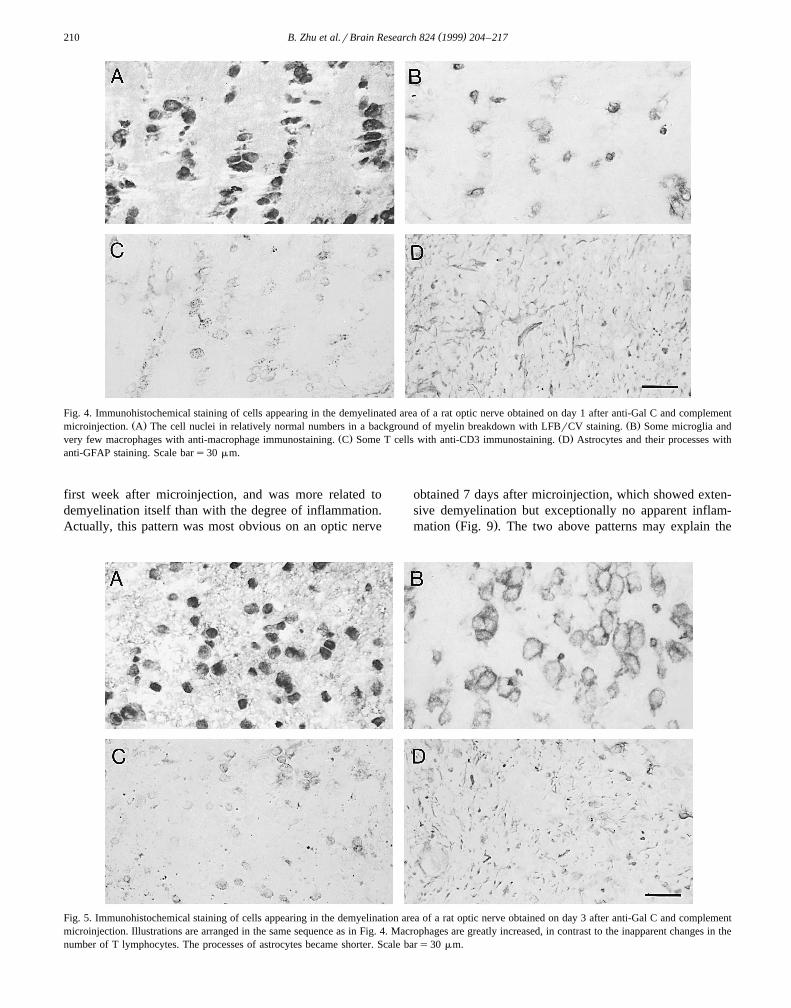
Fig. 4. Immunohistochemical staining of cells appearing in the demyelinated area of a rat optic nerve obtained on day 1 after anti-Gal C and complementŽ . Ž .microinjection. A The cell nuclei in relatively normal numbers in a background of myelin breakdown with LFBrCV staining. B Some microglia and
Ž . Ž .very few macrophages with anti-macrophage immunostaining. C Some T cells with anti-CD3 immunostaining. D Astrocytes and their processes withanti-GFAP staining. Scale bars30 mm.
first week after microinjection, and was more related todemyelination itself than with the degree of inflammation.Actually, this pattern was most obvious on an optic nerve
obtained 7 days after microinjection, which showed exten-sive demyelination but exceptionally no apparent inflam-
Ž .mation Fig. 9 . The two above patterns may explain the
Fig. 5. Immunohistochemical staining of cells appearing in the demyelination area of a rat optic nerve obtained on day 3 after anti-Gal C and complementmicroinjection. Illustrations are arranged in the same sequence as in Fig. 4. Macrophages are greatly increased, in contrast to the inapparent changes in thenumber of T lymphocytes. The processes of astrocytes became shorter. Scale bars30 mm.
( )B. Zhu et al.rBrain Research 824 1999 204–217 211
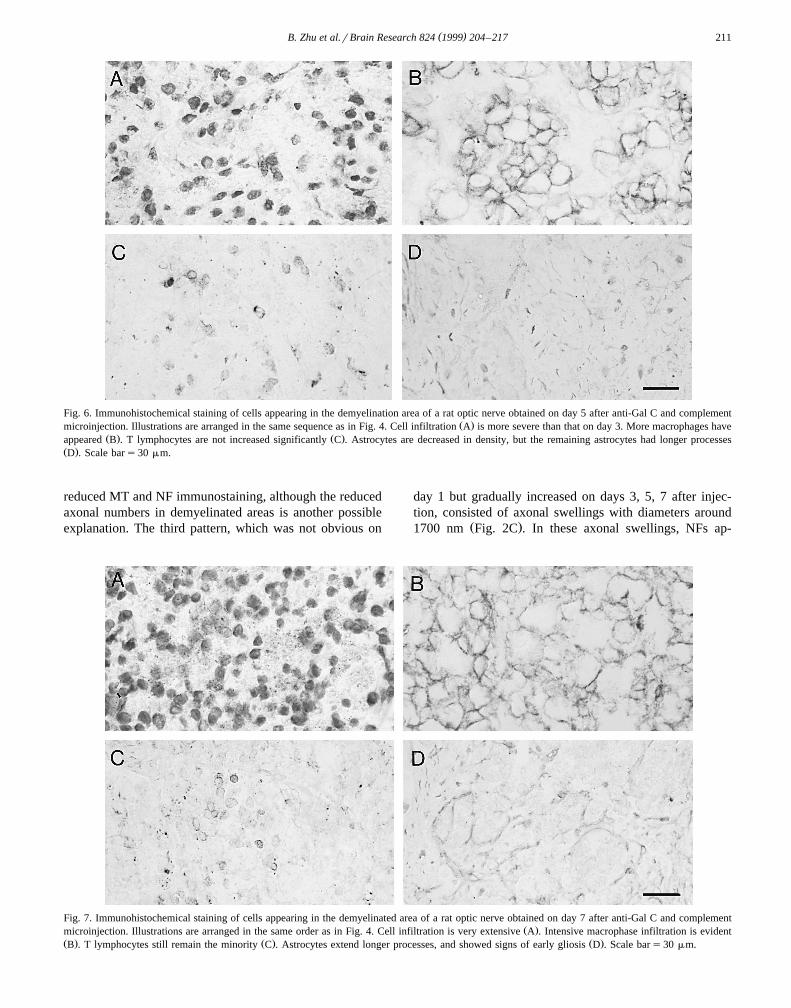
Fig. 6. Immunohistochemical staining of cells appearing in the demyelination area of a rat optic nerve obtained on day 5 after anti-Gal C and complementŽ .microinjection. Illustrations are arranged in the same sequence as in Fig. 4. Cell infiltration A is more severe than that on day 3. More macrophages have
Ž . Ž .appeared B . T lymphocytes are not increased significantly C . Astrocytes are decreased in density, but the remaining astrocytes had longer processesŽ .D . Scale bars30 mm.
reduced MT and NF immunostaining, although the reducedaxonal numbers in demyelinated areas is another possibleexplanation. The third pattern, which was not obvious on
day 1 but gradually increased on days 3, 5, 7 after injec-tion, consisted of axonal swellings with diameters around
Ž .1700 nm Fig. 2C . In these axonal swellings, NFs ap-
Fig. 7. Immunohistochemical staining of cells appearing in the demyelinated area of a rat optic nerve obtained on day 7 after anti-Gal C and complementŽ .microinjection. Illustrations are arranged in the same order as in Fig. 4. Cell infiltration is very extensive A . Intensive macrophase infiltration is evident
Ž . Ž . Ž .B . T lymphocytes still remain the minority C . Astrocytes extend longer processes, and showed signs of early gliosis D . Scale bars30 mm.

( )B. Zhu et al.rBrain Research 824 1999 204–217212
peared in high density with their sidearms outstretched andconnected with those of adjacent NFs, while MTs were
greatly reduced. This change was consistent with day 7Ž .tissue NF and MT immunostaining Fig. 1G and H in
Ž .Fig. 8. Electron micrographs showing the cell infiltration and demyelination on day 1 and day 7 after anti-Gal C and complement microinjection. AŽ . Ž .Extensive vesicular demyelination arrow and a polymorphonuclear cell large arrowhead in a day 1 demyelinated area. Some demyelinated and
Ž . Ž . Ž .demyelinating axons small arrowheads exhibit decreased diameters. B An astrocyte arrowhead , a cell with prominent rough endoplasmic reticulumŽ . Ž .which may be a plasma cell star , and some remyelinated or partially demyelinated axons with very thin myelin arrows in a day 7 demyelinated area.
Scale bars1500 nm.

( )B. Zhu et al.rBrain Research 824 1999 204–217 213
which we found some strong axonal swelling NF staining,but little MT staining.
3.4. Time course analysis of the infiltrated cells in thedemyelinated areas
On day 1 after injection, there was no apparent cellŽinfiltration in most injected optic nerves Fig. 1A, Fig.
.4A . On days 3 and 5 after injection, cell infiltrationŽ .became apparent Fig. 5A, Fig. 6A . It was still less than
Ž .that observed on day 7 after injection Fig. 1E, Fig. 7A .To determine which cell type was associated temporallywith the axonal cytoskeleton changes, we used ED2, CD3Ž . Ža specific T cell surface antigen , and GFAP a specific
.marker for astrocytes antibodies to analyze the cell popu-lation by IHC.
On day 1 after microinjection, the anti-rat macrophageantibody stained sparsely distributed microglia and even
Ž .fewer macrophages Fig. 4B . The number of microgliaseemed to be mildly increased when compared with nor-mal optic nerves. Also, there were a few CD3 positive T
Ž .cells Fig. 4C . Astrocytes and their processes were seen inan almost normal pattern and in normal density on day 1
Ž .Fig. 4D . On days 3, 5, and 7 after microinjection,gradually increased numbers of macrophages were ob-
Žserved in the demyelinated areas Fig. 5B, Fig. 6B, Fig..7B and accounted for about 50–80% of all infiltrated
cells. T cells were only moderately increased in this periodŽ .Fig. 5C, Fig. 6C, Fig. 7C . Astrocytes were decreased in
Ždensity and their processes were shortened on day 3 Fig..5D , but on days 5 and 7, astrocytes began to extend
Ž .longer processes Fig. 6D, Fig. 7D . This change maysuggest that some astrocytes were also injured and mayhave succumbed in the acute stage of demyelination andinflammation, but the surviving astrocytes soon becamereactive and extended longer processes. In the EM study,we did find some polymorphonuclear cells appearing in
Ž .the demyelinating areas on day 1 Fig. 8A , which mightbe related to higher complement activity of the new batchof complement used for that particular experiment. Fromday 3 to day 7, most of the inflammatory cells were
Ž .macrophages Fig. 8B . On day 5, there was evidence ofŽearly gliosis. On day 7, gliosis was more apparent Fig.
.8B, Fig. 9 .The above results suggest that macrophages were the
major cell type of infiltrating cells in the demyelination
Fig. 9. An electron micrograph to illustrate demyelinated axons in a day 7 demyelinated optic nerve. Note that there are no inflammatory cells in this fieldŽ .which is an exception , and both MTs and NFs were present in the axons. The average demyelinated axonal diameter is only about 400 nm, compared to
Ž .about 900 nm in diameter of myelinated axons in the same nerve. Mild gliosis has begun arrow . Scale bars1000 nm.

( )B. Zhu et al.rBrain Research 824 1999 204–217214
areas, and might therefore play a more important role ininflammation-dependent axonal cytoskeleton changes thanother cells.
4. Discussion
4.1. Experimental demyelination in optic nerÕes
The use of microinjection to produce demyelination inthe optic nerve was first reported by Caroll et al. in the ratw x w x5,6 and by Sergott et al. in the guinea pig 60 . Thetechnique has not been widely used since then. Since thismodel has several advantages over other demyelinationmodels, we explored different surgical access methods andvaried injection volumes. We found that dissection alongthe roof of the orbits did not result in retinal ischemia asoccurred with other dissection methods.
Another major concern in using this model to examineaxonal and neuronal changes is the potential of directinjury to the optic nerves by insertion of pipettes and byinjection-induced local pressure. The adult rat optic nerveis about 0.7–0.8 mm in diameter. We found that if the tipsof the glass pipettes were finer than 30 mm, the insertionof the tips of the pipettes into the nerves did not produce adetectable structural change. However, the volume injectedis directly related to the local pressure within the nerves.With the pia mater around the nerves left intact, we testedvarious injection volumes from 0.4 ml to 3 ml, and foundthat a 0.4 ml injection volume reduced the damage zone toless than 0.15 mm in diameter. Thus, we believe that theaxonal cytoskeleton changes which were observed in thedemyelinated areas and which were much more extensivethan this surgically-induced damage zone were not due toinjection-induced injury.
With this approach, we induced extensive demyelina-tion in nerves, as revealed by LFB staining, anti-MBPIHC, and by EM studies. The demyelination zone couldextend 1–2 mm longitudinally and across the whole widthof the optic nerve. On day 1 after the injection, thevesicular breakdown of myelin was very apparent. Such
w xrapid effects were also seen by Sergott et al. 60 . Forstudies in the axonal cytoskeleton changes after demyelina-tion, this is the ideal ‘window’ during which the directeffects of myelin loss on the axons can be analyzed. Invarious EAE models, inflammation in the central nervoussystem always precedes the development of neurologicsigns. Therefore, it would be very difficult to observeinflammation-independent effects of demyelination on cen-tral nervous system axons using EAE models. This limita-tion also applies for the direct studies of nerve tissues fromMS patients. Although microinjection of ethidium bromideor cuprizone into the rat optic nerves may elicit a similar‘window’ period, the diffusion of ethidium bromide orcuprizone into the retina in which the retinal ganglion cellsreside may cause some unexpected neuronal and axonaldamage.
On days 3, 5, and 7 after injection, large areas ofdemyelination were easily be observed. Along with thedemyelination, we observed a gradual increase in cellinfiltration, especially of macrophages. Some T cells werealso present although their numbers were not increasedsignificantly within the first week. On day 7, some oligo-dendrocytes were present and thin sheaths of myelin wereevident. It is probable that this was early remyelination.
4.2. Axonal cytoskeleton changes in the demyelinationareas
It has been hypothesized that inflammation is the maincause of axonal damage in MS and EAE. However, it isnot clear whether demyelination itself would lead to localaxonal cytoskeleton changes. As mentioned above, wewere able to observe the axonal cytoskeleton changes inour optic neuritis model before apparent cell infiltration,i.e., on day 1 after microinjection. This offered the oppor-tunity to observe the direct effects of demyelination on theunderlying axonal cytoskeletons without the complicationsresulting from the arrival of immune-mediating cells. Wefound reduced staining for NFs and MTs on day 1 aftermicroinjection. Under EM, the majority of demyelinatedaxons presented only half of the thickness of myelinatedaxons. Some partially demyelinated axons showed translu-cent axoplasm without apparent NF or MT structures. Wepropose that the axonal cytoskeleton changes can be di-vided into two categories: one inflammation-independent,and the other inflammation-dependent.
Two possible mechanisms may be involved in the in-flammation-independent axonal cytoskeleton changes. Thefirst is the loss of axon–glial interactions. It has beenknown that during neural development, myelin formationinduces the full caliber expansion of mature axonsw x9,10,33,35 . The modulation of axonal caliber by myelinseems to be mediated by phosphorylation of particulardomains of NF proteins and by effects on other NF
w xdynamic behavior 15,45,65 . This process is evident in theperipheral nervous system in the Trembler mouse mutant;in the unmyelinated axons of which the caliber is signifi-cantly reduced, NFs are not phosphorylated and are present
w xin higher density than in wild-type axons 14,15 . Thestability of MTs is decreased and the composition andphosphorylation state of tau protein are also altered inthese axons. Normal sciatic nerve transplantation can exerta profound effect on the underlying neuronal cytoskeleton
w xof grafted Trembler mouse sciatic nerves 14 . In thecentral nervous system, similar effects have been observedin neurodevelopment and when mature oligodendrocytesare transplanted into the CNS of shiverer, jimpy and
w xquaking mutant mice 58 . Interestingly, even when theregions of node of Ranvier were compared with the intern-odal segments of normal axons, they were 40–60% lowerin their level of NF phosphorylation, and thinner in axonal
w xdiameter despite a higher density of NFs 15,32,40,56 .Studies on NF phosphorylation have shown that phospho-

( )B. Zhu et al.rBrain Research 824 1999 204–217 215
Ž .rylation on the KSP Lys–Ser–Pro domain of the tailregions of NF-H and NF-M by proline-directed protein
w xkinases will make NFs extend their sidearms 7,15,30 andw xbecome more stable 47 . Thus, the hypothesis that de-
myelination induces local dephosphorylation on KSP do-mains of NF-H and NF-M, and a consequent withdrawal ofNF sidearms, a decrease in NF stability, and eventually NFdisassembly needs to be tested. Because the extendedsidearms of NFs in the myelinated axons have been ob-
w xserved to slow axonal transport 29,31 , this mechanismcould also explain the consistent finding of increased fast
w xaxonal transport in EAE models 34,48,67 .A second possible mechanism of inflammation-indepen-
dent axonal cytoskeleton changes may involve increasedintra-axonal calcium concentrations. Intra-axonal calciumoverload could be achieved in a number of ways. Onepossibility is an increase in axonal membrane permeabilityafter demyelination. There is some evidence that axonal
w xand myelin cytoplasm are actually connected 16,24 . De-myelination may break this connection and thus affectaxonal membrane integrity, which would be expected tocause an influx of highly concentrated extracellular cal-cium directly into the demyelinated region of the axon.Another possibility is through voltage-dependent calciumchannels andror NaqrCa2q exchanger on the axonalmembrane. In the case of anoxia-induced axonal injury inrat optic nerves, increased calcium influx both throughL-type and N-type Ca2q channels and through NaqrCa2q
w xexchanger has been found to be involved 21,70 . Theintra-axonal calcium overload was responsible for the dis-
w xruption of MTs and NFs in these axons 71 .Within the axon, various protein kinases, phosphatases
and proteases are associated with NFs and MTs, regulatingthe assembly and disassembly of the axonal cytoskeleton.
Ž .Protein kinase C PKC and calciumrcalmodulin-depen-Ž .dent kinase CarCaM can phosphorylate the N-terminal
domain of NF proteins and lead to NF fragmentation andw xdisassembly 25,55,62,74 . PKC and CarCaM can also
phosphorylate tau proteins on Ser-262 and significantlyw xdecrease their stabilization effects on MTs 37,38,63 .
Ž .Phosphatase 2A and 2B calcineurin can be activated bycalcium, depending on the local calcium concentrationw x22,39 . The dephosphorylation on the key domains suchas KSP domain on NF-M and NF-H also leads to cyto-skeleton destabilization and its vulnerability to proteasesw x26,37 . Calcium can activate proteases such as calpains
w x w xwhich directly degrade tau proteins 75 and NFs 44,59 .Our results also showed a gradual increase in inflamma-
Ž .tory cells especially macrophages in the demyelinatedareas on days 3, 5 and 7 after microinjection. At the sametime, the axonal cytoskeleton changes were also moreapparent with immunostaining. On the ultrastructure level,more demyelinated axons with decreased diameters wereobserved. Axonal loss and axonal swellings were alsomore prominent. Some of axonal swellings may representthe occurrence of axonal transection caused by the inflam-
matory cells, especially macrophages. Although the possi-bility certainly exists that inflammation-independent mech-anisms continued to operate in this period, we have theimpression that demyelinated areas with more macrophagesand other inflammatory cells showed more significantaxonal cytoskeleton changes and axonal loss than demyeli-nated areas with fewer or no inflammatory cells. Theseresults are consistent with the recent findings suggestingthat the axonal transection in the lesions of MS waswell-correlated with the degree of inflammation, especially
w xmacrophage infiltration 69 .In the inflammation period, various free radicals includ-
ing reactive oxygen radicals and nitrite radicals which canbe secreted by macrophages may be deleterious to the
w xlocal axons 4 . Peroxynitrate has been shown to nitratetyrosine residues on NF-L and to derange NF phosphoryla-
w xtion and NF assembly 8 . Since free radicals can diffusethrough the axonal membrane, they may have extensiveeffects on axonal lipid and protein peroxidation, on DNAand RNA peroxidation in neuronal nuclei, and even causeneuronal death. The various proteolytic and lipolytic en-zymes and various cytokines secreted from granulocytes
w xand macrophages may also affect axonal integrity 42,68 .In future research, we think it will be of great value to
examine NF and MT phosphorylation on various domains,and to determine if the intra-axonal calcium load is in-creased in demyelinated axons and if calcium channelblockers can relieve the axonal damage. Further detailedstudies will be able to determine if there is upregulation ofcalcium channel expression in the demyelinated axons,how the activities of PKC, CarCaM, and various phos-phatases are changed in the demyelination zone, andwhether inflammatory inhibitors as IFN-b, TGF-b and freeradical scavengers can prevent or reduce the axonal cyto-skeleton changes and axonal damage. Moreover, the studyof how these axonal cytoskeleton changes evolve in thesubacute and chronic phases of demyelination in both ourmodel and in the EAE model is also needed.
Acknowledgements
This work was supported by Medical Research CouncilŽ .of Canada to M.S.C , and the Multiple Sclerosis SocietyŽ .of Canada to D.W.P . We also thank Esther Leung, Jeff
Hollingale, Caron Fournier, and Bill McLauley for techni-cal assistance.
References
w x1 D.L. Arnold, G.T. Riess, P.M. Matthews, G.S. Francis, D.L. Collins,C. Wolfson, J.P. Antel, Use of proton magnetic resonance spec-troscopy for monitoring disease progression in multiple sclerosis,
Ž .Ann. Neurol. 36 1994 76–82.w x2 D. Barnes, P.M. Munro, B.D. Youl, J.W. Prineas, W.I. McDonald,

( )B. Zhu et al.rBrain Research 824 1999 204–217216
The longstanding MS lesion. A quantitative MRI and electronŽ .microscopic study, Brain 114 1991 1271–1280.
w x3 A. Brown, D.E. McFarlin, C.S. Raine, Chronologic neuropathologyof relapsing experimental allergic encephalomyelitis in the mouse,
Ž .Lab. Invest. 46 1982 171–185.w x4 W. Bruck, The role of macrophages in Wallerian degeneration,
Ž .Brain Pathol. 7 1997 741–752.w x5 W.M. Carroll, A.R. Jennings, F.L. Mastaglia, Experimental optic
neuropathy—A model for combined morphological and electrophys-Ž .iological studies, in: J.H. Tyrer, M.J. Eadie Eds. , Clinical and
Experimental Neurology, ADIS Health Science Press, New York,1982, pp. 17–28.
w x6 W.M. Carroll, A.R. Jennings, F.L. Mastaglia, Experimental demyeli-nating optic neuropathy induced by intra-neural injection of galacto-
Ž .cerebroside antiserum, J. Neurol. Sci. 65 1984 125–135.w x7 S.S. Chin, R.K. Liem, Transfected rat high-molecular-weight neuro-
Ž .filament NF-H coassembles with vimentin in a predominantlyŽ .nonphosphorylated form, J. Neurosci. 10 1990 3714–3726.
w x8 S.M. Chou, H.S. Wang, K. Komai, Colocalization of NOS andSOD1 in neurofilament accumulation within motor neurons of amy-otrophic lateral sclerosis: an immunohistochemical study, J. Chem.
Ž .Neuroanat. 10 1996 249–258.w x9 J.S. Cole, A. Messing, J.Q. Trojanowski, V.M. Lee, Modulation of
axon diameter and neurofilaments by hypomyelinating SchwannŽ .cells in transgenic mice, J. Neurosci. 14 1994 6956–6966.
w x10 R.J. Colello, M.E. Schwab, A role for oligodendrocytes in theŽ .stabilization of optic axon numbers, J. Neurosci. 14 1994 6446–
6452.w x11 D. Dahl, G. Perides, A. Bignami, Axonal regeneration in old multi-
ple sclerosis plaques. Immunohistochemical study with monoclonalantibodies to phosphorylated and non-phosphorylated neurofilament
Ž .proteins, Acta Neuropathol. 79 1989 154–159.w x12 C.A. Davie, G.J. Barker, S. Webb, P.S. Tofts, A.J. Thompson, A.E.
Harding, W.I. McDonald, D.H. Miller, Persistent functional deficitin multiple sclerosis and autosomal dominant cerebellar ataxia is
Ž .associated with axon loss, Brain 118 1995 1583–1592.w x13 N. De Stefano, P.M. Matthews, S. Narayanan, G.S. Francis, J.P.
Antel, D.L. Arnold, Axonal dysfunction and disability in a relapse ofmultiple sclerosis: longitudinal study of a patient, Neurology 49Ž .1997 1138–1141.
w x14 S. deWaegh, S.T. Brady, Altered slow axonal transport and regener-ation in a myelin-deficient mutant mouse: the trembler as an in vivo
Ž .model for Schwann cell–axon interactions, J. Neurosci. 10 19901855–1865.
w x15 S. deWaegh, V.M. Lee, S.T. Brady, Local modulation of neurofila-ment phosphorylation, axonal caliber, and slow axonal transport by
Ž .myelinating Schwann cells, Cell 68 1992 451–463.w x16 A. Duncan, M. Ibrahim, M. Berry, A.M. Butt, Transfer of horseradish
peroxidase from oligodendrocyte to axon in the myelinating neonatalŽ .rat optic nerve: artefact or transcellular exchange?, Glia 17 1996
349–355.w x17 J.K. Dyer, J.A. Bourque, J.D. Steeves, Regeneration of brainstem–
spinal axons after immunological disruption of myelin in adult rat,Ž .Exp. Neurol. 154 1998 12–22.
w x20 B. Ferguson, M.K. Matyszak, M.M. Esiri, V.H. Perry, AxonalŽ .damage in acute multiple sclerosis lesions, Brain 120 1997 393–
399.w x21 R. Fern, B.R. Ransom, S.G. Waxman, Voltage-gated calcium chan-
nels in CNS white matter: role in anoxic injury, J. Neurophys. 74Ž .1995 369–377.
w x22 L.M. Fleming, G.V. Johnson, Modulation of the phosphorylationstate of tau in situ: the roles of calcium and cyclic AMP, Biochem. J.
Ž .309 1995 41–47.w x23 L. Frisen, W.F. Hoyt, Insidious atrophy of retinal nerve fibers in
multiple sclerosis. Funduscopic identification in patients with andŽ .without visual complaints, Arch. Ophthalmol. 92 1974 91–97.
w x24 D.D. Gaton, A. Solomon, E. Skutelsky, M. Wolman, Does a cyto-
plasmic bridge connect the CNS axon with the inner loop ofŽ .myelin?, Cell. Mol. Biol. 38 1992 597–599.
w x25 Y. Gonda, K. Nishizawa, S. Ando, S. Kitamura, Y. Minoura, Y.Nishi, M. Inagaki, Involvement of protein kinase C in the regulationof assembly–disassembly of neurofilaments in vitro, Biochem. Bio-
Ž .phys. Res. Commun. 167 1990 1316–1325.w x26 J.A. Greenwood, J.C. Troncoso, A.C. Costello, G.V. Johnson, Phos-
phorylation modulates calpain-mediated proteolysis and calmodulinbinding of the 200-kDa and 160-kDa neurofilament proteins, J.
Ž .Neurochem. 61 1993 191–199.w x27 J. Guy, E.A. Ellis, E. Tark, G.M. Hope, N.A. Rao, Axonal transport
reductions in acute experimental allergic encephalomyelitis: qualita-Ž .tive analysis of the optic nerve, Curr. Eye Res. 8 1989 261–269.
w x28 J. Guy, E.A. Ellis, G.M. Hope, S. Emerson, Maintenance of myeli-nated fibre g ratio in acute experimental allergic encephalomyelitis,
Ž .Brain 114 1991 281–289.w x29 N. Hirokawa, Molecular architecture and dynamics of neuronal
Ž .cytoskeketon, in: R.D. Borgoyen Ed. , The Neuronal Cytoskeleton,Wiley, New York, 1991, pp. 5–74.
w x30 S. Hisanaga, N. Hirokawa, The effects of dephosphorylation on theŽ .structure of the projections of neurofilament, J. Neurosci. 9 1989
959–966.w x31 P.N. Hoffman, R.J. Lasek, The slow component of axonal transport.
Identification of major structural polypeptides of the axon and theirŽ .generality among mammalian neurons, J. Cell Biol. 66 1975
351–366.w x32 S.T. Hsieh, G.J. Kidd, T.O. Crawford, Z. Xu, W.M. Lin, B.D.
Trapp, D.W. Cleveland, J.W. Griffin, Regional modulation of neuro-filament organization by myelination in normal axons, J. Neurosci.
Ž .14 1994 6392–6401.w x33 I. Hutchinson, Transplantation and rejection, in: I. Roitt, J. Brostoff,
Ž .D. Male Eds. , Immunology, Times Mirro International Publishers,Barcelona, 1996, pp. 26–27.
w x34 H.G. Jenkins, H. Ikeda, Tumour necrosis factor causes an increase inaxonal transport of protein and demyelination in the mouse optic
Ž .nerve, J. Neurol. Sci. 108 1992 99–104.w x35 L.L. Kirkpatrick, S.T. Brady, Modulation of the axonal microtubule
Ž .cytoskeleton by myelinating Schwann cells, J. Neurosci. 14 19947440–7450.
w x36 H. Lassmann, G. Suchanek, K. Ozawa, Histopathology and theblood–cerebrospinal fluid barrier in multiple sclerosis, Ann. Neurol.
Ž .36 1994 s42–6.w x37 J.M. Litersky, G.V. Johnson, Phosphorylation of tau in situ: inhibi-
Ž .tion of calcium-dependent proteolysis, J. Neurochem. 65 1995903–911.
w x38 J.M. Litersky, G.V. Johnson, R. Jakes, M. Goedert, M. Lee, P.Seubert, Tau protein is phosphorylated by cyclic AMP-dependentprotein kinase and calciumrcalmodulin-dependent protein kinase IIwithin its microtubule-binding domains at Ser-262 and Ser-356,
Ž .Biochem. J. 361 1996 655–660.w x39 E.M. Mandelkow, J. Biernat, G. Drewes, N. Gustke, B. Trinczek, E.
Mandelkow, Tau domains, phosphorylation, and interactions withŽ . w x wmicrotubules, Neurobiol. Aging 16 1995 355–362, Review 57
xRefs. .w x40 M. Mata, N. Kupina, D.J. Fink, Phosphorylation-dependent neurofil-
ament epitopes are reduced at the node of Ranvier, J. Neurocytol. 21Ž .1992 199–210.
w x41 W.I. McDonald, Rachelle Fishman–Matthew Moore Lecture. Thepathological and clinical dynamics of multiple sclerosis, J. Neu-
Ž . w x w xropath. Exp. Neur. 53 1994 338–343, Review 50 Refs. .w x42 J.E. Merrill, E.N. Benveniste, Cytokines in inflammatory brain
Ž .lesions: helpful and harmful, Trends Neurosci. 19 1996 331–338,w x w xReview 68 Refs. .
w x43 S. Narayanan, L. Fu, E. Pioro, S.N. De, D.L. Collins, G.S. Francis,J.P. Antel, P.M. Matthews, D.L. Arnold, Imaging of axonal damagein multiple sclerosis: spatial distribution of magnetic resonance
Ž .imaging lesions, Ann. Neurol. 41 1997 385–391.

( )B. Zhu et al.rBrain Research 824 1999 204–217 217
w x Ž .44 R.A. Nixon, Proteolysis of neurofilaments, in: C.A. Marotta Ed. ,Neurofilaments, University of Minnesota Press, Minneapolis, 1983,pp. 117–154.
w x45 R.A. Nixon, P.A. Paskevich, R.K. Sihag, C.Y. Thayer, Phosphoryla-tion on carboxyl terminus domains of neurofilament proteins inretinal ganglion cell neurons in vivo: influences on regional neurofil-ament accumulation, interneurofilament spacing, and axon caliber, J.
Ž .Cell Biol. 126 1994 1031–1046.w x47 H.C. Pant, Dephosphorylation of neurofilament proteins enhances
their susceptibility to degradation by calpain, Biochem. J. 256Ž .1988 665–668.
w x48 V.F. Pessoa, H. Ikeda, Increase in axonal transport in demyelinatingoptic nerve fibres in the mouse infected with Semliki Forest virus,
Ž .Brain 107 1984 433–446.w x49 H.C. Powell, R.R. Myers, A.P. Mizisin, T. Olee, S.W. Brostoff,
Response of the axon and barrier endothelium to experimentalallergic neuritis induced by autoreactive T cell lines, Acta Neu-
Ž .ropathol. 82 1991 364–377.w x50 J.W. Prineas, F. Connell, The fine structure of chronically active
Ž .multiple sclerosis plaques, Neurology 28 1978 68–75.w x52 C.S. Raine, A.H. Cross, Axonal dystrophy as a consequence of
Ž .long-term demyelination, Lab. Invest. 60 1989 714–725.w x53 N.A. Rao, Chronic experimental allergic optic neuritis, Invest. Oph-
Ž .thalmol. Vis. Sci. 20 1981 159–172.w x54 N.A. Rao, J. Guy, P.S. Sheffield, Effects of chronic demyelination
on axonal transport in experimental allergic optic neuritis, Invest.Ž .Ophthalmol. Vis. Sci. 21 1981 606–611.
w x55 K.E. Reagan, K.R. Wilmarth, M.A. Friedman, D.M. Abou, In vitrocalcium and calmodulin-dependent kinase-mediated phosphorylationof rat brain and spinal cord neurofilament proteins is increased by
Ž .glycidamide administration, Brain Res. 671 1995 12–20.w x56 A. Reles, R.L. Friede, Axonal cytoskeleton at the nodes of Ranvier,
Ž .J. Neurocytol. 20 1991 450–458.w x57 M. Rodriguez, B. Scheithauer, Ultrastructure of multiple sclerosis,
Ž .Ultrastruct. Pathol. 18 1994 3–13.w x58 I. Sanchez, L. Hassinger, P.A. Paskevich, H.D. Shine, R.A. Nixon,
Oligodendroglia regulate the regional expansion of axon caliber andlocal accumulation of neurofilaments during development indepen-
Ž .dently of myelin formation, J. Neurosci. 16 1996 5095–5105.w x59 W.W. Schlaepfer, U.-J.P. Zimmerman, The degradation of neurofila-
Ž .ments by calpains, in: R.L. Mellellgren, T. Murachi Eds. , Intracel-lular Calcium-dependent Proteolysis, 1990, pp. 241–249.
w x60 R.C. Sergott, M.J. Brown, D.H. Silberberg, R.P. Lisak, Antigalacto-
cerebroside serum demyelinates optic nerve in vivo, J. Neurol. Sci.Ž .64 1984 297–303.
w x62 R.K. Sihag, A.Y. Jeng, R.A. Nixon, Phosphorylation of neurofila-Ž .ment proteins by protein kinase C, FEBS Letters 233 1988 181–
185.w x63 T.J. Singh, J.Z. Wang, M. Novak, E. Kontzekova, I.I. Grundke, K.
Iqbal, Calciumrcalmodulin-dependent protein kinase II phosphory-lates tau at Ser-262 but only partially inhibits its binding to micro-
Ž .tubules, FEBS Letters 387 1996 145–148.w x65 R. Starr, B. Attema, G.H. DeVries, M.J. Monteiro, Neurofilament
phosphorylation is modulated by myelination, J. Neurosci. Res. 44Ž .1996 328–337.
w x66 T. Tabira, K. Sakai, Demyelination induced by T cell lines andclones specific for myelin basic protein in mice, Lab. Invest. 56Ž .1987 518–525.
w x67 E.M. Tansey, H. Ikeda, The relationship between axonal transport ofprotein and demyelination in the optic nerves of mice infected with
Ž .Semliki Forest virus, Brain Res. 397 1986 9–15.w x68 M. Tomita, Y. Fukuuchi, Leukocytes, macrophages and secondary
brain damage following cerebral ischemia, Acta Neurochir. Suppl.Ž . w x w x66 1996 32–39, Review 58 Refs. .
w x69 B.D. Trapp, J. Peterson, R.M. Ransohoff, R. Rudick, S. Mork, L.¨Bo, Axonal transection in the lesions of multiple sclerosis, N. Eng.¨
Ž .J. Med. 338 1998 278–285.w x70 S.G. Waxman, J.A. Black, B.R. Ransom, P.K. Stys, Protection of
the axonal cytoskeleton in anoxic optic nerve by decreased extracel-Ž .lular calcium, Brain Res. 614 1993 137–145.
w x71 S.G. Waxman, J.M. Ritchie, Molecular dissection of the myelinatedŽ .axon, Ann. Neurol. 33 1993 121–136.
w x72 S.R. White, R.M. Bowker, Retrograde transport of horseradishperoxidase is specifically impaired in bulbospinal serotonin axonsduring experimental allergic encephalomyelitis, J. Neuroimmunol.
Ž .18 1988 75–86.w x73 S.R. White, D. Vyas, D. Bieger, Serotonin immunoreactivity in
spinal cord axons and terminals of rodents with experimental aller-Ž .gic encephalomyelitis, Neurosci. 16 1985 701–709.
w x74 J.S. Whitson, A. Kampfl, X. Zhao, C.E. Dixon, R.L. Hayes, Briefpotassium depolarization decreases levels of neurofilament proteins
Ž .in CNS culture, Brain Res. 694 1995 213–222.w x75 L.S. Yang, R.H. Ksiezak, Calpain-induced proteolysis of normal
human tau and tau associated with paired helical filaments, Eur. J.Ž .Biochem. 233 1995 9–17.
Copyright © 2022 FDOKUMEN




















