
Bacillus subtilis spores as vaccine adjuvants: further insights into the mechanisms of action
Upload
independentCategory
view
0download
0
International Journal of Food Microbiology 150 (2011) 59–65
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
Outgrowth inhibition of Clostridium beijerinckii spores by a bacteriocin-producinglactic culture in ovine milk cheese
Sonia Garde a,⁎, Marta Ávila a, Ramón Arias b, Pilar Gaya a, Manuel Nuñez a
a Departamento de Tecnología de Alimentos, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Carretera de La Coruña km 7, 28040 Madrid, Spainb Centro Regional de Selección y Reproducción Animal (CERSYRA) JCCM, Avenida del Vino n° 6, 13300 Valdepeñas, Spain
⁎ Corresponding author. Tel.: +34 91 3474037; fax:E-mail address: [email protected] (S. Garde).
0168-1605/$ – see front matter © 2011 Elsevier B.V. Aldoi:10.1016/j.ijfoodmicro.2011.07.018
a b s t r a c t
a r t i c l e i n f oArticle history:Received 10 May 2011Received in revised form 11 July 2011Accepted 14 July 2011Available online 22 July 2011
Keywords:Clostridium beijerinckiiCheese blowingOutgrowth inhibitionBacteriocin producerNisinLacticin 481
In the manufacture of model cheeses, ovine milk was deliberately contaminated with spores of Clostridiumbeijerinckii INIA 63, a wild isolate from Manchego cheese with late blowing defect, and inoculated with nisin-and lacticin 481-producing Lactococcus lactis subsp. lactis INIA 415 as starter, to test its potential to preventthe late blowing defect, or with L. lactis subsp. lactis INIA 415-2, a spontaneous mutant not producingbacteriocins. Cheeses made individually with the lactococcal strains, without clostridial spores, served ascontrols. Cheese made with clostridial spores and L. lactis subsp. lactis INIA 415-2 showed late blowing defectafter 120 days of ripening. Spoilt cheese also showed lower concentrations of lactic acid, and higher levels ofacetic, propionic and butyric acids, and of other volatile compounds such as 2-propanol and 1-butanol, thancontrol cheese. In addition, cheese made with the bacteriocin producer did not show any late blowingsymptoms, despite its spore counts similar to those of blown cheese, pointing to outgrowth inhibition ofC. beijerinckii spores by bacteriocins. Besides, cheese made with the bacteriocin producer showed similarconcentrations of lactic acid and volatile compounds than control cheese. Inclusion of L. lactis subsp. lactis INIA415 in starter cultures seems a feasible method to prevent late blowing defect in cheese without altering itssensory characteristics.
+34 91 3572293.
l rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Butyric acid fermentation, also known as late blowing defect, isone of the major causes of spoilage in semi-hard and hard ripenedcheeses, resulting in the appearance of texture and flavor defects thatgenerate severe economic losses in the cheese industry (McSweeneyand Fox, 2004). Butyric acid fermentation in cheese is caused by theoutgrowth of species of the genus Clostridium capable of fermentinglactic acid with production of butyric acid, acetic acid, carbon dioxideand hydrogen. Although Clostridium tyrobutyricum, an anaerobe Gram-positive sporeformingbacterium, is considered the primary cause of lateblowing in cheese, other clostridial species such as Clostridiumsporogenes, Clostridium beijerinckii and Clostridium butyricum also havebeen shown to significantly contribute to the appearance of this defectin cheese (Cocolin et al., 2004; Klijn et al., 1995; Le Bourhis et al., 2005,2007).
Clostridium spores coming from soil are able to develop in silage,contaminatemilk during themilkingprocess and survive pasteurization(DasGupta and Hull, 1989; Julien et al., 2008; Te Giffel et al., 2002;Vissers et al., 2006, 2007). Growth of Clostridium in cheese is criticallyinfluenced by lactic acid concentration, salt concentration, pH,moisture
and fat content, ripening time and temperature, and the presence ofother microorganisms (Goudkov and Sharpe, 1965).
Bactofugation and microfiltration of milk, and the addition ofnitrate or lysozyme are common strategies to prevent late blowingdefect (Lodi, 1990; Stadhouders, 1990; van den Berg et al., 2004).However, the reduction in spore numbers achieved by bactofugationmay be insufficient to prevent late blowing defect, microfiltration canbe applied only to skimmilk because milk fat globules are too large topass through the microfiltration membrane, and the use of chemicalsis precluded by the increasing demand for additive-free food products.
An alternative strategy to prevent butyric acid fermentation is theinoculation of milk with bacteriocinogenic lactic acid bacteria (LAB) incheese manufacture. Nevertheless, studies on the use of LAB for suchaim are scarce, and none of the existing studies have been carried out inovine milk cheeses, or investigated the inhibition of species other thanC. tyrobutyricum.Nisin Z-producing Lactococcus lactis ssp. lactis IPLA 729was successfully used to control the growth of C. tyrobutyricum CECT4011 in Vidiago cheese, a semi-hard bovinemilk cheese variety (Rilla etal., 2003), and bacteriocin-producing Lactobacillus gasseri K7 delayedblowing of cheeses artificially inoculatedwith C. tyrobutyricum (BogovicMatijasic et al., 2007). Lacticin 3147-producing L. lactis IFPL 3593controlled the outgrowthofClostridium spores and late blowing in semi-hard cheese (Martínez-Cuesta et al., 2010). Some of these studiesdescribed theeffect ofClostridiumand LABonorganic acidsbut the effecton the volatile fraction of cheese remains unknown.

60 S. Garde et al. / International Journal of Food Microbiology 150 (2011) 59–65
In a previous work, C. beijerinckii INIA 63 was isolated fromManchego cheese with late blowing defect, and characterized withrespect to its production of organic acids and sensitivity to LABbacteriocins together with other 222 Clostridium isolates (Garde et al.,2011). This strain showed the highest production of butyric acidand gas in milk, emerging as the best candidate to reproduce thelate blowing defect in experimental cheese. On the other hand,C. beijerinckii INIA 63 was sensitive to all bacteriocins tested, amongthem nisin Z and lacticin 481.
L. lactis subsp. lactis INIA 415, a nisin Z and lacticin 481 producer,was isolated from rawmilk Manchego cheese (Garde et al., 2001) andhas been successfully used in Hispánico cheese manufacture toaccelerate proteolysis and development of cheese flavor (Ávila et al.,2005; Garde et al., 2002a,b, 2005). In the present work, this strain wasused as starter culture in the manufacture of ovine milk cheeses inorder to prevent the late blowing defect caused by milk inoculationwith C. beijerinckii INIA 63.
2. Materials and methods
2.1. Bacterial strains and propagation
L. lactis subsp. lactis INIA 415, from the INIA culture collection, wasused as bacteriocin-producing mesophilic starter culture. L. lactissubsp. lactis INIA 415-2, a spontaneous nisin- and lacticin 481-resistantmutant not producing bacteriocins, with acid production andproteolytic activities similar to those of the parental strain, was alsoused asmesophilic starter culture. Theyweremaintained at−80 °C inMRS broth (Biolife, Milano, Italy) with 5% glycerol and subculturedtwice in reconstituted skim milk at 30 °C before use in cheesemanufacture. C. beijerinckii INIA 63, an isolate from Manchego cheesewith butyric fermentation defect, was used as blowing agent. It wasmaintained at −80 °C in Reinforced Clostridial Medium (RCM, Difco,Detroit, USA) with 5% glycerol and subcultured in RCM and incubatedat 37 °C for 48 h in anaerobic jars with an H2 plus CO2 generating kit(AnaeroGen, Oxoid, Basingstoke, UK).
2.2. Spore preparation
Spores of C. beijerinckii INIA 63 were prepared by incubation ofBryant and Burkey broth (BBB, Merck, Darmstadt, Germany) culturesat 37 °C under anaerobic conditions for 3 days. After centrifugation(5000×g, 15 min, 20 °C), the pellet was washed twice with steriledistilled water and resuspended in reconstituted skimmed milkbefore use in cheese manufacture. The spore concentration in thesuspension was determined, after heat treatment, by the mostprobable number (MPN) technique. Aliquots (1 mL) from serialdilutions of spore suspension prepared in sterile 0.1% (w/v) peptonesolution, were inoculated into three tube series containing 9 mL ofBBB. All the tubes were overlaid with 2 cm of sterile melted paraffinand heated to 75 °C for 10 min to kill vegetative cells. Incubation wascarried out at 37 °C for up to 7 days and daily tested for gasproduction. Spore counts were expressed as MPN/mL.
2.3. Manufacture of ovine milk cheeses
Model cheeses were manufactured from pasteurized ovine milkfrom the Manchega breed in duplicate experiments, carried out ondifferent days. Each experiment consisted of four vats, each containing5 L of milk, which was heated at 33 °C. Calcium chloride was added at1 g/L to all vats. L. lactis subsp. lactis INIA 415-2 and L. lactis subsp.lactis INIA 415 skimmedmilk cultures were added at 10 mL/L to vats 1and 3, and to vats 2 and 4, respectively. Vats 3 and 4 were deliberatelycontaminated with 2.2×105 spores/mL of C. beijerinckii INIA 63.Rennet (1 mL Maxiren, 165 IMCU/mL, DSM, Delft, The Netherlands)was added to milk 20 min later. After 40 min, the curds were cut into
6–8 mm cubes and scalded at 38 °C for 15 min. Whey was drained offand curds were washed with 1.6 L of distilled and sterilized water at38 °C, and distributed into cylindrical molds. Two cheeses, ofapproximately 500 g in weight, were obtained from each vat. Thecheeses were pressed overnight at 20 °C, salted for 20 min in brine(160 g NaCl/L), vacuum packaged in Cryovac plastic bags and ripenedat 12 °C for 120 days.
In order to elucidate the behavior of C. beijerinckii in cheese and theeffect of bacteriocins produced by L. lactis subsp. lactis INIA 415,cheeses were artificially contaminated with C. beijerinckii spores andmade with L. lactis subsp. lactis INIA 415-2 or L. lactis subsp. lactis INIA415 as described above. After 60 days of ripening at 12 °C, cheeseswhich did not show any symptoms of late blowing defect, wereincubated at 25 °C for 15 days to accelerate the development ofblowing defect.
2.4. Microbiological determinations
Representative cheese samples (5 g) were homogenized with45 mL of a sterile 2% (w/v) sodium citrate solution at 45 °C in aStomacher 400 (A. J. Seward Ltd, London, UK). Decimal dilutions ofcheese homogenates were prepared in sterile 0.1% (w/v) peptonesolution. Lactococci counts were determined in duplicate on platecount agar (Biolife, Milano, Italy) with 0.1% added skim milk (Biolife),after aerobic incubation for 24 h at 30 °C. Spore counts weredetermined by the most probable number (MPN) technique in BBBas described in Section 2.2.
For the determination of bacteriocin activity, cheese samples heldat −40 °C were thawed and 5 g were homogenized in a Stomacher400 with 10 mL sterile 0.02 N HCl at 50 °C. Homogenates werecentrifuged (12,000×g, 20 min, 4 °C) and pH of supernatants wasadjusted to pH 6 with 1 N NaOH. A volume of 50 μL of eachsupernatant was placed in duplicate into wells (5 mm diameter)made in plates of MRS agar (Biolife) inoculated with 0.1% of a 16 hculture of Lactobacillus buchneri St2A as the indicator microorganism.After incubation at 37 °C for 48 h, the diameter of the zone of growthinhibition was measured and bacteriocin activity was expressed inmillimeters. Bacteriocinwas also directly detected by placing pieces ofcheese on plates of MRS agar inoculated with L. buchneri St2A.
2.5. Chemical determinations
Cheese pH was measured in duplicate by means of a Crison pHmeter (model GPL 22, Crison Instruments, Barcelona, Spain) using aCrison penetration electrode (model 52-3.2). Dry matter content wasdetermined in duplicate after drying to constant weight in a vacuumoven at 100 °C.
Lactic acid was extracted from duplicate samples of cheese(Bouzas et al., 1991) and determined by HPLC as previously described(Garde et al., 2011). The results were expressed as g/kg of cheese.
Volatile compounds were extracted using an automatic dynamicheadspace device and analyzed by GC–MS. The analytical procedureand the identification and quantification of volatile compounds werecarried out as described by Garde et al. (2002a). Relative abundancesof compounds were expressed as percentages of their peak areas onthe peak area of the internal standard cyclohexanone.
2.6. Statistical analysis
Statistics were performed by means of SPSS Win 12.0 program(SPSS Inc. Chicago, IL 60611, USA). Multifactor analysis of variancewas carried out by using type of mesophilic starter and cheese age asmain effects. Comparison of means was performed by Tukey's test.Principal component analysis (PCA) with Varimax rotation wascarried out on lactic acid and volatile compounds.

Table 1Lactic acid bacteria (LAB) counts, spore counts and bacteriocin activity (BA) in cheesemanufactured from ovine milk inoculated with Lactococcus lactis subsp. lactis INIA 415-2(bacteriocin non-producer mutant, BNP) or L. lactis subsp. lactis INIA 415 (bacteriocinproducer, BP), and Clostridium beijerinckii INIA 63.
Age(days)
Without C. beijerinckii With C. beijerinckii
BNP BP BNP BP
LABa 1 9.90±0.04b 9.84±0.02b 9.82±0.09b 9.68±0.02a30 9.83±0.03b 9.81±0.04ab 9.85±0.01b 9.71±0.03a60 9.53±0.10bc 9.47±0.15a 9.58±0.11c 9.50±0.08ab90 9.26±0.05a 9.20±0.14a 9.22±0.02a 9.24±0.07a
120 8.98±0.15c 8.85±0.21ab 8.92±0.10bc 8.79±0.16aBAa 1 ND 27.3±1.1 ND 30.0±0.8
30 ND 22.0±0.3 ND 23.0±0.460 ND 21.3±0.8 ND 22.0±0.390 ND 21.5±1.0 ND 19.7±0.4
120 ND 19.2±0.7 ND 18.8±0.4Sporecountsa
1 0.48±0.28a b−0.05a 5.51±0.08b 5.65±0.01b30 0.38±0.23a b−0.05a 5.50±0.06b 5.45±0.07b60 0.28±0.16a 0.36±0.22a 5.34±0.19b 5.23±0.26b90 1.09±0.63a 0.14±0.14a 5.38±0.10b 5.44±0.23b
120 0.98±0.57a b−0.05a 5.50±0.08c 5.05±0.08b
a Mean±SE (n=4) of duplicate determinations in two experiments. LAB counts areexpressed as log cfu/g of cheese and spore counts as log MPN/g of cheese. Bacteriocinactivity was expressed as diameter (mm) of the zone of growth inhibition. ND, notdetected. Means in the same row followed by different letters differ (Pb0.05).
61S. Garde et al. / International Journal of Food Microbiology 150 (2011) 59–65
3. Results and discussion
3.1. Cheese pH and dry matter content
Cheese pH values increased gradually (Pb0.001) during ripening,with no significant influence of the starter type used in cheesemanufacture or the addition of Clostridium spores. The average pHvalue of cheeses at day 1 was 5.42, and it reached an average value of5.73 at 120 days of ripening (data not shown). With respect to drymatter content, no significant differences were found among cheeses(data not shown). Drymatter content increased (Pb0.001) from day 1(56.0%) to day 7 (58.9%) because of curd syneresis, and remainedconstant afterwards.
3.2. Lactic acid bacteria, bacteriocin activity and spore counts
Depending on the type of starter used in cheese manufacture,counts of LAB corresponded to L. lactis subsp. lactis INIA 415(bacteriocin producer, BP) or L. lactis subsp. lactis INIA 415-2(bacteriocin non-producer, BNP). LAB counts decreased (Pb0.001)gradually during ripening (Table 1) in all cheeses. All cheeses showedsimilar LAB counts throughout ripening, independently of the starter
B
A
1
Fig. 1. Cross section (1) and qualitative bacteriocin detection assay (2) of 120-day-old chee(A) Lactococcus lactis subsp. lactis INIA 415-2 (bacteriocin non-producer mutant, BNP) or
used for cheese manufacture. As expected, bacteriocin activity wasonly detected in cheeses made with the BP (Table 1, Fig. 1.2),decreasing (Pb0.001) throughout ripening. The addition of clostridialspores to milk did not affect (PN0.05) LAB counts or bacteriocinactivity in cheese.
In cheeses not contaminatedwith C. beijerinckii, lactate-fermentingspore forming microorganisms were detected, but counts remainedbelow 1.1 log cfu/g throughout ripening (Table 1). In cheesesartificially contaminatedwith C. beijerinckii, spores were concentratedin the curd approximately 2.5-fold with respect to milk, as theybecame entrapped into the casein matrix during whey drainage. Onlythe cheese made with the BNP culture and clostridial spores showedlate blowing after 120 days of ripening, despite the fact that sporecounts were not influenced (PN0.05) by cheese age or starter type(Fig. 1.1). At that time, spore count was only 0.45 log units lower incheese made with the BP culture than in cheese made with the BNPculture.
This narrowdifferencemight result from the fact that germination ofC. beijerinckii spores in cheese made with the BNP culture was followedby very low levels of sporulation of the vegetative cells. Le Bourhis et al.(2007) reported similar spore counts for cheeses made from milkinoculated with a C. sporogenes strain after 1 and 30 days of ripening at23 °C,which showed late blowing defect on day 30. In the presentwork,bacteriocins apparently inhibited outgrowth of C. beijerinckii spores,since spore counts remained fairly constant in cheese made from milkinoculated with the BP culture and clostridial spores throughout theripening period. Similarly, a nisin Z-producing L. lactis strain and alacticin 3147-producing L. lactis strain were successfully used to controlthe growth of C. tyrobutyricum in cheese (Martínez-Cuesta et al., 2010;Rilla et al., 2003) but, unlike our results, a sporicidal effect of bacteriocinson clostridial sporeswas observed by those authors. Both sporicidal andsporostatic effects of nisin have been reported depending on theClostridium strain (De Vuyst and Vandamme, 1994).
In order to elucidate the behavior of C. beijerinckii in cheese and theeffect of bacteriocins produced by L. lactis subsp. lactis INIA 415, 60-day-old cheeses artificially contaminated with C. beijerinckii spores,ripened at 12 °C and without late blowing defect, were incubated at25 °C for 15 days to accelerate the development of blowing defect(Fig. 2). Cheesemadewith the BNP culture showed blowing, voids andslits from day 3 onward (Fig. 2A). In this cheese, spore countsdecreased slightly (0.5 log units) after 7 days of incubation, but theyincreased by 4 log units after 15 days, indicating spore germination,and growth and sporulation of vegetative cells, respectively. However,cheese made with the BP culture did not show any late blowingsymptom (Fig. 2B) and spore counts did not vary during incubation,confirming the sporostatic effect of bacteriocins on C. beijerinckii.Therefore, the use of BP strain L. lactis subsp. lactis INIA 415 in cheesemanufacture controlled effectively the germination of C. beijerinckii
B
A
2
se manufactured from ovine milk inoculated with Clostridium beijerinckii INIA 63 and(B) L. lactis subsp. lactis INIA 415 (bacteriocin producer, BP).

Fig. 2. Photograph of 60-day-old cheese manufactured from ovine milk inoculated with Clostridium beijerinckii INIA 63 and (A) Lactococcus lactis subsp. lactis INIA 415-2 (bacteriocinnon-producer mutant, BNP) or (B) L. lactis subsp. lactis INIA 415 (bacteriocin producer, BP), after further incubation at 25 °C for 15 days.
62 S. Garde et al. / International Journal of Food Microbiology 150 (2011) 59–65
spores and prevented the appearance of the late blowing defect evenunder extreme conditions, favorable for the germination and growthof this clostridial strain.
3.3. Lactic acid and volatile compounds
Lactic acid concentration remained constant during ripening, atapproximately 11 g/kg of cheese, except in cheese made with the BNPculture and C. beijerinckii spores, in which it declined (Pb0.001) after120 days of ripening at concentrations approximately 8 times lowerthan in the rest of cheeses (data not shown), most probably due to itsconsumption by C. beijerinckii during the butyric acid fermentation.
5 10 15 20 25
Tim
Abu
ndan
ce (
x103 )
10
50
100
150
2009
19
23
10
50
100
150
200
1418
31
17
30
20
26
28
32
22
7
15
Fig. 3. Chromatographic profile corresponding to the headspace of 120-day-old cheese(A) Lactococcus lactis subsp. lactis INIA 415-2 (bacteriocin non-producer mutant, BNP) orcyclohexanone. The identity and relative abundance of each peak are described in Table 2.
A total of 57 volatile compounds were identified by GC–MS in 120-day-old ovine milk cheese, including mainly hydrocarbons, alcohols,ketones, aldehydes, acids, andesters (Fig. 3). The relative abundanceof 23volatile compoundswas significantly (Pb0.05) higher in the late blowingcheese, made with the BNP culture and C. beijerinckii spores, because ofthemetabolic activity of thismicroorganism(Table 2). To our knowledge,this is thefirst time that the volatile fraction of a cheesewith late blowingdefect has been studied. As the consequence of butyric acid fermentation,spoilt cheese showed higher concentrations of acetic and butyric acids inaccordance with the lower levels of lactic acid, and it also had higherlevels of propionic acid (Table 2). A correlation between propionic acidfermentation and butyric acid fermentation in late blowing Emmentalcheese artificially contaminated with a C. tyrobutyricum strain was also
30 35 40 45 50
e (min)
B
A
4647
37
36
39
40
45 50
52
53
56
57
48
CY
manufactured from ovine milk inoculated with Clostridium beijerinckii INIA 63 and(B) L. lactis subsp. lactis INIA 415 (bacteriocin producer, BP). CY: internal standard

Table 2Volatile compoundsa significantly affected by late-blowing defect in 120-day-old cheese manufactured from ovine milk inoculated with Lactococcus lactis subsp. lactis INIA 415-2(bacteriocin non-producer mutant, BNP) or L. lactis subsp. lactis INIA 415 (bacteriocin producer, BP), and Clostridium beijerinckii INIA 63.
Compounds (peak number) Without C. beijerinckii With C. beijerinckii
BNP BP BNP BP
Butanal (14) NDa NDa 0.60±0.20b NDa2-Methylbutanal (18) 0.09±0.01a 0.76±0.01b 2.22±0.12d 1.50±0.12cHexanal (31) 0.29±0.01a 0.25±0.02a 0.43±0.04b 0.23±0.02a2-Butanone (17) 0.92±0.02a 2.58±0.06b 105.67±8.82d 18.04±2.44c2,3-Pentanedione (30) 1.50±0.05a 4.69±0.10ab 33.65±0.85c 7.28±0.83b3-Hydroxy-2-pentanone (46) NDa NDa 7.42±0.53b NDa2-Hydroxy-3-pentanone (47) NDa NDa 4.54±0.30b NDa2-Propanol (20) 6.02±0.06b 2.68±0.22a 11.12±0.93c 1.33±0.18a2-Butanol (26) 1.64±0.02b NDa 50.66±1.69c NDa1-Propanol (28) 2.00±0.03a 1.81±0.05a 40.93±3.54b 6.82±0.33a1-Methoxy-2-propanol (36) NDa NDa 9.60±3.00b NDa1-Butanol (37) 8.91±1.22ab 0.99±0.03a 82.43±28.96b 1.39±0.04a2-Methyl-1-butanol (39) 0.85±0.03ab 0.98±0.04b 25.67±0.42c 1.38±0.23a3-Methyl-1-butanol (40) 1.20±0.04ab 1.53±0.08b 13.52±0.24c 0.47±0.12a3-Methyl-3-buten-1-ol (45) 1.08±0.02a 0.89±0.03a 1.45±0.08b 0.85±0.03a1-Octanol (54) 0.16±0.01a 0.15±0.01a 1.81±0.42b 0.12±0.04aAcetic acid (50) 0.19±0.05a 0.17±0.02a 1.68±0.18b 0.22±0.04aPropionic acid (52) NDa NDa 26.25±6.04b NDa2-Methylpropionic acid (53) NDa NDa 0.90±0.10b NDaButyric acid (56) 0.27±0.04a 0.14±0.01a 10.44±2.50b 0.42±0.16a2-Methylbutyric acid (57) NDa NDa 3.57±0.70b NDaEthyl ether (3) NDa NDa 1.13±0.16b NDaPropionic anhydride (22) NDa NDa 6.54±0.29b NDa
a Mean±SE (n=4) of duplicate determinations in two experiments. Volatile compounds are expressed as relative abundances to internal standard cyclohexanone. ND, notdetected. Means in the same row followed by different letters differ (Pb0.05).
63S. Garde et al. / International Journal of Food Microbiology 150 (2011) 59–65
found by Le Bourhis et al. (2007). On the contrary, they observed thatcheese contaminated with a reference strain of C. beijerinckii showedlower levels of propionic acid than control cheese, and suggested that itsless active metabolismmight be due to not being anymorewell adaptedto the cheese environment. Acetic acidhas been associated to vinegar andpungent odor notes, propionic acid to gas, burnt and pungent odor notes,and butyric acid to rancid, fecal and sweaty odor notes (Curioni andBosset, 2002).
C. beijerinckii can switch from acidogenesis to solventogenesis,characterized by the production of compounds like 2-propanol, butanaland 1-butanol (George and Chen, 1983; George et al., 1983; Yan et al.,1988). This fact would explain the higher levels of these compounds in
Principal component 1 (79% va
Prin
cipa
l com
pone
nt 2
(10
% v
aria
nce)
3-Methyl
1-Metho
2,3-Butanedione
2-Methylpro2-Propanone
3-Methylbutanal
Lactic acid
1.0
0.5
0.0
-0.5
-1.0
-1.0 -0.5 0.0
Fig. 4. Principal component analysis plot showing the distribution of lactic acid and volatLactococcus lactis subsp. lactis INIA 415-2 (bacteriocin non-producer mutant, BNP) or L. lact
spoilt cheese (Table 2). Winey and fruity odor descriptors have beenassigned for 2-propanol and 1-butanol, and peppery and pungent odordescriptors for butanal (Barron et al., 2005).
The cheese with late blowing defect also contained higher levels of2-methylbutanal, 2-methyl-1-butanol, 3-methyl-1-butanol, 2-methylpropionic and 2-methylbutyric acids (Table 2). Amino acidmetabolism is involved in the formation of these compounds incheese. Aldehydes 2-methyl-1-butanal and 3-methyl-1-butanal orig-inate from Ile and Leu, respectively, by transamination or Streckerdegradation, and alcohols 2-methyl-1-butanol and 3-methyl-1-butanol from the reduction of the two aldehydes (Curioni and Bosset,2002). High levels of these compounds have been associated with
Hexanal
riance)
-3-buten-1-ol
2-Propanol
Hexanal
Butanal
1-Butanolxy-2-propanol
2-Methylbutanal
panal
2-Butanol
3-Hydroxy-2-pentanone
2-Hydroxy-3-pentanone
2-Methyl-1-butanol
Propionic anhydride
3-Methyl-1-butanol
2-Methylpropanoic acid
1-Propanol
2-Butanone
2-Methylbutyric acid
2,3-Pentanedione
Acetic acid
Ethyl ether
Butyric acid
Propanoic acid
1-Octanol
0.5 1.0
ile compounds of 120-day-old cheese manufactured from ovine milk inoculated withis subsp. lactis INIA 415 (bacteriocin producer, BP) and Clostridium beijerinckii INIA 63.
Principal component 1 (79% variance)
Prin
cipa
l com
pone
nt 2
(10
% v
aria
nce)
1
0
-1
-2
-1 0 1 2
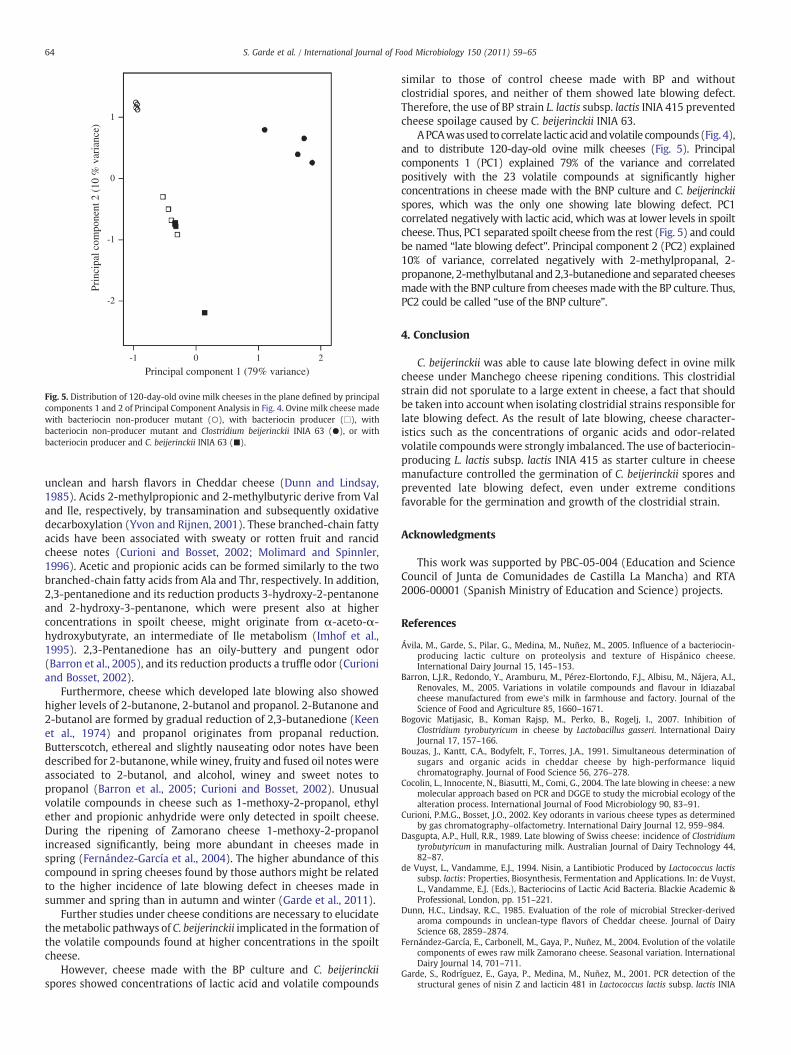
Fig. 5. Distribution of 120-day-old ovine milk cheeses in the plane defined by principalcomponents 1 and 2 of Principal Component Analysis in Fig. 4. Ovine milk cheese madewith bacteriocin non-producer mutant (○), with bacteriocin producer (□), withbacteriocin non-producer mutant and Clostridium beijerinckii INIA 63 (●), or withbacteriocin producer and C. beijerinckii INIA 63 (■).
64 S. Garde et al. / International Journal of Food Microbiology 150 (2011) 59–65
unclean and harsh flavors in Cheddar cheese (Dunn and Lindsay,1985). Acids 2-methylpropionic and 2-methylbutyric derive from Valand Ile, respectively, by transamination and subsequently oxidativedecarboxylation (Yvon and Rijnen, 2001). These branched-chain fattyacids have been associated with sweaty or rotten fruit and rancidcheese notes (Curioni and Bosset, 2002; Molimard and Spinnler,1996). Acetic and propionic acids can be formed similarly to the twobranched-chain fatty acids from Ala and Thr, respectively. In addition,2,3-pentanedione and its reduction products 3-hydroxy-2-pentanoneand 2-hydroxy-3-pentanone, which were present also at higherconcentrations in spoilt cheese, might originate from α-aceto-α-hydroxybutyrate, an intermediate of Ile metabolism (Imhof et al.,1995). 2,3-Pentanedione has an oily-buttery and pungent odor(Barron et al., 2005), and its reduction products a truffle odor (Curioniand Bosset, 2002).
Furthermore, cheese which developed late blowing also showedhigher levels of 2-butanone, 2-butanol and propanol. 2-Butanone and2-butanol are formed by gradual reduction of 2,3-butanedione (Keenet al., 1974) and propanol originates from propanal reduction.Butterscotch, ethereal and slightly nauseating odor notes have beendescribed for 2-butanone, while winey, fruity and fused oil notes wereassociated to 2-butanol, and alcohol, winey and sweet notes topropanol (Barron et al., 2005; Curioni and Bosset, 2002). Unusualvolatile compounds in cheese such as 1-methoxy-2-propanol, ethylether and propionic anhydride were only detected in spoilt cheese.During the ripening of Zamorano cheese 1-methoxy-2-propanolincreased significantly, being more abundant in cheeses made inspring (Fernández-García et al., 2004). The higher abundance of thiscompound in spring cheeses found by those authors might be relatedto the higher incidence of late blowing defect in cheeses made insummer and spring than in autumn and winter (Garde et al., 2011).
Further studies under cheese conditions are necessary to elucidatethemetabolic pathways of C. beijerinckii implicated in the formation ofthe volatile compounds found at higher concentrations in the spoiltcheese.
However, cheese made with the BP culture and C. beijerinckiispores showed concentrations of lactic acid and volatile compounds
similar to those of control cheese made with BP and withoutclostridial spores, and neither of them showed late blowing defect.Therefore, the use of BP strain L. lactis subsp. lactis INIA 415 preventedcheese spoilage caused by C. beijerinckii INIA 63.
APCAwasused to correlate lactic acid andvolatile compounds (Fig. 4),and to distribute 120-day-old ovine milk cheeses (Fig. 5). Principalcomponents 1 (PC1) explained 79% of the variance and correlatedpositively with the 23 volatile compounds at significantly higherconcentrations in cheese made with the BNP culture and C. beijerinckiispores, which was the only one showing late blowing defect. PC1correlated negatively with lactic acid, which was at lower levels in spoiltcheese. Thus, PC1 separated spoilt cheese from the rest (Fig. 5) and couldbe named “late blowing defect”. Principal component 2 (PC2) explained10% of variance, correlated negatively with 2-methylpropanal, 2-propanone, 2-methylbutanal and 2,3-butanedione and separated cheesesmadewith the BNP culture from cheesesmadewith the BP culture. Thus,PC2 could be called “use of the BNP culture”.
4. Conclusion
C. beijerinckii was able to cause late blowing defect in ovine milkcheese under Manchego cheese ripening conditions. This clostridialstrain did not sporulate to a large extent in cheese, a fact that shouldbe taken into account when isolating clostridial strains responsible forlate blowing defect. As the result of late blowing, cheese character-istics such as the concentrations of organic acids and odor-relatedvolatile compounds were strongly imbalanced. The use of bacteriocin-producing L. lactis subsp. lactis INIA 415 as starter culture in cheesemanufacture controlled the germination of C. beijerinckii spores andprevented late blowing defect, even under extreme conditionsfavorable for the germination and growth of the clostridial strain.
Acknowledgments
This work was supported by PBC-05-004 (Education and ScienceCouncil of Junta de Comunidades de Castilla La Mancha) and RTA2006-00001 (Spanish Ministry of Education and Science) projects.
References
Ávila, M., Garde, S., Pilar, G., Medina, M., Nuñez, M., 2005. Influence of a bacteriocin-producing lactic culture on proteolysis and texture of Hispánico cheese.International Dairy Journal 15, 145–153.
Barron, L.J.R., Redondo, Y., Aramburu, M., Pérez-Elortondo, F.J., Albisu, M., Nájera, A.I.,Renovales, M., 2005. Variations in volatile compounds and flavour in Idiazabalcheese manufactured from ewe's milk in farmhouse and factory. Journal of theScience of Food and Agriculture 85, 1660–1671.
Bogovic Matijasic, B., Koman Rajsp, M., Perko, B., Rogelj, I., 2007. Inhibition ofClostridium tyrobutyricum in cheese by Lactobacillus gasseri. International DairyJournal 17, 157–166.
Bouzas, J., Kantt, C.A., Bodyfelt, F., Torres, J.A., 1991. Simultaneous determination ofsugars and organic acids in cheddar cheese by high-performance liquidchromatography. Journal of Food Science 56, 276–278.
Cocolin, L., Innocente, N., Biasutti, M., Comi, G., 2004. The late blowing in cheese: a newmolecular approach based on PCR and DGGE to study the microbial ecology of thealteration process. International Journal of Food Microbiology 90, 83–91.
Curioni, P.M.G., Bosset, J.O., 2002. Key odorants in various cheese types as determinedby gas chromatography–olfactometry. International Dairy Journal 12, 959–984.
Dasgupta, A.P., Hull, R.R., 1989. Late blowing of Swiss cheese: incidence of Clostridiumtyrobutyricum in manufacturing milk. Australian Journal of Dairy Technology 44,82–87.
de Vuyst, L., Vandamme, E.J., 1994. Nisin, a Lantibiotic Produced by Lactococcus lactissubsp. lactis: Properties, Biosynthesis, Fermentation and Applications. In: de Vuyst,L., Vandamme, E.J. (Eds.), Bacteriocins of Lactic Acid Bacteria. Blackie Academic &Professional, London, pp. 151–221.
Dunn, H.C., Lindsay, R.C., 1985. Evaluation of the role of microbial Strecker-derivedaroma compounds in unclean-type flavors of Cheddar cheese. Journal of DairyScience 68, 2859–2874.
Fernández-García, E., Carbonell, M., Gaya, P., Nuñez, M., 2004. Evolution of the volatilecomponents of ewes raw milk Zamorano cheese. Seasonal variation. InternationalDairy Journal 14, 701–711.
Garde, S., Rodríguez, E., Gaya, P., Medina, M., Nuñez, M., 2001. PCR detection of thestructural genes of nisin Z and lacticin 481 in Lactococcus lactis subsp. lactis INIA

65S. Garde et al. / International Journal of Food Microbiology 150 (2011) 59–65
415, a strain isolated from raw milk Manchego cheese. Biotechnology Letters 23,85–89.
Garde, S., Carbonell, M., Fernández-García, E., Medina, M., Nuñez, M., 2002a. Volatilecompounds in Hispánico cheese manufactured using a mesophilic starter, athermophilic starter and bacteriocin-producing Lactococcus lactis subsp. lactis INIA415. Journal of Agricultural and Food Chemistry 50, 6752–6757.
Garde, S., Tomillo, J., Gaya, P., Medina, M., Nuñez, M., 2002b. Proteolysis in Hispánicocheese manufactured using a mesophilic starter, a thermophilic starter andbacteriocin-producing Lactococcus lactis subsp. lactis INIA 415 adjunct culture.Journal of Agricultural and Food Chemistry 50, 3479–3485.
Garde, S., Ávila, M., Medina, M., Nuñez, M., 2005. Influence of a bacteriocin-producinglactic culture on the volatile compounds, odour and aroma of Hispánico cheese.International Dairy Journal 15, 1034–1043.
Garde, S., Arias, R., Gaya, P., Nuñez, M., 2011. Occurrence of Clostridium spp. in ovinemilk and Manchego cheese with late blowing defect: identification andcharacterization of isolates. International Dairy Journal 21, 272–278.
George, H.A., Chen, J.S., 1983. Acidic conditions are not obligatory for onset of butanolformation by Clostridium beijerinckii (synonym, C. butylicum). Applied andEnvironmental Microbiology 46, 321–327.
George, H.A., Johnson, J.L., Moore, W.E.C., Holdeman, L.V., Chen, J.S., 1983. Acetone,isopropanol, and butanol production by Clostridium beijerinckii (syn. Clostridiumbutylicum) and Clostridium aurantibutyricum. Applied and Environmental Micro-biology 45, 1160–1163.
Goudkov, A.V., Sharpe, M.E., 1965. Clostridia in dairying. Journal of Applied Bacteriology28, 63–73.
Imhof, R., Glättli, H., Bosset, J.O., 1995. Volatile organic compounds produced bythermophilic and mesophilic single strain dairy starter cultures. LebensmittelWissenschaft und Technologie 28, 78–86.
Julien, M.C., Dion, P., Lafreniere, C., Antoun, H., Drouin, P., 2008. Sources of clostridia inraw milk on farms. Applied and Environmental Microbiology 74, 6348–6357.
Keen, A.R., Walker, N.J., Peberdy, M.F., 1974. The formation of 2-butanone and 2-butanolin Cheddar cheese. The Journal of Dairy Research 41, 249–257.
Klijn, N., Nieuwenhof, F.F.J., Hollwerf, J.D., Vanderwaals, C.B., Weerkamp, A.H., 1995.Identification of Clostridium tyrobutyricum as the causative agent of late blowing incheese by species-specific PCR amplification. Applied and Environmental Micro-biology 61, 2919–2924.
Le Bourhis, A.G., Saunier, K., Dore, J., Carlier, J.P., Chamba, J.F., Popoff, M.R., Tholozan, J.L.,2005. Development and validation of PCR primers to assess the diversity ofClostridium spp. in cheese by temporal temperature gradient gel electrophoresis.Applied and Environmental Microbiology 71, 29–38.
Le Bourhis, A.G., Dore, J., Carlier, J.P., Chamba, J.F., Popoff, M.R., Tholozan, J.L., 2007.Contribution of C. beijerinckii and C. sporogenes in association with C. tyrobutyricumto the butyric fermentation in Emmental type cheese. International Journal of FoodMicrobiology 113, 154–163.
Lodi, R., 1990. The use of lysozyme to control butyric acid fermentation. Bulletin of theInternational Dairy Federation 251, 51–54.
Martínez-Cuesta, M.C., Bengoechea, J., Bustos, I., Rodríguez, B., Requena, T., Pelaez, C.,2010. Control of late blowing in cheese by adding lacticin 3147-producingLactococcus lactis IFPL 3593 to the starter. International Dairy Journal 20, 18–24.
McSweeney, P.L.H., Fox, P.F., 2004. Metabolism of Residual Lactose and of Lactate andCitrate, In: Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P. (Eds.), Cheese:Chemistry, Physics and Microbiology, Vol. 1: General Aspects, 3rd ed. ElsevierAcademic Press, London, UK, pp. 361–371.
Molimard, P., Spinnler, H.E., 1996. Review: compounds involved in the flavor of surfacemold-ripened cheeses: origins and properties. Journal of Dairy Science 79,169–184.
Rilla, N., Martínez, B., Delgado, T., Rodríguez, A., 2003. Inhibition of Clostridiumtyrobutyricum in Vidiago cheese by Lactococcus lactis ssp. lactis IPLA 729, a nisin Zproducer. International Journal of Food Microbiology 85, 23–33.
Stadhouders, J., 1990. Prevention of butyric acid fermentation by the use of nitrate.Bulletin of the International Dairy Federation 251, 40–46.
Te Giffel, M.C.T., Wagendorp, A., Herrewegh, A., Driehuis, F., 2002. Bacterial spores insilage and raw milk. Antonie van Leeuwenhoek International Journal of Generaland Molecular Microbiology 81, 625–630.
van den Berg, G., Meijer, W.C., Düsterhöft, E.M., Smit, G., 2004. Gouda and RelatedCheeses, In: Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P. (Eds.), Cheese:Chemistry, Physics and Microbiology, Vol. 2: Major Cheese Groups, 3rd ed. ElsevierAcademic Press, London, pp. 103–140.
Vissers, M.M.M., Driehuis, F., Te Giffel, M.C., De Jong, P., Lankveld, J.M.G., 2006.Improving farm management by modeling the contamination of farm tank milkwith butyric acid bacteria. Journal of Dairy Science 89, 850–858.
Vissers, M.M.M., Driehuis, F., Te Giffel, M.C.T., De Jong, P., Lankveld, J.M.G., 2007.Concentrations of butyric acid bacteria spores in silage and relationships withaerobic deterioration. Journal of Dairy Science 90, 928–936.
Yan, R.T., Zhu, C.X., Golemboski, C., Chen, J.S., 1988. Expression of solvent-formingenzymes and onset of solvent production in batch cultures of Clostridiumbeijerinckii (“Clostridium butylicum”). Applied and Environmental Microbiology54, 642–648.
Yvon, M., Rijnen, L., 2001. Cheese flavour formation by amino acid catabolism.International Dairy Journal 11, 185–201.
Copyright © 2022 FDOKUMEN




















