
Neutrophils Induce Sequential Focal Changes in Endothelial Adherens Junction Components: Role of...
16
Neutrophils Induce Sequential Focal Changes in Endothelial Adherens Junction Components: Role of Elastase CARMEN V. IONESCU,* GEDIMINAS CEPINSKAS,* JURATE SAVICKIENE,* MARTIN SANDIG, † AND PETER R. KVIETYS* *Vascular Biology/Inflammation Program, Lawson Health Research Institute, London, ON, Canada; and the † Department of Anatomy and Cell Biology, University of Western Ontario, London, ON, Canada ABSTRACT Objective: In vitro studies have indicated that polymorphonuclear leukocytes (PMNs) traverse endothelial cell monolayers via the paracellular pathway (i.e., through endothelial cell–cell junctions. Herein, we assessed whether the adherens junctions (AJs) are disrupted during PMN transendothelial cell migration. Methods: Human umbilical vein endothelial cells (HUVECs) were grown to confluence on porous membranes and activated with interleukin-1, and PMN transendothelial migration was facilitated by formyl-methionyl-leucyl- phenylalanine. Using dual immunofluorescence staining and laser scanning con- focal microscopy, we assessed the effects of PMN–endothelial cell adhesive in- teractions (i.e., adhesion to and emigration across monolayers) on the AJ com- ponents vascular endothelial (VE)-cadherin, -catenin, -catenin, and -catenin. Results: In the AJ immediately adjacent to the adherent PMN, there was a loss of staining for some of the AJ components. AJ components further away from HUVEC–PMN adhesive interactions were unaffected. An iterative approach in- dicated that the four components were sequentially lost from the AJ. -catenin was lost first, followed by VE-cadherin, -catenin, and, finally, -catenin. In the absence of PMNs, the cross-linking of VE-cadherin, but not platelet endothelial cell adhesion molecule-1 or intercellular adhesion molecule-1, increased the cy- toplasmic accumulation of -catenin. During PMN transendothelial migration, all of the junctional components under study were lost at the immediate site of monolayer penetration. Again, at regions removed from the actual site of PMN penetration of the monolayers, the AJ components were unaffected. PMN-induced disorganization of the AJs was partially prevented by an elastase inhibitor. Conclusions: These findings suggest that adherent PMNs induce a localized, sequential disassembly of AJs, which is partially mediated by PMN-derived elas- tase and involves the initial loss of an intracellular component of AJs (i.e., -catenin). Microcirculation (2003) 10, 205–220. doi:10.1038/sj.mn.7800185 KEY WORDS: -catenin, -catenin, -catenin, inflammation, platelet endothelial cell adhesion molecule-1, transendothelial migration, vascular endothelial-cadherin INTRODUCTION Polymorphonuclear leukocyte (PMN) rolling and subsequent firm adhesion to the endothelium are a prerequisite for PMN transendothelial migration (51,16,6,18,38). A great deal of information has been generated on these initial adhesive interac- tions, and there is a general consensus on the mo- lecular determinants of PMN rolling and adhesion Supported by the Canadian Institutes of Health Research grants MOP-37758 (P.R.K.), MOP-44010 (G.C) and Heart and Stroke Foundation of Ontario grant NA-3181 (M.S.). For reprints of this article, contact Peter R. Kvietys, Vascular Biology/Inflammation Program, Lawson Health Research Insti- tute, 375 South Street, Room C210, London, ON, Canada N6A 4G5; e-mail: [email protected] Received 12 April 2002; accepted 11 November 2002 Microcirculation (2003) 10, 205–220 © 2003 Nature Publishing Group 1073-9688/03 $25.00 www.nature.com/mn
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Neutrophils Induce Sequential Focal Changes in Endothelial Adherens Junction Components: Role of...

Neutrophils Induce Sequential FocalChanges in Endothelial Adherens Junction
Components: Role of ElastaseCARMEN V. IONESCU,* GEDIMINAS CEPINSKAS,*
JURATE SAVICKIENE,* MARTIN SANDIG,† AND PETER R. KVIETYS**Vascular Biology/Inflammation Program, Lawson Health Research Institute,
London, ON, Canada; and the †Department of Anatomy and Cell Biology,University of Western Ontario, London, ON, Canada
ABSTRACT
Objective: In vitro studies have indicated that polymorphonuclear leukocytes(PMNs) traverse endothelial cell monolayers via the paracellular pathway (i.e.,through endothelial cell–cell junctions. Herein, we assessed whether the adherensjunctions (AJs) are disrupted during PMN transendothelial cell migration.Methods: Human umbilical vein endothelial cells (HUVECs) were grown toconfluence on porous membranes and activated with interleukin-1�, and PMNtransendothelial migration was facilitated by formyl-methionyl-leucyl-phenylalanine. Using dual immunofluorescence staining and laser scanning con-focal microscopy, we assessed the effects of PMN–endothelial cell adhesive in-teractions (i.e., adhesion to and emigration across monolayers) on the AJ com-ponents vascular endothelial (VE)-cadherin, �-catenin, �-catenin, and�-catenin.Results: In the AJ immediately adjacent to the adherent PMN, there was a lossof staining for some of the AJ components. AJ components further away fromHUVEC–PMN adhesive interactions were unaffected. An iterative approach in-dicated that the four components were sequentially lost from the AJ. �-cateninwas lost first, followed by VE-cadherin, �-catenin, and, finally, �-catenin. In theabsence of PMNs, the cross-linking of VE-cadherin, but not platelet endothelialcell adhesion molecule-1 or intercellular adhesion molecule-1, increased the cy-toplasmic accumulation of �-catenin. During PMN transendothelial migration,all of the junctional components under study were lost at the immediate site ofmonolayer penetration. Again, at regions removed from the actual site of PMNpenetration of the monolayers, the AJ components were unaffected. PMN-induceddisorganization of the AJs was partially prevented by an elastase inhibitor.Conclusions: These findings suggest that adherent PMNs induce a localized,sequential disassembly of AJs, which is partially mediated by PMN-derived elas-tase and involves the initial loss of an intracellular component of AJs (i.e.,�-catenin).Microcirculation (2003) 10, 205–220. doi:10.1038/sj.mn.7800185
KEY WORDS: �-catenin, �-catenin, �-catenin, inflammation, platelet endothelialcell adhesion molecule-1, transendothelial migration, vascular endothelial-cadherin
INTRODUCTION
Polymorphonuclear leukocyte (PMN) rolling andsubsequent firm adhesion to the endothelium are aprerequisite for PMN transendothelial migration(51,16,6,18,38). A great deal of information hasbeen generated on these initial adhesive interac-tions, and there is a general consensus on the mo-lecular determinants of PMN rolling and adhesion
Supported by the Canadian Institutes of Health Research grantsMOP-37758 (P.R.K.), MOP-44010 (G.C) and Heart and StrokeFoundation of Ontario grant NA-3181 (M.S.).For reprints of this article, contact Peter R. Kvietys, VascularBiology/Inflammation Program, Lawson Health Research Insti-tute, 375 South Street, Room C210, London, ON, Canada N6A4G5; e-mail: [email protected] 12 April 2002; accepted 11 November 2002
Microcirculation (2003) 10, 205–220© 2003 Nature Publishing Group 1073-9688/03 $25.00www.nature.com/mn

(21,30,31,44,47). By contrast, the mechanisms bywhich PMNs traverse the endothelial barrier to gainaccess to the interstitium of inflamed tissue are lesswell defined, and to some extent are still controver-sial. For example, there is some disagreement re-garding the pathway used by PMNs to traverse theendothelial barrier (33). In general, in vivo studieshave indicated that PMNs preferentially use thetranscellular pathway (through endothelial cells)(14), whereas in vitro studies have indicated thatPMNs preferentially use the paracellular pathway(between endothelial cells) (17,38,4). However,there are exceptions to this paradigm, and, at pres-ent, this issue remains unresolved (33). Another con-tentious point is whether PMNs use proteases topenetrate the endothelial lining. Although some evi-dence suggests that PMN transendothelial migrationis not dependent on proteases (15,27), the bulk ofthe evidence indicates that proteases, particularlyPMN-derived elastase, play a critical role in PMNemigration (5,9,8,7,32,52). Recent studies indicatethat PMN do not secrete elastase into the extracel-lular milieu, rather they mobilize elastase to theplasma membrane and localize it to the migratingfront (i.e., pseudopodia) (9).
Our working hypothesis is that PMNs use mem-brane-bound elastase to pass between endothelialcells during diapedesis. This process would requirethe disruption of adhesion junctions holding endo-thelial cells together. There are two adhesion junc-tions that represent relevant barriers to PMN diape-desis: adherens junctions (AJs) and tight junctions(25,11,42). Of these, the AJs have received the mostattention. A typical AJ consists of vascular endothe-lial (VE)-cadheren/catenin complexes (36). The ex-tracellular domains of VE-cadherins interact homo-typically with VE-cadherins on adjacent endothelialcells (39). The intracellular domains are associatedwith either �-catenin or �-catenin. The �-cateninand �-catenin are linked to �-catenin, which con-nects the entire VE-cadherin/catenin complex to theactin cytoskeleton (48). Several lines of evidence in-dicate that these junctions play an important role inPMN diapedesis. Antibodies to VE-cadherin en-hanced PMN migration to an inflammatory stimulus(peritonitis) in vivo (20). Antibodies to VE-cadherinalso can cause endothelial cell retraction and canenhance PMN transendothelial migration in vitro(25). However, efforts to assess AJ integrity directlyduring PMN–endothelial cell adhesive interactionsusing immunofluorescence and biochemical ap-proaches have been hampered by the artifactual re-lease of proteases from PMNs during sample prepa-
ration. (12,41). In addition, PMNs induce changes inAJs rapidly, and these events do not occur simulta-neously within the cell population. Thus, to date,this issue remains largely unresolved.
The major objective of the present study was to as-sess whether PMN–endothelial cell adhesive interac-tions result in alterations in the AJ under conditionsthat would avoid artifactual proteolysis. To this end,we used laser scanning confocal microscopy (LSCM)to assess AJ integrity (immunofluorescence analy-sis). As opposed to biochemical analyses, this ap-proach allowed us to study the AJ components at thesingle-cell level. Our findings are the first to indicatethat when PMNs adhere to human umbilical veinendothelial cell (HUVEC) monolayers grown on per-meable membranes, staining for the various compo-nents of AJs is reduced or lost in the area adjacent toPMN–endothelial cell contact. Staining of the AJcomponents remained continuous at the endothelialcell borders that were not in contact with PMNs.Furthermore, this focal absence of staining in closeproximity to PMN–endothelial cell adhesive interac-tions seemed to be an organized event involving se-quential changes in each of the components of theAJ. �-catenin staining was lost first, followed by VE-cadherin staining, �-catenin staining, and finally,�-catenin staining. The initial loss of an intracellularcomponent (�-catenin) seems to involve PMN-derived elastase and outside-in signaling via VE-cadherin.
MATERIALS AND METHODS
Endothelial Cells
HUVECs were harvested from umbilical cords bycollagenase treatment, as previously described (53).The cell cultures were incubated in complete M199media at 37 °C in a humidified atmosphere with 5%CO2 and were serially passaged by brief trypsiniza-tion (0.25% trypsin in phosphate-buffered saline(PBS) containing 0.02% ethylenediaminetetraaceticacid). HUVECs in the first through second passagewere used in experiments.
Neutrophils
Human neutrophilic PMNs were isolated from thevenous blood of healthy adults using standard dex-tran sedimentation and gradient separation on His-topaque 1077 (Sigma Chemical Co., St. Louis, MO)(53). This procedure yields a PMN population that is95–98% viable (trypan blue exclusion) and 98%pure (acetic acid-crystal violet staining).
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
206

Polymorphonuclear Leukocyte–EndothelialCell Interactions
HUVECs were seeded at high density onto fibronec-tin (25 �g/mL)-coated Falcon cell culture inserts(3-�m diameter pores) and were used when conflu-ent. The HUVEC monolayers were pretreated for 4hours with 10 ng/mL recombinant human interleu-kin (IL)-1� (R&D Systems, Minneapolis, MN) inendothelial growth medium. Subsequently, PMNin M199 were added to the apical aspect of HUVECmonolayers at a PMN/endothelial cell ratio of 10:1,and they were coincubated for 10–20 minutes in thepresence of formyl-methionyl-leucyl-phenylalanine(fMLP; 10−8 M) in the basal compartment. This timeframe was chosen to allow for the capture of PMNsat different stages of interaction with HUVECs (i.e.,adhesion and emigration). In some experiments,PMNs were cocultured with HUVECs in the presenceof an elastase inhibitor, L658,758 (100 �M) (MerckSharp & Dohme Research Laboratory, Hoddesdon,UK). In other experiments, HUVEC monolayersalone, in the absence of PMNs, were treated withpurified neutrophilic elastase (0.5 �g/mL) (Dako,Inc., Carpinteria, CA) for 20 minutes in the absenceor presence of elastase inhibitor L658 758 (100 �M).Subsequently, the inserts were removed and were pro-cessed for either immunofluorescence by LSCM (seebelow) or for the quantitation of PMN transendothelialmigration (as described previously) (9).
Immunofluorescence Analysis of AdherensJunction Proteins
The HUVEC/PMN cocultures were fixed at roomtemperature for 10 minutes in 2% paraformalde-hyde in PBS and were prepared for immunofluores-cence analysis using a previously described immu-nostaining protocol (9,46), with some modificationsto prevent artifactual proteolysis during sample pro-cessing. Fixed HUVEC/PMN cocultures were perme-abilized for 5 minutes with a buffer (pH 7.4) con-taining 15 mM Tris-HCl, 120 mM NaCl, 2.5 mMethylenediaminetetraacetic acid, 2 mM ethylene gly-col tetraacetic acid, 0.5% Triton X-100, 0.1 mMdithiothreitol, and the following protease inhibitors:2 mM agents 4-(2-aminoethyl)benzenesulfonyl-fluoride hydrochloride; 1 mM E-64; and 10 �g/mLeach of pepstatin A, bestatin, leupeptin, and aproti-nin (Sigma Chemical Co.). The samples were thenincubated for 1 hour at room temperature with twoof the following primary antibodies: mouse anti-human VE-cadherin monoclonal antibody, cloneBV6 (Chemicon International Inc., Temecula, CA);goat antihuman VE-cadherin polyclonal antibody
(Santa-Cruz Biotechnology Inc., Santa Cruz, CA);mouse antihuman �-catenin monoclonal antibody,clone 15B8 (Sigma Chemical Co.); rabbit antihuman�-catenin polyclonal antibody (Sigma ChemicalCo.); rabbit antihuman �-catenin polyclonal anti-body (Sigma Chemical Co.); and mouse antihuman�-catenin monoclonal antibody, clone 15F11 (SigmaChemical Co.). The antibodies were prepared in PBScontaining 1% bovine serum albumin (BSA) at aconcentration of 1:100 for anti-VE-cadherin and 1:1000 for the other antibodies. Combinations of fluo-rescently labeled secondary antibodies then wereused at a dilution of 1:300 in PBS containing 1%BSA Alexa Fluor 588 goat antimouse immunoglob-ulin (Ig) G (H+L) conjugated (Molecular Probes,Eugene, OR) and fluorescein isothiocyanate goat an-tirabbit (Invitrogen, Carlsbad, CA), Alexa Fluor 488goat antimouse IgG (H+L) conjugate (MolecularProbes) and Alexa Fluor 568 goat anti-antirabbitIgG (H+L) conjugate (Molecular Probes). Betweenprocedures, the samples were washed in PBS threetimes. Nuclei were labeled with 10 nM Bisbenzimide(Hoechst 33342, Sigma Chemical Co.) in PBS.Samples were mounted in Vectashield (Vector Labo-ratories, Burlingame, CA) on microscope slides, andthe preparations were analyzed by LSCM.
Laser Scanning Confocal Microscopy
Serial optical sections (0.7 �m in thickness) ofsamples were taken in a z direction using a ZeissLSM 410 LSCM (Carl Zeiss Canada Ltd., Don Mills,ON, Canada) equipped with the appropriate epifluo-rescence filters and external krypton-argon laserthat emits excitation wavelengths at 488, 568 and647 nm and an external UV laser that emits excita-tion wavelengths at 351 and 364 nm (Carl ZeissCanada LDT). To prevent “cross-talk” between dif-ferent channels, a sequential scanning approach wasapplied using the software package Zeiss LSM 3.92beta (Carl Zeiss Canada Ltd.). The images presentedare representative of 7–10 experiments and at least10 fields examined per sample.
Immunoblot Analysis
HUVECs were grown to confluence in 35 mm Petridishes and were treated in a similar manner as forthe transendothelial migration studies (i.e., IL-1�for 4 hours followed by fMLP). Subsequently,HUVEC monolayers were treated with purified hu-man neutrophilic elastase (0.5 �g/mL, Dako Inc.)for 20 minutes, were washed in cold PBS, and cyto-plasmic proteins were extracted for 5 minutes in a
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
207

lysis buffer (50 mM Tris-HCl, 140 mM NaCl, 1.6mM MgCl2, 1% Igepal CA-630 (Sigma, St. Louis,MO), and 0.1% Triton X-100) containing the fol-lowing proteinase inhibitors: 2 mM agents 4-(2-aminoethyl)benzenesulfonylfluoride hydrochloride; 1mM E-64; and 10 �g/mL each of pepstatin A, be-statin, leupeptin, and aprotinin. The extracted cyto-plasmic proteins were electrophoresed on 7%sodium dodecyl sulfate polyacrylamide gel electro-phoresis and were transferred to reinforced nitro-cellulose membranes (Schleicher & Schuell, Inc.,Keene, NH). Blots were incubated overnight in ablocking solution (3% BSA and 0.05% Tween-20 inPBS). Subsequently, the membranes were treatedwith mouse antihuman �-catenin monoclonal anti-body (clone 15B8, Sigma), and monoclonal antibodybinding was detected using biotinylated antimouseIgG and Vectastain Elite ABC peroxidase detectionsystem (Vector Laboratories Inc.).
Functional Blocking of Vascular Endothelial–Cadherin Homotypic Adhesion
HUVECs were grown to confluence on fibronectin-coated coverslips. After treatment with 10 ng/mLrecombinant human Il-1� (Pierce Endogen, Rock-ford, IL) for 4 hours, the HUVEC monolayers weretreated with fMLP (10–8 M) and 100 �g/mL rabbitpolyclonal antihuman VE-cadherin (Cedarlane,Hornby, ON, Canada) or mouse monoclonal antihu-man VE-cadherin, clone BV6 (Chemicon Interna-tional Inc.) for 10 minutes. The polyclonal antibodyinterferes with VE-cadherin homotypic adhesion(reversible endothelial cell retraction) (23). Thesamples were processed for immunofluorescence,stained for �-catenin and nuclei, and analyzed byLSCM. In another set of experiments, we assessedthe effects of cross-linking platelet endothelial celladhesion molecule (PECAM)-1 or intercellular ad-hesion molecule (ICAM)-1 on �-catenin accumula-tion in the cytoplasm of HUVECs. PECAM-1 andICAM-1 are HUVEC adhesion molecules that havebeen implicated in PMN–endothelial cell adhesiveinteractions (10,42). In these experiments, HUVECmonolayers were treated with medium alone or withmedium containing mouse monoclonal antihumanPECAM-1 (hec7, gift from Dr. W. Muller) or mousemonoclonal antihuman ICAM-1 clone BBIG41(R&D Systems). These treatments were followed byincubation with Alexa Fluor 488 goat antimouse IgG(H+L) conjugate (Molecular Probes) for 30 minutes.These approaches have been shown previously toelicit ligand-mediated intracellular signaling(3,13,24). The samples then were processed for im-
munohystochemistry, stained for �-catenin and nu-clei, and analyzed by LSCM.
RESULTS
Local Loss of Staining of AJ Components DuringPMN–Endothelial Cell Adhesive Interactions
In accord with our working hypothesis, PMN migra-tion across HUVEC monolayers occurred predomi-nantly via the paracellular pathway. Although about5% of the PMNs were captured migrating through athin portion of the endothelial cell, 95% of the PMNscaptured in the migration process were passing be-tween endothelial cells (data not shown). The majorbarriers encountered by PMNs using the paracellularpathway are the junctional complexes: AJs and tightjunctions. In the present study, we focused our at-tent ion on the effects of PMN adhesion/transendothelial migration on AJs; specifically, withrespect to VE-cadherin, �-catenin, �-catenin, and�-catenin.
The effects of PMN–endothelial cell interactions(i.e., adhesion to and migration through endothelialcell junctions) on the AJ were assessed using an im-munofluorescence protocol and LSCM. This ap-proach allowed for the assessment of changes in thestaining of various components of the AJ at thesingle-cell level. The staining patterns for VE-cadherin, �-catenin, �-catenin, and �-catenin werevisualized in IL-1�-challenged HUVEC monolayersin the absence and presence of PMNs. In the absenceof PMNs, VE-cadherin, �-catenin, �-catenin, and�-catenin staining was localized to the endothelialcell junctions (Fig. 1). The staining was continuous(i.e., there were no discernable gaps noted in thestaining for any of these AJ components). By con-trast, when PMNs were allowed to interact with andmigrate across the IL-1�-challenged HUVEC mono-layers, discontinuities in the staining for the adher-ence junction components were observed (Fig. 2).The staining patterns for VE-cadherin, �-catenin,�-catenin, and �-catenin were interrupted in thoseareas of the endothelial cell junctions with which thePMNs were interacting (i.e., PMN adhesion/transendothelial migration) (Fig. 2, arrows). Stain-ing for the individual components of the AJ re-mained continuous in endothelial cells that werenot directly impacted by PMN–endothelial cellinteractions (Fig. 2, arrowheads). Taken together,these observations suggest that PMN adhesion/transendothelial migration results in the local dis-ruption or disorganization of AJ complexes.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
208

Polymorphonuclear Leukocyte Adhesion toEndothelial Cell Junctions Results in DiscontinuousImmunostaining for �-Catenin, but not VascularEndothelial-Cadherin in the Vicinity ofAdhesion Site
In this set of experiments, we focused our attentionon the early events that occur in the process of PMNtransendothelial migration (i.e., PMN adhesive in-teractions with endothelial cell junctions). Two com-ponents of the AJ (VE-cadherin and �-catenin) andnuclei were labeled for immunofluorescence analy-sis. Serial optical sections were taken by LSCM ofPMNs adherent to IL-1�-stimulated HUVEC mono-layers that had been exposed to an fMLP gradient(i.e., fMLP added to the basal aspect of monolayers).Serial sectioning allowed us to determine accuratelythe exact position of PMNs with respect to HUVECs.Sequential analyses of different channels were per-formed to control cross-talk between the fluorescent
probes. Two populations of adherent PMNs werecaptured: adherent PMNs that were associated withno alterations in the staining for VE-cadherin and�-catenin (85%); and adherent PMNs that were as-sociated with a loss of staining for �-catenin (15%).As shown in Fig. 3, VE-cadherin and �-catenin (redand green fluorescence, respectively) were colocal-ized at the borders of endothelial cells, as indicatedby the yellow fluorescence at these sites. As was ap-parent from the optical section taken at the level ofthe interendothelial junctions located directly under-neath the adherent PMNs (Fig. 3b), the colocaliza-tion of the staining for VE-cadherin and �-cateninwas continuous. In contrast, Fig. 4 shows an ex-ample of an adherent PMN under which there was acontinuous immunostaining for VE-cadherin and an
Figure 2. PMN transendothelial migration induces focaldiscontinuities in the distribution of the AJ components inHUVECs. HUVEC monolayers were stimulated with IL-1� for 4 hours before coincubating with PMNs. PMNswere allowed to migrate for 20 minutes in the presence ofa chemotactic gradient (fMLP). The samples were fixed,permeabilized, and double-immunostained for two AJcomponents. VE-cadherin (red fluorescence) and�-catenin (green fluorescence) are shown in (a) and (b),respectively. �-catenin (red fluorescence) and �-catenin(green fluorescence) are shown in (c) and (d), respec-tively. The images were made using separate channels foreach color, and are shown in black and white. The stain-ing of each of the AJ components was no longer visible inthe interendothelial junctions adjacent to adherent/migrating PMN (arrows). In contrast, AJs located at sitesfurther removed from interacting PMNs retained continu-ous staining for VE-cadherin, �-catenin, �-catenin, and�-catenin (arrowheads). Bars represent 10 �m.
Figure 1. The distribution of AJ components in IL-1�-stimulated HUVECs. HUVEC monolayers grown on per-meable membranes were stimulated with IL-1� for 4hours, and subsequently fMLP was added to the basalaspect of the membranes. Twenty minutes later, thesamples were fixed, permeabilized, and immunostainedfor the AJ components (as described in detail in MATE-RIALS AND METHODS). Nuclei were labeled withHoechst (blue-fluorescence). Optical sections were takenby LSCM (red fluorescence channel) through HUVECmonolayers immunostained for VE-cadherin, �-catenin,�-catenin, and �-catenin. Images are presented in blackand white. All four of the AJ components were localized tothe interendothelial junctions. There were no obvious dis-continuities in the staining for any of the AJ components(arrows). Bars represent 10 �m.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
209

interruption of the staining for �-catenin (Fig. 4b,arrow). Again, there was a continuous colocalizedstaining for VE-cadherin and �-catenin at interen-dothelial sites distant from the adherent PMNs (Fig.4c, arrowhead).
Polymorphonuclear Leukocyte TransendothelialMigration Results in Discontinuous Immunostainingfor Both Vascular Endothelial-Cadherinand �-Catenin
Serial sectioning using LSCM allowed for the cap-ture of PMNs in the process of migration betweenendothelial cells (Fig. 5). PMNs migrating across IL-�-challenged HUVEC monolayers were identified bythe simultaneous presence of the PMN nucleus inthree distinct regions of the monolayer: a portionabove the monolayer (Fig. 5a), a portion betweenadjacent endothelial cells (Fig. 5b), and a portionunderneath the monolayer (Fig. 5c). In the immedi-ate vicinity of the transmigrating PMNs there wasdiscontinuous staining for both VE-cadherin and�-catenin (Fig. 5b). However, continuous colocal-ized staining for VE-cadherin and �-catenin wasnoted at sites further removed from the transmigrat-ing PMNs (Fig. 5b, arrowhead).
Staining of Adherens Junction Components IsAltered Sequentially During PolymorphonuclearLeukocyte–Endothelial Cell Adhesive Interactions
An identical approach to that described above forVE-cadherin and �-catenin was used to assesschanges in the continuity of the immunostaining oftwo different colocalized components of AJs. Repre-sentative experiments are shown in Fig. 6, and theoverall results are summarized in Table 1. Althoughserial optical sectioning was always used to deter-
<
Figure 3. PMN adhesion to HUVECs does not affect thestaining of VE-cadherin or �-catenin. The experimentalconditions were the same as those described in Fig. 2. Thesamples were fixed, permeabilized, and double-immunostained for VE-cadherin and �-catenin (red andgreen fluorescence, respectively). Nuclei were labeled withHoechst (blue-fluorescence). Panels a and b show two op-tical sections of a series taken by LSCM in an apical (a) tobasal (b) direction (z-axis), through cocultures of adher-ent PMNs/HUVECs. The focal plane for each panel isindicated in the diagram below the images. (a) A PMN(arrow) is shown attached to the HUVEC monolayerabove an interendothelial junction. (b) The interendothe-lial junction (arrow) is shown underneath the same PMNs.The yellow fluorescence indicates colocalization of VE-cadherin (red) and �-catenin (green) staining at junctionsbetween endothelial cells. There were no detectablechanges in the staining of VE-cadherin or �-catenin, ei-ther in the interendothelial junction located adjacent toadherent PMN (arrow in b) or in areas more removed fromadherent PMNs (arrowhead in b). Bar represent 10 �m.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
210

mine whether a PMN was adherent or was in theprocess of emigration, only the images obtained atthe level of the interendothelial junctions are shownin Fig. 6. Fig. 6a and a� depict two different adher-ent PMNs under which the colocalized staining for(1) �-catenin and �-catenin and (2) �-catenin andVE-cadherin was continuous. Fig. 6b and b� depict
adherent PMNs under and adjacent to which therewere discontinuities in the colocalized immunostain-ing for one of the pair of adhesion junction compo-nents under study. The extent of the discontinuousstaining varied but was always limited to the endo-thelial cells interacting with the PMNs. As shown inFig. 6b, �-catenin staining (red) was continuous,whereas �-catenin staining (green) was not. Asshown in Fig. 6b�, �-catenin staining (red) was con-tinuous, whereas VE-cadherin staining (green) wasmarkedly reduced. Fig. 6c and c� depict the impactthat PMNs in the process of migration acrossHUVEC monolayers have on the staining of the dif-ferent pairs of AJ components studied. As shown, inthe immediate vicinity of transmigrating PMNs therewere discontinuities in the staining of all the com-ponents examined. As in the previous experiments,the colocalized staining for these pairs of AJ compo-nents remained continuous at sites further removedfrom PMN–endothelial cell interactions.
Additional experiments using the dual-immuno-fluorescence approach revealed that staining of theother AJ components was selective at the PMN–endothelial junction interface. Specifically, (1)�-catenin staining was absent when �-catenin stain-ing was present, (2) VE-cadherin staining was ab-
<
Figure 4. PMN adhesion to HUVECs results in discon-tinuous immunostaining of �-catenin, whereas VE-cadherin staining remains continuous. The experimentalconditions were the same as those described in Fig. 2. Thesamples were fixed, permeabilized, and double-immunostained for VE-cadherin and �-catenin (red andgreen fluorescence, respectively). Nuclei were labeled withHoechst (blue-fluorescence). Serial optical sections of thecoculture of adherent PMNs/HUVECs were taken byLSCM. The images show the same optical section taken atthe level of the interendothelial junctions located imme-diately beneath the adherent PMN (open arrow in c). Thefocal plane is indicated in the diagram below the images.(a) and (b) represent individual channels (a, red; b, green)in black and white, whereas (c) represents a color com-posite of the two. There was significant loss of �-cateninstaining (arrow in b), but not of VE-cadherin (arrow in a)at the interendothelial junctions located underneath andimmediately adjacent to the adherent PMNs. As shown in(c; arrow), there is a localized loss of staining for�-catenin (green fluorescence), but not VE-cadherin (redfluorescence). In addition, there were no changes in VE-cadherin/�-catenin colocalization at the junctions locatedmore distant from the adherent PMNs (arrowhead in c), asindicated by yellow fluorescence (colocalization of the redand green fluorescence). Bar represent 10 �m.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
211

sent when �-catenin staining was present, and (3)�-catenin staining was absent when �-catenin stain-ing was present (data not shown). The results of allthese experiments are summarized in Table 1. Fromthese studies, a general pattern of the effects of PMNadhesion to and migration across HUVECs emerges.Most of the PMNs interacting with HUVECs can becharacterized as follows: (1) PMNs can be capturedadherent to HUVECs without any noticeable alter-ations in the staining for the components studied; or(2) PMNs captured in the process of transendothelialmigration were always associated with the disrup-tion of the staining for all components of the AJsstudied. However, a small population of adherentPMNs (15%) were captured that were associatedwith the loss of one of the two components studied.An iterative approach using dual immunostainingfor two different components of the AJs revealed thatadherent PMNs induce a specific sequential loss ofstaining of the four components of the AJs studied(i.e., �-catenin staining is affected first, followed byVE-cadherin, �-catenin, and, finally, �-cateninstaining).
Role of Elastase in PolymorphonuclearLeukocyte–Endothelial Cell Adhesive Interactions
PMN-derived elastase plays an important role inPMN transendothelial migration (8,9). In the pres-ent study, an elastase inhibitor (L658,758) reducedthe fMLP-induced PMN transendothelial migrationby about 70% (Fig. 7). The few PMNs that werecaptured migrating across HUVEC monolayers byLSCM were always associated with focal disruptionsin the staining for each of the four components of theAJ (data not shown). The fact that PMNs still wereable to migrate in the presence of the elastase inhibi-tor is not entirely surprising. We have shown previ-ously that PMN transendothelial migration can beelastase-independent. In this situation, PMNtransendothelial migration can be prevented by en-dogenous antiproteases (8). Others have shown (50)that in mice deficient in both neutrophilic elastaseand cathepsin G there is still normal recruitment ofPMNs to the site of infection. Taken together, theseobservations indicate that PMNs have redundantmechanisms to ensure appropriate migration to sitesof inflammation, a process that is critical to the sur-vival of the host.
Although PMN–endothelial cell adhesive interac-tions still occurred in the presence of the elastaseinhibitor (albeit to a lesser extent), it was not pos-sible to analyze the VE-cadherin/catenin complexat the discreet locations of PMN interaction with
Figure 5. PMN migration through interendothelial junc-tions alters VE-cadherin and �-catenin immunostaining.The experimental conditions were the same as those de-scribed in Fig. 2. The samples were fixed, permeabilized,and double-immunostained for VE-cadherin and�-catenin (red and green fluorescence, respectively). Nu-clei were labeled with Hoechst (blue-fluorescence). Panelsa, b, and c are optical sections of a series taken by LSCMin an apical (a) to basal (c) direction (z-axis) throughtransmigrating PMNs. The focal plane for each panel isindicated in the diagram below the images. The portion ofa transmigrating PMN that has not penetrated the junction isindicated by the arrow in (a). The arrow in (c) shows theportion of the same PMN that is located underneath theHUVEC monolayer and is entering the membrane pore. In(b), which was taken at the level of the interendothelial junc-tion, note the absence of both VE-cadherin and �-cateninimmunostaining (arrow). VE-cadherin/�-catenin colocal-ization remained continuous at junctions located more dis-tant from the transmigrating PMNs (arrowhead in b), asindicated by the yellow color (colocalization of the red andgreen fluorescence). Bar represent 10 �m.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
212

HUVECs using immunoblot analysis. Thus, we re-sorted to another approach that involved the ampli-fication of the PMN-induced effects on componentsof the AJ by the addition of a greater number ofPMNs to the HUVEC monolayers (PMN/endothelialcell ratio, 30:1). As shown in Fig. 8, when the PMNswere induced to interact (both adhesion andtransendothelial migration) with the IL-1�-challenged HUVECs by the addition of fMLP, therewere widespread discontinuities in the staining forVE-cadherin (Fig. 8a), �-catenin (Fig. 8b),�-catenin (Fig. 8c), and �-catenin (Fig. 8d). Inthe presence of the elastase inhibitor, thesePMN-induced discontinuities in staining were less
prominent for each of the AJ components studied(Fig. 8a� to d�). In addition, in some experimentspunctate staining for �-catenin in the cytoplasm ofHUVECs was noted (Fig. 8b and b�).
Purified Elastase Increases Endothelial CellCytoplasmic Content of �-Catenin
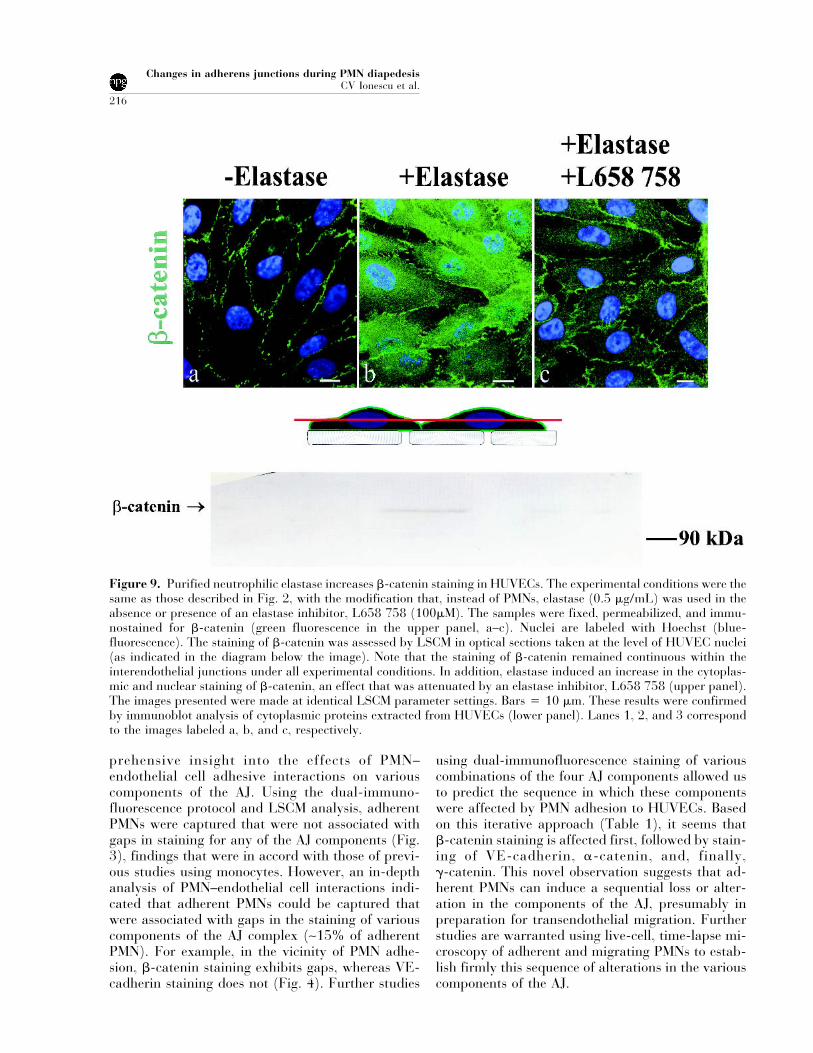
Because �-catenin staining was the first to be lostfrom the junctions and initial experiments withPMNs indicated that �-catenin was present in thecytoplasm (Fig. 8b), we examined the latter phe-nomenon in greater detail. In these experiments, pu-rified human neutrophil elastase (0.5 �g/mL) was
Figure 6. The effects of PMN adhesion and emigration on �-catenin, �-catenin, and VE-cadherin. The experimentalconditions were the same as those described in Fig. 2. The samples were fixed, permeabilized, and double-immunostained for �-catenin and �-catenin (a–c) or �-catenin and VE-cadherin (a�–c�) (red and green fluorescencerespectively for each pair). Nuclei were labeled with Hoechst (blue-fluorescence). Each panel represents an opticalsection through different PMN/HUVEC cocultures that was taken at the level of the interendothelial junctions (indi-cated by the red line in the diagram below the images). In (a) and (a�) note that the junctions adjacent to adherentPMNs (opened arrow) show colocalization of �-catenin/�-catenin and VE-cadherin/�-catenin (arrows). Panels b andb� show the lack of staining of one of the AJ components (arrows) in junctions underneath and in the immediate vicinityof the adherent PMNs (opened arrow). (b) �-Catenin staining is present, but �-catenin staining is absent. (b�)�-Catenin staining is present, but VE-cadherin staining is absent. Panels c and c� show the lack of staining for both AJcomponents (arrows) adjacent to the transmigrating neutrophils (open arrows). Bars represent 10 �m.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
213

added to the endothelial cell monolayers for 20 min-utes, and the appearance of �-catenin in the endo-thelial cytoplasm was assessed by LSCM and immu-noblots. The monolayers were processed for LSCMbefore the appearance of any structural changes inthe monolayers (i.e., endothelial cell retraction) asjudged by light microscopy. Elastase induced an in-crease in cytoplasmic and nuclear staining for�-catenin (Fig. 9, a and b in the upper panel). Thiswas confirmed by immunoblot analysis of cytoplas-mic proteins extracted from HUVECs (Fig. 9, com-pare lanes 1 and 2 in the lower panel). The elastaseinhibitor (L658 758) minimized the extent of cyto-plasmic �-catenin staining induced by elastase (Fig.9c and lane 3 in the bottom panel).
Antibodies Directed Against VascularEndothelial-Cadherin Induce CytoplasmicAccumulation of �-Catenin
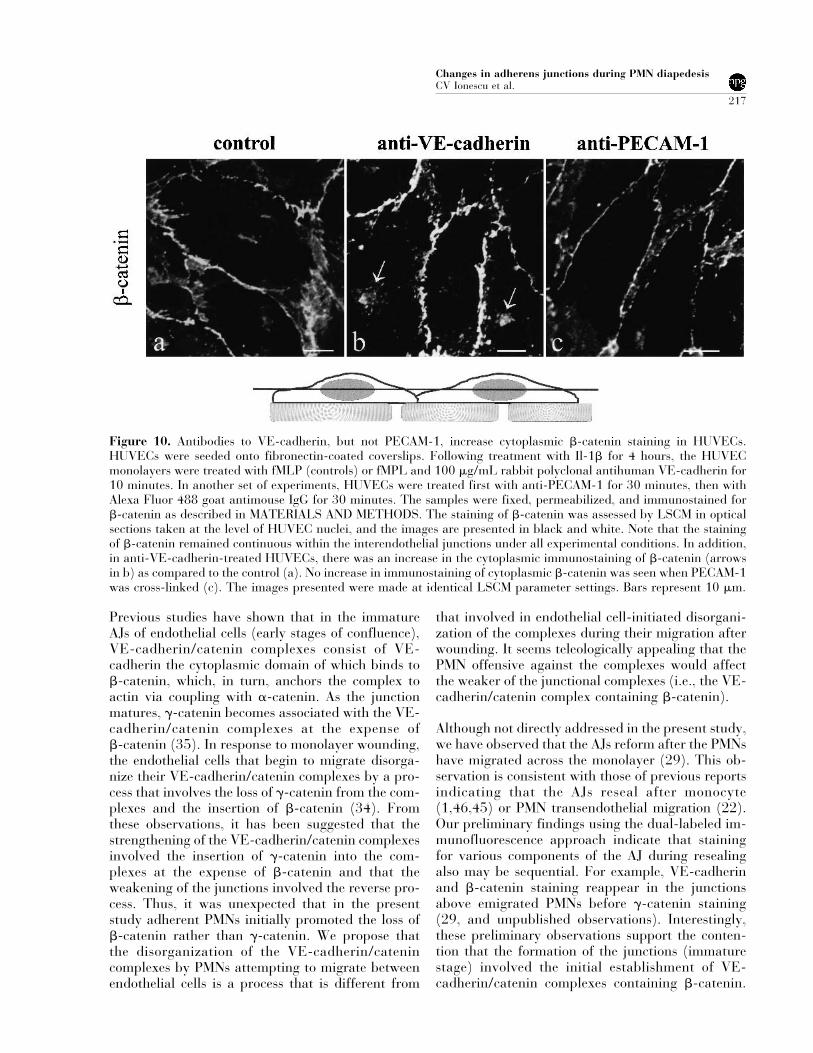
PMN transendothelial migration has been shown tobe dependent on both ICAM-1 and PECAM-1 onendothelial cells (10,42). Cross-linking of these ad-hesion molecules also activates endothelial intracel-lular signaling pathways (3,13,24). The observa-tions that (1) immunostaining for an intracellularcomponent of the AJ (�-catenin) was the first to belost after PMN adhesion to HUVECs (Table 1) and(2) elastase increased cytoplasmic staining for�-catenin suggest that outside-in signaling was in-
duced by the adherent PMNs. However, crosslinkingof PECAM-1 did not result in an increase in cyto-plasmic staining for �-catenin (Fig. 10). Similar re-sults were observed for crosslinking ICAM-1 (datanot shown). By contrast, an antibody directedagainst VE-cadherin induced an increase in cyto-plasmic staining for �-catenin within 10 minutes(Fig. 10).
DISCUSSION
One of the cardinal features of acute inflammation isPMN emigration into the affected tissue. The initialbarrier to PMN emigration is the endothelial liningof the microvasculature. The mechanism(s) bywhich PMNs breach the endothelial barrier is notcompletely understood. In the present study, we fo-cused our attention on the AJ, a major barrier toPMN paracellular transendothelial migration. Tothis end, we used a reductionist approach to assessthe effects of PMN adhesion to and migration acrossHUVEC monolayers that were grown to confluenceon porous filters. Using this model and LSCM, westudied the distribution of interendothelial immuno-fluorescence staining of VE-cadherin, �-catenin,�-catenin, and �-catenin during PMN adhesive in-
Table 1. Effects of PMN adhesion to and migration acrossHUVEC on adherens junction components
VE-cadherin
�-catenin
�-catenin
�-catenin
Adherent 1 + + + +Adherent 2 + −
− +− +
− +− +
+ −Transmigrating − − − −
Dual-immunofluorescence labeling was used to study alterations inthe staining of two different components of adherens junctions. (+)indicates presence of staining, whereas (−) indicates the absence ofstaining. Two populations of adherent PMNs were captured byLSCM: those that were not associated with alterations in the stainingof AJ components (adherent 1:85%) and those that were associatedwith alterations in staining of one of the two components studied(adherent 2:15%). This iterative approach allowed us to predict thesequence in which the different components were affected by adher-ent PMN: �-catenin staining was lost first, followed by VE-cadherinstaining, �-catenin staining, and, finally, �-catenin staining. All ofthe PMNs captured in the process of transendothelial migration wereassociated with the loss of staining for all AJ components studied.
Figure 7. An elastase inhibitor (L658 758) reduces thefMLP-induced PMN transendothelial migration. The ex-perimental conditions were the same as those described inFig. 2. In these experiments 51Cr-labeled PMNs wereadded to IL-1�-stimulated HUVECs grown on permeablesupports in the absence or presence of L658 758 (100�M), and PMN transendothelial migration in response toa fMLP gradient was assessed 30 minutes later. PMNtransendothelial migration was quantitated and was ex-pressed as the percentage of the total number of PMNsadded. Note that the fMLP-induced PMN transendothelialmigration was attenuated by the elastase inhibitor L658758 (n � 3).
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
214

teractions (i.e., adherence to and migration acrossHUVECs). Herein, we provided evidence that (1)there is sequential loss of staining for these AJ com-ponents with �-catenin being affected first, (2)PMN-derived elastase plays a role in the disorgani-zation of the AJs and cytoplasmic accumulation of�-catenin, and (3) antibodies directed to VE-cadherin can induce cytoplasmic accumulation of�-catenin.
In the absence of PMNs, immunofluorescence stain-ing for VE-cadherin, �-catenin, �-catenin, and�-catenin is continuous and is localized to the inter-cellular borders of endothelial cells (Fig. 1). When
PMNs interact with HUVECs, this staining can bediscontinuous (or interrupted) at sites adjacent toPMN–endothelial cell interactions (Fig. 2). We in-terpreted these findings to indicate that at sites ofPMN–endothelial cell interactions there is a loss ofthe component that had been immunofluorescentlystained. One caveat to this interpretation is that theindividual components of AJs may have been alteredin some manner by the PMN–endothelial cell inter-actions to render them undetectable by the antibod-ies used in the assay. Nevertheless, some undefinedalteration in the AJ components occurred only at theimmediate site of PMN–endothelial cell interactions(i.e., the staining for the AJ components remainedcontinuous at interendothelial junctions at sites fur-ther away from the vicinity of PMN–endothelial cellinteractions) (Fig. 2). These observations are thefirst to indicate that PMN-adhesive interactions canresult in focal alterations in AJ components.
Our findings that the continuous endothelial circum-ferential staining for all of the components of AJsstudied (i.e., VE-cadherin, �-catenin, �-catenin, and�-catenin) exhibits gaps at sites of PMN emigrationare consistent with those of previous studies usingmonocytes (1,45,46). In those studies, monocytetransendothelial cell migration was associated withfocal gaps in both VE-cadherin and �-catenin stain-ing. It was also noted that there was no interruptionin VE-cadherin staining when a monocyte was ad-herent to endothelial cells, but was not migrating.Our approach has allowed us to gain a more com-<
Figure 8. Effects of an elastase inhibitor (L658 758) onthe immunofluorescence staining of AJ components inHUVECs during PMN adhesive interactions. The experi-mental conditions were the same as those described in Fig.2. In these experiments, PMNs were added to HUVECs ata high PMN/HUVEC ratio (30:1) in the absence or pres-ence of the elastase inhibitor, L658 758 (100 �M), andthe staining of HUVEC AJ components was assessed 20minutes later. The samples were fixed, permeabilized, anddouble-immunostained for VE-cadherin (a and a�) and�-catenin (b and b�), and for �-catenin (c and c�) and�-catenin (d and d�) (red and green fluorescence, respec-tively, for each pair). The nuclei are labeled with Hoechst(blue-fluorescence). The images were made using separatechannels for each color, and are shown in black and white.Each panel represents an optical section through PMN/HUVEC coculture taken at the level of the interendothe-lial junctions. Note that the interaction of PMNs withHUVECs at a high PMN/HUVEC ratio resulted in wide-spread discontinuities in staining for each of the AJ com-ponents (a, b, c, and d). This effect was reduced by theelastase inhibitor, L658 758 (a�, b�, c�, and d�). Bars rep-resent 10 �m.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
215
prehensive insight into the effects of PMN–endothelial cell adhesive interactions on variouscomponents of the AJ. Using the dual-immuno-fluorescence protocol and LSCM analysis, adherentPMNs were captured that were not associated withgaps in staining for any of the AJ components (Fig.3), findings that were in accord with those of previ-ous studies using monocytes. However, an in-depthanalysis of PMN–endothelial cell interactions indi-cated that adherent PMNs could be captured thatwere associated with gaps in the staining of variouscomponents of the AJ complex (∼15% of adherentPMN). For example, in the vicinity of PMN adhe-sion, �-catenin staining exhibits gaps, whereas VE-cadherin staining does not (Fig. 4). Further studies
using dual-immunofluorescence staining of variouscombinations of the four AJ components allowed usto predict the sequence in which these componentswere affected by PMN adhesion to HUVECs. Basedon this iterative approach (Table 1), it seems that�-catenin staining is affected first, followed by stain-ing of VE-cadherin, �-catenin, and, finally,�-catenin. This novel observation suggests that ad-herent PMNs can induce a sequential loss or alter-ation in the components of the AJ, presumably inpreparation for transendothelial migration. Furtherstudies are warranted using live-cell, time-lapse mi-croscopy of adherent and migrating PMNs to estab-lish firmly this sequence of alterations in the variouscomponents of the AJ.
Figure 9. Purified neutrophilic elastase increases �-catenin staining in HUVECs. The experimental conditions were thesame as those described in Fig. 2, with the modification that, instead of PMNs, elastase (0.5 �g/mL) was used in theabsence or presence of an elastase inhibitor, L658 758 (100�M). The samples were fixed, permeabilized, and immu-nostained for �-catenin (green fluorescence in the upper panel, a–c). Nuclei are labeled with Hoechst (blue-fluorescence). The staining of �-catenin was assessed by LSCM in optical sections taken at the level of HUVEC nuclei(as indicated in the diagram below the image). Note that the staining of �-catenin remained continuous within theinterendothelial junctions under all experimental conditions. In addition, elastase induced an increase in the cytoplas-mic and nuclear staining of �-catenin, an effect that was attenuated by an elastase inhibitor, L658 758 (upper panel).The images presented were made at identical LSCM parameter settings. Bars � 10 �m. These results were confirmedby immunoblot analysis of cytoplasmic proteins extracted from HUVECs (lower panel). Lanes 1, 2, and 3 correspondto the images labeled a, b, and c, respectively.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
216
Previous studies have shown that in the immatureAJs of endothelial cells (early stages of confluence),VE-cadherin/catenin complexes consist of VE-cadherin the cytoplasmic domain of which binds to�-catenin, which, in turn, anchors the complex toactin via coupling with �-catenin. As the junctionmatures, �-catenin becomes associated with the VE-cadherin/catenin complexes at the expense of�-catenin (35). In response to monolayer wounding,the endothelial cells that begin to migrate disorga-nize their VE-cadherin/catenin complexes by a pro-cess that involves the loss of �-catenin from the com-plexes and the insertion of �-catenin (34). Fromthese observations, it has been suggested that thestrengthening of the VE-cadherin/catenin complexesinvolved the insertion of �-catenin into the com-plexes at the expense of �-catenin and that theweakening of the junctions involved the reverse pro-cess. Thus, it was unexpected that in the presentstudy adherent PMNs initially promoted the loss of�-catenin rather than �-catenin. We propose thatthe disorganization of the VE-cadherin/catenincomplexes by PMNs attempting to migrate betweenendothelial cells is a process that is different from
that involved in endothelial cell-initiated disorgani-zation of the complexes during their migration afterwounding. It seems teleologically appealing that thePMN offensive against the complexes would affectthe weaker of the junctional complexes (i.e., the VE-cadherin/catenin complex containing �-catenin).
Although not directly addressed in the present study,we have observed that the AJs reform after the PMNshave migrated across the monolayer (29). This ob-servation is consistent with those of previous reportsindicating that the AJs reseal after monocyte(1,46,45) or PMN transendothelial migration (22).Our preliminary findings using the dual-labeled im-munofluorescence approach indicate that stainingfor various components of the AJ during resealingalso may be sequential. For example, VE-cadherinand �-catenin staining reappear in the junctionsabove emigrated PMNs before �-catenin staining(29, and unpublished observations). Interestingly,these preliminary observations support the conten-tion that the formation of the junctions (immaturestage) involved the initial establishment of VE-cadherin/catenin complexes containing �-catenin.
Figure 10. Antibodies to VE-cadherin, but not PECAM-1, increase cytoplasmic �-catenin staining in HUVECs.HUVECs were seeded onto fibronectin-coated coverslips. Following treatment with Il-1� for 4 hours, the HUVECmonolayers were treated with fMLP (controls) or fMPL and 100 �g/mL rabbit polyclonal antihuman VE-cadherin for10 minutes. In another set of experiments, HUVECs were treated first with anti-PECAM-1 for 30 minutes, then withAlexa Fluor 488 goat antimouse IgG for 30 minutes. The samples were fixed, permeabilized, and immunostained for�-catenin as described in MATERIALS AND METHODS. The staining of �-catenin was assessed by LSCM in opticalsections taken at the level of HUVEC nuclei, and the images are presented in black and white. Note that the stainingof �-catenin remained continuous within the interendothelial junctions under all experimental conditions. In addition,in anti-VE-cadherin-treated HUVECs, there was an increase in the cytoplasmic immunostaining of �-catenin (arrowsin b) as compared to the control (a). No increase in immunostaining of cytoplasmic �-catenin was seen when PECAM-1was cross-linked (c). The images presented were made at identical LSCM parameter settings. Bars represent 10 �m.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
217

We are currently in the process of applying our it-erative approach to address this possibility more de-finitively.
The mechanism(s) by which PMNs induce disrup-tion of the AJ during diapedesis is not entirely clear.In previous studies, we have provided the followinglines of evidence to support a role for PMN-derivedelastase in the disruption of endothelial cell junctionsduring PMN diapedesis. A specific elastase inhibitoror a monoclonal antibody against elastase inhibitsPMN transendothelial migration in response to a va-riety of inflammatory mediators (8,9) (Fig. 7). Theendothelial cell retraction induced by the activatedPMNs can be prevented by elastase inhibitors. Acti-vated PMNs do not simply release elastase into theexternal milieu, rather they mobilize elastase to themembrane and localize it to the migrating front in-teracting with interendothelial cell junctions (9). Fi-nally, others have shown that activated PMNs candegrade both endothelial and epithelial cadherins,an event prevented by elastase inhibitors (5,19). Inthe present study, we provide further support for thecontention that PMN-derived elastase plays a role inthe disruption of the VE-cadherin/catenin com-plexes. When a large number of PMNs were inter-acted with HUVECs, there was widespread loss ofstaining for VE-cadherin, �-catenin, �-catenin, and�-catenin (Fig. 8). This widespread loss of stainingwas partially reversed if an elastase inhibitor wasincluded in the assay. The lack of complete reversalof the widespread loss of staining is most likely dueto redundancies in the mechanisms used by PMNs todisrupt AJs during diapedesis (e.g., recruitment ofother proteases to the migrating front).
That an intracellular component of the AJ (�-catenin) is the first one to disappear from the junc-tion implies that some outside-in signaling may beoccurring during PMN–HUVEC adhesive interac-tions. The results of previous studies have indicatedthat PMN–endothelial cell adhesive interactions in-duce a rise in endothelial cell intracellular calcium(26,49), and chelation of this calcium dramaticallyinhibits PMN transendothelial migration (26). How-ever, how the PMNs induce outside-in signaling isnot entirely clear. Previous studies (28) have indi-cated that the cross-linking of PECAM-1 results inthe phosphorylation of �-catenin, an event that mayinitiate its mobilization from the AJ complex to thecytoplasm. However, in the present study the cross-linking of neither PECAM-1 nor ICAM-1 resulted inthe cytoplasmic accumulation of �-catenin (Fig. 10).These latter observations suggest that these two ad-hesion molecules, which are intimately involved in
PMN transendothelial migration, are not the initia-tors of the outside-in signaling that leads to the cy-toplasmic accumulation of �-catenin. By contrast,antibodies directed to VE-cadherin induced an in-crease in the cytoplasmic accumulation of �-catenin(Fig. 10), indicating that VE-cadherin could be theinitiator of the outside-in signaling. How PMNs in-duce VE-cadherin to initiate this signaling is not en-tirely clear. However, elastase also can induce cyto-plasmic accumulation of �-catenin (Fig. 9). Theseobservations suggest a link between PMN-derivedelastase and VE-cadherin. Interestingly, elastase-mediated proteolysis of VE-cadherin is not a randomprocess but yields fragments of a specific molecularweight (5). Taken together, these observations sug-gest that PMN-derived elastase may partially andsite-specifically cleave VE-cadherin to induce its ac-tivation, a situation not unlike that described forprotease-activated receptors (37). This intriguingpossibility remains to be conclusively proven andwarrants attention.
As mentioned above, the addition of elastase or an-tibodies to VE-cadherin to HUVECs results in therapid appearance of �-catenin in the HUVEC cyto-plasm and nuclei (Figs. 9 and 10). This effect wasmuch more dramatic than when PMNs were used(compare to Fig. 8). This is probably a result ofpurified elastase interacting uniformly with the AJthroughout the monolayer, whereas elastase mobi-lized to the PMN membrane elicits a localized attackon the AJ. Similarly, antibodies to VE-cadherin alsointeracted with VE-cadherin in the intercellularjunctions uniformly rather than site specifically. Theappearance of �-catenin in the cytoplasm can resultin two scenarios. First, �-catenin could be targetedfor degradation. This possibility is supported bystudies (43) indicating that cytoplasmic �-catenin isubiquitinated and degraded by the proteosome path-way. Alternatively, cytoplasmic �-catenin could beinvolved in intracellular signaling. This possibility issupported by the results of the present study, indi-cating that �-catenin is present in HUVEC nuclei(Fig. 9). Interestingly, others (2,40) have implicated�-catenin as a transcriptional activator. Furtherstudies are warranted to address the fate of cytoplas-mic �-catenin (i.e., its degradation or participationin cell signaling).
REFERENCES
1. Allport JR, Muller WA, Luscinskas FW. (2000).Monocytes induce reversible focal changes in vascularendothelial cadherin complex during transendothelialmigration under flow. J Cell Biol 148:203–216.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
218

2. Aplin AE, Howe A, Alahari SK, Juliano RL. (1998).Signal transduction and signal modulation by cell ad-hesion receptors: the role of integrins, cadherins, im-munoglobulin-cell adhesion molecules, and selectins.Pharmacol Rev 50:197–263.
3. Berman ME, Muller WA. (1995). Ligation of platelet/endothelial cell adhesion molecule 1 (PECAM-1/CD31) on monocytes and neutrophils increases bind-ing capacity of leukocyte CR3 (CD11b/CD18). J Im-munol 154:299–307.
4. Burns AR, Walker DC, Brown ES, Thurmon LT,Bowden RA, Keese CR, Simon SI, Entman ML, SmithCW. (1997). Neutrophil transendothelial migration isindependent of tight junctions and occurs preferen-tially at tricellular corners. J Immunol 159:2893–2903.
5. Carden D, Xiao F, Moak C, Willis BH, Robinsonjackson S,Alexander S. (1998). Neutrophil elastase promoteslung microvascular injury and proteolysis of endothe-lial cadherins. Am J Physiol 44:H385–H392.
6. Carlos TM, Harlan JM. (1994). Leukocyte-endo-thelial adhesion molecules. Blood 84:2068–2101.
7. Cepinskas G, Noseworthy R, Kvietys PR. (1997).Transendothelial neutrophil migration: Role of neu-trophil-derived proteases and relationship to transen-dothelial protein movement. Circ Res 81:618–626.
8. Cepinskas G, Noseworthy R, Kvietys PR. (1997).Transendothelial neutrophil migration: Role of neu-trophil-derived proteases and relationship to transen-dothelial protein movement. Circ Res 81:618–626.
9. Cepinskas G, Sandig M, Kvietys PR. (1999). PAF-induced elastase-dependent neutrophil transendothe-lial migration is associated with the mobilization ofelastase to the neutrophil surface and localization tothe migrating front. J Cell Sci 112:1937–1945.
10. Cotran RS, Mayadas-Norton T. (1998). Endothelialadhesion molecules in health and disease. Pathol Biol(Paris) 46:164–170.
11. Dejana E, Valiron O, Navarro P, Lampugnani MG.(1997). Intercellular junctions in the endotheliumand the control of vascular permeability. Ann N YAcad Sci 811:36–43.
12. Del Maschio A, Zanetti A, Corada M, Rival Y, Ruco L,Lampugnani MG, Dejana E. (1996). Polymorpho-nuclear leukocyte adhesion triggers the disorganiza-tion of endothelial cell-to-cell adherens junctions. JCell Biol 135:497–510.
13. Etienne-Manneville S, Manneville JB, Adamson P,Wilbourn B, Greenwood J, Couraud PO. (2000).ICAM-1 coupled cytoskeletal rearrangements andtransendothelial lymphocyte migration involve intra-cellular calcium signaling in brain endothelial celllines. J Immunol 165:3375–3383.
14. Feng D, Nagy JA, Pyne K, Dvorak HF, Dvorak AM.(1998). Neutrophils emigrate from venules by atransendothelial cell pathway in response to FMLP. JExp Med 187:903–915.
15. Furie MB, Naprstek BL, Silverstein SC. (1987). Mi-gration of neutrophils across monolayers of culturedmicrovascular endothelial cells: An in vitro model ofleukocyte extravasation. J Cell Sci 88:161–175.
16. Gahmberg CG, Valmu L, Kotovuori A, Kotovuori P,Hilden TJ, Fagerholm S, Kantor C, Nurminen T, Iha-nus E, Tian L. (1999). Leukocyte adhesion: An inte-grated molecular process at the leukocyte plasmamembrane. Biosci Rep 19:273–281.
17. Garcia JN, Verin AD, Herenyiova M, English D.(1998). Adherent neutrophils activate endothelialmyosin light chain kinase: Role in transendothelialmigration. J Appl Physiol 84:1817–1821.
18. Gautam N, Hedqvist P, Lindbom L. (1998). Kineticsof leukocyte-induced changes in endothelial barrierfunction. Br J Pharmacol 125:1109–1114.
19. Ginzberg HG, Cherapanov V, Dong Q, Cantin A, Mc-Culloch CAG, Shannon PT, Downey GP. (2001).Neutrophil-mediated epithelial injury during trans-migration: role of elastase. Am J Physiol 281:G705–G717.
20. Gotsch U, Borges E, Bosse R, Boggemeyer E, SimonM, Mossmann H, Vestweber D. (1997). VE-cadherinantibody accelerates neutrophil recruitment in vivo. JCell Sci 110:583–588.
21. Granger DN, Kubes P. (1994). The microcirculationand inflammation: modulation of leukocyte-endothelial cell adhesion. J Leukoc Biol 55:662–675.
22. Granger DN, Yuan SY. Zawieja DC. (2000). Ultra-structural basis of leukocyte migration through themicrovascular membrane. In: Granger DN, Schmid-Schonbein GW, editors. Physiology and Pathophysi-ology of Leukocyte Adhesion. New York: Oxford Uni-versity Press, p 185–216.
23. Gulino D, Delachanal E, Concord E, Genoux Y,Morand B, Valiron MO, Sulpice E, Scaife R, AlemanyM, Vernet T. (1998). Alteration of endothelial cellmonolayer integrity triggers resynthesis of vascularendothelium cadherin. J Biol Chem 273:29786–29793.
24. Gurubhagavatula I, Amrani Y, Pratico D, Ruberg FL,Albelda SM, Panettieri RA, Jr. (1998). Engagementof human PECAM-1 (CD31) on human endothelialcells increases intracellular calcium ion concentrationand stimulates prostacyclin release. J Clin Invest 101:212–222.
25. Hordijk PL, Anthony E, Mul FJ, Rientsma R, OomenLM, Roos D. (1999). Vascular-endothelial-cadherinmodulates endothelial monolayer permeability. J CellSci 112:1915–1923.
26. Huang AJ, Manning JE, Bandak TM, Ratau MC, Han-ser KR, Silverstein SC. (1993). Endothelial cell cyto-solic free calcium regulates neutrophil migrationacross monolayers of endothelial cells. J Cell Biol 120:1371–1380.
27. Huber AR, Weiss SJ. (1989). Disruption of the sub-endothelial basement membrane during neutrophil
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
219

diapedesis in an in vitro construct of a blood vesselwall. J Clin Invest 83:1122–1136.
28. Ilan N, Mahooti S, Rimm DL, Madri JA. (1999). PE-CAM-1 (CD31) functions as a reservoir for and amodulator of tyrosine-phosphorylated beta-catenin. JCell Sci 112:3005–3014.
29. Ionescu CV, Cepinskas G, Savickiene J, Kvietys PR.(2001). Sequential recovery of endothelial adherensjunctions components after PMN transendothelial mi-gration (abstract). FASEB J 15:A831.
30. Kishimoto TK, Jutila MA, Berg EL, Butcher EC.(1989). Neutrophil Mac-1 and MEL-14 adhesionproteins inversely regulated by chemotactic factors.Science 245:1238–1241.
31. Kvietys PR, Cepinskas G, Granger DN. (1996). Neu-trophil-endothelial cell interactions during ischemia/reperfusion. In: Kamada T, Shiga T, McCuskey RS,editors. Tissue Reperfusion and Organ function: Is-chemia/Reperfusion Injury. Amsterdam: Elsevier, p179–191.
32. Kvietys PR, Granger DN. (1997). Endothelial cellmonolayers as a tool for studying microvascularpathophysiology. Am J Physiol 273:G1189–G1199.
33. Kvietys PR, Sandig M. (2001). Neutrophil diapedesis:Paracellular or transcellular. News Physiol Sci 16:15–19.
34. Lampugnani MG, Corada M, Andriopoulou P, EsserS, Risau W, Dejana E. (1997). Cell confluence regu-lates tyrosine phosphorylation of adherens junctioncomponents in endothelial cells. J Cell Sci 110:2065–2077.
35. Lampugnani MG, Corada M, Caveda L, Breviario F,Ayalono, Geiger B, Dejana E. (1995). The molecularorganization of endothelial cell to cell junctions: dif-ferential association of plakoglobin, beta-catenin, andalpha-catenin with vascular endothelial cadherin(VE-cadherin). J Cell Biol 129:203–217.
36. Lampugnani MG, Dejana E. (1997). Interendothelialjunctions: Structure, signalling and functional roles.Curr Opin Cell Biol 9:674–682.
37. Lindner JR, Kahn ML, Coughlin SR, Sambrano GR,Schauble E, Bernstein D, Foy D, Hafezi-MoghadamA, Ley K. (2000). Delayed onset of inflammation inprotease-activated receptor-2- deficient mice. J Im-munol 165:6504–6510.
38. Luu NT, Rainger GE, Nash GB. (1999). Kinetics ofthe different steps during neutrophil migrationthrough cultured endothelial monolayers treated withtumour necrosis factor-alpha. J Vasc Res 36:477–485.
39. Maurer P, Hohenester E, Engel J. (1996). Extracel-lular calcium-binding proteins. Curr Opin Cell Biol8:609–617.
40. Miller JR, Hocking AM, Brown JD, Moon RT. (1999).
Mechanism and function of signal transduction by theWnt/beta-catenin and Wnt/Ca2+ pathways. Onco-gene 18:7860–7872.
41. Moll T, Dejana E, Vestweber D. (1998). In vitro deg-radation of endothelial catenins by a neutrophil pro-tease. J Cell Biol 140:403–407.
42. Muller WA, Randolph GJ. (1999). Migration of leu-kocytes across endothelium and beyond: moleculesinvolved in the transmigration and fate of monocytes.J Leukoc Biol 66:698–704.
43. Orford K, Crockett C, Jensen JP, Weissman AM, By-ers SW. (1997). Serine phosphorylation-regulatedubiquitination and degradation of beta- catenin. JBiol Chem 272:24735–24738.
44. Panes J, Granger DN. (1998). Leukocyte-endothelialcell interactions: Molecular mechanisms and implica-tions in gastrointestinal disease. Gastroenterology114:1066–1090.
45. Sandig M, Korvemaker ML, Ionescu CV, Negrou E,Rogers KA. (1999). Transendothelial migration ofmonocytes in rat aorta: Distribution of F-actin, al-pha-catenin, LFA-1, and PECAM-1. Biotech Histo-chem 74:276–293.
46. Sandig M, Negrou E, Rogers KA. (1997). Changes inthe distribution of LFA-1, catenins, and F-actin dur-ing transendothelial migration of monocytes in cul-ture. J Cell Sci 110:2807–2818.
47. Springer TA. (1995). Traffic signals on endotheliumfor lymphocyte recirculation and leukocyte emigra-tion. Annu Rev Physiol 57:827–872.
48. Steinberg MS, McNutt PM. (1999). Cadherins andtheir connections: adhesion junctions have broaderfunctions. Curr Opin Cell Biol 11:554–560.
49. Su WH, Chen H, Huang J, Jen CJ. (2000). Endothe-lial [Ca(2+)](i) signaling during transmigration ofpolymorphonuclear leukocytes. Blood 96:3816–3822.
50. Tkalcevic J, Novelli M, Phylactides M, Iredale JP, Se-gal, AW, Roes J. (2000). Impaired immunity and en-hanced resistance to endotoxin in the absence of neu-trophil elastase and cathepsin G. Immunity 12:201–210.
51. Vestweber D, Blanks JE. (1999). Mechanisms thatregulate the function of the selectins and their li-gands. Physiol Rev 79:181–213.
52. Woodman RC, Reinhardt PH, Kanwar S, JohnstonFL, Kubes P. (1993). Effects of human neutrophil elas-tase (HNE) on neutrophil function in vitro and ininflamed microvessels. Blood 82:2188–2195.
53. Yoshida N, Granger DN, Anderson DC, Rothlein R,Lane C, Kvietys PR. (1992). Anoxia/reoxygenation-induced neutrophil adherence to cultured endothelialcells. Am J Physiol 262:H1891–H1898.
Changes in adherens junctions during PMN diapedesisCV Ionescu et al.
220




















