
The CBFβ Subunit Is Essential for CBFα2 (AML1) Function In Vivo
Upload
independentCategory
view
1download
0
Peter L. Jones and Marlene RabinovitchDennis A. Wigle, Karen E. Thompson, Shmuel Yablonsky, Syed H. E. Zaidi, Claire Coulber,
Artery Smooth Muscle CellsAML1-like Transcription Factor Induces Serine Elastase Activity in Ovine Pulmonary
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1998 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.83.3.2521998;83:252-263Circ Res.
http://circres.ahajournals.org/content/83/3/252World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

AML1-like Transcription Factor Induces Serine ElastaseActivity in Ovine Pulmonary Artery Smooth Muscle Cells
Dennis A. Wigle, Karen E. Thompson, Shmuel Yablonsky, Syed H.E. Zaidi, Claire Coulber,Peter L. Jones, Marlene Rabinovitch
Abstract—In previous studies, we showed that induction of pulmonary artery (PA) smooth muscle cell (SMC) elastaseactivity by serum-treated elastin (STE) requires DNA transcription. We therefore used differential mRNA display toidentify transcripts expressed coincident with elastase induction. Twenty-four individual transcripts were differentiallyexpressed from a screen of'2000 mRNA sequences. An mRNA with sequence homology to the human transcriptionfactor AML1 was identified and subsequently cloned from ovine PA SMCs. Since AML1 binds to a consensus sequencein the promoter of neutrophil elastase, we pursued the possibility that AML1 is a candidate transcription factor for SMCelastase. We documented by immunohistochemistry that serum stimulation induces increased expression of AML1 inthe nucleus of PA SMCs. We also showed that STE induction of elastase activity is associated with early expressionof AML1 mRNA and protein and that AML1 consensus sequence DNA binding activity is increased in nuclear extractsof STE-treated cells. In addition, AML1 antisense oligonucleotides reduced serum induction of elastase activity. Ourstudy thus provides the first functional evidence of AML1 transcriptional activity related to elastase genes and offersnovel insights into the broader biological significance of AML1 in nonmyeloid cells. (Circ Res. 1998;83:252-263.)
Key Words: differential display polymerase chain reactionn pulmonary hypertensionn AML1 transcription
Ultrastructural observations in lung biopsy tissue frompatients with congenital heart defects showing fragmen-
tation of the arterial elastic lamina first suggested thatheightened elastolytic activity may play a critical role in thedevelopment of pulmonary vascular disease.1 Further studiesin experimental animals confirmed high elastin turnover inthe pulmonary artery as a feature of monocrotaline-inducedprogressive pulmonary hypertension.2 Increased vascularelastolytic activity was then documented as preceding thedevelopment of sustained pulmonary hypertension and theinduction of pulmonary artery hypertrophy associated withhypoxia3 or with the injection of the toxin monocrotaline.4 Acause-and-effect relationship between increased vascularelastolytic activity and the initiation of increased pulmonarypressure with its concomitant structural remodeling wassuggested when elastase inhibitors were shown to prevent thedevelopment of experimental pulmonary hypertension.3,5,6Wesubsequently identified, in hypertensive pulmonary arteries, a20-kDa serine elastase that localized to SMCs and appearedto be related to the serine proteinase adipsin.7 These studiessupported previous reports describing the production of asimilar enzyme by aortic SMCs.8,9
Since endothelial injury is an early feature of both pulmo-nary10 and systemic vascular11 diseases, we further investi-gated whether the mechanism accounting for the induction of
EVE might be related to transfer of a serum factor to themedial SMC layer as a consequence of loss of endothelial cellbarrier function. We demonstrated that incubation of PASMCs with either serum12 or STE13 resulted in inducedelastolytic activity. This enzyme has the inhibitor profile of aserine elastase, and its activity appears to require the induc-tion of a tyrosine kinase intracellular signaling pathway.12
Furthermore, similar to neutrophil elastase, this EVE releasespotent mitogenic growth factors, such as basic fibroblastgrowth factor, from the extracellular matrix, thus establishinga mechanistic link between heightened elastase activity andthe SMC proliferative response observed with pulmonaryvascular hypertrophy.14 As a result of elastin degradation, thegeneration of elastin peptides might also stimulate SMCmigration through the induction of fibronectin, linking elas-tase activity with neointimal formation and pulmonary vas-cular disease.15
Further experiments demonstrating that the induction ofSMC elastase activity is inhibited by actinomycin D, as wellas cycloheximide, implied transcription and translation of theelastase gene and/or expression of other gene products relatedto its induction or processing.13 The following investigationused a differential mRNA display to detect transcripts up-regulated coincident with elastase induction. We identifiedone of these transcripts (227 bp) as having homology (90%
Received November 24, 1997; accepted April 15, 1998.From the Division of Cardiovascular Research, Research Institute, The Hospital for Sick Children, and the Departments of Pediatrics and Pathology,
University of Toronto, Toronto, Ontario, Canada.Presented in part at the American Society for Cell Biology, San Francisco, Calif, December 7–11, 1996.Reprint requests to Marlene Rabinovitch, MD, Division of Cardiovascular Research, The Hospital for Sick Children, 555 University Ave, Toronto,
Ontario, Canada M5G 1X8. E-mail [email protected]© 1998 American Heart Association, Inc.
252 by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

over a span of 120 bp) to human AML1 and another as havinga similar 120- to 126-bp region of 88% to 89% homology tohuman lymphotoxin–TNF-a and IL-1a.
AML1 belongs to a family of transcription factors highlyconserved fromDrosophila to human species.16 It was orig-inally identified as the gene altered by the t(8;21) transloca-tion commonly occurring in acute myelogenous leukemia,juxtaposing AML1 with the ETO gene on chromosome 8.17,18
AML1 is a putative transcription factor for the neutrophilelastase gene that contains the appropriate consensus se-quence in its promoter.19 Because of the provocative potentialassociation between the transcription of EVE and AML1, weundertook to investigate its presence and functional signifi-cance in our system. In the present study, we used 59 and 39RACE to confirm the cDNA sequence of the ovine AML1.We documented that there is increased expression of AML1in the nucleus after serum stimulation by using Northern blotand immunocytochemistry. By Western immunoblot, weshowed that AML1 expression is temporally related to EVEactivity. Gel shift analysis was used to document AML1DNA binding activity in SMC nuclear extracts, and antisenseoligonucleotide experiments supported a role for AML1 ininducing EVE activity. The present study provides newinsight into the biological significance of AML1 and theregulation of SMC elastase activity.
Materials and MethodsPA SMC CulturePA SMCs were isolated from the vessels of 100-day-old fetal lambsand from juvenile pigs, as previously described.12 Briefly, the centralpulmonary arteries were removed and rinsed in PBS supplementedwith 300 U/mL penicillin, 300mg/mL streptomycin, and 750 ng/mLamphotericin B (Fungizone, Life Technologies). The endotheliallayer of each artery was removed by scraping the luminal surfacewith a scalpel, and the adventitia was separated from the mediallayer. The media was further dissected into small explants'333 mm in diameter, which showed outgrowth of SMCs after afew days in culture. Cells were cultured in 100-mm dishes usingmedium 199 (Ontario Cancer Institute) with 10% FBS (Intergen) and100 U/mL penicillin, 100mg/mL streptomycin, and 250 ng/mLamphotericin B in a humidified 5% CO2 environment at 37°C. Cellcultures were identified as pure SMCs by uniform immunostainingwith an antibody to smooth musclea-actin and were used forexperimentation at passage 2 or 3.
Measurement of Elastolytic ActivityElastase assays were performed as previously described.12 Briefly,purified elastin from bovine nuchal ligament (Elastin Products Co)
was radiolabeled using [3H]NaBH4 (Dupont/NEN) as described byTakahashi et al.20 [3H]Elastin was reconstituted at 16 mg/mL (spe-cific activity, 2000 cpm/mg elastin) in Tris assay buffer (50 mmol/LTris-HCl, 150 mmol/L NaCl, 10 mmol/L CaCl2, and 0.02% Brij [pH8.0]) and stored at220°C until assay. Before use, [3H]elastin waswashed until the background (nonspecific counts released) was,100 cpm/100mL supernatant. The [3H]elastin suspension was thendiluted to 100 000 cpm/20mL.
To assay both porcine and ovine SMC elastase activity, cells werepassaged into 24-well plates (16-mm diameter) at a density of'105
cells per well. At confluence, cells were washed with PBS and placedin SFM 199 containing 0.1% BSA. After 24 hours of serumstarvation, cells were washed with PBS and incubated with 20mL[3H]elastin (105 cpm) in SFM or with 5% FBS for a further 24-hourperiod. A 600mL aliquot of culture medium was then removed fromeach dish and centrifuged for 5 minutes at 10 000g to pellet insolubleelastin. The resulting supernatant (500mL) was counted to estimatethe release of soluble elastin fragments. Counts resulting from elastindegradation in cell-free wells were subtracted from each data point.All assays were performed in triplicate. Elastase assays wererepeated 3 times with cells from 3 different porcine PA SMCharvests.
Generation of STEPretreating the elastin with FBS has been shown to induce elastinadhesion to SMC surfaces and EVE activity.13 Insoluble elastin wasincubated with FBS at a concentration of 10 mg/mL, rotatedovernight at 37°C, and then washed 4 times with Tris assay buffer toremove all unbound serum factors. The nonradiolabeled STE wasresuspended and incubated with SMCs at a final concentration of 0.5mg/mL, a concentration equal to that used in the elastase assay.
Differential mRNA DisplayDifferential mRNA display was carried out as previously described21
using a kit from Genhunter Corp. Total cellular RNA was preparedby guanidinium and phenol-chloroform extraction22 from 150-mmculture dishes of ovine PA SMCs incubated with either SFM ornonradiolabeled STE for 12 hours after an initial 24-hour period ofserum starvation. RNA samples were treated with DNase for 15minutes at 37°C to remove traces of genomic DNA. Reversetranscription of RNA was performed using poly-T12MN (T12MA,T12MC, T12MG, and T12MT). The resulting cDNA was amplified byPCR in the presence of [35S]dATP and Taq DNA polymerase(Pharmacia) on a Robocycler (Stratagene ) using arbitrary decamersas 59primers and the corresponding T12MN 39 primer. Samples weredenatured at 94°C for 35 seconds, annealed at 40°C for 120 seconds,and extended at 72°C for 35 seconds, for a total of 40 cycles.Radiolabeled PCR products were electrophoresed on a denaturing6% polyacrylamide sequencing gel. Selected bands differentiallyupregulated in response to STE were recovered from sequencinggels, reamplified in a further 40-cycle PCR in the absence of isotope,and electrophoresed on 1.5% low-melting-point agarose gels. PCRproducts were then removed from the gel and purified usingb-agarase 1 (New England Biolabs). Briefly,b-agarase 1 buffer wasadded to each gel fragment, and samples were heated to 70°C for 10minutes until melted. After cooling to 40°C, 1.5 Ub-agarase 1 per100 mL of sample was incubated for 1 hour. Samples were thencooled on ice for 10 minutes and centrifuged at 14 000g for 5minutes to pellet insoluble material. The remaining supernatant wasprecipitated overnight in 2.5 vol ethanol and 0.3 mol/L sodiumacetate, dried, and resuspended in 20mL TE buffer (10 mmol/LTris-Cl [pH 7.6] and 1 mmol/L EDTA [pH 8.0]).
Dot-Blot ScreeningFor dot-blot screening, we used a small aliquot (2mL) of eachindividually amplified PCR product, which was diluted in 100mLH2O, made up to a final concentration of 53 SSC, denatured for 10minutes at 95°C, and then applied to nylon filters (Hybond N,Amersham) using a filtration manifold (Bio-Rad). Both the mem-brane and the underlying filter paper were soaked in 53 SSC for 15
Selected Abbreviations and Acronyms
EMSA 5 electrophoretic mobility shift assayEVE 5 endogenous vascular elastase
IL 5 interleukinPA SMC 5 pulmonary artery SMC
PCR 5 polymerase chain reactionRACE 5 rapid amplification of cDNA ends
SFM 5 serum-free mediumSMC 5 smooth muscle cellSTE 5 serum-treated elastinTNF 5 tumor necrosis factorUTR 5 untranslated region
Wigle et al August 10, 1998 253
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

minutes before assembly of the manifold. Each well was washedonce with 53SSC, and PCR fragments were applied under vacuum.After disassembly of the manifold, membranes were denatured for10 minutes on filter paper soaked in 1.5 mol/L NaCl and 0.5 mol/LNaOH and then neutralized for 5 minutes on filter paper soaked in 1mol/L NaCl and 0.5 mol/L Tris-HCl (pH 7.0). Membranes weredried fully and UV-irradiated to immobilize the DNA.
To check for multiple representation, membranes were screenedwith individually reamplified PCR fragments labeled with [32P]dCTPby random priming (Amersham). Labeling reactions contained 1 mLof the 20-mL reamplification mix. After 1 hour of prehybridization,the PCR fragments were hybridized to the membrane in a solutioncontaining 53 SSC, 50% deionized formamide, 2% SDS, and 100mg/mL salmon sperm DNA. Membranes were stripped for reprobingin 10 mmol/L Tris-HCl and 5 mmol/L EDTA at 65°C for a series offour 30-minute washes.
Cloning and Sequence AnalysisPCR products identified by dot-blot screening as unique sequenceswere again reamplified using the original sample eluted from thedifferential display gel as a template, electrophoresed on 1.5%low-melting-point agarose gels, and purified withb-agarase 1 asdescribed above. Purified PCR products were subcloned into pBSK(Stratagene), by use of TA overhang cloning as previously de-scribed,23 and positive clones were selected by blue/white screening.Insert sequences were obtained using dideoxy sequencing withfluorescein-labeled primers (T7 and T3) on a Pharmacia A.L.F.automated sequencer. Nucleotide sequences were compared withknown sequences by searching GenBank and EMBL databases withBLAST software.24
Northern Blotting and HybridizationTo confirm differential expression of mRNA sequences, Northernblotting was performed using either total RNA (for all nucleotidesequences screened as described above) or purified polyA1 mRNA(for selected sequences as detailed in Results). Total RNA wasprepared according to Chirgwin et al,25 and polyA1 mRNA wasisolated by use of a kit from Invitrogen. Total RNA (20mg) orpolyA1 RNA (5 mg) was electrophoresed on 1% agarose gels andtransferred to nylon membranes (Hybond N) by using standardtechniques.26 Radiolabeling of PCR products or cloned cDNAfragments and hybridization were performed as described above fordot blots. Washes were performed at 50°C (3 times for 20 minuteseach in 13SSC/0.1% SDS followed by 3 times for 20 minutes eachin 0.13SSC/0.1% SDS), and blots were exposed to Kodak X-Omatfilm. RNA loading was assessed by hybridizing the membrane with[32P]dCTP-labeled GAPDH cDNA fragment. Membranes werestripped and reprobed as described above.
mRNA Preparation and Generation of Ovine PASMC AML1 cDNAConfluent PA SMCs in 150-mm culture dishes were serum-starvedfor 24 hours, followed by 12 hours of stimulation with STE. TRIzolReagent (Life Technologies) was used to isolate RNA from theSMCs. The RNA was reverse-transcribed using an adapter-oligo dTprimer, 59-GACTCGAGTCGACATCGAT11-39. This cDNA wasused in 39 RACE with a forward primer, 59-GGTTTCTGTTGTGTTTAATTTC-39, designed from the F12 se-quence. The reverse primer used was a primer to the adapter oligodT, 59-GACTCGAGTCGACATCGA-39. The reaction was as fol-lows: 1 minute at 94°C followed by 39 cycles of denaturation at94°C for 45 seconds, annealing at 55°C for 2 minutes, and extensionat 72°C for 2 minutes, followed by extension at 72°C for anadditional 10 minutes. The resulting PCR products were used in anested PCR using the forward nested primer, 59-ATTTCTCTACAGATTGTATTGT-39, from the F12 sequence. Thereverse primer used was the same as described above. The PCRproduced a single product of'600 bp, which was then subclonedinto pCR2.1 (Invitrogen) plasmid (pSY3) and sequenced on aPharmacia A.L.F. automated DNA sequencer.
In addition, 59 RACE was carried out. A poly dA extension of the59 end of the above reverse-transcribed cDNA was performed usingdATP and terminal deoxynucleotidyl transferase. The PCR reactionused the forward primer 59-GACTCGAGTCGACATCGAT11-39 andthe reverse primer 59-CATATACATATGCTCTACTTCA-39, whichwas from the F12 sequence. The resulting PCR products were usedin a nested PCR using the forward primer to the adapter oligo dT,described above, and a nested reverse primer, 59-ACAATACAATCTGTAGAGAAAT-3 9, which was also from theF12 sequence. A single product of'200 bp was resolved on agarosegel, purified, subcloned, and sequenced as described above.
Northern Blot Using Ovine AML1 cDNAFor Northern blot analyses using the ovine AML1 cDNA, ovinepulmonary artery cells were serum-starved and stimulated with STE,and total RNA was isolated using TRIzol solution. RNA was thenelectrophoresed on formaldehyde agarose gels and transferred toHybond N (Amersham) membranes. Ovine AML1 cDNA 39RACEproduct (a 577-bp sequence) was excised from pSY3 and radiola-beled as described above. Hybridization was performed usingQuikHyb (Stratagene) solution for 1 hour. Membranes were washedtwice for 15 minutes at room temperature with a solution containing23 SSC and 0.1% SDS. High-stringency washes were carried out in0.13 SSC and 0.1% SDS at 60°C.
ImmunostainingFor immunohistochemistry, porcine PA SMCs were plated at 1 to53105 cells/mL onto 20320-mm glass coverslips and grown for 48hours in medium 199 with 10% FBS. Cells were then washed twicein warm PBS, incubated in SFM with 0.1% BSA for 24 hours ofserum starvation, and then either harvested or incubated in medium199 with 10% FBS for 0, 5, 15, and 30 minutes. Cells were washedtwice with warm PBS and then fixed with methanol:acetic acid (2:1)for 5 minutes at220°C. The coverslips were dehydrated and storedat 4°C until use. Rehydration was performed with three 10-minutewashes in PBS and then one 5-minute wash in PBS11% BSA(P-BSA). Blocking was performed for 1 hour at room temperaturewith 10% goat serum in P-BSA, and the primary antibody, apolyclonal antibody produced using a 17-amino-acid N-terminalpeptide of AML1 (N-Arg-Ile-Pro-Val-Asp-Ala-Ser-Thr-Ser-Arg-Arg-Phe-Thr-Pro-Pro-Ser-C, a gift of S. Myers and S. Heibert, St.Jude’s Hospital, Memphis, Tenn) was diluted 1:100 in P-BSA forovernight incubation at 4°C. Coverslips were then washed 3 timesfor 5 minutes each with P-BSA and incubated for 1 hour at roomtemperature with the goat anti-rabbit FITC-conjugated secondaryantibody. One rinse was performed for 5 minutes in P-BSA, followedby two 10-minute washes in PBS. Coverslips were mounted inElvanol and viewed by epifluorescence using appropriate wave-lengths. IgG primary antibody was used as a negative control.
Western ImmunoblottingConfluent monolayers of porcine pulmonary arteries and lamb SMCsin 100-mm dishes were washed twice with SFM, serum-starved for24 hours, and then exposed to STE for varying time periods (0, 1, 5,10, and 30 minutes and 1, 3, 6, and 12 hours). Cells were washedtwice with cold PBS and extracted in hot 2% SDS using a rubberpoliceman. Elastin fragments in the suspension were removed bybrief centrifugation. Samples were normalized for total proteincontent (Bio-Rad), electrophoresed on either 12% or 14% polyacryl-amide Tris-glycine gels, and electroblotted to Immobilon-P mem-branes (Millipore) at 30 V for 1.5 hours. Nonspecific binding wasblocked by incubating the membrane overnight in 5% nonfat milk inPBS with 0.5% Tween 20. This was followed by incubation with thepolyclonal AML1 antibody (1: 200), as described above, and thengoat anti-rabbit secondary antibody (1:5000) (Bio-Rad) and detectedby enhanced chemiluminescence (Amersham) when exposed toKodak X-Omat film. The intensity of the resulting bands wasquantified by scanning soft-laser densitometry. Western immuno-blotting was also carried out to assess expression of AML1 after
254 AML1 and Induction of Vascular Elastase
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

incubation of the control and serum-treated SMCs with antisense,sense, and scrambled oligonucleotides, as described below.
Nuclear Extract PreparationConfluent serum-starved porcine SMCs were treated with either STEor SFM (control) for 30 minutes, and enriched nuclear and cytoplas-mic extracts were prepared as previously described.27 Briefly, afterremoving medium, cells were rinsed in cold PBS and scraped intomicrocentrifuge tubes, and the pellets were washed in PBS. Cellswere lysed in cold hypotonic buffer (10 mmol/L HEPES [pH 7.9],1.5 mmol/L MgCl2, 19 mmol/L KCl, 0.2 mmol/L phenylmethylsul-fonyl fluoride, and 0.5 mmol/L DTT) followed by 10 passes of a typeB Dounce homogenizer (VWR Scientific). Nuclei were pelleted at3300g for 15 minutes at 4°C. The pelleted nuclear proteins werefurther extracted with a high salt solution (20 mmol/L HEPES [pH7.9], 25% glycerol, 1.5 mmol/L MgCl2, 0.8 mol/L NaCl, 0.2 mmol/LEDTA, 0.2 mmol/L phenylmethylsulfonyl fluoride, and 0.5 mmol/LDTT) for 30 minutes and then microcentrifuged at 16 000g for 30minutes. The supernatants were divided into aliquots, rapidly frozenin liquid nitrogen, and stored at270°C. The remaining pellet did notcontain any AML1 immunoreactive proteins, as determined byWestern immunoblotting. The nuclear extracts were then used forWestern immunoblotting with the AML1 antibody or for the EMSA.
Electrophoretic Mobility Shift AssayThe protein-AML1 DNA binding site interaction was analyzed byEMSA according to the protocol described by Meyers et al.16 Thewild-type AML1 binding site double-stranded DNA oligonucleo-tides were prepared by annealing the complementary oligomers59-AATTCGAGTATTGTGGTTAATACG-39 and 59-AATTCG-TATTAACCACAATACTCG-39. The oligomers 59-AATTCGAG-TATTGTTAGTAATACG-39 and 59-AATTCGTATTACTAA-CAATACTCG-39were annealed to form the AML1 mutant bindingsite oligonucleotides. Wild-type and mutant oligonucleotides werelabeled with [a-32P]dATP by use of the Klenow fragment and DNApolymerase. Briefly, the AML1 binding-site reaction contained 10mg total protein from nuclear extracts and 1 ng of radiolabeledoligonucleotides in binding buffer (20 mmol/L HEPES [pH 7.8],1 mmol/L MgCl2, 0.1 mmol/L EGTA, 0.4 mmol/L DTT, 40 mmol/LKCl, 10% glycerol, and 60mg/mL salmon sperm DNA) andproceeded for 30 minutes at room temperature. In competitionstudies, 50 ng of unlabeled double-stranded wild-type or mutantoligonucleotides was preincubated for 30 minutes with the nuclearextracts from STE-treated cells before the addition of the radiola-beled probe. The reactions were resolved on a 5% polyacrylamidenondenaturing gel using TBE buffer (50 mmol/L Tris-borate and1.0 mmol/L EDTA [pH 8.0]), and the gel was dried and exposed toKodak X-Omat film. To confirm that the protein in the nuclearextract was AML1, the nuclear extracts from STE-treated cells werepreincubated with AML1 antibody (1 and 5mL nonimmunopurifiedantisera) before addition of the radiolabeled oligonucleotides. TheAML1 N-terminal peptide was also preincubated with the AML1antibody to further show the specificity of the AML1:DNA gel shiftcomplex.
AML1 Antisense ExperimentsPhosphorothioate oligonucleotides designed for antisense experi-ments were directed to the homologous translation start site of thehuman and mouse AML1 genes and included sense 59-ATGCG-TATCCCCGTAGAT-39, antisense 59-ATCTACGGGGATACG-CAT-39, and scrambled 59-CAGGTCATTGACAGCATG-39 se-quences. The scrambled oligonucleotide contained similar basecomposition but a sequence different from antisense. Oligonucleo-tides were checked in GenBank for potential interference with thetranslation of other mRNA species. For antisense experiments, cellswere plated in 24-well 16-mm dishes and grown until confluence.Cells were then serum-starved for 24 hours in SFM. Initial dose-response experiments using 1, 3, 5, and 10mmol/L oligonucleotideconcentrations determined the 3mmol/L dose to give an appropriateinhibition of elastolytic activity with the addition of the antisense
oligonucleotide. Preincubations of oligonucleotides for 24 hours, 2hours, and 0 hours (addition at time of serum stimulation [0 timepoint]) demonstrated that the latter was sufficient to show the effectof antisense oligonucleotide on elastase induction. Elastase assayswere performed, as described above, after 24 hours of incubationwith oligonucleotides. These experiments were carried out in tripli-cate and repeated with porcine cells from 3 different harvests.
Analysis of DataStatistical analysis was performed on data from elastase assays usinga 1-way ANOVA followed by post hoc comparison of individualgroups using the Fisher protected least significant difference test. Avalue ofP,0.05 was considered statistically significant.
ResultsDifferential Display After STE Induction ofElastolytic ActivityTable 1 summarizes the dot-blot analysis of the 54 differen-tial display fragments identified as potentially upregulatedgenes from a screen of'2000 representative mRNA tran-scripts expressed in PA SMCs after treatment with STE. Atotal of 46 fragments were chosen from the identified 54 fortechnical reasons (eg, retrievability from the filter) andbecause the extent of differential expression was moremarked. Of these 46 fragments, 41 were successfully ream-plified. Only 26 (63.4%) of 41 represented unique sequences.The number of fragments represented in triplicate and dupli-cate is indicated in Table 1. In Figure 1, some of thesequences listed in Table 2 are identified on a representativeportion of the gel to show how they were chosen. Although ithas been suggested that fragments,200 bp are more likely tobe duplicated, we found unique sequences as small as 181 bpand a number of duplicated sequences.200 bp. Twoproducts were lost during the second round of PCR amplifi-cation, ie, after dot blot screening and before subcloning.
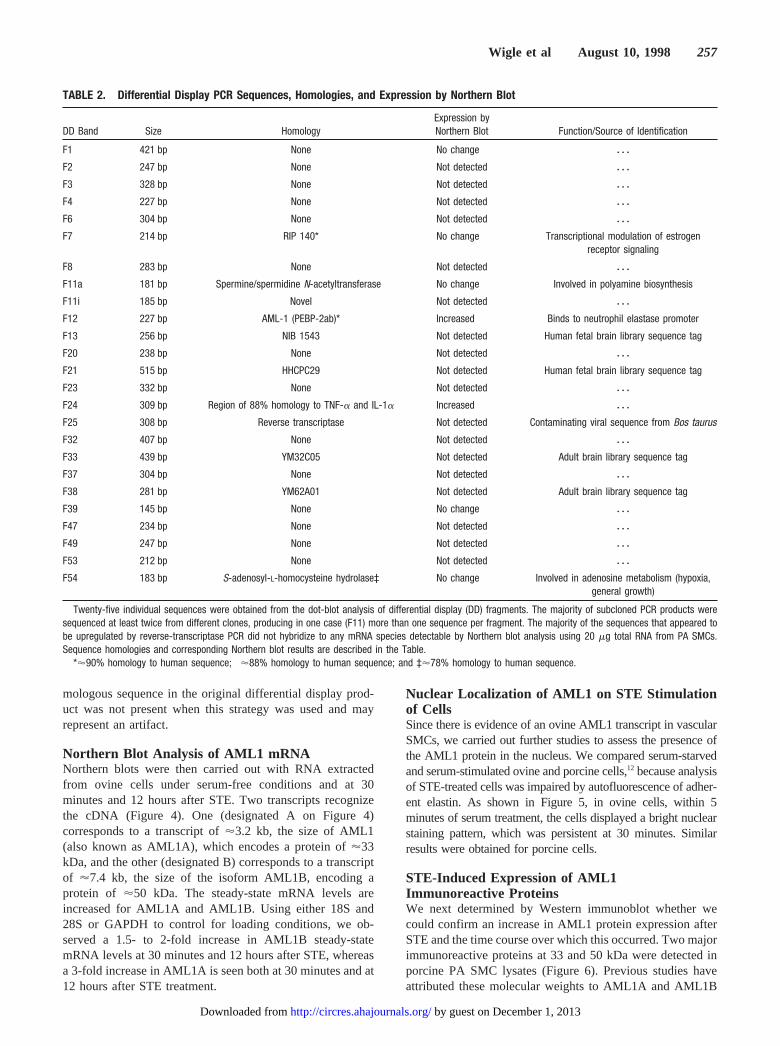
The majority of subcloned PCR products were sequencedat least twice from different clones, producing in one case(F11, Table 2) more than one sequence per fragment. Table 2lists the 25 different sequences obtained and the correspond-ing Northern blot results. As would be expected from an
TABLE 1. Analysis of Differential Display PCR Products
n Percentage
Total fragments identified as differentiallyexpressed
54 z z z
Successful first PCR reamplification 41 z z z
Representation
Individual 15 36.7%
Duplicate 14 34.1%
Triplicate 12 29.3%
Total individual sequences 26 63.4%
Successful second PCR reamplification 24 z z z
Total fragments subcloned andsequenced
24 z z z
Aliquots of individually amplified PCR products differentially expressed wereamplified, applied to nylon filters, and screened for multiple representation byPCR fragments labeled with [32P]dCTP. Individual sequences then underwentsecond PCR reamplification and were subcloned and sequenced as describedin Materials and Methods.
Wigle et al August 10, 1998 255
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

analysis of ovine 39sequences, the majority of genes isolatedshowed no homology to sequences in the GenBank. How-ever, a number of fragments did show homology to sequencetags generated from both human fetal and adult brain cDNAlibraries. The contaminating viral sequence reverse transcrip-tase fromBos taurus(F25) was presumably isolated from theFBS used to make STE. One 309-bp sequence (F24) had aninteresting stretch of homology to the cytokine lymphotoxin–TNF-a (88% homology over 121 bp) and IL-1a (89%homology over 126 bp) (Figure 2). A 71-bp region is 89%homologous to IL-1b. Another 227-bp sequence (F12) had astretch of 123 bp with 90% homology to the transcriptionfactor AML1 (Figure 3A), which is thought to regulatemyeloid differentiation via transcriptional control of genessuch as myeloperoxidase and neutrophil elastase.19 Althoughthere is a DNA recognition sequence for AML1 on theneutrophil elastase promoter, its functional activity vis-a-viscontrol of neutrophil elastase transcription has not beendocumented, nor has AML1 been previously described invascular smooth muscle.
The following 3 gene sequences encoding known proteinswere identified. RIP 140 (F7) is a recently identified nuclearprotein involved in the modulation of transcriptional activa-
tion by the estrogen receptor,28 but it has not previously beendescribed in vascular SMCs.S-Adenosyl-L-homocysteinehydrolase (F54) is involved in adenosine metabolism. It isupregulated with cardiac hypoxia and has been localized tocardiac myocytes.29 Spermine/spermidineN-acetyltransferase(F11a) is an enzyme important in polyamine synthesis. It isincreased before the monocrotaline-induced PA SMC hyper-trophic response.30 The relationship between this enzyme andthe monocrotaline-induced increase in pulmonary artery elas-tase activity, however, is not known.4 Although some of theclones with homology to known genes did not have anidentifiable polyA tail within the insert sequence, all dis-played alignment to the 39UTR of the homologous gene orexpressed sequence tag. The noted exception to this observa-tion was the alignment of F12 to the 59 UTR of the AML1gene, and this will be discussed (Figure 3A).
Northern Blot AnalysisThe majority of the sequences that appeared to be upregulatedby reverse-transcription PCR did not hybridize to any mRNAspecies detectable by Northern blot analysis using 20mg totalRNA from PA SMCs. With other sequence tags that did showhybridization on Northern blot, we could not confirm anappreciable change in mRNA expression after STE treatment.These transcripts were not further pursued. F24, a 309-bpfragment with an interesting region of homology to cytokines,hybridized to a 1.1-kb mRNA species and did appear to beupregulated after STE treatment by'67% (data not shown).Three other sequences identified as having homology toknown genes of potential interest were analyzed further byNorthern blots using polyA1 mRNA (5 mg). The sequenceF7, 90% homologous to human RIP 140 (a 7.5-kb transcriptin the human breast cancer cell line ZR75-1),29 hybridized toan '7-kb transcript on Northern blots of ovine PA SMCRNA but was not upregulated with STE. Two of the othersequences, F11a (with 88% homology to the human spermineN-acetyltransferase species) and F54 (with 78% homologyto the humanS-adenosylhomocysteine hydrolase), wereconfirmed on Northern blot by hybridization to mRNAsof appropriate sizes but, like RIP 140, were not upregulatedwith STE.
AML1 SequenceIt was uncertain whether the sequence tag with homology tohuman AML1 actually encodes this gene product in the ovinespecies. There was only a 123/227-bp stretch of high homol-ogy, and the A-rich region in the 39 end of the transcript wasnot observed in the corresponding human AML1 cDNA.However, the potential functional significance of AML1 inour system led us to pursue studies to document its presencein ovine and porcine SMCs and its relationship to EVEactivity. AML1 has been previously identified only in mam-malian myeloid cells, so studies were carried out to sequencethe ovine AML1 cDNA.
With a strategy using primers derived from the F12sequence and 59 and 39RACE, partial cDNA fragments of211 and 577 bp were obtained. Sequencing of these productsresulted in a total sequence of 767 bp with 70% overallsequence homology to human AML1 (Figure 3). The nonho-
Figure 1. Differential mRNA display of serum-starved (2) andSTE-stimulated (1) cells. Differential mRNA display was carriedout as described in Materials and Methods. F12 and F24(bolded and dark arrows) were identified as upregulated afterSTE treatment and were subsequently isolated and subclonedinto pBSK for further analysis. Other transcripts, either notdetected or not upregulated by STE on Northern blot analysis,are indicated by open arrows, and the numbers relate to thedescription in Table 2.
256 AML1 and Induction of Vascular Elastase
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from
mologous sequence in the original differential display prod-uct was not present when this strategy was used and mayrepresent an artifact.
Northern Blot Analysis of AML1 mRNANorthern blots were then carried out with RNA extractedfrom ovine cells under serum-free conditions and at 30minutes and 12 hours after STE. Two transcripts recognizethe cDNA (Figure 4). One (designated A on Figure 4)corresponds to a transcript of'3.2 kb, the size of AML1(also known as AML1A), which encodes a protein of'33kDa, and the other (designated B) corresponds to a transcriptof '7.4 kb, the size of the isoform AML1B, encoding aprotein of '50 kDa. The steady-state mRNA levels areincreased for AML1A and AML1B. Using either 18S and28S or GAPDH to control for loading conditions, we ob-served a 1.5- to 2-fold increase in AML1B steady-statemRNA levels at 30 minutes and 12 hours after STE, whereasa 3-fold increase in AML1A is seen both at 30 minutes and at12 hours after STE treatment.
Nuclear Localization of AML1 on STE Stimulationof CellsSince there is evidence of an ovine AML1 transcript in vascularSMCs, we carried out further studies to assess the presence ofthe AML1 protein in the nucleus. We compared serum-starvedand serum-stimulated ovine and porcine cells,12 because analysisof STE-treated cells was impaired by autofluorescence of adher-ent elastin. As shown in Figure 5, in ovine cells, within 5minutes of serum treatment, the cells displayed a bright nuclearstaining pattern, which was persistent at 30 minutes. Similarresults were obtained for porcine cells.
STE-Induced Expression of AML1Immunoreactive ProteinsWe next determined by Western immunoblot whether wecould confirm an increase in AML1 protein expression afterSTE and the time course over which this occurred. Two majorimmunoreactive proteins at 33 and 50 kDa were detected inporcine PA SMC lysates (Figure 6). Previous studies haveattributed these molecular weights to AML1A and AML1B
TABLE 2. Differential Display PCR Sequences, Homologies, and Expression by Northern Blot
DD Band Size HomologyExpression byNorthern Blot Function/Source of Identification
F1 421 bp None No change z z z
F2 247 bp None Not detected z z z
F3 328 bp None Not detected z z z
F4 227 bp None Not detected z z z
F6 304 bp None Not detected z z z
F7 214 bp RIP 140* No change Transcriptional modulation of estrogenreceptor signaling
F8 283 bp None Not detected z z z
F11a 181 bp Spermine/spermidine N-acetyltransferase† No change Involved in polyamine biosynthesis
F11i 185 bp Novel Not detected z z z
F12 227 bp AML-1 (PEBP-2ab)* Increased Binds to neutrophil elastase promoter
F13 256 bp NIB 1543 Not detected Human fetal brain library sequence tag
F20 238 bp None Not detected z z z
F21 515 bp HHCPC29 Not detected Human fetal brain library sequence tag
F23 332 bp None Not detected z z z
F24 309 bp Region of 88% homology to TNF-a and IL-1a Increased z z z
F25 308 bp Reverse transcriptase Not detected Contaminating viral sequence from Bos taurus
F32 407 bp None Not detected z z z
F33 439 bp YM32C05 Not detected Adult brain library sequence tag
F37 304 bp None Not detected z z z
F38 281 bp YM62A01 Not detected Adult brain library sequence tag
F39 145 bp None No change z z z
F47 234 bp None Not detected z z z
F49 247 bp None Not detected z z z
F53 212 bp None Not detected z z z
F54 183 bp S-adenosyl-L-homocysteine hydrolase‡ No change Involved in adenosine metabolism (hypoxia,general growth)
Twenty-five individual sequences were obtained from the dot-blot analysis of differential display (DD) fragments. The majority of subcloned PCR products weresequenced at least twice from different clones, producing in one case (F11) more than one sequence per fragment. The majority of the sequences that appeared tobe upregulated by reverse-transcriptase PCR did not hybridize to any mRNA species detectable by Northern blot analysis using 20 mg total RNA from PA SMCs.Sequence homologies and corresponding Northern blot results are described in the Table.
*'90% homology to human sequence; †'88% homology to human sequence; and ‡'78% homology to human sequence.
Wigle et al August 10, 1998 257
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

isoforms, respectively.18 Treatment with STE resulted in arapid and marked elevation of the 33-kDa immunoreactiveprotein between 1 and 5 minutes. This induction lasted up to1 hour, but values were similar to those at baseline by 3 to 6hours. The 50-kDa protein also appeared to be increasedtransiently by STE, although the difference was less than thatobserved with the 33-kDa immunoreactive protein.
The relative amounts of the 2 isoforms and the increasewith STE at 30 minutes corresponds to the mRNA levels. Thepersistent elevation in AML1 mRNA levels for both tran-scripts at 12 hours does not correspond to the drop in proteinalready evident at 6 hours, and this may reflect high turnoveror degradation of the protein or reduced mRNA translationalefficiency.
STE Stimulates the AML1 Consensus DNABinding ActivityElectrophoretic mobility shift assays were therefore carriedout to confirm that AML1 in the nuclear extract of PA SMCs
was of functional significance in that it could form a complexwith its cognate DNA consensus binding sequence. Nuclearextracts from cells treated with SFM (control) or STE for 30minutes were incubated with a radiolabeled 24-bp double-stranded oligonucleotide containing the AML1 DNA bindingsite, as described in Materials and Methods, and run on anondenaturing gel. Figure 7 is an autoradiograph that dem-onstrates at least one slower migrating protein:DNA complex(C2) that is formed in contrast to the free DNA probe. AfterSTE treatment, there is an increase in C2 and an additionalwell-resolved C1 complex formed compared with controlconditions. In competition experiments, a 1-fold (not shown),as well as a 50-fold, increase in unlabeled wild-type oligo-nucleotide, but not the 50-fold mutant oligonucleotide, re-sulted in greater inhibition of complex formation using STEnuclear extracts. In addition, preincubation of increasingconcentrations of AML1 antibody (1 and 5mL of antisera)with the STE-treated nuclear extracts generated a super-shifted band with a corresponding reduction in the previouslyobserved AML1 protein:DNA complexes. The bands werenot shifted with IgG (data not shown). Moreover, preincuba-tion of the AML1 N-terminal peptide (20mL) with theantibody (5mL) before incubation with the nuclear extractand DNA oligonucleotide prevented the supershift. Thesefeatures confirm the specificity of the interaction betweenDNA and AML1 extracted from PA SMC nuclear extracts.
AML1 Antisense Oligonucleotide Inhibition ofElastolytic ActivityTo further determine whether AML1 functioned as a vascularelastase transcription factor, porcine PA SMCs from 3 dif-ferent harvests were treated with AML1 antisense oligonu-cleotides. This attenuated elastase induction by 5% FBSobserved in all 3 experiments implicated the specific involve-ment of AML1 (P,0. 05) (Figure 8A), whereas a scrambledoligonucleotide of similar composition and a sense oligonu-cleotide did not reduce serum induction of elastase activity.Although values for elastase activity appear above basallevels with antisense oligonucleotides, there was no signifi-cant difference in elastase activity. The reduction in serum-induced elastase activity with antisense was accompanied bya comparable reduction on Western immunoblot in the 33kDa, as well as the 50 kDa, AML1 immunoreactive proteins(Figure 8B).
DiscussionWe applied mRNA fingerprinting methodology through dif-ferential display of PCR products in an attempt to identifygenes transcribed in association with production of EVE inPA SMCs. The cells were induced to increase elastaseactivity with STE, with the expectation being that this was amore selective stimulus than serum that also induces elastaseactivity. The objective of these studies was to uncover thevascular elastase mRNA or other mRNA species in whichincreases in steady-state levels were necessary for the tran-scription, translation, processing, and secretion of theenzyme.
Our studies have identified a 227-bp PCR differentialdisplay product that showed homology to the human tran-
Figure 2. Top, Alignment of F24 with TNF-a and IL-1a. F24sequence is aligned to human TNF-a and IL-1a sequences asidentified in GenBank (accession numbers Z14137 and M37210,respectively). The numbers on the right refer to the base pairsidentified in GenBank, and the numbers bolded for F24 refer tothe base pairs sequenced from 59. A 71-bp region of homology(underlined) is also 89% homologous to a sequence in IL-1b.Bottom, Region of homology in the 39 UTR. The bases repre-sented in bold (6881–7001 and 1474–1599) indicate the regionof high homology (120 bp of 88% homology for TNF-a and 126bp of 89% homology for IL-1a).
258 AML1 and Induction of Vascular Elastase
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

scription factor AML1 over a continuous stretch of the120-bp sequence. Since it was likely that the differentialdisplay sequence represented only part of the ovine AML1sequence, we used 59 and 39 RACE to confirm that we couldidentify an ovine AML1 homologue in our cells. Having doneso, we next showed increased expression of AML1A andAML1B mRNA transcript levels in our cells at 30 minutesand 12 hours after STE stimulation. We next addressedwhether expression of AML1 protein correlated with induc-tion of elastase after STE. We showed a transient increase inexpression of AML1 protein by immunohistochemistry usingserum stimulation and by Western immunoblot using STEinduction. The protein levels appeared to correspond to themRNA levels for the AML1 transcripts at 30 minutes butwere reduced by 6 hours. It is possible that AML1 protein israpidly degraded or that the mRNA is inefficiently translatedafter 1 hour. AML1 in nuclear extracts from PA SMCs after30-minute stimulation with STE compared with nonstimu-lated cells showed increased binding complex formation, withits DNA consensus sequence identified on the neutrophilelastase promoter. Using antisense oligonucleotides, we dem-onstrated that inhibition of AML1 translation reduced thesubsequent induction of elastase activity after serum or STEin a manner that correlated with decreased expression of theprotein by Western immunoblot.
A potentially novel gene (F24) with homology to the 39UTR of cytokines that is upregulated (as confirmed byNorthern blot analysis) after stimulation of elastase activitywas also identified (data not shown). A second sequence tagencoded a gene product, RIP-140, involved in the modulationof transcriptional activation by the estrogen receptor. Al-though not upregulated with STE, it was not previouslyknown to be expressed by SMCs, so this observation mayprove to be important in the study of estrogens and vascularpathobiology. Two other sequence tags encoded interestingtranscripts:S-adenosylhomocysteine hydrolase and spermineN-acetyltransferase. Both may play a role in the smoothmuscle proliferation and differentiation associated with pul-monary hypertension and induction of elastase activity, butneither could be confirmed as upregulated with STE onNorthern blot.
The technique of differential mRNA display has rapidlygained acceptance as a powerful tool for the identificationand cloning of differentially expressed genes in a variety ofbiological systems. The method offers a number of technical
Figure 3. Alignment of F12 and ovine AML1 with human AML1.A, F12, a 227-bp sequence, is aligned with the sequence ofhuman AML1. A 90% homology is observed over 120 bp. Thebase-pair numbers on the far right indicate the sequences iden-tified in GenBank (accession number X79549). The A-richsequences in the 39 region of the transcript indicate why thissequence in the 59 UTR was generated by a strategy that isintended to produce sequences upstream from the polyA tail. B,Sequence homology between human AML1 and the 767-bpsequence of ovine AML1 generated by 59 and 39 RACE usingthe strategy described in Materials and Methods is shown. TheF12 sequence homology to human AML1 is underlined. Thestart of 39 and 59 RACE nested primers is shown by asterisks.Sequence homology over this region is 70%. C, The place inthe 59 UTR where homology was noted is indicated.
Wigle et al August 10, 1998 259
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

advantages over other existing techniques for the investiga-tion of differential gene expression. It is fast and relativelyeasy to perform and does not require large initial quantities onRNA. Because even semiquantification of PCR productsrequires stringent internal standards to control for primerattachment and amplification efficiency, the technique will
lead to the sequencing of a large number of “false-positives”in that the increase in mRNA level cannot be confirmed byNorthern blot. Despite this, differential mRNA display hasproven useful when applied to a myriad of investigations,including those directed at the cardiovascular system. Forexample, the system was successfully used to identifyestrogen-regulated genes in human vascular SMCs,31 glucose-induced genes in bovine aortic SMCs,32 and genes expressedafter allograft cardiac transplantation in the rat33 and afterexperimental carotid artery balloon angioplasty.34 We nowreport 2 differentially displayed sequences that appeared to beupregulated by Northern blot in conjunction with increasedelastolytic activity. The first of these genes, F24, contained arather large and interesting region of homology to cytokines.It is possible that this sequence may represent a novelcytokine or, alternatively, an ovine cytokine sequence.
The second differential display fragment revealed a strik-ing stretch of 120-bp homology with human AML1. Thefragment amplified by PCR was, however, not a 39 sequence(as predicted by the experimental design) but a 59 UTRsequence. It was therefore essential to sequence ovine AML1to establish its precise relationship to the transcript identifiedby differential display. Perhaps only fortuitous homology toAML1, a transcription factor known to bind to the neutrophilelastase promoter in immature myeloid cells, led to theinvestigation of AML1 in our system.
The AML1 family of transcription factors is highly homol-ogous to theDrosophila segmentation generunt.35 Thishomology suggests that AML1 is likely to be of significancein the differentiation of a variety of cell types. Two humangenes have been isolated to date, including AML1A andAML1B.18 By interacting with a b component, thesea
Figure 4. STE upregulates ovine AML1 mRNAs. Ovine pulmonary artery SMCs were serum-starved and treated with STE, and totalRNA was isolated at 0 minutes, 30 minutes, and 12 hours. Electrophoresis, transfer, and hybridization were performed as described inMaterials and Methods. Left, top, Two mRNA species hybridized to ovine AML1 cDNA. A and B indicate transcripts of '3.2 and '7.4kb, respectively. Left, middle, RNA loading assessed by reprobing the blot with GAPDH. Left, bottom, Ethidium bromide–stained gelshowing 28S and 18S ribosomal RNAs. Right, Densitometric values obtained from the autoradiograms and normalized to the 0 timepoint.
Figure 5. AML1 nuclear immunostaining. Serum-starved SMCs(A and B) were treated with FBS-supplemented medium for 30minutes (C to F). Double immunofluorescent staining for DAPI(A, C, and E) and AML1 (B and D) was carried out as describedin Materials and Methods. In serum-free conditions, AML1 wasdetected in the cytoplasm and perinuclear region (B), whereasaddition of serum promoted its accumulation in the nucleus (D).No immunostaining of SMCs was observed with control IgG (F).Bar510 mmol/L.
260 AML1 and Induction of Vascular Elastase
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

proteins enhance their ability to bind DNA without changingDNA contact points.36,37 The AML1 mutation occurring inacute myelogenous leukemia, involving the t(8;21) translo-cation juxtaposing AML1 with the ETO gene on chromosome8 (Reference 18), generates a chimeric protein product. Thisproduct retains the known DNA binding activity of AML1 tothe enhancer motif TGTGGT and the region of homology totheDrosophilasegmentation generunt but loses its potentialto transactivate.
There was evidence of increased AML1 in the nucleusafter serum stimulation, and we have shown that MAP kinasekinase activity manifest by phosphorylation of extracellularsignal–regulated kinase-1 is necessary for both increasedexpression of AML1 and induction of elastase activity.38 Todemonstrate the functional activity of AML1, we used oligo-nucleotides encoding the consensus sequence in the neutro-phil elastase promoter in gel mobility shift assays. Since we
have not yet cloned the gene for EVE (the PA SMC serineelastase), we can only speculate that the same promoterelement is present. Although the antisense experimentsshowed a reduction in the level of elastase activity and thusfurther support AML1 as being of functional significance inits induction, it is also possible that AML1 is the transcriptionfactor for other genes required for the processing of thisenzyme. The levels of enzymatic activity observed withantisense were still higher than those under control condi-tions, although the difference was not statistically significant.This is reflected in the lower protein level on Westernimmunoblot and suggests incomplete suppression of mRNAtranslation. This is likely a function of incomplete oligonu-cleotide incorporation or stability. This represents the limita-tion of these experiments, since higher concentrations ofoligonucleotides could be expected to result in cellulartoxicity.
Figure 6. STE regulates the expression of AML1.Serum-starved SMCs were treated with STE (0 to6 hours). Cell lysates were analyzed by SDS-PAGEand immunoblotting with AML1 antibody. Top, Im-munoblot showing densitometric analysis of the33-kDa protein below from 1 of 2 different experi-ments with similar results. PA SMCs express 2major AML1 immunoreactive protein bands at 33and 50 kDa. Molecular weights (MW) from stan-dards are indicated on the left. Bottom, Increase inexpression of the 33-kDa AML1 immunoreactiveprotein appeared within 5 minutes and returned tobaseline by 3 to 6 hours after STE treatment. How-ever, the expression of the 50-kDa immunoreactiveprotein (likely AML1B) also appears to have beeninduced but to a lesser extent than the 33-kDaprotein in STE-treated cells.
Figure 7. Expression of AML1 DNA bindingsite proteins stimulated with STE. Serum-starved SMCs were treated with STE for 30minutes. Nuclear extracts were isolated, andDNA binding to the AML1 consensus site wasassessed in an electrophoretic mobility shiftassay. A radiolabeled wild-type DNA 24-bpoligonucleotide incorporating the AML1 bind-ing site as described in the text was incu-bated with nuclear extracts and analyzed ona 5% polyacrylamide nondenaturing gel. Incompetition experiments to confirm the spec-ificity of the protein:DNA interaction, nuclearextracts from control-treated SMCs preincu-bated with unlabeled DNA containing thewild-type 1-fold (not shown) or 50-foldexcess (wt oligo), but not the mutant AML1-binding site (50-fold excess mut oligo), inhib-ited complex formation. After addition ofAML1 antibody into the binding reaction withSTE-treated nuclear extracts, there was asupershift of the protein:DNA complex rela-tive to STE. This is more apparent withincreasing concentrations of antibody anddisappears when the antibody is preincu-bated with the AML1 N-terminal peptide.
When IgG was substituted for the AML1 antibody, there was no shifted band (not shown). This is a representative gel of 3 inde-pendent experiments with similar results.
Wigle et al August 10, 1998 261
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

It would be of further interest to determine whether thereare other genes upregulated by AML1 in vascular tissueeither in development or in disease states. The fact that micelacking either AML1 or its correspondingb subunit of theAML1-CBF b complex die with massive hemorrhage into thecentral nervous system, peritoneal cavity, and pleural spacemay imply a developmental vascular abnormality in additionto the hematopoietic defects noted.39,40
In summary, the present study used combined Westernimmunoblot and immunohistochemistry to elucidate the nu-
clear localization of AML1 and to establish a temporalassociation between induction of AML1 protein and STEstimulation of PA SMC elastase activity. We also show thatantisense oligonucleotide inhibition of AML1 mRNA trans-lation reduces the STE induction of elastase activity. Thepresent study provides novel insight into the biologicalactivity of a transcription factor hitherto primarily investi-gated in the context of the genetics of myeloid cell differen-tiation and leukemic reactions. It also provides new informa-tion about the regulation of the recently identified vascularelastase, important in vascular development andpathobiology.
AcknowledgmentsThis study was supported by grant T724 from the Heart and StrokeFoundation of Ontario. Dr Thompson is a Fellow and Dr Rabinovitchis a Career Investigator of the Heart and Stroke Foundationof Ontario.
References1. Rabinovitch M, Bothwell T, Hayakawa BN, Williams WG, Trusler GA,
Rowe RD, Olley PM, Cutz E. Pulmonary artery endothelial abnormalitiesin patients with congenital heart defects and pulmonary hypertension: acorrelation of light with scanning electron microscopy and transmissionelectron microscopy.Lab Invest. 1986;55:632–653.
2. Todorovich-Hunter L, Johnson DJ, Ranger P, Keeley FW, RabinovitchM. Altered elastin and collagen synthesis associated with progressivepulmonary hypertension induced by monocrotaline: a biochemical andultrastructural study.Lab Invest. 1988;58:184–195.
3. Maruyama K, Ye C, Woo M, Venkatacharya H, Lines LD, Silver MM,Rabinovitch M. Chronic hypoxic pulmonary hypertension in rats andincreased elastolytic activity.Am J Physiol. 1991;261:H1716–H1726.
4. Todorovich-Hunter L, Dodo H, Ye C, McCready L, Keeley FW,Rabinovitch M. Increased pulmonary artery elastolytic activity inadult rats with monocrotaline-induced progressive hypertensive pul-monary vascular disease compared with infant rats with nonpro-gressive disease.Am Rev Respir Dis. 1992;146:213–223.
5. Ilkiw R, Todorovitch-Hunter L, Maruyama K, Shin J, Rabinovitch M.SC-39026, a serine elastase inhibitor, prevents muscularization of periph-eral arteries, suggesting a mechanism of monocrotaline-induced pulmo-nary hypertension in rats.Circ Res. 1989;64:814–825.
6. Ye C, Rabinovitch M. Inhibition of elastolysis by SC-37698 reducesdevelopment and progression of monocrotaline pulmonary hypertension.Am J Physiol. 1991;261:H1255–H1267.
7. Zhu L, Wigle D, Hinek A, Kobayashi J, Ye C, Zuker M, Dodo H, KeeleyFW, Rabinovitch M. The EVE that governs development and progressionof monocrotaline-induced pulmonary hypertension in rats is a novelenzyme related to the serine proteinase adipsin.J Clin Invest. 1994;94:1163–1171.
8. Leake DS, Hornebeck W, Brechemier D, Rogert L, Peters TJ. Propertiesand subcellular localization of elastase-like activities in arterial smoothmuscle cell in culture.Biochem Biophys Acta. 1983;761:41–47.
9. Hornebeck W, Brechemier D, Soleilhac JM, Bourdillon MC, Robert L.Studies on rat aorta smooth muscle cells’ elastase activity. In: Reddi AH,ed.Extracellular Matrix: Structure and Function. New York, NY: AlanR Liss; 1985;25:269–282.
10. Rosenberg HC, Rabinovitch M. Endothelial injury and vascular reactivityin monocrotaline pulmonary hypertension.Am J Physiol. 1988;255:H1484–H1491.
11. Munro JM, Cotran RS. The pathogenesis of atherosclerosis: atherogenesisand inflammation.Lab Invest. 1988;58:249–261.
12. Kobayashi J, Wigle D, Childs T, Zhu L, Keeley FW, Rabinovitch M.Serum-induced vascular smooth muscle cell elastolytic activity throughtyrosine kinase intracellular signalling.J Cell Physiol. 1994;160:121–131.
13. Thompson K, Kobayashi J, Childs T, Wigle D, Rabinovitch M. Endo-thelial and serum factors which include apolipoprotein A1 tether elastin tosmooth muscle cells inducing serine elastase activity via tyrosine kinase-mediated transcription and translation.J Cell Physiol. 1998;174:78–89.
Figure 8. Antisense oligonucleotide directed against AML1mRNA inhibit serum induction of elastase activity. PorcineSMCs were serum-starved for 24 hours in SFM, at which pointFBS (5%) and phosphorothioate oligonucleotides were added.Elastase assays were performed 24 hours after serum stimula-tion. S, AS, and SCR represent sense, antisense, and scrambledoligonucleotides, respectively. A, The AML1 antisense oligonu-cleotides attenuated the serum induction of elastase activity(P,0.05 vs C, SCR, or S). There was no significant differencebetween basal values in SFM and AS oligonucleotides15%FBS. S as well as SCR oligonucleotides did not reduce theserum induction of elastolytic activity. Experiments wererepeated with cells from 3 different harvests. Bar values repre-sent mean6SEM. B, top, Representative Western blot of theantisense experiment showing proteins that were immunoreac-tive to anti-AML1 antibodies. MW indicates molecular weight. B,bottom, Relative densitometry of the 33-kDa protein after 1-hourtreatment of serum and AML1 oligonucleotides to the SMCsafter 24-hour serum starvation.
262 AML1 and Induction of Vascular Elastase
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from

14. Thompson K, Rabinovitch M. Exogenous leukocyte and endogenouselastases can mediate mitogenic activity in pulmonary artery smoothmuscle cells by release of extracellular matrix-bound basic fibroblastgrowth factor.J Cell Physiol. 1996;166:495–505.
15. Hinek A, Molossi S, Rabinovitch M. Functional interplay betweeninterleukin-1 receptor and elastin binding protein controls fibronectinsynthesis in coronary artery smooth muscle cells.Exp Cell Res. 1996;225:122–131.
16. Meyers S, Downing JR, Hiebert SW. Identification of AML-1 and the(8;21) translocating protein (AML-1ETO) as sequence-specific DNA-binding proteins: therunt homology domain is required for DNA bindingand protein-protein interactions.Mol Cell Biol. 1993;13:6336–6345.
17. Miyoshi H, Kozu T, Shimizu K, Enomoto K, Maseki N, Kaneko Y,Kamada N, Ohki M. The t(8;21) translocation in acute myeloid leukemiaresults in production of an AML1-MTG8 fusion transcript.EMBO J.1993;12:2715–2721.
18. Meyers S, Lenny N, Hiebert SW. The t(8;21) fusion protein interfereswith AML-1B-dependent transcriptional activation.Mol Cell Biol. 1995;15:1974–1982.
19. Nuchprayoon I, Meyers S, Scott LM, Suzow J, Hiebert SW, FriedmanAD. AML1/CBF, altered by chromosomal breaks in myeloid leukemiacells, regulates the murine myeloperoxidase and neutrophil elastase genesin immature myeloid cells.Mol Cell Biol. 1994;14:5558–5568.
20. Takahashi S, Seifter S, Yang FC. A new radioactive assay for enzymeswith elastolytic activity using reduced tritiated elastin: the effect ofsodium dodecylsulfate on elastolysis.Biochim Biophys Acta. 1973;327:138–145.
21. Liang P, Pardee AB. Differential display of eukaryotic messenger RNAby means of the polymerase chain reaction.Science. 1992;257:967–970.
22. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acidguanidinium thiocyanate-phenol-chloroform extraction.Anal Biochem.1987;162:156–159.
23. Marchuk D, Drumm M, Saulino A, Collins A. Construction of T-vectors,a rapid and general system for direct cloning of unmodified PCR prod-ucts.Nucleic Acids Res. 1991;19:1154–1154.
24. Altschul SF, Gish W, Miller W, Meyers EW, Lipman DJ. Basic localalignment search tool.J Mol Biol. 1990;215:403–410.
25. Chirgwin JM, Przybyla AE, MacDonald RJ, Rutter WJ. Isolation ofbiologically active ribonucleic acid from sources enriched in ribonu-clease.Biochemistry. 1979;18:5294–5299.
26. Sambrook J, Fritsch EF, Maniatis T.Molecular Cloning: A LaboratoryManual. 2nd ed. Cold Spring Harbor, NY: Cold Spring Harbor Labo-ratory Press; 1989.
27. Dignam JD, Lebovits RM, Roeder RG. Accurate transcription initiationby RNA polymerase II in a soluble extract from isolated mammaliannuclei.Nucleic Acids Res. 1983;11:1475–1487.
28. Cavailles V, Dauvois S, L’Horset F, Lopez G, Hoare S, Kushner PJ,Parker MG. Nuclear factor RIP140 modulates transcriptional activationby the estrogen receptor.EMBO J. 1995;14:3741–3751.
29. Lloyd HG, Schrader J. Adenosine metabolism in the guinea pig heart: therole of cytosolic S-adenosyl-L-homocysteine hydrolase, 59-nucleotidaseand adenosine kinase.Eur Heart J. 1993;14(suppl I):27–33.
30. Orlinska U, Olson JW, Gebb SA, Gillespie MN. Acetylated polyaminesin lungs from rats with monocrotaline-induced pneumotoxicity.FundamAppl Toxicol. 1989;13:277–284.
31. Koike H, Karas RH, Baur WE, O’Donnell TF, Mendelson ME.Differential-display polymerase chain reaction identifies nucleophosminas an estrogen-regulated gene in human vascular smooth muscle cells.JVasc Surg. 1996;23:477–482.
32. Nishio Y, Aiello LP, King GL. Glucose induced genes in bovine aorticsmooth muscle cells identified by mRNA differential display.FASEB J.1994;8:103–106.
33. Utans U, Liang P, Wyner LR, Karnovsky MJ, Russell ME. Chroniccardiac rejection: identification of five upregulated genes in transplantedhearts by differential display.Proc Natl Acad Sci U S A. 1994;91:6463–6467.
34. Autieri MV, Feuerstein GZ, Yue TL, Ohlstein EH, Douglas SA. Use ofdifferential display to identify differentially expressed mRNAs inducedby rat carotid artery balloon angioplasty.Lab Invest. 1995;72:656–661.
35. Levanon D, Negreanu V, Bernstein Y, Bar-Ami I, Groner Y. AML1,AML2, and AML3, the human members of the “runt domain” genefamily: cDNA structure, expression and chromosomal localization.Genomics. 1994;23:425–432.
36. Ogawa E, Inuzuka M, Maruyama M, Satake M, Naito-Fijimoto M, Ito Y,Shigesada K. Molecular cloning and characterization of PEBP2b, theheterodimeric partner of a noveldrosophila runt-related DNA bindingprotein PEBP2a.Virology. 1993;194:314–331.
37. Wang S, Wang Q, Crute BE, Melnikova IN, Keller SR, Speck NA.Cloning and characterization of subunits of the T-cell receptor and murineleukemia virus enhancer core-binding factor.EMBO J. 1993;13:3324–3339.
38. Thompson K, Rabinovitch M. Induction of vascular elastase is mediatedby integrins and is associated with phosphorylation of focal adhesionkinase and activation of the transcription factor AML-1 [abstract].MolBiol Cell. 1996;7(suppl):588A.
39. Okuda T, van Deursen J, Hiebert SW, Gorsveld G, Downing JR. AML1,the target of multiple chromosomal translocations in human leukemia, isessential for normal fetal liver hematopoiesis.Cell. 1996;84:321–330.
40. Wang Q, Stacy T, Miller JD, Lewis AF, Gu TL, Huang X, Bushweller JH,Bories JC, Alt FW, Ryan G, Liu PP, Wynshaw-Boris A, Binder M,Marın-Padilla M, Sharpe AH, Speck NA. The CBFbeta subunit isessential for CBFalpha2 (AML1) function in vivo.Cell. 1996;87:697–708.
Wigle et al August 10, 1998 263
by guest on December 1, 2013http://circres.ahajournals.org/Downloaded from
Copyright © 2022 FDOKUMEN




















