
New and Old Thoughts on the Segmental Organization of the Forebrain in Lampreys
13
Fax +41 61 306 12 34 E-Mail [email protected] www.karger.com Brain Behav Evol 668 DOI: 10.1159/000XXXXXX New and Old Thoughts on the Segmental Organization of the Forebrain in Lampreys Manuel A. Pombal a Manuel Megías a Sylvia M. Bardet b Luis Puelles b a Neurolam Group, Department of Functional Biology and Health Sciences, Faculty of Biology, University of Vigo, Vigo, b Department of Human Anatomy and Psychobiology, Faculty of Medicine, University of Murcia, Spain netic position, they are of considerable interest to stu- dents of vertebrate evolution and have been the subject of many studies in evolutionary developmental biology and neurobiology. During the last century, a variety of studies were car- ried out on the structure of the brain in lampreys, in a effort to establish a fundamental morphological plan of organization in the central nervous system of vertebrates [Ahlborn, 1883; Johnston, 1902, 1912; Edinger, 1905; von Kupffer, 1906; Schilling, 1907; Sterzi, 1907; Tretjakoff, 1909; Herrick, 1910; Herrick and Obenchain, 1913; Holm- gren, 1922; Kuhlenbeck, 1929, 1977; Saito, 1930; Berg- quist, 1932, 1952, 1954; Heier, 1948; Källén, 1951, 1953; Bergquist and Källén, 1953a, b; Schöber, 1964; Schwab, 1973; Nieuwenhuys, 1977; Nieuwenhuys and Nicholson, 1998; Pombal and Puelles, 1999]. In the pioneering stud- ies, the neural tube of craniates was said to be divided during early ontogenesis into two parts: the archenceph- alon (a prechordal vesicle) and the deuterencephalon (an epichordal vesicle) [von Kupffer, 1906]. The archenceph- alon was thought to become the primary prosencepha- lon, and the deuterencephalon was said to subdivide into a mesencephalon and a rhombencephalon (three-vesicle stage). Ahlborn [1883] had already divided the brain in lampreys (Petromyzon) into prechordal and epichordal parts, although with the border drawn at the fovea isth- mi, i.e., caudal to the mesencephalon. In contrast, von Kupffer [1906] and Sterzi [1907] thought that a trans- verse plane, traced just in front of the notochordal tip, Key Words Agnathans Cyclostomes Lamprey Neuromery Forebrain Secondary prosencephalon Diencephalon Evolution Abstract Ten years ago, we published the first detailed prosomeric map of the forebrain in lampreys. In the interim, the proso- meric model has been modified and simplified to better ex- plain numerous data on the expression patterns of regula- tory genes, as well as data from chemical, hodological and neuroembryological experiments, mostly in amniote verte- brates. In this report we first review the main historical con- cepts of lamprey forebrain organization, relating them to either columnar- or segmental-influenced models and ex- plicit or implicit axial references. Next, our previous proso- meric model of the lamprey forebrain is updated, postulat- ing some new hypotheses on the organization of the secon- dary prosencephalon. Copyright © 2009 S. Karger AG, Basel Historical Overview of Concepts of Lamprey Forebrain Organization Lampreys and hagfishes are the only extant jawless vertebrates (agnathans) and thus represent the sister group of gnathostome vertebrates. Due to their phyloge- Published online: $$$ Manuel A. Pombal Neurolam Group, Department of Functional Biology and Health Sciences Faculty of Biology, University of Vigo ES–36.310 Vigo (Spain) Tel. +34 986 812 390, Fax +34 986 812 556, E-Mail pombalauvigo.es © 2009 S. Karger AG, Basel 0006–8977/09/0000–0000$26.00/0 Accessible online at: www.karger.com/bbe BBE668.indd 1 BBE668.indd 1 11.06.2009 09:18:55 11.06.2009 09:18:55
Transcript of New and Old Thoughts on the Segmental Organization of the Forebrain in Lampreys

Fax +41 61 306 12 34E-Mail [email protected]
Brain Behav Evol 668 DOI: 10.1159/000XXXXXX
New and Old Thoughts on the Segmental Organization of the Forebrain in Lampreys
Manuel A. Pombal a Manuel Megías a Sylvia M. Bardet b Luis Puelles b
a Neurolam Group, Department of Functional Biology and Health Sciences, Faculty of Biology, University of Vigo, Vigo , b Department of Human Anatomy and Psychobiology, Faculty of Medicine, University of Murcia , Spain
netic position, they are of considerable interest to stu-dents of vertebrate evolution and have been the subject of many studies in evolutionary developmental biology and neurobiology.
During the last century, a variety of studies were car-ried out on the structure of the brain in lampreys, in a effort to establish a fundamental morphological plan of organization in the central nervous system of vertebrates [Ahlborn, 1883; Johnston, 1902, 1912; Edinger, 1905; von Kupffer, 1906; Schilling, 1907; Sterzi, 1907; Tretjakoff, 1909; Herrick, 1910; Herrick and Obenchain, 1913; Holm-gren, 1922; Kuhlenbeck, 1929, 1977; Saito, 1930; Berg-quist, 1932, 1952, 1954; Heier, 1948; Källén, 1951, 1953; Bergquist and Källén, 1953a, b; Schöber, 1964; Schwab, 1973; Nieuwenhuys, 1977; Nieuwenhuys and Nicholson, 1998; Pombal and Puelles, 1999]. In the pioneering stud-ies, the neural tube of craniates was said to be divided during early ontogenesis into two parts: the archenceph-alon (a prechordal vesicle) and the deuterencephalon (an epichordal vesicle) [von Kupffer, 1906]. The archenceph-alon was thought to become the primary prosencepha-lon, and the deuterencephalon was said to subdivide into a mesencephalon and a rhombencephalon (three-vesicle stage). Ahlborn [1883] had already divided the brain in lampreys (Petromyzon) into prechordal and epichordal parts, although with the border drawn at the fovea isth-mi, i.e., caudal to the mesencephalon. In contrast, von Kupffer [1906] and Sterzi [1907] thought that a trans-verse plane, traced just in front of the notochordal tip,
Key Words
Agnathans � Cyclostomes � Lamprey � Neuromery � Forebrain � Secondary prosencephalon � Diencephalon � Evolution
Abstract
Ten years ago, we published the first detailed prosomeric map of the forebrain in lampreys. In the interim, the proso-meric model has been modified and simplified to better ex-plain numerous data on the expression patterns of regula-tory genes, as well as data from chemical, hodological and neuroembryological experiments, mostly in amniote verte-brates. In this report we first review the main historical con-cepts of lamprey forebrain organization, relating them toeither columnar- or segmental-influenced models and ex-plicit or implicit axial references. Next, our previous proso-meric model of the lamprey forebrain is updated, postulat-ing some new hypotheses on the organization of the secon-dary prosencephalon. Copyright © 2009 S. Karger AG, Basel
Historical Overview of Concepts of Lamprey
Forebrain Organization
Lampreys and hagfishes are the only extant jawless vertebrates (agnathans) and thus represent the sister group of gnathostome vertebrates. Due to their phyloge-
Published online: $ $ $
Manuel A. Pombal Neurolam Group, Department of Functional Biology and Health Sciences Faculty of Biology, University of Vigo ES–36.310 Vigo (Spain) Tel. +34 986 812 390, Fax +34 986 812 556, E-Mail pombalauvigo.es
© 2009 S. Karger AG, Basel0006–8977/09/0000–0000$26.00/0
Accessible online at:www.karger.com/bbe
BBE668.indd 1BBE668.indd 1 11.06.2009 09:18:5511.06.2009 09:18:55

Pombal /Megías /Bardet /Puelles
Brain Behav Evol 668-T12
separated the mesencephalon from the primary prosen-cephalon (including the prospective diencephalon). Cur-rently, we have come to regard the diencephalon proper (apart the hypothalamus) as being epichordal [Puelles et al., 1987; Puelles and Rubenstein, 1993, 2003; Puelles, 1995; Pombal and Puelles, 1999]. The issue of exactly where the primary rostral tip of the notochord lies has clearly been unresolved for a long time. It is of interest that Johnston [1910] reproduced an observation by Ster-
zi [1907, his fig. 115] of a lamprey larva in which the no-tochord seems to practically touch the prospective hypo-thalamus. A similar, earlier observation by His [1892] even suggests that the notochordal tip might underlie a small part of the hypothalamus, which in our opinion opens the possibility that the entire neural tube is actu-ally epichordal.
The body of work cited on lamprey forebrain, our fo-cus in this paper, can be roughly divided into two camps:
AB anterobasal nucleusac anterior commissureah adenohypophysisAH anterior hypothalamusalv anterior lateral ventricle
(telencephalon)Ce cerebellumch optic chiasmCPa caudal paraventricular nucleusctp commissure of the tuberculum
posteriorDH dorsal hypothalamusdpoc dorsal postoptic commissureds dorsal sacDT dorsal thalamusDTc dorsal thalamus (caudal part)DTm dorsal thalamus (medial part)DTsh dorsal thalamus (subhabenular part)EC epichiasmatic nucleusEMT thalamic eminentiafi foramen interventricularfr fasciculus retroflexusHB habenulahc habenular commissurehp1 hypothalamic prosomere 1
(peduncularor caudal hypothalamus)
hp2 hypothalamic prosomere 2 (prepeduncularor rostral hypothalamus)
Hyp hypothalamusibc interbulbar commissureIII oculomotor nucleusIIIn oculomotor nerveIn olfactory nerveinf infundibulumitc intertectal commissureLGN lateral geniculate nucleuslt lamina terminalisltc commissure of the lamina terminalisM medial palliumM1–3 Müller cell 1–3MAM mammillary region sensu latoMB midbrainMnPO median preoptic nucleusMPO medial preoptic nucleusnac nucleus of the anterior commissure
nh neurohypophysisnibc nucleus of the interbulbar commissurenLLat nucleus of the lateral lobeNMLF nucleus of the medial longitudinal
fasciculusnoa anterior olfactory nucleusnot notochordNPC nucleus of the posterior commissurenpic nucleus of the postinfundibular
commissurenpo preoptic nucleusnpoc nucleus of the postoptic commissureNPR nucleus of the neuroporic recessnprc nucleus of the preinfundibular
commissureNTP nucleus of the tuberculum posteriornTPOC nucleus of the tract of the postoptic
commissureOB olfactory bulbog olfactory glomeruliOT optic tectumot optic tractP pineal organp1–p6 prosomeres 1–6Pa palliumpc posterior commissurepch choroidal plexusPCP paracommissural preoptic nucleusPE prethalamic eminentiaPEP posterior entopeduncular nucleusPH posterior hypothalamusph primordium hippocampipic postinfundibular commissureplv posterior lateral ventricle
(telencephalon)PMg magnocellular preoptic nucleuspoc postoptic commissurePP parapineal organprc preinfundibular commissureps pineal stalkPT pretectumptc posttectal commissurePTh prethalamusPTP paraterminal preoptic nucleusPV paraventricular nucleusPVO hypothalamic periventricular organRh rhombencephalon
rlt recessus lateral tectalRM retromammillary areaRPa rostral paraventricular nucleusS striatumSC suprachiasmatic nucleusSCO subcommisural organSCPO septocommissural preoptic areasd sulcus diencephalicus dorsalisshd sulcus hypothalami dorsalisSHL subhippocampal lobeshp sulcus hippocampishyp sulcus hypothalamicus posteriorsi sulcus intermedius diencephalisiv sulcus infraventricularisslH sulcus limitans of Hisslhp sulcus limitans hippocampiSM nucleus of the stria medullarissm sulcus diencephalicus mediussmth sulcus medius thalamiSp SeptumSPa subpalliumSpC spinal cordSPT subpretectal tegmentumsshp sulcus subhippocampalisssp sulcus septist sulcus thalami dorsalisstha sulcus thalami anteriorstm stria medullarissv sulcus diencephalicus ventralissvt sulcus ventralis telencephalisvth sulcus ventralis thalamiteg mesencephalic tegmentumTEL telencephalonTh thalamusTM tuberomammillary nucleusTN tuberal nucleusTNa tuberal nucleus (anterior part)TNp tuberal nucleus (posterior part)ts torus semicircularisVH ventral hypothalamusVHa ventral hypothalamus (anterior part)VHp ventral hypothalamus (posterior part)vpoc ventral postoptic commissureVT ventral thalamuszl zona limitans intrathalamica
Abbreviations used in this paper
BBE668.indd 2BBE668.indd 2 11.06.2009 09:19:0511.06.2009 09:19:05

Forebrain Organization in Lampreys Brain Behav Evol 668-T1 3
explanations of forebrain organization based on segmen-tal interpretations and explanations based on columnar interpretations. These interpretations were related to al-ternative morphological paradigms of forebrain funda-mental structure, whose popularity surged or ebbed throughout the 20th century. The segmental paradigm was predominant before World War I, and the columnar paradigm became prevalent in the postwar period after World War II. The modern advent of molecular genomic research methods has led to a resurgence of the segmental paradigm, however, due to its greater capacity to inte-grate observed gene expression patterns [Puelles et al., 2004] and details of neuroanatomical and functional subdivisions [Straka et al., 2002, 2006].
Neural tube segments, or neuromeres, were first de-scribed in chicken embryos by von Baer [1828] as repeat-ed, transversally arranged, ring-shaped bulges in the em-bryonic neural tube. The concept was further defined by Orr [1887], who introduced the term ‘neuromere’ and his-tologically characterized the interneuromeric boundar-ies, as well as by Hill [1899, 1900] and von Kupffer [1906], among others. The differentiation of prosomeres, meso-meres, and rhombomeres was introduced by Meek [1907, 1909]. These definitions were systematically elaborated to encompass histogenetic patterns in the developing neural wall of anamniotes and amniotes by Bergquist [1932, 1952] and Bergquist and Källén [1953a, b, 1954]. These authors pointed out that intercrossed early trans-verse (segmental) and longitudinal bands [as identified by Orr, 1887] define stereotyped, square migration areas that first represent proliferative units and, subsequently, radial histogenetic units. Each segment, accordingly, contains a full set of dorsoventral radial zonal units [roof, alar, basal and floor plates of His, 1893a, b].
Although this sort of work interpreted morphological patterns in terms of either gross morphology (segmental outer constrictions and corresponding ventricular ridg-es) or neurogenetic data (areal distribution of mitotic fig-ures and early differentiating postmitotic neurons) [see Puelles et al., 1987 for a review], essential corroboration of the resulting brain models had to wait the advent of mappings of developmental genes [Gaunt et al., 1986; Pu-elles and Rubenstein, 1993]. It was soon discovered that the boundaries of the expression patterns of early devel-opmental genes essentially corroborated the checker-board pattern of segmental and longitudinal subdivisions suggested in previous segmental studies. These recent data and associated functional studies (the examination of anterior-posterior and dorsal-ventral molecular pat-terning mechanisms in model organisms, such as zebra-
fish, frogs, chicks, and mice) finally allowed a glimpse of the complex primary causal events underlying segmental and longitudinal patterning of the neural wall, and of-fered a general explanation of the remarkable cross-spe-cies conservative nature of segmental and columnar zon-al patterns. Indeed, the implicit genomic-based mor-phostasis (canalization) is the root of the fundamental plan of the brain observed throughout the evolution of vertebrates, and it underpins conceivable homologous re-lationships in vertebrates.
The alternative columnar paradigm emerged from the discoveries of Gaskell [1886, 1889], Osborn [1888], Strong [1895], Johnston [1898, 1902, 1905a, b] and Herrick [1899] regarding the dorsoventrally arranged, brainstem ‘func-tional’ columns associated with the diverse components of cranial nerves. These findings had a large impact on the field, as they suggested that neuronal columns were of more importance in adult brains than the seemingly tran-sient neuromeres, which had no specific function attached to them. This led to a large-scale loss of interest in neuro-meres and to Herrick’s [1910] epochal paper postulating a columnar subdivision of the entire forebrain, in series with the columnar subdivision of the brainstem. This pa-per contained two aspects that were soon abandoned: (1) the four longitudinal columns postulated in the dien-cephalon were also held to be continuous, one-to-one, with four columns in the telencephalon, but this was not corroborated by detailed observations (only the hypothal-amus is continuous with the telencephalon); this difficul-ty led to a columnar tradition of dealing separately with telencephalon, diencephalon, midbrain and hindbrain, and the tendency to deal perfunctorily with the interven-ing transitional zones (hemispheric stalk, pretectum, isthmus); (2) a separate problem was that Herrick’s [1910] longitudinal columns (epithalamus, dorsal thalamus, ventral thalamus, and hypothalamus) clearly were or-thogonal (as he depicted them himself) to the then wide-ly accepted forebrain longitudinal axis, defined by His [1892, 1893a, b], which curves around the cephalic flexure and ends under the optic chiasma; this axial reference line was based on the histogenetic boundary between the pre-cocious basal plate and the retarded alar plate. Herrick [1927 and subsequent work] attempted to resolve this dis-crepancy by sidestepping the classical axis, disregarding the cephalic flexure, and postulating a new arbitrary straight axis, parallel to his ‘longitudinal columns’ and the limiting ventricular sulci he described in the dience-phalic wall. This new axial reference virtually passed di-rectly from the hindbrain into the hypothalamus, and ended at the tip of the telencephalon (he later briefly com-
BBE668.indd 3BBE668.indd 3 11.06.2009 09:19:0511.06.2009 09:19:05

Pombal /Megías /Bardet /Puelles
Brain Behav Evol 668-T14
mented that this measure was ‘…controversial, …but con-venient…’ [Herrick, 1948]). Some colleagues, such as Kap-pers [1921, 1947], Röthig [1923], von Haller [1929], Berg-quist and Källén [1954] and Coggeshall [1964] did not subscribe to this view and continued to maintain that Herrick’s diencephalic subdivisions were, in fact, not lon-gitudinal (not columnar) and rather corresponded to pos-sible neuromeric entities ( fig. 1 A) [see also Puelles et al., 2007]. Coghill’s [1924, 1928, 1930] precise observations of spatially patterned neurogenesis in Amblystoma , often cited by Herrick as an embryologic support of his colum-nar model, seem, in retrospect, quite supportive of a neu-romeric interpretation (in the light of the curved axis).
Major support for the columnar model also came from Kuhlenbeck, in a series of systematic reports covering an-amniote and amniote vertebrates [Kuhlenbeck, 1927, 1929, etc.; later recollected in Kuhlenbeck, 1973]. In these papers he basically reconstructed Herrick’s limiting ventricular sulci graphically, accepting the latter’s redefined straight longitudinal axis. The fact that the concept of a ‘limiting sulcus’ was never strictly defined within the columnar school, allowed considerable leeway for reconstructing sulci where one’s expectations suggested they should be (in one place, Kuhlenbeck even mentioned a ‘ridge-like sul-cus’, and he also acknowledged that some supposedly lon-gitudinal sulci run orthogonally into the tegmental inward curvature caused by the cephalic flexure).
The ‘straightened’ columnar model of the forebrain was widely employed during the subsequent era of ex-perimental analysis of neuroanatomical connections coupled with histochemical and immunocytochemical studies (e.g., fig. 1 B). The issue of defining a non-imagi-nary forebrain axis, which is all-important for a valid in-terpretation of the developmental topology of the fore-brain region, and, consequently, for understanding the mechanistic and evolutionary causes of their stereotyped patterning and differentiation, was indeed left by the wayside as regards neuroanatomical studies.
As mentioned above, however, developmental gene markers, and the effects of early anterior-posterior and dorsal-ventral patterning, eventually needed to be inte-grated into the prevailing columnar brain model, and they proved to be profoundly inconsistent with the straight columnar axis. These data clearly supported, in-stead, the segmental models by criteria of parsimony and causal explanatory potency (e.g., most gene patterns cor-relate straightforwardly with the more or less curved lon-gitudinal or transversal boundary lines postulated in the segmental paradigm) [Puelles and Rubenstein, 1993, 2003]. This new scenario also drew attention to previ-ously unsuspected segmental functional aspects of the classic columns (e.g., functionally specialized segmental portions of the primary sensory and motor hindbrain columns or the reticular formation of the brainstem), a part of the reinterpretation of the entire diencephalic ‘col-
Ce
MBMBDT TEL
Rh
Th
Ce
VT
Pa
SPa SpCOB
OBHypHyp
alar
floorbasal
notA B
Fig. 1. Schematic sagittal representations of the central nervous system in vertebrates drawn with the axial flexure in mind ( A ), or disregarding the flexure ( B ). Rostral is to the left. The schema in A is from Kappers [1947], with labels modified. Note that the dor-sal and ventral thalamic regions are clearly represented as mor-phologically transversal entities relative to the curved axis (i.e., perpendicular to the longitudinal alar/basal boundary represent-ed in the figure as a dash-line). The hypothalamus therefore is also implied to be transversal. The axial reference line does not enter the telencephalon, which is thus implied to be dorsal relative to
the hypothalamus. This view is fully consistent with the segmen-tal paradigm. The schema in B is modified from Nauta and Karten [1970]; it was published in the heyday of experimental connectiv-ity studies, and is a good example of conceptual disregard for the axial incurvation at the cephalic flexure. Here the thalamus is lo-cated dorsal to the hypothalamus, as dictated by the columnar paradigm. Both elements lie strictly rostral to the mesencephalon, along a straight axis extending uninterruptedly from the brain-stem into the telencephalon. This view is clearly inspired in the columnar paradigm . Rostral is to the left.
BBE668.indd 4BBE668.indd 4 11.06.2009 09:19:0611.06.2009 09:19:06

Forebrain Organization in Lampreys Brain Behav Evol 668-T1 5
umns’ as neuromeric regions [Marín and Puelles, 1995; Puelles, 1995; Cambronero and Puelles, 2000; Straka et al., 2002, 2006; Marín et al., 2008]. The segments have thus finally come to vindicate their importance at the ba-sis of early brain morphogenetic regionalization, as well as for our understanding of the causal and functional or-ganization of vertebrate brains.
Columnar-Influenced Models of the Forebrain in Lampreys Among the pioneering prewar studies of lamprey fore-
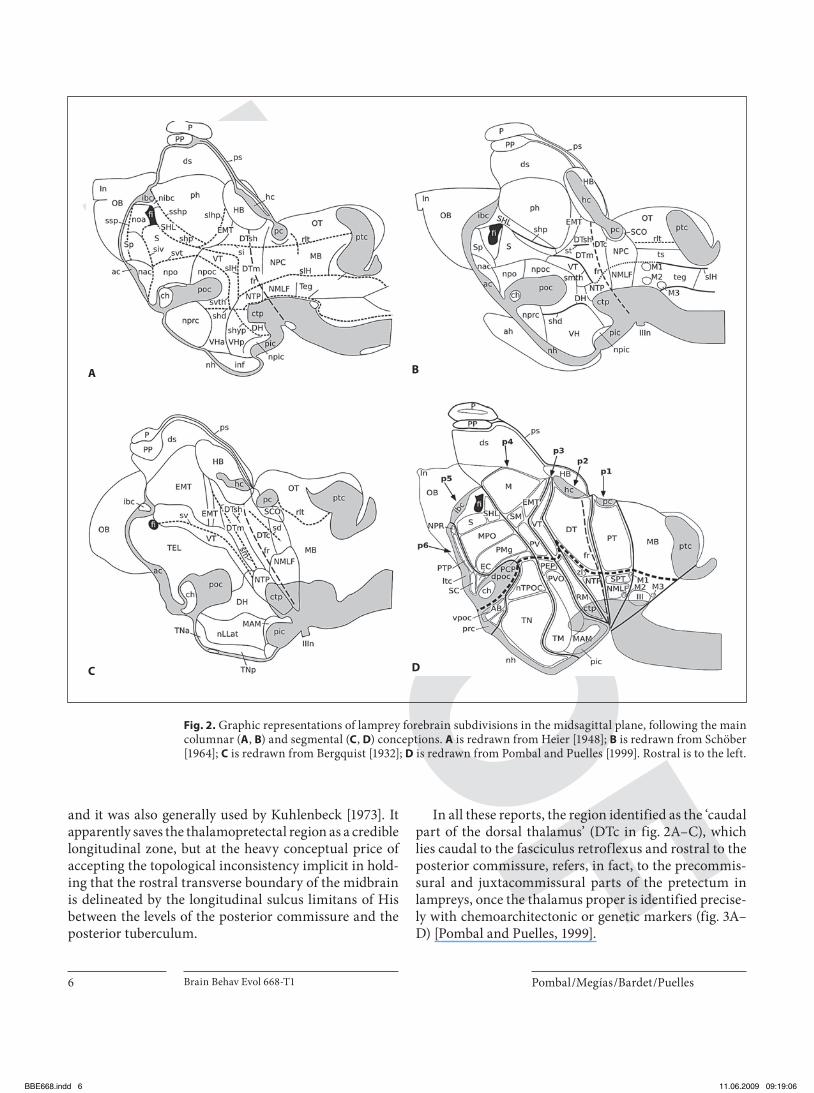
brain organization, those of Herrick [1910] and Herrick and Obenchain [1913] clearly followed the primary colum-nar concept. In the postwar period, in which the columnar concept was prevalent, and continuing until recent times [Nieuwenhuys and Nicholson, 1998], studies on the orga-nization of the forebrain in lampreys generally followed the modified morphological concept introduced by Heier [1948], further elaborated later by Schöber [1964] (see fig. 2 A, B). Heier [1948] explicitly represented the axis of the forebrain as a continuation of the sulcus limitans of His. In his schema, this axis curves around the cephalic flexure until it reaches the region of the posterior tubercu-lum, where it makes a sharp, 90 ° inflexion, and then ex-tends, orthogonally to its previous course, into the floor of the interventricular foramen ( fig. 2 A). Heier [1948] thus seems to have been conscious of the axial problem, and he offered a consensus solution in which he accepted part of the old curved axis, adding to it at the level of the second-ary prosencephalon the straight forebrain axis of Herrick [1922]. As a consequence, the hypothalamus and ventral thalamus are represented as longitudinal (basal) elements, continuous with the telencephalic striatum, whereas the dorsal thalamus and the pretectum appear, in fact, as transverse alar units dorsal to a tubercular-prerubral bas-al plate. There is an ambiguous mixture of columnar and segmental interpretations in Heier s model, which comes rather close to the schema advanced by Kappers [1947; his fig. 66, reproduced here in fig. 1 A], although the relative position of the ventral thalamus differs (compare fig. 1 A and 2 A; we believe that Heier’s [1948] positioning of the ventral thalamus in lampreys is actually in error and is inferior to that suggested by Kappers [1947]).
In Schöber’s [1964] schema ( fig. 2 B), the axial sulcus limitans is traced as a full line only up to the caudal mid-brain; the continuing boundary is traced as a curved, dashed line through the rostral midbrain (corresponding to our pretectal prosomere or synencephalon) and the caudal diencephalon, coming to an end at the tubercu-lum posterior (NTP), essentially as conceived by Kappers
[1947] and Heier [1948]. From that apparent endpoint of Schöber’s basal plate, three sulci proceed in completely different directions. One follows the orthogonal direc-tion used by Heier [1948] into the interventricular fora-men; this extension, however, is no longer interpreted as the sulcus limitans and is first labeled as ‘sulcus medius thalami’ (smth) across the diencephalon (Herrick’s pos-tulated border between ventral and dorsal thalamus), and later as the ‘sulcus hippocampi’ (shp) within the telen-cephalon (separating the unevaginated primordium hip-pocampi from the striatum). Another sulcal boundary was traced as a continuation of the sulcus limitans ros-tralwards along the old axis of His to an ending under-neath the optic chiasma; it is the rostral part of Schöber’s ‘sulcus hypothalami dorsalis’ (shd; fig. 2 B). Finally, the posterior part of the same sulcus appears to extend ros-troventrally around the nucleus of the tuberculum poste-rior (NTP), and then bends ventralward in front of the posterior hypothalamic recess, enclosing the dorsal hy-pothalamus. This last boundary approximates the mam-millary rostral end of the basal plate postulated by Kuhlenbeck [1924, 1927, 1929, 1973], and it also appears in the Heier [1948] schema, identified as ‘sulcus hypo-thalamicus posterior’ (shyp; fig. 2 A). Schöber’s [1964] model, therefore, is likewise a consensus-seeking mix-ture of columnar and segmental interpretations (com-pare fig. 2 B with fig. 1 A), insofar as the hypothalamus and ventral thalamus are, again, conceived explicitly as longitudinal zones; in this case, the apparent error in the identification of the ventral thalamus might lie in the misidentification of Herrick’s ‘sulcus ventralis dienceph-ali’ (corresponding to Heier’s ‘sulcus ventralis thalami’; svth in fig. 2 A) as ‘sulcus medius diencephali’ (i.e., Schö-ber’s ‘sulcus medius thalami’; smth in fig. 2 B), whereas the regions encompassing the dorsal thalamus and pre-tectum implicitly appear, in fact, as transverse segmental zones. Curiously, Schöber’s [1964] model thus pays nom-inal homage to Herrick’s [1922, 1948] schema, but implic-itly admits diverse interpretations of the rostral end of the forebrain axis, and is thus compatible, respectively, with the concepts of segmentationist authors and His [1893a, b], as well as those of Heier [1948] and Kuhlenbeck [1924, 1927, 1929, 1973].
The apparent inconsistency in such partial application of the columnar schema to the forebrain of lampreys was possibly lessened in the mind of these authors and their followers by the attribution to the mesencephalon of the totality or a part of the longitudinal tegmental domain ending rostrally at the posterior tuberculum [e.g., Heier, 1948]. This idea was first introduced by His [1893a, b],
BBE668.indd 5BBE668.indd 5 11.06.2009 09:19:0611.06.2009 09:19:06
Pombal /Megías /Bardet /Puelles
Brain Behav Evol 668-T16
and it was also generally used by Kuhlenbeck [1973]. It apparently saves the thalamopretectal region as a credible longitudinal zone, but at the heavy conceptual price of accepting the topological inconsistency implicit in hold-ing that the rostral transverse boundary of the midbrain is delineated by the longitudinal sulcus limitans of His between the levels of the posterior commissure and the posterior tuberculum.
In all these reports, the region identified as the ‘caudal part of the dorsal thalamus’ (DTc in fig. 2 A–C), which lies caudal to the fasciculus retroflexus and rostral to the posterior commissure, refers, in fact, to the precommis-sural and juxtacommissural parts of the pretectum in lampreys, once the thalamus proper is identified precise-ly with chemoarchitectonic or genetic markers ( fig. 3 A–D) [Pombal and Puelles, 1999].
A B
C D
Fig. 2. Graphic representations of lamprey forebrain subdivisions in the midsagittal plane, following the main columnar ( A , B ) and segmental ( C , D ) conceptions. A is redrawn from Heier [1948]; B is redrawn from Schöber [1964]; C is redrawn from Bergquist [1932]; D is redrawn from Pombal and Puelles [1999]. Rostral is to the left.
BBE668.indd 6BBE668.indd 6 11.06.2009 09:19:0611.06.2009 09:19:06

Forebrain Organization in Lampreys Brain Behav Evol 668-T1 7
Segmental Models of the Lamprey Forebrain A pioneering segmental interpretation of the forebrain
in lampreys was already inherent in the works of von Kupffer [1906], Schilling [1907] and Johnston [1912], but the first detailed segmental analysis of developing and adult lamprey forebrains was reported by Bergquist [1932]. He clearly continued using the curved forebrain axis of His [1893a, b] and followed Palmgren [1921] and Rendahl [1924] in defining the transverse mesencephal-ic-diencephalic boundary just caudal to the posterior commissure. This allowed implicit recognition of three transverse segmental regions with complete sets of four dorsoventral zones (roof, alar, basal and floor plates) in the forebrain; these segmental regions were called by him synencephalon, parencephalon and hypothalamus. In this report he did not officially take a position on the sta-tus of such units as neuromeres, or on their number. Twenty years later, Bergquist [1952, 1954] and Bergquist and Källén [1954] developed their 5-unit neuromeric model of the vertebrate diencephalon, which postulates
two hypothalamic ‘transversal bands’ or neuromeres (comparable to the rostral, or prepeduncular, and the caudal, or peduncular hypothalamic, parts of Puelles and Rubenstein [2003], two parencephalic ‘transversal bands’ (corresponding on one hand to the anterior and posterior parencephalon neuromeres of Rendahl [1924] and, on the other, to the modern prethalamic prosomere, or p3, and the thalamic prosomere, or p2, of Puelles and Rubenstein [2003]), and, finally, one synencephalic ‘transversal band’ (the modern pretectal prosomere, or p1). The ‘synenceph-alon’ of Bergquist [1932] contained only a caudal part of Rendahl’s [1924] synencephalon, however, as he wrongly identified the precommissural pretectum as a caudal part of the dorsal thalamus, lying caudal to the fasciculus ret-roflexus ( fig. 3 A–D). Bergquist clearly did not initially ac-cept Rendahl’s [1924] definition of the thalamopretectal boundary as passing just caudal to the fasciculus retro-flexus, and even in his ultimate paper [1954], this error is carried over to most of the examined specimens, except for the human embryos.
NPCDTc
DTm
OT
Hyp
frIII
VTstmS
alv
alv
OB
OB
In
In
It
fi
EMT
stha
sdfrfr
ch
TEL
Hyp MAM
PTh
TH
stmPE
og
Sp
MPO
sm
VT
DTm
LGN
DTc
NPC
ot
SCO
OT
plv PV
It
PT
III
OT
HBds
alv
PTh
zl
fr
pc
ot
ot
SCO
OT
PT
Th
P
ds
A B
C D
Fig. 3. Diencephalic subdivisions in a lam-prey brain, and position of the thalamus, illustrated by schematic drawings ( A , C)
adapted from Bergquist [1932] and photo-micrographs of sections immunoreacted for calretinin ( B , D ) from Pombal and Pu-elles [1999]. Some major nuclei are repre-sented in the sagittal ( A ) and horizontal ( C ) drawings. In B , note the selective and in-tense calretinin immunoreactivity of the thalamus in the alar plate. In D , note that the plane of section is approximately par-allel to the optic tract and perpendicular to the fasciculus retroflexus, i.e., is strictly horizontally oriented with reference to the bent length axis of the forebrain in lam-preys. Rostral is to the left in A and B and to the top in C and D .
BBE668.indd 7BBE668.indd 7 11.06.2009 09:19:0711.06.2009 09:19:07

Pombal /Megías /Bardet /Puelles
Brain Behav Evol 668-T18
Bergquist [1932, 1952] was particularly interested in the identification of unitary proliferative or migratory (histo-genetic) areas shared among anamniotes, and he investi-gated whether or not these developmental units were de-limited by ventricular sulci (as held by columnar authors). In general, he concluded that early sulci or ventricular con-cavities are of a proliferative nature and lie centered with-in recognizable histogenetic areas (i.e., they correspond to the cavities of the enclosing neuromeres; see fig. 2 C), and only at later stages do some of these proliferative sulci transform or evolve into limiting sulci sensu Herrick [1910] and Kuhlenbeck [1929]. The major limiting sulci defined by these authors in the diencephalon were generally found to be non-limiting relative to histogenetic areas in lam-preys, as well as in all other anamniotes examined. Berg-quist [1932] noted that Herrick [1910] had exclusively ex-amined adult animals. Moreover, he also concluded that these sulci are not longitudinal but represent transverse landmarks (this results automatically from using the axial reference of His [1893a, b] as was obviously understood by Herrick himself [1910, 1922, 1948]). Nevertheless, he main-tained the columnar names sulcus diencephalicus ventra-lis inside the ventral thalamus, sulcus diencephalicus me-dius inside the dorsal thalamus, and sulcus diencephalicus dorsalis inside the synencephalon ( fig. 2 C).
Regarding alar thalamic subdivisions within his parencephalon, Bergquist [1932] identified ‘frontal’, ‘me-dius’, and ‘caudal’ dorsal thalamic regions, separated, re-spectively, by the zona limitans intrathalamica [discov-ered by Rendahl, 1924] and the fasciculus retroflexus. The frontal part corresponds to our prethalamus (old ventral thalamus or alar p3); the medius part represents our thalamus (old dorsal thalamus or alar p2); and the caudal part wrongly refers to the precommisural and jux-tacommissural pretectum [alar p1 of Pombal and Puelles, 1999; the terms ‘prethalamus’ and ‘thalamus’ in present usage were introduced by Puelles and Rubenstein, 2003] ( fig. 3 A–D). Of major interest is Bergquist’s [1932] inter-pretation that the primordium hippocampi of Johnston [1912] actually represents a modified eminentia thalami, placed topologically dorsal to his pars frontalis thalami, that is, dorsal to our prethalamus (the thalamic eminence observed in tetrapods is currently believed to be an inte-gral dorsal part of the prethalamus, and it lies strictly rostral to the thalamic habenular region; Puelles and Ru-benstein [2003] therefore suggested the more accurate name, ‘prethalamic eminence’). Curiously, the term ‘prethalamus’ had already been used by various classic authors, such as Schilling [1907], Sterzi [1907] and Tret-jakoff [1909] for the region subsequently postulated to be
an unevaginated primordium hippocampi by Johnston [1912]. The idea to consider this peculiar region in lam-prey forebrains as a thalamic (prethalamic) eminence was apparently first postulated by Holmgren [1922], on the basis of its morphological position and the fibers of the stria medullaris coursing through it. Such an inter-pretation was thereafter retained by Kuhlenbeck [1929] and Bergquist [1932].
The Prosomeric Model in Lampreys
Transversal Landmarks in the Diencephalon The pretectal- (diencephalic-)-mesencephalic border
is said to pass dorsally behind the posterior commissure, as defined initially by Holmgren’s pupils [Palmgren, 1921; Rendahl, 1924; Bergquist, 1932]. Ventrally, it passes in front of the oculomotor nucleus. In the brain of lampreys there are several pairs of huge reticulospinal cells that have been identified as Müller cells; their relative posi-tions in the brains of various lamprey species (and across the life stages of lampreys) seem to be conserved, and they generally occur in bilateral pairs [see Nieuwenhuys and Nicholson, 1998]. In all lampreys examined there are sev-en pairs of Müller cells, with the three rostralmost pairs traditionally ascribed to the basal mesencephalon [situ-ated within the nucleus reticularis mesencephali; Nieu-wenhuys, 1972]. We believe, however, that Müller cells 1 and 2 lie just rostral to the p1/MB boundary and belong to the pretectal tegmentum [Pombal and Puelles, 1999]. The alar territory of p1 represents the pretectal region, which, in our prosomeric model, is divided into a rostral precommissural part (centered over the synencephalic or metathalamic recess), an intermediate juxtacommissural pretectal domain, and a caudal commissural part, the only component covered by the fibers of the posterior commissure. At alar plate levels, the p1/p2 boundary co-incides with a small cell-poor gap in the periventricular stratum, which is topographically related to the trans-verse trajectory of the fasciculus retroflexus; this tract originates from the habenular nuclei and courses ven-trally towards the floorplate within the caudalmost part of p2 ( fig. 3 A, B). This tract is slightly separated from the pretectal boundary, however, by a strip of thalamic cells ( fig. 3 C, D). The habenular ganglion is delimited from the thalamus by a distinct longitudinal sulcus subhaben-ularis. The transverse border separating the thalamus from the prethalamus is the zona limitans intrathalamica ( fig. 3 , 4 ). Dorsally, this boundary also limits rostrally the habenular region from the prethalamic eminence; ven-
BBE668.indd 8BBE668.indd 8 11.06.2009 09:19:0911.06.2009 09:19:09
Forebrain Organization in Lampreys Brain Behav Evol 668-T1 9
trally it extends into the apex of the cephalic flexure. The rostral boundary of p3 (diencephalic-hypothalamic boundary) is represented by the interprosencephalic boundary (see below). A transverse intrahypothalamic boundary can be identified inside the hypothalamus, just in front of the peduncular tract (see below), separating its peduncular and prepeduncular subdivisions.
The Prosomeric Model in Lampreys Revisited Ten years ago, we published a segmental subdivision of
the prosencephalon in lampreys [Pombal and Puelles, 1999], inspired by the prosomeric model initially reported for tetrapods [Puelles and Rubenstein, 1993; Puelles, 1995]. That model was mostly based on calretinin immu-nocytochemistry (which was used to identify the thala-mus; see fig. 3 B, D), compared to Nissl-stained parallel serial sections, but we also considered other neurochemi-cal and hodological data available at the time. The mini-mal morphogenetic deformation of lamprey forebrains also helped us to establish particular boundaries in the forebrain, such as the zona limitans intrathalamica. Some years later, a modified and simplified segmental model [Puelles and Rubenstein, 2003] was intended to resolve some difficulties found in interpreting the hypothala-mus-telencephalon transition in chickens and mice, con-sistently with respect to the expression pattern of devel-opmental genes. Publication and use of wrong neuromor-phological models is increasingly felt to be a ‘sin’, and neuroanatomic ‘sinners’ should take upon themselves acts of repentance and atonement of the errors possibly caused among naïve readers. Basically, Puelles and Rubenstein [2003] retained only their previous p1–p3 diencephalic prosomeres, whereas the remaining p4–p6 units first pos-tulated in the secondary forebrain [Puelles and Ruben-stein, 1993] were momentarily abandoned, leaving the secondary prosencephalon as a large proneuromere or protosegment with rostral and caudal (prepeduncular and peduncular) hypothalamic parts and an undivided telencephalon. The hypothalamic parts were said to be clearly separated by the transverse intrahypothalamic boundary, demarcated partly by the compact peduncular tract at superficial levels, but this boundary could not be easily prolongated towards the telencephalic roof plate (a criterion for a segmental boundary). The evidence in sup-port of neuromeres in the secondary prosencephalon was thus considered inconclusive. More recently, however, a complementary idea has emerged: the intrahypothalam-ic boundary apparently ends by separating the preoptic area – a part of the molecularly defined subpallium – from the rest of the subpallium; this newly determined course
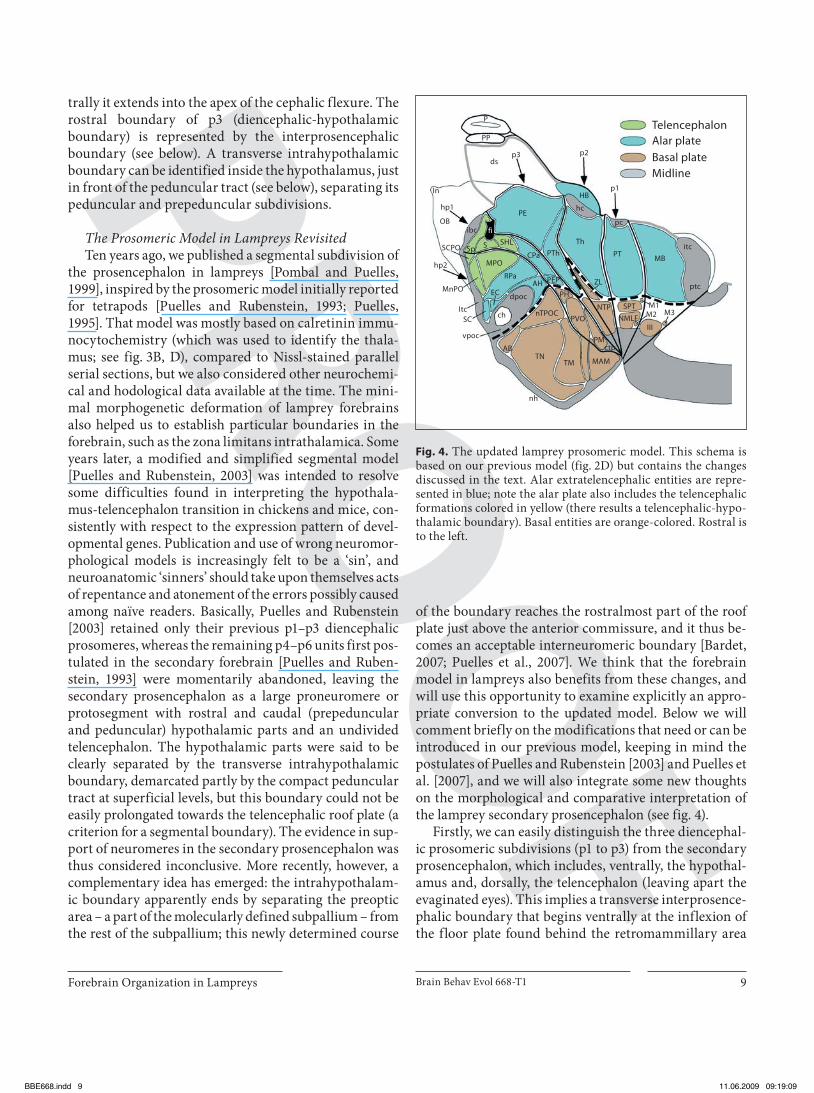
of the boundary reaches the rostralmost part of the roof plate just above the anterior commissure, and it thus be-comes an acceptable interneuromeric boundary [Bardet, 2007; Puelles et al., 2007]. We think that the forebrain model in lampreys also benefits from these changes, and will use this opportunity to examine explicitly an appro-priate conversion to the updated model. Below we will comment briefly on the modifications that need or can be introduced in our previous model, keeping in mind the postulates of Puelles and Rubenstein [2003] and Puelles et al. [2007], and we will also integrate some new thoughts on the morphological and comparative interpretation of the lamprey secondary prosencephalon (see fig. 4 ).
Firstly, we can easily distinguish the three diencephal-ic prosomeric subdivisions (p1 to p3) from the secondary prosencephalon, which includes, ventrally, the hypothal-amus and, dorsally, the telencephalon (leaving apart the evaginated eyes). This implies a transverse interprosence-phalic boundary that begins ventrally at the inflexion of the floor plate found behind the retromammillary area
TelencephalonAlar plateBasal plateMidline
p2
p1HB
hc
dsp3
PP
P
In
hp1
OB
Th
pc
MBPT
ZL ptc
itcPThCPa
S
MPO
SHLSp
PE
fiibc
SCPO
MnPO
hp2
ItcSC
vpoc
ch
dpocEC
RPaAH PEP
nTPOC
PH
AB
TM
nh
MAMTN
PVO
NTP
ctp
SPT
NMLF
M1M2 M3
III
PM
Fig. 4. The updated lamprey prosomeric model. This schema is based on our previous model (fig. 2D) but contains the changes discussed in the text. Alar extratelencephalic entities are repre-sented in blue; note the alar plate also includes the telencephalic formations colored in yellow (there results a telencephalic-hypo-thalamic boundary). Basal entities are orange-colored. Rostral is to the left.
BBE668.indd 9BBE668.indd 9 11.06.2009 09:19:0911.06.2009 09:19:09

Pombal /Megías /Bardet /Puelles
Brain Behav Evol 668-T110
(the retromammillary area encloses the ‘commissure of the posterior tuberculum’, or retromammillary commis-sure; note that in lampreys the bulge called ‘posterior tu-berculum’ apparently lies rostral to this boundary, i.e., within the hypothalamus, although a region lying more caudally in the diencephalic tegmentum is often called by the same name. Compare our nucleus of the tuberculum posterior across the p3 and p2 tegmental regions in fig. 2 D, 4 ) [see also Wulliman and Puelles, 1999 and Wulliman et al., 1999]. The interprosencephalic boundary then passes dorsalward just behind the hypothalamic periventricular organ (PVO), the posterior hypothalamus (PH), the pos-terior entopeduncular area (PEP), and the caudal para-ventricular nucleus (CPa), and ends dorsally between the subhippocampal lobe and the ‘primordium hippocampi’ of Johnston [1912] or the ‘medial pallium’ of Pombal and Puelles [1999], which we identified here as the prethalam-ic eminence (PE), thus finally reaching the choroidal roof plate. We already expressed doubts about the hypothetic ‘hippocampal’ nature of the PE [Pombal and Puelles, 1999], although we conservatively decided to interpret it as ‘medial pallium.’ This attitude is now reinforced, and leads to our viewing this entity as a modified ‘prethalam-ic eminence’, in full agreement with Holmgren [1922], Kuhlenbeck [1929, 1973], Bergquist [1932, 1954], Källén [1951, 1953], and Schwab [1973]. This concept is consistent with its topological position relative to the interventricu-lar foramen and the velum transversum in both lampreys and gnathostomes. The classical ‘eminentia thalami’ re-ceived this name because of its protrusion at the back of the interventricular foramen, at a time when the thalamus and prethalamus were not distinguished from one anoth-er. The evolutionary singularity of an unevaginated hip-pocampal pallium was a genial conjecture of Johnston [1912] which remains an interesting possibility that will probably soon be tested fully with molecular markers. A number of hodological studies have considered the con-nections of our prethalamic eminence to be compatible with Johnston’s conjecture, although often the alternative eminential interpretation was not contemplated [see Nieu-wenhuys and Nicholson, 1998, for review]. In any case, so far there is no strong morphologic, molecular or causal evidence supporting this idea. In fact, the prethalamic eminence in lampreys has been shown to express Lhx1/5 mRNA [Osório et al., 2006], a feature that is typical of the prethalamic eminence in tetrapods [Moreno et al., 2004]. Accordingly, we now include the prethalamic eminence in the dorsal (alar) region of p3, as did Puelles and Ruben-stein [2003]. It seems therefore prudent to fall back on the notion that the telencephalic evagination process in lam-
preys – and therefore the resulting interventricular fora-men – are basically comparable to the condition found in gnathostomes; the telencephalic vesicle in agnathans is unusually small and might actually lack dorsal and me-dial pallium components [Puelles, 2001].
The velum transversum, identified in larval and adult lampreys both by von Kupffer [1906] and Sterzi [1907] as a typical transverse fold of the roof plate between the lam-ina supraneuroporica and the dorsal choroid sac, was held by these authors (and many others thereafter) to lim-it the telencephalon from the diencephalon. We consider it now a roof plate landmark for the p3/p2 boundary, as revealed by the zona limitans ridge that systematically converges with it in tetrapods [L. Puelles, unpublished observations].
The dorsal aspect of the interventricular foramen is limited by the presumptive septal region of the telenceph-alon (Sp), which receives at its caudal end the insertion of the prosencephalic choroidal tela and largely coincides at the local midline with the classic interbulbar commissure; the latter might thus represent the homolog of the septal commissural plate of gnathostomes, possibly including a primordium of the anterior commissure at its rostral end, as well as, eventually, primordial elements of a hippocam-pal commissure at its caudal end. The ventral aspect of the interventricular foramen is occupied by the bulge of the striatum, whose larger part is found inside the evaginated telencephalic vesicle. Tracing of the intrahypothalamic boundary in the modified model (see below) causes slight modification of the shape of the preoptic area and its boundary with the striatum (there is no pallidum in lam-preys, apparently). Moreover, the former paraterminal preoptic area (PTP) and the neuroporic recess area (NPR) are conveniently reclassified as the median preoptic nucle-us (MnPO) and the septocommissural preoptic area (SCPO), respectively. So far, we have not identified a telen-cephalic-hypothalamic transition zone, longitudinally separating the preoptic and paraventricular areas and serving as the boundary between the telencephalon and the hypothalamus, but such a boundary is clearly possible and could be identified eventually with molecular markers (see border between yellow and blue domains in fig. 4 ).
Regarding subdivisions in the hypothalamus, the dis-tinction of rostral (prepeduncular) and caudal (peduncu-lar) hypothalamic parts [Puelles and Rubenstein, 2003] is straightforward, corresponding (albeit roughly, due to er-rors noted above in the identification of the ‘ventral thala-mus’) to the columnar ‘ventral’ and ‘dorsal’ hypothalami, respectively [Heier, 1948; Schöber, 1964]. These hypotha-lamic parts can be now viewed as full neuromeric units
BBE668.indd 10BBE668.indd 10 11.06.2009 09:19:1011.06.2009 09:19:10

Forebrain Organization in Lampreys Brain Behav Evol 668-T1 11
(hypothalamic prosomeres hp1 and hp2), thanks to the new definition of the intrahypothalamic boundary [Bar-det, 2007; Puelles et al., 2007]. The peduncular hypothala-mus (new hp1 prosomere; fig. 4 ), expands dorsally into the evaginated telencephalic vesicle, and largely ends at the septal roof plate (except for a small caudal portion that ends at the choroidal tela in front of the PE). On the other hand, the prepeduncular hypothalamus (new hp2 proso-mere), originates the eye evagination within its alar plate domain and ends dorsally at the unevaginated telencepha-lon impar or preoptic area (enclosing the anterior commis-sure at its septocommissural preoptic roof plate region). Note that, as a result of data collected in chicks by Bardet [2007], we now place most of the mammillary primordium within hp2, and leave the retromammillary area within hp1, differing in this detail from Puelles and Rubenstein [2003]. The ventral parts of hp1 and hp2 are now believed to converge towards the potential tip of the notochord (see comments in the first section). This important change is associated with reinterpretation of the hypothalamic peri-ventricular organ as a longitudinal entity [Bardet, 2007], whereas we initially thought the organ was approximately transversal [Pombal and Puelles, 1999].
The old magnocellular preoptic nucleus (PMg) clearly corresponds to a rostral paraventricular nucleus (RPa) within alar hp2, and caudally it contacts the equally mag-nocellular caudal paraventricular nucleus (CPa) in alar hp1. We are delimiting the CPa slightly differently than previously, shortening it ventrally (leaving space for the new PEP) and expanding it dorsally behind the preoptic area, until it contacts the classic ‘subhippocampal lobe’. The ‘subhippocampal lobe’, which we first postulated might be part of the subpallial amygdala [Pombal and Puelles, 1999], but was found not to express Dlx protein at all [M.A. Pombal, M. Martínez-de-la-Torre, and L. Pu-elles, unpublished observations], thereby invalidating that hypothesis, as Dlx genes are characteristic of the sub-pallium in all vertebrate species examined. The ‘subhip-pocampal lobe’ is now interpreted as an unevaginated part of the pallial amygdala [compare Puelles and Ruben-stein, 2003], corresponding to the pallial extended amyg-dala identified by Puelles et al. [2007] and Bardet et al. [2008] in tetrapods.
Our previous ‘paracommissural preoptic nucleus’ (PCP) can no longer be considered a preoptic entity, due to its position ventral to the paraventricular formations. After being slightly expanded caudalwards under CPa, it nicely corresponds topologically to the concept of a sub-paraventricular area, i.e., the Dlx -expressing ventral li-men of the hypothalamic alar plate according to Puelles
and Rubenstein [2003], Puelles et al. [2004, 2007], and Bardet [2007]. The subparaventricular area develops in tetrapods the suprachiasmatic (SC), epichiasmatic (EC) and anterior hypothalamus (AH) nuclei within hp2 and the posterior entopeduncular area (PEP) within hp1 ( fig. 4 ). Note that the new PEP lies in the added caudal extension of the subparaventricular band, and it substi-tutes part of the ventral end of the former paraventricular nucleus. Our former PEP area is clearly located in the basal plate, and was thus wrongly interpreted as PEP; at the time, we ignored the fact that the corresponding en-topeduncular element of tetrapods is an alar plate deriva-tive. It is here more appropriately renamed posterior hy-pothalamus area (PH).
Finally, various infundibular or tuberal hypothalamic elements that we previously classified as floor plate de-rivatives are now thought to be basal plate formations, keeping in mind that the floor plate of hp2 and hp1 might be quite reduced ( fig. 4 ). The resulting final schema clear-ly emphasizes the morphological axial continuity of the alar and basal longitudinal zones across midbrain, dien-cephalon and hypothalamus, consistent with the restrict-ed axial bending at the cephalic flexure. The segmental or neuromeric anteroposterior subdivisions (p1–p3 in the diencephalon and hp1–hp2 in the secondary prosenceph-alon) are largely wedge-shaped, with constricted ventral portions and expanded dorsal ones, and their mutual boundaries are complete, crossing all the longitudinal zones orthogonally from floor to roof. The hypothalam-ic neuromeres are unique in being related either to the evaginated telencephalic vesicle (hp1) or to the hypophy-sis, eye vesicle and preoptic telencephalon impar (hp2). The developmental peculiarities of hp2 could eventually be clarified, once we better understand the causal pat-terning role of the prechordal plate mesoderm, even if all prosomeres might now, in principle, be conjectured to be epichordal, as initially suggested by His [1892].
Acknowledgements
This study was supported by the Spanish Ministry of Science and Technology (MCyT)-FEDER grant number BFI2003-04479, Spanish MEC-FEDER grant number BFU2006-14127, Xunta de Galicia (Spain) grant number PGIDT-04PXIC31004PN, and Uni-versity of Vigo grant number 07V1A12 to MAP; MCyT grant number BFU2008-04156 and SENECA Foundation contract 04548/GERM/06-10891 to LP; and a predoctoral fellowship from the MCyT to SB.
BBE668.indd 11BBE668.indd 11 11.06.2009 09:19:1011.06.2009 09:19:10

Pombal /Megías /Bardet /Puelles
Brain Behav Evol 668-T112
References
Ahlborn F (1883) Untersuchungen über das Ge-hirn der Petromyzonten. Zschr Wiss Zool 39: 191–294.
Bardet SM (2007) Organización morfológica y citogenética del hipotálamo del pollo sobre base de mapas moleculares. Doctoral thesis (Neuroscience programme). Univ. Murcia (Spain).
Bardet SM, Martínez-de-la-Torre M, Northcutt RG, Rubenstein JLR, Puelles L (2008) Con-served pattern of OTP positive cells in the paraventricular nucleus and other hypotha-lamic sites of tetrapods. Brain Res Bull 75: 231–235.
Bergquist H (1932) Zur Morphologie des Zwi-schenhirns bei niederen Wirbeltieren. Acta Zool 13: 57–304.
Bergquist H (1952) Studies on the cerebral tube in vertebrates. The neuromeres. Acta Zool 33: 117–187.
Bergquist H (1954) Ontogenesis of diencephalic nuclei in vertebrates. A comparative study. Kgl Fysiogr Sällsk Lund Handl NF 65: 1–34.
Bergquist H, Källén B (1953a) Studies on the to-pography of the migration areas in the verte-brate brain. Acta Anat 17: 353–369.
Bergquist H, Källén B (1953b) On the develop-ment of neuromeres to migration areas in the vertebrate cerebral tube. Acta Anat 18: 65–73.
Bergquist H, Källén B (1954) Notes on the early histogenesis and morphogenesis of the cen-tral nervous system in vertebrates. J Comp Neurol 100: 627–659.
Cambronero F, Puelles L (2000) Rostrocaudal nuclear relationships in the avian medulla oblongata: a fate map with quail chick chi-meras. J Comp Neurol 427: 522–545.
Coggeshall RE (1964) A study of diencephalic development in the albino rat. J Comp Neu-rol 122: 241–269.
Coghill GE (1924) Correlated anatomical and physiological studies on the growth of the nervous system of Amphibia. IV. Rates of proliferation and differentiation in the cen-tral nervous system of Amblystoma puncta-tum . J Comp Neurol 37: 71–120.
Coghill GE (1928) Correlated anatomical and physiological studies on the growth of the nervous system of Amphibia. VIII. The de-velopment of the pattern of differentiation in the cerebrum of Amblystoma punctatum . J Comp Neurol 45: 227–247.
Coghill GE (1930) Correlated anatomical and physiological studies on the growth of the nervous system of Amphibia. VIII. The mechanism of association of Amblystoma punctatum . J Comp Neurol 51: 311–375.
Edinger L (1905) Die Deutung des Vorderhirns bei Petromyzon. Anat Anz 26: 633–635.
Gaskell WH (1886) On the structure, distribu-tion and function of the nerves which inner-vate the visceral and vascular systems. J Physiol 7: 1–81.
Gaskell WH (1889) On the relation between the structure, function, distribution and origin of the cranial nerves, together with a theory on the origin of the nervous system of verte-brata. J Physiol 10: 153–211.
Gaunt SJ, Miller JR, Powell DJ, Duboule D (1986) Homoeobox gene expression in mouse em-bryos varies with position by the primitive streak stage. Nature 324: 662–664.
Heier P (1948) Fundamental principles in the structure of the brain. A study of the brain of Petromyzon fluviatilis . Acta Anat 5(suppl 8): 7–213.
Herrick CJ (1899) The cranial and first spinal nerves of Menidia ; a contribution upon the nerve components of the bony fishes. J Comp Neurol 9: 153–455.
Herrick CJ (1910) The morphology of the fore-brain in Amphibia and Reptilia . J Comp Neurol 20: 413–547.
Herrick CJ (1922) Functional factors in the mor-phology of the forebrain of fishes. Madrid: Libro en honor de D. Santiago Ramón y Cajal 1: 143–202.
Herrick CJ (1927) The amphibian forebrain. IV. The cerebral hemispheres of Amblystoma . J Comp Neurol 43: 231–325.
Herrick CJ (1948) The brain of the tiger salaman-der, Ambystoma tigrinum . Chicago: The University of Chicago Press.
Herrick CJ, Obenchain J (1913) Notes on the anatomy of a cyclostome brain: Ichthyomy-zon concolor . J Comp Neurol 23: 635–675.
Hill C (1899) Primary segments of the vertebrate head. Anat Anz 16: 335–369.
Hill C (1900) Developmental history of primary segments of the vertebrate head. Zool Jahrb Abt Anat Ontog 13: 393–446.
His W (1892) Zur allgemeinen Morphologie des Gehirns. Arch Anat Physiol Anat Abt 16: 346–383.
His W (1893a) Über das frontale Ende des Ge-hirnrohres. Arch Anat Physiol Anat Abt 3/4: 157–171.
His W (1893b) Vorschläge zur Eintheilung des Gehirns. Arch Anat Physiol Anat Abt 3/4: 173–179.
Holmgren N (1922) Points of view concerning forebrain morphology of lower vertebrates. J Comp Neurol 34: 391–460.
Johnston JB (1898) The olfactory lobes, fore-brain, and habenular tracts of Acipenser. Zool Bull 1: 221–241.
Johnston JB (1902) The brain of Petromyzon . J Comp Neurol 12: 2–86.
Johnston JB (1905a) The cranial nerve compo-nents of Petromyzon. Morphol Jahrb 34: 149–203.
Johnston JB (1905b) The morphology of the ver-tebrate head, from the viewpoint of the func-tional divisions of the nervous system. J Comp Neurol Psych 15: 175–275.
Johnston JB (1910) A comment upon recent con-tributions on the brain of Petromyzonts. Anat Anz 37: 153–158 and 182–194.
Johnston JB (1912) The telencephalon in cyclo-stomes. J Comp Neurol 22: 341–404.
Källén B (1951) Contributions to the ontogeny of the nuclei and the ventricular sulci in the vertebrate forebrain. Kungl Fysiogr Sällsk Lund Handl NF 62: 5–34.
Källén B (1953) Studies on the topography of the migration areas in the vertebrate brain. Acta Anat 17: 353–369.
Kappers CUA (1921) Die vergleichende Anato-mie des Nervensystems der Wirbeltiere und des Menschen. 2 Vols. Haarlem: De Erven F. Bohn.
Kappers CUA (1947) Anatomie comparée du système nerveux. Haarlem: De Erven F. Bohn and Paris: Masson & Cie.
Kuhlenbeck H (1924) Über die Homologien der Zellmassen im Hemisphärenhirn der Wir-beltiere. Folia Anat Jpn 2: 325–364.
Kuhlenbeck H (1927) Vorlesungen über das Zentralnervensystem der Wirbeltiere. Eine Einführung in die Gehirnanatomie auf ver-gleichender Grundlage. Jena: Fischer.
Kuhlenbeck H (1929) Über die Grundbestand-teile des Zwischenhirnbauplans der Anam-nier. Morphol Jahrb 63: 50–95.
Kuhlenbeck H (1973) The central nervous sys-tem of vertebrates, vol 3, part II. Overall morphologic pattern. Basel: S. Karger.
Kuhlenbeck H (1977) The central nervous sys-tem of vertebrates, vol 5, part I. Derivatives of the prosencephalon: diencephalon and telencephalon. Basel: S. Karger.
Marín F, Puelles L (1995) Morphological fate of rhombomeres in quail/chick chimeras: a seg-mental analysis of hindbrain nuclei. Eur J Neurosci 7: 1714–1738.
Marín F, Aroca P, Puelles L (2008) Hox gene co-linear expression in the avian medulla ob-longata is correlated with pseudorhombo-meric domains. Dev Biol 323: 230–247.
Meek A (1907) The segments of the vertebrate brain and head. Anat Anz 31: 408–415.
Meek A (1909) The encephalomeres and cranial nerves of an embryo of Acanthias vulgaris . Anat Anz 34: 473–475.
Moreno N, Bachy I, Retaux S, González A (2004) LIM-homeodomain genes as developmental and adult genetic markers of Xenopus fore-brain functional subdivisions. J Comp Neu-rol 472: 52–72.
Nauta WJH, Karten HJ (1970) A general profile of the vertebrate brain, with sidelights on the ancestry of cerebral cortex. In: The neurosci-ences: second study program (Schmitt FO, ed), pp 7–26. New York: The Rockefeller Uni-versity Press.
Nieuwenhuys R (1972) Topological analysis of the brain stem of the lamprey Lampetra f lu-viatilis . J Comp Neurol 145: 165–177.
Nieuwenhuys R (1977) The brain of the lamprey in a comparative perspective. Ann NY Acad Sci 299: 97–145.
BBE668.indd 12BBE668.indd 12 11.06.2009 09:19:1011.06.2009 09:19:10

Forebrain Organization in Lampreys Brain Behav Evol 668-T1 13
Nieuwenhuys R, Nicholson C (1998) Lampreys, Petromyzontoidea. In: The central nervous system of vertebrates (Nieuwenhuys R, ten Donkelaar HJ, Nicholson C, eds), vol 1, pp 397–495. Berlin: Springer-Verlag.
Orr HA (1887) Contribution to the embryology of the lizard. J Morphol 1: 311–372.
Osborn HF (1888) A contribution to the internal structure of the amphibian brain. J Morphol 2: 51–96.
Osório J, Megías M, Pombal MA, Retaux S (2006) Dynamic expression of the LIM-homeodo-main gene Lhx15 through larval brain devel-opment of the sea lamprey (P etromyzon ma-rinu s) . Gene Expr Patterns 6: 873–878.
Palmgren A (1921) Embryological and morpho-logical studies on the midbrain and cerebel-lum of vertebrates. Acta Zool Stockh 2: 1–94.
Pombal MA, Puelles L (1999) Prosomeric map of the lamprey forebrain based on calretinin immunocytochemistry, Nissl stain, and an-cillary markers. J Comp Neurol 414: 391–422.
Puelles L (1995) A segmental morphological par-adigm for understanding vertebrate fore-brains. Brain Behav Evol 46: 319–337.
Puelles L (2001) Thoughts on the development, structure and evolution of the mammalian and avian telencephalic pallium. Phil Trans R Soc Lond B Biol Sci 356: 1583–1598.
Puelles L, Rubenstein JL (1993) Expression pat-terns of homeobox and other putative regu-latory genes in the embryonic mouse fore-brain suggest a neuromeric organization. Trends Neurosci 16: 472–479.
Puelles L, Rubenstein JL (2003) Forebrain gene expression domains and the evolving proso-meric model. Trends Neurosci 26: 469–476.
Puelles L, Amat JA, Martínez-de-la-Torre M (1987) Segment-related, mosaic neurogenet-ic pattern in the forebrain and mesencepha-lon of early chick embryos: I. Topography of AChE-positive neuroblasts up to stage HH18. J Comp Neurol 266: 247–268.
Puelles L, Martínez S, Martínez-de-la-Torre M, Rubenstein JLR (2004) Gene maps and relat-ed histogenetic domains in the forebrain and midbrain. In: The rat nervous system (Paxi-nos G, ed), ed 3, pp 3–25. San Diego, CA: Ac-ademic Press.
Puelles L, Martínez-de-la-Torre M, Paxinos G, Watson C, Martínez S (eds) (2007) The chick brain in stereotaxic coordinates: an atlas fea-turing neuromeric subdivisions and mam-malian homologies. San Diego, CA: Aca-demic Press/Elsevier.
Rendahl H (1924) Embryologische und mor-phologische Studien über das Zwischenhirn beim Huhn. Acta Zool Stockh 5: 241–344.
Röthig P (1923) Beiträge zum Studium des Zentralnervesystems der Wirbeltiere. VIII. Über das Zwischenhirn der Amphibien. Arch Mikr Anat 98: 616–645.
Saito T (1930) Über das Gehirn des japanischen Flussneunauges ( Entosphenus japonicus Martens). Fol Anat Jpn 8: 189–263.
Schilling K (1907) Über das Gehirn von Petro-myzon fluviatilis . Abh Senckenberg Natur-forsch Ges Frankfurt 30: 423–446.
Schöber W (1964) Vergleichend-anatomische Untersuchungen am Gehirn der Larven und adulten Tiere von Lampetra fluviatilis (Lin-né, 1758) und Lampetra planeri (Bloch, 1874). J Hirnforsch 7: 107–209.
Schwab ME (1973) Some new aspects about the prosencephalon of Lampetra fluviatilis (L.). A cytoarchitectural and comparative study. Acta Anat 86: 353–375.
Sterzi G (1907) Il sistema nervoso centrale dei vertebrati. Vol. 1, Ciclostomi, Angelo Draghi, Padova.
Straka H, Baker R, Gilland E (2002) The frog as a unique vertebrate model for studying the rhombomeric organization of functionally identified hindbrain neurons Brain Res Bull 57: 301–305.
Straka H, Baker R, Gilland E (2006) Preservation of segmental hindbrain organization in adult frogs. J Comp Neurol 494: 228–245.
Strong OS (1895) The cranial nerves of amphib-ia. A contribution to the morphology of the vertebrata nervous system. J Morphol 10: 101–230.
Tretjakoff D (1909) Das Nervensystem von Am-mocoetes. I. Gehirn. Arch Mikrosk Anat 74: 636–779.
von Baer KE (1828) Über die Entwicklungsge-schichte der Thiere. Beobachtung und Re-flexion, 2 vols. Königsberg: Bornträger.
von Haller V (1929) Die Gliederung des Zwi-schen- und Mittelhirns der Wirbeltiere. Morphol Jahrb 63: 359–407.
von Kupffer K (1906) Die Morphogenie des Cen-tralnervensystems. In: Handbuch der Ver-gleichenden und Experimentellen Entwick-lungslehre der Wirbeltiere (Hertwig O, ed), vol 2, part III, pp 1–272. Jena: Fisher.
Wulliman MF, Puelles L (1999) Postembryonic neural proliferation in the zebrafish fore-brain and its relationship to prosomeric do-mains. Anat Embryol 329: 329–348.
Wulliman MF, Puelles L, Wicht H (1999) Early postembryonic neural development in the zebrafish: a 3-D reconstruction of forebrain proliferation zones shows their relation to prosomeres. Eur J Morphol 37: 117–121.
BBE668.indd 13BBE668.indd 13 11.06.2009 09:19:1011.06.2009 09:19:10




















